
Future trends in neurovirology: Neuronal survival during virus
9
Journal of NeuroVirology, 10: 207–215, 2004 c 2004 Journal of NeuroVirology ISSN: 1355-0284 print / 1538-2443 online DOI: 10.1080/13550280490468438 2003 Pioneer in NeuroVirology Review Future trends in neurovirology: Neuronal survival during virus infection and analysis of virus-specific T cells in central nervous system tissues Michael BA Oldstone Division of Virology, Department of Neuropharmacology, The Scripps Research Institute, La Jolla, California, USA The recent ability to quantitate and localize the interaction of antigen-specific T cells in the central nervous system during infectious and autoimmune dis- eases will greatly enhance our understanding of the molecular and biologic basis of virus-induced diseases of the nervous and other systems. Journal of NeuroVirology (2004) 10, 207–215. Keywords: MMHC expression; neurons; TAP expression; TCR: T cell receptor; trafficking of antigen-specific T cells in the CNS1; virus Setting the stage Viral diseases of the central nervous system (CNS) have been known since antiquity. Over 2300 years before the first viruses were isolated, Homer men- tioned the Greek word for rabies, “lyssa,” in The Il- liad (reviewed in Kaplan and Meslin, 1996; Waterson and Wilkinson, 1978). About 400 years later, Aris- totle presented a clear picture of rabies in animals, although he failed to realize that humans could be similarly infected. But not until the 16th century did Fracastoro (1546) provide a clear description of rabies infection of man. Even before those early published depictions of viral disease, the first suspected case of poliomyelitis came from an Egyptian stele dating from the 18th dynasty of 1580 to 1350 BC (reviewed in Koprowski, 1996; Oldstone, 1998). Ancient hiero- glyphic records describing the pharaoh Ramses with a limb withered during childhood suggest that po- liomyelitis virus infection caused the deformity, al- though trauma, birth defect, or vascular insufficiency remain as possibilities. Isolation of the rabies and po- liomyelitis agents and identification of both as a virus would await the early 20th century. The understand- ing that disease occurred in organs or tissues such as Address correspondence to Michael B. A. Oldstone, MD, Divi- sion of Virology, Department of Neuropharmacology, The Scripps Research Institute, 10550 N. Torrey Pines Road, La Jolla, CA 92037, USA. E-mail: [email protected] This is publication number 16404-NP from the Department of Neuropharmacology, The Scripps Research Institute, La Jolla, CA. This work was supported in part by USPHS grants AI09484, AI36222, DK58541, and AI45927. Received 5 April 2004; accepted 5 April 2004. the brain rather than disturbing the balance among the four humors—fluids of the body: blood, phlegm, and yellow and black bile—derived from anatomic observations of only 233 years ago. So the science of pathology and modern medicine began, as exem- plified by the work of Giovanni Battista Morgagni in 1761. His publication, “On the sites and causes of dis- eases as disclosed by anatomic dissection,” compared alterations in the disease-ridden tissues of patients to the state of healthy, normal tissues. Neurovirology: virus-neuron interactions and major histocompatibility complex (MHC) expression in the CNS Neurotropic viruses can enter the CNS by two main routes, as outlined in Table 1. Most viruses gain ac- cess to the CNS by a hematogenous, or blood-borne route, which requires them to cross from a blood ves- sel into the brain parenchyma via the blood-brain barrier. These viruses can travel in plasma, bound to cells (i.e., red blood cells) or be contained inside cells (i.e., lymphocytes and/or macrophages). Other viruses, such as rabies, travel in a retrograde fashion along axon processes, transmitted trans-synaptically into the CNS. Neurons are essential to the host’s survival and are irreplaceable once destroyed, the exception be- ing olfactory neurons. Therefore, neurotropic viral infections that directly cause neuronal death, or in- duce an aggressive antineuronal or antiviral immune response should permanently deplete the body’s store of vital cells. Likely through selection and be- cause of their essential role in host survival, CNS
Transcript of Future trends in neurovirology: Neuronal survival during virus

Journal of NeuroVirology, 10: 207–215, 2004c© 2004 Journal of NeuroVirologyISSN: 1355-0284 print / 1538-2443 onlineDOI: 10.1080/13550280490468438
2003 Pioneer in NeuroVirology Review
Future trends in neurovirology: Neuronal survivalduring virus infection and analysis of virus-specificT cells in central nervous system tissues
Michael BA Oldstone
Division of Virology, Department of Neuropharmacology, The Scripps Research Institute, La Jolla, California, USA
The recent ability to quantitate and localize the interaction of antigen-specificT cells in the central nervous system during infectious and autoimmune dis-eases will greatly enhance our understanding of the molecular and biologicbasis of virus-induced diseases of the nervous and other systems. Journal ofNeuroVirology (2004) 10, 207–215.
Keywords: MMHC expression; neurons; TAP expression; TCR: T cell receptor;trafficking of antigen-specific T cells in the CNS1; virus
Setting the stage
Viral diseases of the central nervous system (CNS)have been known since antiquity. Over 2300 yearsbefore the first viruses were isolated, Homer men-tioned the Greek word for rabies, “lyssa,” in The Il-liad (reviewed in Kaplan and Meslin, 1996; Watersonand Wilkinson, 1978). About 400 years later, Aris-totle presented a clear picture of rabies in animals,although he failed to realize that humans could besimilarly infected. But not until the 16th century didFracastoro (1546) provide a clear description of rabiesinfection of man. Even before those early publisheddepictions of viral disease, the first suspected caseof poliomyelitis came from an Egyptian stele datingfrom the 18th dynasty of 1580 to 1350 BC (reviewedin Koprowski, 1996; Oldstone, 1998). Ancient hiero-glyphic records describing the pharaoh Ramses witha limb withered during childhood suggest that po-liomyelitis virus infection caused the deformity, al-though trauma, birth defect, or vascular insufficiencyremain as possibilities. Isolation of the rabies and po-liomyelitis agents and identification of both as a viruswould await the early 20th century. The understand-ing that disease occurred in organs or tissues such as
Address correspondence to Michael B. A. Oldstone, MD, Divi-sion of Virology, Department of Neuropharmacology, The ScrippsResearch Institute, 10550 N. Torrey Pines Road, La Jolla, CA 92037,USA. E-mail: [email protected]
This is publication number 16404-NP from the Departmentof Neuropharmacology, The Scripps Research Institute, La Jolla,CA. This work was supported in part by USPHS grants AI09484,AI36222, DK58541, and AI45927.
Received 5 April 2004; accepted 5 April 2004.
the brain rather than disturbing the balance amongthe four humors—fluids of the body: blood, phlegm,and yellow and black bile—derived from anatomicobservations of only 233 years ago. So the scienceof pathology and modern medicine began, as exem-plified by the work of Giovanni Battista Morgagni in1761. His publication, “On the sites and causes of dis-eases as disclosed by anatomic dissection,” comparedalterations in the disease-ridden tissues of patients tothe state of healthy, normal tissues.
Neurovirology: virus-neuron interactions andmajor histocompatibility complex (MHC)expression in the CNS
Neurotropic viruses can enter the CNS by two mainroutes, as outlined in Table 1. Most viruses gain ac-cess to the CNS by a hematogenous, or blood-borneroute, which requires them to cross from a blood ves-sel into the brain parenchyma via the blood-brainbarrier. These viruses can travel in plasma, boundto cells (i.e., red blood cells) or be contained insidecells (i.e., lymphocytes and/or macrophages). Otherviruses, such as rabies, travel in a retrograde fashionalong axon processes, transmitted trans-synapticallyinto the CNS.
Neurons are essential to the host’s survival andare irreplaceable once destroyed, the exception be-ing olfactory neurons. Therefore, neurotropic viralinfections that directly cause neuronal death, or in-duce an aggressive antineuronal or antiviral immuneresponse should permanently deplete the body’sstore of vital cells. Likely through selection and be-cause of their essential role in host survival, CNS

Tracking virus and immune response interactions with neurons208 MBA Oldstone
Table 1 Examples of the route taken by neurotropic viruses toaccess the central nervous system
Hematogenous Neural pathwayHerpes simplex Herpes simplexCytomegalovirus Varicella zosterEpstein-Barr virus RabiesPoliovirus PoliovirusMeasles ReovirusCoxsackievirusesHuman immunodeficiency virus (HIV)MumpsEchovirusesLymphocytic choriomeningitisArboviruses
neurons rarely express class I or II MHC molecules(Horwitz et al, 1996, 1997; Joly et al, 1991; Keaneet al, 1992; Mucke and Oldstone, 1992). Some excep-tions are a limited number of selected neurons thatexpress MHC molecules at minimal levels (Redwineet al, 2001), MHC molecules expressed during neu-ron development and believed to play a role in synap-tic plasticity (Corriveau et al, 1998; Huh et al, 2000),MHC expression in mature neurons that have lostelectrical activity (i.e., dying or dead) (Hausmannet al, 2001; Neumann et al, 1995), and those identi-fied in graft-versus-host studies (Hickey and Kimura,1987).
MHC class I molecules must be present on vi-rally infected target cells for their recognition anddestruction by cytotoxic T lymphocytes (CTLs). Inthe absence of MHC antigen presentation, CTLs failto see virally infected neurons despite the presenceof viral proteins on their surfaces. Interestingly, evenwhen interferon (IFN)-γ , a strong up-regulator ofMHC molecules is overexpressed in the CNS in exper-iments that use a transgenic technique (Horwitz et al,1999, 1997), neither MHC class I nor MHC class II pro-tein molecules can be induced on neurons. Moreover,these MHC protein molecules, although not consti-tutively expressed on either astrocytes or oligoden-drocytes, can be markedly induced by IFN-γ only onoligodendrocytes but not astrocytes. Thus, in murinemodels only vital oligodendrocytes, but not neuronsnor astrocytes, would likely have the ability to act astargets for antigen-specific T cells. MHC moleculesare constitutively expressed on microglia and bloodvessels and up-regulated by IFN. In addition to fail-ing to express MHC proteins on their surfaces, neu-rons lack other molecules necessary for antigen pro-cessing, as shown in vitro. When a virus infects acell, the virus uncoats and synthesizes viral proteins,which are either incorporated into progeny virus ordegraded into small peptides of 9 to 11 amino acids,mediated by the proteasome complex. These pep-tides are then shuttled into the endoplasmic retic-ulum by TAP1 and TAP2 proteins, where they en-counter the host cell–encoded class I MHC: the heavychain and the light chain (β2-microglobulin). Thecomplex consisting of the class I heavy chain, the β2-
microglobulin, and the viral peptide is transportedto the cell surface, where the receptor of a specificCTL recognizes the complex, leading to the infectedcell’s death (reviewed in Whitton and Oldstone,2001).
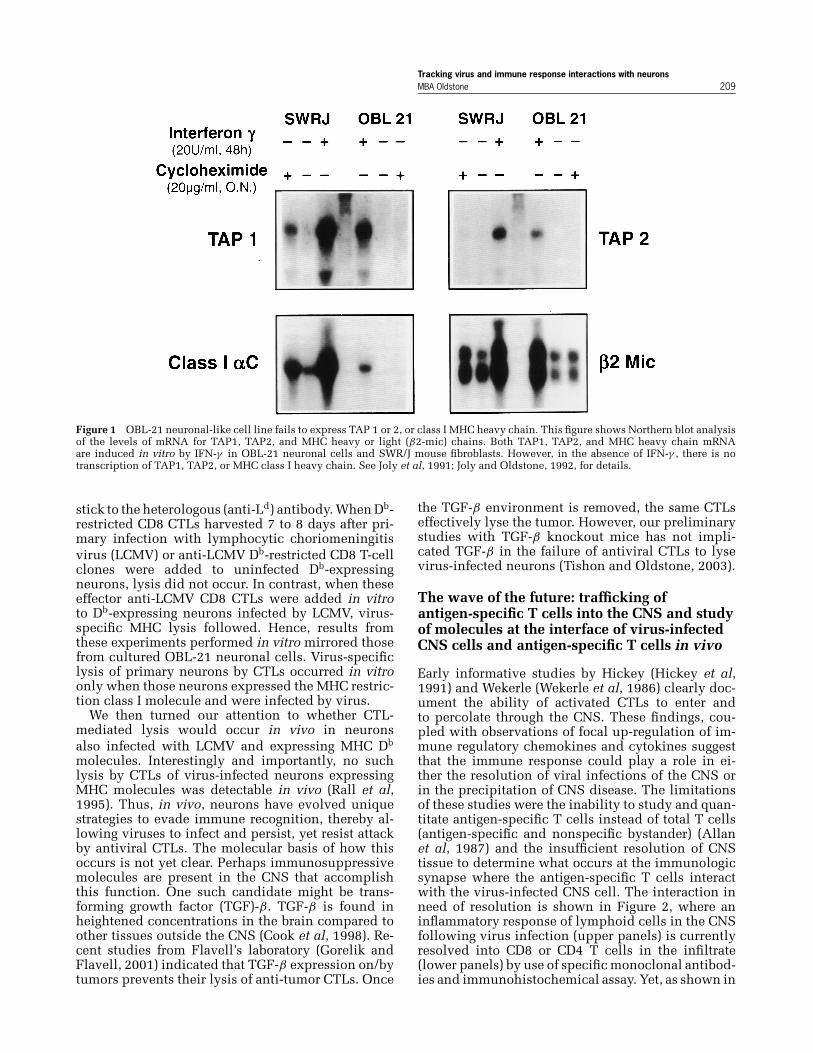
Study of OBL-21 neuronal cells in vitro indicatesthat at least two defects have the power to makeantigen presentation of viral peptides to the CTLsunlikely (Joly et al, 1991; Joly and Oldstone, 1992).These neuronal cells fail to transcribe the mRNAencoding the class I MHC heavy chain and fail totranscribe the mRNA encoding the TAP1 and TAP2molecules (Figure 1). The consequence is that thesecultured neurons are unable to present viral peptidein the context of class I MHC and, therefore, do notserve as targets for CTL-mediated lysis (Joly et al,1991; Joly and Oldstone, 1992). However, whenOBL-21 cells are treated with IFN-γ , a molecule thatup-regulates endogenous MHC, or are transfectedwith a retroviral vector encoding a heterologous MHCmolecule (Db), surface expression of the class I MHCmolecule occurs so that CTL-mediated lysis becomespossible. Thus, reconstitution of class I MHC in vitrorestores lysability of neurons by CTLs. However,in vivo treatment with excessive amounts of IFN-γ –induced focally in the CNS fails to induce MHCmolecules (Horwitz et al, 1999, 1997). Further, unlikethe situation in vitro, complementation of MHC classI molecules into and on neurons by means of trans-genic techniques fails to make such virally infectedneurons expressing MHC class I molecules suscep-tible to CTL-mediated lysis in vivo (Rall et al, 1995).
We generated transgenic mice expressing a classI MHC molecule driven by a neuronal-specific eno-lase promoter (Rall et al, 1995). Our initial thoughtwas that mouse neurons now expressing a functionalclass I MHC molecule should serve as targets forvirus-specific CTL-mediated lysis. Further, such amodel would allow one to address questions of sen-sitized CTLs’ movement across the blood-brain bar-rier and the consequences to a host of expressingMHC molecules in neurons with and without viralinfection and in the presence of an antiviral CTL re-sponse. The MHC class I Db mRNA was detected inbrains of transgenic mice but not in their other tis-sues such as thymus, spleen, pancreas, lung, liver,or testes, or in the peripheral nervous system (sci-atic and optic nerves). Protein synthesis of the trans-gene was identified in primary hippocampal neu-rons with a monoclonal antibody that recognized thecorrect folding of Db-containing Db-restricted pep-tides. When cultured, isolated neurons were placedonto cover slips precoated either with an antibodydirected against the transgene (Db) or against a het-erologous class I MHC gene (Ld), only transgenicneurons in contact with the anti-Db antibody ad-hered and aborized (Rall et al, 1995). Nontransgenicneurons did not stick to either the anti-Db– or theanti-Ld–coated slips, and transgenic neurons did not
Tracking virus and immune response interactions with neuronsMBA Oldstone 209
Figure 1 OBL-21 neuronal-like cell line fails to express TAP 1 or 2, or class I MHC heavy chain. This figure shows Northern blot analysisof the levels of mRNA for TAP1, TAP2, and MHC heavy or light (β2-mic) chains. Both TAP1, TAP2, and MHC heavy chain mRNAare induced in vitro by IFN-γ in OBL-21 neuronal cells and SWR/J mouse fibroblasts. However, in the absence of IFN-γ , there is notranscription of TAP1, TAP2, or MHC class I heavy chain. See Joly et al, 1991; Joly and Oldstone, 1992, for details.
stick to the heterologous (anti-Ld) antibody. When Db-restricted CD8 CTLs harvested 7 to 8 days after pri-mary infection with lymphocytic choriomeningitisvirus (LCMV) or anti-LCMV Db-restricted CD8 T-cellclones were added to uninfected Db-expressingneurons, lysis did not occur. In contrast, when theseeffector anti-LCMV CD8 CTLs were added in vitroto Db-expressing neurons infected by LCMV, virus-specific MHC lysis followed. Hence, results fromthese experiments performed in vitro mirrored thosefrom cultured OBL-21 neuronal cells. Virus-specificlysis of primary neurons by CTLs occurred in vitroonly when those neurons expressed the MHC restric-tion class I molecule and were infected by virus.
We then turned our attention to whether CTL-mediated lysis would occur in vivo in neuronsalso infected with LCMV and expressing MHC Db
molecules. Interestingly and importantly, no suchlysis by CTLs of virus-infected neurons expressingMHC molecules was detectable in vivo (Rall et al,1995). Thus, in vivo, neurons have evolved uniquestrategies to evade immune recognition, thereby al-lowing viruses to infect and persist, yet resist attackby antiviral CTLs. The molecular basis of how thisoccurs is not yet clear. Perhaps immunosuppressivemolecules are present in the CNS that accomplishthis function. One such candidate might be trans-forming growth factor (TGF)-β. TGF-β is found inheightened concentrations in the brain compared toother tissues outside the CNS (Cook et al, 1998). Re-cent studies from Flavell’s laboratory (Gorelik andFlavell, 2001) indicated that TGF-β expression on/bytumors prevents their lysis of anti-tumor CTLs. Once
the TGF-β environment is removed, the same CTLseffectively lyse the tumor. However, our preliminarystudies with TGF-β knockout mice has not impli-cated TGF-β in the failure of antiviral CTLs to lysevirus-infected neurons (Tishon and Oldstone, 2003).
The wave of the future: trafficking ofantigen-specific T cells into the CNS and studyof molecules at the interface of virus-infectedCNS cells and antigen-specific T cells in vivo
Early informative studies by Hickey (Hickey et al,1991) and Wekerle (Wekerle et al, 1986) clearly doc-ument the ability of activated CTLs to enter andto percolate through the CNS. These findings, cou-pled with observations of focal up-regulation of im-mune regulatory chemokines and cytokines suggestthat the immune response could play a role in ei-ther the resolution of viral infections of the CNS orin the precipitation of CNS disease. The limitationsof these studies were the inability to study and quan-titate antigen-specific T cells instead of total T cells(antigen-specific and nonspecific bystander) (Allanet al, 1987) and the insufficient resolution of CNStissue to determine what occurs at the immunologicsynapse where the antigen-specific T cells interactwith the virus-infected CNS cell. The interaction inneed of resolution is shown in Figure 2, where aninflammatory response of lymphoid cells in the CNSfollowing virus infection (upper panels) is currentlyresolved into CD8 or CD4 T cells in the infiltrate(lower panels) by use of specific monoclonal antibod-ies and immunohistochemical assay. Yet, as shown in

Tracking virus and immune response interactions with neurons210 MBA Oldstone
Figure 2 Histologic picture of cells infiltrating the CNS as judged by H and E stain (upper panels) and immunohistochemical analysis oftotal CD4 or CD8 T cells (lower panels). Missing is knowledge of which of the cells is antigen-specific to the disease process under studyas compared to nonspecific T cells or macrophages, NK cells, etc., that are called into the inflammatory nidus.
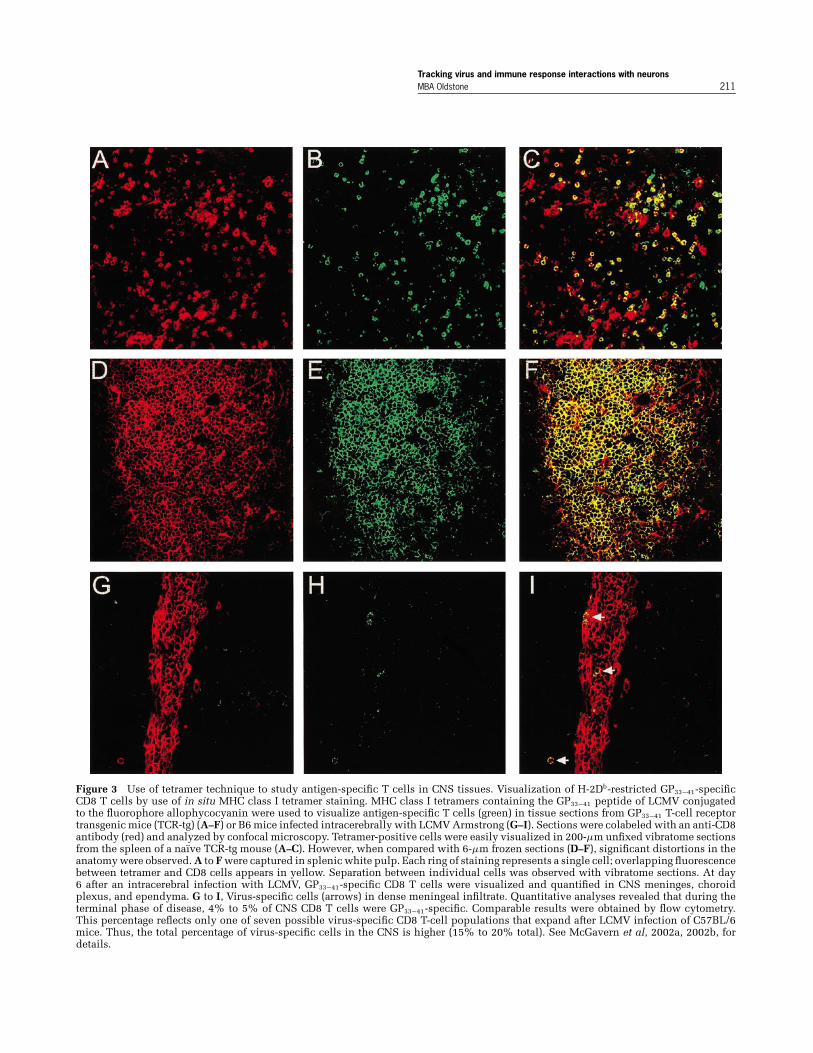
the lower panel of Figure 3, when the number of anti-gen (virus)-specific T cells is defined, they make uponly a minority, between 2% to 4%, of the total T-cellresponse to a single immunodominant T-cell epitope(Figure 3) or up to 15% to 20% for all the dominantand subdominant T-cell epitopes restricted by theMHC allele. The importance of these antigen-specificT cells is that, when they are blocked in the appro-priate model, immunopathologic disease does not oc-cur. Hence, only these few cells are the focus of ourattention, not the vast majority of nonactive, nonpar-ticipating bystander T cells (McGavern and Truong,2004). It is these antigen (virus)-specific T cells thatone must follow in their generation, expansion, andtrafficking to the CNS and as they interact with virus-infected cells in the CNS. Identifying the moleculesrequired to turn on or turn off effector function is thechallenge of the future. The novel technology devel-oped and refined just over a year ago in 2002 solely byDorian McGavern in my laboratory (McGavern et al,2002a; McGavern and Truong, 2004; McGavern et al,2002b) (Figure 4) allows the visualization and iden-tification at high resolution of interactions between
either antigen (virus)-specific CD8 or CD4 T cells andvirus-infected cells in vivo (Figures 3, 4).
Earlier strategies to mark antigen-specific lympho-cytes as they moved to the CNS or other tissues(Flugel et al, 2001; Flugel and Bradl, 2001; Reinhardtet al, 2001; Skinner et al, 2000) primarily usedone of two techniques. The first was in vitro ma-nipulation to label T cells of interest, i.e., CD4 Tcells that cause allergic encephalitis were transducedwith a retroviral vector engineered to express greenfluorescent protein (GFP) (Flugel et al, 2001). Thesecond implemented a model in which ovalbumin(OVA)-specific CD4 Th1.1 T cells were transferredinto Th1.2 congenic mice then treated with a mono-clonal antibody to detect the Th1.1 marker (Reinhardtet al, 2001). Other studies used tetramers to visual-ize OVA-labeled T cells (Skinner et al, 2000). Thesestudies provided the foundation for the innovativemethod later developed by McGavern (McGavernet al, 2002a) (Figure 5). The strength of McGavern’sstudies is twofold. First, he has extended the ear-lier tetramer strategy (Skinner et al, 2000) by de-veloping techniques to use six micron sections
Tracking virus and immune response interactions with neuronsMBA Oldstone 211
Figure 3 Use of tetramer technique to study antigen-specific T cells in CNS tissues. Visualization of H-2Db-restricted GP33−41-specificCD8 T cells by use of in situ MHC class I tetramer staining. MHC class I tetramers containing the GP33−41 peptide of LCMV conjugatedto the fluorophore allophycocyanin were used to visualize antigen-specific T cells (green) in tissue sections from GP33−41 T-cell receptortransgenic mice (TCR-tg) (A–F) or B6 mice infected intracerebrally with LCMV Armstrong (G–I). Sections were colabeled with an anti-CD8antibody (red) and analyzed by confocal microscopy. Tetramer-positive cells were easily visualized in 200-µm unfixed vibratome sectionsfrom the spleen of a naı̈ve TCR-tg mouse (A–C). However, when compared with 6-µm frozen sections (D–F), significant distortions in theanatomy were observed. A to F were captured in splenic white pulp. Each ring of staining represents a single cell; overlapping fluorescencebetween tetramer and CD8 cells appears in yellow. Separation between individual cells was observed with vibratome sections. At day6 after an intracerebral infection with LCMV, GP33−41-specific CD8 T cells were visualized and quantified in CNS meninges, choroidplexus, and ependyma. G to I, Virus-specific cells (arrows) in dense meningeal infiltrate. Quantitative analyses revealed that during theterminal phase of disease, 4% to 5% of CNS CD8 T cells were GP33−41-specific. Comparable results were obtained by flow cytometry.This percentage reflects only one of seven possible virus-specific CD8 T-cell populations that expand after LCMV infection of C57BL/6mice. Thus, the total percentage of virus-specific cells in the CNS is higher (15% to 20% total). See McGavern et al, 2002a, 2002b, fordetails.

Tracking virus and immune response interactions with neurons212 MBA Oldstone
Figure 4 The use of GFP × TCR technique to study antigen-specific T cells in the CNS. The upper three panel rows show the antigen-specific CD8 T cells (first rows, in blue), the presence of viral antigen in CNS infected leptomeningeal cells (second row, in red), andstaining for molecules of interest at the immunologic synapse in the CNS: adhesion molecule LFA-1, T-cell activation molecule LCK, andT-cell effector molecule perforin (PFP) (third row, in green) using monoclonal antibodies to these molecules labeled with a fluorochromedye. The last row (right row) shows the merge. The two enlarged panels at the bottom provide the initial documentation that one antigen-specific CD8 T cells (lower left, in red) can engage multiple, in this case three, virus-infected target cells (lower right, in green). Theimmune synapse is the yellow color. The large panel on the lower right shows antigen-specific CD8 T cells (blue) that release perforin(PFP: yellow) onto two virus-infected target cells (red). Thus, antigen-specific T cells have a greater hit in this one snapshot in time thanpreviously believed. See McGavern et al, 2002a, for details. Our and other in vitro kinetic studies have shown that one antigen-specificCD8 T cell can attack and lyse one target, then disengages to attack kinetically multiple targets, also emphasizing the effectiveness of asingle antigen-specific T cell. See McGavern et al, 2002a, 2002b, for details.
(instead of 200-µ sections used by Skinner et al), se-lecting two dyes that enhance absorption and emis-sion at the same or closely related wavelengths,thereby potentiating the signal and employing con-focal microscopy, which dramatically improves theresolution of cells under investigation. The result ishigher resolution microscopy than was previouslypossible, and this enabled fine dissection of the im-
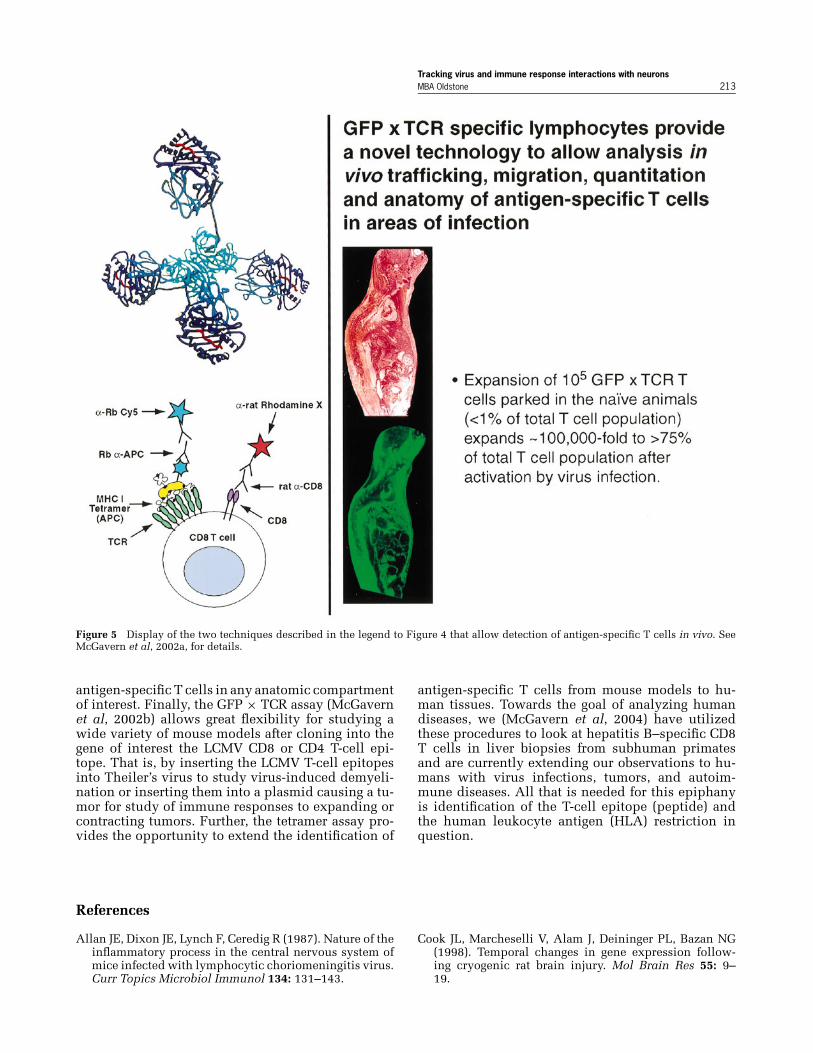
mune synapse in vivo. In addition, the invention ofthe GFP × TCR (T cell receptor) adoptive transfer pro-cedure (Figures 4, 5), in conjunction with a peripheralvirus infection (virus recognized by adoptively trans-ferred TCR CD8 or CD4 T cell) allows enormous ex-pansion of antigen (virus)-specific T cells (from <1%of the total T-cell repertoire to >75%). We now havethe ability to dissect cellular trafficking and localize
Tracking virus and immune response interactions with neuronsMBA Oldstone 213
Figure 5 Display of the two techniques described in the legend to Figure 4 that allow detection of antigen-specific T cells in vivo. SeeMcGavern et al, 2002a, for details.
antigen-specific T cells in any anatomic compartmentof interest. Finally, the GFP × TCR assay (McGavernet al, 2002b) allows great flexibility for studying awide variety of mouse models after cloning into thegene of interest the LCMV CD8 or CD4 T-cell epi-tope. That is, by inserting the LCMV T-cell epitopesinto Theiler’s virus to study virus-induced demyeli-nation or inserting them into a plasmid causing a tu-mor for study of immune responses to expanding orcontracting tumors. Further, the tetramer assay pro-vides the opportunity to extend the identification of
References
Allan JE, Dixon JE, Lynch F, Ceredig R (1987). Nature of theinflammatory process in the central nervous system ofmice infected with lymphocytic choriomeningitis virus.Curr Topics Microbiol Immunol 134: 131–143.
antigen-specific T cells from mouse models to hu-man tissues. Towards the goal of analyzing humandiseases, we (McGavern et al, 2004) have utilizedthese procedures to look at hepatitis B–specific CD8T cells in liver biopsies from subhuman primatesand are currently extending our observations to hu-mans with virus infections, tumors, and autoim-mune diseases. All that is needed for this epiphanyis identification of the T-cell epitope (peptide) andthe human leukocyte antigen (HLA) restriction inquestion.
Cook JL, Marcheselli V, Alam J, Deininger PL, Bazan NG(1998). Temporal changes in gene expression follow-ing cryogenic rat brain injury. Mol Brain Res 55: 9–19.

Tracking virus and immune response interactions with neurons214 MBA Oldstone
Corriveau RA, Huh GS, Shatz CJ (1998). Regulation of classI MHC gene expression in the developing and matureCNS by neural activity. Neuron 21: 505–520.
Flugel A, Berkowicz T, Ritter T, Labeur M, Jenne DE, Li Z,Ellwart JW, Willem M, Lassmann H, Wekerle H (2001).Migratory activity and functional changes of green flu-orescent effector cells before and during experimentalautoimmune encephalomyelitis. Immunity 14: 547–560.
Flugel A, Bradl M (2001). New tools to trace populations ofinflammatory cells in the CNS. Glia 36: 125–136.
Gorelik L, Flavell RA (2001). Immune-mediated eradica-tion of tumors through the blockade of transforminggrowth factor-beta signaling in T cells. Nat Med 7: 1118–1122.
Hausmann J, Schamel K, Staeheli P (2001). CD8(+) Tlymphocytes mediate Borna disease virus-induced im-munopathology independently of perforin. J Virol 75:10460–10466.
Hickey WF, Hsu BL, Kimura H (1991). T lymphocyte entryinto the central nervous system. J Neurosci Res 28: 254–260.
Hickey WF, Kimura H (1987). Graft versus host disease elic-its expression of class I and class II MHC antigens andpresence of scattered T lymphocytes in rat central ner-vous system. Proc Natl Acad Sci U S A 84: 2082–2086.
Horwitz MS, Evans CF, McGavern DB, Rodriguez M,Oldstone MBA (1997). Primary demyelination in trans-genic mice expressing interferon-gamma. Nat Med 3:1037–1041.
Horwitz MS, Evans CF, Oldstone MBA (1999). Expres-sion of major histocompatibility complex class I andII molecules in vivo by oligodendrocytes during viral-mediated central nervous system autoimmune disease.Lab Invest 79: 235–242.
Huh GS, Boulanger LM, Du H, Riquelme PA, Brotz TM,Shatz CJ (2000). Functional requirement for class I MHCin CNS development and plasticity. Science 290: 2051.
Joly E, Mucke L, Oldstone MBA (1991). Viral persistence inneurons explained by lack of major histocompatibilitycomplex class I expression. Science 253: 1283–1285.
Joly E, Oldstone MBA (1992). Neuronal cells are deficientin loading peptides onto MHC class I molecules. Neuron8: 1185–1190.
Kaplan MM, Meslin FX (1996). A brief history of ra-bies. In: Microbe hunters—then and now. Koprowski H,Oldstone MBA (eds). Bloomington, IL: Medi-Ed Press,pp 47–55.
Keane RW, Tallent MW, Podack ER (1992). Resistance andsusceptibility of neural cells to lysis by cytotoxic lym-phocytes and by cytolytic granules. Transplantation 54:520–562.
Koprowski H (1996). Poliomyelitis: visit to ancient his-tory. In: Microbe Hunters—then and now. Koprowski H,Oldstone MBA (eds). Bloomington, IL: Medi-Ed Press,pp 141–152.
McGavern D, Chisari FV, Oldstone MBA (2004). Unpub-lished observations.
McGavern D, Christen U, Oldstone MBA (2002a). Molecu-lar anatomy of antigen-specific CD8+ T cell engagementand synapse formation in vivo. Nat Immunol 3: 918–925.
McGavern D, Truong P (2004). Rebuilding an immune-mediated CNS disease: antigen-specific but not by-stander T cells participate in disease pathogenesis. J Im-munol In press.
McGavern DB, Homann D, Oldstone MBA (2002b). T cellsin the central nervous system: The delicate balance be-
tween viral clearance and disease. J Infect Dis 186: 145–151.
Mucke L, Oldstone MBA (1992). The expression of MHCclass I antigens in the brain differs markedly in acute andpersistent infections with lymphocytic choriomeningi-tis virus. J Neuroimmunol 36: 193–198.
Neumann H, Cavalie A, Jenne DE, Wekerle H (1995). In-duction of MHC class I genes in neurons. Science 270:720–721.
Oldstone MBA (1998). Viruses, plagues, and history.New York, NY: Oxford University Press.
Rall G, Mucke L, Oldstone MBA (1995). Consequencesof cytotoxic T lymphocyte interaction with major his-tocompatibility complex class I-expressing neurons invivo. J Exp Med 182: 1201–1212.
Redwine JM, Buchmeier MJ, Evans CF (2001). In vivo ex-pression of major histocompatibility complex moleculeson oligodendrocytes and neurons during viral infection.Am J Pathol 159: 1219–1224.
Reinhardt RL, Khoruts A, Merica R, Zell T, Jenkins MK(2001). Visualizing the generation of memory CD4 Tcells in the whole body. Nature 410: 101–105.
Skinner PJ, Daniels MA, Schmidt CS, Jameson SC, HaaseAT (2000). Cutting edge: in situ tetramer staining ofantigen-specific T cells in tissues. J Immunol 165: 613–617.
Tishon A, Oldstone MBA (2003). Unpublished observa-tions.
Waterson A, Wilkinson L (1978). An introduction to the his-tory of virology. Cambridge, UK: Cambridge UniversityPress.
Wekerle H, Linington C, Lassmann H, Meyermann R (1986).Cellular immune reactivity within the CNS. Trends Neu-rosci 9: 271–277.
Whitton JL, Oldstone MBA (2001). The immune re-sponse to viruses. In: Fields virology. Knipe D,Howley P, Griffin D, Lamb R, Martin M, Straus S (eds).Philadelphia: Lippincott Williams & Wilkins, pp 285–320.
Biography
Michael BA Oldstone, MD, was presented the 2003 Pi-oneer in NeuroVirology Award at the 5th International

Tracking virus and immune response interactions with neuronsMBA Oldstone 215
Symposium on NeuroVirology, which was held September2–6, 2003 in Baltimore, Maryland, USA. The InternationalSociety for NeuroVirology presented this award of distinc-tion to Dr. Oldstone for his leadership, outstanding schol-arly accomplishments, and contributions to the field ofneurovirology.
Dr. Oldstone received his medical degree from the Uni-versity of Maryland School of Medicine, after which hecompleted his intern and residency training at Univer-sity Hospital in Baltimore, Maryland. As a postdoctoralfellow, he trained under Dr. Frank Nixon at the ScrippsClinic and Research Foundation in La Jolla, California,currently known as The Scripps Research Institute.
Dr. Oldstone is a Professor in the Departments of Neu-ropharmacology and Immunology at The Scripps ResearchInstitute and the Head of the Division of Virology. The Pio-neer in NeuroVirology Award represents an impressive listof honors bestowed upon Dr. Oldstone, among them, theJ. Allyn Taylor International Prize in Medicine, Rouse-Whipple Award for Research Excellence, Abraham FlexnerAward, and The Cotzias Award for Research Excellence inNervous System Disease. Additionally, he was elected tothe Institute of Medicine, the National Academy of Sci-ences, and the Association of American Physicians.
For more information and a list of past Pioneers, visitwww.isnv.org.


















