Glutamate and capsaicin effects on trigeminal nociception I: Activation and peripheral sensitization...
10
Research Report Glutamate and capsaicin effects on trigeminal nociception I: Activation and peripheral sensitization of deep craniofacial nociceptive afferents David K. Lam, Barry J. Sessle, James W. Hu ⁎ Faculty of Dentistry, University of Toronto 124 Edward Street, Toronto, Ontario, Canada M5G 1G6 ARTICLE INFO ABSTRACT Article history: Accepted 10 November 2008 Available online 19 November 2008 We have examined the effect of the peripheral application of glutamate and capsaicin to deep craniofacial tissues in influencing the activation and peripheral sensitization of deep craniofacial nociceptive afferents. The activity of single trigeminal nociceptive afferents with receptive fields in deep craniofacial tissues were recorded extracellularly in 55 halothane-anesthetized rats. The mechanical activation threshold (MAT) of each afferent was assessed before and after injection of 0.5 M glutamate (or vehicle) and 1% capsaicin (or vehicle) into the receptive field. A total of 68 afferents that could be activated by blunt noxious mechanical stimulation of the deep craniofacial tissues (23 masseter, 5 temporalis, 40 temporomandibular joint) were studied. When injected alone, glutamate and capsaicin activated and induced peripheral sensitization reflected as MAT reduction in many afferents. Following glutamate injection, capsaicin-evoked activity was greater than that evoked by capsaicin alone, whereas following capsaicin injection, glutamate-evoked responses were similar to glutamate alone. These findings indicate that peripheral application of glutamate or capsaicin may activate or induce peripheral sensitization in a subpopulation of trigeminal nociceptive afferents innervating deep craniofacial tissues, as reflected in changes in MAT and other afferent response properties. The data further suggest that peripheral glutamate and capsaicin receptor mechanisms may interact to modulate the activation and peripheral sensitization in some deep craniofacial nociceptive afferents. Crown Copyright © 2008 Published by Elsevier B.V. All rights reserved. Keywords: Excitatory amino acid TRPV1 Pain Temporomandibular joint 1. Introduction There is emerging evidence that both peripheral excitatory amino acid (EAA) and vanilloid (TRPV1) receptor mechanisms may modulate nociceptive processing of input from deep craniofacial tissues. Glutamate, the endogenous agonist for EAA receptors, is a well-documented central excitatory neurotransmitter but a number of studies indicate a role for peripheral glutamate receptors in the transduction of nocicep- tive information (Yu et al. 1996; Lawand et al. 2000; McNearney et al. 2000, 2004; Carlton 2001; Carlton et al. 2003; Lam et al. 2005b). We have identified a novel peripheral nociceptive role for glutamate in the craniofacial region. Intramuscular (masseter) or temporomandibular joint (TMJ) injection of glutamate reflexly evokes a dose-dependent increase in rat jaw muscle activity (Cairns et al. 1998), activates and sensitizes mechanosensitive nociceptive afferents through the activation of peripheral EAA receptors (Cairns et al. 2001a, BRAIN RESEARCH 1251 (2009) 130 – 139 ⁎ Corresponding author. Fax: +1 416 979 4936. E-mail address: [email protected] (J.W. Hu). 0006-8993/$ – see front matter. Crown Copyright © 2008 Published by Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2008.11.029 available at www.sciencedirect.com www.elsevier.com/locate/brainres
-
Upload
david-k-lam -
Category
Documents
-
view
222 -
download
1
Transcript of Glutamate and capsaicin effects on trigeminal nociception I: Activation and peripheral sensitization...

B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res
Research Report
Glutamate and capsaicin effects on trigeminal nociception I:Activation and peripheral sensitization of deep craniofacialnociceptive afferents
David K. Lam, Barry J. Sessle, James W. Hu⁎
Faculty of Dentistry, University of Toronto 124 Edward Street, Toronto, Ontario, Canada M5G 1G6
A R T I C L E I N F O
⁎ Corresponding author. Fax: +1 416 979 4936.E-mail address: [email protected] (J.W
0006-8993/$ – see front matter. Crown Copyrdoi:10.1016/j.brainres.2008.11.029
A B S T R A C T
Article history:Accepted 10 November 2008Available online 19 November 2008
We have examined the effect of the peripheral application of glutamate and capsaicin todeep craniofacial tissues in influencing the activation and peripheral sensitization of deepcraniofacial nociceptive afferents. The activity of single trigeminal nociceptive afferentswith receptive fields in deep craniofacial tissues were recorded extracellularly in 55halothane-anesthetized rats. The mechanical activation threshold (MAT) of each afferentwas assessed before and after injection of 0.5 M glutamate (or vehicle) and 1% capsaicin (orvehicle) into the receptive field. A total of 68 afferents that could be activated by bluntnoxious mechanical stimulation of the deep craniofacial tissues (23 masseter, 5 temporalis,40 temporomandibular joint) were studied. When injected alone, glutamate and capsaicinactivated and induced peripheral sensitization reflected as MAT reduction in manyafferents. Following glutamate injection, capsaicin-evoked activity was greater than thatevoked by capsaicin alone, whereas following capsaicin injection, glutamate-evokedresponses were similar to glutamate alone. These findings indicate that peripheralapplication of glutamate or capsaicin may activate or induce peripheral sensitization in asubpopulation of trigeminal nociceptive afferents innervating deep craniofacial tissues, asreflected in changes in MAT and other afferent response properties. The data furthersuggest that peripheral glutamate and capsaicin receptor mechanisms may interact tomodulate the activation and peripheral sensitization in some deep craniofacial nociceptiveafferents.
Crown Copyright © 2008 Published by Elsevier B.V. All rights reserved.
Keywords:Excitatory amino acidTRPV1PainTemporomandibular joint
1. Introduction
There is emerging evidence that both peripheral excitatoryamino acid (EAA) and vanilloid (TRPV1) receptor mechanismsmay modulate nociceptive processing of input from deepcraniofacial tissues. Glutamate, the endogenous agonist forEAA receptors, is a well-documented central excitatoryneurotransmitter but a number of studies indicate a role forperipheral glutamate receptors in the transduction of nocicep-
. Hu).
ight © 2008 Published by
tive information (Yu et al. 1996; Lawand et al. 2000; McNearneyet al. 2000, 2004; Carlton 2001; Carlton et al. 2003; Lam et al.2005b). We have identified a novel peripheral nociceptive rolefor glutamate in the craniofacial region. Intramuscular(masseter) or temporomandibular joint (TMJ) injection ofglutamate reflexly evokes a dose-dependent increase in ratjaw muscle activity (Cairns et al. 1998), activates andsensitizes mechanosensitive nociceptive afferents throughthe activation of peripheral EAA receptors (Cairns et al. 2001a,
Elsevier B.V. All rights reserved.

Table 1 –Mechanoreceptive field (RF) and baseline meanpooledorthodromic conductionvelocity (CV)andmechanicalactivation threshold (MAT) of deep craniofacial nociceptiveafferents
RF location Aδ-fiber afferent C-fiber afferentMean±SE Mean±SE
n MeanCV (m/s)
MeanMAT (g)
n MeanCV (m/s)
MeanMAT (g)
TMJ 31 9.8±1.1 29.6±3.5 9 1.6±0.1 30.0±8.8Masseter 19 8.8±1.2 27.6±5.3 4 1.7±0.3 21.1±17.1Temporalis 4 16.9±1.9 24.7±9.9 1 1.7 4.9Pooled 54 9.8±0.8 28.5±2.8 14 1.6±0.1 24.9±7.2
131B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
b, 2002, 2003a,b; Dong et al. 2006, 2007), and activates andinduces central sensitization of brainstem nociceptive neu-rons in the trigeminal subnucleus caudalis/upper cervical cord(Vc/UCC) (Lam et al. 2008). The peripheral EAA receptor, N-methyl-D-aspartate (NMDA), in particular may play an impor-tant role in glutamate-induced effects on nociceptive afferentssince NMDA receptor antagonists applied locally into masti-catorymuscles significantly decrease glutamate-evoked affer-ent activation and sensitization as well as jaw muscle activity(Cairns et al. 1998, 2003a, 2007; Dong et al. 2006, 2007).Similarly, glutamate injection into the human massetermuscle causes pain that may be attenuated by co-injectionof an NMDA receptor antagonist (Cairns et al. 2001a, 2003a,b,2006; Svensson et al. 2003, 2005).
Another peripheral receptor involved in craniofacial noci-ceptive mechanisms is the TRPV1 receptor. This receptor isactivated by application to peripheral tissues of the inflam-matory irritant capsaicin, protons or noxious heat (Caterina etal. 1997; Tominaga et al. 1998; Dray 2005). Capsaicin canactivate (Liu and Simon 1994, 1996, 2003) and sensitizetrigeminal afferents (Strassman et al. 1996) and also activatebrainstem nociceptive neurons (Carstens et al. 1998; Zanottoet al. 2007). Furthermore, capsaicin injection into the rat TMJreflexly evokes a dose-dependent increase in jaw muscleactivity (Tang et al. 2004; Lam et al. 2005a), activates andinduces central sensitization of brainstem nociceptive neu-rons in the Vc/UCC (Lam et al. 2008), and in addition producesan inflammatory response within these tissues that can beblocked by TRPV1 receptor antagonists (Hu et al. 2005a).Capsaicin injected into human craniofacial regions can alsoinduce secondary hyperalgesia, allodynia and jawmuscle painassociated with changes in jaw motor function (Sohn et al.2000, 2004; Wang et al. 2002; Gazerani and Arendt-Nielsen2005; Gazerani et al. 2006).
However, there has been little investigation of possibleinteractions between peripheral EAA and TRPV1 receptors andtheir effects on nociceptive processing. There is evidence thatEAA receptors may indirectly modulate nociception viainteractions with TRPV1 in the central nervous system. Bothcapsaicin-evoked release of spinal substance P (Afrah et al.2001) and capsaicin-evoked activity at the periaqueductal greylevel (Palazzo et al. 2002; Xing and Li 2006) may be dependenton glutamate-evoked activation of NMDA receptors. A role forNMDA receptor modulation of capsaicin-induced c-fos expres-sion within the rat Vc has also been demonstrated (Mitsikos-tas et al. 1998). Similarly, the presence of EAA and TRPV1receptors on primary afferents and findings that their agonistshave effects when injected into peripheral tissues raise thepossibility that the actions of peripherally applied glutamateand capsaicin could involve interactions between EAA andTRPV1 receptors in these tissues. Indeed, jaw muscle activityreflexly evoked by the TMJ application of capsaicin can beattenuated by pre-injection into the TMJ of NMDA receptorantagonists (Lam et al. 2005a), suggesting that peripheral EAAreceptors may interact with peripheral TRPV1 receptormechanisms. While it is also well known that capsaicin cansensitize or desensitize nociceptive primary afferents (Bau-mann et al. 1991; Craft and Porreca 1992; LaMotte et al. 1992;Liu and Simon 1996; Sikand and Premkumar 2007), it is notknown whether glutamate or capsaicin would sensitize or
desensitize each other's effects on afferent responses. Thus,the aim of the present study was to examine the effect of theperipheral application of glutamate and capsaicin in deepcraniofacial tissues in influencing the activation and periph-eral sensitization of deep craniofacial nociceptive primaryafferents. The data have been briefly presented in abstractform (Lam et al. 2004a,b).
2. Results
2.1. Neuronal properties
A total of 68 trigeminal nociceptive primary afferents withreceptive fields (RFs) in deep craniofacial tissues wererecorded: 54 Aδ-fiber and 14 C-fiber afferents. The estimatedantidromic (Aδ-fiber: mean±SE= 10.2±0.9 m/s, n=48; C-fiber:1.7±0.2 m/s, n=12) and orthodromic (Aδ-fiber: 9.8±0.8 m/s,n=54; C-fiber: 1.6±0.1 m/s, n=14) conduction velocities (CVs)were not statistically different (pN0.05, Mann–Whitney U test).The majority of afferents could be activated antidromicallyand orthodromically (60/68; 48 Aδ-fiber, 12 C-fiber); theremainder were activated only orthodromically (8/68; 6 Aδ-fiber, 2 C-fiber). A small proportion (10%, 7/68, 6 Aδ-fiber, 1 C-fiber) were spontaneously active (0.38±0.12 spikes/second)prior to injection of receptor agonists. Table 1 displays thebaseline mean CV andmechanical activation threshold (MAT)for afferents with RFs in the TMJ (n=40), masseter (n=23) andtemporalis (n=5).
There were no differences between the Aδ-fiber and C-fiberafferents in baseline MAT (pN0.05, t-test) (Table 1) or inresponses (Rmag, Rdur, Rlat and Pfreq) to glutamate andcapsaicin (pN0.05, Mann–Whitney U test; pN0.05, Fisher exacttest) (data not shown) and as a result, the data were pooledtogether for analysis of glutamate and capsaicin-inducedactivation and sensitization. Examples of typical afferent RFand response properties are shown in Figs. 1A and B.
2.2. Glutamate effects
2.2.1. ActivationInjection of glutamate alone (but not vehicle alone, 0/4)activated 43% (12/28) of the afferents tested (9/23 Aδ-fibers,CV=8.2±1.5 m/s; 3/5 C-fibers, CV=1.6±0.3 m/s) (e.g. Fig. 1B)(p<0.001, Fisher exact test), with the activation response

Fig. 1 – (A) Example of typical mechanoreceptive field and response properties of deep craniofacial nociceptive afferent toinjection of glutamate vehicle followed by capsaicin (CV=2.8m/s, MAT=38 g, Aδ-fiber TMJ afferent). (a) Deepmechanoreceptivefield (white circle) of nociceptive afferent involving the TMJ region indicated by arrow, (b) Afferent mechanical activationthreshold determined with von Frey device, (c) No response in this afferent was evoked by injection of glutamate vehicle intothe TMJ, (d) Afferent response evoked by injection of capsaicin following glutamate vehicle into the TMJ. (B) Example of typicalmechanoreceptive field and response properties of deep craniofacial nociceptive afferent to injection of glutamate followed bycapsaicin (CV=2.0 m/s, MAT=12.7 g, C-fiber TMJ afferent). (a) Deep mechanoreceptive field (white circle) of nociceptive afferentinvolving the TMJ region indicated by arrow, (b) Afferent mechanical activation threshold determined with von Frey device,(c) Afferent response evoked by injection of glutamate into the TMJ, (d) Afferent response evoked by injection of capsaicinfollowing glutamate into the TMJ, (e) Stimulation of Vc/UCC (50 μs, 50 μA, 100 Hz) evoked an antidromic action potential(latency: 6.0 ms). By measuring the distance between the recording electrode and the stimulating electrode in Vc/UCC anddividing by the antidromic latency, the CV of this afferent was estimated to be 2.0 m/s, (f) Blunt mechanical stimulation of theTMJ tissuewas used to evoke orthodromic spikes that served as a trigger for electrical stimulation of Vc/UCC (antidromic spike).Shortening the delay between the orthodromically evoked spike and the electrical stimulus applied to Vc/UCC resulted in acollision, as evidenced by the disappearance of the antidromic spike*.
132 B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
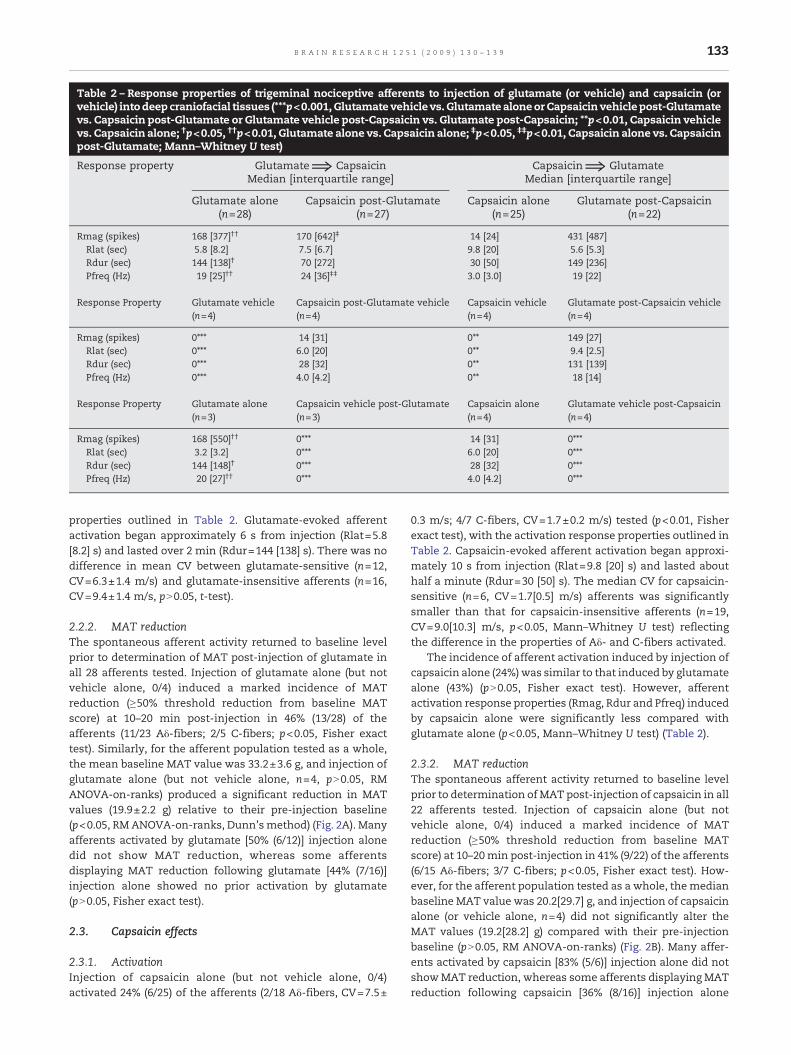
Table 2 – Response properties of trigeminal nociceptive afferents to injection of glutamate (or vehicle) and capsaicin (orvehicle) intodeep craniofacial tissues (***p<0.001, Glutamate vehicle vs. Glutamate aloneor Capsaicin vehiclepost-Glutamatevs. Capsaicin post-Glutamate or Glutamate vehicle post-Capsaicin vs. Glutamate post-Capsaicin; **p<0.01, Capsaicin vehiclevs. Capsaicin alone; †p<0.05, ††p<0.01, Glutamate alone vs. Capsaicin alone; ‡p<0.05, ‡‡p<0.01, Capsaicin alone vs. Capsaicinpost-Glutamate; Mann–Whitney U test)
Response property Glutamate⇒ Capsaicin Capsaicin⇒ GlutamateMedian [interquartile range] Median [interquartile range]
Glutamate alone(n=28)
Capsaicin post-Glutamate(n=27)
Capsaicin alone(n=25)
Glutamate post-Capsaicin(n=22)
Rmag (spikes) 168 [377]†† 170 [642]‡ 14 [24] 431 [487]Rlat (sec) 5.8 [8.2] 7.5 [6.7] 9.8 [20] 5.6 [5.3]Rdur (sec) 144 [138]† 70 [272] 30 [50] 149 [236]Pfreq (Hz) 19 [25]†† 24 [36]‡‡ 3.0 [3.0] 19 [22]
Response Property Glutamate vehicle(n=4)
Capsaicin post-Glutamate vehicle(n=4)
Capsaicin vehicle(n=4)
Glutamate post-Capsaicin vehicle(n=4)
Rmag (spikes) 0⁎⁎⁎ 14 [31] 0⁎⁎ 149 [27]Rlat (sec) 0⁎⁎⁎ 6.0 [20] 0⁎⁎ 9.4 [2.5]Rdur (sec) 0⁎⁎⁎ 28 [32] 0⁎⁎ 131 [139]Pfreq (Hz) 0⁎⁎⁎ 4.0 [4.2] 0⁎⁎ 18 [14]
Response Property Glutamate alone(n=3)
Capsaicin vehicle post-Glutamate(n=3)
Capsaicin alone(n=4)
Glutamate vehicle post-Capsaicin(n=4)
Rmag (spikes) 168 [550]†† 0⁎⁎⁎ 14 [31] 0⁎⁎⁎Rlat (sec) 3.2 [3.2] 0⁎⁎⁎ 6.0 [20] 0⁎⁎⁎Rdur (sec) 144 [148]† 0⁎⁎⁎ 28 [32] 0⁎⁎⁎Pfreq (Hz) 20 [27]†† 0⁎⁎⁎ 4.0 [4.2] 0⁎⁎⁎
133B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
properties outlined in Table 2. Glutamate-evoked afferentactivation began approximately 6 s from injection (Rlat=5.8[8.2] s) and lasted over 2 min (Rdur=144 [138] s). There was nodifference in mean CV between glutamate-sensitive (n=12,CV=6.3±1.4 m/s) and glutamate-insensitive afferents (n=16,CV=9.4±1.4 m/s, pN0.05, t-test).
2.2.2. MAT reductionThe spontaneous afferent activity returned to baseline levelprior to determination of MAT post-injection of glutamate inall 28 afferents tested. Injection of glutamate alone (but notvehicle alone, 0/4) induced a marked incidence of MATreduction (≥50% threshold reduction from baseline MATscore) at 10–20 min post-injection in 46% (13/28) of theafferents (11/23 Aδ-fibers; 2/5 C-fibers; p<0.05, Fisher exacttest). Similarly, for the afferent population tested as a whole,the mean baseline MAT value was 33.2±3.6 g, and injection ofglutamate alone (but not vehicle alone, n=4, pN0.05, RMANOVA-on-ranks) produced a significant reduction in MATvalues (19.9±2.2 g) relative to their pre-injection baseline(p<0.05, RMANOVA-on-ranks, Dunn'smethod) (Fig. 2A). Manyafferents activated by glutamate [50% (6/12)] injection alonedid not show MAT reduction, whereas some afferentsdisplaying MAT reduction following glutamate [44% (7/16)]injection alone showed no prior activation by glutamate(pN0.05, Fisher exact test).
2.3. Capsaicin effects
2.3.1. ActivationInjection of capsaicin alone (but not vehicle alone, 0/4)activated 24% (6/25) of the afferents (2/18 Aδ-fibers, CV=7.5±
0.3 m/s; 4/7 C-fibers, CV=1.7±0.2 m/s) tested (p<0.01, Fisherexact test), with the activation response properties outlined inTable 2. Capsaicin-evoked afferent activation began approxi-mately 10 s from injection (Rlat=9.8 [20] s) and lasted abouthalf a minute (Rdur=30 [50] s). The median CV for capsaicin-sensitive (n=6, CV=1.7[0.5] m/s) afferents was significantlysmaller than that for capsaicin-insensitive afferents (n=19,CV=9.0[10.3] m/s, p<0.05, Mann–Whitney U test) reflectingthe difference in the properties of Aδ- and C-fibers activated.
The incidence of afferent activation induced by injection ofcapsaicin alone (24%)was similar to that induced by glutamatealone (43%) (pN0.05, Fisher exact test). However, afferentactivation response properties (Rmag, Rdur and Pfreq) inducedby capsaicin alone were significantly less compared withglutamate alone (p<0.05, Mann–Whitney U test) (Table 2).
2.3.2. MAT reductionThe spontaneous afferent activity returned to baseline levelprior to determination of MAT post-injection of capsaicin in all22 afferents tested. Injection of capsaicin alone (but notvehicle alone, 0/4) induced a marked incidence of MATreduction (≥50% threshold reduction from baseline MATscore) at 10–20min post-injection in 41% (9/22) of the afferents(6/15 Aδ-fibers; 3/7 C-fibers; p<0.05, Fisher exact test). How-ever, for the afferent population tested as a whole, themedianbaseline MAT value was 20.2[29.7] g, and injection of capsaicinalone (or vehicle alone, n=4) did not significantly alter theMAT values (19.2[28.2] g) compared with their pre-injectionbaseline (pN0.05, RM ANOVA-on-ranks) (Fig. 2B). Many affer-ents activated by capsaicin [83% (5/6)] injection alone did notshowMAT reduction, whereas some afferents displaying MATreduction following capsaicin [36% (8/16)] injection alone
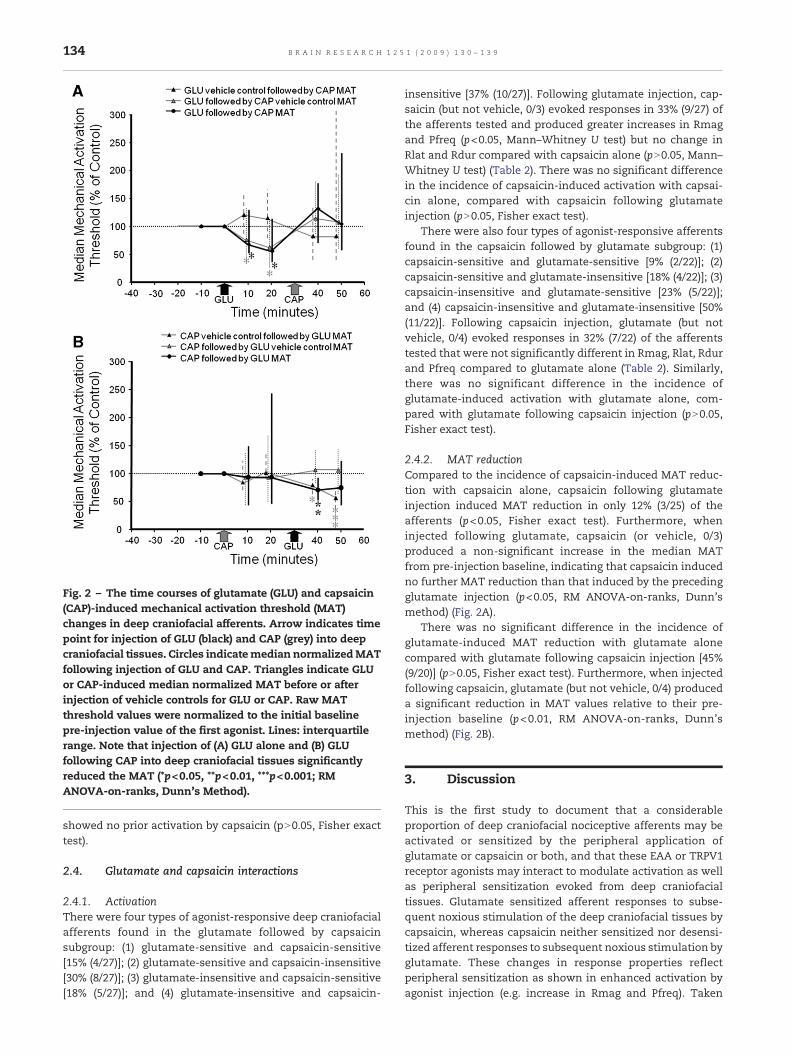
Fig. 2 – The time courses of glutamate (GLU) and capsaicin(CAP)-induced mechanical activation threshold (MAT)changes in deep craniofacial afferents. Arrow indicates timepoint for injection of GLU (black) and CAP (grey) into deepcraniofacial tissues. Circles indicatemedian normalizedMATfollowing injection of GLU and CAP. Triangles indicate GLUor CAP-induced median normalized MAT before or afterinjection of vehicle controls for GLU or CAP. Raw MATthreshold values were normalized to the initial baselinepre-injection value of the first agonist. Lines: interquartilerange. Note that injection of (A) GLU alone and (B) GLUfollowing CAP into deep craniofacial tissues significantlyreduced the MAT (*p<0.05, **p<0.01, ***p<0.001; RMANOVA-on-ranks, Dunn's Method).
134 B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
showed no prior activation by capsaicin (pN0.05, Fisher exacttest).
2.4. Glutamate and capsaicin interactions
2.4.1. ActivationThere were four types of agonist-responsive deep craniofacialafferents found in the glutamate followed by capsaicinsubgroup: (1) glutamate-sensitive and capsaicin-sensitive[15% (4/27)]; (2) glutamate-sensitive and capsaicin-insensitive[30% (8/27)]; (3) glutamate-insensitive and capsaicin-sensitive[18% (5/27)]; and (4) glutamate-insensitive and capsaicin-
insensitive [37% (10/27)]. Following glutamate injection, cap-saicin (but not vehicle, 0/3) evoked responses in 33% (9/27) ofthe afferents tested and produced greater increases in Rmagand Pfreq (p<0.05, Mann–Whitney U test) but no change inRlat and Rdur compared with capsaicin alone (pN0.05, Mann–Whitney U test) (Table 2). There was no significant differencein the incidence of capsaicin-induced activation with capsai-cin alone, compared with capsaicin following glutamateinjection (pN0.05, Fisher exact test).
There were also four types of agonist-responsive afferentsfound in the capsaicin followed by glutamate subgroup: (1)capsaicin-sensitive and glutamate-sensitive [9% (2/22)]; (2)capsaicin-sensitive and glutamate-insensitive [18% (4/22)]; (3)capsaicin-insensitive and glutamate-sensitive [23% (5/22)];and (4) capsaicin-insensitive and glutamate-insensitive [50%(11/22)]. Following capsaicin injection, glutamate (but notvehicle, 0/4) evoked responses in 32% (7/22) of the afferentstested that were not significantly different in Rmag, Rlat, Rdurand Pfreq compared to glutamate alone (Table 2). Similarly,there was no significant difference in the incidence ofglutamate-induced activation with glutamate alone, com-pared with glutamate following capsaicin injection (pN0.05,Fisher exact test).
2.4.2. MAT reductionCompared to the incidence of capsaicin-induced MAT reduc-tion with capsaicin alone, capsaicin following glutamateinjection induced MAT reduction in only 12% (3/25) of theafferents (p<0.05, Fisher exact test). Furthermore, wheninjected following glutamate, capsaicin (or vehicle, 0/3)produced a non-significant increase in the median MATfrom pre-injection baseline, indicating that capsaicin inducedno further MAT reduction than that induced by the precedingglutamate injection (p<0.05, RM ANOVA-on-ranks, Dunn'smethod) (Fig. 2A).
There was no significant difference in the incidence ofglutamate-induced MAT reduction with glutamate alonecompared with glutamate following capsaicin injection [45%(9/20)] (pN0.05, Fisher exact test). Furthermore, when injectedfollowing capsaicin, glutamate (but not vehicle, 0/4) produceda significant reduction in MAT values relative to their pre-injection baseline (p<0.01, RM ANOVA-on-ranks, Dunn'smethod) (Fig. 2B).
3. Discussion
This is the first study to document that a considerableproportion of deep craniofacial nociceptive afferents may beactivated or sensitized by the peripheral application ofglutamate or capsaicin or both, and that these EAA or TRPV1receptor agonists may interact to modulate activation as wellas peripheral sensitization evoked from deep craniofacialtissues. Glutamate sensitized afferent responses to subse-quent noxious stimulation of the deep craniofacial tissues bycapsaicin, whereas capsaicin neither sensitized nor desensi-tized afferent responses to subsequent noxious stimulation byglutamate. These changes in response properties reflectperipheral sensitization as shown in enhanced activation byagonist injection (e.g. increase in Rmag and Pfreq). Taken

135B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
together, these findings suggest that both peripheral EAA andTRPV1 receptor mechanisms may be involved in the nocicep-tive processing of deep craniofacial nociceptive afferents andmay interact to modulate the activation and peripheralsensitization in some nociceptive afferents supplying deepcraniofacial tissues.
Properties of deep craniofacial afferents projecting to Vc/UCC and their CVs are consistent with previous findings(Cairns et al. 2001a,b, 2002, 2003a; Dong et al. 2006, 2007). Ourfindings of glutamate-induced activation and peripheralsensitization in trigeminal nociceptive afferents are consis-tent with previous evidence of glutamate-evoked dose-dependent increases in jaw muscle activity, activation andsensitization of deep nociceptive afferents, and pain in humanmasticatory muscles (Cairns et al. 1998, 2001a,b, 2002, 2003a,b,2006, 2007; Svensson et al. 2003, 2005; Dong et al. 2006, 2007).Likewise, the incidence and discharge pattern of capsaicin-induced trigeminal afferent activation in the present study areconsistent with those found in mechanosensitive spinal(Baumann et al. 1991; LaMotte et al. 1992) and trigeminal(Strassman et al. 1996; Ikeda et al. 1997) afferents in previousstudies. In general, capsaicin tended to primarily activate veryslowly conducting afferents and the discharge was irregularand the majority of afferents ceased discharging within thefirst 60 s. Thus, it seems that the majority of trigeminalnociceptive afferents with deep craniofacial RFs in the presentstudy responded too weakly or transiently to be able toaccount for the magnitude and duration of capsaicin-inducedcraniofacial pain in humans (Sohn et al. 2000, 2004;Wang et al.2002; Gazerani and Arendt-Nielsen 2005; Gazerani et al. 2006).When injected alone, the capsaicin-induced MAT reductionwas not as robust as that induced by glutamate alone. Whilethe non-significant reduction in MAT for the afferent popula-tion as a whole is consistent with previous studies suggestingcapsaicin-evoked mechanical sensitization is due to centralrather than peripheral sensitization (Baumann et al. 1991;LaMotte et al. 1992), this non-significant capsaicin-evokedMAT reduction may be due, in part, to the lower afferentresponses (Rmag, Rdur and Pfreq) evoked by capsaicin alonecompared to glutamate alone. Nevertheless, the significantincidence of MAT reduction induced by capsaicin alonedemonstrates that some trigeminal nociceptive afferents canbe readily sensitized by the peripheral application of capsaicinand these findings are consistent with our previous evidenceof capsaicin-evoked dose-dependent increases in jaw muscleactivity (Tang et al. 2004). Taken together, the above findingsindicate that peripheral TRPV1 receptors in addition toperipheral EAA receptors in deep craniofacial tissues mayplay an important role in nociceptive processing.
The finding that glutamate or capsaicin may induce MATreduction without prior afferent activation may be explainedby glutamate and capsaicin acting on peripheral non-neuronalcells in addition to neuronal cells and causing them viaparacrine activation to release mediators that sensitize thedeep craniofacial afferents (Lam et al. 2005a,b). There has beenrecent compelling evidence for the expression and function ofglutamate and capsaicin in signaling processes in severaltypes of non-neuronal cells (Meddings et al. 1991; Skerry andGenever 2001; Kato et al. 2003; Rizvi and Luqman 2003; Li et al.2005; Xin et al. 2005). Thus it may be possible that the
activation of glutamate and capsaicin receptors on non-neuronal cells may result in the release of various othersensitizers including bradykinin, amines, prostanoids, growthfactors, chemokines, cytokines, protons and ATP (Sikand andPremkumar 2007; Woolf and Ma, 2007) that may sensitize thedeep craniofacial afferents.
3.1. Glutamate and capsaicin receptor interactions in deepcraniofacial tissues
Capsaicin-evoked activation of trigeminal nociceptive affer-ents following glutamate injection were significantlyenhanced compared to capsaicin alone, suggesting thatglutamate may sensitize the nociceptive afferents and pro-ducemore immediate (e.g. decrease Rlat), larger (e.g. increasedRmag and Pfreq) and more prolonged (e.g. increased Rdur)responses to subsequent noxious stimuli (e.g. to capsaicin).These results contrast with those in a recent rat behavioralmodel in which masseteric injection of glutamate andcapsaicin, in alternating order 10 min apart, evoked compar-able nocifensive responses regardless of injection sequence(Ro and Capra 2006) but are consistent with findings that pre-injection of NMDA receptor antagonists into the TMJ regionattenuates jawmuscle activity evoked by capsaicin (Lam et al.2005a). A protein that is likely to mediate the interactionsbetween peripheral NMDA and TRPV1 receptors is the Ca2+-calmodulin-dependent kinase II (CaMKII) which is persistentlyactivated after NMDA receptor stimulation (see Yamakura andShimoji 1999; Petrenko et al. 2003; Paoletti and Neyton 2007)and phosphorylation of TRPV1 by CaMKII is required for itsligand binding (Jung et al. 2004; Suh and Oh 2005; Tominagaand Tominaga 2005). Although no studies to date havedemonstrated the co-localization of peripheral NMDA andTRPV1 receptors on the same trigeminal primary afferentterminal, nociceptive responses could be enhanced if thesame nociceptive afferent expresses both EAA and TRPV1receptors. Taken together, these findings suggest that theactivation and/or sensitization of peripheral EAA receptorsmay be important in the mechanisms whereby capsaicin viaTRPV1 receptors evoke nociceptive trigeminal responses.
In addition to possible interactions between ionotropicreceptors, there is evidence of a major coupling between G-protein-coupled receptors and some TRP channels in themembrane such as TRPA1 and TRPV1 (Sikand and Premkumar2007;Woolf andMa 2007). For example, TRPA1may function asa receptor-operated channel for bradykinin by allowing Ca2+
influx following activation of the B2 receptor (Bautista et al.2006). Bradykinin can also significantly potentiate TRPV1activity by activating the Ca2+/phospholipid-dependent kinase(PKC) pathway (Sikand and Premkumar 2007). Activation of thePKC pathway has also been shown to lower the heat thresholdof TRPV1 below body temperature and sensitize TRPV1receptor responses to capsaicin (Premkumar and Ahern 2000;Crandall et al. 2002). The mechanism behind this effect isthought to involve direct phosphorylation resulting in PKC-dependent insertion of TRPV1 receptors into the neuronalmembrane (for review, see Hucho and Levine 2007). This typeof couplingmayalso exist betweenTRPV1andotherG-protein-coupled receptors (Sikand and Premkumar 2007;Woolf andMa2007), such as themetabotropic EAA receptors, and provide an

136 B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
additionalmeans for interactions between peripheral EAA andTRPV1 receptors.
Peripheral glutamate receptor mechanisms may not onlymodulate capsaicin-evoked activity but they may also mod-ulate capsaicin-induced MAT reduction. Capsaicin neithersensitized nor desensitized glutamate-induced MATresponses since glutamate-induced MAT reduction in thenociceptive afferents following capsaicin injection wereunchanged compared to glutamate alone. However, thecapsaicin-induced median MAT following glutamate injectionwas increased and the incidence of MAT reduction wassignificantly lower compared to capsaicin alone. This findingrepresents an apparent paradox where glutamate maysensitize a trigeminal afferent to subsequently producegreater capsaicin-evoked activity yet at the same time, itmay also result in an attenuation of capsaicin-induced MATreduction. One possible explanation for this paradox is thatthe greater capsaicin-evoked afferent activity following glu-tamate may cause an excess influx of calcium ions into theafferent and result in desensitization of the trigeminalnociceptive afferent to mechanical stimuli. This finding isconsistent with previous studies in which capsaicin mayeither sensitize or desensitize spinal (Baumann et al. 1991;LaMotte et al. 1992; Simone et al. 1997; Serra et al. 2004) ortrigeminal (Liu and Simon 1996; Strassman et al. 1996)nociceptive afferents to mechanical stimuli (i.e. sensitizationwith low concentrations and desensitization with high con-centrations). Another possibility is that the trigeminal noci-ceptive afferents may be resistant to the subsequentsensitizing effects of capsaicin application due to a possibleceiling effect from the prior glutamate-induced peripheralsensitization. That is, the nociceptive afferents may havereached theirmaximal reductions inMAT following glutamateinjection such that they cannot be sensitized further bycapsaicin activation 30 min post-glutamate injection.
4. Experimental procedures
4.1. Animal preparation
Adult male (n=55, 250–400 grams (g)) Sprague–Dawley ratswere prepared for acute in vivo recording activity of trigeminalafferents under surgical anesthesia (O2: 1 L/min; halothane:1.5–2.5%; Cairns et al. 2001a,b). A tracheal cannula wasinserted and artificial ventilation initiated. The rat's headwas then placed in a stereotaxic frame and the skin over thedorsal surface of the skull was reflected. A trephination wasmade on the left side of the skull to allow a recordingmicroelectrode to be lowered through the brain and into thetrigeminal ganglion. An incision was also made in the skinoverlying the Vc/UCC region, a C1 laminectomy was per-formed, and the dura was reflected to expose the Vc/UCC andfacilitate placement of a stimulating microelectrode in the leftVc/UCC (Cairns et al. 2001a,b; Hu et al. 2005b).
After completion of all surgical procedures, the halothanelevel was slowly reduced to a level (1–1.3%) that was justsufficient to produce reflex suppression of the hindlimb tonoxious pressure applied to the hindpaw to ensure that anadequate level of anesthesia was maintained for the duration
of the experiment. Heart rate and body core temperature werecontinuously monitored throughout the experiment and keptwithin the physiological range of 330–430/min and 37–37.5 °C,respectively. All procedures were approved by the Universityof Toronto Animal Care Committee in accordance with theregulations of the Ontario Animal Research Act (Canada).
4.2. Recording and stimulating procedures
Extracellular activity of single trigeminal nociceptive primaryafferents with RFs in deep craniofacial tissues (masseter ortemporalis muscles, or TMJ) was recorded with an epoxy-resin-coated tungstenmicroelectrode. One hour after comple-tion of surgery, the microelectrode was slowly lowered intothe brain under stereotaxic guidance (anterior 3.5–4 mm,lateral 3–4 mm) until afferent discharges were observed inresponse to light brush stimuli applied to the craniofacialregion; these dischargeswere usually found 7–8mmbelow thecortical surface. A round dental burnisher (1-mm diameter)was then applied as a noxious mechanical search stimulus(∼100 g) over the craniofacial cutaneous tissues in an attemptto identify trigeminal afferents with deep craniofacial noci-ceptive RFs (Cairns et al. 2001b). This∼100 g forcewas found tobe noxious to the experimenter when applied to his TMJ regionor dorsal surface of his hand.
When an afferent that responded to direct, blunt noxiousmechanical stimuli was found, a careful assessment of its RFwas made to ascertain that the afferent was responding todeep as opposed to cutaneous stimulation. The skin overlyingthe RF was pulled gently away from contact with the deeptissue, and brush, pinch, and pressure stimuli were applieddirectly to the skin surface. If the afferent did not respond toany of these cutaneous stimuli, then the afferent wasconsidered to have a deep mechanonociceptive RF. The RFand MAT of the afferent was assessed at the time intervalsspecified under the experimental paradigm (see below). Thedeep RF of each afferent was determined through the use of around dental burnisher. The size and location of the afferent'sdeep RF was also outlined on a life-size drawing of the rat'shead. The MAT of the afferent's deep RF was determined withan electronic von Frey device (Model 735, 1.0-mm diameterprobe tip, Somedic Sales AB, Sweden) applied to the center ofthe RF and was defined as the force (g) required to evoke thefirst spike, or a firing rate of greater than 2 standard deviationsabove baseline afferent activity when the afferent wasspontaneously active, measured at the afferent's deep RF sitewith a ramp of gradually increasing force. The reproducibilityof the electronic von Frey device in determiningMAT has beendemonstrated previously (Möller et al. 1998; Cairns et al. 2002).
To test whether an afferent with a deep craniofacial RFprojected to the caudal brainstem, electrical stimuli (50 μsbiphasic pulse, range 10–80 μA, 0.5 Hz) were applied to astimulating electrode lowered into the left Vc/UCC (0–6 mmcaudal to obex). The stimulating electrode was movedmediolaterally (0.1-mm steps) and rostrocaudally (0.5-mmsteps) in the Vc/UCC until electrical stimulation evokedantidromic responses as determined by classical criteria forantidromic activation (all-or-none at threshold, invariantlatency, high-frequency following (≥100 Hz), and collision)(Price et al. 1976; Cairns et al. 1996, 2001a,b). The initial

137B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
electrical stimuli were applied 6 mm caudal to the obex. Ifstimulation at this location did not evoke an antidromic actionpotential, the stimulating electrode was moved rostrallytoward the obex until either an antidromic action potentialwas evoked or electrical stimuli had been applied unsuccess-fully up to the level of the obex. Antidromic action potentialswere collided with the orthodromic action potentials evokedby mechanical stimulation of the deep craniofacial RF, toconfirm the projection of the deep nociceptive afferent to thecaudal brainstem. At the end of the study, the antidromic CVof the afferent was determined by calculating the straight-linedistance between the stimulatingmicroelectrode placed in theVc/UCC and the recording microelectrode, divided by theantidromic latency. The skin overlying the deep craniofacialRF of the afferent was also surgically excised and it wasconfirmed that mechanical and/or electrical (50–100 μs bipha-sic pulse, range 10–80 μA, 0.5 Hz) stimuli applied directly to thedeep tissue could also evoke activity in the afferent. Ifelectrical stimulation applied to the deep tissue RF evokedan orthodromic action potential of invariant latency (<0.2 msvariability) with the ability to follow high-frequency electricalstimuli (≥100 Hz), then the conduction distance between thestimulation location and the trigeminal ganglion was esti-mated and an orthodromic CV calculated. For afferents thatcould not be demonstrated to project to the Vc/UCC, onlyorthodromic CVs were determined.
4.3. Receptor agonists
Glutamate (0.5M; 10 μL; Sigma Chemical Company, St. Louis,MO), 1% capsaicin (10% capsaicin in ethanol:Tween-80: sterilenormal saline in a 1:1:8 ratio by volume; 10 μL; Calbiochem, LaJolla, CA) or vehicle (isotonic saline as control for glutamate orethanol:Tween 80:sterile normal saline in a 1:1:8 ratio byvolume as control for capsaicin; 10 μL) was injected into thedeep craniofacial afferent RF at 30 min intervals in bothexperimental subgroups of rats. The concentrations of gluta-mate and capsaicin were chosen on the basis of their efficacyin evoking jawmuscle activity and inflammation and we havepreviously documented that such injected solutions arelocalized to the site of injection (Cairns et al. 1998, 2001a,b,2002, 2003a; Tang et al. 2004; Hu et al. 2005a). All solutionswere adjusted to approximate physiologic pH (7.2–7.6).
4.4. Experimental paradigm
Experimental animals were divided into subgroups accordingto the sequence of injection of receptor agonists: afferentresponse properties of rats with glutamate (or vehicle)injection followed by capsaicin (or vehicle) injection in onesubgroup of rats were comparedwith properties of afferents ina second subgroup of rats with capsaicin (or vehicle) injectionfollowed by glutamate (or vehicle) injection. The followingexperimental paradigm was applied to each of the subgroups:10 min after a nociceptive afferent was classified on the basisof deep RF, antidromic CV and response properties as an Aδ(CV≥2.5–30 m/s) or C (CV<2.5 m/s) fiber nociceptive afferent(Price et al. 1976; Cairns et al. 2001a,b), the baseline MAT (in g)was determined by averaging the threshold for three con-secutive mechanical stimuli applied at 1-minute intervals.
The needle tip of a catheter (a 27-gauge needle connected bypolyethylene tubing to a Hamilton syringe, 100 μL) wascarefully inserted into the deep tissue RF of the afferent. Itwas observed that insertion of the catheter used to inject thereceptor agonist evoked a spike discharge in all nociceptiveafferents identified in order to confirm that the injection sitewas within the afferent RF. Baseline afferent activity wasrecorded for 10 min prior to injection of the first receptoragonist or vehicle control into the deep tissue RF. The receptoragonist or vehicle control was then slowly injected into thedeep tissue (over a 5-second period). The following fourresponse properties were assessed over the next 10 minperiod: (1) Response magnitude (Rmag): the total number ofevoked spikes, or a firing rate of greater than 2 standarddeviations above baseline afferent activity when the afferentwas spontaneously active, following agonist or vehicle injec-tion, (2) Response duration (Rdur): the total time (seconds) fromthe first spike following agonist or vehicle injection to the lastspike, (3) Response latency (Rlat): the total time (seconds) fromagonist or vehicle injection to the first spike following agonistinjection, and (4) Peak frequency (Pfreq): the highest firing ratein a 1 s period (Hz) during the Rdur. The needle was thenwithdrawn at the end of the 10 min period and an assessmentwas made at this 10 min time point and again at 20 min afterthe injection of the receptor agonist or vehicle control todetermine if any MAT changes from baseline had occurred.MAT reduction was defined as ≥50% threshold reduction frombaseline MAT score measured at the center of the RF site. RawMAT threshold values measured post-injection were normal-ized to the initial baseline pre-injection value to illustratepopulation responses. The same protocol described abovewasused for injection of the second receptor agonist (or vehicle)30 min post-injection of the first receptor agonist (or vehicle).
In 13 rats, it was possible to examine the effect of injectedreceptor agonists on more than one afferent on the ipsilateral(left) side because the RFs of the afferents were at differentdeep tissue sites or opposite ends of the samemuscle. In thesecases, which helped minimize the total number of animalsused in the present study, a minimum of 2 h elapsed betweeninjections into the RFs. At the end of each experiment, ratswere euthanized with the agent T61 (Hoechst, Canada).The brain was removed and it was confirmed that microelec-trode tracks were visible on the surface of the trigeminalganglion.
4.5. Data analysis
Recorded afferent activity was stored electronically andanalyzed off-line. Most of the population data are reportedasmean±SE. However, if not normally distributed, populationdata are reported as median values with interquartile rangesindicated in square brackets; median [IQR]. Mann–Whitney Utest, t-test, Fisher exact test, and RM ANOVA-on-ranks wereused as appropriate (p<0.05 considered to reflect statisticalsignificance).
Acknowledgments
Support contributed by CIHR MOP-43905 and NIH DE15420.

138 B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
R E F E R E N C E S
Afrah, A.W., Stiller, C.O., Olgart, L., Brodin, E., Gustafsson, H., 2001.Involvement of spinal N-methyl-D-aspartate receptors incapsaicin-induced in vivo release of substance P in the ratdorsal horn. Neurosci. Lett. 316, 83–86.
Baumann, T.K., Simone, D.A., Shain, C.N., LaMotte, R.H., 1991.Neurogenic hyperalgesia: the search for the primary cutaneousafferent fibers that contribute to capsaicin-induced pain andhyperalgesia. J. Neurophysiol. 66, 212–227.
Bautista, D.M., Jordt, S.E., Nikai, T., Tsuruda, P.R., Read, A.J.,Poblete, J., Yamoah, E.N., Basbaum, A.I., Julius, D., 2006. TRPA1mediates the inflammatory actions of environmental irritantsand proalgesic agents. Cell 124, 1269–1282.
Cairns, B.E., Fragoso, M.C., Soja, P.J., 1996. Active-sleep-relatedsuppression of feline trigeminal sensory neurons: evidenceimplicating presynaptic inhibition via a process of primaryafferent depolarization. J. Neurophysiol. 75, 1152–1162.
Cairns, B.E., Sessle, B.J., Hu, J.W., 1998. Evidence that excitatoryamino acid receptors within the temporomandibular jointregion are involved in the reflex activation of the jaw muscles.J. Neurosci. 18, 8056–8064.
Cairns, B.E., Hu, J.W., Arendt-Nielsen, L., Sessle, B.J., Svensson, P.,2001a. Sex-related differences in human pain and rat afferentdischarge evoked by injection of glutamate into the massetermuscle. J. Neurophysiol. 86, 782–791.
Cairns, B.E., Sessle, B.J., Hu, J.W., 2001b. Characteristics ofglutamate-evoked temporomandibular joint afferent activityin the rat. J. Neurophysiol. 85, 2446–2454.
Cairns, B.E., Gambarota, G., Svensson, P., Arendt-Nielson, L., Berde,C.B., 2002. Glutamate-induced sensitization of rat massetermuscle fibers. Neuroscience 109, 389–399.
Cairns, B.E., Svensson, P., Wang, K., Hupfeld, S., Graven-Nielsen,T., Sessle, B.J., Berde, C.B., Arendt-Nielsen, L., 2003a.Activation of peripheral NMDA receptors contributes tohuman pain and rat afferent discharges evoked by injectionof glutamate into the masseter muscle. J. Neurophys. 90,2098–2105.
Cairns, B.E., Wang, K., Hu, J.W., Sessle, B.J., Arendt-Nielsen, L.,Svensson, P., 2003b. The effect of glutamate-evoked massetermuscle pain on the human jaw-stretch reflex differs in menand women. J. Orofac. Pain 17, 317–325.
Cairns, B.E., Svensson, P., Wang, K., Castrillon, E., Hupfeld, S.,Sessle, B.J., Arendt-Nielsen, L., 2006. Ketamine attenuatesglutamate-induced mechanical sensitization of the massetermuscle in human males. Exp. Brain Res. 169, 467–472.
Cairns, B.E., Dong, X., Mann, M.K., Svensson, P., Sessle, B.J.,Arendt-Nielsen, L., McErlane, K.M., 2007. Systemicadministration of monosodium glutamate elevatesintramuscular glutamate levels and sensitizes rat massetermuscle afferent fibers. Pain 132, 33–41.
Carlton, S.M., 2001. Peripheral excitatory amino acids. Curr. Opin.Pharmacol. 1, 52–56.
Carlton, S.M., McNearney, T., Cairns, B.E., 2003. Peripheralglutamate receptors: Novel targets for Analgesics?In: Dostrovsky, J.O., Carr, D.B., Koltzenburg, M. (Eds.),Proceedings of the 10th World Congress on Pain. IASP Press,Seattle, WA, pp. 125–139.
Carstens, E., Kuenzler, N., Handwerker, H.O., 1998. Activation ofneurons in rat trigeminal subnucleus caudalis by differentirritant chemicals applied to oral or ocular mucosa.J. Neurophysiol. 80, 465–492.
Caterina, M.J., Schumacher, M.A., Tominaga, M., Rosen, T.A.,Levine, J.D., Julius, D., 1997. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389,816–824.
Craft, R.M., Porreca, F., 1992. Treatment parameters ofdesensitization to capsaicin. Life Sci. 51, 1767–1775.
Crandall, M., Kwash, J., Yu, W., White, G., 2002. Activation ofprotein kinase C sensitizes human VR1 to capsaicin and tomoderate decreases in pH at physiological temperatures inXenopus oocytes. Pain 98, 109–117.
Dong, X.D., Mann, M.K., Sessle, B.J., Arendt-Nielsen, L., Svensson,P., Cairns, B.E., 2006. Sensitivity of rat temporalis muscleafferent fibers to peripheral N-methyl-D-aspartate receptoractivation. Neuroscience 141, 939–945.
Dong, X.D., Mann, M.K., Kumar, U., Svensson, P., Arendt-Nielsen,L., Hu, J.W., Sessle, B.J., Cairns, B.E., 2007. Sex-relateddifferences in NMDA-evoked rat masseter muscle afferentdischarge result from estrogen-mediated modulation ofperipheral NMDA receptor activity. Neuroscience 146,822–832.
Dray, A., 2005. Pharmacology of inflammatory pain. In: Merskey,H., Loeser, J.D., Dubner, R. (Eds.), The Paths of Pain 1975–2005.IASP Press, Seattle, WA, pp. 177–190.
Gazerani, P., Arendt-Nielsen, L., 2005. The impact of ethnicdifferences in response to capsaicin-induced trigeminalsensitization. Pain 117, 223–229.
Gazerani, P., Staahl, C., Drewes, A.M., Arendt-Nielsen, L., 2006. Theeffects of Botulinum Toxin type A on capsaicin-evoked pain,flare, and secondary hyperalgesia in an experimental humanmodel of trigeminal sensitization. Pain 122, 315–325.
Hu, J.W., Fiorentino, P.M., Cairns, B.E., Sessle, B.J., 2005a. TRPV1Receptor Mechanisms Involved in Capsaicin-Induced Oedemain the Temporomandibular Joint Region. Oral Bio. Med. 2,241–248.
Hu, J.W., Sun, K.Q., Vernon, H., Sessle, B.J., 2005b. Craniofacialinputs to upper cervical dorsal horn: implications for somato-sensory information processing. Brain Res. 1044, 93–106.
Hucho, T., Levine, J.D., 2007. Signaling pathways in sensitization:toward a nociceptor cell biology. Neuron 55, 365–376.
Ikeda, H., Tokita, Y., Suda, H., 1997. Capsaicin-sensitive A deltafibers in cat tooth pulp. J. Dent. Res. 76, 1341–1349.
Jung, J., Shin, J.S., Lee, S.Y., Hwang, S.W., Koo, J., Cho, H., Oh, U.,2004. Phosphorylation of vanilloid receptor 1 byCa2+/calmodulin-dependent kinase II regulates its vanilloidbinding. J. Biol. Chem. 279, 7048–7054.
Kato, S., Aihara, E., Nakamura, A., Xin, H., Matsui, H., Kohama, K.,Takeuchi, K., 2003. Expression of vanilloid receptors in ratgastric epithelial cells: role in cellular protection. Biochem.Pharmacol. 66, 1115–1121.
Lam, D.K., Sessle, B.J., Hu, J.W., 2004a. Glutamate and capsaicin-evoked activity in deep craniofacial trigeminal nociceptiveafferents. Program No. 3817. 2004 Abstract Viewer/ItineraryPlanner. International Association for Dental Research.
Lam, D.K., Sessle, B.J., Hu, J.W., 2004b. Glutamate andcapsaicin-induced activation and peripheral sensitisation indeep craniofacial trigeminal nociceptive primary afferents.Program No. 294.6. 2004 Abstract Viewer/Itinerary Planner.Society for Neuroscience, Washington, DC. Online.
Lam, D.K., Sessle, B.J., Cairns, B.E., Hu, J.W., 2005a. PeripheralNMDA receptor modulation of jaw muscle electromyographicactivity induced by capsaicin injection into thetemporomandibular joint of rats. Brain Res. 1046, 68–76.
Lam, D.K., Sessle, B.J., Cairns, B.E., Hu, J.W., 2005b. Neuralmechanisms of temporomandibular joint and masticatorymuscle pain: a possible role for peripheral glutamatereceptor mechanisms. Pain Res. Manag. 10,145–152.
Lam, D.K., Sessle, B.J., Hu, J.W., 2008. Glutamate and capsaicineffects on trigeminal nociception II: Activation and centralsensitization in brainstem neurons with deep craniofacialafferent input. Brain Res.
LaMotte, R.H., Lundberg, L.E., Torebjork, H.E., 1992. Pain,hyperalgesia and activity in nociceptive C units in humansafter intradermal injection of capsaicin. J. Physiol. 448,749–764.

139B R A I N R E S E A R C H 1 2 5 1 ( 2 0 0 9 ) 1 3 0 – 1 3 9
Lawand, N.B., McNearney, T., Westlund, K.N., 2000. Amino acidrelease into the knee joint: key role in nociception andinflammation. Pain 86, 69–74.
Li, T., Ghishan, F.K., Bai, L., 2005. Molecular physiology of vesicularglutamate transporters in the digestive system. World J.Gastroenterol. 11, 1731–1736.
Liu, L., Simon, S.A., 1994. A rapid capsaicin-activated current in rattrigeminal ganglion neurons. Proc. Natl. Acad. Sci. U. S. A. 91,738–741.
Liu, L., Simon, S.A., 1996. Capsaicin-induced currents with distinctdesensitization and Ca2+ dependence in rat trigeminal gang-lion cells. J. Neurophysiol. 75, 1503–1514.
Liu, L., Simon, S.A., 2003. Modulation of IA currents by capsaicin inrat trigeminal ganglion neurons. J. Neurophysiol. 89,1387–1401.
McNearney, T., Speegle, D., Lawand, N., Lisse, J., Westlund, K.N.,2000. Excitatory amino acid profiles of synovial fluid frompatients with arthritis. J. Rheumatol. 27, 739–745.
McNearney, T., Baethge, B.A., Cao, S., Alam, R., Lisse, J.R.,Westlund, K.N., 2004. Excitatory amino acids, TNF-alpha, andchemokine levels in synovial fluids of patients with activearthropathies. Clin. Exp. Immunol. 137, 621–627.
Meddings, J.B., Hogaboam, C.M., Tran, K., Reynolds, J.D., Wallace,J.L., 1991. Capsaicin effects on non-neuronal plasmamembranes. Biochim. Biophys. Acta 1070, 43–50.
Mitsikostas, D.D., Sanchez del Rio, M., Waeber, C., Moskowitz, M.A., Cutrer, F.M., 1998. The NMDA receptor antagonist MK-801reduces capsaicin-induced c-fos expression within rattrigeminal nucleus caudalis. Pain 76, 239–248.
Möller, K.A., Johansson, B., Berge, O.G., 1998. Assessingmechanicalallodynia in the rat paw with a new electronic algometer.J. Neurosci. Methods 84, 41–47.
Palazzo, E., de Novellis, V., Marabese, I., Cuomo, D., Rossi, F.,Berrino, L., Rossi, F., Maione, S., 2002. Interaction betweenvanilloid and glutamate receptors in the central modulation ofnociception. Eur. J. Pharmacol. 439, 69–75.
Paoletti, P., Neyton, J., 2007. NMDA receptor subunits: function andpharmacology. Curr. Opin. Pharmacol. 7, 39–47.
Petrenko, A.B., Yamakura, T., Baba, H., Shimoji, K., 2003. The role ofN-methyl-D-aspartate (NMDA) receptors in pain: a review.Anesth. Analg. 97, 1108–1116.
Premkumar, L.S., Ahern, G.P., 2000. Induction of vanilloid receptorchannel activity by protein kinase C. Nature 408, 985–990.
Price, D.D., Dubner, R., Hu, J.W., 1976. Trigeminothalamic neuronsin nucleus caudalis responsive to tactile, thermal, andnociceptive stimulation of monkey's face. J. Neurophysiol. 39,936–953.
Rizvi, S.I., Luqman, S., 2003. Capsaicin-induced activation oferythrocyte membrane sodium/potassium and calciumadenosine triphosphatases. Cell. Mol. Biol. Lett. 8, 919–925.
Ro, J.Y., Capra, N.F., 2006. Assessing mechanical sensitivityof masseter muscle in lightly anesthetized rats: a modelfor craniofacial muscle hyperalgesia. Neurosci. Res. 56,119–123.
Serra, J., Campero, M., Bostock, H., Ochoa, J., 2004. Two types ofC nociceptors in human skin and their behaviour in areas ofcapsaicin-induced secondary hyperalgesia. J. Neurophysiol. 91,2770–2781.
Sikand, P., Premkumar, L.S., 2007. Potentiation of glutamatergicsynaptic transmission by protein kinase C-mediated
sensitization of TRPV1 at the first sensory synapse. J. Physiol.581, 631–647.
Simone, D.A., Marchettini, P., Ochoa, J.L., 1997. Primary afferentnerve fibers that contribute to muscle pain sensation inhuman. Pain Forum 6, 207–212.
Skerry, T.M., Genever, P.G., 2001. Glutamate signalling innon-neuronal tissues. Trends Pharmacol Sci. 22, 174–181.
Sohn, M.K., Graven-Nielsen, T., Arendt-Nielsen, L., Svensson, P.,2000. Inhibition of motor unit firing during experimentalmuscle pain in humans. Muscle Nerve 23, 1219–1226.
Sohn, M.K., Graven-Nielsen, T., Arendt-Nielsen, L., Svensson, P.,2004. Effects of experimental muscle pain on mechanicalproperties of single motor units in human masseter. Clin.Neurophysiol. 115, 76–84.
Strassman, A.M., Raymond, S.A., Burstein, R., 1996. Sensitizationof meningeal sensory neurons and the origin of headaches.Nature 384, 560–564.
Suh, Y.G., Oh, U., 2005. Activation and activators of TRPV1 andtheir pharmaceutical implication. Curr. Pharm. Des. 11,2687–2698.
Svensson, P., Cairns, B.E., Wang, K., Hu, J.W., Graven-Nielsen, T.,Arendt-Nielsen, L., Sessle, B.J., 2003. Glutamate-evoked painandmechanical allodynia in the humanmasseter muscle. Pain101, 221–227.
Svensson, P., Wang, K., Arendt-Nielsen, L., Cairns, B.E., Sessle, B.J.,2005. Pain effects of glutamate injections into human jaw orneck muscles. J. Orofac. Pain 19, 109–118.
Tang, M.L., Haas, D.A., Hu, J.W., 2004. Capsaicin-induced jointinflammation is not blocked by local anesthesia. Anesth. Prog.51, 2–9.
Tominaga, M., Tominaga, T., 2005. Structure and function ofTRPV1. Pflugers Arch. 451, 143–150.
Tominaga, M., Caterina, M.J., Malmberg, A.B., Rosen, T.A., Gilbert,H., Skinner, K., Raumann, B.E., Basbaum, A.I., Julius, D., 1998.The cloned capsaicin receptor integrates multiplepain-modulating stimuli. Neuron 21, 531–543.
Wang, K., Arendt-Nielsen, L., Svensson, P., 2002.Capsaicin-induced muscle pain alters the excitability of thehuman jaw-stretch reflex. J. Dent. Res. 81, 650–654.
Woolf, C.J., Ma, Q., 2007. Nociceptors–noxious stimulus detectors.Neuron 55, 353–364.
Xin, H., Tanaka, H., Yamaguchi, M., Takemori, S., Nakamura, A.,Kohama, K., 2005. Vanilloid receptor expressed in thesarcoplasmic reticulum of rat skeletal muscle. Biochem.Biophys. Res. Commun. 332, 756–762.
Xing, J., Li, J., 2006. TRPV1 receptor mediates glutamatergicsynaptic input to dorsolateral periaqueductal gray (dl-PAG)neurons. J. Neurophysiol. 97, 503–511.
Yamakura, T., Shimoji, K., 1999. Subunit- and site-specificpharmacology of the NMDA receptor channel. Prog Neurobiol.59, 279–298.
Yu, X.M., Sessle, B.J., Haas, D.A., Izzo, A., Vernon, H., Hu, J.W., 1996.Involvement of NMDA receptor mechanisms in jawelectromyographic activity and plasma extravasation inducedby inflammatory irritant application to temporomandibularjoint region of rats. Pain 68, 169–178.
Zanotto, K., Merrill, A.W., Carstens, M.I., Carstens, E., 2007.Neurons in superficial trigeminal subnucleus caudalisresponsive to oral cooling, menthol and other irritant stimuliJ. Neurophysiol. 97, 966–978.