
Histological investigations on the Tyrolean Ice Man
12
Histological Investigations on the Tyrolean Ice Man M.W. HESS, 1 * G. KLIMA, 1 K. PFALLER, 1 K.H. KU ¨ NZEL, 2 AND O. GABER 2 1 Institute of Histology and Embryology, University of Innsbruck, A-6010 Innsbruck, Austria 2 Institute of Anatomy, University of Innsbruck, A-6010 Innsbruck, Austria KEY WORDS neolithic glacier mummy; specimen preparation; ultrastructure; cryopreservation ABSTRACT The 5,200-year-old Tyrolean Ice Man discovered in 1991 in the O ¨ tztal Alps is the world’s most ancient known human glacier mummy. Histological investigation was aimed at 1) optimizing specimen preparation and 2) documenting the preservation state of (sub)cellular components. Minute pieces of frozen tissue were removed endoscopically from rib bone and cartilage, major blood vessels, oral cavity and alimentary tract, liver, spleen, diaphragm, respiratory system, femoral muscle and nerve, sympathetic trunk, brain, and skin. Double fixation with glutaraldehyde followed by osmium tetroxide and embedding in Epon/Araldite epoxy resins proved to be the method of choice for both light and transmission electron microscopy combined with classical histochemistry. In particular, mild evacuation of the desiccated tissue was determined to be essential to ensure homogeneous infiltration with fixatives and resins; as a result, sections of excellent quality could be obtained with any kind of sample. With regard to the preservation degree of (sub)cellular components, distinct tissue-specific patterns were observed. There were highly intact skeletal and connective tissues proper, however, most interestingly, there were remarkably intact nervous tissue components as well. By contrast, epithelial, muscle, and reticular connective tissues as well as blood had generally disintegrated due to autolysis, freeze/thaw damage, and adipocere formation. For a tentative interpretation of these patterns, we considered general aspects of cryopreservation, such as physicochemical properties of subcellular constituents and tissue physiology. Am J Phys Anthropol 106:521–532, 1998. r 1998 Wiley-Liss, Inc. In 1991, an exceptionally well-preserved, prehistoric male corpse was discovered in a glacial field in the O ¨ tztal Alps at 3,200 meters above sea level near the Italian- Austrian border. For details on the recovery, see Seidler et al. (1992). The frozen body proved to be almost completely mummified, presumably as a result of processes akin to freeze drying (Ambach et al., 1992). Based on accelerator mass spectrometry and con- ventional archaeological dating, its absolute age was estimated to be approximately 5,200 years, i.e., an individual from the Late Neo- lithic Age (Bonani et al., 1992). Hence, the Tyrolean Ice Man, also known as the ‘‘Man from Hauslabjoch,’’ ‘‘Similaun Man’’, or ‘‘O ¨ tzi,’’ is the most ancient human glacier mummy discovered to date. The aims of the histological investigations on the Tyrolean Ice Man were 1) to optimize specimen preparation according to the tis- sues’ specificities, and 2) to document the preservation state of (sub)cellular compo- Dedicated to Prof. Werner Platzer (Head and Chairman of the Institute of Anatomy, Medical Faculty, University of Innsbruck) on the occasion of his retirement. *Correspondence to: M.W. Hess, Institute of Histology and Embryology, University of Innsbruck, Mu ¨ llerstrasse 59, A-6010 Innsbruck, Austria. E-mail: [email protected] Received 11 July 1997; accepted 27 April 1998. AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 106:521–532 (1998) r 1998 WILEY-LISS, INC.
Transcript of Histological investigations on the Tyrolean Ice Man

Histological Investigations on the Tyrolean Ice ManM.W. HESS,1* G. KLIMA,1 K. PFALLER,1 K.H. KUNZEL,2AND O. GABER2
1Institute of Histology and Embryology, University of Innsbruck,A-6010 Innsbruck, Austria2Institute of Anatomy, University of Innsbruck, A-6010 Innsbruck, Austria
KEY WORDS neolithic glacier mummy; specimen preparation;ultrastructure; cryopreservation
ABSTRACT The 5,200-year-old Tyrolean Ice Man discovered in 1991 inthe Otztal Alps is the world’s most ancient known human glacier mummy.Histological investigation was aimed at 1) optimizing specimen preparationand 2) documenting the preservation state of (sub)cellular components.Minute pieces of frozen tissue were removed endoscopically from rib bone andcartilage, major blood vessels, oral cavity and alimentary tract, liver, spleen,diaphragm, respiratory system, femoral muscle and nerve, sympathetictrunk, brain, and skin. Double fixation with glutaraldehyde followed byosmium tetroxide and embedding in Epon/Araldite epoxy resins proved to bethe method of choice for both light and transmission electron microscopycombined with classical histochemistry. In particular, mild evacuation of thedesiccated tissue was determined to be essential to ensure homogeneousinfiltration with fixatives and resins; as a result, sections of excellent qualitycould be obtained with any kind of sample. With regard to the preservationdegree of (sub)cellular components, distinct tissue-specific patterns wereobserved. There were highly intact skeletal and connective tissues proper,however, most interestingly, there were remarkably intact nervous tissuecomponents as well. By contrast, epithelial, muscle, and reticular connectivetissues as well as blood had generally disintegrated due to autolysis,freeze/thaw damage, and adipocere formation. For a tentative interpretationof these patterns, we considered general aspects of cryopreservation, such asphysicochemical properties of subcellular constituents and tissue physiology.Am J Phys Anthropol 106:521–532, 1998. r 1998 Wiley-Liss, Inc.
In 1991, an exceptionally well-preserved,prehistoric male corpse was discovered in aglacial field in the Otztal Alps at 3,200meters above sea level near the Italian-Austrian border. For details on the recovery,see Seidler et al. (1992). The frozen bodyproved to be almost completely mummified,presumably as a result of processes akin tofreeze drying (Ambach et al., 1992). Basedon accelerator mass spectrometry and con-ventional archaeological dating, its absoluteage was estimated to be approximately 5,200years, i.e., an individual from the Late Neo-lithic Age (Bonani et al., 1992). Hence, theTyrolean Ice Man, also known as the ‘‘Man
from Hauslabjoch,’’ ‘‘Similaun Man’’, or‘‘Otzi,’’ is the most ancient human glaciermummy discovered to date.
The aims of the histological investigationson the Tyrolean Ice Man were 1) to optimizespecimen preparation according to the tis-sues’ specificities, and 2) to document thepreservation state of (sub)cellular compo-
Dedicated to Prof. Werner Platzer (Head and Chairman of theInstitute of Anatomy, Medical Faculty, University of Innsbruck)on the occasion of his retirement.
*Correspondence to: M.W. Hess, Institute of Histology andEmbryology, University of Innsbruck, Mullerstrasse 59, A-6010Innsbruck, Austria. E-mail: [email protected]
Received 11 July 1997; accepted 27 April 1998.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 106:521–532 (1998)
r 1998 WILEY-LISS, INC.

nents. We employed chemical fixation fol-lowed by resin embedding or cryomicrotomyand classical histochemistry for light micros-copy (LM) and transmission electron micros-copy (TEM). When our data are comparedwith the literature on other mummies, thevery particular conditions of preservation ineach case must be considered. For instance,permafrost mummies, such as 15th centuryGreenland mummies (Hart-Hansen et al.,1991) and pre-Columbian Andean mummiesfrom El Plomo or Nevado Amapato (Rein-hard, 1996), may compare well with the IceMan (for review, see Zimmermann, 1996).By contrast, there is less congruence withfinds from the deserts of Taklimakan orNorthern Chile or with bog bodies fromNorthern Europe (for review, see Spindler etal., 1996). In these cases, decomposition wasimpeded by exposure to salt, dry sand, ortannic acids, respectively; analogous mecha-nisms of preservation and, thus, restrictedcomparability hold true for artificially em-balmed mummies.
In the present contribution, we show rep-resentative data on a broad range of tissuesamples obtained from the Tyrolean Ice Man.The distinct preservation patterns that wereobserved proved to be highly specific. Wetherefore attempt to interpret some of themby considering general aspects of cryopreser-vation (e.g., physicochemical properties ofsubcellular constituents and tissue physiol-ogy).
Results of this study were presented inpart in abstract form (Hess et al., 1996a,b).
MATERIALS AND METHODSSampling
Minute quantities of tissue were removedendoscopically under sterile conditions byemploying titanium tools, as described indetail by Gaber et al. (1995), Gunkel et al.(1997), and Platzer et al. (1998). The totalweight of samples taken for biological andmedical analyses was approximately 10 g,and the mummy’s weight was 13.29 kg(Kunzel, unpublished observations).
According to topographic and macroscopiccriteria, samples for histological investiga-tion were obtained from rib bone and carti-lage, vena cava, thoracic aorta, oral cavity,stomach, small and large intestine, liver,
spleen, diaphragm, nasal cavity, larynx, lung[see also the analytical electron microscopicwork by Pabst and Hofer (1998) on the IceMan’s lung deposits], femoral muscle, femo-ral nerve, sympathetic trunk, cerebral cor-tex, and skin (in total, more than 50 tissuesamples). Control samples were obtainedfrom a present-day human glacier corpse(i.e., a mountain climber who had been lostin the 1920s in the Otztal Alps and releasedfrom the glacier approximately 70 yearslater).
Fixation
Immediately after sampling, the more orless frozen tissue pieces were dissected intosmall cubes 1–4 mm side length. Subse-quently, the specimens were immersed incold fixative (4% weight/volume formalde-hyde and/or 2.5% volume/volume glutaralde-hyde in 0.2 M sodium cacodylate buffer, pH7.2, at 4°C) and kept for approximately 90seconds under mild vacuum to remove airfrom the leather-like tissue (evacuation byhand within a syringe). This procedure iscommon for botanical specimens, facilitat-ing homogeneous infiltration with fixativesand embedding media (Hayat, 1989). Afteraldehyde fixation (12–24 hours at 4–25°C),the specimens were optionally postfixed with1% (weight/volume) aqueous osmium-tetrox-ide (OsO4; 8 hours at 4°C) followed by etha-nol and/or acetone dehydration.
Embedding and microtomy
Pilot experiments with paraffin embed-ding produced unsatisfactory results in termsof sectioning properties and identification ofstructural details as well as poor or uncer-tain reliability of staining specificity; thus,we generally used resin embedding. Speci-mens were infiltrated with resin monomersat room temperature for 3 days with con-tinual motion. Epoxy or acrylic resins wereheat cured (Epon, Araldite, or Unicryl: Brit-ish Biocell International, Cardiff, UnitedKingdom; LR white: London Resin Com-pany, London, United Kingdom). Resinblocks were trimmed with a motor-drivenabrasive wheel (Reichert Ultratrim, Leica;Vienna, Austria). Semithin sections (0.2–0.5µm) or ultrathin sections (80–120 nm) werecut from block faces of 4 mm or 2 mm side
522 M.W. HESS ET AL.

length, respectively, by using an ultramicro-tome (Reichert Ultracut-S, Leica) equippedwith diamond knives (Diatome, Biel, Swit-zerland) at a cutting speed of 0.4–1.0 mmper second (note: cross links between mum-mified tissues and the embedding resins areonly weak; to prevent tissue componentsfrom breaking off the resin, it proved essen-tial to avoid trimming by hand with razorblades or cutting sections thicker than 0.5µm). Semithin sections were transferred witha wire loop onto glass slides; in the case offragile embeddings, sections were allowed toattach onto the glass slide merely by dryingat room temperature followed by heating theglass to approximately 90°C. Hence, thefragmentation and/or floating away of deli-cate sections prior to or during staining wascompletely avoided. Ultrathin sections werecollected onto uncoated or Formvar carbon-coated copper or gold 200-mesh thin bargrids.
Cryomicrotomy
0.5–2.5 µm semithin cryosections (Toku-yasu, 1973) were cut with glass knives atfrom -70°C to -90°C by using a ReichertUltracut-S ultramicrotome equipped with aReichert FC-S cryochamber.
Staining and microscopy
Semithin sections. Staining was per-formed as follows (see also, Pearse, 1980):1) Toluidine blue; 2) Toluidine blue followedby acid fuchsin (see pp. 326–327 in Hayat,1989); 3) Victoria blue B for elastic material(see Snodgress et al., 1972) applied to deplas-tinized epoxy sections (Hayat, 1989); 4) enzy-mic digestion of elastin with elastase typeIII (Montes et al., 1985) applied to deplas-tinized and optionally oxidized epoxy sec-tions followed by general or elastin-specificstaining; 5) Coomassie brilliant blue forproteins (Fisher, 1968) applied to epoxy sec-tions or cryosections; 6) Sudan black B forlipid localization on cryosections; and 7) 0.1µg/ml diamidino-2-phenylindole (DAPI;Manzini et al., 1983) for DNA localization oncryosections. The sections were viewed witha Photomikroskop III or a Axiophot (Zeiss,Oberkochen, Germany).
Ultrathin sections. The following proce-dures were used for processing of ultrathinsections: 1) overall staining with 0.5%weight/volume aqueous uranyl acetate andalkaline lead citrate (Reynolds, 1963); 2)periodic acid-thiocarbohydrazide-silver pro-teinate stained polysaccharides with 1–2glycol groups (Thiery, 1967; Courtoy andSimar, 1974; Erdos, 1986), as described pre-viously (Hess and Hesse, 1994); 3) thiocarbo-hydrazide-silver proteinate staining, whichindicated unsaturated lipids (Rowley andDahl, 1977), as described previously (Hessand Hesse, 1994). Sections were examinedwith an electron microscope at 60, 80, or 100kV (EM 10A, Zeiss; CM120 Philips, Eind-hoven, Netherlands).
Note that, for reasons of brevity and leg-ibility, we usually used the term ‘‘cells’’instead of ‘‘remnants/residues/remains ofcells’’ in the following text.
RESULTS AND DISCUSSIONGeneral methodological remark
For conservational reasons, the removalof tissue samples from this unique corpsewas limited to the absolutely indispensableminimum. This curatorial decision was sup-ported by the very first histological analysisof the digestive tract and liver, in which poorpreservation was observed. Considering thelack of preservation of visceral tissues, to-gether with the minute size of the speci-mens, our approach was not able to revealany possible pathological changes.
Technical notes
Double fixation with aldehydes and os-mium-tetroxide followed by Epon/Aralditeembedding proved to be the preparationmethod of choice for LM and TEM analysis.Excellent, reproducible results were ob-tained with any kind of tissue. Special proce-dures to soften desiccated tissue, as intro-duced by Ruffer (1921) or Sandison (1955),for instance, were not necessary. However,sufficient evacuation of samples prior tofixation combined with prolonged resin infil-tration was found to be essential in order toobtain homogeneous blocks and, in turn,coherent sections. No further improvementwas achieved by embedding in acrylic resins
523HISTOLOGY OF THE TYROLEAN ICE MAN

instead of Epon/Araldite, which is consis-tent with a remark by Williams et al. (1995)on skin samples from the Ice Man. Spurr’slow-viscosity epoxy resin mixture (Spurr,1969) was not tested because of its potentialextractive properties (see Hess, 1990).
Due to the specimens’ properties, folds insemithin and ultrathin sections were inher-ent in all preparations.
Histological observations
Due to the mummy’s low actual weight(approximately 13 kg), as expected, the softtissues displayed signs of considerableshrinkage, e.g., extremely dense arrange-ment of collagen fibrils. A certain rehydra-tion of the mummified tissues, however, wasachieved by exposing the specimens over-night to aqueous fixatives.
Concerning the preservation state of (sub)-cellular constituents, it has to be empha-sized that the patterns observed throughoutall samples analyzed are highly regular.Furthermore, the degree of preservation ap-pears to be specific for the various types oftissues/cells/subcellular constituents.
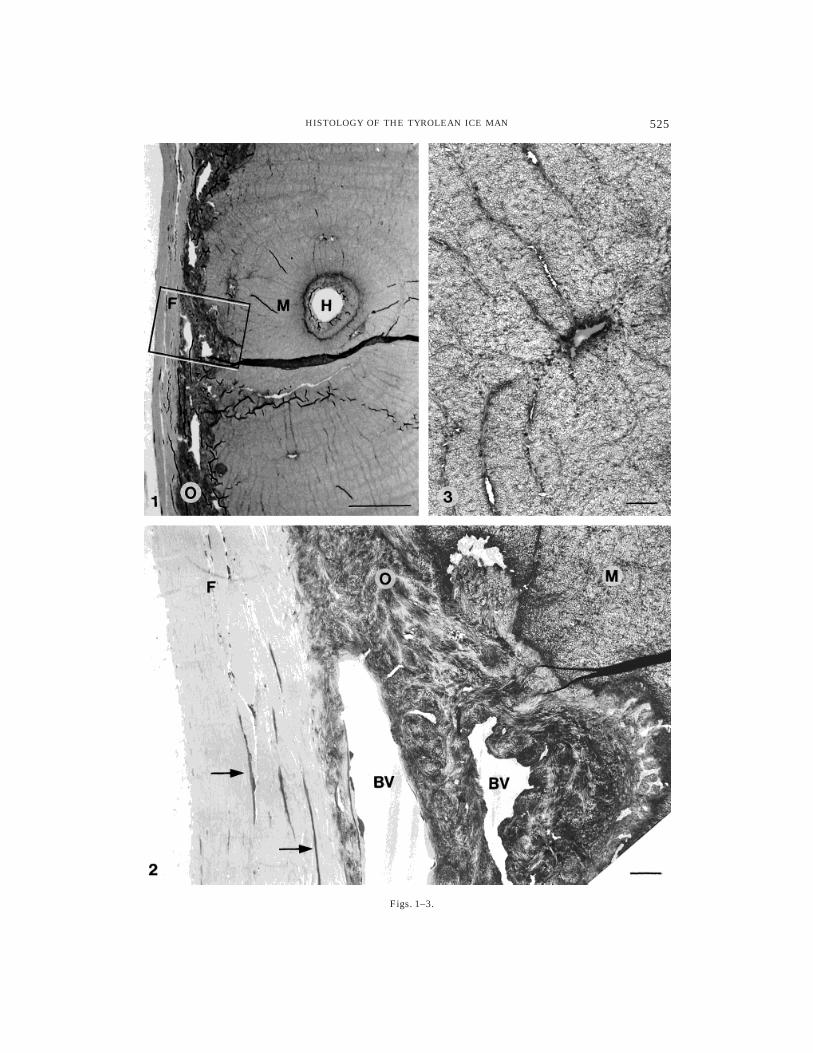
Skeletal and connective tissues proper(e.g., compact bone, hyaline cartilage,elastic arteries, large veins). The grossarchitecture of rib bone (Figs. 1, 2) andcartilage as well as of major blood vessels,such as thoracic aorta (Fig. 7) or vena cava,is easily recognized (for details, see figurelegends; note: reticular connective tissuesare discussed below).
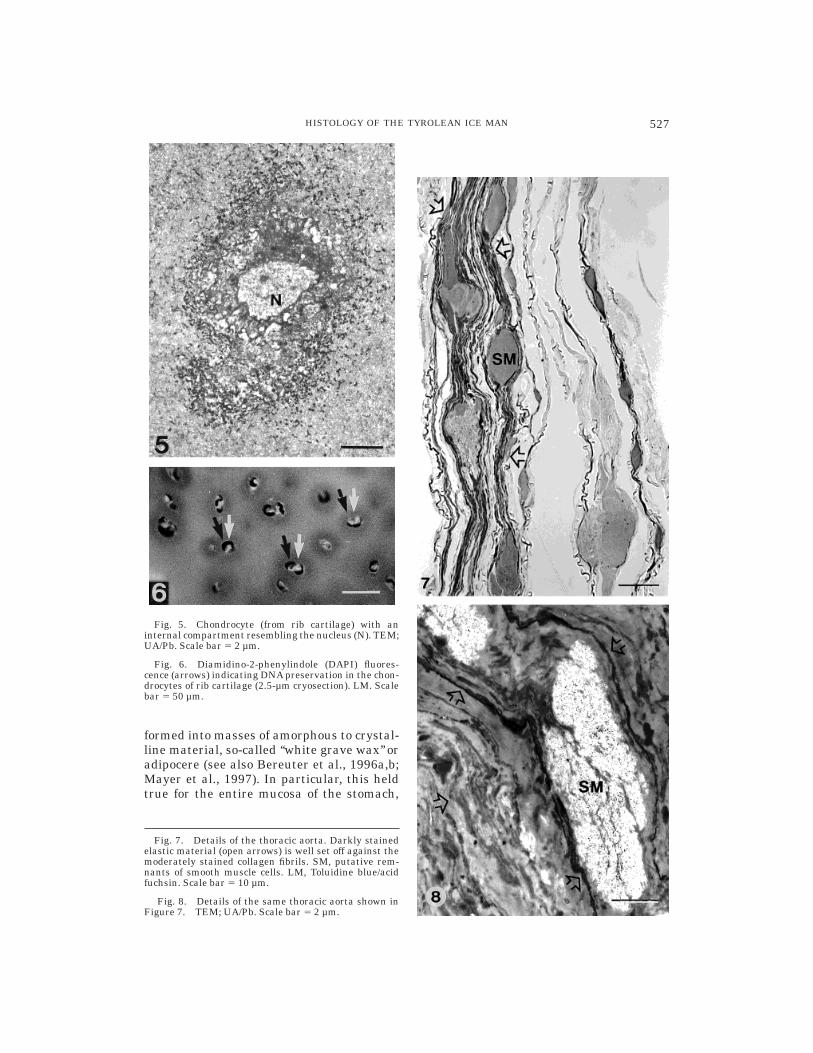
Extracellular components, in particularcollagen and elastin, are well preserved andcould always be identified. Collagen fibrilsstill display specific ultrastructural fea-tures, e.g., approximately 64/67 nm periodic-ity (Fig. 4), and birefringence under polar-ized light. Elastic material appearedcompletely amorphous, and no peripheralmicrofibrils were seen (Fig. 8). Its chemicalnature was assessed histochemically. Com-monly used dyes, such as orcein or resorcin-fuchsin, had proven to be nonspecific in pilotexperiments with paraffin and deplastinizedresin sections, a feature already known fromother ancient tissues (Sandison, 1963). Victo-ria blue B as well as sequential stainingwith Toluidine blue and acid fuchsin (Hayat,
1989) gave reliable results, as verified byelastase digestion according to Montes et al.(1985). Hayat’s recipe proved to be particu-larly apt for making a clear distinction be-tween elastic material and collagen on black-and-white photomicrographs (Fig. 7).
With regard to the cells of these connec-tive tissues, we observed generally well-defined, spindle-shaped structures withamorphous contents, presumably fibrocytes,located between densely packed collagenfibrils (Figs. 2, 4); thus, similar elongatestructures occurring within the perineu-rium of the femoral nerve likely representperineural cells. Osteocytes are no longervisible, except for their outlines (Fig. 3).Chondrocytes, which were identified clearlyin rib cartilage, consistently showed distinctsubcellular compartments resembling nu-clei (Fig. 5). However, the internal organiza-tion of cytoplasm and nucleoplasm had notbeen preserved. Moderate, although dis-tinct, DAPI fluorescence observed withinthe chondrocytes strongly indicated DNA,possibly nuclear DNA. (Fig. 6; note thatHandt et al., 1994, studied material fromthe same rib sample.)
Nervous tissue (e.g., peripheral nerves,vegetative ganglia, brain). Tissuesamples obtained from the Ice Man’s femo-ral nerve and sympathetic trunk regularlydisplayed the concentric membrane lamel-lae of myelin sheaths (Figs. 9, 11, 12) embed-ded within collagenous endoneurium andsurrounded by flattened perineural cells.Myelinated nerve fibers of the diaphragm,skin (Fig. 10), and cerebral cortex also provedto be preserved. Note, however, that some
Fig. 1. Cross section through an osteon from rib bonewith adjacent periosteum. F, stratum fibrosum of perios-teum; H, Haversian canal; M, mineralized bone matrix;O, stratum osteogenicum of periosteum. The windowframes the corresponding area shown in Figure 2.
Light microscopy (LM), Toluidine blue staining (Tol).Scale bar 5 25 µm.
Fig. 2. Rib bone periosteum: fibrocytes (arrows)within the stratum fibrosum and blood vessels (BV)within the stratum osteogenicum can be seen. Transmis-sion electron microscopy (TEM), uranyl acetate and leadstaining (UA/Pb). Scale bar 5 2.5 µm.
Fig. 3. Rib bone showing a ghost of an osteocyte(tangential section) with clearly visible canaliculi. TEM;(UA/Pb). Scale bar 5 2.5 µm.
524 M.W. HESS ET AL.
Figs. 1–3.
525HISTOLOGY OF THE TYROLEAN ICE MAN

nerve fiber remnants showed completelydisordered ultrastructure. We assume thatrepeated freezing and thawing of the IceMan during the preceding 5,000 years (in-cluding the recovery and subsequent storageof the corpse at -6°C) accounted for localstructural breakdown of myelin sheaths andrearrangement (‘‘recrystallization’’) of my-elin constituents (for details on the latterphenomenon, see, e.g., Erk et al., 1996).Patterns that resembled somewhat the nodesof Ranvier were observed occasionally,Schmidt-Lanterman clefts could not be iden-tified at all. Stains for lipid and protein(Sudan black and Coomassie blue, respec-tively) applied to semithin cryosectionsand/or resin sections reacted positively withthe myelin remains (see, e.g., Fig. 13); at theelectron microscopic level, unsaturated lip-ids and periodic acid-Schiff-reactive carbohy-drates could be localized (see, e.g., Fig. 12).These photomicrographs show for the firsttime, to our knowledge, subcellular compo-nents of nervous tissue preserved over mil-lennia in a naturally mummified human(see Doran et al., 1986; Gerszten and Mar-tinez, 1995). Photomicrographs showing rec-ognizable structural patterns of mummified
myelin have only been published from mod-ern (approximately 50-year-old) samples(Radanov et al., 1992); Aichel (1927) brieflymentioned recognizable myelin sheaths inan approximately 1,500-year-old bog corpsebut did not provide drawings or photo-graphs; Gouzhang et al. (1979) publishedmicrographs from an approximately 2,000-year-old female corpse showing remains ofperipheral nerve fibers, but stated that ‘‘. . .details of their submicroscopic structurescould not be distinguished.’’ The nerve cellbodies observed within a ganglion from thesympathetic trunk (Fig. 11) or the cerebralcortex no longer showed internal structures,nor did the residues of axoplasm (Fig. 10) orSchwann cells’ cytoplasm that were occasion-ally identified as well.
Epithelial tissue (e.g., in digestive andrespiratory tract; glands, endothelia),muscle tissue (e.g., smooth muscle ofblood vessels and digestive tract, skel-etal muscle), reticular connective tissue(e.g., spleen, bone marrow), and blood.Epithelial and reticular connective tissuesas well as blood cells generally suffered fromconspicuous disintegration. They were trans-
Fig. 4. Collagen fibres with typical periodicity of approx. 64 nm (stratum fibrosum of rib boneperiosteum). The arrow indicates a fibrocyte. TEM; UA/Pb. Scale bar 5 0.2 µm.
526 M.W. HESS ET AL.
formed into masses of amorphous to crystal-line material, so-called ‘‘white grave wax’’ oradipocere (see also Bereuter et al., 1996a,b;Mayer et al., 1997). In particular, this heldtrue for the entire mucosa of the stomach,
Fig. 7. Details of the thoracic aorta. Darkly stainedelastic material (open arrows) is well set off against themoderately stained collagen fibrils. SM, putative rem-nants of smooth muscle cells. LM, Toluidine blue/acidfuchsin. Scale bar 5 10 µm.
Fig. 8. Details of the same thoracic aorta shown inFigure 7. TEM; UA/Pb. Scale bar 5 2 µm.
Fig. 5. Chondrocyte (from rib cartilage) with aninternal compartment resembling the nucleus (N). TEM;UA/Pb. Scale bar 5 2 µm.
Fig. 6. Diamidino-2-phenylindole (DAPI) fluores-cence (arrows) indicating DNA preservation in the chon-drocytes of rib cartilage (2.5-µm cryosection). LM. Scalebar 5 50 µm.
527HISTOLOGY OF THE TYROLEAN ICE MAN
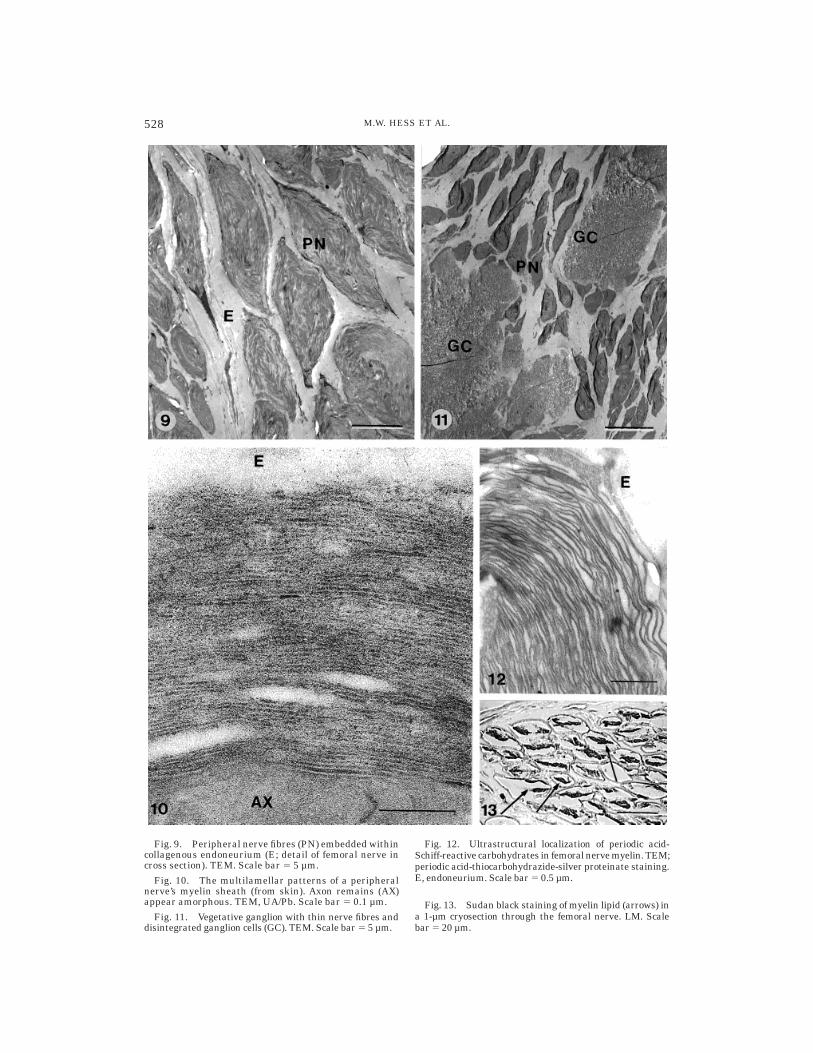
Fig. 9. Peripheral nerve fibres (PN) embedded withincollagenous endoneurium (E; detail of femoral nerve incross section). TEM. Scale bar 5 5 µm.
Fig. 10. The multilamellar patterns of a peripheralnerve’s myelin sheath (from skin). Axon remains (AX)appear amorphous. TEM, UA/Pb. Scale bar 5 0.1 µm.
Fig. 11. Vegetative ganglion with thin nerve fibres anddisintegrated ganglion cells (GC). TEM. Scale bar 5 5 µm.
Fig. 12. Ultrastructural localization of periodic acid-Schiff-reactive carbohydrates in femoral nerve myelin. TEM;periodic acid-thiocarbohydrazide-silver proteinate staining.E, endoneurium. Scale bar 5 0.5 µm.
Fig. 13. Sudan black staining of myelin lipid (arrows) ina 1-µm cryosection through the femoral nerve. LM. Scalebar 5 20 µm.
528 M.W. HESS ET AL.

the small and large intestine, as well as thelung, and, likewise, for the liver paren-chyma (Fig. 14), peripheral blood, bone mar-row, and the splenic pulp. These findings
contrast in part with reports of remarkablywell-preserved blood and epidermal cells inartificial and natural dry mummies (see,e.g., Lewin, 1967; Zimmerman, 1973; Riddleet al., 1975; Perrin et al., 1994; Gerszten etal., 1997). The only examples of recognizableepithelial remains in the Ice Man are theclearly defined cellular patterns of the haircuticle (Fig. 15).
With regard to muscle tissue, the outlinesof cells/fibers sometimes appeared still dis-cernible, but the cells’ internal architecturewas generally destroyed (see, e.g., Figs. 7, 8).It is of note that Zimmerman and Tedford(1976) could show cross striations preservedin skeletal muscle from a 21,000-year-oldmammoth.
Control samples of any kind of soft tissueobtained from a present-day glacier corpseshowed almost complete structural break-down as a consequence of adipocere forma-tion.
Hypotheses dealing with thetissue-specific preservation patterns
of the Ice Man’s tissues
It seems reasonable to characterize brieflythe major mechanisms that account gener-ally for tissue disintegration in glacier-corpses. In chronological order, these are 1)autolysis and bacterial activity (biologicaldestruction; gram-negative bacteria were ob-served in the digestive tract and within thelumen of a Haversian canal of rib bone); 2)freeze/thaw-damage, i.e., (intra)cellular icecrystallization, which causes membrane dis-ruption and, in turn, confluence of (sub)cellu-lar compartments (physical destruction); and3) transformation of molecules into adipo-cere (chemical destruction; note: the mecha-nisms that account for this phenomenon,which has been known for centuries, are stillenigmatic in detail; for reviews, see Be-reuter et al., 1996a; Mayer et al., 1997).These destructive forces act in a cumulativeway (in this context, see also Andreasen etal., 1991).
In the following, we consider some proper-ties of (sub)cellular constituents that pre-sumably contributed to impede disintegra-tion of the Ice Man’s body. First, structuresthat are distant from sites of enzymic activ-ity were clearly not affected by autolysis,
Fig. 14. Crystalline adipocere observed in liversamples. TEM; UA/Pb. Scale bar 5 5 µm.
Fig. 15. Inner hair cuticle (HC) with preserved cellu-lar patterns. Double arrow marks a pigment granule.TEM; UA/Pb. Scale bar 5 1 µm.
529HISTOLOGY OF THE TYROLEAN ICE MAN

e.g., myelin sheaths and extracellular fibers.Low metabolic activity, too, may have lim-ited autolysis, for instance, in chondrocytes.Second, freeze/thaw damage was limitedlocally by the chemical composition of subcel-lular constituents: that is, high proportionsof lipids and/or low water content of themyelin sheaths and dense collagenous tis-sue, respectively. Analogously, the specificchemical composition of cytoplasm andnucleus (i.e., different freezing properties)may account for the preservation of thenuclear compartment as a distinct struc-tural entity in chondrocytes. Third, oneshould mention the physical properties ofsubcellular components: Highly orderedstructures, such as myelin (being consideredas ‘‘paracrystalline’’: Fernandez-Moran,1962) and collagen fibrils, essentially with-stood destruction resulting from crystalliza-tion processes such as ice crystallization andadipocere formations. Finally, the isolatedlocation of some kinds of cells should also beconsidered. The template of extracellularmatrix and fibers, within which fibrocytes,perineural cells, and chondrocytes are em-bedded, may have helped to retain theirmore or less disintegrated residues in place,that is, to preserve at least the individualcontours of these cells.
On the other hand, metabolically activecells that are rich in protein and/or water,such as internal epithelia (e.g., glands), thenerve cell bodies, reticular connective tis-sues, blood, and muscle, as well as thedigestive tract with its bacteria, generallydid not withstand structural disintegrationand/or transformation processes (i.e., autoly-sis, bacterial decomposition, freeze/thawdamage, adipocere formation). Together, ourhistological data, in general, are consistentwith reports on the molecular state of the IceMan’s tissues (i.e., partially well-preservedlipoidal components: Williams et al., 1995;Makristathis et al., 1996; more or less de-graded proteins: Lubec et al., 1994; andmore or less degraded nucleic acids: Handtet al., 1994).
CONCLUSIONS AND OUTLOOK
Slight modifications of standard prepara-tion procedures for electron microscopy al-lowed detailed microscopic analysis of a
broad range of tissue samples obtained fromthe Tyrolean Ice Man. According to ultra-structural criteria, this naturally mummi-fied, prehistoric human, in part, remainedremarkably well preserved over millennia.Whether tissue components still exhibit lec-tin-binding or immunoreactivity (seeFulcheri, 1995) is currently under investiga-tion. Apart from the morphological findings,methodological notes may be of interest forothers who are investigating similar finds.
ACKNOWLEDGMENTS
We are grateful to W. Platzer (Institute ofAnatomy, University of Innsbruck) and M.Pavelka (Institute of Histology and Embryol-ogy, University of Innsbruck) for their con-tinuous support. Special thanks for helpfuldiscussion are due to B. Afzelius (Stock-holm), P. Debbage (Innsbruck), R. Hermann(Zurich), H. Hohenberg (Hamburg), E. Kir-schning (Hamburg), H. Lassmann (Wien), E.Linder (Stockholm), J. Loidl (Wien), M.Michel (Lausanne), M. Pavelka (Innsbruck),W. Pfaller (Innsbruck), H. Plattner (Kon-stanz), W. Schmidt (Innsbruck), H. Sitte(Homburg/Saar), M. Spycher (Zurich), D.Studer (Bern), W. Voorhout (Utrecht), and P.Walther (Zurich). Technical assistance of theinstitutes’ staffs, especially the skillful workof K. Gutleben and photographic artwork ofR. Haring, is acknowledged.
LITERATURE CITED
Aichel OH (1927) Uber Moorleichen, nebst Mitteilungeines neuen Falles (2.5 jahriges Madchen von Rost inDithmarschen. Anthropologischer Anzeiger 4(Suppl.II):57–73.
Ambach E, Tributsch W, and Ambach W (1992) Ismummification possible in snow. Forens. Sci. Int.54:191–192.
Andreasen C, Gullov HC, Hart-Hansen JP, Lyberth J,and Tauber H (1991) The find. In Hart-Hansen JP,Meldgaard J, and Nordqvist J (eds.): The GreenlandMummies. London: The Trustees of the British Mu-seum, Inc., pp. 37–52.
Bereuter TL, Lorbeer E, Reiter C, Seidler H, andUnterdorfer H (1996a) Post-mortem alteration of hu-man lipids-part I: Evaluation of adipocere formationand mummification by desiccation. In Spindler K,Wilfing H, Rastbichler-Zissernig E, zur Nedden D,and Nothdurfter H (eds.): Human Mummies: A GlobalSurvey of their Status and the Techniques of Conser-vation. Wien: Springer, Inc., pp. 265–273.
Bereuter TL, Reiter C, Seidler H, and Platzer W (1996b)Post-mortem alteration of human lipids-part II: Lipidcomposition of a skin sample from the Iceman. InSpindler K, Wilfing H, Rastbichler-Zissernig E., zurNedden D, and Nothdurfter H (eds.): Human Mum-mies: A Global Survey of their Status and the Tech-
530 M.W. HESS ET AL.

niques of Conservation. Wien: Springer, Inc., pp.275–278.
Bonani G, Ivy SD, Niklaus TR, Suter M, Housley RA,Bronk CR, van Klinken GJ, and Hedges REM (1992)Altersbestimmung von Milligrammproben der OtztalerGletscherleiche mit der Beschleunigermassenspe-ktrometriemethode (AMS). In Hopfel F, Platzer W,and Spindler K (eds.): Der Mann im Eis, Band 1,Veroffentlichungen der Universitat Innsbruck 187.Wien: Springer, Inc., pp. 108–116.
Courtoy R, and Simar LJ (1974) Importance of controlsfor the demonstration of carbohydrates in electronmicroscopy with the silver methenamine or the thiocar-bohydrazide-silver proteinate methods. J. Microsc.Oxford 100:199–211.
Doran GH, Dickel DN, Ballinger WE, Agee OF, LaipisPJ, and Hauswirth WW (1986) Anatomical, cellularand molecular analysis of 8,000-year-old human braintissue from the Windover archeological site. Nature323:803–806.
Erdos GW (1986) Localization of carbohydrate contain-ing molecules. In Aldrich HC and Todd WJ (eds.):Ultrastructure Techniques for Microorganisms. NewYork: Plenum Press, Inc., pp. 399–420.
Erk I, Michel M, and Lepault J (1996) Electron cryo-microscopy of vitrified bulk biological specimens: Idealand real structures of water-lipid phases. J. Microsc.Oxford 182:15–23.
Fernandez-Moran H (1962) Cell membrane ultrastruc-ture. Low-temperature electron microscopy and x-raydiffraction studies of lipoprotein components in lamel-lar systems. Circulation 26:1039–1065.
Fisher DB (1968) Protein staining of ribboned Eponsections for light microscopy. Histochemie 16:92–96.
Fulcheri E (1995) Immunohistochemistry: a new out-look in histopaleopathology. J. Biol. Res. Boll. Soc. It.Biol. Sper. 71:105–110.
Gaber O, Kunzel KH, Maurer H, Bodner E, Thumfart W,Glaser K, Gunkel A, Truppe MJ, Freysinger W, andTiefenbrunner F (1995) Endoscopic collection of speci-mens of the Ice Man [abstract]. Acta Anat. 152:285.
Gerszten PC, and Martınez AJ (1995) The neuropathol-ogy of South American mummies. Neurosurgery 36:756–761.
Gerszten PC, Gerszten E, and Allison MJ (1997) Ultra-structure of a well preserved lymphocyte from amummified human. J. Electron Microsc. 46:443–445.
Gouzhang Z, Wenhui F, Yiheng B, Jinian X, and YunshuY (1979) Microscopic and submicroscopic studies onthe peripheral nerve and the skeletal muscle of thefemale cadaver found in the Han tomb no. 1. ScientiaSinica 22:1095–1098.
Gunkel AR, Freysinger W, Thumfart WF, Truppe MJ,Gaber O, Kunzel KH, Platzer W, and Tiefenbrunner F(1997) Otorhinolaryngologic computer-assisted biop-sies of the Iceman. Arch. Otolaryngol. Head NeckSurg. 123:253–256.
Handt, O, Richards M, Trommsdorff M, Kilger C, Sima-nainen J, Georgiev O, Bauer K, Stone A, Hedges R,Schaffner W, Utermann G, Sykes B, and Paabo S(1994) Molecular genetic analyses of the Tyrolean IceMan. Science 264:1775–1178.
Hart-Hansen JP, Meldgaard J, and Nordqvist J (1991)The Greenland Mummies. London: The Trustees ofthe British Museum, Inc.
Hayat MA (1989) Principles and Techniques of ElectronMicroscopy. Biological Applications, 3rd ed. Hound-smills: Macmillan Press, Inc.
Hess MW (1990) Spurr-resin affects the ultrastructureof freeze-fixed/freeze-substituted pollen grains. Ultra-microscopy 34:219–221.
Hess MW, and Hesse M (1994) Ultrastructural observa-tions on anther tapetum development of freeze-fixed
Ledebouria socialis Roth (Hyacinthaceae). Planta 192:421–430.
Hess MW, Klima G, Gaber O, Kunzel KH, Pavelka M,and Platzer W (1996a) Histologische Beobachtungenan der jungsteinzeitlichen Gletschermumie vom Haus-labjoch. 18. Alpenlandisches Anatomentreffen, 16–19Mai, 1996, Munchen [abstract]. Ann. Anat. 178:571.
Hess MW, Klima G, Pfaller K, Gaber O, Kunzel KH,Pavelka M, and Platzer W (1996b) The Neolithicglacier-mummy from the Hauslabjoch (Schnalstal,Autonome Provinz Bozen-Sudtirol, Italy). Aspects ofultrastructure preservation [abstract]. Electron Mi-croscopy 1996: Proceedings of the XI European Con-gress on Microscopy Dublin, Ireland 26–30 August1996 (in press).
Lewin PK (1967) Paleo-electron microscopy of mummi-fied tissue. Nature 213:416–417
Lubec G, Weninger M, and Anderson SR (1994) Racem-ization and oxidation studies of hair protein in theHomo tiroliensis. FASEB J. 8:1166–1169.
Makristathis A., Mader R, Varmuza K, Simonitsch I,Scharzmeier J, Seidler H, Platzer W, Unterndorfer H,and Scheithauer R (1996) Comparison of the lipidprofile of the Tyrolean Iceman with bodies recoveredfrom glaciers. In Spindler K, Wilfing H, Rastbichler-Zissernig E., zur Nedden D, and Nothdurfter H (eds.):Human Mummies: A Global Survey of Their Statusand the Techniques of Conservation. Wien: Springer,Inc., pp. 279–281.
Manzini G, Barcellona ML, Avitabile M, and Quadrifo-glio F (1983) Interaction of diamidino-2-phenylindole(DAPI) with natural and synthetic nucleic acids.Nucleic Acids Res. 11:8861–8876.
Mayer BX, Reiter C, and Bereuter TL (1997) Investiga-tion of the triacylglycerol composition of iceman’smummified tissue by high-temperature gas chromatog-raphy. J. Chromatogr. B. Biomed. Sci. Appl. 692:1–6.
Montes GS, Krisztan RM, and Junqueira LCU (1985)Preservation of elastic system fibers and of collagenmolecular arrangement and stainability in an Egyp-tian mummy. Histochemistry 83:117–119.
Pabst MA, and Hofer F (1998) Deposits of differentorigin in the lung of the 5300-year-old Tyrolean IceMan. Am. J. Phys. Anthropol. (in press).
Pearse AGE (1980) Histochemistry Theoretical and Ap-plied, 4th ed. Edinburgh: Churchill Livingstone, Inc.
Perrin C, Noly V, Mourer R, and Schmitt D (1994)Preservation des structures cutanees des momiesd’egypte-etude ultrastructurale. Ann. Dermatol. Vene-reol. 121:470–475.
Platzer W, Kunzel KH, Gaber O, and Maurer H (1998)Endoskopische Untersuchungstechnik und Probenent-nahme beim Eismann. In: The Man in the Ice, Vol. 5,Wien: Springer, Inc. (in press).
Radanov S, Stoev S, Davidov M, Nachev S, Stanchev N,and Kirova E (1992) A unique case of naturallyoccurring mummification of human brain tissue. Int.J. Leg. Med. 105:173–175.
Reinhard J (1996) Peru’s ice maidens. Natl. Geographic189/6:62–81.
Reynolds ES (1963) The use of lead citrate at high pH asan electron opaque stain in electron microscopy. J.Cell Biol. 17:208–212.
Riddle JM, Ho KL, Chason JL, and Schwyn RC (1975)Peripheral blood elements found in an Egyptianmummy: A three-dimensional view. Science 192:374–375.
Rowley JR, and Dahl AO (1977) Pollen development inArtemisia vulgaris with special reference to glycoca-lyx material. Pollen Spores 19:169–184.
Ruffer MA (1921) Studies in the Paleopathology ofEgypt. Chicago: University of Chicago Press, Inc.
531HISTOLOGY OF THE TYROLEAN ICE MAN

Sandison AT (1955) The histological examination ofmummified material. Stain Technol. 30:277–283.
Sandison AT (1963) Staining of vascular elastic fibres inmummified and dried human tissues. Nature 198:597.
Seidler H, Bernhard W, Teschler-Nicola M, Platzer W,zur Nedden D, Henn R, Oberhauser A, and Sjovold T(1992) Some anthropological aspects of the prehistoricTyrolean Ice Man. Science 258:455–457.
Snodgress AB, Dorsey CH, Bailey GWD, and DicksonLG (1972) Conventional histopathologic staining meth-ods compatible with Epon-embedded, ossmicated tis-sue. Lab. Invest. 26:329–337.
Spindler K, Wilfing H, Rastbichler-Zissernig E., zurNedden D, and Nothdurfter H (1996) Human Mum-mies: A Global Survey of their Status and the Tech-niques of Conservation. Wien: Springer, Inc.
Spurr AR (1969) A low-viscosity epoxy resin embeddingmedium for electron microscopy. J. Ultrastruct. Res.26:31–43.
Thiery JP (1967) Mise en evidence des polysaccharides
sur coupes fines en microscopie electronique. J. Micros-copie. Paris 6:987–1018.
Tokuyasu KT (1973) A technique for ultracryotomy ofcell suspensions and tissues. J. Cell Biol. 57:551–565.
Williams AC, Edwards HGM, and Barry BW (1995) The‘‘Iceman’’: Molecular structure of 5200-year-old skincharacterised by Raman spectroscopy and electronmicroscopy. Biochim. Biophys. Acta 1246:98–105.
Zimmerman MR (1973) Blood cells preserved in amummy 2000 years old. Science 180:303–304.
Zimmerman MR (1996) Mummies of the arctic regions.In Spindler K, Wilfing H, Rastbichler-Zissernig E., zurNedden D, and Nothdurfter H (eds.): Human Mum-mies: A Global Survey of their Status and the Tech-niques of Conservation. Wien: Springer, Inc., pp.83–92.
Zimmerman MR, and Tedford RH (1976) Histologicstructures preserved for 21,300 years. Science 194:183–184.
532 M.W. HESS ET AL.


















