
FINAL TECHNICAL REPORT - gov.uk TECHNICAL REPORT ... tissues into water using a Baermann funnel, ......
30
1 CROP PROTECTION PROGRAMME Identification of resistance to major nematode pests of yams (Dioscorea spp.) in West Africa R6694 (ZA0021) FINAL TECHNICAL REPORT 1 APRIL 1996 – 31 MARCH 2000 RICHARD PLOWRIGHT & CHARLES KWOSEH CABI Bioscience Bakeham Lane Egham Surrey TW20 9TY. "This publication is an output from a research project funded by the United Kingdom Department for International Development for the benefit of developing countries. The views expressed are not necessarily those of DFID.” [R6694 Crop Protection Programme]
-
Upload
truongnguyet -
Category
Documents
-
view
217 -
download
1
Transcript of FINAL TECHNICAL REPORT - gov.uk TECHNICAL REPORT ... tissues into water using a Baermann funnel, ......

1
CROP PROTECTION PROGRAMME
Identification of resistance to major nematode pests of yams (Dioscorea
spp.) in West Africa
R6694 (ZA0021)
FINAL TECHNICAL REPORT
1 APRIL 1996 – 31 MARCH 2000
RICHARD PLOWRIGHT & CHARLES KWOSEH
CABI Bioscience
Bakeham Lane
Egham Surrey TW20 9TY.
"This publication is an output from a research project funded by the United Kingdom Department for International Development for the benefit of developing countries. The views expressed are not necessarily those of DFID.” [R6694 Crop Protection Programme]

2
EXECUTIVE SUMMARY
This DFID, CPP project was a collaboration between CABI Bioscience and the Tuber and
Root improvement programme of the International Institute of Tropical Agriculture in
Nigeria. The project sought to identify sources of resistance to the principal nematode
pests of yams (Scutellonema bradys and Meloidogyne incognita) which cause dry rot and
galling of yam tubers in West Africa and are a priority for yam breeders at IITA. The
project focused on development of yam nematode resistance screening techniques for field
and screenhouse and carried out extensive screening of germplasm from West Africa. An
important aim was to achieve a better understanding of the genetic and environmental
components of variability in the host: parasite interaction of yams and yam nematodes
through strategic research. The project involved survey work, which examined the
distribution of nematodes in Ghana and assessed farmer’s perceptions of nematode losses.
It also involved field, screenhouse and glasshouse trials in Nigeria, Ghana and UK.
Yam nematodes are widespread in Ghana, where they are well known to farmers and
where attempts to control nematodes using fallow and selecting good planting material are
commonplace. Farmers in Ghana consider losses from S. bradys to be more important
than those from M. incognita. Farmers grow a large number of varieties and whilst they
might be willing to test new, potentially resistant, varieties, it is important that such should
fit into this diverse background.
Experiments on precision established that Latin square field designs are more favourable
than an Alpha array, but neither dealt satisfactorily with the heterogeneity of natural
nematode populations and thus screening protocols which used artificial inoculum
(nematode infected yam peelings) were more reliable. Five or more replications are
recommended in order to approach meaningful separation of quantitative variants. Data
from experiments using young yam seedlings should be interpreted with care since this
work identified discrepancies between the results of such experiments and expression in
the field.
Most of the yam species and accessions screened have been found to be susceptible to
nematodes. There was variation in susceptibility within Dioscorea rotundata, but the
functional importance of this variation in terms of increased yield and reduced storage
losses requires more work. Dioscorea dumetorum has resistance to both S. bradys and M.
incognita, but barriers to interspecific crosses to D. rotundata preclude the exploitation of
this resistance at present.
Monoxenic culture techniques for the mass rearing of S. bradys have successfully been
developed, which can assist further detailed pot experimentation.
Pratylenchus coffeae which is widespread in Ghana on Musa spp. can multiply in roots of
a number of species of Dioscorea, namely D. alata, D. rotundata, D. cayenensis, D.
esculenta, D. dumetorum, D. bulbifera, but does not multiply readily in yam tubers and
hence is not a threat to yam production. P. coffeae in Ghana appears to be a Musa host
race.

3
BACKGROUND
Yams, Dioscorea spp. are important economic crops grown mainly in tropical countries
especially, in the yam belt or yam zone of West Africa. Many farm-families depend on the
tubers for food, cash, local food security and other traditional uses. The tubers provide a
substantial intake of vitamins (thiamine and vitamin C) and iron and also protein. Yam
peels and over matured tubers are used to feed domestic animals such as pigs, goats and
chickens. It is gradually being realised as an important non-traditional crop for export.
However, yams are severely damaged by plant parasitic nematodes reducing yield, food
quality, and market value. At least three potential nematode pest species namely
Scutellonema bradys, Meloidogyne spp. and Pratylenchus coffeae are known to exist.
Reduction of 20-30% in tuber weight at harvest has been reported. Also, nematode
infection contributes to long term storage losses, which have been estimated as 50%, and
loss can be total.
Chemicals, cultural methods such as hot water treatment of seed yams, crop rotation,
biological control, the use of resistant varieties and integrated management, can control
plant-parasitic nematodes. Most of the management options are limited in use due to high
costs, time, feasibility and adverse effects on the environment and mammalian toxicity.
Most plant-parasitic nematodes by their nature and habitat are restricted in their movement
from field to field and area to area and new physiological races that break resistance
probably would be slow. Yam cultivars with resistance to nematodes would provide an
attractive nematode management option appropriate to smallholder farm families. There is
therefore the need to begin to search for nematode resistance in yams.
PROJECT PURPOSE
The main project purpose was to identify sources of resistance to yam nematodes for use in
Yam Improvement Programmes aimed at reducing the yield and storage losses in yam
caused by nematodes. This addressed the DFID Crop Protection Programme’s
Forest/Agriculture Interface, Purpose 1, output of improved methods of managing the
major nematode pests of root and tuber crops. The project aimed to achieve a better
understanding of the genetic and environmental
components of variability in the host: parasite interaction of yams and yam nematodes
through strategic research. The project focused on development of yam nematode
resistance screening techniques and screening of yam germplasm for nematode resistance
then examined the correlation between nematode host status and other plant characteristics.
The findings should provide recommendations for plant breeders.

4
DETAILS OF COMPLETED RESEARCH ACTIVITIES
OUTPUT 1. TECHNIQUES FOR SCREENING YAMS FOR RESISTANCE TO
NEMATODES REFINED
MONOXENIC CULTURE OF THE YAM NEMATODE, S. BRADYS
Yam nematodes have not previously been cultured under monoxenic conditions, but with
careful maintenance, monoxenic cultures of migratory nematodes can provide a reliable
supply of inoculum of consistent quantity and quality as are required for screening. In this
work, the suitability of excised tuber tissues as a tissue substrate for S. bradys was
investigated. White yam tuber was washed and cut into pieces with skin intact and then
dipped in Bio- Supercarb for 15 min. The pieces were air dry and then cut into slices of
about 3-6 g each with skin intact. The skin of each slice was removed and then sterilised in
sodium hypochlorite (1 % available chlorine) for 15 min. The slices were rinsed six times
with sterilised distilled water and then dabbed dry on filter paper. The sterilised yam slices
were plated on 1 % water agar medium in Petri dishes. The plates had been put in a drying
cabinet at 30 C for 10 min to dry off excess water. The plates were kept in the culture room
at 25 C for about three weeks to callus before inoculated with S. bradys. S. bradys was
sterilised with 5 drops of 0.1 % malachite green (technical grade) for 5 min and then rinsed
10 times with sterilised distilled water. Each plate containing the callus yam slice was
inoculated with 20 or 30 active juvenile and adult stages of S. bradys. The plates were
sealed with insulating tape and kept in the culture room at 25 C.S. bradys were then
extracted from both the yam tissue and agar of the plates by a modified Baermann funnel
method after five months.
A means to suppress contaminating fungi in yam tissue callus was needed and Bio-
Supercarb, a systematic fungicide, which contains carbendazin, was used. The mortality of
the fungicide was tested on S. bradys. Three levels of concentration of the fungicide, 0.5,
1.1, and 2.2 and litres of water were used and water as the control. About 5 ml of the
various fungicide concentrations and water were separately dispensed into sterilised watch
glasses. Twenty active stages of S. bradys were put in each watch glass; there were three
replicates. S. bradys was obtained from yam bin cultures at International Institute of
Parasitology (IIP), UK. The nematodes were observed daily for three days and the effect of
treatment on them assessed by recording the number of live and dead nematodes. A
nematode was considered alive when active.
REFINEMENT OF A PROTOCOL FOR SCORING SYMPTOMS OF NEMATODE INJURY AND
QUANTIFYING NEMATODE DENSITIES
Experiments were done to examine the rate of emergence of S. bradys from chopped yam
tissues into water using a Baermann funnel, in order to establish optimum period of
incubation with regard to nematode recovery and the deterioration of yam tissue. A novel
method for scoring nematode symptoms on yam tubers was also devised and tested.
Although not critically evaluated experimentally, different methods were operated

5
concurrently in the same laboratory at IITA allowing full discussion of the practical
elements of each.
The novel method began with a random selection of tubers from each accession.
Symptoms of dry rot, galling as well as tissue softness and cracks were then recorded for
each tuber (0-3) based on the Table 1.1.
Table 1.1 Description of infection types and scores for yam nematodes
Infection Softened yam
Tuber tissue (%)
Root/tuber
galling
Tuber
Cracking
Root
Necrosis (%) score Dry/wet rot
0 0 None None None None
1 < 25 Light Light Light Light
2 26 – 50 Moderate Moderate Moderate Moderate
3 > 50 Severe Severe Severe Severe
Based on the scoring, the selected tubers of each accession were grouped according to
symptom types. A sub-sample of one to four tubers for each accession was selected from
the groupings. Each of the selected tubers was separately peeled (strip) from the proximal
to the distal end at three places of about equal distance from one another. The peelings of
each tuber were bulked, chopped to about 3-4 mm width and 2 cm length and thoroughly
hand-mixed. A 5 g sub-sample was taken for nematode extraction by a modified Baermann
funnel method. The nematodes were identified and numbers were counted. Sub-samples of
3 g in muslin bags were taken to assess the population density of Meloidogyne species. The
tissues in muslin bags were boiled in acid fuchsin stain for 15 min. and each muslin bag
was macerated for 20s at low speed. Stained Meloidogyne species in juvenile stages and
adults were counted.
COMPARISON OF FIELD EXPERIMENTAL DESIGNS
Two field experiments were set up at IITA research farm, Ibadan, to compare the
efficiency of Latin square arrangement and Alpha array experimental designs for screening
for yam nematode resistance. The experiments were established with alternating
allocations of naturally infested soil and naturally infested soil plus artificial infestation
with S. bradys infected yam tuber peelings. The naturally infested field had been cropped
to yam for three consecutive seasons.
STUDIES ON THE INVASION AND POPULATION DYNAMICS OF S.BRADYS IN YAM TUBERS.
A study of the population dynamics of yam nematode in tubers was done to examine the
role of both the rate of nematode infection and multiplication on observed differences in
susceptibility. This field trial studied the invasion and population dynamics of S. bradys in
resistant and susceptible yam tubers at IITA.
INVESTIGATION OF THE INOCULUM TYPE AND SIZE OF S. BRADYS
A range of nematode delivery systems are available for inoculating nematodes. This work
was done to optimise the delivery and quantity of inoculated nematodes in yam nematode

6
screening protocols. Two types and three sizes of inoculum were used in this study in the
screenhouse at IITA. The types of inoculum are nematode in water suspension and
chopped yam peelings. The inoculum sizes were 1 g, 25 g and 100 g of chopped S. bradys
infected yam peelings and 120, 2,800 and 10,000 S. bradys active stages representing the
number of nematodes in the peelings respectively. Two contrasting IITA D. alata
accessions, TDa 294 and TDa 92-2 were used. The plants were potted in about 800 ml
heat-sterilised 2:1 topsoil-cocopeat mix and then were inoculated after two weeks. A
simple line screening design with five replicates was used. The plants were assessed 9
weeks after inoculation by determining any visual nematode damage and reproduction in
roots and tubers of all entries in the screen.
INFLUENCE OF REPLICATION ON PRECISION IN ESTIMATING QUANTITATIVE DIFFERENCES
IN SUSCEPTIBILITY OF YAM TO NEMATODES
There are currently no good sources of resistance in Disoscorea rotundata to yam
nematode, although work, to date, suggests that quantitative variation might play a role.
This work was done to examine the level of replication that might be required to enable
selection of quantitative variants. A set of 21 potted tissue culture plants of two contrasting
D. rotundata accessions, TDr 87/00571 and TDr 87/00211 were used in this trial in the
screenhouse at IITA. The plants were potted in steam sterilised soil-cocopeat mix. The
established plants were inoculated with about 1,700 active stages of S. bradys (35 g
infected yam peelings) per pot. The simple line design was used. Entries were assessed
eight weeks after inoculation by recording any visual nematode injury and the number of S.
bradys in roots or tubers. The plants were randomly harvested as groups of 3, 5 and 13
representing the number of replicates for each entry.
OUTPUT 2 YAM GERMPLASM SCREENED FOR RESISTANCE TO
NEMATODES
PRELIMINARY SCREENING FOR NEMATODE RESISTANCE OF IITA YAM GERMPLASM
At the outset of this research project an assessment of nematode resistance in stored yam
tubers at IITA was made. This was a collection of germplasm which had not been exposed
experimentally to nematode infection. For this reason any potentially resistant tubers
identified, had to be treated with caution. A total of 306 D. rotundata and 18 D. alata
accessions were screened at International Institute of Tropical Agriculture (IITA), Ibadan,
Nigeria. The tubers were harvested in 1996 from the research farm at IITA headquarters,
Ibadan, Nigeria. The plots were planted with cassava or yam in 1994 and mucuna fallow in
1995. The harvested accessions were either stored in a storeroom maintained at 19 C or
kept in an open-air yam barn under ambient conditions for about three months. Each
accession was randomly selected and symptoms of dry rot, galling and tissue softness as
well as cracking of each tuber were scored (0-3). Based on the scoring, the selected tubers
of each accession were grouped according to symptom types. A sub-sample of one to four
tubers for each accession was selected from the groupings. Each of the selected tubers was
separately peeled (strip) from the proximal to the distal end at three places of about equal
distance from one another. The peelings of each tuber were bulked, chopped to about 3-4
mm width and 2 cm length and thoroughly mixed. A 5g sub-sample was taken for
nematode extraction by a modified Baermann funnel method. The nematodes were

7
identified and numbers were counted. Sub-samples of 3 g in muslin bags were taken to
assess the population density of Meloidogyne species. The tissues in muslin bags were
boiled in acid fuchsin stain for 15 min. and each muslin bag was macerated for 20 s at low
speed. Stained Meloidogyne species in juvenile stages and adults were counted.
SCREENHOUSE SCREENING OF SELECTED GENOTYPES OF IITA YAM ACCESSIONS,
NIGERIA.
Based on the preliminary assessment of nematode infestation in yams of the IITA yam
breeding programme, 23 genotypes of D. rotundata and 17 wild and domesticated
Dioscorea species were selected for screening in pots. The wild and some domesticated
yams were collected from Cameroon, Benin Republic, Republic of Guinea, Cote d'Ivoire,
and Burkina Faso all in West Africa by the Biotechnology Research Unit of IITA. Uniform
plants from yam minisetts pre-sprouted in sterilised moist coco-peat (shredded coconut
husk) were used for screening for resistance to S. bradys and Meloidogyne spp. A
randomised complete block design with eight replicates was used. Established plants in the
pots were inoculated with about 360 active stages of S. bradys per pot and 500 juveniles of
Meloidogyne species per pot for each experiment. All entries were assessed five months
after inoculation by recording visual nematode damage and the number of nematodes in
roots or tubers.
FIELD SCREENING OF GHANAIAN LANDRACES FOR RESISTANCE TO S. BRADYS AND
MELOIDOGYNE SPECIES
Forty Ghanaian yam landraces identified by farmers during the Farmer participatory
appraisal (See output 3) were screened for resistance at Crops Research Institute in
Kumasi.
GLASSHOUSE INVESTIGATION OF THE VARIATION IN SUSCEPTIBILITY OF YAM SPECIES TO
S. BRADYS, UK.
This experiment was set up at IIP, UK in the glasshouse. The yam plants were obtained
from yam vines except D. dumetorum that was from seeds. They were screened in vacapots
containing about 200 ml sterilised soil-sand mix. The established plants were inoculated
with about 500 active stages of S. bradys. All the plants in the screen were assessed 9
weeks after inoculation by recording any apparent nematode damage and reproduction of
the nematodes in the roots or tubers.
FIELD SCREENING OF OPEN-POLLINATED YAM SEEDLING POPULATION FOR S. BRADYS
RESISTANCE.
The open-pollinated yam seedlings were obtained from accession TDr 95/05576 (female)
and suspected male, D. rotundata var. Puna. The seeds were sown in moistened peat
pellets in the glasshouse at IITA. The seedlings were transplanted to the field at 2 to 3 leaf
stage (5–6 weeks old) in ridges at 1 m x 0.25 m and then a shed was raised over them. 16-
16-16 NPK fertiliser was applied using the ring method. The seedlings were staked
immediately they started to crawl. Six weeks after transplanting, the seedlings were

8
inoculated with about 1,700 S. bradys (25 g yam peelings). All the plants in the screen
were assessed seven months after inoculation by recording any apparent nematode damage
and reproduction of the nematodes in tubers. Each seedling was considered genotypically
different and there was no replication.
SCREENHOUSE SCREENING OF YAM PARENTAL LINES FOR RESISTANCE TO S. BRADYS
Yam parental lines of the Cellular and Molecular Technologies Laboratories, IITA were
tested in a pot experiment in the screenhouse for their reaction to S. bradys. Tissue culture
plants of ten parental lines (five each of D. alata and D. rotundata) were screened. The
tissue culture plants were potted in steam sterilised soil-cocopeat mix. The established
plants were inoculated with about1,700 active stages of S. bradys (35 g chopped S. bradys
infected yam peelings) per pot.
SCREENHOUSE SCREENING OF PARENTS AND PROGENIES OF INTRASPECIFIC
HYBRIDISATIONS FOR RESISTANCE TO NEMATODES
Intraspecific hybrids of TDr 87/00211 (male) and TDr 87/00571 (female) were screened in
pot experiments in the screenhouse for their reaction to S. bradys and Meloidogyne
incognita at IITA, Nigeria. Fifty-nine genotypes of intraspecific hybrids (Tissue culture
plants) and their parents were screened for their reaction to S. bradys and twenty-five for
Meloidogyne incognita. The tissue culture plants were potted in sterilised soil-cocopeat
mix. The established plants were inoculated with about 780 active stages of S. bradys (25 g
S. bradys infected yam peelings) per pot. For the Meloidogyne species trial, each pot was
inoculated with 400 juveniles of Meloidogyne species. The plants were assessed eight
weeks after inoculation by recording visual nematode injury and the number of nematodes
in roots or tubers.
OUTPUT 3 NEMATODE VARIABILITY ON YAM KNOWN
PARTICIPATORY RURAL APPRAISAL: NEMATODE PESTS PROBLEMS OF YAM IN GHANA
A farmer participatory appraisal of pests and diseases in stored yams was made in Ghana
in 1998 to examine farmers’ perceptions of nematode disease problems on yams. A total of
17 districts and farmers from 32 towns and villages were considered. The surveyed area is
within the Forest-transitional and Guinea-Savannah agroecological zones. The farmers
provided information on the type of varieties they grow, cropping system, planting
material, storage methods, and pests’ problems of yams and control. A total of 243
respondents (farmers) contributed. Dry rot and galling of tubers were identified. Nematode
infected yam tubers from all the sites visited were collected. Local and traditional yam
varieties were also collected for controlled nematode resistance screening.
IDENTIFICATION AND DISTRIBUTION OF YAM NEMATODES IN GHANA
Nematode populations in yam tissues collected during the rural appraisal were extracted
and identified to species level.

9
HOST RACE STATUS OF PRATYLENCHUS COFFEAE ON YAM IN GHANA
Pratylenchus coffeae is widespread in Ghana, where it is a pest of Musa, but it was not
found infecting yam although, it is known to be a pest of yam in Pacific rim countries and
central America. This experiment was done to examine race specific variation in P. coffeae
in Ghana.
This trial was set up in the screenhouse at UST, Kumasi. Plants from yam minisetts and
tissue culture plants of Musa species were potted in sterilised soil-cocopeat mix. The
established plants were inoculated with about 800 active stages of P. coffeae (45 g infected
roots and corms of Musa). The plants were assessed eight weeks after inoculation by
recording visual nematode injury and the number of nematodes in roots or tubers.
Concurrently glasshouse experiments were done in the UK comparing the multiplication of
populations of P. coffeae from Belize and Ghana on Musa cv Grande Naine, Dioscorea
rotundata and D. togoensis.
RESEARCH RESULTS
OUTPUT 1. TECHNIQUES FOR SCREENING YAMS FOR RESISTANCE TO
NEMATODES REFINED
MONOXENIC CULTURE OF THE YAM NEMATODE, S. BRADYS
S. bradys populations in the inoculated yam tuber slices after treatment increased by ca
100 to 247 and 340 to 820 fold for 20 or 30 S. bradys after five months respectively (Table
1.2). Mean reproduction increased by 196 fold for 20 S. bradys and 600 fold for 30 S.
bradys inoculated (Table 1.2). The outer surfaces of the yam slices were darkened. This
may be due to necrosis of invaded tissues, whereas the inner parts had fresh tissues. All the
culture plates were free from contaminating microorganisms. Dry rot symptoms
characteristic of S. bradys infection in yam tissues were not observed. This is probably
because of the humid condition in the Petri plates. Also, nematodes are known to behave
differently in callus plant tissues. In previous work at IITA culturing of Scutellonema
bradys in the greenhouse was successful on sweet potato and cowpea, but not on sprouting
yam tuber pieces. However, this technique using sterilised yam slices on water agar
medium in Petri dishes can support high reproduction of S. bradys and constitutes a
suitable method for mass-production of the nematodes for screening for host plant
resistance. It is impossible to track the fate of inoculated nematodes, but it is always the
case that a proportion of nematodes are not potent. These results suggest that raising the
inoculum level from 20–30 dramatically improved the multiplication of nematodes in the
time frame of the experiment.

10
Table 1.2. S. bradys reproduction in yam tuber slices five months after inoculation
Plate no.
Weight of yam
Slices/ g
Initial S. bradys
Population (Pi)
Mean no.
S. bradys (Pf)
Pf/Pi
1 3.0 20.0 2040.0 102.0
2 5.0 20.0 5230.0 261.5
3 3.0 20.0 3454.0 172.7
4 4.0 20.0 4930.0 246.5
5 6.0 30.0 24600.0 820.0
6 6.0 30.0 22800.0 760.0
7 3.0 30.0 10250.0 341.7
8 4.0 30.0 14400.0 480.0
Table 1.3. Effect of Bio-Supercarb concentration on S. bradys with initial inoculation
of 20 nematodes
Dosage
*Mean no. S.
bradys alive
Mean no. S.
bradys dead
% Live
nematodes
% Dead
Nematodes
Recommended (1.1) 19.0 1.0 95.0 5.0
Low (0.5) 20.0 0.0 100.0 0.0
High (2.2) 20.0 0.0 100.0 0.0
Tap water (control) 20.0 0.0 100.0 0.0
*Mean of three replicates
No death was observed in the low, high and control treatments after three days however,
one death was observed in the recommended dosage (Table 1.3). Analysis of variance
showed that there were no significant differences between treatments. It is very difficult to
axenise yam tissues for monoxenic culture, but Bio-Supercarb can be used to treat yam
slices for the suppression of fungi growth during the monoxenic culture of S. bradys.
REFINEMENT OF A PROTOCOL FOR SCORING SYMPTOMS OF NEMATODE INJURY AND
QUANTIFYING NEMATODE DENSITIES
The 0-3 rating of dry rot symptoms (Table 1.1) worked well as a means of assessment and
it was not thought that the creation of further divisions of symptom severity would lead to
increased accuracy. Peeling yam tissue from the proximal to the distal end reduces bias in
selecting areas of tissue for nematode population estimates, but smaller samples of peel
taken systematically over the tuber would probably work just as well.
Results from Baermann funnel extracts in water showed that most nematodes escaped from
macerated yam peelings, within two days of setting up the incubation at ambient

11
temperatures. Incubated tissues rapidly deteriorated after 2 days, but the extra work
required for daily removal and replenishment of extracts, which does increase their
longevity, did not recover significantly more nematodes and did not justify the extra work
involved. This method would in any case be impossible within very large screening trials.
COMPARISON OF FIELD EXPERIMENTAL DESIGNS
The dry rot scores and nematode counts for the tested accessions are given in Tables 1.4
&1.5. The consistency and reliability of field screening was greatly improved by artificial
inoculation of nematodes and such methods, i.e. which do not rely on natural infection, are
recommended. Neither design eliminated the occurrence of false negative (resistant)
reactions as a result of ’escapes’ from natural nematode infection. Such escapes can occur
because of the heterogeneous distribution of nematodes in the soil. Generally, the Latin
square arrangement recorded more consistent dry rot rating and high nematode counts than
the Alpha array and gave small (4%) improvements in efficiency over the Alpha array.
Latin square which also benefits from being relatively simple to use can be recommended.
Table 1.4. Effect of a Latin square design on the reaction of ten IITA yam parental lines to S.
bradys eight weeks after storage (Inoculated with ca 2,500 S. bradys [50 g infected yam
peelings])
*Mean dry rot index Mean no. S. bradys/ 5 g tuber peelings
Naturally dArtificially Naturally infested soil Artificially infested Soil
Yam Infested Infested kTrans-
formed s.e
Trans-
Formed s.e Accession
Soil Soil No. No.
TDr 131a 0.3 3.0 80.0 5.7 6.8 1481.4 37.9 3.5
TDr 89/01438b 3.0 3.0 845.7 23.7 8.0 800.6 26.4 3.5
TDr 93-28a 1.6 3.0 367.0 16.1 6.0 716.2 25.1 3.5
TDr 93-72a 2.6 3.0 741.6 25.2 6.0 395.6 19.1 3.5
TDr 89/02677b 2.6 3.0 1101.6 31.2 6.0 849.0 27.0 3.5
TDr 93-82a 2.5 3.0 521.3 19.7 6.8 447.5 20.6 3.9
TDr 93-1a 1.8 3.0 724.0 23.5 6.0 146.0 11.8 3.5
TDr 93-31a 2.5 3.0 641.3 22.6 6.8 147.7 11.9 4.6
TDr 93-83a 1.0 3.0 431.3 17.5 6.8 86.3 8.0 3.5
TDr 93-2a 2.2 3.0 954.0 26.4 6.0 435.0 19.5 3.5
CV 50.9 0.0 62.7 36.3 cRoot MSE 1.0 0.0 13.5 7.8
*Average of 5 replicates, s.e: Standard error. TDr: Dioscorea rotundata, alandrace,
bBreeder’s line
kSquare root (Mean + 0.5) and SAS adjusted for missing data.
CRoot mean square error.
dNaturally
infested soil plus chopped S. bradys infested yam peelings.

12
Table 1.5. Effect of an Alpha array design on the reaction of ten IITA yam parental
lines to S. bradys eight weeks after storage (Inoculated with ca 2,500 S. bradys [50 g
infected yam peelings]) Mean no. S. bradys/ 5 g tuber peelings
*Mean dry rot index Naturally infested soil Artificially infested soil
Yam Naturally dArtificially
kTrans-
formed s.e
Trans-
formed s.e Accession infested soil infested soil No. No.
TDr 131a 1.6 3.0 375.8 16.7 5.6 1198.3 26.6 7.9
TDr89/01438b 1.9 3.0 460.4 16.9 4.7 470.0 20.1 7.9
TDr 93-28a 1.3 3.0 288.3 15.4 7.5 898.7 25.4 5.0
TDr 93-72a 2.1 2.3 759.3 23.9 4.7 178.3 11.8 8.0
TDr 89/ 02677b 2.3 2.7 739.3 26.6 5.1 329.0 18.0 8.0
TDr 93-82a 1.5 3.0 590.0 17.6 9.3 570.6 22.1 4.6
TDr 93-1a 2.3 2.3 137.3 10.3 7.6 164.7 10.5 5.4
TDr 93-31a 1.3 3.0 251.7 11.7 7.5 265.0 14.8 5.5
TDr 93-83a 0.9 2.3 147.4 8.8 4.4 393.3 19.0 8.0
TDr 93-2a 0.0 2.3 20.0 3.1 7.6 318.5 16.4 6.8
CV 70.6 29.6 72.9 67.5 cRoot MSE 1.1 0.8 11.8 12.6
*Average of 10 replicates, s.e: Standard error. TDr: Dioscorea rotundata, aLandrace,
bBreeder’s
line. k
Square root (Mean + 0.5) and SAS adjusted for missing data. CRoot mean square error.
dNaturally infested soil plus chopped S. bradys infested yam peelings.
STUDIES ON THE INVASION AND POPULATION DYNAMICS OF S.BRADYS IN YAM TUBERS.
Nematode population at harvest and subsequent dry rot symptoms ranged from 0 to 2.8 and
0 to 560 respectively. At 8 weeks after harvest, the range was 0 to 3 for dry rot and 0 to
800 for number of nematodes. Generally, symptoms and nematode population density
increased with time in the susceptible yam accessions, except in TK106 which had little or
no living tissue left for invasion. There was no change in dry rot symptom expression and
invasion in the resistant yam accession, D. dumetorum (Table 1.6). In susceptible yam
tubers the S. bradys had invaded at harvest and continued to reproduce in storage. This
work cannot detail the mechanism of resistance in D. dumetorum, but it is likely that post-
infectional characteristics in the yam tuber, confer resistance. It is less likely that
nematodes are completely prevented from entering the D. dumetorum tuber.
Table 1.6. Invasion and population dynamics of S. bradys in yam tubers
(Means of 6 replicates and inoculated with ca 2,500 S. bradys [50 g yam peelings])
Yam accession
(all D. rotundata
except otherwise stated)
Dry rot score
Mean no. S. bradys/5 g tuber peelings
Harvest After 8 wk Harvest After 8 wk
TDr 89/01438 1.7 1.8 210.0 490.0
CR1/0051 2.8 3.0 180.0 800.0
D. dumetorum 0.0 0.0 0.0 0.0
TDr 131 2.2 2.4 60.0 680.0
TK 106 2.0 2.7 560.0 60.0

13
INVESTIGATION OF THE INOCULUM TYPE AND SIZE OF S. BRADYS
In most cases, the S. bradys infected yam peelings recorded higher nematode numbers than
with the nematode in water suspension (Table 1.7). There was no significant difference
between the inoculum size in peelings however, nematode numbers increased from the
lowest weight of 1g to the highest of 100 g peelings. Nematode suspensions did not give
consistent reproduction of S. bradys in both yam accessions. The coefficient of variation
(CV) was high for suspension, for example, 68.8 and 80.2 for number of S. bradys in roots
and tubers as against 46.4 to 67.8 for peelings respectively. Based on the results, S. bradys
infected peelings are a better source of inoculum, if available, for screening for nematode
resistance in yam.
Table 1.7. Efficiency of inoculum type and size of S. bradys in resistance screening Mean tuber
Cracking
Index
Mean no. S. bradys/ 5 g tissue
Yam
accession
Inoculum Tuber
peelings
Type Size Roots Range Range
TDa 294 Peels 1.0 g 0.5 (0.6) 306.0 65.0-675.0 213.0 0.0-820.0
TDa 294 Suspension 120.0 1.0 (0.0) 9.0 0.0-35.0 0.0 0.0-0.0
TDa 294 Peels 25.0 g 0.0 (0.0) 429.0 150.0-707.0 85.0 60.0-100.0
TDa 294 Suspension 2800.0 0.3 (0.6) 119.0 15.0-220.0 11.0 0.0-30.0
TDa 294 Peels 100.0 g 0.7 (0.6) 461.0 287.0-635.0 368.0 55.0-680.0
TDa 294 Suspension 10000.0 1.0 (0.0) 132.0 0.0-215.0 34.0 0.0-90.0
TDa 92-2 Peels 1.0 g 1.0 (0.0) 136.0 15.0-310.0 95.0 30.0-245.0
TDa 92-2 Suspension 120.0 1.5 (0.6) 3.0 0.0-10.0 16.0 10.0-25.0
TDa 92-2 Peels 25.0 g 0.1 (0.4) 421.0 116.0-1160.0 430.0 80.0-1413.0
TDa 92-2 Suspension 2800.0 1.0 (0.0) 30.0 0.0-105.0 32.0 0.0-85.0
TDa 92-2 Peels 100.0 g 0.3 (0.5) 437.0 0.0-1080.0 518.0 20.0-890.0
TDa 92-2 Suspension 10000.0 0.7 (0.6) 55.0 0.0-155.0 71.0 0.0-190.0
Standard deviation in parentheses. Mean of 5 replicates.
INFLUENCE OF REPLICATION ON PRECISION IN ESTIMATING QUANTITATIVE DIFFERENCES
IN SUSCEPTIBILITY OF YAM TO NEMATODES
The number of S. bradys/5g tissue ranged from 135 to 929 for roots and 1 to 55 for tubers
for TDr 87/00571 and 265 to 1889 for roots and 4 to 40 for tubers for TDr 87/00211 (Table
1.8), with significant differences (P=0.05) between the replications. Increasing replication
improves the precision of quantitative differences in susceptibility. In practice, practical
considerations will govern the level of replication, but the results suggest that greater than
5 replicates give better separation of quantitative differences in susceptibility in yam to S.
bradys.

14
Table 1.8. Influence of replication on quantitative difference in susceptibility of yam to S.
bradys (Inoculated with 1700 S. bradys)
Mean tuber
cracking
index
Mean no. S. bradys/ 5 g tissue
Replicate
number
Yam
accession
Tuber
peelings
s.e Roots s.e s.e
3 TDr 87/00211 1.0 (0.0) 0.3 135.0 (82.9) 6.8 2.0 (4.0) 27.5
5 TDr 87/00211 1.4 (0.6) 0.2 343.0 (164.9) 5.2 1.0 (0.6) 21.3
13 TDr 87/00211 0.5 (0.5) 0.1 929.0 (642.3) 3.3 55.0 (58.4) 13.2
3 TDr 87/00571 1.0 (0.0) 0.4 265.0 (243.0) 12.2 4.0 (4.9) 1.9
5 TDr 87/00571 2.3 (1.2) 0.3 956.0 (484.5) 9.4 27.0 (23.1) 1.5
13 TDr 87/00571 2.0 (0.8) 0.2 1889.0 (1715.3) 7.0 40.0 (38.6) 0.9
Standard deviation in parenthesis. s.e : standard error

15
OUTPUT 2 YAM GERMPLASM SCREENED FOR RESISTANCE TO
NEMATODES
PRELIMINARY SCREENING FOR NEMATODE RESISTANCE OF IITA YAM
The highest number of nematodes recorded was 4,500/5 g for S. bradys and 250/5 g for
Meloidogyne species. S. bradys was the most prevalent nematode occurring in 46% of
accessions. 10.2 % and 0.3 % of the accessions were infested with Meloidogyne and
Pratylenchus species respectively. No linear relationship was observed between symptom
ratings and nematode population densities in tubers for either nematode. Based on the
number of nematodes per unit tissue selection criteria, the following number of accessions
were placed in each reaction category: 0-10 = Resistant (R), 11-25 = Partially resistant
(PR), 26–50 = Moderately susceptible (MS), > 50 = Susceptible (S) and Sensitive = severe
symptom and low nematode density.
Table 2.1. IITA yam accessions grouped under reaction categories
No. of accessions for nematodes reactions
S. bradys Meloidogyne species Pratylenchus
Reaction
category D. rotundata D. alata D. rotundata D. alata Yams
R 160.0 13.0 202.0 18.0 0.0
PR 43.0 2.0 43.0 0.0 0.0
MS 48.0 0.0 24.0 0.0 2.0
S 52.0 3.0 9.0 0.0 6.0
Sensitive 3.0 0.0 28.0 0.0 0.0
SCREENING OF SELECTED GENOTYPES OF IITA YAM ACCESSIONS FOR RESISTANCE TO S.
BRADYS AND MELOIDOGYNE INCOGNITA, NIGERIA.
In field screening all genotypes selected were susceptible to S. bradys, but there were
significant differences (P<0.01) between accessions. There was a strong linear relationship
between dry rot symptoms and S. bradys population. The number of S. bradys/5 g ranged
from 0 to 1,825. In most cases, higher dry rot scores were associated with higher the
numbers of S. bradys (Table 2.2).

16
Table 2.2. Selected IITA yam accessions and their reaction to S. bradys
(Mean of unequal number of replicates (3-12) and inoculated with about 800 S. bradys
(50g infected yam peels)
Yam accession
Dry rot index (0-3)
Harvest After 8 weeks No. S. bradys/5 g after 8 weeks
TDa 92-2 1.0 2.9 1825.0
TDa 95-92 1.6 3.0 1167.0
TDr 93-91 0.8 2.2 1086.0
TDa 96-12 1.3 2.8 1048.0
TDa 297 1.0 2.8 840.0
TDr 93-22 2.0 2.3 529.0
IN94R-23 0.5 2.2 488.0
TDr 131 1.0 2.6 427.0
TDa 93-36 1.8 2.9 408.0
TDa 96-5 1.3 2.5 393.0
TDr 93-49 1.2 2.3 327.0
TDr 93-82 0.7 2.7 285.0
IN94R-5 0.5 2 0 260.0
TDr 93-74 1.2 2.2 211.0
TDa 95-27 2.0 2.5 209.0
TDr 93-9 3.0 3.0 160.0
TDr 93-72 2.0 3.0 128.0
TDr 93-1 1.0 3.0 105.0
TDr 93-39 0.0 0.0 0.0
In general, D. alata (TDa) had higher population densities of S. bradys than D. rotundata
(TDr) ranging from 209 to 1825 and 0 to 1086 respectively. All IITA D. alata and D.
rotundata accessions screened are susceptible to S. bradys.

17
Table 2.3. Reaction of IITA yam accessions to Meloidogyne incognita in pot trials
(Means of 8 replicates and inoculated with about 400 Meloidogyne juveniles [J2])
Yam accession (all D. rotundata
except otherwise specified)
Tuber galling
(0-3)
Root galling
(0-3)
Meloidogyne J2/5 g tissue
Roots Tuber
1010 1.0 0.4 22.0 23.0
D. mangenotiana (Guinea forest) 0.1 0.6 268.0 13.0
TDr 93-82 0.5 0.9 5.0 6.0
TDr 179 0.4 0.3 4.0 4.0
TDr 89/02565 0.4 0.4 12.0 3. 0
KK102 1.0 0.8 15.0 3.0
TDr 131 0.4 0.4 3.0 2.0
TDr 89/02677 0.8 0.3 47.0 2.0
TDr 93-25 0.5 0.6 16.0 1.0
2008 0.4 0.6 13.0 0.0
1024 0.3 0.1 3.0 0.0
1020 0.5 0.8 2.0 0.0
1009Tm101 0.1 0.3 0.0 0.0
BP 129 0.1 0.3 3.0 0.0
Ka 120 0.1 1.0 8.0 0.0
Yb 114 0.1 1.0 0.0 0.0
KP 106 0.0 0.0 9.0 0.0
NF 104 0.3 0.6 3.0 0.0
TDr 93-1 0.1 0.9 2.0 0.0
TDr 93-28 0.3 0.5 3.0 0.0
TDr 93-31 0.0 0.3 0.0 0.0
TDr 93-47 0.0 0.3 0.0 0.0
TDr 93-72 0.4 0.3 3.0 0.0
TDr 93-74 0.4 0.4 0.0 0.0
TDr 95-78 0.1 0.5 5.0 0.0
TDr 93-79 0.3 0.4 8.0 0.0
TDr 95-83 0.3 0.6 53.0 0.0
TDr 93-89 0.1 0.0 2.0 0.0
TDr 87/00158 0.1 0.9 0.0 0.0
TDr 89/01438 0.0 0.1 0.0 0.0
TDr 89/02461 0.8 0.1 0.0 0.0
TDr 89/02494 0.4 0.8 10.0 0.0
IN94R-5 0.0 0.4 0.0 0.0
TDr 95-158 0.3 0.8 0.0 0.0
IN94R-102 0.0 0.4 1.0 0.0
D. praehensilis (Moyenne in Guinea) 0.1 0.8 0.0 0.0
BP128 0.3 0.6 7.0 0.0
Baniakpa 0.0 0.1 0.0 0.0
D. praehensilis (Guinea forest) 0.0 0.3 0.0 0.0
D. Praehensilis (Benin Republic) 0.3 0.4 0.0 0.0
Standard deviation 0.6 0.5 0.9 0.1
Analysis of variance showed highly significant differences (P = 0.01) between yam
accesssions. The mean number of Meloidogyne incognita ranged from 0 to 268/ 5g roots
and 0 to 23/5 g tuber (Table 2.3). There was no apparent correlation between galling in
roots and tubers and numbers of nematodes in root and tubers. There was no linear
relationship between susceptibility in roots and tubers. These are results are difficult to
interpret especially regarding classification because of the poor reaction of the yams to M.

18
incognita. This is probably because of the high temperatures recorded in the glasshouse
resulting in killing of the nematodes or making them impotent.
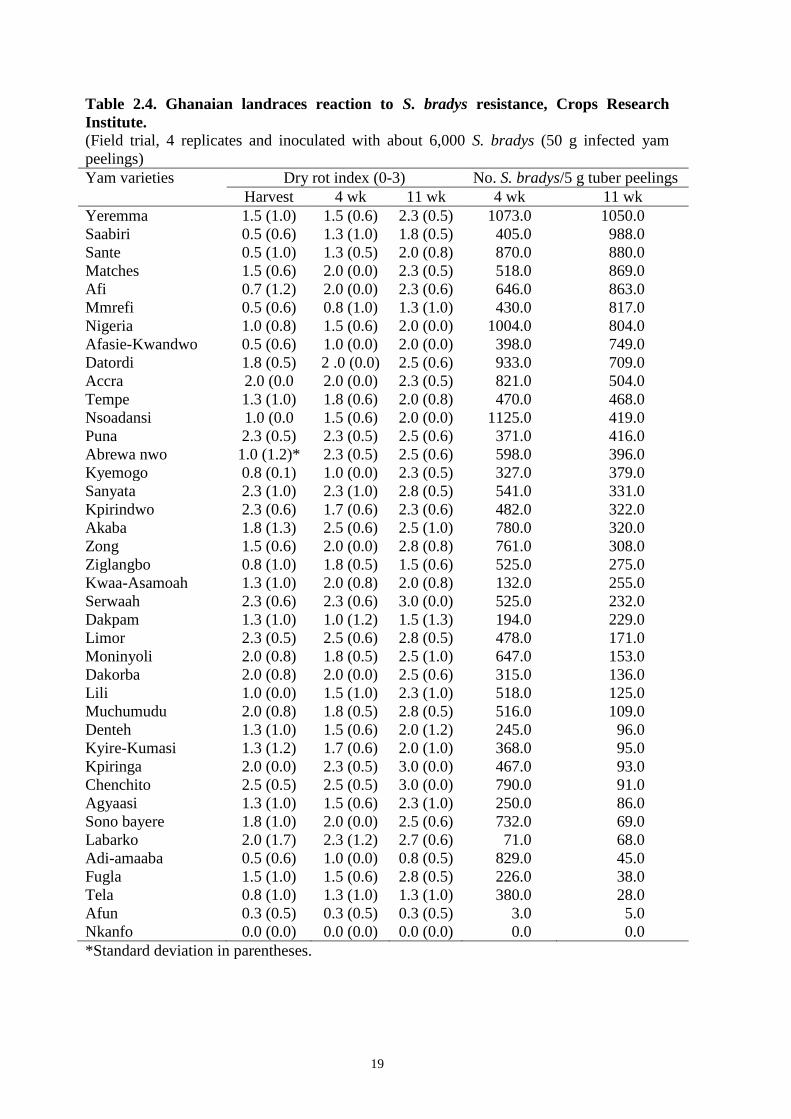
SCREENING OF GHANAIAN LANDRACES FOR RESISTANCE TO S. BRADYS AND
MELOIDOGYNE SPECIES
Based on the farmer participatory appraisal, 40 Ghanaian yam landraces were screened.
There were highly significant differences (P = 0.01) between yam varieties. There was
weight reduction in the stored yam tubers, and dry rot scores also increased within the
storage period. There was a linear relationship between the weight reduction in tubers
infected with S. bradys and the increased dry rot symptoms. It is noteworthy that the
severity of both dry rot and root knot nematode galls increased during storage. This
implies that both species continue to multiply in storage. Gall scores did not correlate with
tuber weight loss and confirm farmer perceptions that root knot nematode is a less
important nematode in terms of crop loss. Dry rot and S. bradys population density
revealed a strong negative correlation (r =0.42, P = 0.01). S. bradys counts at four weeks
ranged from 0 to 951/5 g and from 0 to 675/5 g at 11 weeks after harvest (Table 2.4).
Again, severely infected yam tubers recorded high nematode populations. However, some
severely infected tubers had low S. bradys population probably because these tubers were
either dried out or completely destroyed by dry and wet rot with very little or no living
tissue remaining. Ten contrasting varieties were selected for further studies and results
confirmed that D. dumentorum var. nkanfo and D. cayenensis var. Afun are resistant to S.
bradys. D. dumentorum var. nkanfo is also resistant to Meloidogyne incognita.
19
Table 2.4. Ghanaian landraces reaction to S. bradys resistance, Crops Research
Institute.
(Field trial, 4 replicates and inoculated with about 6,000 S. bradys (50 g infected yam
peelings)
Yam varieties Dry rot index (0-3) No. S. bradys/5 g tuber peelings
Harvest 4 wk 11 wk 4 wk 11 wk
Yeremma 1.5 (1.0) 1.5 (0.6) 2.3 (0.5) 1073.0 1050.0
Saabiri 0.5 (0.6) 1.3 (1.0) 1.8 (0.5) 405.0 988.0
Sante 0.5 (1.0) 1.3 (0.5) 2.0 (0.8) 870.0 880.0
Matches 1.5 (0.6) 2.0 (0.0) 2.3 (0.5) 518.0 869.0
Afi 0.7 (1.2) 2.0 (0.0) 2.3 (0.6) 646.0 863.0
Mmrefi 0.5 (0.6) 0.8 (1.0) 1.3 (1.0) 430.0 817.0
Nigeria 1.0 (0.8) 1.5 (0.6) 2.0 (0.0) 1004.0 804.0
Afasie-Kwandwo 0.5 (0.6) 1.0 (0.0) 2.0 (0.0) 398.0 749.0
Datordi 1.8 (0.5) 2 .0 (0.0) 2.5 (0.6) 933.0 709.0
Accra 2.0 (0.0 2.0 (0.0) 2.3 (0.5) 821.0 504.0
Tempe 1.3 (1.0) 1.8 (0.6) 2.0 (0.8) 470.0 468.0
Nsoadansi 1.0 (0.0 1.5 (0.6) 2.0 (0.0) 1125.0 419.0
Puna 2.3 (0.5) 2.3 (0.5) 2.5 (0.6) 371.0 416.0
Abrewa nwo 1.0 (1.2)* 2.3 (0.5) 2.5 (0.6) 598.0 396.0
Kyemogo 0.8 (0.1) 1.0 (0.0) 2.3 (0.5) 327.0 379.0
Sanyata 2.3 (1.0) 2.3 (1.0) 2.8 (0.5) 541.0 331.0
Kpirindwo 2.3 (0.6) 1.7 (0.6) 2.3 (0.6) 482.0 322.0
Akaba 1.8 (1.3) 2.5 (0.6) 2.5 (1.0) 780.0 320.0
Zong 1.5 (0.6) 2.0 (0.0) 2.8 (0.8) 761.0 308.0
Ziglangbo 0.8 (1.0) 1.8 (0.5) 1.5 (0.6) 525.0 275.0
Kwaa-Asamoah 1.3 (1.0) 2.0 (0.8) 2.0 (0.8) 132.0 255.0
Serwaah 2.3 (0.6) 2.3 (0.6) 3.0 (0.0) 525.0 232.0
Dakpam 1.3 (1.0) 1.0 (1.2) 1.5 (1.3) 194.0 229.0
Limor 2.3 (0.5) 2.5 (0.6) 2.8 (0.5) 478.0 171.0
Moninyoli 2.0 (0.8) 1.8 (0.5) 2.5 (1.0) 647.0 153.0
Dakorba 2.0 (0.8) 2.0 (0.0) 2.5 (0.6) 315.0 136.0
Lili 1.0 (0.0) 1.5 (1.0) 2.3 (1.0) 518.0 125.0
Muchumudu 2.0 (0.8) 1.8 (0.5) 2.8 (0.5) 516.0 109.0
Denteh 1.3 (1.0) 1.5 (0.6) 2.0 (1.2) 245.0 96.0
Kyire-Kumasi 1.3 (1.2) 1.7 (0.6) 2.0 (1.0) 368.0 95.0
Kpiringa 2.0 (0.0) 2.3 (0.5) 3.0 (0.0) 467.0 93.0
Chenchito 2.5 (0.5) 2.5 (0.5) 3.0 (0.0) 790.0 91.0
Agyaasi 1.3 (1.0) 1.5 (0.6) 2.3 (1.0) 250.0 86.0
Sono bayere 1.8 (1.0) 2.0 (0.0) 2.5 (0.6) 732.0 69.0
Labarko 2.0 (1.7) 2.3 (1.2) 2.7 (0.6) 71.0 68.0
Adi-amaaba 0.5 (0.6) 1.0 (0.0) 0.8 (0.5) 829.0 45.0
Fugla 1.5 (1.0) 1.5 (0.6) 2.8 (0.5) 226.0 38.0
Tela 0.8 (1.0) 1.3 (1.0) 1.3 (1.0) 380.0 28.0
Afun 0.3 (0.5) 0.3 (0.5) 0.3 (0.5) 3.0 5.0
Nkanfo 0.0 (0.0) 0.0 (0.0) 0.0 (0.0) 0.0 0.0
*Standard deviation in parentheses.
20
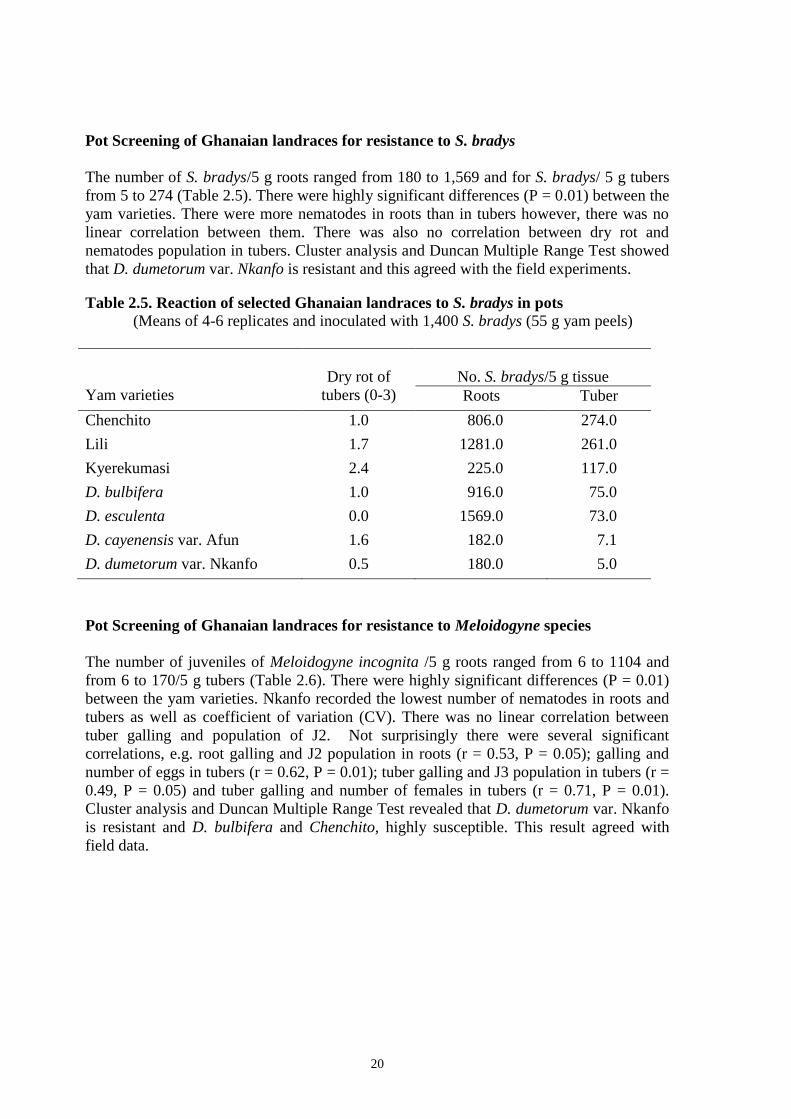
Pot Screening of Ghanaian landraces for resistance to S. bradys
The number of S. bradys/5 g roots ranged from 180 to 1,569 and for S. bradys/ 5 g tubers
from 5 to 274 (Table 2.5). There were highly significant differences (P = 0.01) between the
yam varieties. There were more nematodes in roots than in tubers however, there was no
linear correlation between them. There was also no correlation between dry rot and
nematodes population in tubers. Cluster analysis and Duncan Multiple Range Test showed
that D. dumetorum var. Nkanfo is resistant and this agreed with the field experiments.
Table 2.5. Reaction of selected Ghanaian landraces to S. bradys in pots
(Means of 4-6 replicates and inoculated with 1,400 S. bradys (55 g yam peels)
Yam varieties
Dry rot of
tubers (0-3)
No. S. bradys/5 g tissue
Roots Tuber
Chenchito 1.0 806.0 274.0
Lili 1.7 1281.0 261.0
Kyerekumasi 2.4 225.0 117.0
D. bulbifera 1.0 916.0 75.0
D. esculenta 0.0 1569.0 73.0
D. cayenensis var. Afun 1.6 182.0 7.1
D. dumetorum var. Nkanfo 0.5 180.0 5.0
Pot Screening of Ghanaian landraces for resistance to Meloidogyne species
The number of juveniles of Meloidogyne incognita /5 g roots ranged from 6 to 1104 and
from 6 to 170/5 g tubers (Table 2.6). There were highly significant differences (P = 0.01)
between the yam varieties. Nkanfo recorded the lowest number of nematodes in roots and
tubers as well as coefficient of variation (CV). There was no linear correlation between
tuber galling and population of J2. Not surprisingly there were several significant
correlations, e.g. root galling and J2 population in roots (r = 0.53, P = 0.05); galling and
number of eggs in tubers (r = 0.62, P = 0.01); tuber galling and J3 population in tubers (r =
0.49, P = 0.05) and tuber galling and number of females in tubers (r = 0.71, P = 0.01).
Cluster analysis and Duncan Multiple Range Test revealed that D. dumetorum var. Nkanfo
is resistant and D. bulbifera and Chenchito, highly susceptible. This result agreed with
field data.

21
Table 2.6. Reaction of selected Ghanaian landraces to Meloidogyne incognita in pots
(Means of 4-6 replicates and inoculated with 10000 Meloidogyne J2 (10 g galled tomato
roots)
Yam
varieties
Tuber
galling
(0-3)
Root
galling
(0-3)
J2/5 g
roots
Eggs/5 g
tuber
J2/5 g
tuber
J3/5 g
tuber
Females/5 g
tuber
Nkanfo 1.0 0.5 6.0 3.0 6.0 0.0 0.0
Afun 1.6 1.2 58.0 69.0 36.0 19.0 16.0
D. bulbifera 0.5 1.3 1104.0 1 0.0 170.0 0.0 3.0
Yeremma 1.6 2.0 821.0 55.0 13.0 10.0 8.0
Chenchito 2.4 2.4 634.0 322.0 99.0 162.0 60.0
GLASSHOUSE INVESTIGATION OF THE VARIATION IN SUSCEPTIBILITY OF YAM SPECIES TO
S. BRADYS, UK.
Table 2.7. Reaction of S. bradys in roots and tubers of yam species
(Mean of 4 replicates and inoculated with 500 S. bradys)
Mean no. S. bradys/5 g tissue
Yam species Roots Tuber
D. dumetorum 10630.0 800.0
D. abyssinica 1035.0 625.0
D. rotundata 89/01892 950.0 255.0
D. rotundata 91/00658 295.0 225.0
D. preussii 140.0 5.0
D. togoensis 140.0 0.0
There were no significant differences between treatments. The mean number of S. bradys
ranged from 140 to 10,630/5 g roots and 0 to 800/5 g tuber (Table 2.7). Again, there were
more nematodes in the roots than tubers. None of the entries exhibited symptoms of
nematode injury, furthermore there was no apparent relationship between susceptibility in
roots and tubers, suggesting that susceptibility in roots of yams is not clearly linked to the
susceptibility of tubers. D. togoensis and D. preussii were found to be potentially resistant
and D. dumetorum and D. abyssinica were highly susceptible. The results of this test
contrast sharply with others indicating that D. dumetorum has resistance to both S. bradys
and Meloidogyne incognita. This appears to indicate that resistance to nematodes is not
likely to be species specific.
FIELD SCREENING OF OPEN-POLLINATED SEEDLING POPULATION FOR S. BRADYS
RESISTANCE.
The dry rot symptoms ranged between 0 to 3 while the number of S. bradys was between 0
to 1,985/ 5 g tuber. Cluster analysis and Duncan Multiple Range test for dry rot scores and
the number of S. bradys/5 g tuber showed that 6 % of the genotypes are potentially
resistant whilst 94 % are highly susceptible.

22
POT SCREENING OF YAM PARENTAL LINES FOR RESISTANCE TO S. BRADYS
All the parental lines were susceptible to S. bradys and there were no significant
differences between them. The number of S. bradys/5 g roots ranged from 507 to 1,518
and for number of S. bradys/5 g from 9 to 138 (Table 2.8).
Table 2.8. Variation in susceptibility of yam parental lines to S. bradys
(Means of 4-7 replicates and inoculated with about 1,700 S. bradys)
Tuber cracking
(0-3)
S. bradys/5 g tissue
Yam parental lines Roots Tuber
TDr 93-1 1.1 1479.o 18.0
TDr 93-2 1.6 1225.0 104.0
TDa 92-2 0.7 792.0 42.0
TDa 95/00328 1.4 680.0 138.0
TDr 87/00571 1.6 1109.0 24.0
TDa 87/01091 1.0 716.0 13.0
TDa 95-310 1.0 507.0 9.0
TDa 85/00257 1.0 835.0 33.0
TDr 93-50 2.0 1518.0 59.0
TDr 87/00211 0.9 558.0 24.0
All the parental lines are susceptible in roots and tuber to S. bradys. However, variation in
susceptibility among the lines was observed. There was no correlation between the number
of S. bradys in roots and that in tubers.
POT SCREENING OF PARENTS AND PROGENIES OF INTRASPECIFIC HYBRIDISATIONS FOR
RESISTANCE TO NEMATODES
There were no significant differences between the genotypes regarding the number of S.
bradys in tubers. Again, no correlation was found between the numbers of nematodes in
roots and that in tubers. All the hybrids and their parents in the screen were susceptible to
S. bradys although there was quantitative variation in susceptibility. Selected contrasting
hybrids screened on the field were all susceptible and agreed with the pot trial. This
suggests that resistance of yam to S. bradys is not likely to be controlled by recessive
genes. The number of S. bradys/5 g root ranged from 16 to 4935 and that of S. bradys/5g
tuber from 0 to 259 (Table 2.9). Higher numbers of the nematodes were again found in
roots than in tubers.

23
Table 2.9. Reaction of yam intraspecific hybrids to S. bradys
(Means of 4–8 replicates inoculated with about 780 S. bradys)
Parents and hybrids
Tuber
cracking (0-3)
Mean no. S. bradys/5 g tissue
Roots Tuber
CR1/0096 0.7 450.0 16.0
CR1/0051 0.8 1098.0 50.0
CR/002 1.0 1795.0 42.0
CR1/0036 1.3 222.0 17.0
CR1/0058 1.0 1590.0 123.0
CR1/00102 1.0 470.0 8.0
CR1/00192 1.5 1036.0 238.0
CR1/0027 2.1 4935.0 259.0
CR1/0020 0.6 132.0 1.0
CR1/0029 1.0 781.0 11.0
CR1/00193 0.5 583.0 3.0
CR1/OO92 1.0 1658.0 16.0
CR1/0022 0.4 210.0 12.0
CR1/00172 1.8 1464.0 116.0
CR1/0095 1.5 1301.0 25.0
CR1/001 3.0 1362.0 180.0
CR1/0034 1.0 222.0 21.0
CR1/0048 2.0 1491.0 186.0
CR1/0065 1.3 237.0 12.0
CR1/0098 1.8 959.0 107.0
CR1/0044 1.3 259.0 25.0
CR1/0039 1.4 67.0 18.0
CR1/0024 0.5 1742.0 25.0
CR1/0091 1.8 1293.0 107.0
CR1/00146 0.7 890.0 23.0
CR1/0046 2.8 662.0 66.0
CR1/0025 0.5 1297.0 8.0
CR1/0038 1.3 368.0 38.0
CR1/OO64 1.2 325.0 58.0
CRI/0041 1.0 1169.0 30.0
CR1/00113 1.4 936.0 134.0
CR1/00130 0.3 209.0 23.0
CR1/0075 1.3 282.0 34.0
CR1/00138 1.2 728.0 91.0
CR1/0056 0.3 904.0 7.0
CR1/0035 1.3 40.0. 38.0
CR1/00173 1.3 316.0 83.0
CR1/0069 1.3 979.0 29.0
CR1/00129 0.8 208.0 7.0
CR1/0054 1.0 4689.0 152.0
CR1/0016 1.0 1288.0 0.0
CR1/0053 1.4 1060.0 31.0
CR1/00101 1.0 55.0 68.0

24
Table 2.9 (cont.)
Parents and hybrids
Tuber
cracking (0-3)
Mean no. S. bradys/5 g tissue
Roots Tuber
CR1/0040 0.6 1217.0 13.0
CR1/0037 1.0 536.0 8.0
CR1/0052 1.0 1681.0 51.0
CR1/0028 0.3 650.0 30.0
CR1/00105 1.3 304.0 4.0
CR1/00143 3.0 771.0 50.0
CR1/00126 1.0 175.0 45.0
CR1/0074 1.5 1161.0 147.0
CR1/00106 1.0 624.0 26.0
CR1/0093 1.0 610.0 71.0
CR1/0011 0.8 1745.0 42.0
87/00571 2.0 1194.0 14.0
87/00211 0.7 735.0 22.0
CR1/00139 2.7 446.0 116.0
CR1/0030 1.8 2741.0 115.0
CR1/0088 1.0 16.0 130.0
The parent (TDr 87/00211) was highly susceptible to Meloidogyne incognita (Table 2.10).
There was variation in susceptibility between hybrids, but very careful and precise
screening and selection would be required to utilise this variation with confidence.
Procedurally, it is appropriate to use all stages of Meloidogyne species for assessment of
susceptibility in yam tubers as there were significant differences (P = 0.01) between egg,
J3 and female counts in tubers.
Table 2.10. Reaction of yam intraspecific hybrids to Meloidogyne incognita
(Means of 4-8 replicates and inoculated with about 400 Meloidogyne J2s)
Yam hybrid
and parent
Tuber
galling
(0-3)
Root
Galling
(0-3)
Root
knot J2/5
g root
Root
knot
eggs/5 g
tuber
Root knot
J2/5 g
tuber
Root
knot
J3/5 g
tuber
Root
knot
female/5
g tuber
CR1/0090 2.0 1.7 1753.0 2042.0 94.0 131.0 388.0
TDr 87/00211 3.0 2.0 8808.0 0.0 74.0 124.0 173.0
CR1/0042 2.3 1.3 2423.0 280.0 93.0 25.0 141.0
CR1/0021 2.0 1.3 5404.0 84.0 33.0 31.0 132.0
CR1/00147 1.8 0.8 8780.0 89.0 31.0 56.0 128.0
CR1/00107 3.0 1.0 2896.0 38.0 8.0 13.0 96.0
CR1/00112 2.0 0.5 3994.0 83.0 24.0 13.0 65.0
CR1/0080 2.0 0.5 1108.0 0.0 4.0 17.0 56.0
CR1/0062 0.5 0.5 1172.0 15.0 11.0 12.0 49.0
CR1/0073 0.0 0.5 4621.0 65.0 0.0 16.0 49.0
CR1/0070 2.0 0.7 2436.0 15.0 2.0 7.0 46.0
CR1/0079 1.4 1.0 2263.0 15.0 0.0 20.0 42.0
CR1/0031 1.2 0.7 1321.0 39.0 17.0 3.0 32.0
CR1/0051 2.0 1.0 1033.0 0.0 3.0 3.0 24.0

25
Table 2.10 (cont.)
CR1/0033 1.0 0.2 1508.0 16.0 10.0 3.0 23.0
CR1/003 1.3 1.3 2710.0 12.0 4.0 7.0 20.0
CR1/00132 1.0 0.3 778.0 97.0 8.0 1.0 16.0
CR1/007 0.5 0.5 328.0 2.0 2.0 2.0 12.0
CR1/00118 1.5 0.5 674.0 9.0 21.0 15.0 12.0
CR1/0089 0.8 0.8 1293.0 37.0 15.0 2.0 10.0
CR1/00111 0.8 0.3 142.0 35.0 5.0 3.0 9.0
CR1/00123 1.8 0.3 690.0 15.0 10.0 0.0 8.0
CR1/00128 1.0 0.7 678.0 10.0 24.0 0.0 7.0
CR1/0077 1.0 1.0 1549.0 2.0 0.0 2.0 5.0
CR1/006 2.0 0.5 410.0 0.0 0.0 0.0 0.0
Field Screening of Cellular and Molecular technologies laboratory (IITA) accessions
and interspecific hybrids for resistance to S. bradys
The population density of S. bradys ranged from 17 to 2,502/5 g yam peelings (Table
2.11). There were highly significant differences (P = 0.01) between the accessions. There
was correlation (r= 0.39, P<0.01) between S. bradys population and dry rot symptom
expression. There was also correlation between all the plant characteristics like softness,
crack and population density. All the interspecific hybrids were highly susceptible to the
nematode. The wild accession, D. dumetorum was found to be resistant to S. bradys.
Table 2.11. Reaction of yam interspecific hybrids and accessions to S. bradys (Mean of 10 replicates after 8 weeks and inoculated with 2,500 S. bradys (50 g infected peels)
Yam accession
Tuber
cracking
(0-3)
Tuber
softness
(0-3)
Dry rot
index
(0-3)
Mean no. S. bradys/5 g
tuber peelings
TDr 98/00216 2.9 1.1 3.0 2502.0
TDr 89/00217 2.8 1.0 3.0 1156.0
TDr 97/01316 3.0 0.5 3.0 1105.0
TDr 97/01314 2.5 0.7 2.5 1028.0
Kappe 138 2.3 0.6 2.6 827.0
TDr 97/01306 2.5 0.9 2.6 654.0
TDr 97/01275 2.4 0.4 2.9 189.0
BP 128 2.4 0.6 2.9 134.0
D. dumetorum 0.0 0.0 0.0 17.0
Field screening of yam intraspecific hybrids for resistance to S. bradys
S. bradys population ranged for 373 to 1,054/5 g yam peelings and dry rot ranged from 2.6
to 3 (Table 2.12). There were no significant differences between clones. This implies that
all the clones are susceptible to S. bradys. The parents of the hybrids were also susceptible.

26
Table 2.12 Reaction of intraspecific hybrids to S. bradys 8 weeks after harvest
(Mean of 10 replicates and inoculated with 2,500 S. bradys (50g infected yam peels)
Yam hybrids
Tuber
cracking
(0-3)
Tuber
softness
(0-3)
Dry rot
(0-3)
Mean no. S. bradys/5g
CR1/0083 2.2 1.0 3.0 628.0
CR1/0074 2.1 0.9 2.9 696.0
CR1/0057 2.2 1.5 3.0 373.0
CR1/0024 2.1 1.1 3.0 746.0
CR1/0051 2.6 1.4 3.0 1054.0
CR1/0036 2.4 1.0 3.0 1023.0
CR1/0099 2.3 1.6 3.0 500.0
CR1/0075 1.5 1.2 2.6 973.0
CR1/0037 2.6 0.9 3.0 617.0
CR1/00143 2.7 1.0 3.0 472.0
OUTPUT 3 NEMATODE VARIABILITY ON YAM KNOWN
PARTICIPATORY RURAL APPRAISAL: NEMATODE PESTS PROBLEMS OF YAM IN GHANA
The symptoms of nematode injury were clearly identified by farmers. Most farmers were
very familiar with the dry rot of tubers caused by migratory endoparasitic nematodes. In
parts of the country, more than 90% of farmers had local names for the disease symptoms
e.g. nkronsa nkronsa or edwie (rashes). Farmers could readily identify tubers with dry rot
symptoms and these were rejected at planting or consumed early. Farmers estimated losses
from dry rot to be 21% (0-100) in the Forest zone and 30% (2-100) in the Savannah.
Losses from root knot nematode were estimated as 11% (0-40) in the forest and zero in the
savannah (Table 3.1). The prevalence of the symptoms was 83% of sites in the forest zone
and 100% in the savannah (Table 3.2). The dominant component crops are also shown. At
all sites, dry rot symptoms were associated with S. bradys and galling with Meloidogyne
incognita. In most years farmers grew 10-15 yam varieties mostly of Dioscorea rotundata,
but also D. alata, D. cayenensis, D. dumetorum and D. bulbifera. All were thought by
farmers to be susceptible to dry rot, but some varieties e.g. D. rotundata var. Lili and
species of D. alata were said to store better. Traditional yam varieties were often described
to be free of dry rot. There can be few cases where a nematode problem is culturally so
important and so well understood by farmers. This suggests that farmers would adopt a
resistant variety, if such is identified or developed. However, farmers grow a large number
of yam varieties for different reasons like taste and yield, so any new variety should fit this
background.
Table 3.1. Farmer’s estimates of the proportion of tubers affected by dry rot and root
knot nematode in Ghana
Agroecological zone Dry rot disease (%) Root knot galling (%)
Mean Range Mean Range
Forest-transitional 21 0-100 11 0-40
Guinea-Savannah 30 2-100 0 0

27
Table 3.2 Prevalence of dry rot and root knot nematodes in Ghana
Agroecological zone
Dry rot
(%)
RKN
(%)
Dominant component crops
Forest-transitional 81 72 Cassava, chilli/maize
Guinea-Savannah 100 24 Millet, Cassava, maize/sorghum
IDENTIFICATION AND DISTRIBUTION OF YAM NEMATODES IN GHANA
The survey of stored tubers in Ghana revealed that the prevalence of nematode was high
(>70% of sites) in both the forest and savannah agroecological zones. Root knot nematodes
were found at all sites in the Savannah, but were less prevalent in the forest zone.
Meloidogyne incognita was the species of root knot encountered and dry rot symptoms
were entirely associated with S. bradys. No other migratory endoparasite was encountered.

28
HOST RACE STATUS OF PRATYLENCHUS COFFEAE ON YAM IN GHANA
The number of P.coffeae/5g roots ranged from 0 to 743 and from 0 to 14 / 5g tubers in the
yam (Table 21). There was significant difference (P = 0.05) between the yam amd Musa
varieties. The Musa species recorded the highest number of nematodes in roots however,
Musa cv. Brodeyuo (true horn) was more susceptible. There was no correlation between
per cent root necrosis and P. coffeae population. Trials on the pest status of P. ccffeae on
yams in Ghana have been interesting.
Table 3.3 Host race status of P. coffeae on yam in Ghana
(Mean of 4-6 replicates and inoculated with 800 P. coffeae)
Yam and Musa varieties %Root
necrosis
Crack
(0-3)
Tuber
dry rot
(0-3)
No.
P.coffeae/5g
root
No.
P.coffeae/5g
tuber
D. dumetorum cv.
Nkanfo
44 0.2 0 5 0
D. cayenensis cv. Afun 43.3 0 0 35 0
D. bulbifera 62.5 0.3 0 73 0
D. alata cv. Yeremma 70 1 0 0 0
D. rotundata cv. Lili 67.5 1 0 27 10
D. esculenta 22.5 0 0 3 14
Musa cv. Asamienu 5 - - 190 -
Musa cv. Brodeyuo 17.5 - - 743 -
Table 3.4 Host race differentiation of P. coffeae from Ghana and Belize
Host species P. coffeae ex Ghana
(nematodes/g root)
P. coffeae ex Belize
(nematodes/g root)
Dioscorea togoensis 3.0 30.0
Dioscorea rotundata 1.0 341.0
Musa cv Grande Naine 256.0 6.0
P. coffeae is known to be a pest of yams in the Caribbean, Pacific and Central America but
nothing was known of its pest status on yam in Ghana, although it is widespread on
plantain. P. coffeae can multiply in roots of a number of species of Dioscorea (Table 3.3).
However, the nematode is unlikely to be a threat to yam production because tubers do not
generally support nematode reproduction. Pot studies in the UK (Table 3.4) reveal a clear
host preference between geographically distinct nematode isolates and shown that the
Ghanaian population of P. coffeae, is a Musa host race and contrasts with Central
American populations which do cause dry rot in yams. Musa varieties appear to differ in
their susceptibility to Ghanaian populations of P. coffeae; Grande Naine is the least
susceptible of the 3 varieties tested.

29
PROJECT PUBLICATIONS
Book chapter
Kwoseh, C.K., Plowright, R.A. and Bridge, J. (2001) Scutellonema bradys on Dioscorea
spp. In: Evaluating Plants for Resistance and Tolerance to Plant Parasitic Nematodes,
Starr, J.L, Cook,R. & Bridge, J. (Eds.). CAB International, Wallingford (In press)
Conference and Workshop Proceedings
Plowright, R.A. and Kwoseh, C.K. (1998) Farmers perceptions of nematode disease in
yams in Ghana and the prevalence of endoparasitic nematodes in stored stored tubers.
Proceedings the 24 International Nematology symposium, 4-9 August 1998, Dundee.
Kwoseh, C.K., Plowright, R. A., Stanfield, J. & Asiedu, R. (1998) Culturing Scutellonema
bradys on yam tuber slices. In: Proceeding of the 7th Triennial Symposium of the
International Society for Tropical Root Crops - Africa Branch, Cotonou, Benin, 11 - 17
October 1998. Poster.
Kwoseh, C.K., Plowright, R.A. and Bridge, J. (2000) Nematode pests of yams (Dioscorea
spp.) in Ghana. Proceeding of Workshop: Uptake by Farmers of Yam Research
Recommendations, Kumasi, Ghana, 16 March 2000, Natural Resources Institute,
Greenwich University, UK.
PhD Thesis
Kwoseh, C.K. (2000) Identification of resistance to major nematode pests of yams
(Dioscorea spp.) in West Africa. PhD thesis. Reading University. 200pp.
Internal Presentation
Kwoseh, C.K. 2000. The yam nematode (Scutellonema bradys) in West Africa and the
potential for its control through resistance. Presentation at DFID’s Crop Protection
Programmes’ Root Crops Cluster meeting, Natural Resources Institute, Greenwich
University, 19 September, 2000.
Reports
Meerman, J.C., Speijer, P.R., Asiedu, R. (1999) Establishment of the geographic
distribution of yam nematode pests in Nigeria, Ghana and the Republic of Benin. - in
collaboration with C. Kwoseh, J. Mudiope, R. Plowright, T.E. Sangoyomi. IITA annual
report for 1999.
Meerman, J.C., Speijer, P.R., Asiedu, R. (1999) Improvement of the methodologies for
screening of yam host plant response to nematodes. – in collaboration with R. Plowright,
C.Kwoseh. IITA annual report for 1999.
Meerman, J.C., Speijer, P.R., Asiedu, R. (1999) Screening for resistance in yam
germplasm to nematodes affecting yam - in collaboration with R. Plowright, C.Kwoseh*,
J. Mudiope. IITA annual report for 1999.

30
CONTRIBUTION OF OUTPUTS
This work has contributed greatly to the understanding of important pests of yams in West
Africa. Good sources of resistance have been identified in Dioscorea dumetorum but barriers
to interspecific hybridisation mean that this can not be exploited in the short term. Although
quantitative variation in susceptibility has been identified, it would seem unlikely that useful
nematode resistance will be found in D. rotundata itself. The development of novel cultural
control methods with farmers, based around the issues of clean seed and crop rotation is likely
to be more fruitful in the immediate future, particularly since farmers already utilise such
methods.










![[Clarinet_Institute] Baermann, Carl - Clarinet Method, Op.63 (Part 3)](https://static.fdocuments.us/doc/165x107/55721355497959fc0b9218eb/clarinetinstitute-baermann-carl-clarinet-method-op63-part-3.jpg)







