
GENERAL INTRODUCTION INDEX - research.vu.nl matter.pdf · Nowadays, neuroscience techniques allow...
31
8 GENERAL INTRODUCTION INDEX General introduction ........................................................................................................ 9 The brain: a very complex puzzle.................................................................................... 9 Brain puzzle pieces: the key players of brain activity ..................................................... 9 Inhibitory pieces: GABA receptors ............................................................................... 10 Excitatory pieces: Glutamatergic receptors ................................................................... 11 AMPA receptors ......................................................................................................... 11 The hippocampus........................................................................................................... 13 Excitation and inhibition balance .................................................................................. 14 Synaptic plasticity ......................................................................................................... 15 More pieces of the puzzle: Auxiliary proteins............................................................... 18 TARPs .................................................................................................................. 20 Cornichons ........................................................................................................... 21 Shisas ................................................................................................................... 22 Aim of this thesis ........................................................................................................... 23 Bibliography .................................................................................................................. 24
-
Upload
phamnguyet -
Category
Documents
-
view
224 -
download
0
Transcript of GENERAL INTRODUCTION INDEX - research.vu.nl matter.pdf · Nowadays, neuroscience techniques allow...

8
GENERALINTRODUCTIONINDEX
General introduction ........................................................................................................ 9 The brain: a very complex puzzle.................................................................................... 9 Brain puzzle pieces: the key players of brain activity ..................................................... 9 Inhibitory pieces: GABA receptors ............................................................................... 10 Excitatory pieces: Glutamatergic receptors ................................................................... 11
AMPA receptors ......................................................................................................... 11 The hippocampus ........................................................................................................... 13 Excitation and inhibition balance .................................................................................. 14 Synaptic plasticity ......................................................................................................... 15 More pieces of the puzzle: Auxiliary proteins............................................................... 18
TARPs .................................................................................................................. 20 Cornichons ........................................................................................................... 21 Shisas ................................................................................................................... 22
Aim of this thesis ........................................................................................................... 23 Bibliography .................................................................................................................. 24

9
1GENERALINTRODUCTION
The brain: a very complex puzzle The brain is a fascinating and the most complex organ of the body. It has triggered many scientists to devote their time and energy to reveal its nature and how it functions. It was initially studied by anatomists such as the Spanish Nobel prize winner Ramón y Cajal (1906) and many others who were captivated by the beauty of its nature [1, 2]. Cajal’s drawings depict the complexity of the different brain areas, the variety of cell types, but at the same time, his drawings also showed certain order and structure inside an apparent chaos. The high complexity of this massive puzzle that is the brain underlies its capability of performing a multitude of tasks. At a macroscopic level, the brain puzzle pieces could be the different brain areas; visual cortex, motor cortex, hippocampus, amygdala, prefrontal cortex, thalamus, to name a few. Each brain area has its roles and functions; vision, motor movement, memory, fear, and all the rest of functions that are controlled by the brain. Zooming in at the microscopic level, we find smaller puzzle pieces; the diversity of cell types that exist in the different brain areas such as pyramidal neurons, purkinje neurons, and a variety of interneurons and glia cells, among others. The puzzle gets even more complex when we zoom in at the molecular level and we take into consideration the many different molecules that co-exist in each cell, and even more specifically, in each subcellular region. Synapses are of major interest since they are the sites where cells communicate with each other. Synapses undergo dynamic molecular changes that are responsible for the most relevant property of the brain: its plasticity. In this thesis I took on the challenge of investigating the role of several of these molecular puzzle pieces in neuronal functioning. A century ago, Ramon y Cajal was able to discern many anatomical and cellular details with a rudimentary microscopy. Nowadays, neuroscience techniques allow us to analyze the involvement of the several subunits that make up neurotransmitter receptors, discover the protein-to-protein interactions that occur inside the neurons, their effects on neuronal activity and on even higher level such as behavior, memory and learning. Using these modern techniques, we investigated the involvement of different molecular subunits and auxiliary proteins that make up and affect the communication between brain cells and plasticity in the brain. Brain puzzle pieces: the key players of brain activity Brain activity is tightly regulated by very complex molecular mechanisms. The way the brain generates and processes the information has an electrochemical origin. The neurons communicate with each other by means of neurotransmitters. The release and reception of the neurotransmitters is tightly regulated by intricate protein machineries that regulate neuronal activity. In a broader view, brain activity can be seen as the regulation between excitation and inhibition. There are excitatory neurons that, by definition, release excitatory neurotransmitters, such as glutamate, dopamine,

10
acetylcholine, norepinephrine, epinephrine (adrenaline), and serotonin. Similarly, there are inhibitory neurons that release inhibitory transmitters, such as glycine and gamma-aminobutyric acid (GABA), the latter being the most abundant inhibitory neurotransmitter in the brain. In this thesis, I will focus on mechanisms underlying the fast excitatory neurotransmission mediated by AMPA receptors (AMPARs) and the fast inhibitory neurotransmission mediated by GABAA receptors (GABAARs). Inhibitory pieces: GABA receptors GABAARs are ionotropic inhibitory receptors that upon activation allow chloride influx into the cells, which in turn results in hyperpolarization of the adult neurons [3]. GABAA receptors are pentamers composed of a combination of subunits from α1-6, β1-3, γ1-3 and δ [4-7] (figure 1). There is another type of GABARs which are the metabotropic GABABRs, which are involved in intracellular signalling via G-protein coupled receptors linked to potassium channels. In the 80s, Barnard et. al. [8-12] identified the α- and β-subunit of the GABAARs. The binding sites for gamma-aminobutyric acid (GABA) agonists reside on the β-subunits, while the benzodiazepine binding sites reside on the α-subunits. Both subunits are strongly linked allosterically [8]. GABAAR composition varies from brain region and even between neurons within a given region [13-15]. In situ hybridization and immunocytochemical studies revealed that α1 & α2 are widely expressed throughout the brain [16-19] whereas others, like α6, are only expressed in specific populations of neurons, in particular on the cerebellar granule neuron [15, 20] and in particular the subunit α5 is located at extrasynaptic sites [17, 21]. Coexpression of α and β subunits in various combinations reproduced many properties of native GABAARs but not the sensitivity to benzodiazepines. Coexpression of the subunit γ was necessary in order to obtain GABAA-benzodiazepine-sensitive receptors [13, 22]. Moreover, coexpression of γ-subunit variants (γ1, γ 2, γ 3), together with α and β subunits, results in varying degrees of modulation by benzodiazepine receptor ligands [23-27] .
Figure 1. GABA receptor subunit composition and agonists binding sites. Taken from Jacob et. al. 2008 [3] There are pharmacological differences due to subunit composition of the receptors. GABAARs expressing the α1 subunit show higher affinity for GABA than those expressing α2 or α3 subunits [24]. Other pharmacological properties of the receptors

11
might also be affected if the subunit composition is altered. An example would be the loss of zolpidem efficacy in the hippocampus from mutant mice lacking the γ2 GABAA subunit [28]. Moreover, receptors which are composed of α3 subunits, in combination with α1 and α2subunits, yield increased responses to benzodiazepines than do receptors that contain only α1 or α2 subunits. The role of the diverse GABAARs may vary depending on their subunit composition. For example, neurons expressing α1- GABAARs have been found to mediate sedation, whereas those expressing α2-GABAA receptors mediate anxiolysis. [17]. The described subunit heterogeneity of GABAA receptors that provides such diversity of function and pharmacology sensitivity has clinical implications. Deficits in the functional expression of GABAARs are critical in epilepsy, anxiety disorders, cognitive deficits, schizophrenia, depression, and substance abuse [3, 29-34]. Many of these diseases are treated with drugs that act on the GABAergic system. Excitatory pieces: Glutamatergic receptors. Glutamate is released from presynaptic neurons and binds to the ionotropic glutamate receptors (iGluRs) and to the metabotropic receptors (mGluRs). There are three types of iGluRs: alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA), N-methyl-D-aspartate (NMDA) and Kainate (KA) receptors [35, 36]. The metabotropic receptors (mGluRs) have a slower response than the ionotropic receptors and are coupled to GTP-binding proteins (G-proteins) that regulate the production of intracellular messengers [37]. NMDARs are tetrameric ion channels assembled from GluN1-3 subunits with splice variants GluN2A-D, and GluN3A-B [38-40]. NMDARs have a delayed activation that is determined by the coexistence of glutamate binding and a membrane depolarization status that would release the magnesium block that they have by default while at rest. They are permeable to calcium and sodium, contributing to the depolarization of the neuron. Kainate receptors (KARs) are tetramers formed from the subunits GluA5-7, KA1-2. KARs are also permeable to sodium and calcium like AMPAR but they have slower kinetics. Presynaptic KARs modulate GABA release and therefore, have an effect in inhibition. AMPA receptors AMPARs mediate fast excitatory neurotransmission. AMPARs are tetrameric ion channels assembled from GluA1, GluA2, GluA2L (a long splice variant of GluA2), GluA3, GluA4 and GluA4c (short c-tail splice variant of GluA4) subunits (formerly referred as GluR1-4 or GluRA-D) assembled in a dimer of dimer manner [41-43] to form either heteromeric or homomeric receptors. Heteromeric AMPARs typically consist of two GluA2 subunits and either two GluA1, GluA3 or GluA4 subunits. The homomeric receptors are less abundant; they consist of 4 subunits of the same type GluA1, GluA2, GluA3 or GluA4. Great understanding of the structure and assembly of the AMPAR subunits came from the first X-ray structure of a full-length homomeric GluA2 AMPAR [42-45] (figure 2).
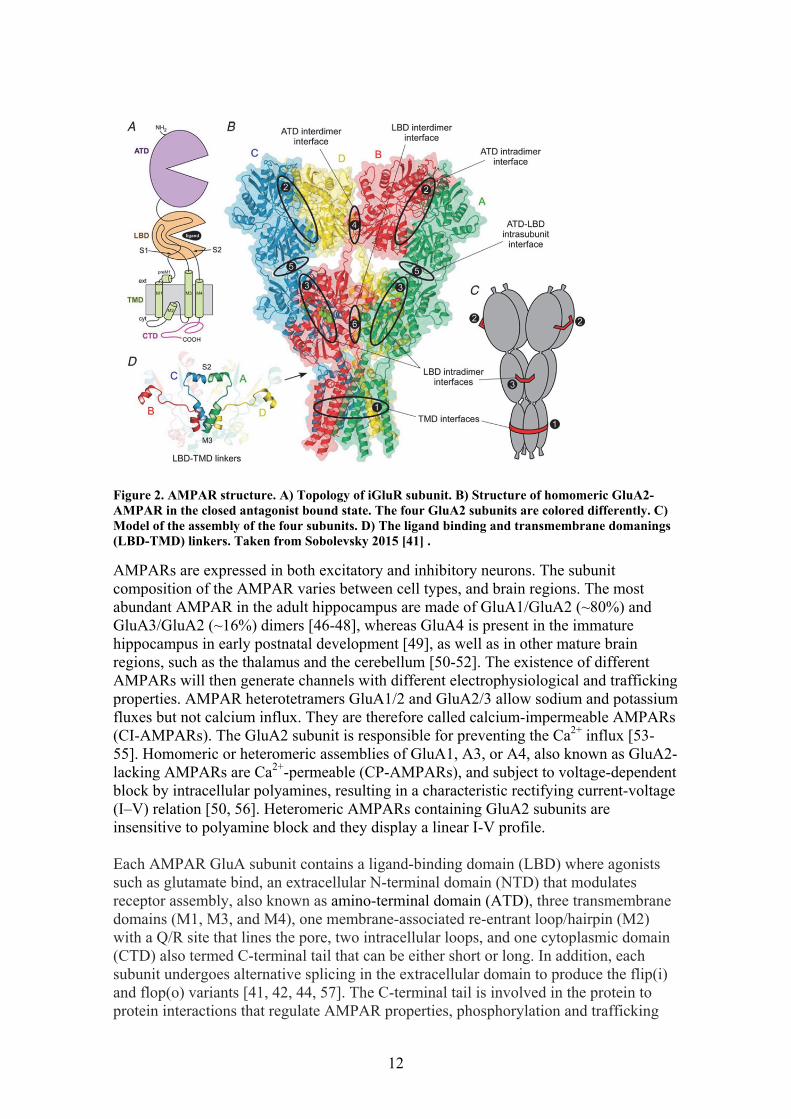
12
Figure 2. AMPAR structure. A) Topology of iGluR subunit. B) Structure of homomeric GluA2-AMPAR in the closed antagonist bound state. The four GluA2 subunits are colored differently. C) Model of the assembly of the four subunits. D) The ligand binding and transmembrane domanings (LBD-TMD) linkers. Taken from Sobolevsky 2015 [41] .
AMPARs are expressed in both excitatory and inhibitory neurons. The subunit composition of the AMPAR varies between cell types, and brain regions. The most abundant AMPAR in the adult hippocampus are made of GluA1/GluA2 (~80%) and GluA3/GluA2 (~16%) dimers [46-48], whereas GluA4 is present in the immature hippocampus in early postnatal development [49], as well as in other mature brain regions, such as the thalamus and the cerebellum [50-52]. The existence of different AMPARs will then generate channels with different electrophysiological and trafficking properties. AMPAR heterotetramers GluA1/2 and GluA2/3 allow sodium and potassium fluxes but not calcium influx. They are therefore called calcium-impermeable AMPARs (CI-AMPARs). The GluA2 subunit is responsible for preventing the Ca2+ influx [53-55]. Homomeric or heteromeric assemblies of GluA1, A3, or A4, also known as GluA2-lacking AMPARs are Ca2+-permeable (CP-AMPARs), and subject to voltage-dependent block by intracellular polyamines, resulting in a characteristic rectifying current-voltage (I–V) relation [50, 56]. Heteromeric AMPARs containing GluA2 subunits are insensitive to polyamine block and they display a linear I-V profile. Each AMPAR GluA subunit contains a ligand-binding domain (LBD) where agonists such as glutamate bind, an extracellular N-terminal domain (NTD) that modulates receptor assembly, also known as amino-terminal domain (ATD), three transmembrane domains (M1, M3, and M4), one membrane-associated re-entrant loop/hairpin (M2) with a Q/R site that lines the pore, two intracellular loops, and one cytoplasmic domain (CTD) also termed C-terminal tail that can be either short or long. In addition, each subunit undergoes alternative splicing in the extracellular domain to produce the flip(i) and flop(o) variants [41, 42, 44, 57]. The C-terminal tail is involved in the protein to protein interactions that regulate AMPAR properties, phosphorylation and trafficking

13
[46]. Long-tailed AMPARs are composed from the GluA1, GluA2L, and GluA4 subunits and short-tailed AMPARs are composed from GluA2, GluA3, and GluA4c. Those AMPARs composed of short C-terminal subunits (GluA2/3) cycle continuously cycling in and out of the synapse, whereas those containing long C-terminal subunits (GluA1/2 and GluA2/4) are delivered to synapses upon synaptic activity [58, 59]. Glutamate and other agonists like AMPA, bind to the AMPAR at the ligand-binding domain (LBD) causing opening of the pore. In addition, there are binding regions at the interface between subunits, within the pore, and in the region between the extracellular domain and the transmembrane segments. Ligands that target these sites can be classified into four categories: (1) competitive agonists and antagonists, (2) positive allosteric modulators, (3) negative allosteric modulators and (4) pore blockers. The kinetics of the AMPARs are determined by the different kinetics and interactions of the ligands. Moreover, it was recently discovered that the kinetics of the AMPARs are also tuned by the different AMPAR auxiliary proteins [60-63] . AMPARs are synthesized in the endoplasmic reticulum (ER) inside the neurons. They travel through the cytosol to reach the synapses where they are needed to mediate fast excitatory transmission. Export of AMPARs from the ER is thought to require the interaction of the cytoplasmic C-terminal domain of the AMPAR subunits with other auxiliary proteins. The GluA2 C-terminus has a PDZ motif that interacts with several PDZ domain-containing proteins, including PICK1 [64, 65], which is thought to be necessary for GluA2 exit from the ER. The GluA1 C-terminus also contains a PDZ domain which interacts with SAP97 [66, 67]. Finally, AMPAR exit from the ER and the acquisition of mature glycosylation at the Golgi complex is assisted by a family of transmembrane AMPAR auxiliary proteins [68, 69]. Further traveling and membrane localization of AMPARs at the synapses is also assisted by a protein machinery [70]. AMPARs get inserted into the membranes by exocytosis and get back into the cytosol by endocytosis where they will be further degraded and recycled [71-74]. AMPARs are very mobile receptors that undergo lateral diffusion to move from extrasynaptic sites to the synaptic sites where they are required to mediate fast excitatory transmission. Imaging studies using fluorescent recovery after photo bleaching and quantum dots technique, revealed that approximately half of the postsynaptic AMPARs were exchangeable with extrasynaptic receptors [75-78]. The availability of AMPAR at the synapse will determine the synaptic strength. Thus, AMPAR status, biophysical properties and AMPA trafficking are key mechanisms for synaptic plasticity. The hippocampus The hippocampus is a brain area responsible for memory formation and spatial navigation. It’s particularly well organized anatomy makes it especially suitable for network experiments. The hippocampus is divided into the following areas: Dentate Gyrus (DG) and the Cornu Ammonis 1, 2 and 3 (CA1,2,3) areas. The hippocampus is connected to multiple brain areas, including several cortices, amygdala and thalamus [79, 80], reflecting it’s great importance. The principal connections within the hippocampus are the mossy fibbers, which connect DG axons with CA3 neurons, the Schaffer collateral fibbers, which connect CA3 axons with CA1 neurons and the perforant path fibers that connect the entorhinal cortex (EC) with the DG neurons. There is a feedback loop hippocampus-EC created with the axons that project from CA1 and CA3 back to the EC.

14
Glutamatergic and gabaergic transmission has been extensively studied in the hippocampus. The molecular and functional properties of both ionotropic glutamatergic and gabaergic receptors have been characterized [39, 81-84]. The different neuronal subtypes of the hippocampus have been classified depending on their morphology, molecular contents and neurotransmitter release In particular, the hippocampus CA1 region contains up to 21 subtypes of interneurons and 2-3 types of pyramidal neurons (figure 3) [81].
Figure 3. Neuronal types in the hippocampal CA1 region. (Image taken from Klausberger Science 2008 [81]) The hippocampus has served as a model to study plasticity for decades. Its layered organized anatomy makes it very suitable for in vitro studies where specific cell types and anatomical regions can be easily identified. Moreover, in vitro studies provide the possibility to control pharmacology applications and understand neuronal function. Excitation and inhibition balance When excitation and inhibition are in balance, the overall activity of the brain fluctuates creating electromagnetic waves known as brain oscillations. Brain oscillations reflect a specific synchrony of the underlying neural network activity. Brain oscillations are classified in frequency bands as: delta (δ, 0.1 ~ 3 Hz), theta (θ, 5 ± 3 Hz ), alpha (α, 10 ± 3 Hz), beta (β, 13 ~ 35 Hz), gamma (40 ~ 100 Hz) and fast ripples (100 ~ 200 Hz). Human brain oscillations were described for the first time by Hans Berger in 1924 [85-87]. He described the alpha rhythm as the 10 Hz oscillation observed in the EEG measurements performed on his son while he was sitting quietly with closed eyes. He already suspected that the observed brain oscillations had their origin inside the cellular and molecular machinery of the recorded cortical tissue. In the 1990’s, Salmelin and Hari [88-90] recorded electromagnetic brain signals from healthy humans and characterized the observed alpha (~10 Hz) as well as the beta oscillations

15
(13 ~ 35 Hz), which were associated with conscious awake activity and motor function. Higher frequency termed gamma oscillations (40–100 Hz) were firstly studied in the cat visual cortex [91] and were found to be associated with higher cognitive tasks and attention [92-96]. A specific oscillation was found in the hippocampus and entorhinal cortex associated with exploratory behavior and rapid eye movement during sleep [97-99] and defined as the theta rhythm (~5 Hz). AMPA and GABA receptors are both essential for the occurrence of brain oscillations, proved by the fact that blocking any of them resulted in the extinction of the brain network oscillations [100]. Oscillatory activity relies on AMPAR-mediated fast synaptic excitation for both the pyramidal cells and the interneurons [101-103] and specially on the feedback loop involving perisomatic inhibition onto the pyramidal neurons [100, 104-106]. Several hippocampal GABAergic neurons have been identified as key players of specific oscillations (figure 3) [81, 107-111]. Hippocampal interneurons were classified according to their protein expression, their morphology, their localization inside the hippocampus, and their firing correlation to the different oscillatory activity [81, 105, 111-119]. Among all interneurons, a sub-type of interneuron plays a key role in generating and maintaining the specific frequency range of beta oscillations: the perisomatic-targeting interneurons which are mainly parvalbumin positive (PV+). In particular, in the hippocampus, the most abundant PV+ interneuron are the basket cells [100, 110, 120, 121]. Another prominent type of interneurons are the cholecystokinin (CCK+) interneurons which are also involved in oscillatory network activity [122-125]. Not only the different receptors determine the overall neuronal activity, but their particular subunit composition also plays a key role on tuning brain activity. Depending on the subunit composition, different activity patterns and oscillations can take place. For example, if the GABAA receptors lack the β3 subunit, hippocampal theta and gamma oscillations get weaker [126]. On the contrary, higher expression of the GABAergic subunits β2 and β3 resulted in faster synaptic kinetics and faster oscillations [109]. In particular in CA1, PV+ basket cells mainly contain α1 GABAARs, while CCK+ interneurons contain α2/α3 GABAARs [127, 128]. Therefore, altering GABAAR activity by modifying receptor subunits would produce changes in oscillatory activity such as the frequency adaptation alteration seen when the α1-GABAR subunit is missing [129, 130]. To have a proper functioning of the brain, all these brain pieces, i.e. excitatory and inhibitory receptors, must be very well fine-tuned. The importance of keeping a proper balance between excitation and inhibition is clearly shown when such balance is perturbed. Disruption of the excitatory-inhibitory balance might lead to dramatic malfunctioning of the neuronal network. The network and behavioral consequences of a pharmacological or pathological excitation/inhibition misbalance can cause severe alterations. The misbalance might translate into an hyper-excited network, epileptic-like network [131-134]. Therefore, it’s of great importance to discern the role of each tiny brain puzzle piece, cause disruption or alterations in any brain piece could potentially have malfunctioning consequences. Synaptic plasticity

16
Synapses are highly specialized areas where neurotransmitters (NTs) are released into the synaptic cleft from the presynaptic neuron by means of an extensive molecular machinery of vesicle exocytosis involving many proteins [135]. At the postsynaptic neuron sites, the NTs are received and bound to the corresponding receptors. The information is thereby transferred from the presynaptic neuron to the postsynaptic neuron by means of electrochemical communication. The postsynaptic neuron will integrate the incoming inputs and act accordingly; i.e. depolarizing and firing specific patterns of action potentials if mainly excitatory inputs were received, or hyperpolarizing and inhibiting the postsynaptic firing if inhibitory inputs govern. Synapses undergo plasticity. They are capable of changing their efficacy and strength by means of altering their molecular contents. In 1949, Donald Hebb proposed a set of rules for synaptic plasticity [136]. Hebbian theory postulated that “when an axon of cell A is near enough to excite a cell B and repeatedly or persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A' s efficiency, as one of the cells firing B, is increased” . Or in short: “Cells that fire together, wire together”. Hebb set the basis for what we now call spike-timing-dependent plasticity (STDP). In line with Hebb’s predictions, synapse strength can be modified depending on the millisecond timing of action potential firing and the sign of synaptic plasticity depends on the spike order of presynaptic and postsynaptic neurons [137-140]. Bi and Poo [141] investigated the specific timing requirements between pre- and postsynaptic neurons in STDP. By varying the timing and order of pre and postsynaptic spiking, they found that critical time windows exist for synaptic modification on the order of tens of milliseconds. The relative spike timing between two neurons will determine the direction of the plasticity, either towards potentiation or towards depression [142, 143]. Plasticity is classified depending on the duration of the changes occurred. If plasticity changes last few to several hundreds of milliseconds, plasticity would fall into the category of short-term plasticity (STP), and if plasticity lasts over several dozens of minutes, hours or even days, it would then be considered long term plasticity. In 1973, Bliss and colleagues [144] reported long-lasting activity-dependent synaptic strengthening in hippocampal synapses, defined then as long-term potentiation (LTP). Interestingly, plasticity changes are bidirectional and reversible. The opposite synaptic process can also occur, causing long-term depression (LTD) [145, 146]. Similarly, for the case of short-term plasticity, the paired-pulse-ratio (PPR) quantification will determine the synaptic change: the augmentation of the synaptic strength would then be termed paired-pulse facilitation (PPF) and on the contrary, the decrease in synaptic strength will be reflected as paired-pulse depression (PPD). The frequency of the synaptic activity determines the direction of the plasticity. High frequency stimulation (HFS) induces LTP, while low frequency stimulation (LFS) induces LTD, i.e. 1 Hz during 15 minutes [142, 147, 148]. Typically, HFS, also known as tetanic stimulation, requires activation of the presynaptic fibres at 100 Hz 3-4 times in a minute. Another broadly used LTP induction protocol is the theta burst stimulation which consists of short trains of 5 pulses at 100 Hz, repeated 10 times at 5 Hz (theta frequency) and repeated 3-4 times. Theta stimulation is thought to be more physiological-like than the tetanus stimulation, based on the fact that hippocampal pyramidal neurons fire bursts of action potentials at theta frequency when animals are engaged in learning and exploring the environment [97, 99].

17
All these plasticity changes rely on complex molecular machineries involving many membrane receptors, vesicle release proteins, synaptic density proteins, kinases and other molecules such as ions. All of these pieces are required players for the synapse to undergo plasticity. To highlight a few, calcium is a key regulator [149, 150] at both the pre- and post-synaptic sites since it regulates vesicle release as well as the intracellular signaling cascade on the postsynaptic site. The amount of neurotransmitter released determines the degree of synaptic strength presynaptically. On the postsynaptic side, ionotropic NMDARs and AMPARs play a key role in long term plasticity. Their number, availability and functionality determine the degree of synaptic strength postsynaptically [151, 152]. It is well established that NMDARs are essential for LTP induction and maintenance over time [153, 154]. Upon glutamate activation of AMPARs the postsynaptic neuron depolarizes, which in turn will cause the removal of the magnesium block at the NMDARs, and therefore calcium will enter the cells activating multiple signaling cascades intracellularly [151, 152, 155-157]. The amount of AMPARs at the synapses was shown to vary during plasticity: potentiated synapses contained more AMPARs and depressed synapses showed less AMPARs (figure 4) [148, 151, 152]. These studies indicated that AMPARs were crucial in determining long lasting plasticity.
Figure 4. LTD and LTP trigger the removal and insertion, respectively, of AMPAR at the synapses. (Image taken from Fleming & England review 2010 [151]) It is not only the ionotropic receptors themselves, but their subunit composition that is crucial in determining the degree of plasticity at the synapses. For example, the long subunits of AMPARs (GluA1, GluA2-long, GluA4) are inserted into the synaptic sited in an activity-dependent manner [46, 49, 58, 158]. In particular, the GluA1 subunit plays a crucial role in plasticity processes like LTP [159, 160]. In the hippocampus, GluA1 lacking mice showed impaired LTD and LTP related to an impaired insertion of AMPARs-GluA1 in the synapses. Specific mutations at the phosphorylation sites of GluA1 produced impaired LTP and LTD [161-163]. Moreover, these plasticity mechanisms require activation of several protein phosphatases and protein kinases in the downstream signalling. LTP in CA1 depends on phosphorylation of GluA1 [164], Ca2+-calmodulin-dependent protein kinase II (CaMKII) [165, 166] and cAMP-dependent protein kinase (PKA) [155, 167-169]. LTD, on the other hand, is accompanied by a dephosphorylation of GluA1 [164], phosphorylation of GluA2 [65, 170]. Altering AMPAR phosphorylation regulates synaptic transmission by either changing single channel properties or receptor trafficking [58, 171, 172]. It is thought that those intracellular signaling pathways control the insertion of AMPAR into the synaptic sites [152, 173]. Overall, AMPARs have been proven to be crucial for LTP and LTD plasticity processes [174].

18
Similarly to glutamatergic synapses, GABAergic synapses undergo plasticity changes [175]. Changes of the GABAAR availability at the synaptic sites as well as changes on the GABAARs’ subunit composition modulate the synaptic strength of GABAergic synapses postsynaptically, whereas the amount of GABA released regulates GABAergic plasticity presynaptically. It has been found that altering α1-GABAAR subunit expression in the brain, alters synaptic plasticity and excitability [176]. Such α1-mediated plasticity has been observed during normal brain development, in animal models of epilepsy, and upon withdrawal from alcohol and benzodiazepines. More pieces of the puzzle: Auxiliary proteins To make the puzzle even more complex, and therefore more exciting and challenging to disentangle, the above described ionotropic glutamate receptors do not exist in isolation in the membrane. It has been found that several membrane receptors are surrounded by a variety of cytoplasmic and transmembrane proteins which interact with several receptors. In particular, the recent discovery that AMPAR are surrounded by a complex network of proteins changed our vision of the synapse into a much more complex one. It is now well established that these auxiliary proteins interact with AMPARs in such a way that they are capable of modulating several biophysical and physiological properties of the AMPARs [61, 72, 177-182]. According to S. Tomita [63], the following criteria have to be fulfill in order to fall into the category of ligand-gated ion channel auxiliary subunit (also termed auxiliary protein): (1) to be a non-pore-forming protein and therefore do not have ion-channel conductance, (2) to have a direct and stable interaction with a pore-forming subunit of the ion channel, (3) to modulate channel properties and/or trafficking in heterologous cells, (4) to be necessary for in vivo native ion channel functioning. Three main families of AMPARs’ auxiliary proteins have been discovered and characterized: transmembrane AMPAR regulatory proteins (TARPs), cornichon proteins and cystine-knot AMPAR auxiliary protein (CKAMP) (figures 5 & 6). The first AMPA auxiliary protein discovered was stargazin (γ2) by Letts et. al. in 1998 [183]. The cerebellar granule cells of the stargazer mouse displayed loss of functional AMPARs at the surface. The mouse that genetically lacked TARP-γ2 suffered from ataxia and epileptic seizures [184, 185]. To date, the following specific AMPA auxiliary proteins have been discovered: stargazin (TARP γ2), TARPs γ3-γ5,γ7-γ8, cornichons 1-4, Shisas 6-7, and Shisa 9 which was initially referred as CKAMP44 (Cys-knot AMPA auxiliary protein with a molecular weight of 44 kDa) [60]. Shisa 4 and 8 are also expressed in the brain and their functionality remains to be studied.

19
Figure 5. Schematic structures of a ionotropic glutamate receptor and three transmembrane auxiliary proteins families. Image adapted from Haering et. al 2014 [186]

20
Figure 6. Diagram of AMPARs and their interacting proteins. AMPARs interact with several transmembrane proteins as well as with post-synaptic intracellular proteins creating a complex network that modulates AMPAR functioning. Adapted from Choquet et. al. 2010 [71] TARPs TARPs are non-pore-forming integral membrane proteins with four transmembrane domains that interact with AMPARs. The stargazin TARP, also named γ2, has more family members: γ3, γ4, γ5, γ7 and γ8. The proteins γ1 and γ6 are not considered TARPs because they do not modulate AMPAR activity. TARPs are classified into two types depending on their PDZ domain: Type I TARPs (γ2, γ3, γ4, γ8) have a typical PDZ binding domain at the C-terminus, whereas Type II TARPs (γ5 & γ7) have a non-canonical PDZ binding domain at the C-terminus [63, 187, 188]. TARPs are widely expressed in the central nervous system. They are expressed differentially all over the brain, and particularly, γ2 is expressed in every type of neuron [189]. Almost all tissues and cell types in the brain express more than one type I TARP subunit, with the exception of cerebellar granule cells, which express only one type I TARP, γ2, and one type II TARP, γ7 [189]. Most AMPARs in the cerebellum have at least one associated TARP [187]. Since the initial discovery of stargazin protein, it has been shown that TARPs play a role in AMPAR maturation [190, 191], AMPAR trafficking [192-197] to the cell surface, AMPAR clustering and subcellular localization [188, 198, 199]. In particular, type I TARPs modulate both trafficking and functional channel properties of AMPARs [200], whereas Type II TARPs modulate only channel properties of AMPARs [181, 201]. Moreover, TARPs also modulate the pharmacology of AMPARs [200, 202-204]. For example, stargazin γ2 lowers the blocking ability of the quinoxalinedione-based glutamatergic antagonists CNQX and NBQX [202, 205, 206]. With respect to the functional properties of AMPARs, TARPs affect several properties of AMPAR functioning like: AMPARs’ kinetics, i.e. changes in the ESPCs’

21
deactivation times, the desensitization times, the recovery from desensitization, and single channel conductance [61, 207-212]. TARPs have been found to be involved in synaptic plasticity [180, 193, 199, 210, 213, 214]. TARPs affect short-term plasticity as well as long term plasticity as it has been been found for TARP γ8. Lack of TARP γ8 resulted in reduced STP and also reduced LTP in the hippocampus [199, 210]. It is then well established that TARPs are indeed AMPAR auxiliary proteins and that they take part in AMPARs’ life cycle and functioning. Cornichons Cornichons are a distinct family of transmembrane AMPAR auxiliary proteins. Mammalian cornichons are homologous to the cornichons found in drosophila and yeast [215]. Cornichons also fulfill the criteria set by S. Tomita [63] to be auxiliary proteins of ionotropic receptors; they are non-pore-forming proteins, they do not have any ion-channel conductance, they have a direct and stable interaction with the AMPAR, and they have been found to modulate the biophysical properties and trafficking of AMPAR [62, 186, 215-218]. Therefore, cornichons are necessary for in vivo native AMPAR functioning. In particular, the isoforms cornichons-2 and cornichons-3, but not the cornichons-1 slow the deactivation and desensitization of AMPAR-mediated currents, independently of the subunit composition of the receptors (GluA2 containing or not). Cornichons increase surface expression of AMPARs [215] Moreover, cornichon-2 and cornichon-3 also affect the functional properties of AMPARs, They enhance the sensitivity of AMPARs to glutamate, they enhance single-channel conductance, they alter channel gating by slowing the deactivation and desensitization times, and they also enhance calcium permeability of CP-AMPARs. Cornichon-2/3 decrease the block by intracellular polyamines of CP- AMPARs [62]. It was initially thought that the functional role of cornichons in neurons was limited to intracellular trafficking of AMPARs from the ER to the Golgi apparatus [216]. It was later found that in hippocampus and in cerebellar neurons, cornichons were also involved in the trafficking of AMPARs to the synapses but in a collaborative manner together with TARP-γ8. Hippocampal AMPAR-mediated synaptic transmission was reduced in the cornichon-2/3 conditional knockout mice. This deficit was thought to be because of the selective loss of surface GluA1-containing AMPARs [218]. Neurons lacking cornichon-2/3 showed faster kinetics, attributed to the loss of GluA1/2 receptors, which deactivate more slowly than the remaining GluA2/3 receptors. A direct role of cornichons in plasticity has not yet been described. Cornichons might affect plasticity indirectly by competing with TARPs on their binding site at AMPARs. It was found that cornichon-2/3 interaction with non-GluA1 subunits was prevented by TARP-γ8 [218]. Thus, the current view of the AMPAR functioning and trafficking includes interactions between the different AMPAR subunits with TARPs, cornichons, and most probably also with the newly characterized Shisa proteins.

22
Shisas The third family of AMPAR auxiliary proteins is the Shisa family. The first Shisa to be discovered and characterized was Shisa9, initially named CKAMP44 from Cystine-knot AMPAR auxiliary protein with a molecular weight of 44 kDa [60]. There are currently 9 identified shisa family members, but only the Shisas 4,6-9 are expressed in the brain. In particular, Shisa9 is more abundant in the dentate gyrus (DG) in the hippocampus and in cerebellum, and is also present in the cortex [60]. Shisa6 is specifically expressed in cerebellum and co-expressed in the hippocampus together with Shisa7 [219]. The latter is also broadly expressed through cortical regions and not in cerebellum. Shisa 4 and 8 are also expressed in the brain and its functionality remains to be studied. The characterization of Shisas as AMPAR auxiliary proteins is very recent. To date, three family members (Shisa6, Shisa7 and Shisa9) have been found to fulfill the criteria to be ion-channel auxiliary proteins. . The initial report on Shisa9 (CKAMP44) [60] showed that Shisa9 interacts with GluA1 and GluA2, and that overexpression of Shisa9 lowers AMPAR steady-state currents. Shisa9 also affected the functional properties of AMPARs by increasing receptor desensitization, as well as the recovery from desensitization. Moreover, Shisa9 is also involved in short-term plasticity since overexpression of Shisa9 results in lower paired-pulse ratio from the AMPAR-mediated EPSCs in hippocampal pyramidal CA1 neurons. Very recently, another study came up [210] showing that similarly to cornichons, Shisa9 acts in a collaborative manner with TARP γ8. The coexpression of both auxiliary subunits is necessary for the efficient targeting of AMPARs to the cell surface of DG granule cells. Shisa9 and TARP γ-8 are proposed to be contained in the same AMPAR complex based on electrophysiological and biochemical evidence [210]. With respect to plasticity, it was found that TARP-γ8 but not Shisa9 is required for LTP [210]. Another Shisa family protein, Shisa6, has been very recently characterized as an AMPAR auxiliary protein [219, 220]. Shisa6 has been shown to interact with GluA1-3 receptors and affect AMPAR functioning in several ways: lack of Shisa6 in hippocampal pyramidal CA1 neurons results in faster AMPAR kinetics and it results in impaired short term plasticity. Moreover, Shisa6 protects AMPARs from desensitization. Similarly to Shisa9, Shisa6 interacts with postsynaptic density proteins by its C-terminal PDZ domain, which results in confinement of AMPARs at the postsynaptic sites. The newly discovered Shisa proteins have opened up a new vision of AMPARs functioning and tuning by showing that AMPARs’ functioning depends on a complex network of interacting proteins.

23
Aim of this thesis This thesis focuses on specific features of excitatory and inhibitory hippocampal synaptic transmission. In particular, this thesis contains two main topics: 1) the role of the GABAergic α1 and α2 subunits on synaptic transmission and hippocampal network oscillations and 2) the role of the two newly discovered AMPAR auxiliary proteins Shisa7 and Shisa9 in excitatory synaptic plasticity and network activity. The following research questions were addressed:
1. What is the role on synaptic and network activity of the α1 and α2 GABAAR subunits?
2. Do interactions of the C-terminal of the Shisa9 protein affect AMPAR functioning and synaptic properties? Do such interactions affect the fast network oscillations?
3. Is Shisa7 an AMPAR auxiliary protein? Does Shisa7 affect AMPAR functioning? Does Shisa 7 affect the synaptic properties? Does Shisa7 affect glutamatergic plasticity? Does Shisa7 have a role in memory?
To address these questions we used a multidisciplinary approach combining molecular assays, electrophysiological techniques of patch-clamp and multi-electrode arrays for single cell and field potential recordings, respectively, on in vitro mice hippocampal brain preparations. The study was supported by molecular assays and completed with mice behavior paradigms such as fear conditioning in order to test if AMPAR auxiliary proteins have a role in hippocampal memory formation. The combined performed studies gave us a broader overview of the role of the AMPAR auxiliary proteins Shisa9 and Shisa7 as well as the differential role of the GABAergic subunits α1 and α2.

24
Bibliography 1. Sotelo C (2003) Viewing the brain through the master hand of Ramon y Cajal. Nat Rev Neurosci 4: 71-77. 2. Garcia-Lopez P, Garcia-Marin V and Freire M (2010) The histological slides and drawings of cajal. Front Neuroanat 4: 9. 3. Jacob TC, Moss SJ and Jurd R (2008) GABA(A) receptor trafficking and its role in the dynamic modulation of neuronal inhibition. Nat Rev Neurosci 9: 331-343. 4. Pirker S, Schwarzer C, Wieselthaler A, Sieghart W and Sperk G (2000) GABA(A) receptors: immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience 101: 815-850. 5. Sieghart W and Sperk G (2002) Subunit composition, distribution and function of GABA(A) receptor subtypes. Curr Top Med Chem 2: 795-816. 6. Wisden W, Laurie DJ, Monyer H and Seeburg PH (1992) The distribution of 13 GABAA receptor subunit mRNAs in the rat brain. I. Telencephalon, diencephalon, mesencephalon. J Neurosci 12: 1040-1062. 7. Olsen RW and Tobin AJ (1990) Molecular biology of GABAA receptors. Faseb J 4: 1469-1480. 8. Casalotti SO, Stephenson FA and Barnard EA (1986) Separate subunits for agonist and benzodiazepine binding in the gamma-aminobutyric acidA receptor oligomer. J Biol Chem 261: 15013-15016. 9. Barnard EA, Darlison MG, Fujita N, Glencorse TA, Levitan ES, et al. (1988) Molecular biology of the GABAA receptor. Adv Exp Med Biol 236: 31-45. 10. Barnard EA and Seeburg PH (1988) Structural basis of the GABA-activated chloride channel: molecular biology and molecular electrophysiology. Adv Biochem Psychopharmacol 45: 1-18. 11. Barnard EA (1995) The molecular biology of GABAA receptors and their structural determinants. Adv Biochem Psychopharmacol 48: 1-16. 12. Barnard EA, Darlison MG and P S (1987) Molecular biology of the GABAA receptor: the receptor channel superfamily. Trends in neurosciences. 13. Shivers BD, Killisch I, Sprengel R, Sontheimer H, Kohler M, et al. (1989) Two novel GABAA receptor subunits exist in distinct neuronal subpopulations. Neuron 3: 327-337. 14. Somogyi P, Takagi H, Richards JG and Mohler H (1989) Subcellular localization of benzodiazepine/GABAA receptors in the cerebellum of rat, cat, and monkey using monoclonal antibodies. J Neurosci 9: 2197-2209. 15. Baude A, Sequier JM, McKernan RM, Olivier KR and Somogyi P (1992) Differential subcellular distribution of the alpha 6 subunit versus the alpha 1 and beta 2/3 subunits of the GABAA/benzodiazepine receptor complex in granule cells of the cerebellar cortex. Neuroscience 51: 739-748. 16. Kasugai Y, Swinny JD, Roberts JD, Dalezios Y, Fukazawa Y, et al. (2010) Quantitative localisation of synaptic and extrasynaptic GABAA receptor subunits on hippocampal pyramidal cells by freeze-fracture replica immunolabelling. Eur J Neurosci 32: 1868-1888. 17. Mohler H (2006) GABA(A) receptor diversity and pharmacology. Cell Tissue Res 326: 505-516. 18. Nusser Z, Roberts JD, Baude A, Richards JG, Sieghart W, et al. (1995) Immunocytochemical localization of the alpha 1 and beta 2/3 subunits of the GABAA

25
receptor in relation to specific GABAergic synapses in the dentate gyrus. Eur J Neurosci 7: 630-646. 19. Nusser Z, Sieghart W and Somogyi P (1998) Segregation of different GABAA receptors to synaptic and extrasynaptic membranes of cerebellar granule cells. J Neurosci 18: 1693-1703. 20. Nusser Z, Sieghart W, Stephenson FA and Somogyi P (1996) The alpha 6 subunit of the GABAA receptor is concentrated in both inhibitory and excitatory synapses on cerebellar granule cells. J Neurosci 16: 103-114. 21. Towers SK, Gloveli T, Traub RD, Driver JE, Engel D, et al. (2004) Alpha 5 subunit-containing GABAA receptors affect the dynamic range of mouse hippocampal kainate-induced gamma frequency oscillations in vitro. J Physiol 559: 721-728. 22. Cope DW, Wulff P, Oberto A, Aller MI, Capogna M, et al. (2004) Abolition of zolpidem sensitivity in mice with a point mutation in the GABAA receptor gamma2 subunit. Neuropharmacology 47: 17-34. 23. Sarto I, Klausberger T, Ehya N, Mayer B, Fuchs K, et al. (2002) A novel site on gamma 3 subunits important for assembly of GABA(A) receptors. J Biol Chem 277: 30656-30664. 24. Scholze P, Ebert V and Sieghart W (1996) Affinity of various ligands for GABAA receptors containing alpha 4 beta 3 gamma 2, alpha 4 gamma 2, or alpha 1 beta 3 gamma 2 subunits. Eur J Pharmacol 304: 155-162. 25. Korpi ER, Mihalek RM, Sinkkonen ST, Hauer B, Hevers W, et al. (2002) Altered receptor subtypes in the forebrain of GABA(A) receptor delta subunit-deficient mice: recruitment of gamma 2 subunits. Neuroscience 109: 733-743. 26. Pritchett DB, Sontheimer H, Shivers BD, Ymer S, Kettenmann H, et al. (1989) Importance of a novel GABAA receptor subunit for benzodiazepine pharmacology. Nature 338: 582-585. 27. Nicoll RA, Eccles JC, Oshima T and Rubia F (1975) Prolongation of hippocampal inhibitory postsynaptic potentials by barbiturates. Nature 258: 625-627. 28. Cope DW, Halbsguth C, Karayannis T, Wulff P, Ferraguti F, et al. (2005) Loss of zolpidem efficacy in the hippocampus of mice with the GABAA receptor gamma2 F77I point mutation. Eur J Neurosci 21: 3002-3016. 29. Loup F, Picard F, Andre VM, Kehrli P, Yonekawa Y, et al. (2006) Altered expression of alpha3-containing GABAA receptors in the neocortex of patients with focal epilepsy. Brain 129: 3277-3289. 30. Loup F, Wieser HG, Yonekawa Y, Aguzzi A and Fritschy JM (2000) Selective alterations in GABAA receptor subtypes in human temporal lobe epilepsy. J Neurosci 20: 5401-5419. 31. Loup F, Picard F, Yonekawa Y, Wieser HG and Fritschy JM (2009) Selective changes in GABAA receptor subtypes in white matter neurons of patients with focal epilepsy. Brain 132: 2449-2463. 32. Wendling F, Bartolomei F, Bellanger JJ and Chauvel P (2002) Epileptic fast activity can be explained by a model of impaired GABAergic dendritic inhibition. Eur J Neurosci 15: 1499-1508. 33. Yee BK, Keist R, von Boehmer L, Studer R, Benke D, et al. (2005) A schizophrenia-related sensorimotor deficit links alpha 3-containing GABAA receptors to a dopamine hyperfunction. Proc Natl Acad Sci U S A 102: 17154-17159. 34. Lewis DA, Cho RY, Carter CS, Eklund K, Forster S, et al. (2008) Subunit-selective modulation of GABA type A receptor neurotransmission and cognition in schizophrenia. The American journal of psychiatry 165: 1585-1593.

26
35. Ozawa S, Kamiya H and Tsuzuki K (1998) Glutamate receptors in the mammalian central nervous system. Prog Neurobiol 54: 581-618. 36. Simeone TA, Sanchez RM and Rho JM (2004) Molecular biology and ontogeny of glutamate receptors in the mammalian central nervous system. J Child Neurol 19: 343-360; discussion 361. 37. Nistri A, Ostroumov K, Sharifullina E and Taccola G (2006) Tuning and playing a motor rhythm: how metabotropic glutamate receptors orchestrate generation of motor patterns in the mammalian central nervous system. J Physiol 572: 323-334. 38. Ehlers MD, Tingley WG and Huganir RL (1995) Regulated subcellular distribution of the NR1 subunit of the NMDA receptor. Science 269: 1734-1737. 39. Fritschy JM, Weinmann O, Wenzel A and Benke D (1998) Synapse-specific localization of NMDA and GABA(A) receptor subunits revealed by antigen-retrieval immunohistochemistry. J Comp Neurol 390: 194-210. 40. Sobolevsky AI, Prodromou ML, Yelshansky MV and Wollmuth LP (2007) Subunit-specific contribution of pore-forming domains to NMDA receptor channel structure and gating. J Gen Physiol 129: 509-525. 41. Sobolevsky AI (2015) Structure and gating of tetrameric glutamate receptors. J Physiol 593: 29-38. 42. Wollmuth LP and Sobolevsky AI (2004) Structure and gating of the glutamate receptor ion channel. Trends Neurosci 27: 321-328. 43. Sobolevsky AI, Rosconi MP and Gouaux E (2009) X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor. Nature 462: 745-756. 44. Jin R, Singh SK, Gu S, Furukawa H, Sobolevsky AI, et al. (2009) Crystal structure and association behaviour of the GluR2 amino-terminal domain. Embo J 28: 1812-1823. 45. Sobolevsky AI, Yelshansky MV and Wollmuth LP (2004) The outer pore of the glutamate receptor channel has 2-fold rotational symmetry. Neuron 41: 367-378. 46. Gomes AR, Correia SS, Carvalho AL and Duarte CB (2003) Regulation of AMPA receptor activity, synaptic targeting and recycling: role in synaptic plasticity. Neurochem Res 28: 1459-1473. 47. Wenthold RJ, Petralia RS, Blahos J, II and Niedzielski AS (1996) Evidence for multiple AMPA receptor complexes in hippocampal CA1/CA2 neurons. J Neurosci 16: 1982-1989. 48. Lu W, Shi Y, Jackson AC, Bjorgan K, During MJ, et al. (2009) Subunit composition of synaptic AMPA receptors revealed by a single-cell genetic approach. Neuron 62: 254-268. 49. Zhu JJ, Esteban JA, Hayashi Y and Malinow R (2000) Postnatal synaptic potentiation: delivery of GluR4-containing AMPA receptors by spontaneous activity. Nat Neurosci 3: 1098-1106. 50. Keinanen K, Wisden W, Sommer B, Werner P, Herb A, et al. (1990) A family of AMPA-selective glutamate receptors. Science 249: 556-560. 51. Monyer H, Seeburg PH and Wisden W (1991) Glutamate-operated channels: developmentally early and mature forms arise by alternative splicing. Neuron 6: 799-810. 52. Okamoto K and Sekiguchi M (1991) Synaptic receptors and intracellular signal transduction in the cerebellum. Neurosci Res 9: 213-237. 53. Tanaka H, Grooms SY, Bennett MV and Zukin RS (2000) The AMPAR subunit GluR2: still front and center-stage. Brain Res 886: 190-207. 54. Hanley JG (2014) Subunit-specific trafficking mechanisms regulating the synaptic expression of Ca(2+)-permeable AMPA receptors. Semin Cell Dev Biol 27: 14-22.

27
55. Yuan T and Bellone C (2013) Glutamatergic receptors at developing synapses: the role of GluN3A-containing NMDA receptors and GluA2-lacking AMPA receptors. Eur J Pharmacol 719: 107-111. 56. Rossi B, Maton G and Collin T (2008) Calcium-permeable presynaptic AMPA receptors in cerebellar molecular layer interneurones. J Physiol 586: 5129-5145. 57. Mayer ML (2006) Glutamate receptors at atomic resolution. Nature 440: 456-462. 58. Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, et al. (2000) Driving AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain interaction. Science 287: 2262-2267. 59. Shi S, Hayashi Y, Esteban JA and Malinow R (2001) Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell 105: 331-343. 60. von Engelhardt J, Mack V, Sprengel R, Kavenstock N, Li KW, et al. (2010) CKAMP44: a brain-specific protein attenuating short-term synaptic plasticity in the dentate gyrus. Science 327: 1518-1522. 61. Milstein AD, Zhou W, Karimzadegan S, Bredt DS and Nicoll RA (2007) TARP subtypes differentially and dose-dependently control synaptic AMPA receptor gating. Neuron 55: 905-918. 62. Coombs ID, Soto D, Zonouzi M, Renzi M, Shelley C, et al. (2012) Cornichons modify channel properties of recombinant and glial AMPA receptors. J Neurosci 32: 9796-9804. 63. Yan D and Tomita S (2012) Defined criteria for auxiliary subunits of glutamate receptors. J Physiol 590: 21-31. 64. Xia J, Zhang X, Staudinger J and Huganir RL (1999) Clustering of AMPA receptors by the synaptic PDZ domain-containing protein PICK1. Neuron 22: 179-187. 65. Chung HJ, Xia J, Scannevin RH, Zhang X and Huganir RL (2000) Phosphorylation of the AMPA receptor subunit GluR2 differentially regulates its interaction with PDZ domain-containing proteins. J Neurosci 20: 7258-7267. 66. Douyard J, Shen L, Huganir RL and Rubio ME (2007) Differential neuronal and glial expression of GluR1 AMPA receptor subunit and the scaffolding proteins SAP97 and 4.1N during rat cerebellar development. J Comp Neurol 502: 141-156. 67. Zhou W, Zhang L, Guoxiang X, Mojsilovic-Petrovic J, Takamaya K, et al. (2008) GluR1 controls dendrite growth through its binding partner, SAP97. J Neurosci 28: 10220-10233. 68. Cuadra AE, Kuo SH, Kawasaki Y, Bredt DS and Chetkovich DM (2004) AMPA receptor synaptic targeting regulated by stargazin interactions with the Golgi-resident PDZ protein nPIST. J Neurosci 24: 7491-7502. 69. Stricker NL and Huganir RL (2003) The PDZ domains of mLin-10 regulate its trans-Golgi network targeting and the surface expression of AMPA receptors. Neuropharmacology 45: 837-848. 70. Ehlers MD, Mammen AL, Lau LF and Huganir RL (1996) Synaptic targeting of glutamate receptors. Curr Opin Cell Biol 8: 484-489. 71. Choquet D (2010) Fast AMPAR trafficking for a high-frequency synaptic transmission. Eur J Neurosci 32: 250-260. 72. Opazo P and Choquet D (2011) A three-step model for the synaptic recruitment of AMPA receptors. Mol Cell Neurosci 46: 1-8. 73. Zhou Q, Xiao M and Nicoll RA (2001) Contribution of cytoskeleton to the internalization of AMPA receptors. Proc Natl Acad Sci U S A 98: 1261-1266. 74. Ehlers MD (2000) Reinsertion or degradation of AMPA receptors determined by activity-dependent endocytic sorting. Neuron 28: 511-525.

28
75. Heine M, Groc L, Frischknecht R, Beique JC, Lounis B, et al. (2008) Surface mobility of postsynaptic AMPARs tunes synaptic transmission. Science 320: 201-205. 76. Czondor K, Mondin M, Garcia M, Heine M, Frischknecht R, et al. (2012) Unified quantitative model of AMPA receptor trafficking at synapses. Proc Natl Acad Sci U S A 109: 3522-3527. 77. Groc L, Heine M, Cognet L, Brickley K, Stephenson FA, et al. (2004) Differential activity-dependent regulation of the lateral mobilities of AMPA and NMDA receptors. Nat Neurosci 7: 695-696. 78. Adesnik H, Nicoll RA and England PM (2005) Photoinactivation of native AMPA receptors reveals their real-time trafficking. Neuron 48: 977-985. 79. van Strien NM, Cappaert NL and Witter MP (2009) The anatomy of memory: an interactive overview of the parahippocampal-hippocampal network. Nat Rev Neurosci 10: 272-282. 80. Amaral DG and Witter MP (1989) The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience 31: 571-591. 81. Klausberger T and Somogyi P (2008) Neuronal diversity and temporal dynamics: the unity of hippocampal circuit operations. Science 321: 53-57. 82. Prenosil GA, Schneider Gasser EM, Rudolph U, Keist R, Fritschy JM, et al. (2006) Specific subtypes of GABAA receptors mediate phasic and tonic forms of inhibition in hippocampal pyramidal neurons. J Neurophysiol 96: 846-857. 83. Lissin DV, Gomperts SN, Carroll RC, Christine CW, Kalman D, et al. (1998) Activity differentially regulates the surface expression of synaptic AMPA and NMDA glutamate receptors. Proc Natl Acad Sci U S A 95: 7097-7102. 84. Gomperts SN, Carroll R, Malenka RC and Nicoll RA (2000) Distinct roles for ionotropic and metabotropic glutamate receptors in the maturation of excitatory synapses. J Neurosci 20: 2229-2237. 85. Gloor P (1969) Hans Berger and the discovery of the electroencephalogram. Electroencephalogr Clin Neurophysiol: Suppl 28:21-36. 86. Gloor P (1969) The work of Hans Berger. Electroencephalogr Clin Neurophysiol 27: 649. 87. Berger H (1924) Hans Bergers entdeckung des elektrenkephalogramms Und seine ersten befunde Archiv Für Psychiatrie Und Nervenkrankheiten 6: 227:283. 88. Salmelin R and Hari R (1994) Characterization of spontaneous MEG rhythms in healthy adults. Electroencephalogr Clin Neurophysiol 91: 237-248. 89. Salmelin R and Hari R (1994) Spatiotemporal characteristics of sensorimotor neuromagnetic rhythms related to thumb movement. Neuroscience 60: 537-550. 90. Hari R and Salmelin R (1997) Human cortical oscillations: a neuromagnetic view through the skull. Trends Neurosci 20: 44-49. 91. Ghose GM and Freeman RD (1992) Oscillatory discharge in the visual system: does it have a functional role? J Neurophysiol 68: 1558-1574. 92. Basar E, Schurmann M, Basar-Eroglu C and Karakas S (1997) Alpha oscillations in brain functioning: an integrative theory. Int J Psychophysiol 26: 5-29. 93. Basar E, Basar-Eroglu C, Karakas S and Schurmann M (2000) Brain oscillations in perception and memory. Int J Psychophysiol 35: 95-124. 94. Schurmann M, Basar-Eroglu C and Basar E (1997) Gamma responses in the EEG: elementary signals with multiple functional correlates. Neuroreport 8: 1793-1796. 95. Basar E, Basar-Eroglu C, Karakas S and Schurmann M (2001) Gamma, alpha, delta, and theta oscillations govern cognitive processes. Int J Psychophysiol 39: 241-248.

29
96. Basar-Eroglu C, Struber D, Schurmann M, Stadler M and Basar E (1996) Gamma-band responses in the brain: a short review of psychophysiological correlates and functional significance. Int J Psychophysiol 24: 101-112. 97. Buzsaki G (2005) Theta rhythm of navigation: link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 15: 827-840. 98. Royer S, Zemelman BV, Losonczy A, Kim J, Chance F, et al. (2012) Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nat Neurosci 15: 769-775. 99. Geisler C, Diba K, Pastalkova E, Mizuseki K, Royer S, et al. (2010) Temporal delays among place cells determine the frequency of population theta oscillations in the hippocampus. Proc Natl Acad Sci U S A 107: 7957-7962. 100. Mann EO, Suckling JM, Hajos N, Greenfield SA and Paulsen O (2005) Perisomatic feedback inhibition underlies cholinergically induced fast network oscillations in the rat hippocampus in vitro. Neuron 45: 105-117. 101. Palhalmi J, Paulsen O, Freund TF and Hajos N (2004) Distinct properties of carbachol- and DHPG-induced network oscillations in hippocampal slices. Neuropharmacology 47: 381-389. 102. Bartos M, Vida I, Frotscher M, Meyer A, Monyer H, et al. (2002) Fast synaptic inhibition promotes synchronized gamma oscillations in hippocampal interneuron networks. Proc Natl Acad Sci U S A 99: 13222-13227. 103. Stark E, Roux L, Eichler R, Senzai Y, Royer S, et al. (2014) Pyramidal cell-interneuron interactions underlie hippocampal ripple oscillations. Neuron 83: 467-480. 104. Hajos N and Paulsen O (2009) Network mechanisms of gamma oscillations in the CA3 region of the hippocampus. Neural Netw 22: 1113-1119. 105. Tukker JJ, Fuentealba P, Hartwich K, Somogyi P and Klausberger T (2007) Cell type-specific tuning of hippocampal interneuron firing during gamma oscillations in vivo. J Neurosci 27: 8184-8189. 106. Mann EO, Radcliffe CA and Paulsen O (2005) Hippocampal gamma-frequency oscillations: from interneurones to pyramidal cells, and back. J Physiol 562: 55-63. 107. Heistek TS, Ruiperez-Alonso M, Timmerman AJ, Brussaard AB and Mansvelder HD (2012) alpha2-containing GABAA receptors expressed in hippocampal region CA3 control fast network oscillations. J Physiol 591: 845-858. 108. Mann EO and Paulsen O (2007) Role of GABAergic inhibition in hippocampal network oscillations. Trends Neurosci 30: 343-349. 109. Heistek TS, Jaap Timmerman A, Spijker S, Brussaard AB and Mansvelder HD (2010) GABAergic synapse properties may explain genetic variation in hippocampal network oscillations in mice. Front Cell Neurosci 4: 18. 110. Hajos N, Palhalmi J, Mann EO, Nemeth B, Paulsen O, et al. (2004) Spike timing of distinct types of GABAergic interneuron during hippocampal gamma oscillations in vitro. J Neurosci 24: 9127-9137. 111. Hartwich K, Pollak T and Klausberger T (2009) Distinct firing patterns of identified basket and dendrite-targeting interneurons in the prefrontal cortex during hippocampal theta and local spindle oscillations. J Neurosci 29: 9563-9574. 112. Klausberger T, Magill PJ, Marton LF, Roberts JD, Cobden PM, et al. (2003) Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo. Nature 421: 844-848. 113. Klausberger T, Marton LF, Baude A, Roberts JD, Magill PJ, et al. (2004) Spike timing of dendrite-targeting bistratified cells during hippocampal network oscillations in vivo. Nat Neurosci 7: 41-47.

30
114. Somogyi P and Klausberger T (2005) Defined types of cortical interneurone structure space and spike timing in the hippocampus. J Physiol 562: 9-26. 115. Fuentealba P, Tomioka R, Dalezios Y, Marton LF, Studer M, et al. (2008) Rhythmically active enkephalin-expressing GABAergic cells in the CA1 area of the hippocampus project to the subiculum and preferentially innervate interneurons. J Neurosci 28: 10017-10022. 116. Viney TJ, Lasztoczi B, Katona L, Crump MG, Tukker JJ, et al. (2013) Network state-dependent inhibition of identified hippocampal CA3 axo-axonic cells in vivo. Nat Neurosci 16: 1802-1811. 117. Lasztoczi B and Klausberger T (2014) Layer-specific GABAergic control of distinct gamma oscillations in the CA1 hippocampus. Neuron 81: 1126-1139. 118. Ciocchi S, Passecker J, Malagon-Vina H, Mikus N and Klausberger T (2015) Brain computation. Selective information routing by ventral hippocampal CA1 projection neurons. Science 348: 560-563. 119. Forro T, Valenti O, Lasztoczi B and Klausberger T (2015) Temporal organization of GABAergic interneurons in the intermediate CA1 hippocampus during network oscillations. Cereb Cortex 25: 1228-1240. 120. Oren I, Mann EO, Paulsen O and Hajos N (2006) Synaptic currents in anatomically identified CA3 neurons during hippocampal gamma oscillations in vitro. J Neurosci 26: 9923-9934. 121. Gulyas AI, Szabo GG, Ulbert I, Holderith N, Monyer H, et al. (2010) Parvalbumin-containing fast-spiking basket cells generate the field potential oscillations induced by cholinergic receptor activation in the hippocampus. J Neurosci 30: 15134-15145. 122. Somogyi P, Hodgson AJ, Smith AD, Nunzi MG, Gorio A, et al. (1984) Different populations of GABAergic neurons in the visual cortex and hippocampus of cat contain somatostatin- or cholecystokinin-immunoreactive material. J Neurosci 4: 2590-2603. 123. Nunzi MG, Gorio A, Milan F, Freund TF, Somogyi P, et al. (1985) Cholecystokinin-immunoreactive cells form symmetrical synaptic contacts with pyramidal and nonpyramidal neurons in the hippocampus. J Comp Neurol 237: 485-505. 124. Maccaferri G, Roberts JD, Szucs P, Cottingham CA and Somogyi P (2000) Cell surface domain specific postsynaptic currents evoked by identified GABAergic neurones in rat hippocampus in vitro. J Physiol 524 Pt 1: 91-116. 125. Somogyi J, Baude A, Omori Y, Shimizu H, El Mestikawy S, et al. (2004) GABAergic basket cells expressing cholecystokinin contain vesicular glutamate transporter type 3 (VGLUT3) in their synaptic terminals in hippocampus and isocortex of the rat. Eur J Neurosci 19: 552-569. 126. Hentschke H, Benkwitz C, Banks MI, Perkins MG, Homanics GE, et al. (2009) Altered GABAA,slow inhibition and network oscillations in mice lacking the GABAA receptor beta3 subunit. J Neurophysiol 102: 3643-3655. 127. Baude A, Bleasdale C, Dalezios Y, Somogyi P and Klausberger T (2007) Immunoreactivity for the GABAA receptor alpha1 subunit, somatostatin and Connexin36 distinguishes axoaxonic, basket, and bistratified interneurons of the rat hippocampus. Cereb Cortex 17: 2094-2107. 128. Milenkovic I, Vasiljevic M, Maurer D, Hoger H, Klausberger T, et al. (2013) The parvalbumin-positive interneurons in the mouse dentate gyrus express GABAA receptor subunits alpha1, beta2, and delta along their extrasynaptic cell membrane. Neuroscience 254: 80-96.

31
129. Baker C, Sturt BL and Bamber BA (2010) Multiple roles for the first transmembrane domain of GABAA receptor subunits in neurosteroid modulation and spontaneous channel activity. Neurosci Lett 473: 242-247. 130. Szabadics J, Varga C, Molnar G, Olah S, Barzo P, et al. (2006) Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits. Science 311: 233-235. 131. Bragin A, Penttonen M and Buzsaki G (1997) Termination of epileptic afterdischarge in the hippocampus. J Neurosci 17: 2567-2579. 132. Buzsaki G (1986) Hippocampal sharp waves: their origin and significance. Brain Res 398: 242-252. 133. Buzsaki G, Smith A, Berger S, Fisher LJ and Gage FH (1990) Petit mal epilepsy and parkinsonian tremor: hypothesis of a common pacemaker. Neuroscience 36: 1-14. 134. Menendez de la Prida L and Trevelyan AJ (2011) Cellular mechanisms of high frequency oscillations in epilepsy: on the diverse sources of pathological activities. Epilepsy Res 97: 308-317. 135. Fon EA and Edwards RH (2001) Molecular mechanisms of neurotransmitter release. Muscle Nerve 24: 581-601. 136. Morris RG (1999) D.O. Hebb: The Organization of Behavior, Wiley: New York; 1949. Brain research bulletin 50: 437. 137. Levy WB and Steward O (1983) Temporal contiguity requirements for long-term associative potentiation/depression in the hippocampus. Neuroscience 8: 791-797. 138. Gustafsson B, Wigstrom H, Abraham WC and Huang YY (1987) Long-term potentiation in the hippocampus using depolarizing current pulses as the conditioning stimulus to single volley synaptic potentials. J Neurosci 7: 774-780. 139. Abraham WC, Gustafsson B and Wigstrom H (1987) Long-term potentiation involves enhanced synaptic excitation relative to synaptic inhibition in guinea-pig hippocampus. J Physiol 394: 367-380. 140. Magee JC and Johnston D (1997) A synaptically controlled, associative signal for Hebbian plasticity in hippocampal neurons. Science 275: 209-213. 141. Bi GQ and Poo MM (1998) Synaptic modifications in cultured hippocampal neurons: dependence on spike timing, synaptic strength, and postsynaptic cell type. J Neurosci 18: 10464-10472. 142. Feldman DE (2012) The spike-timing dependence of plasticity. Neuron 75: 556-571. 143. Abbott LF and Nelson SB (2000) Synaptic plasticity: taming the beast. Nat Neurosci 3 Suppl: 1178-1183. 144. Bliss TV and Gardner-Medwin AR (1973) Long-lasting potentiation of synaptic transmission in the dentate area of the unanaestetized rabbit following stimulation of the perforant path. J Physiol 232: 357-374. 145. Neves G, Cooke SF and Bliss TV (2008) Synaptic plasticity, memory and the hippocampus: a neural network approach to causality. Nat Rev Neurosci 9: 65-75. 146. Whitlock JR, Heynen AJ, Shuler MG and Bear MF (2006) Learning induces long-term potentiation in the hippocampus. Science 313: 1093-1097. 147. Bear MF and Malenka RC (1994) Synaptic plasticity: LTP and LTD. Curr Opin Neurobiol 4: 389-399. 148. Malenka RC and Bear MF (2004) LTP and LTD: an embarrassment of riches. Neuron 44: 5-21. 149. Rizo J and Sudhof TC (2012) The membrane fusion enigma: SNAREs, Sec1/Munc18 proteins, and their accomplices--guilty as charged? Annu Rev Cell Dev Biol 28: 279-308.

32
150. Rizo J and Xu J (2015) The Synaptic Vesicle Release Machinery. Annu Rev Biophys 44: 339-367. 151. Fleming JJ and England PM (2010) AMPA receptors and synaptic plasticity: a chemist's perspective. Nat Chem Biol 6: 89-97. 152. Citri A and Malenka RC (2008) Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33: 18-41. 153. Nicoll RA, Oliet SH and Malenka RC (1998) NMDA receptor-dependent and metabotropic glutamate receptor-dependent forms of long-term depression coexist in CA1 hippocampal pyramidal cells. Neurobiol Learn Mem 70: 62-72. 154. Malenka RC and Nicoll RA (1993) NMDA-receptor-dependent synaptic plasticity: multiple forms and mechanisms. Trends Neurosci 16: 521-527. 155. Baudry M, Zhu G, Liu Y, Wang Y, Briz V, et al. (2014) Multiple cellular cascades participate in long-term potentiation and in hippocampus-dependent learning. Brain Res. 156. Malenka RC, Kauer JA, Perkel DJ and Nicoll RA (1989) The impact of postsynaptic calcium on synaptic transmission--its role in long-term potentiation. Trends Neurosci 12: 444-450. 157. Malenka RC, Kauer JA, Perkel DJ, Mauk MD, Kelly PT, et al. (1989) An essential role for postsynaptic calmodulin and protein kinase activity in long-term potentiation. Nature 340: 554-557. 158. Man HY, Sekine-Aizawa Y and Huganir RL (2007) Regulation of {alpha}-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor trafficking through PKA phosphorylation of the Glu receptor 1 subunit. Proc Natl Acad Sci U S A 104: 3579-3584. 159. Makino Y, Johnson RC, Yu Y, Takamiya K and Huganir RL (2011) Enhanced synaptic plasticity in mice with phosphomimetic mutation of the GluA1 AMPA receptor. Proc Natl Acad Sci U S A 108: 8450-8455. 160. Diering GH, Gustina AS and Huganir RL (2014) PKA-GluA1 coupling via AKAP5 controls AMPA receptor phosphorylation and cell-surface targeting during bidirectional homeostatic plasticity. Neuron 84: 790-805. 161. Lee HK, Takamiya K, He K, Song L and Huganir RL (2010) Specific roles of AMPA receptor subunit GluR1 (GluA1) phosphorylation sites in regulating synaptic plasticity in the CA1 region of hippocampus. J Neurophysiol 103: 479-489. 162. Lee HK and Kirkwood A (2011) AMPA receptor regulation during synaptic plasticity in hippocampus and neocortex. Semin Cell Dev Biol 22: 514-520. 163. Boehm J, Kang MG, Johnson RC, Esteban J, Huganir RL, et al. (2006) Synaptic incorporation of AMPA receptors during LTP is controlled by a PKC phosphorylation site on GluR1. Neuron 51: 213-225. 164. Lee HK, Barbarosie M, Kameyama K, Bear MF and Huganir RL (2000) Regulation of distinct AMPA receptor phosphorylation sites during bidirectional synaptic plasticity. Nature 405: 955-959. 165. Kirkwood A, Silva A and Bear MF (1997) Age-dependent decrease of synaptic plasticity in the neocortex of alphaCaMKII mutant mice. Proc Natl Acad Sci U S A 94: 3380-3383. 166. Hardingham N, Glazewski S, Pakhotin P, Mizuno K, Chapman PF, et al. (2003) Neocortical long-term potentiation and experience-dependent synaptic plasticity require alpha-calcium/calmodulin-dependent protein kinase II autophosphorylation. J Neurosci 23: 4428-4436.

33
167. Hasegawa Y, Mukai H, Asashima M, Hojo Y, Ikeda M, et al. (2014) Acute modulation of synaptic plasticity of pyramidal neurons by activin in adult hippocampus. Front Neural Circuits 8: 56. 168. Zhu G, Liu Y, Wang Y, Bi X and Baudry M (2015) Different patterns of electrical activity lead to long-term potentiation by activating different intracellular pathways. J Neurosci 35: 621-633. 169. Esteban JA, Shi SH, Wilson C, Nuriya M, Huganir RL, et al. (2003) PKA phosphorylation of AMPA receptor subunits controls synaptic trafficking underlying plasticity. Nat Neurosci 6: 136-143. 170. Kim CH, Chung HJ, Lee HK and Huganir RL (2001) Interaction of the AMPA receptor subunit GluR2/3 with PDZ domains regulates hippocampal long-term depression. Proc Natl Acad Sci U S A 98: 11725-11730. 171. Soderling TR and Derkach VA (2000) Postsynaptic protein phosphorylation and LTP. Trends Neurosci 23: 75-80. 172. Lee HK, Takamiya K, Han JS, Man H, Kim CH, et al. (2003) Phosphorylation of the AMPA receptor GluR1 subunit is required for synaptic plasticity and retention of spatial memory. Cell 112: 631-643. 173. Derkach VA, Oh MC, Guire ES and Soderling TR (2007) Regulatory mechanisms of AMPA receptors in synaptic plasticity. Nat Rev Neurosci 8: 101-113. 174. Huganir RL and Nicoll RA (2013) AMPARs and synaptic plasticity: the last 25 years. Neuron 80: 704-717. 175. Fritschy JM and Brunig I (2003) Formation and plasticity of GABAergic synapses: physiological mechanisms and pathophysiological implications. Pharmacology & therapeutics 98: 299-323. 176. Kralic JE, Korpi ER, O'Buckley TK, Homanics GE and Morrow AL (2002) Molecular and pharmacological characterization of GABA(A) receptor alpha1 subunit knockout mice. The Journal of pharmacology and experimental therapeutics 302: 1037-1045. 177. Tomita S, Sekiguchi M, Wada K, Nicoll RA and Bredt DS (2006) Stargazin controls the pharmacology of AMPA receptor potentiators. Proc Natl Acad Sci U S A 103: 10064-10067. 178. Cheng J, Dong J, Cui Y, Wang L, Wu B, et al. (2012) Interacting partners of AMPA-type glutamate receptors. J Mol Neurosci 48: 441-447. 179. Nicoll RA, Tomita S and Bredt DS (2006) Auxiliary subunits assist AMPA-type glutamate receptors. Science 311: 1253-1256. 180. Ziff EB (2007) TARPs and the AMPA receptor trafficking paradox. Neuron 53: 627-633. 181. Kato AS, Zhou W, Milstein AD, Knierman MD, Siuda ER, et al. (2007) New transmembrane AMPA receptor regulatory protein isoform, gamma-7, differentially regulates AMPA receptors. J Neurosci 27: 4969-4977. 182. Milstein AD and Nicoll RA (2008) Regulation of AMPA receptor gating and pharmacology by TARP auxiliary subunits. Trends Pharmacol Sci 29: 333-339. 183. Letts VA, Felix R, Biddlecome GH, Arikkath J, Mahaffey CL, et al. (1998) The mouse stargazer gene encodes a neuronal Ca2+-channel gamma subunit. Nat Genet 19: 340-347. 184. Menuz K and Nicoll RA (2008) Loss of inhibitory neuron AMPA receptors contributes to ataxia and epilepsy in stargazer mice. J Neurosci 28: 10599-10603. 185. Osten P and Stern-Bach Y (2006) Learning from stargazin: the mouse, the phenotype and the unexpected. Curr Opin Neurobiol 16: 275-280.

34
186. Haering SC, Tapken D, Pahl S and Hollmann M (2014) Auxiliary subunits: shepherding AMPA receptors to the plasma membrane. Membranes (Basel) 4: 469-490. 187. Vandenberghe W, Nicoll RA and Bredt DS (2005) Stargazin is an AMPA receptor auxiliary subunit. Proc Natl Acad Sci U S A 102: 485-490. 188. Bats C, Soto D, Studniarczyk D, Farrant M and Cull-Candy SG (2012) Channel properties reveal differential expression of TARPed and TARPless AMPARs in stargazer neurons. Nat Neurosci 15: 853-861. 189. Shi Y, Lu W, Milstein AD and Nicoll RA (2009) The stoichiometry of AMPA receptors and TARPs varies by neuronal cell type. Neuron 62: 633-640. 190. Jackson AC and Nicoll RA (2011) The expanding social network of ionotropic glutamate receptors: TARPs and other transmembrane auxiliary subunits. Neuron 70: 178-199. 191. Meng H, Walker N, Su Y and Qiao X (2006) Stargazin mutation impairs cerebellar synaptogenesis, synaptic maturation and synaptic protein distribution. Brain Res 1124: 197-207. 192. Milstein AD and Nicoll RA (2009) TARP modulation of synaptic AMPA receptor trafficking and gating depends on multiple intracellular domains. Proc Natl Acad Sci U S A 106: 11348-11351. 193. Jackson AC and Nicoll RA (2011) Stargazin (TARP gamma-2) is required for compartment-specific AMPA receptor trafficking and synaptic plasticity in cerebellar stellate cells. J Neurosci 31: 3939-3952. 194. Straub C and Tomita S (2012) The regulation of glutamate receptor trafficking and function by TARPs and other transmembrane auxiliary subunits. Curr Opin Neurobiol 22: 488-495. 195. Sumioka A (2013) Auxiliary subunits provide new insights into regulation of AMPA receptor trafficking. J Biochem 153: 331-337. 196. Barker CA, Zeng X, Bettington S, Batsanov AS, Bryce MR, et al. (2007) Porphyrin, phthalocyanine and porphyrazine derivatives with multifluorenyl substituents as efficient deep-red emitters. Chemistry 13: 6710-6717. 197. Chen L, El-Husseini A, Tomita S, Bredt DS and Nicoll RA (2003) Stargazin differentially controls the trafficking of alpha-amino-3-hydroxyl-5-methyl-4-isoxazolepropionate and kainate receptors. Mol Pharmacol 64: 703-706. 198. Chen L, Chetkovich DM, Petralia RS, Sweeney NT, Kawasaki Y, et al. (2000) Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 408: 936-943. 199. Rouach N, Byrd K, Petralia RS, Elias GM, Adesnik H, et al. (2005) TARP gamma-8 controls hippocampal AMPA receptor number, distribution and synaptic plasticity. Nat Neurosci 8: 1525-1533. 200. Tomita S, Adesnik H, Sekiguchi M, Zhang W, Wada K, et al. (2005) Stargazin modulates AMPA receptor gating and trafficking by distinct domains. Nature 435: 1052-1058. 201. Kato AS, Siuda ER, Nisenbaum ES and Bredt DS (2008) AMPA receptor subunit-specific regulation by a distinct family of type II TARPs. Neuron 59: 986-996. 202. Menuz K, Stroud RM, Nicoll RA and Hays FA (2007) TARP auxiliary subunits switch AMPA receptor antagonists into partial agonists. Science 318: 815-817. 203. Soto D, Coombs ID, Kelly L, Farrant M and Cull-Candy SG (2007) Stargazin attenuates intracellular polyamine block of calcium-permeable AMPA receptors. Nat Neurosci 10: 1260-1267.

35
204. Jackson AC, Milstein AD, Soto D, Farrant M, Cull-Candy SG, et al. (2011) Probing TARP modulation of AMPA receptor conductance with polyamine toxins. J Neurosci 31: 7511-7520. 205. Maclean DM and Bowie D (2011) Transmembrane AMPA receptor regulatory protein regulation of competitive antagonism: a problem of interpretation. J Physiol 589: 5383-5390. 206. Cokic B and Stein V (2008) Stargazin modulates AMPA receptor antagonism. Neuropharmacology 54: 1062-1070. 207. Turetsky D, Garringer E and Patneau DK (2005) Stargazin modulates native AMPA receptor functional properties by two distinct mechanisms. J Neurosci 25: 7438-7448. 208. Priel A, Kolleker A, Ayalon G, Gillor M, Osten P, et al. (2005) Stargazin reduces desensitization and slows deactivation of the AMPA-type glutamate receptors. J Neurosci 25: 2682-2686. 209. Sager C, Terhag J, Kott S and Hollmann M (2009) C-terminal domains of transmembrane alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA) receptor regulatory proteins not only facilitate trafficking but are major modulators of AMPA receptor function. J Biol Chem 284: 32413-32424. 210. Khodosevich K, Jacobi E, Farrow P, Schulmann A, Rusu A, et al. (2014) Coexpressed auxiliary subunits exhibit distinct modulatory profiles on AMPA receptor function. Neuron 83: 601-615. 211. Soto D, Coombs ID, Gratacos-Batlle E, Farrant M and Cull-Candy SG (2014) Molecular mechanisms contributing to TARP regulation of channel conductance and polyamine block of calcium-permeable AMPA receptors. J Neurosci 34: 11673-11683. 212. Yamazaki M, Ohno-Shosaku T, Fukaya M, Kano M, Watanabe M, et al. (2004) A novel action of stargazin as an enhancer of AMPA receptor activity. Neurosci Res 50: 369-374. 213. Bats C, Farrant M and Cull-Candy SG (2013) A role of TARPs in the expression and plasticity of calcium-permeable AMPARs: evidence from cerebellar neurons and glia. Neuropharmacology 74: 76-85. 214. Yokoi N, Fukata M and Fukata Y (2012) Synaptic plasticity regulated by protein-protein interactions and posttranslational modifications. Int Rev Cell Mol Biol 297: 1-43. 215. Schwenk J, Harmel N, Zolles G, Bildl W, Kulik A, et al. (2009) Functional proteomics identify cornichon proteins as auxiliary subunits of AMPA receptors. Science 323: 1313-1319. 216. Shi Y, Suh YH, Milstein AD, Isozaki K, Schmid SM, et al. (2010) Functional comparison of the effects of TARPs and cornichons on AMPA receptor trafficking and gating. Proc Natl Acad Sci U S A 107: 16315-16319. 217. Drummond JB, Simmons M, Haroutunian V and Meador-Woodruff JH (2012) Upregulation of cornichon transcripts in the dorsolateral prefrontal cortex in schizophrenia. Neuroreport 23: 1031-1034. 218. Herring BE, Shi Y, Suh YH, Zheng CY, Blankenship SM, et al. (2013) Cornichon proteins determine the subunit composition of synaptic AMPA receptors. Neuron 77: 1083-1096. 219. Klaassen RV, Stroeder J, Coussen F, Hafner AS, Petersen JD, et al. (2016) Shisa6 traps AMPA receptors at postsynaptic sites and prevents their desensitization during synaptic activity. Nat Commun 7.

36
220. Farrow P, Khodosevich K, Sapir Y, Schulmann A, Aslam M, et al. (2015) Auxiliary subunits of the CKAMP family differentially modulate AMPA receptor properties. Elife 4.

37
Chapter2
Alpha2‐containingGABAARsexpressedinhippocampal
regionCA3controlfastnetworkoscillations.Tim S. Heistek, Marta Ruiperez-Alonso, A. Jaap Timmerman, Arjen B. Brussaard and Huibert D. Mansvelder
Dept. Integrative Neurophysiology, Center for Neurogenomics and Cognitive Research, Neuroscience Campus Amsterdam, VU University, Amsterdam, The Netherlands.

38


















