
Evidence for a polyspermy block at the level of sperm-egg plasma membrane fusion in Urechis caupo
8
Evidence for a Polyspermy Block at the Level of Sperm-Egg Plasma Membrane Fusion in Urechis caupo MILES PAUL AND MEREDITH GOULD-SOMERO Department of Bzology, Uniuersity of Victorza, Victoria, British Coliimbrn V8W2Y2 trnd Department of Biology, University of California, Son Bego, Ln Jolln, California 92037 ABSTRACT The results of sperm binding experiments reveal no change in the sperm binding properties of the egg surface coat at fertilization of UTechis caupo eggs. When fertilized eggs are reinseminated, sperm continue to attach to the egg surface coat. The acrosomal tubules of supernumerary sperm are observed in the perivitelline space closely apposed to the egg membrane. Thus, the polyspermy block in Urechis eggs involves neither alteration of sperm bind- ing sites nor inhibition of the acrosome reaction. Our results suggest that the block is at the level of sperm-egg membrane fusion. In animal eggs monospermic fertiliza- tion is prerequisite for normal development, and mechanisms are present which pre- vent more than one sperm from partici- pating in zygote formation. Several differ- ent strategies which ensure monospermic fertilization are known. In some cases many sperm normally penetrate the egg (“physiological polyspermy”), but follow- ing successful fusion of one of these with the female pronucleus, the rest degener- ate or are relegated to the periphery of the blastoderm. Physiological polyspermy, observed in many large yolky eggs such as those of elasmobranchs, urodeles, birds, and reptiles, may be necessary to ensure that at least one sperm nucleus will en- counter the egg nucleus (for a review, see Rothschild , ’56). In a large number of species, however, there are mechanisms which exclude all but the fertilizing sperm from the egg. The most thoroughly studied polyspermy blocks of this type are those in which the contents of cortical granules are discharged and cause an alteration in the egg surface coat, thus decreasing its capacity to bind sperm or its susceptibility to sperm pene- tration. This type of polyspermy block is found among the echinoids (Carroll and Epel, ’75; Tegner and Epel, ’73; Vacquier et al., ’72), anurans (Wyrick et al., ’74) and mammals (Barros and Yanagimachi, ’71; Gwatkin et al., ’73). There must also be other mechanisms for preventing penetration of supernumer- J. EXP. ZOOL., 196: 105-112 ary sperm since cortical granules either are not present in some eggs, e.g., tunic- ates (Ursprung and Schabtach, ’64) or do not discharge their contents in response to insemination, e.g., the bivalves Mytilus (Humphreys, ’67) and Spisula (Rebhun, ’62). There are several obvious possibili- ties. First, there could be some alteration in the egg surface layers resulting in a barrier to sperm attachment or penetra- tion, but which does not involve the con- tents of cortical granules. Second, there might be some process which prevents fusion of the sperm and egg plasma mem- branes. Such a process, which could be transient and insufficient in itself, may be present even in those eggs which have the cortical granule mediated processes. A third possibility could involve a change in the egg cortex which prevents the for- mation of a penetration cone. Eggs of Urechis caupo have a rapid block to polyspermy which begins to develop within 10 seconds of insemination (16°C) and is essentially complete by one minute (Paul, ’75a). Although Urechis eggs have cortical granules, there is no detectable change in these granules until at least four minutes after insemination when the elevation of the surface coat (also called the chorion or vitelline layer) begins, and even then the “cortical reaction” involves only limited and prolonged extrusion of granules into the perivitelline space. Indi- rect evidence suggests release of granule contents, but the mechanism by which 105
-
Upload
miles-paul -
Category
Documents
-
view
212 -
download
0
Transcript of Evidence for a polyspermy block at the level of sperm-egg plasma membrane fusion in Urechis caupo

Evidence for a Polyspermy Block at the Level of Sperm-Egg Plasma Membrane Fusion in Urechis caupo
M I L E S P A U L AND M E R E D I T H G O U L D - S O M E R O D e p a r t m e n t of Bzology, Un iuers i t y of Victorza, V ic tor ia , Bri t ish Coliimbrn V 8 W 2 Y 2 trnd D e p a r t m e n t of Biology, Un iver s i t y of Cali fornia, S o n Bego , Ln Jol ln , California 92037
ABSTRACT The results of sperm binding experiments reveal no change in the sperm binding properties of the egg surface coat at fertilization of UTechis caupo eggs. When fertilized eggs are reinseminated, sperm continue to attach to the egg surface coat. The acrosomal tubules of supernumerary sperm are observed in the perivitelline space closely apposed to the egg membrane. Thus, the polyspermy block in Urechis eggs involves neither alteration of sperm bind- ing sites nor inhibition of the acrosome reaction. Our results suggest that the block is at the level of sperm-egg membrane fusion.
In animal eggs monospermic fertiliza- tion is prerequisite for normal development, and mechanisms are present which pre- vent more than one sperm from partici- pating in zygote formation. Several differ- ent strategies which ensure monospermic fertilization are known. In some cases many sperm normally penetrate the egg (“physiological polyspermy”), but follow- ing successful fusion of one of these with the female pronucleus, the rest degener- ate or are relegated to the periphery of the blastoderm. Physiological polyspermy, observed in many large yolky eggs such as those of elasmobranchs, urodeles, birds, and reptiles, may be necessary to ensure that at least one sperm nucleus will en- counter the egg nucleus (for a review, see Ro thschild , ’56).
In a large number of species, however, there are mechanisms which exclude all but the fertilizing sperm from the egg. The most thoroughly studied polyspermy blocks of this type are those in which the contents of cortical granules are discharged and cause a n alteration in the egg surface coat, thus decreasing its capacity to bind sperm or its susceptibility to sperm pene- tration. This type of polyspermy block is found among the echinoids (Carroll and Epel, ’75; Tegner and Epel, ’73; Vacquier et al., ’72), anurans (Wyrick et al., ’74) and mammals (Barros and Yanagimachi, ’71; Gwatkin et al., ’73).
There must also be other mechanisms for preventing penetration of supernumer-
J. EXP. ZOOL., 196: 105-112
ary sperm since cortical granules either are not present in some eggs, e.g., tunic- ates (Ursprung and Schabtach, ’64) or do not discharge their contents in response to insemination, e.g., the bivalves Myt i lus (Humphreys, ’67) and Spisula (Rebhun, ’62). There are several obvious possibili- ties. First, there could be some alteration in the egg surface layers resulting in a barrier to sperm attachment or penetra- tion, but which does not involve the con- tents of cortical granules. Second, there might be some process which prevents fusion of the sperm and egg plasma mem- branes. Such a process, which could be transient and insufficient in itself, may be present even in those eggs which have the cortical granule mediated processes. A third possibility could involve a change in the egg cortex which prevents the for- mation of a penetration cone.
Eggs of Urechis caupo have a rapid block to polyspermy which begins to develop within 10 seconds of insemination (16°C) and is essentially complete by one minute (Paul, ’75a). Although Urechis eggs have cortical granules, there is no detectable change in these granules until at least four minutes after insemination when the elevation of the surface coat (also called the chorion or vitelline layer) begins, and even then the “cortical reaction” involves only limited and prolonged extrusion of granules into the perivitelline space. Indi- rect evidence suggests release of granule contents, but the mechanism by which
105

106 MILES P A U L AND MEREDITH GOULD-SOMERO
this occurs has not yet been established (Gould-Somero and Holland, ’75b). In any case, the “cortical reaction” is too slow to account for the polyspermy block in these eggs.
We have investigated the possibility that the polyspermy block in Urechis eggs in- volves some change in the extracellular surface coat, thus affecting sperm attach- ment or penetration through it. We find that there is no such change in this struc- ture. Furthermore, our observations imply that the polyspermy block in Urechis eggs is at the level of sperm-egg membrane fusion.
MATERIAL AND METHODS
Adult Urechis caupo were obtained from Elkhorn Slough (Moss Landing) or Bodega Bay, California, and maintained in glass tubes in fresh or recirculated seawater. Gametes were obtained and handled as previously described (Gould, ’67; Paul, ’75a). For electron microscopy, eggs were fixed and processed according to the method of Gould-Somero and Holland (‘75a) and examined on a Philips E M 300 electron microscope. Polyspermy was determined as previously described (Paul, ’75a).
In the assay of sperm binding to which we refer as the “continuous sperm bind- ing assay,” we used the “subtraction method’ of Vacquier and Payne (‘73) as follows. After equal volumes of egg sus- pension (10% vlv) and sperm suspension (“dry” sperm diluted 1:600) were mixed, aliquots were removed at the indicated times and added to an equal volume of formal- dehyde-sea water (15 ml 37% formalde- hyde + 85 ml sea water). To estimate the number of sperm which were not bound, eggs and bound sperm were allowed to settle at 1 x g , and the turbidity of the supernatant sea water was measured at
Controls (no sperm binding) were prepared by adding egg and sperm sus- pensions separately to formaldehyde-sea water. The turbidity was directly propor- tional to sperm concentration (determined with a hemocytometer) over the range of sperm concentrations in these experi- ments. The numbers of sperm bound per egg are calculated as the difference be- tween “total sperm” and “unbound sperm” in each case.
Attempts to measure sperm binding by
the “pronase method” of Vacquier and Payne (’73) were unsuccessful. In this method the eggs are washed to remove the unattached sperm in the supernatant sea water, then the bound sperm are released with pronase. However, in Urechis the sperm apparently do not adhere as tight- ly as in sea urchins, since the number of bperm remaining attached to the eggs de- creases with each successive wash.
We aIso investigated the possibility of a change in the capacity of the egg to bind additional sperm at various times after in- semination (“pulse binding assay”). In these experiments 2 92 egg suspensions were inseminated with a dilute sperm sus- pension (final sperm dilution, 1 :20,000). At the indicated times, aliquots were re- moved, concentrated by centrifugation, and added to a concentrated sperm suspen- sion (final egg dilution, 4% ; final sperm dilution, 1.1,200). After one or two min- utes, the mixture was added to an equal volume of formaldehyde-sea water and pro- cessed as described above.
RESULTS
T h e capaci ty of the surface coat to bind sperm does no t change after
fertilization Results of the continuous sperm bind-
ing assay revealed no significant release of bound sperm from the egg surface coat through 16 minutes in three of four ex- periments performed with different batches of eggs (A, fig. 1). We observed sperm de- tachment, however, after 3 0 4 5 minutes. In one of four experiments (B, fig. 1) sperm began to detach earlier. We attribute this early detachment to some change in the sperm (“ageing”) and not to a change in the binding capacity of the egg sur- face coat, since reinsemination of these same eggs resulted in binding of the new sperm (B’, fig. 1). This conclusion is con- firmed by a series of “pulse binding as- says.’’ In these experiments, eggs re- ceive an initial insemination with dilute sperm and are reinseminated with fresh sperm at intervals thereafter. All eggs were activated by the initial insemination. The capacity of eggs to bind fresh sperm does not change through 42 minutes after insemination (fig. 2). Indeed, we have ob- served that sperm continue to bind to cleav- age stages and even trochophore larvae.

POLYSPERMY BLOCK IN U R E C H I S EGGS 107
500-1 “iF\ U cl c 0 3 0 0 200
E 100
II 9111rll
5 10 15 20 25 30 0
r n i n
Fig. 1 The numbers of sperm bound to eggs as a function of time after insemination, following insemination at T = 0 (continuous sperm binding assay). In experiment A, no sperm detached through 16 minutes after insemination (the same result was obtained in two other similar experiments). In ex- periment B, sperm remained bound through eight minutes after insemination, then began to detach. Thirty minutes after insemination in experiment B , a sample of eggs was washed and reinseminated with fresh sperm to give the point B’; this demon- strates that the detachment observed was not due to a change in the eggs’ binding capacity but rather to sperm “ageing.”
(In Urechis, as in many other protostome embryos, the larvae do not hatch from the surface coat.)
The results of both of the above assays of sperm binding show that there is no change in the egg surface coat which af- fects the binding of sperm. However, we have observed that bound sperm can be dislodged from the surface if the eggs are subjected to turbulence (such as vortex stirring). This detachment is time depend- ent; sperm are not dislodged prior to five minutes after insemination, but can be completely removed by 10-1 5 minutes. Nev- ertheless, upon reinsemination, fresh sperm adhere to the eggs and, again, cannot be removed from the egg surface by vortex stirring until after five minutes. We inter- pret this as an “ageing” process of the sperm or sperm-egg interface but not a specific change in the properties of the surface coat.
Observations of bound sperm by electron microscopy
Heavily inseminated eggs may be sur- rounded by hundreds of supernumerary sperm. In these eggs, large numbers of acrosomal tubules penetrate through the surface coat into the perivitelline space
and lie close to the egg plasma membrane. In one heavily inseminated sample of eggs, 12-20 supernumerary sperm with acro- soma1 tubules in the perivitelline space were observed in single thin sections of eggs, though the eggs were only slightly polyspermic (average, 4 sperm per egg). We thus estimate that a minimum of sev- eral hundred such sperm must be as- sociated with each egg. A typical super- numerary sperm is shown in figure 3.4 (2 minutes postinsemination). Identification of these sperm as nonpenetrating sperm is unambiguous since penetration cones as- sociated with penetrating sperm are exten- sively developed by two minutes (fig. 3B), although the sperm are still external to the egg surface coat at this time. From this observation alone, it is clear that the block to polyspermy in Urechis eggs cannot act at a level which prevents the penetration of supernumerary acrosomal tubules into the perivitelline space.
Observations of eggs reinseminated at 10 and 20 minutes after an initial insem- ination provide further evidence that there is no change in the properties of the surface coat with respect to penetration of acrosomal tubules. In these experiments eggs were initially inseminated with a dilute sperm suspension followed by rein- semination with a concentrated sperm sus- pension at 10 or 20 minutes and fixed for electron microscopy two minutes later. A control group was inseminated only once
1 I r 2 4 6 8 10 12 14 42
m i n
Fig. 2 The numbers of sperm bound to pre- viously inseminated eggs during 2-minute “pulses“ with fresh sperm. A and B represent two separate experiments. No decrease in the eggs‘ sperm bind^ ing capacity is seen through 42 minutes after in^ semination. Abscissa. time after the initial insem- ination at which the 2-minute pulse ended; e.g.. the 5-minute point shows sperm bound during a 3 5 minute pulse after the initial insemination.

108 MILES PAUL AND MEREDITH GOULD-SOMERO
Fig. 3 Sperm-egg associations two minutes after insemination. A. Supernumerary sperm. 'The acrosomal tubule (at) penetrates through the surface coat (sc) and lies closely apposed to the egg plasma membrane. mv, microvilli in perivitelline space; mvt. microvillar tips pinched off at the end of oogenesis (Gould-Somero and Holland, '75a). B . Penetrating sperm as- sociated with a prominent sperm penetration cone (pc) which extends through the surface coat (sc). cg. cortical granules; j. egg jelly. A, X 28,000. B, x 32,000.
with the dilute sperm suspension and fixed at 10 or 20 minutes. We observed numer- ous sperm attached to the reinseminated eggs. Most of these sperm attached to the eggs during the second insemination, since we only rarely observed sperm on the sur- face of the control eggs. A large propor- tion of the supernumerary sperm were associated with an acrosomal tubule pene- trating into the perivitelline space.
In Urechis eggs the surface coat appears wrinkled during its elevation since initial-
ly elevation is localized. Acrosomal tubules are observed closely apposed to the egg surface in regions of slight elevation (fig. 4A). Their approximation to the egg mern- brane is limited to its microvillar exten- sions in regions of greater elevation (fig. 4B). In these regions the distal part of the acrosomal tubules usually remains just be- low the surface coat. In the perivitelline space the acrosomal tubule of nonpene- trating sperm generally appears as an ex- panded vesicle with invaginations (figs.
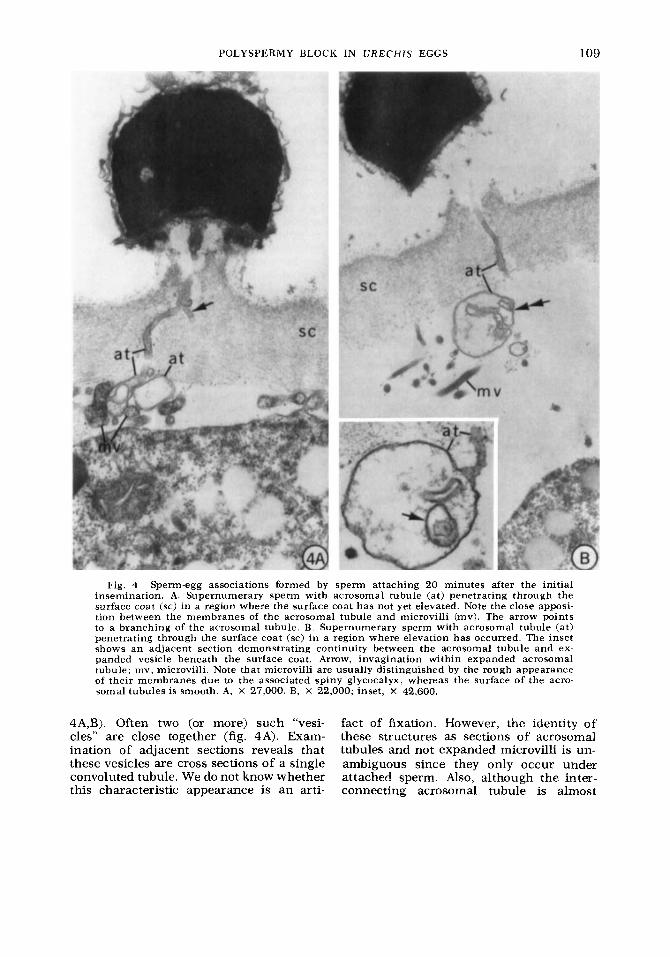
POLYSPERMY BLOCK IN U R E C H I S EGGS 109
Fig. 4 Sperm-egg associations formed by sperm attaching 20 minutes after the initial insemination. A. Supernumerary sperm with acrosomal tubule (at) penetrating through the surface coat (sc) in a region where the surface coat has not yet elevated. Note the close apposi- tion between the membranes of the acrosomal tubule and microvilli (mv). The arrow points to a branching of the acrosomal tubule. B. Supernumerary sperm with acrosomal tubule (at) penetrating through the surface coat (sc) i n a region where elevation has occurred. The inset shows an aaacen t section demonstrating continuity between the acrosomal tubule and ex- panded vesicle beneath the surface coat. Arrow, imagination within expanded acrosomal tubule; mv, microvilli. Note that microvilli are usually distinguished by the rough appearance of their membranes due to the associated spiny glycocalyx, whereas the surface of the acro- soma1 tubules is smooth. A, X 27,000. B, X 22,000; inset, X 42,600,
4A,B). Often two (or more) such “vesi- fact of fixation. However, the identity of cles” are close together (fig. 4A). Exam- these structures as sections of acrosomal ination of adjacent sections reveals that tubules and not expanded microvilli is un- these vesicles are cross sections of a single ambiguous since they only occur under convoluted tubule. We do not know whether attached sperm. Also, although the inter- this characteristic appearance is an arti- connecting acrosomal tubule is almost

110 MILES PAUL AND MEREDITH GOULD-SOMERO
never continuous in any one section, ex- amination of close or adjacent sections reveals the continuity of the vesicles with structures which are clearly acrosomal tubules (fig. 4B, inset).
Although we have examined many hun- dreds of such acrosomal tubules which are closely associated with either the egg sw- face proper or its microvillar extensions, we have never seen a fusion of the mem- branes. This implies that membrane fusion is a rare event and therefore that the polyspermy block affects this level of sperm Penetration.
DISCUSSION
In broad outline the sequence of events in gamete interaction leading to fertilization (sperm-egg contact, membrane fusion, and sperm penetration) appears similar in most animals for which details of this process are known (Austin, '68; Colwin and Col- win, '67). The most obvious stages at which polyspermy blocks might act to prevent penetration of supernumerary sperm are (1) sperm attachment, (2) the acrosome reaction and penetration through the egg surface coat(s), ( 3 ) the juxtaposition of sperm and egg plasma membranes, (4) fu- sion of sperm and egg plasma membranes, and (5) formation of the penetration cone.
In the case of Urechis , the results of our sperm binding assays indicate that the block does not act at the level of sperm attachment (1 above) for there is no de- tectable change in the ability of the sur- face coat to bind sperm following insem- ination. We have also demonstrated that the acrosome is able to penetrate the sur- face coat well after insemination. This eliminates the possibility (2 above) that the block acts by inhibiting the acrosome re- action or preventing penetration of the acrosomal tubule through the surface coat. Also, the observations (Gould-Somero and Holland, '75b) that there are neither ob- vious changes in the structure of the surface coat nor any exocytosis during the period in which the polyspermy block is established (Paul, '75a) are consistent with the apparent lack of functional modifica- tions indicated by the observations de- scribed above. Thus, the polyspermy block in Urechis is fundamentally different from that in the echinoderms (Carroll and Epel, '75; Vacquier et al., '72), the ham-
ster and probably several other mammals (Barros and Yanagimachi, '71: Gwatkin et al, '73) and anurans (Wyrick et al, '74) in which cortical granule discharge results in alteration of the surface coats to prevent further sperm binding and pene- tration.
Tyler ('65) proposed that withdrawal of microvilli from the egg surface coat might provide a mechanism for the polyspermy block in Urechis eggs. Microvilli may be required for the fusion process since their low radius of curvature would reduce the electrostatic repulsion between membranes thus permitting the close approximation required for membrane fusion (Poste and Allison, '73). Whether sperm-egg fusion in Urechis occurs via microvilli is not yet known. However, there is no change in the relation of microvilli to the surface coat during the first four minutes after insem- ination; after four minutes, the microvilli no longer penetrate as deeply into the ele- vating surface coat although they continue to extend throughout the perivitelline space. Our results demonstrate that prior to surface coat elevation, the acrosomal tubules of supernumerary sperm lie in close apposition to the plasma membrane of microvilli as well as intervening regions of the egg surface. Even after the surface coat has elevated, acrosomal tubules of supernumerary sperm become closely as- sociated with microvilli in the perivitelline space. We assume these supernumerary acrosomal tubules are potentially capable of fusion but that fusion is prevented by changes in the egg. (We have also ob- served that sperm are able to penetrate partially activated eggs in which elevation of the surface coat has already occurred; in these eggs the microvilli are withdrawn from the surface coat.)' Thus, several lines of evidence eliminate the possibility that the polyspermy block prevents the close juxtaposition of the sperm and egg plasma membranes.
In Urechis eggs, a prominent sperm penetration cone forms while the sperm nucleus is still outside the surface coat (fig. 3B). This cone is probably necessary for successful migration of the sperm nu- cleus which does not begin until after four minutes. If plasma membrane fusion
I M . Paul. i n preparation

POLYSPERMY BLOCK IN U R E C H I S EGGS 1 1 1
precedes formation of the entrance cone, a polyspermy block could act after sperm- egg fusion by preventing cone formation. Although cytoplasmic continuity would have been established, the nuclei of super- numerary sperm would not penetrate into the deeper cytoplasm. In this case, we would expect to see fusions between su- pernumerary sperm and egg membranes in the heavily inseminated samples exam- ined. However, we have not seen any clear instances of fusion among hundreds of acrosomal tubules observed in the perivi- telline space closely apposed to the egg plasma membrane.
By eliminating the other levels which the polyspermy block might affect, we are left with alternative 4 above, that the poly- spermy block in Urechis eggs acts by some mechanism preventing sperm-egg mem- brane fusion. The only alternative to this is that the acrosomal tubule of the fertil- izing sperm initiates formation of a pene- tration cone prior to membrane fusion, and the polyspermy block then acts to prevent formation of additional, "super- numerary" cones. Although we know of no precedent for this sequence of events in the fertilization of any other egg, this possibility cannot be eliminated at present.
Polyspermy blocks at the level of sperm- egg fusion have been suggested for sever- al other organisms, including those with cortical granule mediated blocks. For ex- ample, Rothschild and Swann ('52) have considered that the cortical reaction in sea urchin eggs is too slow to be effective in itself, and may be preceded by a "fast" block at the level of sperm-egg fusion. (However, this has been disputed [Gins- burg, '631 .) The block in the eggs of some mammals (the most thoroughly invest- tigated is the rabbit) acts at the level of fusion, although cortical granule discharge and alteration of the zona pellucida oc- cur as well (Chang and Hunt, '56; Con- rad et al., '71; Cooper and Bedford, '71; Overstreet and Bedford, '74). Also, mature toad eggs inseminated after the vitelline layer has been removed are monospermic while immature eggs under the same con- ditions are polyspermic, suggesting a poly- spermy block at the plasma membrane (Katagiri, '74). An example of an egg which establishes a polyspermy block that is not associated with any known altera-
tion in the surface coat is that of Spisula (Ziomek and Epel, '75). In this case also, the block is established in the absence of the surface coat and, in addition, is sensitive to the drug cytochalasin-B. How- ever, neither the mechanism of the block nor the level of sperm penetration which is blocked is known.
The block to sperm-egg fusion in Ure- chis might be effected by some altera- tion in the surface coat or contents of the perivitelline space that we have so far been unable to detect. A direct demon- stration that the polyspermy block is inde- pendent of the surface coat would be to show that the block is established in the absence of this structure, as has been demonstrated with Spisula eggs (Ziomek and Epel, '75). Washing in hypotonic glu- cose containing 5 mM EGTA (Gould-So- mero and Holland, '75b; also Engstrom, '71)z is the best method available for denuding Urechis eggs. Such eggs do not develop a polyspermy block. However, this pro- cedure renders the egg plasma membrane extremely fragile and may otherwise sig- nificantly alter its properties. Thus, the interpretation of these results remains ambiguous.
A variety of hypotheses has been pro- posed to explain the fusion of biological membranes at the molecular level (Poste and Allison, '73). Unfortunately, the little that is known at present about the molecular mechanisms involved in membrane fusion in general, let alone the specific fusion of the sperm and egg plasma membranes, limits useful speculation as to how the polyspermy blocks may affect this process. Models of membrane fusion, nevertheless, provide useful working hypothesis. If poly- spermy blocks act at the level of sperm- egg membrane fusion, as our results in- dicate in the case in Urechis eggs, then the molecular mechanism may be one which stabilizes the membrane against fusion. For example, this might be ac- complished by decreasing the motional freedom of membrane macromolecules due to calcium binding (Poste and Allison, '73). The possibility that calcium ion is specifically involved in the control of mem- brane fusion provides a useful working
2 The method used to remove the S p i s u l n egg surface coat (Ziomek and Epel. '75) i s not effective with Urrclz is eggs.

112 MILES PAUL AND MEREDITH GOULD-SOMERO
hypothesis for our effort to understand the nature of the polyspermy block in Urec his eggs.
ACKNOWLEDGMENTS
We thank C. L. Singla for technical as- sistance in the preparation of samples for electron microscopy, L. Holland for tech- nical assistance and help in preparation of the manuscript, R. Eggleston and T. Ronan for supplying us with adult Ure- chis, and D. H. Paul and D. Epel for critically reading the manuscript. This work was supported by grants from the National Research Council of Canada (NRCA6946) to M. Paul and from the Na- tional Institutes of Health (HD-03015) and the Population Council, New York (M74.85) to M. Gould-Somero.
LITERATURE CITED Austin, C. R. 1968 Ultrastructure of Fertiliza-
tion. Holt, Rinehart and Winston, New York. Barros, C., and R. Yanagimachi 1971 Induction
of zona reaction in golden hamster eggs by cor- tical granule material. Nature, (London), 233:
Carroll, E. G., and D. Epel 1975 Isolation and biological activity of the proteases released by sea urchin eggs following fertilization. Dev. Biol.,
268-269.
44: 2 2 3 2 . Chang, M. C., and D. M. Hunt 1956 Effects of
proteolytic enzymes on the zona pellucida of fer- tilized and unfertilized eggs. Expt. Cell Res., 1 1 : 497499 .
Colwin, L. H., and A. L. Colwin 1967 Membrane fusion in relation to sperm-egg association. In: Fertilization. C . B. Metz and A. Monroy, eds. Academic Press, Inc., New York, I: 295-367.
Conrad, K., J. Buckley and R. Stambaugh 1971 Studies on the nature of the block to polyspermy in rabbit ova. J. Reprod. Fert., 27: 133-136.
Cooper, G. W., and J. M. Bedford 1971 Charge density change in the vitelline surface following fertilization of the rabbit egg. J. Reprod. Fert., 25: 431436 .
Engstrom, W. S. 1971 Removal of the fertiliza- tion membrane of fertilized eggs of Urechis caupo and development of “membraneless” embryos. Biol. Bull., 140: 369-375.
Ginsburg, A. S. 1963 Mechanism of blockade of polyspermia in echinoderms. Doklady Adad. Nauk SSSR,152: 5 0 1 5 0 4 .
Gould, M. C. 1967 Echiuroid worms: Urechis. In: Methods in Developmental Biology. F. H. Wilt and N. K. Wessells, eds. Thomas Y. Crowell Co., New York, pp. 163-171.
Gould-Somero, M.,and L. Holland 1975a Oocyte
differentiation in Urechis caupo (Echiura): a fine structural study. J. Morph., 147: 475406 .
1975b Fine structural investigation of the insemination response in Urechis caupo. Dev. Biol., 46 : 358-369
Gwatkin, R. B. L., D. T. Williams, J. F. Hartmann and M. J. Kniazuk 1973 The mna reaction of hamster and mouse eggs. Production in vitro by a trypsin-like protease from cortical granules. J. Reprod. Fert., 32: 259-265.
Humphreys, W. J . 1967 Fine structure of cor- tical granules in eggs and gastrulae of Myti lus edulis. J. Ultrastruct. Res., 17: 314326 .
Katagiri, C. 1974 A high frequency of fertiliza- tion in premature and mature coelomic toad eggs after enzymic removal of vitelline membrane. J. Embryol. exp. Morph., 31: 573587 .
Overstreet, J. W., and J. M. Bedford 1974 Com- parison of the penetrability of the egg vestments in follicular oocytes, unfertilized and fertilized ova of the rabbit. Dev. Biol., 41 : 185-192.
Paul, M. 1975a The polyspermy block in eggs of Urechis caupo: evidence for a “rapid” block. Expt. Cell a s . , 90: 137-143.
Release of acid and changes in light-scattering properties following fertilization of Urechis caupo eggs. Dev. Biol., 43: 299412 .
Poste, G., and A. C. Allison 1973 Membrane fusion. Biochim. Biophys. Acta, 300: 421465 .
Rebhun, L. 1. 1962 Electron microscope studies on the vitelline membrane of the surf clam, Spisula solidissima. J. Ultrastruct. Res., 6: 107- 122.
Rothschild, Lord 1956 Fertilization. Holt, Rine- hart and Winston, New York.
Rothschild, Lord, and M. M. Swann 1952 The fertilization reaction in the sea urchin. The block to polyspermy. J. Expt. Biol., 29: 469483 .
Tegner, M. J., and D. Epel 1973 Sea urchin sperm- egg interactions: studies with the scanning elec- tron microscope. Science, 179: 685-688.
Tyler, A. 1965 The biology and chemistry of fertilization. Am. Nat., 99: 309334 ,
Ursprung, H., and E. Schabtach 1964 The fine structure of the egg of a tunicate, Ascidia nigra. J. Exp. Zool., 156: 253-268.
1972 Protease activity establishes the block against polyspermy in sea urchin eggs. Nature (Lon- don), 240: 352-353.
Vacquier, V. D., and J. E. Payne 1973 Methods for quantitating sea urchin sperm-egg binding. Expt. Cell Res., 82: 227-235.
1974 Agglutination of jelly coat and cortical granule components and the block to polyspermy in the amphibian Xenopus laevis. Proc. Natl. Acad. Sci. (U.S.A.), 71: 2067-2071.
Ziomek, C. A., and D. Epel 1975 A rapid cyto- chalasin-B sensitive alteration of the surface of Spisula solidissima eggs results in the block to polyspermy. Science (Washington, D.C.), 189:
1975b.
Vacquier, V. D., M. J. Tegner and D. Epel
Wyrick, R. E., T. Nishiharaand J. L. Hedrick
139-14 1 .


















