
Gut infl transfer between pathogenic and commensal ... · determined by 16S rRNA sequencing (Fig....
6
Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae Bärbel Stecher a,b,1 , Rémy Denzler a , Lisa Maier a , Florian Bernet a , Mandy J. Sanders c , Derek J. Pickard c , Manja Barthel a , Astrid M. Westendorf d , Karen A. Krogfelt e , Alan W. Walker c , Martin Ackermann f,g , Ulrich Dobrindt h,i , Nicholas R. Thomson c , and Wolf-Dietrich Hardt a a Institute of Microbiology, ETH Zürich, 8093 Zürich, Switzerland; b Max-von-Pettenkofer Institute, Ludwig-Maximilians-Universität Munich, 80336 Munich, Germany; c Welcome Trust Sanger Institute, Genome Campus, Hinxton, Cambridge CB10 1SA, United Kingdom; d Institute of Medical Microbiology, University Hospital, University Duisburg-Essen, 45122 Essen, Germany; e Department of Surveillance and Research, Statens Serum Institut, 2300 S Copenhagen, Denmark; f Department of Environmental Sciences, ETH Zürich, 8092 Zürich, Switzerland; g Department of Environmental Microbiology, Eawag, 8600 Dubendorf, Switzerland; h Institute for Molecular Infectious Biology, University of Würzburg, 97080 Würzburg, Germany; and i Institute for Hygiene, University of Münster, 48149 Münster, Germany Edited by John J. Mekalanos, Harvard Medical School, Boston, MA, and approved December 12, 2011 (received for review August 18, 2011) The mammalian gut harbors a dense microbial community inter- acting in multiple ways, including horizontal gene transfer (HGT). Pangenome analyses established particularly high levels of genetic flux between Gram-negative Enterobacteriaceae. However, the mechanisms fostering intraenterobacterial HGT are incompletely understood. Using a mouse colitis model, we found that Salmo- nella-inflicted enteropathy elicits parallel blooms of the pathogen and of resident commensal Escherichia coli. These blooms boosted conjugative HGT of the colicin-plasmid p2 from Salmonella enter- ica serovar Typhimurium to E. coli. Transconjugation efficiencies of ∼100% in vivo were attributable to high intrinsic p2-transfer rates. Plasmid-encoded fitness benefits contributed little. Under normal conditions, HGT was blocked by the commensal microbiota inhib- iting contact-dependent conjugation between Enterobacteriaceae. Our data show that pathogen-driven inflammatory responses in the gut can generate transient enterobacterial blooms in which conjugative transfer occurs at unprecedented rates. These blooms may favor reassortment of plasmid-encoded genes between pathogens and commensals fostering the spread of fitness-, viru- lence-, and antibiotic-resistance determinants. bacterial evolution | hospital-acquired infection | mucosal immune response | plasmid spread T he mammalian gut harbors a very dense microbial commu- nity, the “microbiota” (>10 12 bacteria/g), which has profound effects on the host’s nutrition, physiology, and immune system (1). In humans, the microbiota is generally composed of several hundred different bacterial phylotypes. Its composition differs between individuals forming a “collective microbiome” of more than 100,000 genes (2, 3). Much of the microbiota’s genome plasticity is thought to be attributable to horizontal gene transfer (HGT), the most effective mechanism of which is conjugation, the exchange of plasmids (4–6). Conjugational plasmid transfer is consistently fueling the emergence of hypervirulent or antibi- otic-resistant pathogens (7, 8), as illustrated by Escherichia coli O104:H4, which has recently caused an outbreak in Germany (9). However, key questions about the maintenance and the transfer of conjugative plasmids in natural bacterial populations have remained unanswered. Here, we describe the discovery of a mechanism driving efficient conjugation between Enter- obacteriaceae in the host’s intestine. Enterobacteriaceae include many pathogenic as well as commensal species (i.e., Salmonella enterica and E. coli ). Plasmid profiling, enterobacteriaceal genome sequencing, and microbiome analyses identified a large and highly diverse plasmid-encoded accessory gene pool indicative of efficient conjugative HGT (10). However, in the normal gut, the Enter- obacteriaceae are generally present in very low densities (<<10 8 cfu/g). These densities are way too low for efficient conjugative plasmid transfer, as this process hinges on direct physical contact between the donor and the acceptor bacterium (11). In the mammalian intestine, the vast majority of intestinal bacteria consist of obligate anaerobic members of the Firmicutes and Bacteroidetes phyla. It is thought that these anaerobes keep the overall density of facultative anaerobic bacteria (i.e., Enter- obacteriaceae) rather low (<<10 8 cfu/g), a condition termed colonization resistance (CR) (12–14). Thus, in complex bacterial ecosystems, low densities of donor and recipient bacteria may lower the frequency of direct bacterial encounters and thus de- crease the chance of conjugation-mediated HGT. Several studies reported inefficient enterobacterial HGT in the normal mam- malian gut (15–17), whereas others identified higher rates of HGT (18, 19). This suggested that particular conditions exist that might favor plasmid exchange between Enterobacteriaceae. The factors influencing the efficiency of conjugative HGT in this bacterial family are incompletely understood to date. Inflammatory host responses triggered by the gut immune system (in inflammatory bowel disease patients) or by pathogens such as Salmonella spp. or pathogenic E. coli strains can suppress the anaerobic microbiota and boost enterobacterial colonization densities (20–24). Here, we show that disease-triggered entero- bacterial blooms can fuel HGT between two prominent members of this family, Salmonella enterica serovar Typhimurium (S. Tm) and E. coli. Results and Discussion We have frequently observed parallel blooms (≥10 8 cfu/g each) of S. Tm strain SL1344 (S. Tm wt ) and commensal E. coli in our experiments using the streptomycin mouse model for Salmonella diarrhea. Fig. 1 shows the presence of commensal E. coli recorded during routine screening of S. Tm wt infected mice by plating on selective agar (Fig. 1A) (21, 25, 26). In some cases, E. coli accounted for >80% of the total intestinal bacteria as Author contributions: B.S., R.D., and W.-D.H. designed research; B.S., R.D., L.M., F.B., M.J.S., D.J.P., M.B., U.D., and N.R.T. performed research; A.M.W. contributed new reagents/ana- lytic tools; B.S., R.D., L.M., F.B., M.B., K.A.K., A.W.W., M.A., U.D., and N.R.T. analyzed data; and B.S., M.A., and W.-D.H. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Freely available online through the PNAS open access option. Database deposition: The 16S rRNA gene sequences reported in this paper have been deposited in the GenBank [accession nos. HE582402–HE582620 (ENA)]. 1 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1113246109/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1113246109 PNAS | January 24, 2012 | vol. 109 | no. 4 | 1269–1274 MICROBIOLOGY
-
Upload
nguyenphuc -
Category
Documents
-
view
221 -
download
0
Transcript of Gut infl transfer between pathogenic and commensal ... · determined by 16S rRNA sequencing (Fig....

Gut inflammation can boost horizontal genetransfer between pathogenic andcommensal EnterobacteriaceaeBärbel Stechera,b,1, Rémy Denzlera, Lisa Maiera, Florian Berneta, Mandy J. Sandersc, Derek J. Pickardc, Manja Barthela,Astrid M. Westendorfd, Karen A. Krogfelte, Alan W. Walkerc, Martin Ackermannf,g, Ulrich Dobrindth,i,Nicholas R. Thomsonc, and Wolf-Dietrich Hardta
aInstitute of Microbiology, ETH Zürich, 8093 Zürich, Switzerland; bMax-von-Pettenkofer Institute, Ludwig-Maximilians-Universität Munich, 80336 Munich,Germany; cWelcome Trust Sanger Institute, Genome Campus, Hinxton, Cambridge CB10 1SA, United Kingdom; dInstitute of Medical Microbiology, UniversityHospital, University Duisburg-Essen, 45122 Essen, Germany; eDepartment of Surveillance and Research, Statens Serum Institut, 2300 S Copenhagen, Denmark;fDepartment of Environmental Sciences, ETH Zürich, 8092 Zürich, Switzerland; gDepartment of Environmental Microbiology, Eawag, 8600 Dubendorf,Switzerland; hInstitute for Molecular Infectious Biology, University of Würzburg, 97080 Würzburg, Germany; and iInstitute for Hygiene, University of Münster,48149 Münster, Germany
Edited by John J. Mekalanos, Harvard Medical School, Boston, MA, and approved December 12, 2011 (received for review August 18, 2011)
The mammalian gut harbors a dense microbial community inter-acting in multiple ways, including horizontal gene transfer (HGT).Pangenome analyses established particularly high levels of geneticflux between Gram-negative Enterobacteriaceae. However, themechanisms fostering intraenterobacterial HGT are incompletelyunderstood. Using a mouse colitis model, we found that Salmo-nella-inflicted enteropathy elicits parallel blooms of the pathogenand of resident commensal Escherichia coli. These blooms boostedconjugative HGT of the colicin-plasmid p2 from Salmonella enter-ica serovar Typhimurium to E. coli. Transconjugation efficiencies of∼100% in vivo were attributable to high intrinsic p2-transfer rates.Plasmid-encoded fitness benefits contributed little. Under normalconditions, HGT was blocked by the commensal microbiota inhib-iting contact-dependent conjugation between Enterobacteriaceae.Our data show that pathogen-driven inflammatory responses inthe gut can generate transient enterobacterial blooms in whichconjugative transfer occurs at unprecedented rates. These bloomsmay favor reassortment of plasmid-encoded genes betweenpathogens and commensals fostering the spread of fitness-, viru-lence-, and antibiotic-resistance determinants.
bacterial evolution | hospital-acquired infection | mucosal immuneresponse | plasmid spread
The mammalian gut harbors a very dense microbial commu-nity, the “microbiota” (>1012 bacteria/g), which has profound
effects on the host’s nutrition, physiology, and immune system(1). In humans, the microbiota is generally composed of severalhundred different bacterial phylotypes. Its composition differsbetween individuals forming a “collective microbiome” of morethan 100,000 genes (2, 3). Much of the microbiota’s genomeplasticity is thought to be attributable to horizontal gene transfer(HGT), the most effective mechanism of which is conjugation,the exchange of plasmids (4–6). Conjugational plasmid transferis consistently fueling the emergence of hypervirulent or antibi-otic-resistant pathogens (7, 8), as illustrated by Escherichia coliO104:H4, which has recently caused an outbreak in Germany(9). However, key questions about the maintenance and thetransfer of conjugative plasmids in natural bacterial populationshave remained unanswered. Here, we describe the discovery ofa mechanism driving efficient conjugation between Enter-obacteriaceae in the host’s intestine. Enterobacteriaceae includemany pathogenic as well as commensal species (i.e., Salmonellaenterica and E. coli). Plasmid profiling, enterobacteriaceal genomesequencing, and microbiome analyses identified a large and highlydiverse plasmid-encoded accessory gene pool indicative of efficientconjugative HGT (10). However, in the normal gut, the Enter-obacteriaceae are generally present in very low densities (<<108
cfu/g). These densities are way too low for efficient conjugativeplasmid transfer, as this process hinges on direct physical contactbetween the donor and the acceptor bacterium (11).In the mammalian intestine, the vast majority of intestinal
bacteria consist of obligate anaerobic members of the Firmicutesand Bacteroidetes phyla. It is thought that these anaerobes keepthe overall density of facultative anaerobic bacteria (i.e., Enter-obacteriaceae) rather low (<<108 cfu/g), a condition termedcolonization resistance (CR) (12–14). Thus, in complex bacterialecosystems, low densities of donor and recipient bacteria maylower the frequency of direct bacterial encounters and thus de-crease the chance of conjugation-mediated HGT. Several studiesreported inefficient enterobacterial HGT in the normal mam-malian gut (15–17), whereas others identified higher rates ofHGT (18, 19). This suggested that particular conditions exist thatmight favor plasmid exchange between Enterobacteriaceae. Thefactors influencing the efficiency of conjugative HGT in thisbacterial family are incompletely understood to date.Inflammatory host responses triggered by the gut immune
system (in inflammatory bowel disease patients) or by pathogenssuch as Salmonella spp. or pathogenic E. coli strains can suppressthe anaerobic microbiota and boost enterobacterial colonizationdensities (20–24). Here, we show that disease-triggered entero-bacterial blooms can fuel HGT between two prominent membersof this family, Salmonella enterica serovar Typhimurium (S. Tm)and E. coli.
Results and DiscussionWe have frequently observed parallel blooms (≥108 cfu/g each)of S. Tm strain SL1344 (S. Tmwt) and commensal E. coli in ourexperiments using the streptomycin mouse model for Salmonelladiarrhea. Fig. 1 shows the presence of commensal E. colirecorded during routine screening of S. Tmwt infected mice byplating on selective agar (Fig. 1A) (21, 25, 26). In some cases,E. coli accounted for >80% of the total intestinal bacteria as
Author contributions: B.S., R.D., andW.-D.H. designed research; B.S., R.D., L.M., F.B., M.J.S.,D.J.P., M.B., U.D., and N.R.T. performed research; A.M.W. contributed new reagents/ana-lytic tools; B.S., R.D., L.M., F.B., M.B., K.A.K., A.W.W., M.A., U.D., and N.R.T. analyzed data;and B.S., M.A., and W.-D.H. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.
Database deposition: The 16S rRNA gene sequences reported in this paper have beendeposited in the GenBank [accession nos. HE582402–HE582620 (ENA)].1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1113246109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1113246109 PNAS | January 24, 2012 | vol. 109 | no. 4 | 1269–1274
MICRO
BIOLO
GY
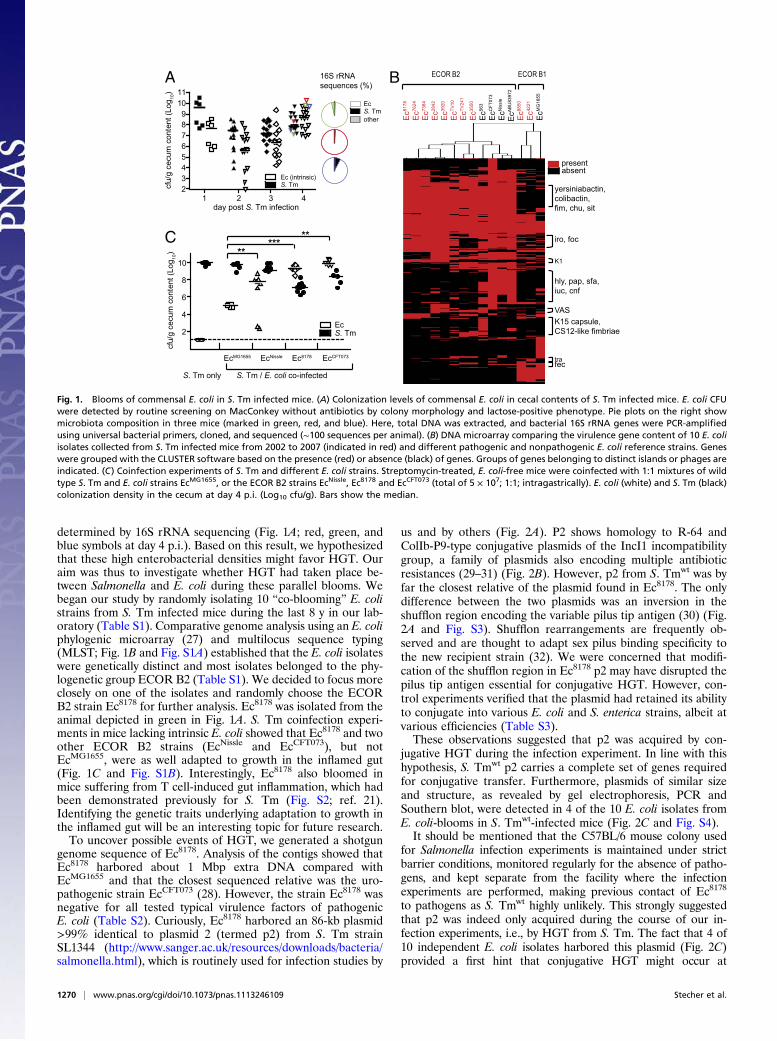
determined by 16S rRNA sequencing (Fig. 1A; red, green, andblue symbols at day 4 p.i.). Based on this result, we hypothesizedthat these high enterobacterial densities might favor HGT. Ouraim was thus to investigate whether HGT had taken place be-tween Salmonella and E. coli during these parallel blooms. Webegan our study by randomly isolating 10 “co-blooming” E. colistrains from S. Tm infected mice during the last 8 y in our lab-oratory (Table S1). Comparative genome analysis using an E. coliphylogenic microarray (27) and multilocus sequence typing(MLST; Fig. 1B and Fig. S1A) established that the E. coli isolateswere genetically distinct and most isolates belonged to the phy-logenetic group ECOR B2 (Table S1). We decided to focus moreclosely on one of the isolates and randomly choose the ECORB2 strain Ec8178 for further analysis. Ec8178 was isolated from theanimal depicted in green in Fig. 1A. S. Tm coinfection experi-ments in mice lacking intrinsic E. coli showed that Ec8178 and twoother ECOR B2 strains (EcNissle and EcCFT073), but notEcMG1655, were as well adapted to growth in the inflamed gut(Fig. 1C and Fig. S1B). Interestingly, Ec8178 also bloomed inmice suffering from T cell-induced gut inflammation, which hadbeen demonstrated previously for S. Tm (Fig. S2; ref. 21).Identifying the genetic traits underlying adaptation to growth inthe inflamed gut will be an interesting topic for future research.To uncover possible events of HGT, we generated a shotgun
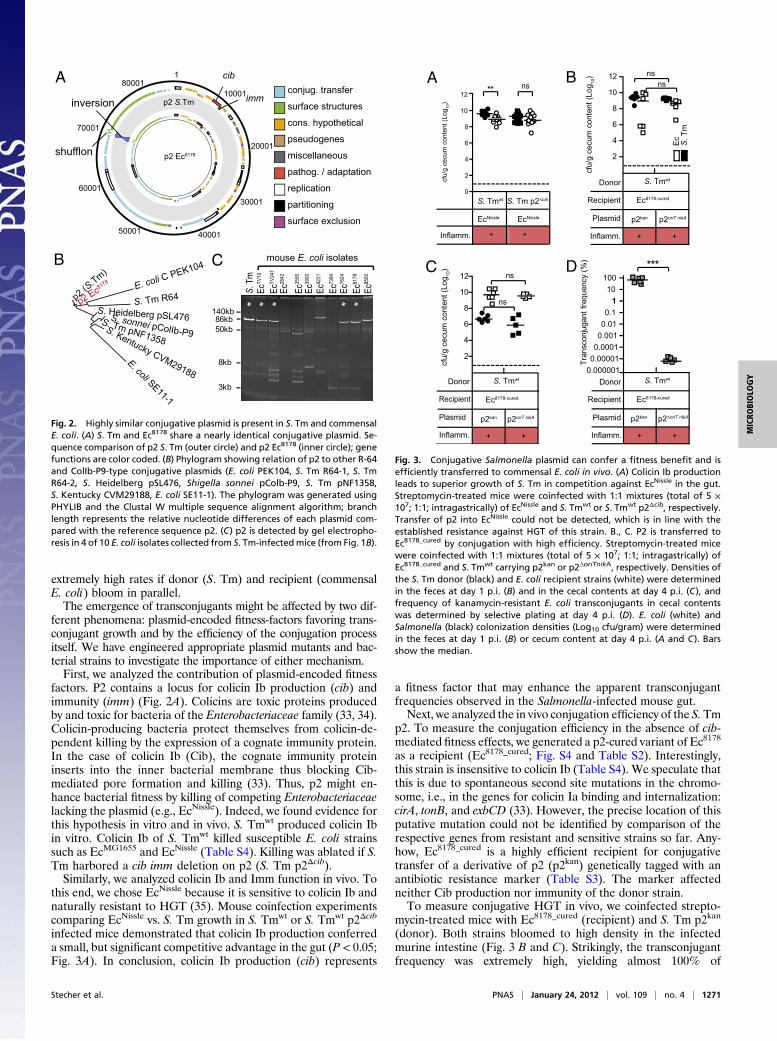
genome sequence of Ec8178. Analysis of the contigs showed thatEc8178 harbored about 1 Mbp extra DNA compared withEcMG1655 and that the closest sequenced relative was the uro-pathogenic strain EcCFT073 (28). However, the strain Ec8178 wasnegative for all tested typical virulence factors of pathogenicE. coli (Table S2). Curiously, Ec8178 harbored an 86-kb plasmid>99% identical to plasmid 2 (termed p2) from S. Tm strainSL1344 (http://www.sanger.ac.uk/resources/downloads/bacteria/salmonella.html), which is routinely used for infection studies by
us and by others (Fig. 2A). P2 shows homology to R-64 andColIb-P9-type conjugative plasmids of the IncI1 incompatibilitygroup, a family of plasmids also encoding multiple antibioticresistances (29–31) (Fig. 2B). However, p2 from S. Tmwt was byfar the closest relative of the plasmid found in Ec8178. The onlydifference between the two plasmids was an inversion in theshufflon region encoding the variable pilus tip antigen (30) (Fig.2A and Fig. S3). Shufflon rearrangements are frequently ob-served and are thought to adapt sex pilus binding specificity tothe new recipient strain (32). We were concerned that modifi-cation of the shufflon region in Ec8178 p2 may have disrupted thepilus tip antigen essential for conjugative HGT. However, con-trol experiments verified that the plasmid had retained its abilityto conjugate into various E. coli and S. enterica strains, albeit atvarious efficiencies (Table S3).These observations suggested that p2 was acquired by con-
jugative HGT during the infection experiment. In line with thishypothesis, S. Tmwt p2 carries a complete set of genes requiredfor conjugative transfer. Furthermore, plasmids of similar sizeand structure, as revealed by gel electrophoresis, PCR andSouthern blot, were detected in 4 of the 10 E. coli isolates fromE. coli-blooms in S. Tmwt-infected mice (Fig. 2C and Fig. S4).It should be mentioned that the C57BL/6 mouse colony used
for Salmonella infection experiments is maintained under strictbarrier conditions, monitored regularly for the absence of patho-gens, and kept separate from the facility where the infectionexperiments are performed, making previous contact of Ec8178
to pathogens as S. Tmwt highly unlikely. This strongly suggestedthat p2 was indeed only acquired during the course of our in-fection experiments, i.e., by HGT from S. Tm. The fact that 4 of10 independent E. coli isolates harbored this plasmid (Fig. 2C)provided a first hint that conjugative HGT might occur at
A
2
4
6
8
10
cfu/
g ce
cum
con
tent
(Log
10)
*****
C
S. Tm only
EcMG1655 EcNissle Ec8178
S. Tm / E. coli co-infected
**
EcCFT073
23456789
1011
cfu/
g ce
cum
con
tent
(Log
10)
day post S. Tm infection1 2 3 4
S. TmEc
S. TmEc
ECOR B2 ECOR B1
Ec42
21
EcM
G16
55
Ec88
50E
cAB
U83
972
EcN
issl
eE
cCFT
073
Ec35
50
Ec56
3
EcTV
241
EcTV
10E
c3920
Ec28
42E
c7384
Ec76
24E
c8178
absentpresent
16S rRNAsequences (%)
other
S. TmEc (intrinsic)
yersiniabactin,colibactin,fim, chu, sit
iro, foc
K1
hly, pap, sfa,iuc, cnf
VASK15 capsule,CS12-like fimbriae
trafec
B
Fig. 1. Blooms of commensal E. coli in S. Tm infected mice. (A) Colonization levels of commensal E. coli in cecal contents of S. Tm infected mice. E. coli CFUwere detected by routine screening on MacConkey without antibiotics by colony morphology and lactose-positive phenotype. Pie plots on the right showmicrobiota composition in three mice (marked in green, red, and blue). Here, total DNA was extracted, and bacterial 16S rRNA genes were PCR-amplifiedusing universal bacterial primers, cloned, and sequenced (∼100 sequences per animal). (B) DNA microarray comparing the virulence gene content of 10 E. coliisolates collected from S. Tm infected mice from 2002 to 2007 (indicated in red) and different pathogenic and nonpathogenic E. coli reference strains. Geneswere grouped with the CLUSTER software based on the presence (red) or absence (black) of genes. Groups of genes belonging to distinct islands or phages areindicated. (C) Coinfection experiments of S. Tm and different E. coli strains. Streptomycin-treated, E. coli-free mice were coinfected with 1:1 mixtures of wildtype S. Tm and E. coli strains EcMG1655, or the ECOR B2 strains EcNissle, Ec8178 and EcCFT073 (total of 5 × 107; 1:1; intragastrically). E. coli (white) and S. Tm (black)colonization density in the cecum at day 4 p.i. (Log10 cfu/g). Bars show the median.
1270 | www.pnas.org/cgi/doi/10.1073/pnas.1113246109 Stecher et al.
extremely high rates if donor (S. Tm) and recipient (commensalE. coli) bloom in parallel.The emergence of transconjugants might be affected by two dif-
ferent phenomena: plasmid-encoded fitness-factors favoring trans-conjugant growth and by the efficiency of the conjugation processitself. We have engineered appropriate plasmid mutants and bac-terial strains to investigate the importance of either mechanism.First, we analyzed the contribution of plasmid-encoded fitness
factors. P2 contains a locus for colicin Ib production (cib) andimmunity (imm) (Fig. 2A). Colicins are toxic proteins producedby and toxic for bacteria of the Enterobacteriaceae family (33, 34).Colicin-producing bacteria protect themselves from colicin-de-pendent killing by the expression of a cognate immunity protein.In the case of colicin Ib (Cib), the cognate immunity proteininserts into the inner bacterial membrane thus blocking Cib-mediated pore formation and killing (33). Thus, p2 might en-hance bacterial fitness by killing of competing Enterobacteriaceaelacking the plasmid (e.g., EcNissle). Indeed, we found evidence forthis hypothesis in vitro and in vivo. S. Tmwt produced colicin Ibin vitro. Colicin Ib of S. Tmwt killed susceptible E. coli strainssuch as EcMG1655 and EcNissle (Table S4). Killing was ablated if S.Tm harbored a cib imm deletion on p2 (S. Tm p2Δcib).Similarly, we analyzed colicin Ib and Imm function in vivo. To
this end, we chose EcNissle because it is sensitive to colicin Ib andnaturally resistant to HGT (35). Mouse coinfection experimentscomparing EcNissle vs. S. Tm growth in S. Tmwt or S. Tmwt p2Δcib
infected mice demonstrated that colicin Ib production conferreda small, but significant competitive advantage in the gut (P < 0.05;Fig. 3A). In conclusion, colicin Ib production (cib) represents
a fitness factor that may enhance the apparent transconjugantfrequencies observed in the Salmonella-infected mouse gut.Next, we analyzed the in vivo conjugation efficiency of the S.Tm
p2. To measure the conjugation efficiency in the absence of cib-mediated fitness effects, we generated a p2-cured variant of Ec8178
as a recipient (Ec8178_cured; Fig. S4 and Table S2). Interestingly,this strain is insensitive to colicin Ib (Table S4). We speculate thatthis is due to spontaneous second site mutations in the chromo-some, i.e., in the genes for colicin Ia binding and internalization:cirA, tonB, and exbCD (33). However, the precise location of thisputative mutation could not be identified by comparison of therespective genes from resistant and sensitive strains so far. Any-how, Ec8178_cured is a highly efficient recipient for conjugativetransfer of a derivative of p2 (p2kan) genetically tagged with anantibiotic resistance marker (Table S3). The marker affectedneither Cib production nor immunity of the donor strain.To measure conjugative HGT in vivo, we coinfected strepto-
mycin-treated mice with Ec8178_cured (recipient) and S. Tm p2kan
(donor). Both strains bloomed to high density in the infectedmurine intestine (Fig. 3 B and C). Strikingly, the transconjugantfrequency was extremely high, yielding almost 100% of
1
10001
20001
30001
4000150001
60001
70001
80001
p2 Ec8178
p2 S.Tm imm
cib
p2 (S.Tm)
p2 Ec8178 E. coli C PEK104
E. coli SE11-1
S. Tm R64S. Heidelberg pSL476S. sonnei pColIb-P9S. Tm pNF1358S. Kentucky CVM29188
86kb
8kb
3kb
50kb
140kb
mouse E. coli isolates* * * **
S. T
mEc
TV10
EcTV
241
Ec28
42
Ec35
50
Ec39
20
Ec42
21
Ec73
94
Ec76
24
Ec81
78
Ec85
50
pathog. / adaptation
conjug. transfer
surface structures
cons. hypothetical
pseudogenes
miscellaneous
replication
partitioning
surface exclusion
shufflon
inversion
A
CB
Fig. 2. Highly similar conjugative plasmid is present in S. Tm and commensalE. coli. (A) S. Tm and Ec8178 share a nearly identical conjugative plasmid. Se-quence comparison of p2 S. Tm (outer circle) and p2 Ec8178 (inner circle); genefunctions are color coded. (B) Phylogram showing relation of p2 to other R-64and ColIb-P9-type conjugative plasmids (E. coli PEK104, S. Tm R64-1, S. TmR64-2, S. Heidelberg pSL476, Shigella sonnei pColb-P9, S. Tm pNF1358,S. Kentucky CVM29188, E. coli SE11-1). The phylogram was generated usingPHYLIB and the Clustal W multiple sequence alignment algorithm; branchlength represents the relative nucleotide differences of each plasmid com-pared with the reference sequence p2. (C) p2 is detected by gel electropho-resis in 4 of 10 E. coli isolates collected from S. Tm-infectedmice (from Fig. 1B).
2
4
6
8
10
12
***
cfu/
gce
cum
cont
ent (
Log 10
)
Donor
Recipient
Inflamm.
S. Tmwt
++
Ec8178-cured
Plasmid p2kan
ns
p2oriT nikA
ns
0
2
4
6
8
10
12
cfu/
gce
cum
cont
ent (
Log 10
)
** ns
S. Tm p2∆cib
EcNissle
S. Tmwt
EcNissle
0.0000010.000010.00010.0010.010.11
10100
Tran
scon
juga
ntfre
quen
cy (%
)
Donor
Recipient
Inflamm.
S. Tmwt
++
Ec8178-cured
Plasmid p2kan p2∆oriT nikA
Inflamm. ++
2
4
6
8
10
12
S. Tmwt
++
Ec8178-cured
p2kan
ns
p2oriT nikA
ns
S. T
mEc
Donor
Recipient
Inflamm.
Plasmid
A B
C D
cfu/
gce
cum
cont
ent (
Log 10
)
Fig. 3. Conjugative Salmonella plasmid can confer a fitness benefit and isefficiently transferred to commensal E. coli in vivo. (A) Colicin Ib productionleads to superior growth of S. Tm in competition against EcNissle in the gut.Streptomycin-treated mice were coinfected with 1:1 mixtures (total of 5 ×107; 1:1; intragastrically) of EcNissle and S. Tmwt or S. Tmwt p2Δcib, respectively.Transfer of p2 into EcNissle could not be detected, which is in line with theestablished resistance against HGT of this strain. B., C. P2 is transferred toEc8178_cured by conjugation with high efficiency. Streptomycin-treated micewere coinfected with 1:1 mixtures (total of 5 × 107; 1:1; intragastrically) ofEc8178_cured and S. Tmwt carrying p2kan or p2ΔoriTnikA, respectively. Densities ofthe S. Tm donor (black) and E. coli recipient strains (white) were determinedin the feces at day 1 p.i. (B) and in the cecal contents at day 4 p.i. (C), andfrequency of kanamycin-resistant E. coli transconjugants in cecal contentswas determined by selective plating at day 4 p.i. (D). E. coli (white) andSalmonella (black) colonization densities (Log10 cfu/gram) were determinedin the feces at day 1 p.i. (B) or cecum content at day 4 p.i. (A and C). Barsshow the median.
Stecher et al. PNAS | January 24, 2012 | vol. 109 | no. 4 | 1271
MICRO
BIOLO
GY

Ec8178_cured harboring p2kan by day 4 p.i. (Fig. 3D). No trans-conjugants were detected in control infections using a S. Tmwt
donor strain carrying an oriT-deficient, nonmobilizable plasmidvariant (p2ΔoriTnikA; Fig. 3 B and C). In conclusion, the intrinsicrate of conjugative HGT of p2 is extremely high in vivo, even ifa colicin-resistant E. coli strain is used as recipient.We also wanted to determine whether other p2 genes besides
cib would confer any detectable fitness benefit for the plasmid-bearing strain (i.e., S. Tm) in competition against plasmid-freeEc8178 in vivo. To test this, we had to use the conjugation-de-fective plasmid variant p2ΔoriTnikAΔcib to exclude conjugativetransfer to Ec8178 during the competition experiment. We coin-fected streptomycin-treated mice with S. Tm p2ΔoriTnikAΔcib andeither Ec8178 or Ec8178-cured. The competitive index of S. Tm andE. coli did not differ significantly between both experiments. Thisverified that p2 does not confer any other detectable benefit tothe recipient than those attributable to the cib/imm cassette (Fig.S5). Thus, in conclusion, extremely high transconjugant fre-quencies are observed in this system even in the absence ofplasmid-conferred fitness benefits.Our data suggested that conjugative HGT rates can be ex-
tremely high in pathogen-triggered enterobacterial blooms.However, it remained unclear whether high donor and recipientdensities per se were sufficient. Alternatively, the inflammatorymilieu in the gut might enhance HGT rates by some unknownmechanism. To address this, we analyzed conjugative transfer ofp2 from S. Tm strains into Ec8178_cured under three differentconditions:
i) In the first group, we coinfected animals with Ec8178_cured andan avirulent S. Tm SL1344 derivative (S. Tmavir p2cm), whichdoes not trigger gut inflammation (Table S2 and refs. 21 and36). For this group, we used low-complexity microbiota(LCM) mice, a colony harboring the commensals of the Al-tered Schaedler flora (37) in the gut. This gut flora allowedefficient Salmonella and E. coli gut colonization even in theabsence of enteropathy or antibiotic treatment (26) (Fig. 4 Aand C).
ii) In the second group, we infected streptomycin treated mice(normal gut microbiota) with the same strains as describedabove (i). In line with earlier work (21), the mice did notdevelop enteropathy, and regrowth of the normal gut micro-biota of these mice reduced enterobacterial colonization lev-els by day 4 p.i. (Fig. 4 A and C).
iii) In the third group, we coinfected streptomycin treated mice(normal gut microbiota) with Ec8178_cured and S. Tmwt (p2cm).This elicited gut inflammation and triggered parallel bloomsof the donor and the recipient (Fig. 4 A and C). The slightdifferences in total population sizes of donor and recipientsmight be attributable to minor differences in the bloomingkinetics of both strains.
In the first and the third group, conjugation efficienciesreached close to 100% and were significantly higher than in thesecond group (P < 0.05; Fig. 4B). Thus, a high rate of conjugativeHGT required high donor- and acceptor densities. Of note,100% transconjugation was achieved in LCM mice coinfectedwith the avirulent S. Tm strain. Therefore, HGT efficiency is notdirectly attributable to gut inflammation but rather to the hightiters of donor and recipient bacteria.To further substantiate this correlation, we manipulated the
density of the S. Tm donor strain in the inflamed gut and ana-lyzed the effects on the conjugation efficiency. To alter thedensity of the colicin-deficient S. Tm donor strain (S. Tmtet
p2Δcib), we used a conjugation-inert but otherwise geneticallyidentical S. Tm strain as “competitor” (S. Tm p2ΔoriTnikAΔcib) atvarious different concentrations. In addition, all S. Tm strainsinvolved were deficient in colicin production. We coinfected
groups of mice with Ec8178_cured (recipient) and the indicatedratio of S. Tmtet p2Δcib (donor) and S. Tm p2ΔoriTnikAΔcib (com-petitor; 10-, 100-, 1,000-, 10,000-, and 100,000-fold excess overthe donor strain). These experiments revealed a positive corre-lation between the donor density and the yield of trans-conjugants at day 1 and 4 p.i. (Fig. 5 A and B; P < 0.05; Fig. S6 Aand B). Similar results were observed in corresponding in vitroexperiments (Fig. S6C). However, only 1% transconjugantscould be maximally observed. We speculate that the extremelyhigh transconjugant frequencies seen in vivo could be due tosuccessive rounds of HGT during the 4-d-long infection periodor to very high local E. coli/S. Tm densities at some site of thehosts intestine.In conclusion, we report the highly efficient HGT of a natural
S. Tm plasmid to resident commensal E. coli in vivo. The ex-traordinary rate of transconjugant formation is partly explainedby the cib-mediated fitness conferred by the plasmid but mostlyby the high intrinsic efficiency of p2 transfer and by the capacityof the pathogen to trigger enteric inflammation. Inflammationelicits concomitant Salmonella and E. coli blooms, which canraise donor and acceptor densities to values >100-fold above
10μm
α E
. col
i O7
DN
A
1
10
100
Tran
scon
juga
ntfre
quen
cy (%
) **
α S
. Tm
O4,
5 D
NA
2
4
6
8
10
Log 10
cfu/
gce
cum
cont
ent
****
Inflamm.
S. Tmavir
--
Ec8178_cured
Donor
Recipient
Plasmid p2cm
+
CONLCM CON Microbiota
S. Tmavir S. Tmwt Donor
Recipient
Inflamm.
S. Tmavir
--
Ec8178_cured
Plasmid p2cm
+
Microbiota
S. Tmavir S. Tmwt
inflammation: nocomp. microbiota: no
inflammation: nocomp. microbiota: yes
inflammation: yescomp. microbiota: no
S. TmEc
CONLCM CON
A
C
B
Fig. 4. Inflammation boosts conjugative p2 transfer by increasing density ofdonors and recipients. (A) Streptomycin-treated low complexity microbiota(LCM; Left) or conventional (CON; Center and Right) mice were coinfectedwith 1:1 mixtures (total of 5 × 107; 1:1; intragastrically) of Ec8178_cured and S.Tmwt or S. Tmavir strains carrying p2cm, respectively. Densities of the S. Tmdonor and E. coli recipient strains were determined in the cecal contents atday 4 p.i. (B) The frequency of chloramphenicol-resistant E. coli trans-conjugants in cecal contents was determined by selective plating at day 4 p.i.Bars show the median. (C) Competing gut microbiota hinders bacterial contactas shown by immunofluorescent staining of Ec8178 and S. Tm in the cecal lu-men of a representative mouse of the experiment shown in a. Serial sectionsof PFA-fixed cecum tissue were stained with α-E. coli O7 (recolored blue) orα-S. Tm O4.5 antiserum (recolored magenta). Microbiota and host nuclearDNA was stained with Sytox green (recolored yellow). Arrows point at gran-ulocytes in the cecal lumen of mice with gut inflammation. (Scale bar: 10 μm.)
1272 | www.pnas.org/cgi/doi/10.1073/pnas.1113246109 Stecher et al.

those typically encountered in the normal mammalian intestine(14, 38). This fuels the reassortment of genetic material betweendifferent Enterobacteriaceae and suggests that infected patientsmight enhance the spread of plasmid-encoded fitness-, virulence-and antibiotic resistance-determinants (e.g., extended-spectrumβ-lactamases; ref. 39). Our findings shift the current paradigm ofthe “separate” evolution of pathogens (i.e., Salmonella) andcommensals (i.e., E. coli): Both are interconnected: Episodes ofpathogen-inflicted disease boost not only the evolution ofpathogens, but also that of the commensals. Our findings mightbe of central relevance for explaining the notorious emergence ofnew pathogenic and commensal strains with altered phenotypes.Finally, our data may add to the question of how plasmids are
maintained in bacterial populations over evolutionary time(“plasmid persistence puzzle”; see SI Text for detailed discussion)(40). Based on theoretical considerations, carriage of beneficialgenes is insufficient to account for plasmid maintenance. Due tofitness costs, plasmids should be lost and beneficial genes shouldmove to the chromosome. To explain the paradox of plasmidexistence, several mechanisms had been proposed: high-rateconjugative HGT (“infectious transmission”), plasmid shuttlingbetween bacterial strains, ecotypes or species (40) and the exis-tence of “amplifyer strains,” exhibiting extraordinarily high trans-conjugation efficiencies (41). However, so far, the evidencesupporting these hypotheses was mostly limited to theoreticalmodels and in vitro data. Our analysis of p2 transfer in theinfected gut provides evidence that “infectious transmission” aswell as plasmid shuttling between bacterial strains contribute toplasmid maintenance in nature. This must occur at very highfrequencies, as 4 of 10 independent strains analyzed in our initialexperiments had acquired p2 (Fig. S4). Thus, plasmid transfer inenterobacterial blooms comprising different bacterial species ismost likely a frequent event. Pathogenesis-inflicted blooms maybe an important feature of enterobacterial plasmid biology.
Materials and MethodsBacteria and Plasmids. Bacterial strains and plasmids used are listed in TableS2. For infections, E. coli and S. Tm strains were grown as described (42).Generation of bacterial mutants and plasmids, E. coli phylogenic microarray,genome sequencing, sequence comparisons and alignments, broad-rangebacterial 16S rRNA gene sequence analysis and plasmid isolation proceduresare described in SI Materials and Methods.
Isolation and Characterization of E. coli Strains From Frozen Tissue. Sections ofcecum tissues embedded in O.C.T. (Sakura) were prepared on a previously
sterilized cryotome. Sectionswere suspended in PBS and live bacteria recoveredby plating on MacConkey agar plates. Lactose-positive colonies were furthercharacterized using the Enterotube system (Becton Dickinson). ECOR typingwas performed using a well established triplex-PCR (43). MLST analysis wasdone as described by sequencing of the adk, fumC, gyrB, icd,mdh, purA, rEcA,and ST gene fragments and corresponding allocation to sequence types (STs).
Animal Experiments. All mice used in the study were on C57BL/6 backgroundand bred at the Rodent Center at the ETH Zürich under SPF conditions in in-dividually ventilated cages. LCM mice colonized with the Altered Schaedlerflora were described (26). Conventional, E. coli-free mice (CON) were initiallyobtained from Janvier, bred at the RCHCI and routinely tested for the absenceof E. coli in the feces. For S. Tm infections, mice were treated with strepto-mycin (20 mg per animal 24 h before Salmonella infection) and infectedby gavage with 5 × 107 cfu S. Tm or mixtures of S. Tm and E. coli as described.Live bacterial loads in the cecal content were determined by plating onMacConkey-agar (Oxoid) with respective antibiotics (streptomycin 100 μg/mL;kanamycin 30 μg/mL and chloramphenicol 30 μg/mL; ampicillin 100 μg/mL;tetracycline 12 μg/mL). Histology was done at necropsy. Briefly, tissues wereembedded in O.C.T. (Sakura) and flash frozen. H&E-stained cecum cryosectionswere scored as described: Evaluating submucosal edema, PMN infiltration,goblet cells, and epithelial damage yielded a total score of 0–13 points.
For induction of acute colitis, CD8+ T cells from CL4-TCR transgenic micethat express an α/βT cell receptor recognizing an epitope of the HA proteinpresented by MHC class I (the H-2Kd:HA512-520 complex) were adoptivelytransferred into VILLIN-HA mice that express the A/PR8/34 HA epitope frominfluenza virus A under control of the enterocyte-specific villin promoter (44).Single-cell suspensions were prepared from the spleen of CL4-TCR transgenicmice. Cell suspensions were depleted of CD4+, CD11b+, CD45R+, DX5+, andTer-119+ cells by using the MACS CD8 T cell isolation kit (Miltenyi Biotec).CL4-TCR T cells were purified by negative selection according to the manu-facturer’s instructions. Isolated CD8+ T cells were washed once in PBS andresuspended (4 × 107 cells per mL of PBS). Then, 4 × 106 purified CL4-TCRtransgenic T cells were injected i.v. into VILLIN-HA transgenic mice. Diseasesymptoms (weight loss and diarrhea) were observed 4–5 d after adoptivetransfer. Animal experiments were approved (licenses 201/2004 and 201/2007Kantonales Veterinäramt Zürich) and performed according to local guide-lines (TschV, Zurich) and the Swiss animal protection law (TschG).
Immunofluorescence. For detecting S. Tm and Ec8178 in the gut lumen in situ,cecal tissues were recovered and treated as described (45). Briefly, the tissuewas fixed in paraformaldehyde (4% in PBS, pH 7.4 overnight, 4 °C), washedwith PBS, equilibrated in PBS (20% sucrose, 0.1% NaN3 overnight, 4 °C), em-bedded in O.C.T. (Sakura), snap-frozen in liquid nitrogen and stored at −80 °C.Cryosections (7 μm) were air-dried for 2 h at room temperature, fixed in 4%paraformaldehyde (5 min), washed, and blocked in 10% (wt/vol) normal goatserum in PBS for 1 h. S. Tm and Ec8178 were detected by staining for 1 h witha polyclonal rabbit α-Salmonella-O-antigen group B serum (factors 4, 5;Difco; 1:300) or with a polyclonal rabbit α-E. coli-O7 serum (Statens SerumInstitut; 1:100) and DyLight-549-conjugated secondary goat-α-rabbit antibody(Jackson). DNA was stained with Sytox-green (1:10,000; Invitrogen). Sectionswere mounted with Vectashield hard set (Vector Laboratories) and analyzedusing a Leica SP5 confocal laser scanning microscope (Leica Microsystems).
Statistical Analysis. Statistical analysis was performed using the exact Mann-Whitney U test (Graphpad Prism Version 5.01). P values less than 0.05 (two-tailed) were considered statistically significant.
Colicin Assay. For measuring production of colicin and colicin sensitivity, thecolicin-producing strain was grown overnight as small spot (ø5 mm) on LBagar containing 2 μg/mL Mitomycin C (Sigma). The plate was overlaid withthe tester strain in Top-agar (0.75% agarose). Growth of the tester strainwas analyzed after 24 h. Halo formation around the producer indicatedsensitivity of the tester strain.
Mating Experiments. An overnight culture of donor and recipient strains wasgrown in LB media with appropriate antibiotics (37 °C, 180 rpm). The re-cipient strain was marked with antibiotic resistance by either carryingpWKS30 (46) (Amp) or pACYC184 (New England Biolabs; Cm). A total of 1 mLof each overnight culture was washed three times in PBS and concentrated(300 μL PBS final). Donor and recipient strains were mixed at a ratio of 1:1.Eighty microliters of the mixture was spread on LB Agar plate (surfacemating), and matings were incubated for 4 h at 37 °C. Bacteria were re-covered and plated onto respective selective agar to quantify trans-conjugant frequencies (cfu of plasmid-positive/total recipient cfu).
0.0000010.000010.00010.0010.010.1
110
100day 4 p.i.
2 3 4 5 6 7 8 9 10
Donor density (cfu/g Log10) Donor density (cfu/g Log10)
day 1 p.i.Tr
ansc
on. f
requ
ency
(%)
2 3 4 5 6 7 8 9 10
A B
Fig. 5. Intestinal density of S. Tm plasmid donors correlates with E. colitransconjugant frequency. Streptomycin-treated conventional mice (n = 20)were coinfected with mixtures of Ec8178_cured amp (recipient; ampR), S. Tmtet
p2Δcib (donor; tetR; plasmid p2: kanR) and increasing amounts of S. Tmwt
p2ΔoriTnikAΔcib (competitor; cmR) at no (n = 4), 10-fold (n = 3), 100-fold (n = 3),1,000-fold (n = 3), 10,000-fold (n = 3), and 100,000-fold (n = 4) excess over thedonor strain (total of 5 × 107; intragastrically). The density of the S. Tm donor(x axis) was plotted against E. coli transconjugant frequencies (fraction ofampR kanR E. coli of total ampR E. coli; y axis) as determined in the feces atday 1 p.i. (A) and cecal contents at day 4 p.i. (B). A linear relationship be-tween cfu and transconjugant frequency could be predicted by linear re-gression analysis (P < 0.05).
Stecher et al. PNAS | January 24, 2012 | vol. 109 | no. 4 | 1273
MICRO
BIOLO
GY

ACKNOWLEDGMENTS. We are grateful to Yvonne Lötscher (Zürich), ManuelDiehl, Lubov Nedialkova (Münich), and Barbara Plaschke (Würzburg) for ex-perimental support. We thank members of the W.-D.H. laboratory, SamBrown, and Mathias Heikenwälder for helpful comments and discussion.
Work was supported by UBS AG (Zürich) on behalf of a customer (W.-D.H.),Wellcome Trust Grant 076964 (to N.R.T., D.J.P., M.J.S., and A.W.W.), andDeutsche Forschungsgemeinschaft DFG SFB 479 TP A1 and DO 789/4-1 (toU.D.) and DFG STE 1971/2-1 (to B.S.).
1. Round JL, Mazmanian SK (2009) The gut microbiota shapes intestinal immune re-sponses during health and disease. Nat Rev Immunol 9:313–323.
2. Qin J, et al.; MetaHIT Consortium (2010) A human gut microbial gene catalogue es-tablished by metagenomic sequencing. Nature 464:59–65.
3. Arumugam M, et al. (2011) Enterotypes of the human gut microbiome. Nature 470:510–512.
4. Treangen TJ, Rocha EP (2011) Horizontal transfer, not duplication, drives the ex-pansion of protein families in prokaryotes. PLoS Genet 7:e1001284.
5. Hehemann JH, et al. (2009) Transfer of carbohydrate-active enzymes from marinebacteria to Japanese gut microbiota. (Translated from eng). Nature 464:908–912.
6. Nelson KE, et al.; Human Microbiome Jumpstart Reference Strains Consortium (2010)A catalog of reference genomes from the human microbiome. Science 328:994–999.
7. Paulsen IT, et al. (2003) Role of mobile DNA in the evolution of vancomycin-resistantEnterococcus faecalis. Science 299:2071–2074.
8. Johnson TJ, et al. (2010) Horizontal gene transfer of a ColV plasmid has resulted ina dominant avian clonal type of Salmonella enterica serovar Kentucky. PLoS ONE 5:e15524.
9. Bielaszewska M, et al. (2011) Characterisation of the Escherichia coli strain associatedwith an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbio-logical study. Lancet Infect Dis 11:671–676.
10. Tenaillon O, Skurnik D, Picard B, Denamur E (2010) The population genetics ofcommensal Escherichia coli. Nat Rev Microbiol 8:207–217.
11. Scott KP (2002) The role of conjugative transposons in spreading antibiotic resistancebetween bacteria that inhabit the gastrointestinal tract. Cell Mol Life Sci 59:2071–2082.
12. Bohnhoff M, Drake BL, Miller CP (1954) Effect of streptomycin on susceptibility ofintestinal tract to experimental Salmonella infection. Proc Soc Exp Biol NY 86:132–137.
13. van der Waaij D, Berghuis-de Vries JM, Lekkerkerk Lekkerkerk-v (1971) Colonizationresistance of the digestive tract in conventional and antibiotic-treated mice. J Hyg(Lond) 69:405–411.
14. Berg RD (1996) The indigenous gastrointestinal microflora. Trends Microbiol 4:430–435.
15. Licht TR, Christensen BB, Krogfelt KA, Molin S (1999) Plasmid transfer in the animalintestine and other dynamic bacterial populations: the role of community structureand environment. Microbiology 145:2615–2622.
16. Daniels JB, Call DR, Besser TE (2007) Molecular epidemiology of blaCMY-2 plasmidscarried by Salmonella enterica and Escherichia coli isolates from cattle in the PacificNorthwest. Appl Environ Microbiol 73:8005–8011.
17. Smet A, et al. (2011) In situ ESBL conjugation from avian to human Escherichia coliduring cefotaxime administration. J Appl Microbiol 110:541–549.
18. Moubareck C, Bourgeois N, Courvalin P, Doucet-Populaire F (2003) Multiple antibioticresistance gene transfer from animal to human enterococci in the digestive tract ofgnotobiotic mice. Antimicrob Agents Chemother 47:2993–2996.
19. Faure S, Perrin-Guyomard A, Delmas JM, Chatre P, Laurentie M (2010) Transfer ofplasmid-mediated CTX-M-9 from Salmonella enterica serotype Virchow to Enter-obacteriaceae in human flora-associated rats treated with cefixime. AntimicrobAgents Chemother 54:164–169.
20. Lupp C, et al. (2007) Host-mediated inflammation disrupts the intestinal microbiotaand promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2:119–129.
21. Stecher B, et al. (2007) Salmonella enterica serovar Typhimurium exploits in-flammation to compete with the intestinal microbiota. (Translated from Eng). PLoSBiol 5:e244.
22. Pédron T, Sansonetti P (2008) Commensals, bacterial pathogens and intestinal in-flammation: an intriguing ménage à trois. Cell Host Microbe 3:344–347.
23. Winter SE, et al. (2010) Gut inflammation provides a respiratory electron acceptor forSalmonella. Nature 467:426–429.
24. Darfeuille-Michaud A, et al. (2004) High prevalence of adherent-invasive Escherichiacoli associated with ileal mucosa in Crohn’s disease. Gastroenterology 127:412–421.
25. Barthel M, et al. (2003) Pretreatment of mice with streptomycin provides a Salmonellaenterica serovar Typhimurium colitis model that allows analysis of both pathogen andhost. Infect Immun 71:2839–2858.
26. Stecher B, et al. (2010) Like will to like: abundances of closely related species canpredict susceptibility to intestinal colonization by pathogenic and commensal bac-teria. PLoS Pathog 6:e1000711.
27. Brzuszkiewicz E, et al. (2006) How to become a uropathogen: comparative genomicanalysis of extraintestinal pathogenic Escherichia coli strains. Proc Natl Acad Sci USA103:12879–12884.
28. Welch RA, et al. (2002) Extensive mosaic structure revealed by the complete genomesequence of uropathogenic Escherichia coli. Proc Natl Acad Sci USA 99:17020–17024.
29. Kim SR, Komano T (1997) The plasmid R64 thin pilus identified as a type IV pilus. JBacteriol 179:3594–3603.
30. Komano T, Yoshida T, Narahara K, Furuya N (2000) The transfer region of IncI1plasmid R64: Similarities between R64 tra and legionella icm/dot genes.Mol Microbiol35:1348–1359.
31. Sampei G, et al. (2010) Complete genome sequence of the incompatibility group I1plasmid R64. Plasmid 64:92–103.
32. Komano T, Kubo A, Nisioka T (1987) Shufflon: Multi-inversion of four contiguous DNAsegments of plasmid R64 creates seven different open reading frames. Nucleic AcidsRes 15:1165–1172.
33. Cascales E, et al. (2007) Colicin biology. Microbiol Mol Biol Rev 71:158–229.34. Kerr B, Riley MA, Feldman MW, Bohannan BJ (2002) Local dispersal promotes bio-
diversity in a real-life game of rock-paper-scissors. Nature 418:171–174.35. Sonnenborn U, Schulze J (2009) The non-pathogenic Escherichia coli strain Nissle 1917 -
features of a versatile probiotic. Microb Ecol Health Dis 21:122–158.36. Hapfelmeier S, et al. (2005) The Salmonella pathogenicity island (SPI)-2 and SPI-1 type
III secretion systems allow Salmonella serovar typhimurium to trigger colitis viaMyD88-dependent and MyD88-independent mechanisms. J Immunol 174:1675–1685.
37. Dewhirst FE, et al. (1999) Phylogeny of the defined murine microbiota: AlteredSchaedler flora. Appl Environ Microbiol 65:3287–3292.
38. Stecher B, Hardt WD (2010) Mechanisms controlling pathogen colonization of thegut. Curr Opin Microbiol 14:82–91.
39. Pfeifer Y, Cullik A, Witte W (2010) Resistance to cephalosporins and carbapenems inGram-negative bacterial pathogens. Int J Med Microbiol 300:371–379.
40. Bergstrom CT, Lipsitch M, Levin BR (2000) Natural selection, infectious transfer andthe existence conditions for bacterial plasmids. Genetics 155:1505–1519.
41. Dionisio F, Matic I, Radman M, Rodrigues OR, Taddei F (2002) Plasmids spread veryfast in heterogeneous bacterial communities. Genetics 162:1525–1532.
42. Hapfelmeier S, et al. (2004) Role of the Salmonella pathogenicity island 1 effectorproteins SipA, SopB, SopE, and SopE2 in Salmonella enterica subspecies 1 serovarTyphimurium colitis in streptomycin-pretreated mice. Infect Immun 72:795–809.
43. Clermont O, Bonacorsi S, Bingen E (2000) Rapid and simple determination of theEscherichia coli phylogenetic group. Appl Environ Microbiol 66:4555–4558.
44. Westendorf AM, et al. (2006) Autoimmune-mediated intestinal inflammation-impactand regulation of antigen-specific CD8+ T cells. Gastroenterology 131:510–524.
45. Stecher B, et al. (2008) Motility allows S. Typhimurium to benefit from the mucosaldefence. Cell Microbiol 10:1166–1180.
46. Wang RF, Kushner SR (1991) Construction of versatile low-copy-number vectors forcloning, sequencing and gene expression in Escherichia coli. Gene 100:195–199.
1274 | www.pnas.org/cgi/doi/10.1073/pnas.1113246109 Stecher et al.


















