FACTORS INFLUENCING GROWTH AND POLYSACCHARIDE FORMATION …jb.asm.org/content/88/5/1433.full.pdf ·...
9
-JOURNAL OF BACTFRIOLOGY V0ol. 88, No. 5, 1). 1433-1441 NovemIiber, 1964 Copyright © 1964 Amiierican Society foir Microbiology Printed in U.S.A. FACTORS INFLUENCING GROWTH AND POLYSACCHARIDE FORMATION BY STRAINS OF CHROMOBACTERIUM VIOLA CEUM WILLIANI A. CORPE Department of Botany, Barnardl College, Columbia University, New Y-ork, N\ew Y-ork Received for publication 8 June 1964 ABSTRACT COR1'E, WILLIAM A. (Columbia University, New York, N.Y.). Factors influencing growth and polysaccharide formation by strains of Chromt o- bacteriumn ziolaceutmi. J. Bacteriol. 88:1433-1441. 1964.-Gelatinous Chromtiobacteriumn violaceuml strains elaborate a fibrous exopolysaccharide that forms a matrix in which growing cells are em- bedded. Stable, nongelatinous variants arising from gelatinous cultures were of two types: (i) those that had lost the ability to produce polysac- charide, and (ii) those that had lost the capacity to bind the polymer into a mnatrix. Gelatinous strains growing in Tryptone did not produce a matrix, but rather elaborated polysaccharide free into the medium. The effect of Tryptone was not one of selection of nongelatinous variants. Growth and polysaccharide synthesis in a defined medium were greatest when amino acids were substituted for ammonia as a nitrogen source. Best growth and polysaccharide yield occurred when the ratio of carbohydrate to nitrogen was in the order of 10:1. Depolymerization of formed polymer occurred in cultures grown with Casamino Acids as the nitro- gen source. Calcium ion stimulated polysaccharide formation. Iron, although stimulating growth, inhibited polysaccharide formation. Gelatinous strains of Chromobacterium vio- laceum produce an extracellular heteropolysac- charide (Corpe, 1960) which is firmly associated with the cells, causing agar colonies to be raised, rubbery, and adherent (Corpe, 1953). Within 2 months after isolating the soil bacteria into labo- ratory media, nongelatinous strains began to appear, suggesting that the polysaecharide-cell relationship had been modified in some manner, or that the organism had lost its polysaccharide- producing ability. Wilkinson, Duguid, and Edmunds (1954) de- scribed the production of slime by a noncapsu- lated variant of Aerobacter aerogenes which had been isolated from a capsulated culture. Both capsule and slime polysaceharides were similar chemically and antigenically, but the property responsible for binding the polymer into a co- hesive capsule had been lost. The physical displacement of polysaccharide from a capsule to a dispersible slime, though probably unrelated to polysaccharide formation, may indicate a change in the chemistry of the cell surface, as some of the experiments rel)orted in this paper suggest. MATERIALS AND 'METHODS Cultures. The organisms used in this study were all typically gelatinous when initially isolated from soil (Corpe, 1953). After several months of cultivation on laboratory media, nongelatinous colonies began to appear. Such variants could be isolated quite readily from these cultures, but frequently reverted to a mixture of colony types. By repeated selection, stable gelatinous and nongelatinous cultures were obtained. The variant colony types used in this study are de- scribed in Table 1. Aledia. Complex, undefined media were pre- pared from protein hydrolysates, leptone, Tryptone, and Vitamin Free Casamino Acids, as well as yeast extract (Difco). These ingredients were dissolved in distilled water in various con- centrations, in mixtures or alone. In most experi- ments, glucose was used in 1 % (w/v) concentra- tion. The medium was routinely adjusted to pH 7.0. The salt mixture used in defined media con- tained (in grams per liter): KH2PO4, 1.36; Na2HPO04, 2.13; MIgSO4.7H20, 0.2; FeSO4 7H20, 0.0005; and CaCl2, 0.005. The salts were dis- solved in distilled water, adjusted to pH 7.0, and autoclaved in tubes or flasks. Ammonium sulfate or other nitrogen source was added as sterile solution to give a final concentration of 0.05 to 0.1 % (w/v). Glucose served as the carbon source, and was also added aseptically to the sterile salts solution to give a final concentration of 1% (w/v). 1433 on June 5, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of FACTORS INFLUENCING GROWTH AND POLYSACCHARIDE FORMATION …jb.asm.org/content/88/5/1433.full.pdf ·...

-JOURNAL OF BACTFRIOLOGYV0ol. 88, No. 5, 1). 1433-1441 NovemIiber, 1964Copyright © 1964 Amiierican Society foir Microbiology
Printed in U.S.A.
FACTORS INFLUENCING GROWTH AND POLYSACCHARIDEFORMATION BY STRAINS OF CHROMOBACTERIUM
VIOLACEUMWILLIANI A. CORPE
Department of Botany, Barnardl College, Columbia University, New Y-ork, N\ew Y-ork
Received for publication 8 June 1964
ABSTRACT
COR1'E, WILLIAM A. (Columbia University, NewYork, N.Y.). Factors influencing growth andpolysaccharide formation by strains of Chromto-bacteriumn ziolaceutmi. J. Bacteriol. 88:1433-1441.1964.-Gelatinous Chromtiobacteriumn violaceumlstrains elaborate a fibrous exopolysaccharide thatforms a matrix in which growing cells are em-bedded. Stable, nongelatinous variants arisingfrom gelatinous cultures were of two types: (i)those that had lost the ability to produce polysac-charide, and (ii) those that had lost the capacityto bind the polymer into a mnatrix. Gelatinousstrains growing in Tryptone did not produce amatrix, but rather elaborated polysaccharide freeinto the medium. The effect of Tryptone was notone of selection of nongelatinous variants. Growthand polysaccharide synthesis in a defined mediumwere greatest when amino acids were substitutedfor ammonia as a nitrogen source. Best growth andpolysaccharide yield occurred when the ratio ofcarbohydrate to nitrogen was in the order of 10:1.Depolymerization of formed polymer occurred incultures grown with Casamino Acids as the nitro-gen source. Calcium ion stimulated polysaccharideformation. Iron, although stimulating growth,inhibited polysaccharide formation.
Gelatinous strains of Chromobacterium vio-laceum produce an extracellular heteropolysac-charide (Corpe, 1960) which is firmly associatedwith the cells, causing agar colonies to be raised,rubbery, and adherent (Corpe, 1953). Within 2months after isolating the soil bacteria into labo-ratory media, nongelatinous strains began toappear, suggesting that the polysaecharide-cellrelationship had been modified in some manner,or that the organism had lost its polysaccharide-producing ability.
Wilkinson, Duguid, and Edmunds (1954) de-scribed the production of slime by a noncapsu-lated variant of Aerobacter aerogenes which hadbeen isolated from a capsulated culture. Bothcapsule and slime polysaceharides were similar
chemically and antigenically, but the propertyresponsible for binding the polymer into a co-hesive capsule had been lost.The physical displacement of polysaccharide
from a capsule to a dispersible slime, thoughprobably unrelated to polysaccharide formation,may indicate a change in the chemistry of thecell surface, as some of the experiments rel)ortedin this paper suggest.
MATERIALS AND 'METHODS
Cultures. The organisms used in this study wereall typically gelatinous when initially isolatedfrom soil (Corpe, 1953). After several months ofcultivation on laboratory media, nongelatinouscolonies began to appear. Such variants could beisolated quite readily from these cultures, butfrequently reverted to a mixture of colony types.By repeated selection, stable gelatinous andnongelatinous cultures were obtained. Thevariant colony types used in this study are de-scribed in Table 1.
Aledia. Complex, undefined media were pre-pared from protein hydrolysates, leptone,Tryptone, and Vitamin Free Casamino Acids, aswell as yeast extract (Difco). These ingredientswere dissolved in distilled water in various con-centrations, in mixtures or alone. In most experi-ments, glucose was used in 1 % (w/v) concentra-tion. The medium was routinely adjusted to pH7.0.The salt mixture used in defined media con-
tained (in grams per liter): KH2PO4, 1.36;Na2HPO04, 2.13; MIgSO4.7H20, 0.2; FeSO4 7H20,0.0005; and CaCl2, 0.005. The salts were dis-solved in distilled water, adjusted to pH 7.0,and autoclaved in tubes or flasks. Ammoniumsulfate or other nitrogen source was added assterile solution to give a final concentration of0.05 to 0.1% (w/v). Glucose served as the carbonsource, and was also added aseptically to thesterile salts solution to give a final concentrationof 1% (w/v).
1433
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

J. BACTERIOL.
TABLE 1. Growth characteristics of stable gelatinousand nongelatinous variants of Chromobacterium
violaceurm
Type Description
GelatinousG
Colonies
Broth (static)
Broth (shaken)
NongelatinousNGM
Colonies
Broth (static)
Broth (shaken)
NongelatinousNGS
Colonies
Broth (static)
Broth (shaken)
Well raised, rubbery, ad-herent to agar surface
Rubbery, entire pellicle,broth clear, polysac-charide confined to thepellicle
Jelly-like, turbid mass ofcells and polysaccharidewhich were not sepa-
rable in the centrifuge
Flat edges, slightly raisedin the center, stickymucoid texture
Light ring of growth atthe surface of the me-
dium, broth turbid,polysaccharide remainsin supernatant fluidwhen cells removed bycentrifugation
Cultures turbid and vis-cous but cells removedby centrifugation, poly-saccharide free
Flat, smooth surface, but-tery texture
Cultures turbid, not vis-cous, no gelatinous pol-ysaccharide in the me-
dium or on the cellsCultures uniformly tur-
bid, nonviscous, no ge-
latinous polysaccharidein the medium or on thecells
Flasks containing 50 ml of complete mediumwere inoculated and incubated at 30 C on a
rotary shaker. The inoculum was grown for 24hr in synthetic medium, centrifuged, washedonce in sterile phosphate buffer (pH 7.0), andresuspended in buffer at the same concentrationas was the original culture. Absorbance of the
culture was about 0.300 at 600 m,u. Portions(0.5 ml) were used to inoculate flasks.
Methods. Growth of cultures was measuredturbidimetrically at 600 m,. Plate counts wereperformed on cultures grown in peptone-glucosebroth. Dilutions were made in sterile tap water,and were plated on the surface of peptone-glu-cose agar, by use of a sterile glass spreader.Colonies were counted and inspected for theirgelatinous or nongelatinous character (Corpe,1953).Bound gelatinous polysaccharide was measured
roughly by centrifuging a 10-ml volume ofshaken culture at 20,000 X g in a centrifuge(model AX; Lourdes Instrument Corp., Brook-lyn, N.Y.) for 15 min in a 15-ml tube, and byestimating the total volume of the gelatinouscell mass in milliliters. Free polysaccharideproduced by NGM stiains was precipitated fromculture supernatant fluids with 4 volumes of 95%(w/v) ethanol. The resulting precipitate wasdispersed in water, and was reprecipitated,drained, and dried in air. The precipitate wassuspended with warming in a measured volume ofwater and was cooled. Suitable portions wereanalyzed for hexose by reaction with anthrone(Neish, 1952). The method was used also todetermine the glucose remaining in the culturemedium. All anthrone carbohydrate determina-tions were done in triplicate.
Another crude measure of polysaccharideformation by NGM strains was done by deter-mining the viscosity of whole cultuies. For thispurpose, a 5-ml pipette which had a water flowtime of 13.05 sec was selected. Viscosity ofcultures was determined as the flow time ofculture divided by the flow time of water oruninoculated medium.
Free polysaccharide isolated from NGMcultures was isolated and purified by the methodspreviously applied to the gelatinous polymer(Corpe, 1960).Ammonia was determined quantitatively by
use of the micro-Kjeldahl distillation and titra-tion procedure described by Kabat and Mayer(1961). Total nitrogen was determined by themethod of Johnson (1941).
a-Keto acids were detected qualitatively bychromatography of their 2, 4-dinitrophenylhy-drazones with the use of methods described byCavallini, Frontali, and Toschi (1949).Fragments of gelatinous colonies were prepared
for electron microscopic examination by grinding
1434 CORPE
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
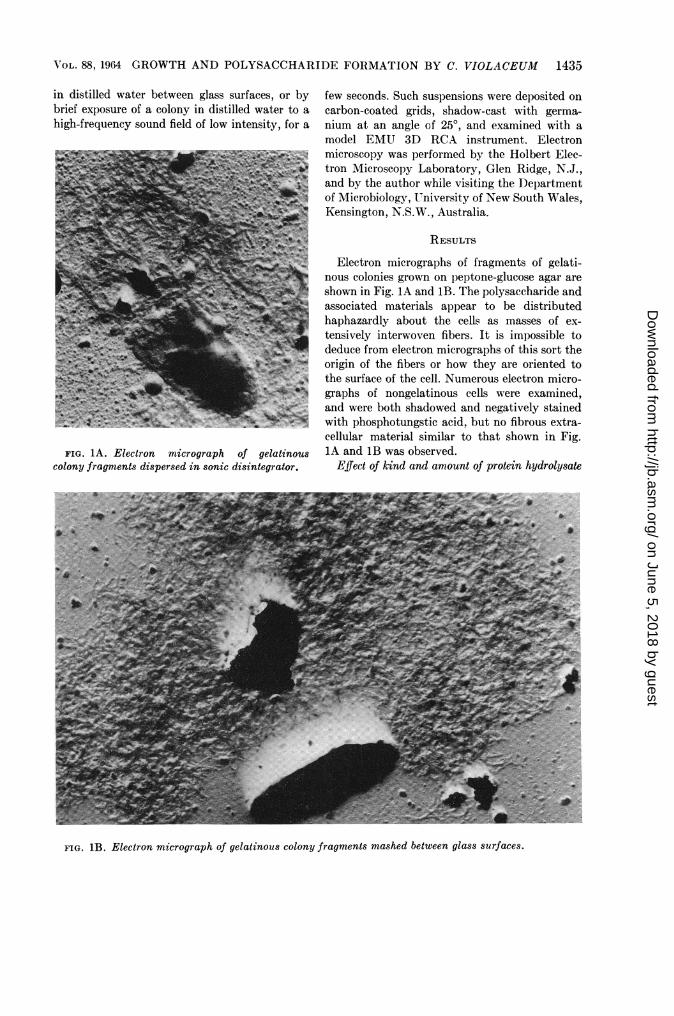
VOL. 88, 1964 GROWTH AND POLYSACCHARIDE FORMATION BY C. VIOLACEUM 1435
in distilled water between glass surfaces, or bybrief exposure of a colony in distilled water to ahigh-frequency sound field of low intensity, for a
FIG. 1A. Electron micrograph of gelatinouscolony fragments dispersed in sonic disintegrator.
few seconds. Such suspensions were deposited oncarbon-coated grids, shadow-cast with germa-nium at an angle of 25°, and examined with amodel EMU 3D RCA instrument. Electronmicroscopy was performed by the Holbert Elec-tron Microscopy Laboratory, Glen Ridge, N.J.,and by the author while visiting the Departmentof Microbiology, University of New South Wales,Kensington, N.S.W., Australia.
RESULTS
Electron micrographs of fragments of gelati-nous colonies grown on peptone-glucose agar areshown in Fig. IA and 1B. The polysaccharide andassociated materials appear to be distributedhaphazardly about the cells as masses of ex-tensively interwoven fibers. It is impossible todeduce from electron micrographs of this sort theorigin of the fibers or how they are oriented tothe surface of the cell. Numerous electron micro-graphs of nongelatinous cells were examined,and were both shadowed and negatively stainedwith phosphotungstic acid, but no fibrous extra-cellular material similar to that shown in Fig.1A and 1B was observed.
Effect of kind and amount of protein hydrolysate
FIG. 1B. Electron micrograph of gelatinous colony fragments mashed between glass surfaces.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

TABLE 2. Effect of protein hydrolysates on growth and gelatinous polysaccharide formation by 32G*
Glucose 1% (w/v) plus
Hydrolysate concn Peptone Tryptone Yeast extract Vitamin Free(%,W/v)smnoAcd
OD Gelatinous OD Gelatinous OD Gelatinous OD Gelatinouspolysaccharide polysaccharide polysaccharide polysaccharide
0.2 1.5 ++ 0.71 + 2.0 +++ 2.0 +0.5 1.7 +++ 1.35 + 0.72 + 1.75 +1.0 2.0 +++ 1.60 _ 0.58 + 0.002.0 2.0 + 1.70 0.46 0.004.0 1.7 + 1.50 _ 0.00 0.00
* All media were inoculated with 18-hr peptone-glucose broth culture and were incubated with shak-ing at 30 C for 72 hr. Optical densities (OD) were measured against an appropriate medium blank,and the amount of gelatinous polysaceharide-cell mass was estimated visually and was scored. Sym-bols: -, no polysaccharide; +, polysaccharide present.
TABLE 3. Effect of two protein hydrolysates on
growth and polysaccharide formation bygelatinous variants
Hydrolysate
Peptone +glucose
Tryptone +glucose
Age Strain
hr
17
41
88
168
184590
30G32G30G32G30G32G30G32G32G32G32G
Opticaldensity
0.7551.532.02.02.02.02.02.00.3972.02.0
Gelati-nous cells+ poly-saccha-ride*
ml
0.381.821.552.983.947.559.5010.00.100.250.25
Superna-tant fluidpolysac-charide*(gg/ml asfucose)
-t
NoneNone
None
29.087.592.0
* Volume of cell-bound polysaccharide per 10ml of culture.
t A negative sign indicates no test was made.
on grotwth and polysaccharide formation. Severalmedia, each of which contained glucose plus a
different protein hydrolysate in distilled water,were prepared. Cultures were incubated for up
to 6 days; optical density of the culture and theconcentration of the gelatinous polysaccharidewere measured at intervals (Table 2). All of thehydrolysates and yeast extract were, in varyingdegree, inhibitory to gelatinous polysaccharideformation when used in high concentration.Vitamin Free Casamino Acids at concentrationsof 1 % (w/v) or above were clearly inhibitory to
cell growth as well, but the higher concentrationsof peptone and Tryptone were only slightlyinhibitory. Yeast extract also adversely affectedthe growth of cells and gelatinous polysaccharideformation when used in concentrations above0.2% (w/v).When peptone-glucose broth was supplemented
with yeast extract or Casamino Acids to the ex-tent of 1% (w/v), growth and gelatinous poly-saccharide formation were inhibited.The influences of Tryptone and peptone on
growth and polysaccharide formation by gelati-nous (G) strains are compared in Table 3. Thepolysaccharide was bound to the cells only inpeptone + glucose medium. Removal of the cellsfrom peptone + glucose broth cultures by centrif-ugation effectively removed all of the polysac-charide as well. In Tryptone + glucose broth,the G strains produced polysaccharide which wasfree and could be recovered from culture super-natant fluids after removal of cells by centrifuga-tion.
Free polysaccharide, whether isolated fromcultures of NGM strains or from G strains grownon Tryptone medium, was heavily contaminatedwith protein. Solutions of crude polysaccharideshowed absorption at 280 m, but none at 260m,u. Polysaccharide precipitated from dialyzedculture supernatant fluids with 4 volumes ofacetone or 95% ethanol contained 8 to 10%total nitrogen. Fractional precipitation of thepolysaccharide with acetone, followed by depro-teinization with chloroform and butanol (10:1),lowered the nitrogen content to 5 to 7 %. A morevigorous purification procedure (Corpe, 1960)
1436 CORPE J. BACTERIOL.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 88, 1964 GROWTH AND POLYSACCHARIDE FORMATION BY C. VIOLACEUM 1437
was required to reduce the nitrogen content to1.9 to 3%O, a value consistent with that reportedfor gelatinous polysaccharide by Martin andRichards (1963).As mentioned previously, NGS strains, al-
though producing no gummy exopolysaccharide,did accumulate an alcohol-insoluble material intheir culture supernatant fluids. The materialgave a positive test with anthrone, indicating thepresence of a carbohydrate. The latter waschemically and physically distinct from thegummy exopolysaccharide of G and NGMstrains, but was apparently the same as the so-called flocculent polysaccharide recovered fromgelatinous strains and described previously(Corpe, 1960). A similar material was observedin culture filtrates of NGM strains as well. Thesepreparations contained 6-deoxyhexose and hep-tose, making them distinct chemically from thegelatinous or gummy polymer. They were alsoshown to contain about 45% protein on a dryweight basis (Corpe, unpublished data). Theflocculent polysaccharide appeared only whenalcoholic supernatant fluids were chilled or whenthe concentration of alcohol increased, or both;therefore, contamination of the gummy polysac-charides by this anthrone-positive material wasnot significant.The possibility that loss of the gelatinous
character in Tryptone was caused by selectionof nongelatinous mutants was ruled out, becausegelatinous growth appeared without a time lagwhen Tryptone cultures were transferred topeptone + glucose broth. Periodic plating ofTryptone + glucose broth cultures on peptone +glucose agar, until there were as few as ten viablecells per milliliter, yielded, invariably, gelatinouscolonies.
Freshly isolated strains of gelatinous chromo-bacteria did not invariably respond to Tryptoneas described here for the stable gelatinous mutant.Gelatinous colonies were observed on Tryptoneagar, and gelatinous growth was recognized inTryptone broth, but the length of time the growthremains gelatinous is short, and colonies changefrom their characteristic rubbery texture to astate which might be called "meaty." Suchcolonies are not adherent to the agar, and can bebroken up easily. This event was interpreted asthe result of autolysis because, microscopically,the cells are involuted and the colonies are sterile.The same sort of thing sometimes occurs inpeptone + glucose broth or agar, but only after
32 G
0
6 32 NGS
w
5 _
25 50 75 100 125 150 175TIME (his)
FIG. 2. Viable counts of gelatinous (32G) andnongelatinous (32NGS) variants grown in peptone +glucose broth on a shaker at 30 C.
a period of 2 weeks of growth at optimal temper-ature, rather than 3 to 4 days as is the case inTryptone medium.Of the three mutant types of chromobacteria
described here, the NGS type was by far themost difficult to maintain in culture. Viabilitycurves of an NGS strain and a G strain are shownin Fig. 2. Very early and rapid death of the NGStype is indicated. The rate of death of G strainswas not as fast; furthermore, viable cells werefound in cultures of this sort after 1 month at 30C. Tryptone used in 1% concentration in anotherwise similar medium is even less favorablefor survival of both NGS and G cultures.Some attempt was made to demonstrate an
enzyme in Tryptone-grown cells and culturefiltrates which would release polysaccharidefrom cells. Gelatinous colonies were suspended in6-day Tryptone culture supernatant fluid; ortris(hydroxymethyl)aminomethane (tris)-buff-ered suspensions of alumina-ground cells fromthese cultures. No disruption of the gelatinouscolony was observed in either instance. The trisbuffer (0.05 M) was adjusted to pH 8.7, identicalwith that of the culture at the time of harvest.In another experiment, a thick gelatinous pep-tone + glucose broth culture was pasteurizedin a 60 C water bath for 1 hr, dialyzed overnightagainst distilled water, and diluted to give alightly viscous suspension. Portions (5 ml) ofthe suspension were mixed with Tryptone culturesupernatant fluid or ground cells as described
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

J. BACTERIOL.
~200
~~~~~~~~~~~~~~1.04150 L
300 60 9 20 10
U-
~0.5
30 60 90 12D 190
TIME (hrs)
FIG. 3. Effect of iron (Fe++) and calcium (Ca++)on polysaccharide formation by an NGM variant indefined medium. The curve drawn with dotted linesrepresents growth of the organism in complete me-
dium.
1.0 -2-0z
141 *7540
D
0
0~~~~~.oo50_-~~~~~~~~@ \\\\XI25E~~~~~1.50-5 i-0 15 2.0
GLUCOSE, (Mgn/mi)
FIG. 4. Effect of carbohydrate concentration on
growth and polysaccharide formation by an NGMvariant in defined medium after 68 hr at 30 C. Theconcentration of NH4N was 0.1 mg/ml.
above. There was no change in viscosity, andcentrifugation at 20,000 X g for 30 min did notseparate cells from the polysaccharide over andabove that observed in controls, indicating thatno dissociation of polymer from the cells hadoccurred.
Growth and polysaccharide formation in de-fined media. When calcium was withheld fromthe salts mixture, polysaccharide yield by 32NGM was greatly reduced (Fig. 3). When ironwas also omitted, a somewhat greater amount ofpolysaccharide was produced. In the presence ofadded calcium, relatively large amounts ofpolysaccharide were formed but, again, a better
yield and more rapid production occurred wheniron was omitted. The final yield of cells in thesecultures was approximately the same, though theaddition of iron permitted a more rapid attain-ment of final cell yield. Increased concentrationsof calcium and iron salts above that describedgave no further increase in polysaccharide syn-
thesis or growth.Magnesium omission from the salt mixture had
a depressing effect on growth in both rate andfinal cell yield. It could not be replaced by eithercalcium or manganese. The addition of manganeseand molybdenum had no effect on either growthor polysaccharide yield.The main consideration in experiments with
defined media was to determine whether thekind and amount of various nitrogen sources
would affect polysaccharide yield. When am-
monium sulfate concentration was held constantat 0.1 mg/ml and the glucose level was increasedfrom 0.1 to 20 mg/ml, considerable differencesin growth response and polysaccharide formationwere observed (Fig. 4). Growth was best with 5to 10 mg/ml of glucose, and about 10 mg/ml ofglucose were best for polysaccharide formation.With the glucose concentration held at 5 mg/mland the ammonium nitrogen level varied, bestgrowth occurred with 1 to 2 mg/ml, but bestpolysaccharide yield occurred at 0.1 mg/ml(Fig. 5). Similar results were obtained when
30-g
200 1 -10-i
a
7
6
5
0.1 0.2 0.3 0.4 0.5 0.6
[NH:] Mgm N/mi
FIG. 5. Growth and polysaccharide formation byan NGM variant in defined medium containing 1%(w/v) glucose and various concentrations of am-
monium nitrogen. Flasks were incubated with shak-ing at 30 C for 65 hr.
1438 CORPE
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

AVOL. 88, 1964 GROWTH AND POLYSACCHARIDE FORMATION BY C. VIOLACEUM 1439
glutamate nitrogen was used as the nitrogensource (Fig. 6). Best growth and largest polysac-charide yield per unit volume of culture occurredwhen 0.1 mg of glutamate nitrogen and 10 mg ofglucose per milliliter were supplied. The largestpolysaccharide yield per unit weight of cellmaterial, however, occurred when 0.05 mg/ml ofglutamate nitrogen was supplied.
After growth of the organism, the glucose-ammonium salts medium had a pH of 4.8 to 5.0,and had accumulated a-ketoglutaric acid but noother carboxylic acid. The addition of excessNH4+ )revented accumulation of the acid andstimulated cell yield, but also reduced the yieldof polysaccharide.When glutamic acid was used in levels above
0.05 mg of glutamate nitrogen per milliliter, theculture became alkaline because of accumulationof ammonia in the medium, glucose disappeared,and a-ketoglutarate accumulated in only traceamounts. Yields of polysaccharide were notdecreased when the glucose + ammonium saltsmedium was maintained above pH 7.0.Ammonia, glutamate, and Vitamin Free Casa-
mino Acids as nitrogen sources for growth andpolysaccharide production by 32 NGM arecompared in Fig. 7. Glutamate and Casamino
'A---- --- --- -
0.2
FIG.6. Growth accandplehride frainb
La.~~~~~~~~~~~~~~~~~=
1000 7
0.2~~~~~~~~~~~~~~~~~0
FIG.6. Groth and poyacarddomainb
at30CforGOhr.0 C
2.0
tw
D
C-).LU.000
2.2
,2.0_
1.8
1.6 2
1.4 8U,
11.2
20 40 60 80 100 20
TIME (hrs)
FIG. 7. Comparative effect of glutamate (A),ammonia (E), and Casamino Acids (0) on growth(solid lines) and polysaccharide formation (dottedlines). The defined medium contained 1% (w/v)carbohydrate plus 0.16 mg/ml of nitrogen.
Acids were used more readily, in that final cellyield was attained faster than when ammonia wassupplied as the nitrogen source; however, themaximal cell yield was approximately the samein all three. Increase in viscosity of the cultureswas used as an index of polysaccharide produc-tion; the most noticeable events were that bothammonia and glutamate cultures took a longertime to reach a level of viscosity attained earlyin the Casamino Acid culture, and that theviscosity of the Casamino Acid culture fell offquite perceptibly after a maximal viscosity hadbeen reached. This culture also showed a dropin optical density, which was interpreted tenta-tively as an indication of autolysis.Another experiment was done with the use of
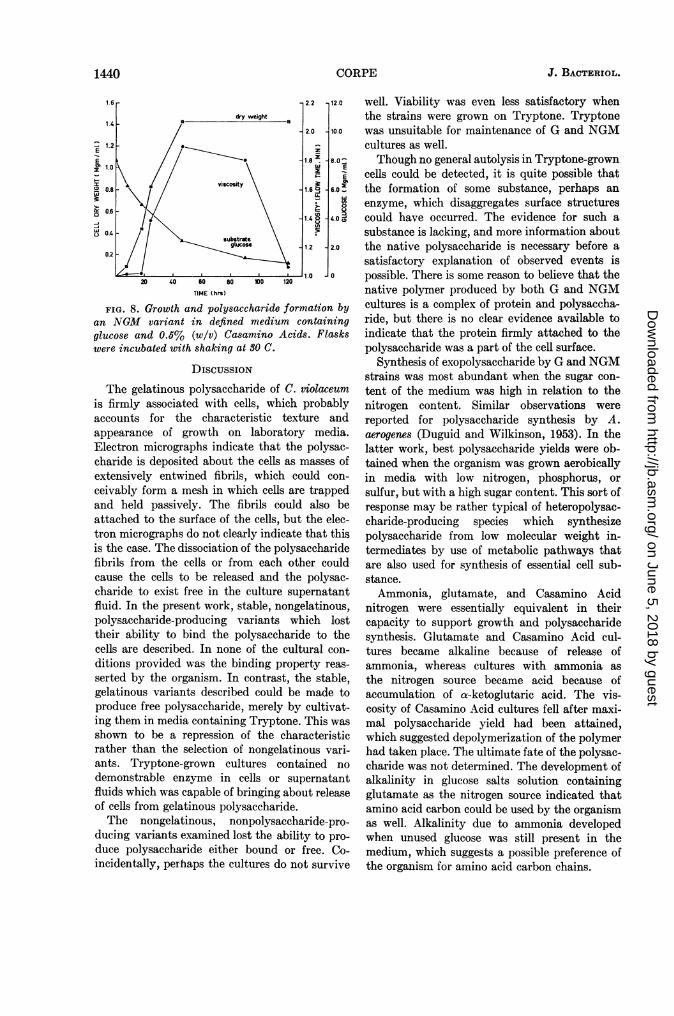
an increased amount of Casamino Acids (0.5%)over that which had been used in the previousexperiment (0.2%; Fig. 8). Though there was aconsiderable drop in the viscosity of the culture,there was no increase in alcohol-soluble carbohy-drate. Though a physical change is evident, theresults do not indicate what chemical changesmight have occurred. Change in polysaccharideviscosity was not accompanied by decrease inoptical density or in dry weight of cells. Allcultures grown on amino acid media becamestrongly alkaline if the amino acids were used inexcessive amounts or if the level of glucose waslow.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
J. BACTERIOL.
-g
3.0:-j
-J
20 40 60 80 100 120
TIME (hrs)
FIG. 8. Growth and polysaccharide formation byan NGM variant in defined medium containingglucose and 0.5% (w/v) Casamino Acids. Flaskswere incubated with shaking at 30 C.
DIscussIoN
The gelatinous polysaccharide of C. violaceumis firmly associated with cells, which probablyaccounts for the characteristic texture andappearance of growth on laboratory media.Electron micrographs indicate that the polysac-charide is deposited about the cells as masses ofextensively entwined fibrils, which could con-
ceivably form a mesh in which cells are trappedand held passively. The fibrils could also beattached to the surface of the cells, but the elec-tron micrographs do not clearly indicate that thisis the case. The dissociation of the polysaccharidefibrils from the cells or from each other couldcause the cells to be released and the polysac-charide to exist free in the culture supernatantfluid. In the present work, stable, nongelatinous,polysaccharide-producing variants which losttheir ability to bind the polysaccharide to thecells are described. In none of the cultural con-
ditions provided was the binding property reas-
serted by the organism. In contrast, the stable,gelatinous variants described could be made toproduce free polysaccharide, merely by cultivat-ing them in media containing Tryptone. This was
shown to be a repression of the characteristicrather than the selection of nongelatinous vari-ants. Tryptone-grown cultures contained no
demonstrable enzyme in cells or supernatantfluids which was capable of bringing about releaseof cells from gelatinous polysaccharide.The nongelatinous, nonpolysaccharide-pro-
ducing variants examined lost the ability to pro-
duce polysaccharide either bound or free. Co-incidentally, perhaps the cultures do not survive
well. Viability was even less satisfactory whenthe strains were grown on Tryptone. Tryptonewas unsuitable for maintenance of G and NGMcultures as well.Though no general autolysis in Tryptone-grown
cells could be detected, it is quite possible thatthe formation of some substance, perhaps anenzyme, which disaggregates surface structurescould have occurred. The evidence for such asubstance is lacking, and more information aboutthe native polysaccharide is necessary before asatisfactory explanation of observed events ispossible. There is some reason to believe that thenative polymer produced by both G and NGMcultures is a complex of protein and polysaccha-ride, but there is no clear evidence available toindicate that the protein firmly attached to thepolysaccharide was a part of the cell surface.
Synthesis of exopolysaccharide by G and NGMstrains was most abundant when the sugar con-tent of the medium was high in relation to thenitrogen content. Similar observations werereported for polysaccharide synthesis by A.aerogenes (Duguid and Wilkinson, 1953). In thelatter work, best polysaccharide yields were ob-tained when the organism was grown aerobicallyin media with low nitrogen, phosphorus, orsulfur, but with a high sugar content. This sort ofresponse may be rather typical of heteropolysac-charide-producing species which synthesizepolysaccharide from low molecular weight in-termediates by use of metabolic pathways thatare also used for synthesis of essential cell sub-stance.Ammonia, glutamate, and Casamino Acid
nitrogen were essentially equivalent in theircapacity to support growth and polysaccharidesynthesis. Glutamate and Casamino Acid cul-tures became alkaline because of release ofammonia, whereas cultures with ammonia asthe nitrogen source became acid because ofaccumulation of a-ketoglutaric acid. The vis-cosity of Casamino Acid cultures fell after maxi-mal polysaccharide yield had been attained,which suggested depolymerization of the polymerhad taken place. The ultimate fate of the polysac-charide was not determined. The development ofalkalinity in glucose salts solution containingglutamate as the nitrogen source indicated thatamino acid carbon could be used by the organismas well. Alkalinity due to ammonia developedwhen unused glucose was still present in themedium, which suggests a possible preference ofthe organism for amino acid carbon chains.
1440 CORPE
J 0
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 88, 1964 GROWTH AND POLYSACCHARIDE FORMATION BY C. VIOLACEUM 1441
Production of alkalinity in glucose-amino acidmedia was observed in Pseudomonas species(Rhodes, 1959; Gaby, Logan, and Whitaker,1962), and probably occurs in other species ofthis physiological type as well.
Synthesis of polysaccharide by chromobacteriawas stimulated by the presence of calcium inthe growth medium, which was most evidentwhen iron was omitted. The omission of iron,however, slowed the growth rate of the organism,although the final cell yield was not dramaticallyaffected. Martin and Richards (1963) includedCaCO3 in peptone + glucose medium used inproduction of gelatinous polysaccharide by agelatinous strain. The salt was undoubtedlystimulatory because of the presence of calcium,rather than any buffering capacity it mightsupply to the medium.
ACKNOWLEDGMENTS
This investigation was supported by PublicHealth Service research grant AI-04280 fromthe National Institute of Allergy and InfectiousDiseases. The author is grateful to A. D. Brownfor having read the manuscript while in prepa-ration and for his suggestions.
LITERATURE CITED
CAVALLINI, D., N. FRONTALI, AND G. ToSCHI.1949. Keto-acid content of human blood andurine. Nature 164:792-793.
CORPE, W. A. 1953. Variation in pigmentation andmorphology of colonies of gelatinous strainsof Chromobacterium species from soil. J.Bacteriol. 66:470-477.
CORPE, W. A. 1960. The extracellular polysac-charide of gelatinous strains of Chromobac-terium violaceum. Can. J. Microbiol. 6:153-163.
DUGUID, J. P., AND J. F. WILKINSON. 1953. Theinfluence of cultural conditions on polysac-charide production by Aerobacter aerogenes.J. Gen. Microbiol. 9:174-189.
GABY, W. L., C. LOGAN, AND S. WHITAKER. 1962.Catabolism of nitrogenous compounds byPseudomonas. J. Gen. Microbiol. 28:379-384.
JOHNSON, M. J. 1941. Isolation and properties of apure yeast polypeptidase. J. Biol. Chem. 137:575.
KABAT, E. A., AND M. M. MAYER. 1961. Experi-mental immunochemistry, 2nd ed. Charles CThomas, Publisher, Springfield, Ill.
MARTIN, J. P., AND S. J. RICHARDS. 1963. Decom-position and binding action of a polysac-charide from Chromobacterium violaceum. J.Bacteriol. 85:1288-1294.
NEISH, A. C. 1952. Analytical methods for bac-terial fermentations, 2nd revision. Nat]. Res.Council Can. Rept. no. 46-8-3.
RHODES, M. E. 1959. The characterization ofPseudomonas fluorescens. J. Gen. Microbiol.21:221-263.
WILKINSON, J. F., J. P. DUGUID, AND P. N. ED-MUNDS. 1954. The distribution of polysac-charide production in Aerobacter and Escher-ichia strains and its relation to antigeniccharacter. J. Gen. Microbiol. 11:59-72.
on June 5, 2018 by guesthttp://jb.asm
.org/D
ownloaded from