
Identification and Characterization of NDT 9513727 [N bis...
12
Identification and Characterization of NDT 9513727 [N,N- bis(1,3-Benzodioxol-5-ylmethyl)-1-butyl-2,4-diphenyl-1H- imidazole-5-methanamine], a Novel, Orally Bioavailable C5a Receptor Inverse Agonist □ S Robbin M. Brodbeck, Daniel N. Cortright, Andrzej P. Kieltyka, Jianying Yu, Carolyn O. Baltazar, Marianne E. Buck, Robin Meade, George D. Maynard, Andrew Thurkauf, Du-Shieng Chien, Alan J. Hutchison, and James E. Krause Departments of Molecular Biology and Biochemistry (R.M.B., D.N.C., A.P.K., J.Y., C.O.B., M.E.B., R.M., J.E.K.), Drug Metabolism (D.-S.C.), and Medicinal Chemistry (G.D.M., A.T., A.J.H.), Neurogen Corporation, Branford, Connecticut Received May 28, 2008; accepted August 26, 2008 ABSTRACT The complement system represents an innate immune mech- anism of host defense that has three effector arms, the C3a receptor, the C5a receptor (C5aR), and the membrane attack complex. Because of its inflammatory and immune-enhancing properties, the biological activity of C5a and its classical recep- tor have been widely studied. Because specific antagonism of the C5aR could have therapeutic benefit without affecting the protective immune response, the C5aR continues to be a prom- ising target for pharmaceutical research. The lack of specific, potent and orally bioavailable small-molecule antagonists has limited the clinical investigation of the C5aR. We report the dis- covery of NDT 9513727 [N,N-bis(1,3-benzodioxol-5-ylmethyl)- 1-butyl-2,4-diphenyl-1H-imidazole-5-methanamine], a small- molecule, orally bioavailable, selective, and potent inverse agonist of the human C5aR. NDT 9513727 was discovered based on the integrated use of in vitro affinity and functional assays in conjunction with medicinal chemistry. NDT 9513727 inhibited C5a-stimulated responses, including guanosine 5-3- O-(thio)triphosphate binding, Ca 2 mobilization, oxidative burst, degranulation, cell surface CD11b expression and chemotaxis in various cell types with IC 50 s from 1.1 to 9.2 nM, respectively. In C5a competition radioligand binding experiments, NDT 9513727 exhibited an IC 50 of 11.6 nM. NDT 9513727 effectively inhibited C5a-induced neutropenia in gerbil and cynomolgus macaque in vivo. The findings suggest that NDT 9513727 may be a promising new entity for the treatment of human inflammatory diseases. The complement system is part of the innate immune system that plays an important role in host defense mecha- nisms and consists of more than 30 blood-borne and cellular proteins activated via three biochemical cascades of activa- tion, named the classical, alternative, and mannose-binding lectin pathways. Effector mechanisms of the complement system include two anaphylotoxin G-protein-coupled recep- tors, the C3a receptor and the C5aR (also known as CD88), and the membrane attack complex, C5b-9. C5a, a 74-amino acid four-helix bundle glycoprotein, is generated after local complement activation brought about by divergent stimuli including immune complexes, infectious agents, various bio- chemicals including polysaccharides and the amyloidogenic -peptide, and physical injury (Makrides, 1998). The C5aR is expressed on several cells of myeloid lineage including neu- trophils and mast cells, on Kupfer and stellate cells of the liver, and on subsets of smooth muscle, endothelial, epithe- lial, lymphocytic, astrocytic, and neuronal cells (Gerard and Gerard, 1994; Zwirner et al., 1999; Schieferdecker et al., 2001). C5aR is a G-protein-coupled receptor that functionally couples ligand binding to intracellular responses, including decreases in intracellular cAMP levels, increases in intracellular calcium . R.M.B. and D.N.C. contributed equally to this work. Article, publication date, and citation information can be found at http://jpet.aspetjournals.org. doi:10.1124/jpet.108.141572. □ S The online version of this article (available at http://jpet.aspetjournals.org) contains supplemental material. ABBREVIATIONS: C5aR, C5a receptor; AMD, age-related macular degeneration; r, rat; h, human; NDT 9517011, 4-(2-{[cyclohexyl(phenyl)meth- yl]amino}-2-oxoethyl)-N-(diphenylmethyl)-2-piperazinecarboxamide; CHO, Chinese hamster ovary; PBS, phosphate-buffered saline; BSA, bovine serum albumin; GTPS, guanosine 5-3-O-(thio)triphosphate; GTPS[ 35 S], guanosine 5-O-(3-[ 35 S]thiotriphosphate; KRH, Krebs-Ringer-HEPES; FLIPR, fluorometric imaging plate reader; m.o.i., multiplicity of infection; ROS, reactive oxygen species; Bay K8644, S-()-1,4-dihydro-2,6- dimethyl-5-nitro-4-(2-[trifluoromethyl]phenyl)-3-pyridine carboxylic acid methyl ester. 0022-3565/08/3273-898–909$20.00 THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 327, No. 3 Copyright © 2008 by The American Society for Pharmacology and Experimental Therapeutics 141572/3402437 JPET 327:898–909, 2008 Printed in U.S.A. 898 http://jpet.aspetjournals.org/content/suppl/2008/08/27/jpet.108.141572.DC1 Supplemental material to this article can be found at: at ASPET Journals on May 16, 2018 jpet.aspetjournals.org Downloaded from
-
Upload
truongduong -
Category
Documents
-
view
215 -
download
1
Transcript of Identification and Characterization of NDT 9513727 [N bis...

Identification and Characterization of NDT 9513727 [N,N-bis(1,3-Benzodioxol-5-ylmethyl)-1-butyl-2,4-diphenyl-1H-imidazole-5-methanamine], a Novel, Orally Bioavailable C5aReceptor Inverse Agonist□S
Robbin M. Brodbeck, Daniel N. Cortright, Andrzej P. Kieltyka, Jianying Yu,Carolyn O. Baltazar, Marianne E. Buck, Robin Meade, George D. Maynard,Andrew Thurkauf, Du-Shieng Chien, Alan J. Hutchison, and James E. KrauseDepartments of Molecular Biology and Biochemistry (R.M.B., D.N.C., A.P.K., J.Y., C.O.B., M.E.B., R.M., J.E.K.),Drug Metabolism (D.-S.C.), and Medicinal Chemistry (G.D.M., A.T., A.J.H.), Neurogen Corporation, Branford, Connecticut
Received May 28, 2008; accepted August 26, 2008
ABSTRACTThe complement system represents an innate immune mech-anism of host defense that has three effector arms, the C3areceptor, the C5a receptor (C5aR), and the membrane attackcomplex. Because of its inflammatory and immune-enhancingproperties, the biological activity of C5a and its classical recep-tor have been widely studied. Because specific antagonism ofthe C5aR could have therapeutic benefit without affecting theprotective immune response, the C5aR continues to be a prom-ising target for pharmaceutical research. The lack of specific,potent and orally bioavailable small-molecule antagonists haslimited the clinical investigation of the C5aR. We report the dis-covery of NDT 9513727 [N,N-bis(1,3-benzodioxol-5-ylmethyl)-1-butyl-2,4-diphenyl-1H-imidazole-5-methanamine], a small-
molecule, orally bioavailable, selective, and potent inverseagonist of the human C5aR. NDT 9513727 was discoveredbased on the integrated use of in vitro affinity and functionalassays in conjunction with medicinal chemistry. NDT 9513727inhibited C5a-stimulated responses, including guanosine 5�-3-O-(thio)triphosphate binding, Ca2� mobilization, oxidative burst,degranulation, cell surface CD11b expression and chemotaxis invarious cell types with IC50s from 1.1 to 9.2 nM, respectively. InC5a competition radioligand binding experiments, NDT 9513727exhibited an IC50 of 11.6 nM. NDT 9513727 effectively inhibitedC5a-induced neutropenia in gerbil and cynomolgus macaque invivo. The findings suggest that NDT 9513727 may be a promisingnew entity for the treatment of human inflammatory diseases.
The complement system is part of the innate immunesystem that plays an important role in host defense mecha-nisms and consists of more than 30 blood-borne and cellularproteins activated via three biochemical cascades of activa-tion, named the classical, alternative, and mannose-bindinglectin pathways. Effector mechanisms of the complementsystem include two anaphylotoxin G-protein-coupled recep-tors, the C3a receptor and the C5aR (also known as CD88),
and the membrane attack complex, C5b-9. C5a, a 74-aminoacid four-helix bundle glycoprotein, is generated after localcomplement activation brought about by divergent stimuliincluding immune complexes, infectious agents, various bio-chemicals including polysaccharides and the amyloidogenic�-peptide, and physical injury (Makrides, 1998). The C5aR isexpressed on several cells of myeloid lineage including neu-trophils and mast cells, on Kupfer and stellate cells of theliver, and on subsets of smooth muscle, endothelial, epithe-lial, lymphocytic, astrocytic, and neuronal cells (Gerard andGerard, 1994; Zwirner et al., 1999; Schieferdecker et al., 2001).C5aR is a G-protein-coupled receptor that functionally couplesligand binding to intracellular responses, including decreases inintracellular cAMP levels, increases in intracellular calcium
.R.M.B. and D.N.C. contributed equally to this work.Article, publication date, and citation information can be found at
http://jpet.aspetjournals.org.doi:10.1124/jpet.108.141572.□S The online version of this article (available at http://jpet.aspetjournals.org)
contains supplemental material.
ABBREVIATIONS: C5aR, C5a receptor; AMD, age-related macular degeneration; r, rat; h, human; NDT 9517011, 4-(2-{[cyclohexyl(phenyl)meth-yl]amino}-2-oxoethyl)-N-(diphenylmethyl)-2-piperazinecarboxamide; CHO, Chinese hamster ovary; PBS, phosphate-buffered saline; BSA, bovineserum albumin; GTP�S, guanosine 5�-3-O-(thio)triphosphate; GTP�S[35S], guanosine 5�-O-(3-[35S]thiotriphosphate; KRH, Krebs-Ringer-HEPES;FLIPR, fluorometric imaging plate reader; m.o.i., multiplicity of infection; ROS, reactive oxygen species; Bay K8644, S-(�)-1,4-dihydro-2,6-dimethyl-5-nitro-4-(2-[trifluoromethyl]phenyl)-3-pyridine carboxylic acid methyl ester.
0022-3565/08/3273-898–909$20.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 327, No. 3Copyright © 2008 by The American Society for Pharmacology and Experimental Therapeutics 141572/3402437JPET 327:898–909, 2008 Printed in U.S.A.
898
http://jpet.aspetjournals.org/content/suppl/2008/08/27/jpet.108.141572.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

levels, and activation of phosphatidylinositol 3-kinase and mi-togen-activated protein kinase activities (Gerard and Gerard,1994). The cellular consequences of C5aR activation includeclassic neutrophil activation events, such as chemotaxis, cellu-lar adhesion, and degranulation. In addition, C5aR activationhas been shown to influence cytokine and chemokine gene ex-pression and cellular apoptosis (Guo et al., 2000; Grant et al.,2002).
Because of the proinflammatory nature of C5a, C5aR an-tagonists have been sought as potential therapeutic agentsfor disorders, such as rheumatoid arthritis, asthma, respira-tory distress syndrome, ischemia-reperfusion injury, sepsis,psoriasis, inflammatory bowel disease, and neuroinflamma-tory disorders (Guo and Ward, 2005). Several research ad-vances, including C5aR knockout mice, recombinant C5a pro-tein antagonists, and C5a and C5 convertase-neutralizingantibodies, have provided important evidence for the involve-ment of the C5aR in various inflammatory disorders (Wanget al., 1995; Pellas et al., 1998; Gerard and Gerard, 2002; Guoand Ward, 2005; Solomon et al., 2005; Kohl, 2006). In thecase of rheumatoid arthritis, the role of the C5aR in diseasepathogenesis and progression has been elucidated using theK/BxN T-cell receptor transgenic mouse model of inflamma-tory synovitis (Ji et al., 2002). In a series of elegant studiesusing several complement pathway deficiencies in mice, theseinvestigators demonstrated that the inflammatory arthriticstate was critically dependent on C5aR, whereas disruption ofthe C5b-9 complex had no effect on disease pathogenesis. Hu-man genetic studies implicate increased complement activity inthe pathophysiology of age-related macular degeneration(Hageman et al., 2006). Utility of C5aR antagonism in AMD issupported by a recent study in C5a receptor-null mice (Nozakiet al., 2006).
Additional evidence for the utility of a C5aR blocker as atherapeutic comes from the activity of peptide-based C5aRantagonists in disease models and inflammatory states, suchas reverse passive Arthus and dermal inflammation, anti-gen-induced arthritis, sepsis and related models of endotoxicshock, ischemia-reperfusion injury, and inflammatory boweldisease (Guo and Ward, 2005; Solomon et al., 2005; Kohl,2006).
Potent, selective, and orally bioavailable small-moleculeC5aR antagonists will be invaluable in the study of the roleof C5aR in human inflammatory pathophysiology. Herein,we describe the properties of a compound from our small-molecule C5aR discovery program, NDT 9513727. We iden-tified C5aR modulators using radioligand binding. Subse-quently, we differentiated chemical series based on the lackof agonistic properties at the C5aR, a process that led to theidentification of NDT 9513727. We then used several in vitrofunctional assays to demonstrate that NDT 9513727 is apotent competitive antagonist with properties consistentwith inverse agonism. Finally, NDT 9513727 displays activ-ity in a human ex vivo blood assay and in C5a-inducedneutropenia gerbil and macaque models in vivo.
Materials and MethodsMaterials. Recombinant expression of rat (r) and human (h) C5a
and hC5a desArg was performed in an Escherichia coli system asdescribed previously (Okinaga et al., 2003). Recombinant (r) humanC5a was purchased from Sigma-Aldrich (St. Louis, MO). NDT
9513727 was synthesized at Neurogen according to procedures re-ported in Thurkauf et al. (2002) and Luke et al. (2003); concentra-tions of this compound are stated as the free-base equivalent. Thecyclic peptide C5a antagonists AcF[OPdCHAWr] and F[OPdChaWr](Finch et al., 1999) were synthesized as described previously (Waterset al., 2005). The small-molecule agonist NDT 9517011 was synthe-sized at Neurogen.
Cell Lines and Cell Culture. CHO, U937, HL-60, and AtT-20cells were obtained from the American Type Cell Culture (Manas-sass, VA). For the functional studies described below, U937 cellswere differentiated in RPMI 1640 containing 10% fetal bovine serumand 1 mM dibutyryl cAMP for 48 h at 37°C, and HL-60 cells weredifferentiated in RPMI 1640 containing 20% fetal bovine serum and1 mM dibutyryl cAMP for 48 h at 37°C. AtT-20 cells were cultured inF10 medium containing 15% horse serum and 2.5% fetal bovineserum. hC5aR was isolated from a human fetal brain cDNA libraryby standard hybridization techniques, and its sequence was deter-mined. The hC5aR cDNA was cloned into the pcDNA3.1 mammalianexpression plasmid (Invitrogen, Carlsbad, CA) and transfected intoCHO cells. A CHO cell line expressing high levels of hC5aR wassubsequently selected by G418 (Sigma-Aldrich) resistance. The CHOcell line expressing C5L2 was described previously (Okinaga et al.,2003).
Recombinant Expression of the Human C5a Receptor inBaculovirus-Infected Sf9 Cells. The hC5aR cDNA was clonedinto the baculovirus expression vector, pBacPak9 (Clontech, Moun-tain View, CA). Specific C5a receptor- and G-protein subunit-con-taining baculoviral expression vectors were cotransfected along withBaculoGold DNA (BD Biosciences Pharmingen, San Diego, CA) intoSf9 cells. The Sf9 cell culture supernatant was harvested 3 daysafter transfection. The recombinant virus-containing supernatantwas serially diluted in Hink’s TNM-FH insect medium (JRH Bio-sciences, Lenexa, KS)-supplemented Grace’s salts and with 4.1 mML-Gln, 3.3 g/l lactalbumin hydrolysate, 3.3 g/l ultrafiltered yeasto-late, and 10% heat-inactivated fetal bovine serum (hereafter “insectmedium”) and plaque assayed to titer the viral stock for recombinantplaques. Recombinant baculoviral clones were identified, passage 3baculoviral stock was titered via plaque assay, and a multiplicity ofinfection (m.o.i.) and incubation time course experiment was carriedout to determine conditions for optimal receptor expression. Resultsfrom these studies showed that an m.o.i. of 0.1 and a 72-h incubationwere parameters that achieved optimal hC5a receptor expression inup to 1 liter of Sf9 cultures.
Log-phase Sf9 cells were infected with recombinant baculovirusfollowed by culture in insect medium at 27°C. Infections were carriedout either with the hC5a receptor alone or with the hC5a receptorin combination with G-protein-encoding virus stocks obtainedfrom BioSignal Packard, Inc. (Montreal, QC, Canada). The het-erotrimeric G-protein subunits expressing viral stocks were ratG�i2-encoding virus stock (BioSignal Packard, Inc. no. V5J008),bovine G�1-encoding virus stock (BioSignal Packard, Inc. no.V5H012), and human G�2-encoding virus stock (BioSignal Pack-ard, Inc. no. V6B003). The infections were carried out at an m.o.i.of 0.1:1.0:0.5:0.5 for hC5aR, G�i2, G�1, and G�2, respectively, andharvested 72 h after infection. A cell suspension aliquot wasanalyzed for viability by trypan blue dye exclusion, and the re-maining Sf 9 cells were harvested via centrifugation (3000 rpm/10min/4°C).
Affinity Binding with [125I]hC5a. Sf9 cell pellets expressinghC5aR, G�i2, �1, and �2 were resuspended in homogenization buffer(10 mM HEPES, 250 mM sucrose, 0.5 �g/ml leupeptin, 2 �g/mlaprotinin, 200 �M phenylmethylsulfonyl fluoride, and 2.5 mMEDTA, pH 7.4) and homogenized using a Polytron homogenizer(setting 5 for 30 s). The homogenate was centrifuged (536g/10 min/4°C) to pellet the nuclei, and the supernatant containing plasmamembranes was decanted to a clean centrifuge tube, centrifuged(48,000g/30 min, 4°C), and resuspended in 30 ml of homogenizationbuffer. This centrifugation and resuspension step was repeated
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 899
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

twice. The final pellet was resuspended in ice-cold Dulbecco’s PBScontaining 5 mM EDTA and stored in frozen aliquots at �80°C untilneeded. The protein concentration of the resulting membrane prep-aration was measured using the Bradford protein assay (Bio-Rad,Hercules, CA). P2 membranes were resuspended by Dounce homog-enization in binding buffer (50 mM HEPES, pH 7.6, 120 mM NaCl,1 mM CaCl2, 5 mM MgCl2, 0.1% BSA, pH 7.4, 0.1 mM bacitracin, 100KIU/ml aprotinin). P2 membrane preparations were obtained fortissue binding studies. Canine, rat, rabbit, mouse, gerbil, and cyno-molgus macaque monkey (Macaque fasicularis) lung tissues wereobtained from Covance Research Products (Princeton, NJ) or Pel-Freeze, Inc. (Rogers, AR) and were stored at �80°C until preparationof membranes. Lung was minced into small pieces, thawed in ice-coldphosphate-buffered saline, rinsed with the same buffer, and homog-enized at setting 5 in a Polytron homogenizer in a buffer containing1 mM phenanthrolene in 50 mM Tris, pH 7.0. The homogenate wascentrifuged at 2300 rpm in a Sorvall 5C rotor (Sorvall, Newton, CT)for 30 min at 4°C, and the supernatant was harvested and filteredthrough gauze. The filtered supernatant was centrifuged at 20,000rpm in the same rotor for 90 min. The supernatant was decanted,and the pellet was resuspended in the buffer described above. Thesuspension was centrifuged 10 min at 20,000 rpm, and the P2 mem-branes were resuspended at a protein concentration of 2 mg/ml in 25mM HEPES, pH 7.2, containing 0.2 mg/ml each of aprotinin andleupeptin.
For saturation binding analysis, membranes (5 �g of protein) wereadded to polypropylene tubes containing 0.005 to 0.5 nM [125I]hC5a(recombinant; PerkinElmer Life and Analytical Sciences, Waltham,MA) and the binding buffer with the final concentration of additivesas stated above. Nonspecific binding was determined in the presenceof 300 nM hC5a (Sigma-Aldrich) and accounted for less than 10% oftotal binding.
For competition analysis, Sf9 membranes (5 �g) and lung mem-branes (150 �g) were added to polypropylene tubes containing 0.030nM [125I]C5a for Sf9 membrane binding studies or 0.050 to 0.10 nM[125I]C5a for tissue binding analysis. NDT 9513727 was added toseparate assay tubes at concentrations ranging from 10�10 to 10�5 Mto yield a final volume of 0.250 ml. Nonspecific binding was deter-mined in the presence of 300 nM hC5a (Sigma-Aldrich) and ac-counted for less than 10% of total binding. After a 2-h incubation atroom temperature, reactions were terminated by rapid vacuum fil-tration. Samples were filtered over presoaked (in 1.0% polyethyleni-mine for 2 h before use) GF/C Whatman filters and rinsed two timeswith 5 ml of cold binding buffer without BSA, bacitracin, or aproti-nin. Remaining bound radioactivity was quantified by gamma count-ing. IC50 and Hill coefficient (nH) were determined by plotting thedata as a log-logit function, and the 50% inhibition point was deter-mined by using the linear portion of data plot. The Hill coefficientwas determined as the slope factor of the resultant linear trans-formed data.
GTP�[35S] Binding Assays. GTP�[35S] binding activity wasmeasured using a modification of a previously described method(Weiland and Jacobs, 1994). Sf9 cells recombinantly expressing thehuman C5aR and the G-protein subunits �i2, �1, and �2 were used toprepare P2 membranes as described in the affinity binding protocol.On the assay day, thawed membrane preparations were resuspendedin GTP[35S] binding assay buffer (50 mM Tris, pH 7.0, 120 mM NaCl,2 mM MgCl2, 2 mM EGTA, 0.1% BSA, 0.1 mM bacitracin, 100KIU/ml aprotinin, 5 �M GDP) and added to reaction tubes at aconcentration of 30 �g/reaction tube. After adding NDT 9513727 atconcentrations ranging from 0.01 nM to 10 �M, reactions were ini-tiated by the simultaneous addition of 100 pM GTP�[35S] and humanrecombinant hC5a (Sigma-Aldrich) ranging in concentration from0.001 nM to 10.0 �M (final assay volume of 0.250 ml). After 60-minincubation at room temperature, the reaction was terminated byvacuum filtration over GF/C filters (presoaked in wash buffer con-taining 0.1% bovine serum albumin) with ice-cold wash buffer (50mM Tris, pH 7.0, containing 120 mM NaCl). Bound GTP�[35S] was
determined by liquid scintillation spectrometry. Nonspecific bindingwas defined by 10 �M GTP�S and represented less than 5% of thetotal binding.
Inverse agonist activity of NDT 9513727 was assessed in a modi-fied GTP�[35S] binding assay, wherein the baseline activity waselevated by lowering the GDP concentration in the assay to 1.0 �Mand performing an NDT 9513727 dose-response study in the absenceof C5a. NDT 9513727 was assessed as to its mechanistic type ofantagonism by evaluation in the GTP�[35S] binding assay usingthe Schild analysis (Arunlakshana and Schild, 1959). The Schildanalysis evaluated C5a-stimulated GTP�[35S] binding dose-responsecurves at four different NDT 9513727 concentrations.
Intracellular Calcium Mobilization. Differentiated U937 cellswere pelleted by centrifugation and resuspended in Krebs-Ringer-HEPES (KRH) solution (25 mM HEPES, 5 mM KCl, 0.96 mMNaH2PO4, 1 mM MgSO4, 2 mM CaCl2, 5 mM glucose, pH 7.4)containing Fluo-3 (Invitrogen, Eugene, OR) calcium-sensitive dye(10 �g/ml). The cell suspension was incubated at 37°C for 30 min.Cells were pelleted, washed, repelleted, and resuspended at a den-sity of 1 � 106 cells/ml. Cells were seeded into 96-well plates suitablefor use in the fluorometric imaging plate reader (FLIPR, model no. 1;Molecular Devices, Sunnyvale CA). Fluorescence responses, mea-sured by excitation at 480 nM and emission at 530 nM, were moni-tored upon the addition of 0.01 to 30 nM human C5a to an FLIPR.
Differentiated U937 cells were incubated with varying concentra-tions of NDT 9513727, ranging from 0.1 nM to 4 �M, for 1 h beforethe addition of 0.3 nM hC5a (an EC50 concentration). The fluores-cence response, upon the subsequent addition of the EC50 concentra-tion of hC5a, was then monitored using an FLIPR. The IC50 and Hillcoefficient of the C5a response were determined as with the radioli-gand binding data.
C5a-Induced U937 and Neutrophil Cell Chemotaxis Assays.NDT 9513727 was evaluated for inhibition of C5a-induced chemo-taxis of either freshly prepared human neutrophils or differentiatedU937 cells. Fresh human neutrophils were isolated from whole bloodusing a Polymorphprep gradient (Nycomed, Oslo, Norway). Fortymilliliters of human blood yielded approximately 30 million neutro-phils with greater than 95% viability as assessed by trypan bluestaining. Cells were sedimented by centrifugation and incubatedwith 8 �g/ml calcein-AM fluorescent dye (Molecular Probes) for 30min at 37°C. Cells were sedimented by centrifugation, resuspendedin phenol red-free RPMI medium (Invitrogen) containing 0.1% fetalbovine serum, sedimented again by centrifugation, and resuspendedat a concentration of 3 � 106 cells/ml in phenol red-free RPMI 1640medium containing 0.1% fetal bovine serum. One hundred fifty-microliter cell suspensions were mixed in assay tubes containing 1.5�l of NDT 9513727 (10�11 to 10�6 M final concentrations) in di-methyl sulfoxide. Cells were incubated with compound for 30 min atroom temperature. Neuroprobe ChemoTx 96-well plates (NeuroProbe, Gaithersburg, MD) were used to assess hC5a-activated che-motaxis. The wells in the plate contained either 0.4 or 0.5 nM C5a(for U937 and human neutrophils, respectively), concentrations ofC5a previously determined to be EC50 to achieve a maximal chemo-taxis response in these assays. An 8-�m pore diameter membranewas placed on top of the wells, and aliquots of cell suspension weretransferred in quadruplicate onto spots on the membrane directlyover each well. The plate was placed at 37°C for 1 h. The tops of theplate membranes were then rinsed with phosphate-buffered salineand gently wiped with cheesecloth to eliminate residual cells that didnot migrate. Fluorescence measurements were made in the platewells with the Fluoroskan Ascent 96-well plate fluorometer (excita-tion 485 nm, emission 530 nm; Thermo Electron Corp, Hartford, CT).The extent of fluorescence present provides an index of the extent ofcellular chemotaxis. The data were plotted as a function of drugconcentration to determine the IC50 using a logistic equation, withthe aid of the KaleidaGraph Program 3.0 from Synergy Software.
C5a-Induced Cell Degranulation Assay. U937 cells were dif-ferentiated as described above for 72 h. Cells were centrifuged and
900 Brodbeck et al.
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

resuspended in HEPES buffer containing 0.5 mM glucose at 107
cells/ml. Cells were then equilibrated for 30 min at 37°C. Approxi-mately 3.5 million cells were added to 1.5 ml tubes containing 3.5 �lof NDT 9513727 (10�11 to 10�6 M final concentrations). Cells with orwithout the presence of drug were incubated at room temperature for30 min. One hundred microliters per well of cells was added intriplicate to a 96-well Falcon clear bottom plate containing 5.0 �g ofcytochalasin B and 5 nM C5a, which was previously determined asthe EC50 dose for �-glucuronidase release. Plates were incubated for30 min at 37°C. After incubation, 100 �l of supernatant was carefullyremoved and transferred to a fresh 96-well clear bottom plate (con-taining 50 �M fluorescein di-�-D-glucuronide) for the determinationof �-glucuronidase activity. Plates were incubated for 16 h at 37°C.Released fluorescein was measured using the Fluoroskan platereader (excitation, 485 nm; emission, 530 nm). The data were plottedas a function of drug concentration to determine the IC50 using alogistic equation as described above.
Reactive Oxygen Species Assay. Differentiated HL-60 cellswere washed several times and resuspended in KRH containing 10�M dihydrorhodamine 123 (Molecular Probes). Aliquots of the cellu-lar suspension were transferred to individual wells in a 96-wellplate. Some wells contained varying concentrations (0.01–100 nM) ofNDT 9513727, and others did not contain drug. Recombinant hC5awas diluted in KRH buffer containing 200 nM phorbol 12-myristate13-acetate and added to each of the wells. The C5a concentration inall wells containing NDT 9513727 was 2.5 nM, the EC50 for C5a inthis assay. Plates were incubated at 37°C for 30 min. Fluorescencewas then measured using the FluorSkan (excitation, 488 nm; emis-sion, 530 nM) and normalized to negative control wells by subtract-ing the background fluorescence observed (no C5a added). Normal-ized data were plotted as a function of drug concentration todetermine the IC50 using a logistic equation, with the aid of theKaleidaGraph Program 3.0 from Synergy Software (Reading, PA).The assay typically resulted in a signal to background noise ofbetween 60 to 90 relative fluorescence units using maximal C5aconcentrations of 100 nM.
Granulocyte CD11b Flow Cytometry Assay. Human bloodwas collected in Vacutainers containing 143 USP units of sodiumheparin anticoagulant (BD Biosciences, San Jose, CA) and processedwithin 4 h of collection. These experiments were performed at roomtemperature to minimize the conversion of C5a to C5a desArg byplasma carboxypeptidase activity. BioHit precision micropipets wereused for accuracy and reproducibility. Using 12 � 75-mm polysty-rene tubes, drug or dimethyl sulfoxide vehicle was added to 100 �l ofwhole blood (1% dimethyl sulfoxide final) for 15 min before theaddition of recombinant human C5a (Sigma-Aldrich). C5a stock so-lution was prepared in PBS containing 0.25 mg/ml bovine serumalbumin (Sigma-Aldrich). For C5a concentration-response analysis,1 to 300 nM C5a was added to stimulate granulocytes for 2 to 5 min.For drug inhibition experiments, 10 nM C5a (approximate EC40) wasadded. Direct immunofluorescent granulocyte labeling was accom-plished with conjugated monoclonal antibodies CD11b-PE-Cy5 andCD15-fluorescein isothiocyanate from BD Biosciences Pharmingen.Twenty microliters of a neat working solution of each antibody wasadded to each sample and incubated for 10 min. Subsequently, thesamples were lysed for 10 min in 2 ml of NH4Cl, pH 7.3, buffer andcentrifuged at 300g for 5 min, washed in 1 ml of 0.1% sodiumazide/PBS buffer, recentrifuged, and fixed in 1% paraformaldehyde/0.1% sodium azide/PBS buffer before flow cytometry analysis. Sam-ples were run on a Beckman Coulter EPICS XL-MCL Flow Cytom-eter (Beckman Coulter, Fullerton, CA) and analyzed using EXPO 32ADC software. The collection protocol was designed to sort cell pop-ulations on the distribution of forward light scatter verses side lightscatter adjusted to detect fluorochrome emission in the range of 680(PE-Cy5) and 525 nm (fluorescein isothiocyanate). Cytometry datacollection was terminated after 5000 CD15-positive cells were de-tected. The data were analyzed by calculating the mean CD11b-PE-Cy5 fluorescence detected on strongly forward and side light scatter
cells (deemed granulocytes). The mean CD11b-PE-Cy5 fluorescencewas calculated from two independent measurements of each hC5aconcentration tested and plotted as a function of hC5a concentrationto determine EC50. For NDT 9513727 data, percentage inhibition ofC5a-stimulated response was calculated from two independent mea-surements of each drug concentration tested and plotted as a func-tion of drug concentration to determine IC50.
Pharmacokinetics of NDT 9513727 in Rats and Monkeys.NDT 9513727 was formulated in PEG 400/water [50/50 (v/v)] for i.v.studies and in 0.5% methylcellulose/0.1% triacetin/water for oralstudies. Drug solution was administered to male Sprague-Dawleyrats or male cynomolgus macaque monkeys by bolus intravenousinjection or oral gavage. Blood samples (0.4 ml each for rat, 1.0 mlfor monkey) were collected in 1.5-ml tubes containing heparin fromrats via an indwelling jugular catheter or from monkeys via thesaphenous or femoral vein into tubes containing potassium EDTA.Blood samples were kept on ice and centrifuged within 30 min ofcollection. The plasma fractions were frozen at �20°C. NDT 9513727was extracted from plasma by protein precipitation with acetonitrileand analyzed by a validated liquid chromatography/tandem massspectrometry method. Pharmacokinetic parameters were deter-mined for each individual animal using the WinNonlin program(Pharsight, Mountain View, CA) and then averaged.
Human C5a-Induced Neutropenia in Gerbils. All experi-ments in Mongolian gerbils were performed in accordance with theapproval of the Animal Care and Use Committees of Wesleyan Uni-versity and Neurogen Corporation and in accordance with the Guidefor the Care and Use of Laboratory Animals (1996). Six-week-oldMongolian gerbils (Charles River Laboratories, Inc., Wilmington,MA) were housed for 2 days before experimentation. The method ofSumichika et al. (2002) was used with a few modifications. In brief,Mongolian gerbils were anesthetized with 60 mg/kg i.m. Telazol(Webster Veterinary Supply, Sterling, MA) and were placed on aheating pad to keep body temperature approximately 37°C through-out the experiment, and a catheter was placed in the jugular vein. Atapproximately 15 min after anesthesia administration, an initial100-�l blood sample was collected via the jugular catheter into asodium EDTA tube. This was defined as sample t �5 min. Simi-larly, a second blood sample was collected 5 min later, t 0 min.hC5a (diluted in 50 �l of saline) was administered immediately afterthe t 0 min blood sample via the jugular catheter. Blood sampleswere collected as described at t 1, 3, 5, and, in C5a dose-responseexperiments, 10 min. Blood samples were kept on ice and analyzedas rapidly as possible on a HemaVet 950 blood analyzer (DrewScientific, Oxford, CT). For experiments with NDT 9513727, drugwas weighed and added to the appropriate volume of 0.5% methyl-cellulose in water and probe sonicated. Triacetin was added toachieve 0.1% final concentration, and the drug suspension wasstirred with a magnetic stir bar overnight. NDT 9513727 was stablein dosing solution up to 1 week after preparation. Food was removedfrom the animal cages for 1 h before drug administration. NDT9513727 or 0.5% methylcellulose/0.1% triacetin control solution wasadministered to gerbils by oral gavage at a volume of 10 ml/kg. Theobserved concentration of neutrophils varied between subjects. NDT9513727 was administered 2 h before hC5a challenge. The neutro-phil concentrations observed in each of the blood samples wereexpressed as a percentage of baseline concentration, which was cal-culated as the average of the t �5 min and t 0 min bloodneutrophil concentrations. Unpaired Student’s t test was used todetermine significance versus vehicle.
Human C5a-Induced Neutropenia in Cynomolgus Macaque.All experiments performed in cynomolgus macaque (Macaca fascicu-laris) were performed at Covance Research Products with the approvalof Covance Research Products Animal Care and Use Committee and incompliance with the Guide for the Care and Use of Laboratory Animals.Animals were housed in groups until the day before the experiment,when they were housed in individual cages in a climate controlledenvironment. Two males and two females weighing 3 to 4 kg were used
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 901
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

in each experiment, except in the 75 mg/kg study (two males, onefemale). Animals were given ad libitum access to water during theexperiments, but no food was provided during the period of 8 h before toapproximately 5 h after drug administration. NDT 9513727 was sus-pended in 0.5% methylcellulose/0.1% triacetin in water and stirredovernight in the dark before administration. Drug solution or vehiclewas administered by nasogastric gavage in a volume of 10 ml/kg bodyweight. Approximately 3 to 3.5 h postdrug administration, animalswere anesthetized with ketamine (0.3 ml of ketamine and 0.2 ml ofxylazine i.m.), and an intravenous catheter was introduced. While an-imals were anesthetized, a constant saline drip was maintained, andbody temperature was kept at 37 °C with a warming pad. Fifteenminutes after intravenous placement, a blood sample was removedfrom the saphenous vein by venipuncture and collected in a tube con-taining sodium EDTA anticoagulant. Subsequent blood samples werecollected in a similar fashion at 30, 31, 35, 40, and 60 min after intra-venous placement. Approximately 4-h postdrug administration (30 minafter intravenous placement), a solution of 30 �g/ml hC5a in sterilesaline, was injected intravenously to achieve a final dose of 10 �g/kg.Blood samples were mixed gently, kept briefly on ice, and analyzedquickly on a HemaVet blood analyzer. Neutrophil levels were analyzedas described above. These experiments were a single crossover, paireddesign such that each subject was tested twice, once after oral NDT9513727 and once after oral vehicle, such that data were analyzed bypaired Student’s t test for statistical significance.
ResultsIdentification of NDT 9513727. The human brain C5a
receptor cloned was identical to the neutrophil receptor re-ported by both Boulay et al. (1991) and Gerard and Gerard(1991). The hC5aR was either expressed by itself or coex-pressed with the G-protein subunits consisting of rat G�i2,human G�1, and bovine G�2. When assessed by saturationbinding using [125I]C5a, membranes from Sf9 cells that wereinfected with hC5aR alone (i.e., in the absence of G-proteins)possessed two marked affinity states of 0.14 and 0.78 nMwith receptor densities of 109 and 287 fmol/mg, respectively.On the other hand, when Sf9 cells were coinfected withhC5aR and G�i2, G�1, and G�2, the two affinity states re-mained the same, but the measurable receptor density was4–5 fold higher (455 versus 1541 fmol/mg, data not shown).The presence of NaCl in the radioligand binding assay bufferwas critical to the identification of the antagonist lead series.The lead optimization strategy, highlighted by Hutchisonand Krause (2004), identified the novel nonpeptide C5aRantagonist NDT 9513727, shown in Fig. 1.
Radioligand Binding Analysis of NDT 9513727 atC5aR. NDT 9513727 displaced [125I]C5a binding with anIC50 value of 11.6 � 1.0 nM, n 31 (Fig. 2). Although NDT9513727 was found to have potent activity in both gerbil andmonkey (cynomolgus macaque), lung membrane prepara-tions (6.4 and 7.3 nM, respectively, Table 1), it displayed
minimal activity in rat, mouse, and dog lung preparations(IC50 values greater than 10 �M, Table 1). Binding of NDT9513727 to hC5aR was, like other reported small-moleculehC5aR antagonists, dependent on W213 in transmembranedomain V (Waters et al., 2005; data not shown).
In Vitro Functional Activity of NDT 9513727. Coex-pression of C5aR with G-proteins in baculovirus-infected Sf9cells was also critical for establishing the GTP�S functionalassay. When the same two batches of membranes(Sf9.hC5aR and Sf9.hC5aR.i2�1�2 membranes) were assayedusing hC5a-stimulated GTP�[35S] binding, the EC50 remainedunchanged, but the maximum agonist-stimulated response wasincreased roughly 6-fold in the Sf9.hC5aR.i2�1�2 membranes(data not shown). All subsequent GTP�[35S] binding experi-ments were performed with membranes obtained from C5aRand G-protein coexpressed baculovirus-infected Sf9 cells.
In the presence of 10 nM hC5a (EC50 18 nM � 1.7, n 4), NDT 9513727 reversed the hC5a agonist effect in a dose-dependent fashion, with an IC50 9.2 � 0.9 nM, n 22 (Fig.3A). Although NDT 9513727 consistently and dose-depen-dently lowered the GTP�[35S] binding signal below the base-line, AcF[OPdChaWr] only decreased the hC5a-stimulatedGTP�[35S] binding response to baseline, with an IC50 Fig. 1. Structure of NDT 9513727.
Fig. 2. NDT 9513727 has affinity for the hC5aR. Both hC5a (f) and NDT9513727 (F) dose-dependently compete for [125I]C5a binding using Sf9cell membranes expressing the recombinant hC5aR along with theG-proteins �i2, �1, and �2. The data are presented as the percentagespecific [125I]hC5a binding as a function of ligand concentration. Theligand dose-responses yield an IC50 0.29 � 0.1 nM for hC5a, with annH 0.6 � 0.03, and 11.6 � 1.0 nM for NDT 9513727, with an nH 0.8 �0.6 nM. The data are representative of mean � S.E.M., n 6 for hC5a,and n 31 for NDT 9513727.
TABLE 1C5a receptor sequence identity across species and competition bindingactivity of NDT 9513727
Species Receptor Identity to Human NDT 9513727* (IC50)
%
Human 100 11.6 nMMacaque 95 7.3 nMCanine 68 �10 �MRat 64 �10 �MMouse 65 �10 �MGerbil 66 6.4 nM
* 125I�hC5a radioligand binding used hC5a as the competing ligand for all tissuepreparations.
902 Brodbeck et al.
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

222.7 � 41.7 nM, n 7. The GTP�[35S] binding assay can berun three different modes: agonist mode, antagonist mode asdescribed above, and inverse agonist mode. In agonist mode,ligands can be assessed for their ability to functionally acti-vate the hC5aR. Screening compounds with the affinity assaydescribed above resulted in the discovery of both nonpep-tide agonists and antagonists as determined by the agonistmode GTP�[35S] binding assay. Compared with hC5a, NDT9517011 demonstrated significantly greater efficacy (maxi-mal efficacy 400% above basal compared with the 100%above basal for hC5a; Fig. 4). Although the superagonistNDT 9517011 was clearly more efficacious than the full pep-tide agonist hC5a, its potency was significantly lower(EC50 2580 versus 18 nM for hC5a; Fig. 4). When thehC5aR was activated by the nonpeptide superagonist NDT9517011, the nonpeptide antagonist NDT 9513727 hadnearly the same functional potency as it had for hC5a func-tion in the GTP�[35S] assay (data not shown). Agonist mode
testing of NDT 9513727 indicated that not only did it notpossess any detectable agonist activity, it dose-dependentlylowered the hC5aR GTP�[35S] binding signal below baseline,indicative of inverse agonism (Fig. 4). To more conclusivelydemonstrate the inverse agonist property of NDT 9513727,the GTP�[35S] binding assay was modified to allow ligandtesting in the absence of hC5a (inverse agonist mode; Fig.3B). Although both NDT 9513727 and AcF[OPdChaWr] dose-dependently reversed the hC5a-stimulated GTP�[35S] bind-ing response (Fig. 3A), only NDT 9513727 dose-dependentlyreduced the constitutive GTP�[35S] activity of hC5aR (IC50 5.7 � 1.6, n 5; Fig. 3B).
To determine whether NDT 9513727 was a competitiveantagonist, multiple hC5a dose responses were run using theagonist mode GTP�[35S] binding assay in the presence ofincreasing concentrations of NDT 9513727 (0, 3, 10, 33, and100 nM, Fig. 5). The data illustrate that at doses of at least100 nM, the effect of NDT 9513727 on hC5aR responses wassurmountable by high doses of hC5a. In addition, a parallelrightward shift in hC5a dose-response curves indicates thatthe NDT 9513727 mechanism of action is competitive innature. A Schild plot of the data from Fig. 5 yielded a slope of1.09, Kd 3.7 nM (r2 0.96), consistent with competitiveantagonism (this experiment was repeated once with similarresults). When AcF[OPdChaWr] was similarly assessed, arightward parallel shift in hC5a dose-response activity wasseen, suggesting that the peptide antagonist also functions asa surmountable, competitive antagonist (data not shown).
Species selectivity is evident for both peptide and nonpep-tide ligands for the C5a receptor. The competition bindingdata presented in Table 1 demonstrates that NDT 9513727has limited, if any, affinity for rC5aR. The fact that hC5a alsohas limited affinity for the rat receptor complicates the eval-uation of NDT 9513727 affinity at the rC5aR (Fig. 6; data not
Fig. 3. NDT 9513727 functions as an inverse agonist. The inverse agonistproperties of NDT 9513727 were demonstrated using two independentassays. A, antagonism of C5a-stimulated GTP�[35S] binding. Sf9 cellmembranes expressing hC5aR along with G�i2, �1, and �2 were stimu-lated with 10 nM hC5a (EC50 18.0 nM � 1.7, n 4) and that GTP�[35S]binding signal was antagonized by either NDT 9513727 (F) or AcF[OP-dChaWr] (f). Although NDT 9513727 dose-dependently reversed theC5a-induced response below the baseline (IC50 9.2 � 0.9 nM, n 18),AcF[OPdChaWr] blocked the GTP�[35S] binding signal only to baseline(IC50 222.3 nM � 41.7, n 7). B, dose-dependent inhibition of hC5aRfunctional activity in the absence of C5a stimulation. The baselineGTP�[35S] binding activity was dose-dependently decreased by NDT9513727 (F) with an IC50 5.7 � 1.6 (n 5), whereas AcF[OPdChaWr](f) had no detectable activity. Representative data examples are shown.
Fig. 4. Intrinsic activity of both peptide and nonpeptide C5aR ligands asmeasured by GTP�[35S] binding. The GTP�[35S] binding assay run in“agonist mode” can detect both increases (agonist) and decreases (inverseagonist) in receptor-mediated nucleotide exchange activity. Sf9 cell mem-branes expressing the hC5aR along with the G-proteins �i2, �1, and �2 aretreated with recombinant hC5a (f), NDT 9513727 (F), or NDT 9517011(E), and data are reported as changes in GTP�[35S] binding relative tobasal activity as a function of ligand dose. Recombinant hC5a and NDT9517011 both function as agonists with EC50 values of 18.0 � 1.7 nM (n 4) and 2578 � 698 nM (n 3), respectively. NDT 9513727 dose-depen-dently decreases the baseline, with an IC50 2.4 � 0.8 nM (n 4). Theexperimental results shown are representative of the several performed.
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 903
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

shown). Sf9 membranes expressing rC5aR along with theG-proteins �i2, �1, and �2 were used in an agonist modeGTP�[35S] binding assay to determine potency and efficacy ofboth rC5a and hC5a. Although rC5a is a potent (8.3 � 1.0nM; n 5) and efficacious peptide at the rat receptor, thehuman peptide has limited activity at the rat receptor (Fig.6A). Based on the aforementioned peptide species selectivity,to directly determine the potency of NDT 9513727 at therC5aR, 10 nM rC5a was used to stimulate GTP�[35S] bindingthat was subsequently challenged with NDT 9513727. NDT9513727 was found to have no significant activity at therC5aR at doses up to 10 �M (Fig. 6B). The hC5L2R displays49% identity with the hC5aR (Cain and Monk, 2002; Okinagaet al., 2003) and binds both hC5a and hC5a desArg with highaffinity (Table 2). No binding activity, however, was observedfor NDT 9513727 at the C5L2R (Table 2).
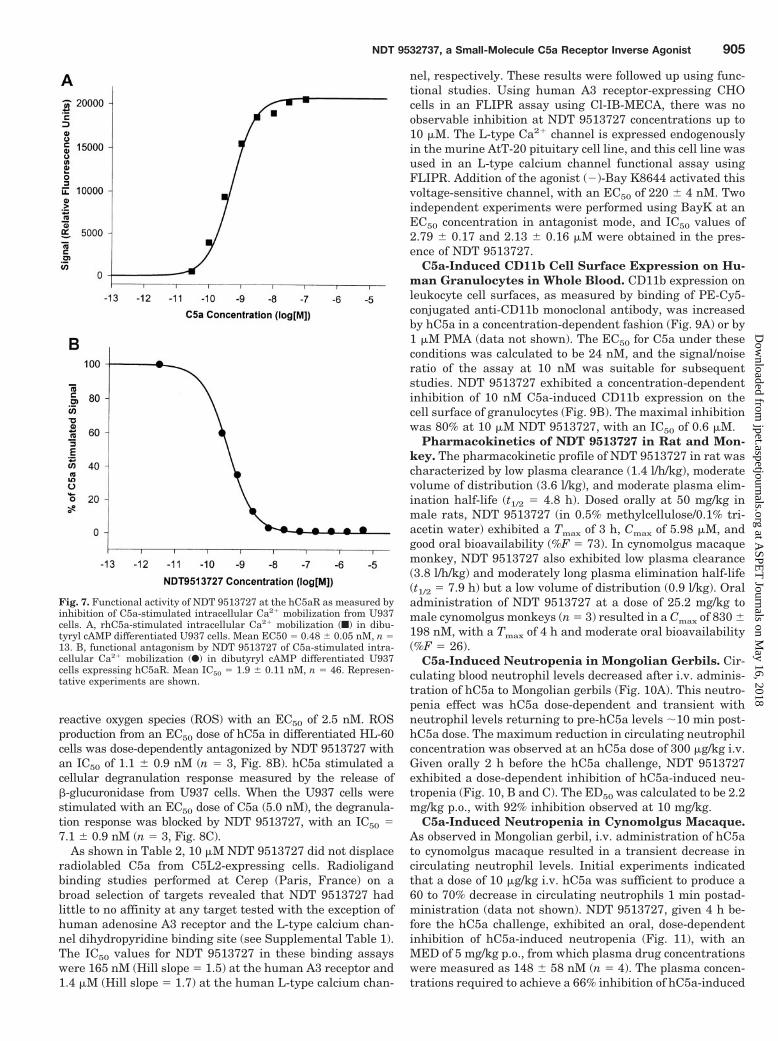
hC5a dose-dependently stimulated increases of intracellu-lar calcium mobilization in dibutyryl cAMP-differentiatedU937 cells, which endogenously express the C5a receptor. Inthis whole-cell assay, hC5a functioned with an EC50 of 0.48 �0.05 nM, n 13 (Fig. 7A). NDT 9513727 dose-dependentlyreversed an EC50 dose of hC5a, with an IC50 1.9 � 0.11 nM,n 46 (Fig. 7B). Similar to the results shown for theGTP�[35S] assay in Fig. 5, the evaluation of NDT 9513727 ina Schild-type analysis in the calcium mobilization assay wasalso consistent with that of a competitive antagonist (datanot shown).
C5a activatesseveral proinflammatory functions of neutro-phils. In these studies, hC5a stimulated a chemotactic responsein both freshly prepared human polymorphonuclear neutro-phils and human U937 cells with an EC50 maximal chemotaxisresponse of 0.5 and 0.4 nM, respectively. As shown in Fig. 8A,NDT 9513727 blocked hC5a-stimulated chemotaxis activity in
human polymorphonuclear neutrophils and U937 cells with anIC50 of 4.7 � 0.8 nM (n 8) and 4.3 � 0.3 nM (n 42),respectively. HL-60 cells were stimulated by hC5a to release
TABLE 2Ligand potencies at hC5a and hC5L2 receptor as measured byradioligand binding with 125I�hC5a
LigandRecombinantly Expressed Receptor*
hC5aR hC5L2R
hC5a** 0.05 � 0.01 nM 0.85 � 0.07 nMhC5a desArg** 14.1 � 1.7 nM 16.2 � 3.0 nMNDT 9513727 11.6 � 5.3 nM �10 �M
* The CHO cell expression system was used for all examples except NDT 9513727activity at the hC5aR, which used baculovirus-expressed C5aR. All values shown areIC50 values.
** Synthesized and purified using His-tag methodologies (see Materials andMethods).
Fig. 5. NDT 9513727 is a competitive antagonist of the hC5aR. Antago-nism by NDT 9513727 (f, 3 nM; E, 10 nM; �, 33 nM; x, 100 nM) ofGTP�[35S] binding stimulated by hC5a (F, control) in membranes fromSf9 cells recombinantly expressing hC5aR and G�i2, �1, and �2. Note thatNDT 9513727 is a surmountable antagonist because the highest concen-trations of NDT 9513727 resulted in responses up to 310 and 340% abovebaseline at the maximal hC5a concentration.
Fig. 6. Species selectivity is evident with both peptide and nonpeptideligands for the C5aR. The GTP�[35S] binding assay run in “agonist mode”was used to assess peptide potency and efficacy at the rC5aR. A, Sf9 cellmembranes expressing recombinant rC5aR along with the G-proteins �i2,�1, and �2 were stimulated with either recombinant hC5a (f) or rC5a-His(F), and data are shown as changes in GTP�[35S] binding relative to basalactivity as a function of ligand concentration. rrC5a-His functions as anagonist at the rC5aR, with an EC50 of 8.3 nM � 1.0 (n 5), but hC5a hasno agonist activity at the rat receptor. B, to directly test the speciesselectivity of NDT 9513727, rC5aR was stimulated with 10 nM rC5a, andthe GTP�[35S] binding signal was challenged with increasing concentra-tions of NDT 9513727. NDT 9513727 had no activity at the rC5aR up to10 �M. Representative experiments are shown.
904 Brodbeck et al.
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from
reactive oxygen species (ROS) with an EC50 of 2.5 nM. ROSproduction from an EC50 dose of hC5a in differentiated HL-60cells was dose-dependently antagonized by NDT 9513727 withan IC50 of 1.1 � 0.9 nM (n 3, Fig. 8B). hC5a stimulated acellular degranulation response measured by the release of�-glucuronidase from U937 cells. When the U937 cells werestimulated with an EC50 dose of C5a (5.0 nM), the degranula-tion response was blocked by NDT 9513727, with an IC50 7.1 � 0.9 nM (n 3, Fig. 8C).
As shown in Table 2, 10 �M NDT 9513727 did not displaceradiolabled C5a from C5L2-expressing cells. Radioligandbinding studies performed at Cerep (Paris, France) on abroad selection of targets revealed that NDT 9513727 hadlittle to no affinity at any target tested with the exception ofhuman adenosine A3 receptor and the L-type calcium chan-nel dihydropyridine binding site (see Supplemental Table 1).The IC50 values for NDT 9513727 in these binding assayswere 165 nM (Hill slope 1.5) at the human A3 receptor and1.4 �M (Hill slope 1.7) at the human L-type calcium chan-
nel, respectively. These results were followed up using func-tional studies. Using human A3 receptor-expressing CHOcells in an FLIPR assay using Cl-IB-MECA, there was noobservable inhibition at NDT 9513727 concentrations up to10 �M. The L-type Ca2� channel is expressed endogenouslyin the murine AtT-20 pituitary cell line, and this cell line wasused in an L-type calcium channel functional assay usingFLIPR. Addition of the agonist (�)-Bay K8644 activated thisvoltage-sensitive channel, with an EC50 of 220 � 4 nM. Twoindependent experiments were performed using BayK at anEC50 concentration in antagonist mode, and IC50 values of2.79 � 0.17 and 2.13 � 0.16 �M were obtained in the pres-ence of NDT 9513727.
C5a-Induced CD11b Cell Surface Expression on Hu-man Granulocytes in Whole Blood. CD11b expression onleukocyte cell surfaces, as measured by binding of PE-Cy5-conjugated anti-CD11b monoclonal antibody, was increasedby hC5a in a concentration-dependent fashion (Fig. 9A) or by1 �M PMA (data not shown). The EC50 for C5a under theseconditions was calculated to be 24 nM, and the signal/noiseratio of the assay at 10 nM was suitable for subsequentstudies. NDT 9513727 exhibited a concentration-dependentinhibition of 10 nM C5a-induced CD11b expression on thecell surface of granulocytes (Fig. 9B). The maximal inhibitionwas 80% at 10 �M NDT 9513727, with an IC50 of 0.6 �M.
Pharmacokinetics of NDT 9513727 in Rat and Mon-key. The pharmacokinetic profile of NDT 9513727 in rat wascharacterized by low plasma clearance (1.4 l/h/kg), moderatevolume of distribution (3.6 l/kg), and moderate plasma elim-ination half-life (t1/2 4.8 h). Dosed orally at 50 mg/kg inmale rats, NDT 9513727 (in 0.5% methylcellulose/0.1% tri-acetin water) exhibited a Tmax of 3 h, Cmax of 5.98 �M, andgood oral bioavailability (%F 73). In cynomolgus macaquemonkey, NDT 9513727 also exhibited low plasma clearance(3.8 l/h/kg) and moderately long plasma elimination half-life(t1/2 7.9 h) but a low volume of distribution (0.9 l/kg). Oraladministration of NDT 9513727 at a dose of 25.2 mg/kg tomale cynomolgus monkeys (n 3) resulted in a Cmax of 830 �198 nM, with a Tmax of 4 h and moderate oral bioavailability(%F 26).
C5a-Induced Neutropenia in Mongolian Gerbils. Cir-culating blood neutrophil levels decreased after i.v. adminis-tration of hC5a to Mongolian gerbils (Fig. 10A). This neutro-penia effect was hC5a dose-dependent and transient withneutrophil levels returning to pre-hC5a levels 10 min post-hC5a dose. The maximum reduction in circulating neutrophilconcentration was observed at an hC5a dose of 300 �g/kg i.v.Given orally 2 h before the hC5a challenge, NDT 9513727exhibited a dose-dependent inhibition of hC5a-induced neu-tropenia (Fig. 10, B and C). The ED50 was calculated to be 2.2mg/kg p.o., with 92% inhibition observed at 10 mg/kg.
C5a-Induced Neutropenia in Cynomolgus Macaque.As observed in Mongolian gerbil, i.v. administration of hC5ato cynomolgus macaque resulted in a transient decrease incirculating neutrophil levels. Initial experiments indicatedthat a dose of 10 �g/kg i.v. hC5a was sufficient to produce a60 to 70% decrease in circulating neutrophils 1 min postad-ministration (data not shown). NDT 9513727, given 4 h be-fore the hC5a challenge, exhibited an oral, dose-dependentinhibition of hC5a-induced neutropenia (Fig. 11), with anMED of 5 mg/kg p.o., from which plasma drug concentrationswere measured as 148 � 58 nM (n 4). The plasma concen-trations required to achieve a 66% inhibition of hC5a-induced
Fig. 7. Functional activity of NDT 9513727 at the hC5aR as measured byinhibition of C5a-stimulated intracellular Ca2� mobilization from U937cells. A, rhC5a-stimulated intracellular Ca2� mobilization (f) in dibu-tyryl cAMP differentiated U937 cells. Mean EC50 0.48 � 0.05 nM, n 13. B, functional antagonism by NDT 9513727 of C5a-stimulated intra-cellular Ca2� mobilization (F) in dibutyryl cAMP differentiated U937cells expressing hC5aR. Mean IC50 1.9 � 0.11 nM, n 46. Represen-tative experiments are shown.
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 905
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

neutropenia (25 mg/kg p.o. dose) were 410 � 218 nM (n 4).The inhibition of neutropenia observed in the 75 mg/kg dosegroup (plasma exposure 832 � 370 nM, n 3) did notreach statistical significance (p 0.088).
DiscussionNDT 9513727 is an orally active, potent, and selective
antagonist of the hC5a receptor. In recombinant or nativelyexpressed hC5aR radioligand binding or functional assaysystems, NDT 9513727 displays subnanomolar to single-digitnanomolar antagonist activity. The in vitro functional assaysranged from a direct assessment of GTP for GDP exchange ata G-protein � subunit coupled to the C5aR, to downstreamreceptor-mediated events including intracellular calcium mo-bilization, and functional cellular responses including de-granulation, oxidative burst, and chemotaxis. The GTP�[35S]
Fig. 8. NDT 9513727 antagonism of oxidative burst, degranulation andchemotactic inflammatory functions in human cells that endogenously ex-press the C5aR. A, NDT 9513727 dose-dependently antagonized hC5a-stim-ulated chemotaxis in both freshly purified human neutrophils (F) and hu-man U937 cells (f). In both cells systems, an EC50 dose of hC5a was used tostimulate a chemotactic response for antagonism studies, 0.5 and 0.4 nM forhuman neutrophils and U937 cells, respectively. NDT 9513727 antagonizedchemotaxis with an IC50 4.7 � 0.8 nM, nH 0.9 � 0.04 (n 8), and IC50 4.3 nM � 0.3, nH 0.9 � 0.01 (n 42) for human neutrophils and U937cells, respectively. B, NDT 9513727 antagonism of C5a-stimulated produc-tion of ROS. An EC50 dose of C5a (2.5 nM) was blocked by NDT 9513727,with an IC50 1.1 � 0.9 nM (n 3). C, NDT 9513727 antagonism ofC5a-stimulated release of �-glucuronidase from U937 cells. An EC50 dose ofC5a (5.0 nM) was blocked by NDT 9513727, with an IC50 7.1 � 0.9 nM(n 3). Representative experiments are shown.
Fig. 9. NDT 9513727 inhibition of hC5a-induced CD11b expression onhuman blood granulocytes. Aliquots of freshly collected human bloodwere exposed to varying concentrations of hC5a (A) or NDT 9513727 plus10 nM hC5a (B). hC5a elicited a concentration-dependent increase inCD11b surface expression on granulocytes as determined by flow cytom-etry analysis. The calculated EC50 was 24.4 � 2.9 nM (S.E.M., n 7).NDT 9513727 inhibited hC5a-induced CD11b expression in a concentra-tion-dependent fashion, with an IC50 of 0.6 �M � 0.2 nM (S.E.M., n 7).Representative experiments are shown.
906 Brodbeck et al.
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

binding assay allowed for the simultaneous discovery ofsmall-molecule agonists and inverse agonists, as shown inFig. 4. A representative small-molecule agonist, 4-(2-{[cyclo-hexyl(phenyl)methyl]amino}-2-oxoethyl)-N-(diphenylmethyl)-2-piperazinecarboxamide, although clearly weak with regardto potency, demonstrates superagonism as compared withhC5a. Importantly, the effect of this superagonist in theGTP�[35S] binding assay was antagonized by NDT 9513727,with virtually identical pharmacology as compared withhC5a.
Studies on the pharmacologic mechanism of action of NDT9513727 indicate that it functions in a manner consistentwith competitive antagonism and is surmountable by hC5a.The fact that NDT 9513727 is an inverse agonist at thehC5aR complicates the further analysis of these data with aSchild regression because the curves do not conform to thestrict requirements of Schild analysis. Examination of datafor two peptidic antagonists, AcF[OPdChaWr] and F[OPd-ChaWr], reveals more easily interpreted profiles; neitherpeptidic antagonist demonstrated any detectable inverseagonism (Fig. 3; data not shown), and Schild analyses clearlyindicated that both peptides were surmountable antagonistswith parallel, right-shifted dose-response curves. Previously,F[OPdChaWr] was assessed in a Schild analysis using gran-ulocytes and umbilical artery cell preparations (Paczkowskiet al., 1999). In these whole-cell functional assays evaluatingmyeloperoxidase release or spasmogenic responses, the pep-tide antagonist appeared insurmountable in nature. It ispossible that differences in the receptor systems used andendpoints assessed in these studies explain the apparentdivergent results.
NDT 9513727 acts as an inverse agonist, in contrast toF[OPdChaWr] (Paczkowski et al., 1999), which functions as aneutral antagonist. Theoretically, inverse agonists may pro-vide therapeutic advantages over neutral antagonists. Forclinical situations in which GPCR mutations cause ailmentsbecause of excessively high constitutive activity, the poten-tial utility of inverse agonists is clear (de Ligt et al., 2000;Seifert and Wenzel-Seifert, 2002). Beyond these orphan in-dications, however, the benefit of inverse agonists over neu-tral antagonists is more speculative. One analysis shows thatinverse agonists predominate in the current world of GPCRmodulators in accord with the theoretical predictions(Kenakin, 2004). One might suspect that the greatest likeli-hood for neutral antagonism might be with peptide antago-nists that would be predicted to interact with the receptor asthe peptide agonist does, thereby merely blocking its effect as
Fig. 10. NDT 9513727 inhibition of hC5a-induced neutropenia in Mon-golian gerbils. A, hC5a induced a dose-dependent, transient neutropeniain Mongolian gerbils: F, saline; f, 30 �g/kg hC5a; Œ, 100 �g/kg hC5a; �,200 �g/kg hC5a; x, 300 �g/kg hC5a. B, NDT 9513727 p.o. dose-depen-dently inhibited 200 �g/kg hC5a-stimulated neutropenia: E, 0.5% meth-ylcellulose/0.1% triacetin; f, 0.3 mg/kg NGD NDT 9513727; �, 1 mg/kgNDT 9513727; �, 3 mg/kg NDT 9513727; X, 10 mg/kg NDT 9513727; F,30 mg/kg NDT 9513727. C, data shown in B are presented as neutropeniarelative to baseline neutrophil levels as a function of oral administrationof 0.5% methylcellulose/0.1% triacetin vehicle (control) or dose of NDT9513727. �, significance from control, with p � 0.05 (unpaired Student’st test).
Fig. 11. NDT 9513727 inhibition of hC5a-induced neutropenia in cyno-molgus macaques. In this paired study, average neutropenia relative tobaseline neutrophil levels is shown as a function of oral dose of 0.5%methylcellulose/0.1% triacetin vehicle (filled bars) or dose of NDT9513727 (open bars). Error bars, S.E.M.; 5 (n 4 subjects), 25 (n 6),and 75 (n 3) mg/kg. �, significance from control with p � 0.05 (pairedStudent’s t test). The 75 mg/kg group achieved p 0.088 versus itscontrol group.
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 907
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

opposed to having intrinsic activity. The data shown in Fig. 3,which compare the relative inverse agonism of NDT 9513727with the peptide antagonist AcF[OPdChaWr], support theabove-stated assumption.
NDT 9513727 displays species selectivity as shown in Ta-ble 1. Although the gerbil receptor is only 66% identical to thehC5aR (Waters et al., 2005), NDT 9513727 maintains highpotency at the gerbil receptor (6.4 versus 11.6 nM at thehC5aR). The rodent receptors (mouse and rat) and the caninereceptor, although also having roughly 65% identity to thehC5aR, demonstrate no detectable affinity. Although speciesselectivity issues are not uncommon with small-moleculeGPCR antagonists, particularly with chemokine and relatedreceptors, the situation is further complicated by comple-ment C5a agonist species selectivity. Both rC5a and hC5ahave comparable binding affinity at the hC5aR (0.05 � 0.01and 0.14 � 0.01 nM, respectively) and in other hC5aR in vitrotests. However, although rC5a has high potency and func-tional activity at the rC5aR, it has at least 3 orders of mag-nitude weaker activity at the hC5aR. NDT 9513727, in addi-tion to having no detectable activity for rat, mouse, or canineC5aR in the competition assay (Table 1), exhibited no detect-able functional GTP�[35S] binding activity at the rC5aRwhen the rC5a was used (Fig. 3).
A two-site model of C5a agonist binding to the C5aR hasbeen strongly supported by experimental data (Chenowethand Hugli, 1980; Siciliano et al., 1994; Chen et al., 1998;Gerber et al., 2001), in which the relatively large C5a 4-helixbundle ligand has a strong ionic interaction with N-terminalreceptor domain site one (residues 21–30), which accounts forabout 50% of the binding energy. Subsequently, the four-helix bundle-N-terminal receptor domain complex positionsthe C5a carboxyl-terminal domain in proximity with inter-helical regions of the receptor for site two binding, resultingin receptor and G-protein activation. The intramolecular siteof NDT 9513727 action on C5aR has not been established.The similar potency of NDT 9513727 on functional responsesand binding supports the notion that the small-moleculeantagonist may prevent receptor activation by interactingeither directly or allosterically with site two of the C5aR.This model may explain the observations that some small-molecule C5a antagonist compounds demonstrated incom-plete displacement in affinity assays and that a solid corre-lation between functional activity and affinity is not observedwith all test compounds identified in the discovery program(data not shown).
NDT 9513727 was also evaluated in a battery of more than50 neurotransmitter and hormone receptor binding assaysand was found to display little to no activity. The mostsignificant was at the A3 adenosine receptor, where an IC50
of 165 nM was observed in a binding assay, but no A3 activitywas observed in a functional calcium mobilization assay atconcentrations up to 10 �M. Functional antagonism of theL-type calcium channel was observed at 2 to 3 �M, which wassome 250 to 2200-fold weaker than the range of functionalC5a antagonist activity (at GTP�S and ROS generation, re-spectively). No binding activity was observed at C5L2 (Table2), a C5aR homolog (originally named gpr77) isolated fromhuman and other species that displays 49% identity with thehC5aR (Cain and Monk, 2002; Okinaga et al., 2003).
Both in vivo and ex vivo approaches were utilized to inves-tigate NDT 9513727 under physiological conditions. Acute
neutropenia has been used to demonstrate the in vivo effi-cacy of C5aR antagonists in rodents (Pellas et al., 1998). Thisapproach takes advantage of the fact that C5aR activationmediates increased neutrophil adhesion via increased cellsurface integrin receptor expression (Crowell and Van Epps,1990), specifically the CD11b/CD18 complex, Mac1. Becauseof the species selectivity of NDT 9513727, gerbil and primatemodels were used to investigate neutropenia in vivo. In ger-bil, NDT 9512737 blocked C5a-stimulated neutropenia afteroral dosing, displaying an ED50 of 2.2 mg/kg and 92% inhi-bition at 10 mg/kg. In cynomolgus macaque, NDT 9513727exhibited 66% inhibition of C5a-stimulated neutropenia at 25mg/kg p.o. Efficacy in animals given 75 mg/kg p.o. NDT9513727 was not substantially better than 25 mg/kg p.o.,which may be attributed to the lower number of subjects inthe higher dose group, lack of dose-proportional increase inplasma exposure, and/or variability in plasma exposurewithin dose groups. In monkey and rat, NDT 9513727 exhib-ited low plasma clearance and low volume of distribution andvariable oral bioavailability. The compound also exhibitsvery high human plasma protein binding (�99%; data notshown). Hence, the lack of dose-effect correlation between thegerbil and monkey models may be related to species differ-ences in plasma protein binding and/or oral bioavailability.
C5a-mediated up-regulation of CD11b on human bloodgranulocytes ex vivo can be directly measured in undilutedhuman blood by flow cytometry. In this test, NDT 9513727displayed concentration-dependent activity, with an IC50 of0.6 �M. In a similar vein, reduced activity of NDT 9513727was observed in the FLIPR assay in the presence of addedhuman serum (data not shown). The modest potency of NDT9513727 in vivo and ex vivo compared with the serum-free invitro whole-cell assays is likely because of the high plasmaprotein binding exhibited by the compound (as noted above),although other factors cannot be completely ruled out. In anycase, the concentration required to achieve approximately50% inhibition is similar in both the cynomolgus macaqueneutropenia and human granulocyte CD11b expressionassays, suggesting that these factors are similar in bothspecies.
NDT 9513727 is a novel, potent, small-molecule C5a recep-tor antagonist that exhibits characteristics of an inverse ag-onist. It is highly selective for the primate and gerbil C5areceptor subtypes and is otherwise selective versus all othertargets against which it has been tested. NDT 9513727 isorally bioavailable, with desirable pharmacokinetics in mul-tiple species, and it effectively inhibits C5a-induced activityin vivo and in human whole blood. NDT 9513727 is a prom-ising experimental therapeutic for the treatment of humaninflammatory disease.
Acknowledgments
We thank Lauren Danner and Aaron Walker for expert cell cul-ture support; Dawn Alderman, Charles Cheng, and Marci Crandallfor functional assay assistance; Dave Matson for helpful discussions;and Dr. Steve Pearson, Dr. Karen Armstrong, Dennis Ybarra, andAmando Elizondo at Covance Research Products for collaboration onthe cynomolgus macaque monkey experiments. We dedicate thismanuscript to Xiao-Shu He (deceased March 18, 2005), who firstsynthesized NDT 9513727. Her contributions made this publicationpossible.
908 Brodbeck et al.
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from

ReferencesArunlakshana O and Schild HO (1959) Some quantitative uses of drug antagonists.
Br J Pharmacol 14:48–58.Boulay F, Mery L, Tardif M, Brouchon L, and Vignais P (1991) Expression cloning of
a receptor for C5a anaphylatoxin on differentiated HL-60 cells. Biochemistry30:2993–2999.
Cain SA and Monk PN (2002) The orphan receptor C5L2 has high affinity bindingsites for complement fragments C5a and C5a des-Arg(74). J Biol Chem 277:7165–7169.
Chen Z, Zhang X, Gonnella NC, Pellas TC, Boyar WC, and Ni F (1998) Residues21–30 within the extracellular N-terminal region of the C5a receptor represent abinding domain for the C5a anaphylatoxin. J Biol Chem 273:10411–10419.
Chenoweth DE and Hugli TE (1980) Human C5a and C5a analogs as probes of theneutrophil C5a receptor. Mol Immunol 17:151–161.
Crowell RE and Van Epps DE (1990) Nonsteroidal antiinflammatory agents inhibitupregulation of CD11b, CD11c, and CD35 in neutrophils stimulated by formyl-methionine-leucine-phenylalanine. Inflammation 14:163–171.
de Ligt RA, Kourounakis AP, and IJzerman AP (2000) Inverse agonism at G protein-coupled receptors: (patho)physiological relevance and implications for drug discov-ery. Br J Pharmacol 130:1–12.
Finch AM, Wong AK, Paczkowski NJ, Wadi SK, Craik DJ, Fairlie DP, and Taylor SM(1999) Low-molecular-weight peptidic and cyclic antagonists of the receptor for thecomplement factor C5a. J Med Chem 42:1965–1974.
Gerard C and Gerard NP (1994) C5A anaphylatoxin and its seven transmembrane-segment receptor. Annu Rev Immunol 12:775–808.
Gerard NP and Gerard C (1991) The chemotactic receptor for human C5a anaphy-latoxin. Nature 349:614–617.
Gerard NP and Gerard C (2002) Complement in allergy and asthma. Curr OpinImmunol 14:705–708.
Gerber BO, Meng EC, Dotsch V, Baranski TJ, and Bourne HR (2001) An activationswitch in the ligand binding pocket of the C5a receptor. J Biol Chem 276:3394–3400.
Grant EP, Picarella D, Burwell T, Delaney T, Croci A, Avitahl N, Humbles AA,Gutierrez-Ramos JC, Briskin M, Gerard C, et al. (2002) Essential role for the C5areceptor in regulating the effector phase of synovial infiltration and joint destruc-tion in experimental arthritis. J Exp Med 196:1461–1471.
Guo RF, Huber-Lang M, Wang X, Sarma V, Padgaonkar V, Craig RA, RiedemannNC, McClintock SD, Hlaing T, Shi MM, et al. (2000) Protective effects of anti-C5ain sepsis-induced thymocyte apoptosis. J Clin Invest 106:1271–1280.
Guo RF and Ward PA (2005) Role of C5a in inflammatory responses. Annu RevImmunol 23:821–852.
Hageman GS, Hancox LS, Taiber AJ, Gehrs KM, Anderson DH, Johnson LV, RadekeMJ, Kavanagh D, Richards A, Atkinson J, et al. (2006) Extended haplotypes in thecomplement factor H (CFH) and CFH-related (CFHR) family of genes protectagainst age-related macular degeneration: characterization, ethnic distributionand evolutionary implications. Ann Med 38:592–604.
Hutchison AJ and Krause JE (2004) The discovery of small molecule C5a antago-nists. Annu Rep Med Chem 39:139–147.
Institute of Laboratory Animal Resources (1996) Guide for the Care and Use ofLaboratory Animals, 7th ed, Institute of Laboratory Animal Resources, Commis-sion on Life Sciences, National Research Council, Washington, DC.
Ji H, Ohmura K, Mahmood U, Lee DM, Hofhuis FM, Boackle SA, Takahashi K,Holers VM, Walport M, Gerard C, et al. (2002) Arthritis critically dependent oninnate immune system players. Immunity 16:157–168.
Kenakin T (2004) Efficacy as a vector: the relative prevalence and paucity of inverseagonism. Mol Pharmacol 65:2–11.
Kohl J (2006) Drug evaluation: the C5a receptor antagonist PMX-53. Curr Opin MolTher 8:529–538.
Luke GP, Maynard G, Mitchell S, Thurkauf A, Xie L, Zhang L, Zhang S, Zhao H,Chenard BL, Gao Y, et al. (2003) inventors; Neurogen Corporation, assignee. Newaryl imidazoles and related compounds as c5a receptor modulators. World Patentnumber WO/2003/082829. 2003 Sept 10.
Makrides SC (1998) Therapeutic inhibition of the complement system. PharmacolRev 50:59–87.
Nozaki M, Raisler BJ, Sakurai E, Sarma JV, Barnum SR, Lambris JD, Chen Y,Zhang K, Ambati BK, Baffi JZ, et al. (2006) Drusen complement components C3aand C5a promote choroidal neovascularization. Proc Natl Acad Sci U S A 103:2328–2333.
Okinaga S, Slattery D, Humbles A, Zsengeller Z, Morteau O, Kinrade MB, BrodbeckRM, Krause JE, Choe HR, Gerard NP, et al. (2003) C5L2, a nonsignaling C5Abinding protein. Biochemistry 42:9406–9415.
Paczkowski NJ, Finch AM, Whitmore JB, Short AJ, Wong AK, Monk PN, Cain SA,Fairlie DP, and Taylor SM (1999) Pharmacological characterization of antagonistsof the C5a receptor. Br J Pharmacol 128:1461–1466.
Pellas TC, Boyar W, van Oostrum J, Wasvary J, Fryer LR, Pastor G, Sills M, Braun-walder A, Yarwood DR, Kramer R, et al. (1998) Novel C5a receptor antagonistsregulate neutrophil functions in vitro and in vivo. J Immunol 160:5616–5621.
Schieferdecker HL, Schlaf G, Jungermann K, and Gotze O (2001) Functions ofanaphylatoxin C5a in rat liver: direct and indirect actions on nonparenchymal andparenchymal cells. Int Immunopharmacol 1:469–481.
Seifert R and Wenzel-Seifert K (2002) Constitutive activity of G-protein-coupledreceptors: cause of disease and common property of wild-type receptors. NaunynSchmiedebergs Arch Pharmacol 366:381–416.
Siciliano SJ, Rollins TE, DeMartino J, Konteatis Z, Malkowitz L, Van Riper G, BondyS, Rosen H, and Springer MS (1994) Two-site binding of C5a by its receptor: analternative binding paradigm for G protein-coupled receptors. Proc Natl Acad SciU S A 91:14446–14450.
Solomon S, Kassahn D, and Illges H (2005) The role of complement and the Fcgamma R system in the pathogenesis of arthritis. Arthritis Res Ther 7:129–135.
Sumichika H, Sakata K, Sato N, Takeshita S, Ishibuchi S, Nakamura M, KamahoriT, Ehara S, Itoh K, Ohtsuka T, et al. (2002) Identification of a potent and orallyactive non-peptide C5a receptor antagonist. J Biol Chem 277:49403–49407.
Thurkauf A, Zhang X, He X-s, Zhao H, Peterson J, Maynard G, and Ohliger R (2002)inventors; Neurogen Corporation, assignee. High affinity small molecule c5a re-ceptor modulators. World Patent number WO/2002/049993. 2002 Jun 27.
Wang Y, Rollins SA, Madri JA, and Matis LA (1995) Anti-C5 monoclonal antibodytherapy prevents collagen-induced arthritis and ameliorates established disease.Proc Natl Acad Sci U S A 92:8955–8959.
Waters SM, Brodbeck RM, Steflik J, Yu J, Baltazar C, Peck AE, Severance D, ZhangLY, Currie K, Chenard BL, et al. (2005) Molecular characterization of the gerbilC5a receptor and identification of a transmembrane domain V amino acid that iscrucial for small molecule antagonist interaction. J Biol Chem 280:40617–40623.
Weiland T and Jacobs KH (1994) Measurement of receptor-stimulated guanosine5�-O-(gamma-thio)triphosphate binding by G proteins. Methods Enzymol 237:3–13.
Zwirner J, Fayyazi A, and Gotze O (1999) Expression of the anaphylatoxin C5areceptor in non-myeloid cells. Mol Immunol 36:877–884.
Address correspondence to: Dr. James E. Krause, ARMGO Pharma, Inc.,3960 Broadway, New York, NY 10032. E-mail: [email protected]
NDT 9532737, a Small-Molecule C5a Receptor Inverse Agonist 909
at ASPE
T Journals on M
ay 16, 2018jpet.aspetjournals.org
Dow
nloaded from


















