
Evidence for a foot-inhibiting substance in hydra
9
Cell differentiation, 5 (1976) 151--159 151 © Elsevier/North-Holland Biomedical Press, Amsterdam -- Printed in The Netherlands EVIDENCE FOR A FOOT-INHIBITING SUBSTANCE IN HYDRA Tobias SCHMIDT and H. Chica SCHALLER European Molecular Biology Laboratory, Heidelberg, West Germany Accepted 20 July 1976 Crude extracts of hydra contain a substance or substances which specifically inhibit foot formation in regenerating hydra. This foot inhibitor(s) is probably of low molecular weight, and it is present in the animal in an inactive, structure-bound form as a gradient from foot to head. The foot region contains 60% of this inhibitor in a 7--8 fold higher concentration than the other regions. The foot inhibitor was purified chemically from the head activator (Schaller, 1973) and the 'head' inhibitor (Berking, 1974). It is suggested that the substance under investigation is a new morphogen, responsible for foot inhibition in hydra. Because of its simple morphology and its high regeneration capacity, hy- dra is an ideal system to study processes of morphogenesis and differentia- tion. In regeneration experiments the polarity of the animal is strictly main- tained. Many models have been proposed to explain this phenomenon (Bur- nett, 1968; Webster, 1971; Wolpert, 1971; Gierer et al., 1972). Experimen- tal evidence seems to favour the notion that in hydra, polarity is controlled by substances emanating either from the head or the foot region. So far two substances have been described, one stimulatory (Schaller, 1973) and one inhibitory (Berking, 1974), which may control head formation in hydra and which are probably responsible for the gradients of activation and in- hibition extending from head to foot in hydra. MacWilliams et al. (1968, 1970, 1974) showed by transplantation experi- ments that the foot region has control functions comparable to and probably independent from the head region. We show in this paper that a foot-inhibit- ing substance can be isolated from crude extracts of hydra and that this sub- stance may be responsible for the gradient in foot inhibition extending from foot to head in hydra. MATERIALS AND METHODS Animals Hydra attenuata were mass-cultured in a medium slightly modified accord- ing to Lenhoff (1970) containing 1 mM CaCl:, 1 mM NaC1, 0.1 mM KC1, 0.1 mM MgC12, 0.125 mM NaHCO3 in deionised water, pH 7.8, at 20 +- 2°C.
-
Upload
tobias-schmidt -
Category
Documents
-
view
215 -
download
1
Transcript of Evidence for a foot-inhibiting substance in hydra

Cell differentiation, 5 (1976) 151--159 151 © Elsevier/North-Holland Biomedical Press, Amsterdam -- Printed in The Netherlands
E V I D E N C E F O R A F O O T - I N H I B I T I N G S U B S T A N C E IN H Y D R A
Tobias SCHMIDT and H. Chica SCHALLER
European Molecular Biology Laboratory, Heidelberg, West Germany
Accepted 20 July 1976
Crude extracts of hydra contain a substance or substances which specifically inhibit foot formation in regenerating hydra. This foot inhibitor(s) is probably of low molecular weight, and it is present in the animal in an inactive, structure-bound form as a gradient from foot to head. The foot region contains 60% of this inhibitor in a 7--8 fold higher concentration than the other regions. The foot inhibitor was purified chemically from the head activator (Schaller, 1973) and the 'head' inhibitor (Berking, 1974). It is suggested that the substance under investigation is a new morphogen, responsible for foot inhibition in hydra.
Because o f its s imple m o r p h o l o g y and its high r egene ra t ion capac i ty , hy- dra is an ideal s y s t e m to s t u d y processes of m o r p h o g e n e s i s and d i f fe ren t ia - t ion . In r egene ra t ion e x p e r i m e n t s the po l a r i t y o f the an imal is s t r ic t ly main- ta ined. Many mode l s have been p r o p o s e d to expla in this p h e n o m e n o n (Bur- ne t t , 1968 ; Webster , 1971; Wolper t , 1971; Gierer e t al., 1972) . E x p e r i m e n - tal ev idence seems to f avour the n o t i o n t h a t in hyd ra , po la r i ty is con t ro l l ed b y subs tances e m a n a t i n g e i ther f r o m the head or the f o o t region. So far t w o subs tances have been descr ibed , one s t i m u l a t o r y (Schaller , 1973) and one i nh ib i t o ry (Berking, 1974) , which m a y con t ro l head f o r m a t i o n in h y d r a and which are p r o b a b l y respons ib le fo r the gradients o f ac t iva t ion and in- h ib i t ion e x t e n d i n g f r o m head to f o o t in hydra .
MacWill iams e t al. ( 1968 , 1970 , 1974) s howed by t r a n s p l a n t a t i o n exper i - m e n t s t ha t the f o o t region has con t ro l func t ions c o m p a r a b l e to and p r o b a b l y i n d e p e n d e n t f r o m the head region. We show in this p a p e r t ha t a foo t - inh ib i t - ing subs t ance can be i so la ted f r o m c rude ex t r ac t s o f h y d r a and t h a t this sub- s tance m a y be respons ib le fo r the grad ien t in f o o t inh ib i t ion e x t e n d i n g f r o m f o o t to head in hydra .
MATERIALS AND METHODS
Animals
Hydra at tenuata were mass -cu l tu red in a m e d i u m sl ightly m o d i f i e d accord- ing to L e n h o f f (1970) con ta in ing 1 mM CaCl : , 1 mM NaC1, 0.1 mM KC1, 0.1 mM MgC12, 0 .125 mM NaHCO3 in de ionised wate r , p H 7.8, a t 20 +- 2°C.
152
T h e y were fed dai ly wi th naupl i i o f Artemia salina and washed 6- -8 h af ter feeding.
Crude extracts
H y d r a or h y d r a t issue were son ica ted in disti l led wa te r unde r ice-cooling for 3--5 min with a Branson B12 u l t r a son ica to r at 4 0 - - 5 0 wat ts . The absorp- t ion at 280 n m was m eas u red to e s t ima te the c o n c e n t r a t i o n of the crude ex- t ract .
Statistical analysis
I n h i b i t o r y e f fec t s were ascer ta ined by c o m p a r i n g m e a n values b y means of the t- test or b y c o m p a r i n g abso lu te n u m b e r s by means o f the ×2.test .
RESULTS
Assay for foot inhibition
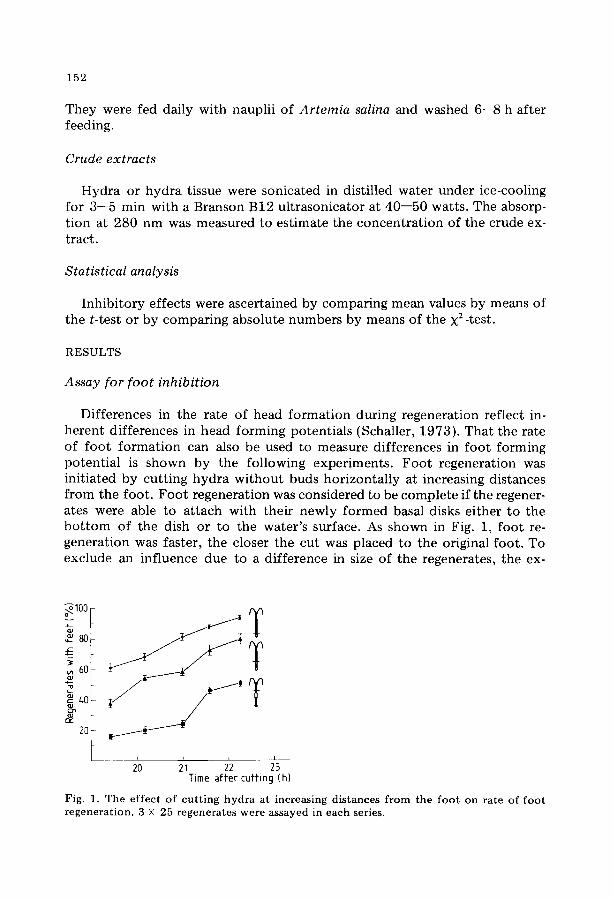
Dif fe rences in the ra te o f head f o r m a t i o n dur ing regenera t ion ref lec t in- he ren t d i f fe rences in head f o r m i n g po ten t i a l s (Schaller , 1973) . T h a t the ra te of f o o t f o r m a t i o n can also be used to measu re d i f fe rences in f o o t f o r m i n g po t en t i a l is shown b y the fo l lowing expe r imen t s . F o o t r egenera t ion was in i t ia ted by cu t t ing h y d r a w i t h o u t buds h o r i z o n t a l l y at increasing dis tances f r o m the foo t . F o o t r egenera t ion was cons idered to be c o m p l e t e if the regener- ates were able to a t t ach wi th the i r newly f o r m e d basal disks e i ther t o the b o t t o m o f the dish or to the wa te r ' s surface. As shown in Fig. 1, f o o t re- gene ra t ion was fas ter , the closer the cu t was p laced to the original foo t . To exc lude an inf luence due to a d i f fe rence in size of the regenera tes , the ex-
°•--100 t
"I 2'0 2hi 2'2 2L3 Time after cuffing (h)
Fig. 1. The effect of cutting hydra at increasing distances from the foot on rate of foot regeneration. 3 X 25 regenerates were assayed in each series.
153
~100 I
}.o T T I
21 2'2 2'3 24 Time affer cuffing (h)
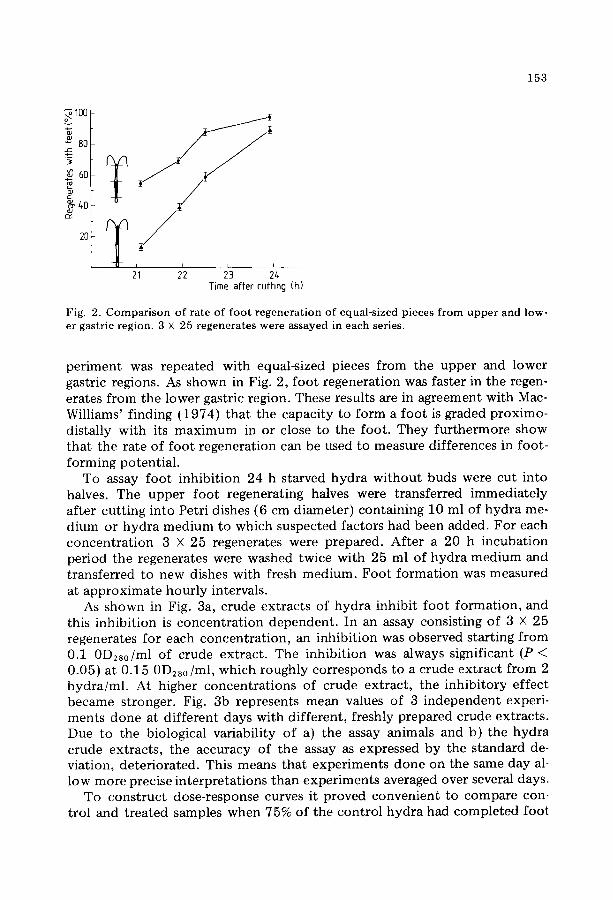
Fig. 2. C o m p a r i s o n of ra te of f oo t r egene ra t i on of equal-sized pieces f rom u p p e r and low- er gastric region. 3 x 25 regenera tes were assayed in each series.
periment was repeated with equal-sized pieces from the upper and lower gastric regions. As shown in Fig. 2, foot regeneration was faster in the regen- erates from the lower gastric region. These results are in agreement with Mac- Williams' finding (1974) that the capacity to form a foot is graded proximo- distally with its maximum in or close to the foot. They furthermore show that the rate of foot regeneration can be used to measure differences in foot- forming potential.
To assay foot inhibition 24 h starved hydra without buds were cut into halves. The upper foot regenerating halves were transferred immediately after cutting into Petri dishes (6 cm diameter) containing 10 ml of hydra me- dium or hydra medium to which suspected factors had been added. For each concentration 3 X 25 regenerates were prepared. After a 20 h incubation period the regenerates were washed twice with 25 ml of hydra medium and transferred to new dishes with fresh medium. Foot formation was measured at approximate hourly intervals.
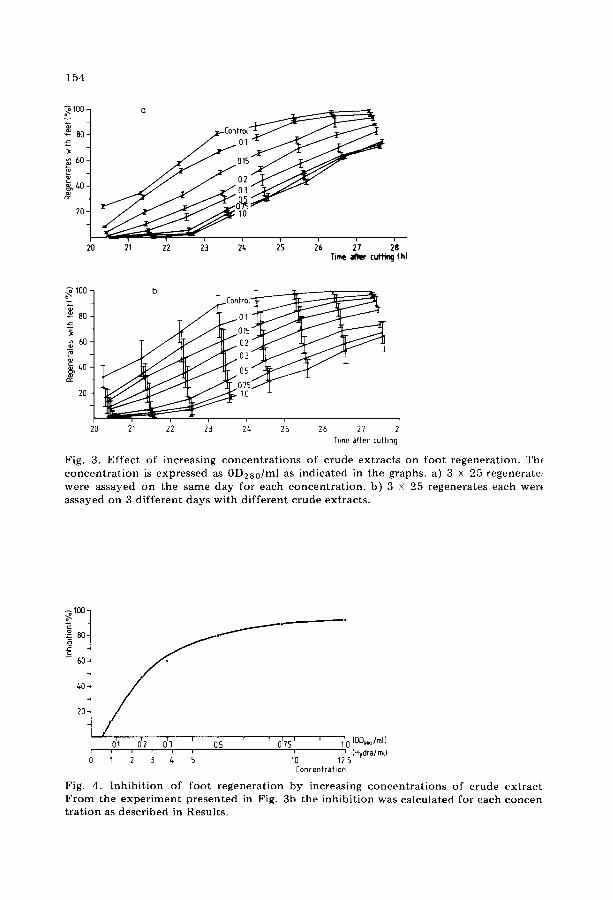
As shown in Fig. 3a, crude extracts of hydra inhibit foot formation, and this inhibition is concentration dependent. In an assay consisting of 3 X 25 regenerates for each concentration, an inhibition was observed starting from 0.1 0D280/ml of crude extract. The inhibition was always significant (P < 0.05) at 0.15 0D280/ml, which roughly corresponds to a crude extract from 2 hydra/ml. At higher concentrations of crude extract, the inhibitory effect became stronger. Fig. 3b represents mean values of 3 independent experi- ments done at different days with different, freshly prepared crude extracts. Due to the biological variability of a) the assay animals and b) the hydra crude extracts, the accuracy of the assay as expressed by the standard de- viation, deteriorated. This means that experiments done on the same day al- low more precise interpretations than experiments averaged over several days.
To construct dose-response curves it proved convenient to compare con- trol and treated samples when 75% of the control hydra had completed foot
154
~ 1 0 0 -
-
8 0 - .a=
-
60 -
~ - m
20-
20 ~ ;2 ;3 2~ ;s h 2'7 2'8 Time ~ cuWmg (h)
o~ 100
80 .zz
60 L
20
b
20 21 22
C o n f r o t ~
015
O3 05
i i i i r
24 25 26 27 2 Time affer cutting
Fig. 3. Ef fec t of increas ing c o n c e n t r a t i o n s of c rude ex t rac t s on foo t regenera t ion . Th( c o n c e n t r a t i o n is expressed as 0D2s0 /ml as ind ica ted in the graphs, a) 3 x 25 regeneratet were assayed on the same day for each c o n c e n t r a t i o n , b) 3 x 25 regenera tes each wer( assayed on 3 d i f f e ren t days wi th d i f f e ren t c rude ex t rac ts .
1 0 0 -
_ 8 0 -
6 0 -
- °
60-. /
20-
0'I 0'2 0~3 ' 0~5 ' r 0'75' ' 110 (OD,ao/ml}
; ; ~ ~ ~ ; ~'o ~2's (H~I~) Con(:enfrafion
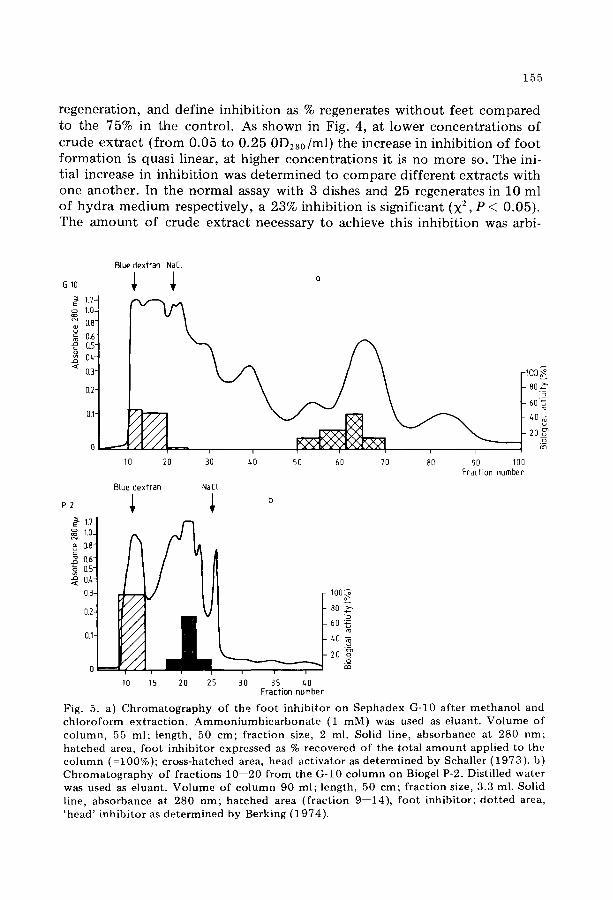
Fig. 4. I n h i b i t i o n of f oo t r egene ra t ion by increasing c o n c e n t r a t i o n s of c rude ex t r ac t F r o m the e x p e r i m e n t p r e sen t ed in Fig. 3b the i nh ib i t i on was ca lcula ted for each concert t r a t i on as descr ibed in Resul ts .
155
regenerat ion, and def ine inhib i t ion as % regenerates w i t h o u t feet compared to the 75% in the control . As s h o w n in Fig. 4, at lower concentrat ions of crude extract ( from 0 .05 to 0 .25 0D280/ml) the increase in inhib i t ion o f f oo t format ion is quasi linear, at higher concentrat ions it is no more so. The ini- tial increase in inh ib i t ion was determined to compare different extracts with one another. In the normal assay with 3 dishes and 25 regenerates in 10 ml o f hydra medium respect ively , a 23% inhibi t ion is s ignif icant (X 2, P < 0 .05) . The a m o u n t o f crude extract necessary to achieve this inh ib i t ion was arbi-
8(ue dextran NaCt
+1o ; ; ° ~ 1.7+ o 1,0-
~' 0.8-
g 0.6- .o 0.s-
0.(,-
< 0.3.
0.2 +
0.1-
0 ~
10 20 30 L,O 50 60 70 80 90 100 Fraction number
Btue dextran Na Ct
1.7- 1.0-
CL8- A 0.6" 0.5"
Q 0.~-
0.3- ~ I /F 100~ I
0.1' [- ~.0
""-------~ --...-F 20 ~ 0 ' i i i / i i i
10 15 20 25 30 35 /+0 Fraction number
Fig. 5. a) Chromatography of the foot inhibitor on Sephadex G-10 after methanol and chloroform extraction. Ammoniumbicarbonate (1 raM) was used as eluant. Volume of column, 55 ml; length, 50 era; fraction size, 2 m]. Solid line, absorbanee at 280 nm; hatched area, foot inhibi tor expressed as % recovered of the total amount applied to the column (=100%); cross-hatched area, head activator as determined by Schaller (1973). b) C h r o m a t o g r a p h y o f f r ac t i ons 1 0 - - 2 0 f r o m the O-10 c o l u m n o n Bioge] P-2. Dist i l led w a t e r w a s used as e luan t . V o l u m e o f c o l u m n 90 ml ; l eng th , 50 c m ; f r ac t ion size, 3.3 ml. Solid line, a b s o r b a n c e at 280 n m ; h a t c h e d area ( f r a c t i o n 9 - -14) , f o o t i nh ib i to r ; d o t t e d area, ' h e a d ' i n h i b i t o r as d e t e r m i n e d by Berk ing (1974) .
i ~oo~ 8o.~
4o- d

156
trarily defined as one biological unit (1 BU). Accordingly the specific activ- ity of a given extract is expressed as BU/0D280.
Purification of the foot inhibitor from other hydra morphogens
The foot- inhibi tory activity was purified according to the following scheme: 1) Crude extracts were prepared by homogenising and sonicating hydra in distilled water. 2) The freeze-dried crude extracts were extracted extensively with methanol , and the methanol insoluble substances were re- moved by fil tration through paper. 3) The methanol filtrate was evaporated to dryness, redissolved in distilled water, and extracted with chloroform. 4) The water phase of the chloroform extract ion was concentrated by evapora- tion and chromatographed on a Sephadex G-10 column (Fig. 5a). 5) The ac- tive fractions from the G-10 column were combined, concentrated by evap- orat ion and chromatographed on a Biogel P-2 column (Fig. 5b).
The foot-inhibiting activity is soluble in methanol and insoluble in chloro- form and ether. It is not sedimentable by centrifugation in distilled water at 38,000 rpm for 3 h. These findings indicate that the substance(s) responsible for the inhibition is of relatively low molecular weight. By chromatography on G-10 (Fig. 5a) the foot inhibitor(s) is separated from the head activator (Schaller, 1973); by chromatography on P-2 (Fig. 5b) it is separated from the 'head ' inhibitor (Berking, 1974). The above purification procedure leads to a 25--30 fold enrichment of the foot inhibitor(s) compared to crude ex- tracts with a 50% recovery.
The purified foot inhibitor had no effect on head formation, if gastric regions regenerating a head were incubated in the first 20 h after cutting at concentrat ions corresponding to 1, 3, and 10 BU of foo t inhibitor. Neither the rate of head formation, nor the average number of tentacles regenerated per head was different f rom untreated controls. A negative effect on bud format ion both in regenerating and intact animals was found starting from a concentra t ion of 1 BU.
Distribution of the foot-inhibiting activity in hydra
Preliminary experiments had shown that the foot-inhibiting activity dif- fuses rapidly into the medium, when hydra were cut into pieces. To prevent such a loss, hydra pieces were transferred into methanol immediately after cutting. After sonication of the pieces in methanol and incubation for 6 h at 55°C, the methanol crude extract was filtered through paper and the filtrate was evaporated to dryness, redissolved in water and extracted once with chloroform. To study the distribution of the foot- inhibi tory activity in hy- dra, animals wi thout buds were cut into 4 roughly equal-sized pieces; these pieces were prepared as described above, and the concentra t ion of inhibitor was determined for each region. It was found that an effect corresponding to I BU was obtained with the extract of 5 heads, 5 upper gastric regions, 4
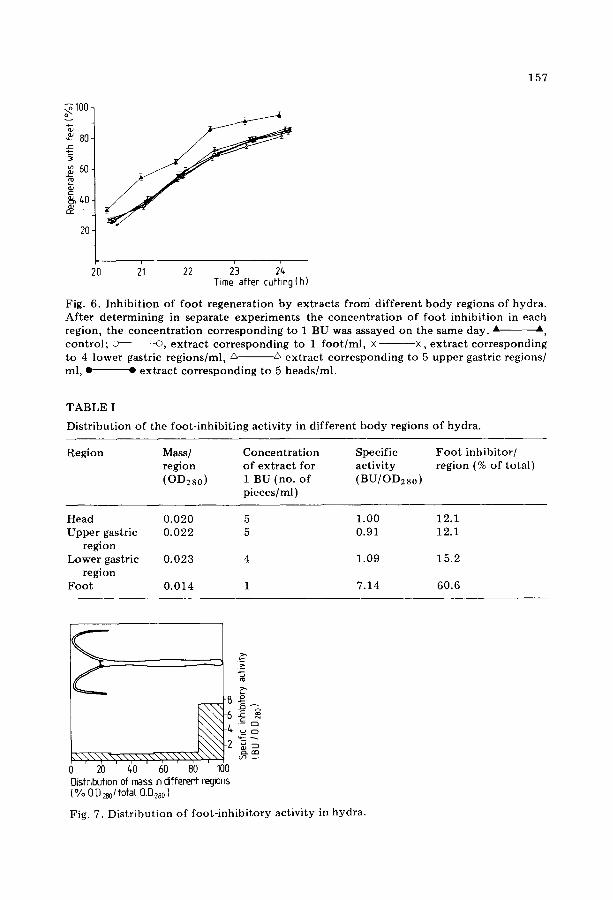
157
o~100-
380 . ~ -
~ 60.
g, t,O
20
20 2'1 2'2 2'3 2'• Time after cuffing (h)
Fig. 6. I n h i b i t i o n of f oo t r egene ra t i on by ex t r ac t s f rom d i f f e ren t b o d y regions of hydra . Af t e r d e t e r m i n i n g in separa te e x p e r i m e n t s the c o n c e n t r a t i o n of f oo t i nh ib i t i on in each region, the c o n c e n t r a t i o n c o r r e s p o n d i n g to 1 BU was assayed on the same day. _A _A, c o n t r o l ; o o, e x t r a c t c o r r e s p o n d i n g to 1 f o o t / m l , × - ×, e x t r a c t c o r r e s p o n d i n g to 4 lower gastric r eg ions /ml , A -~ e x t r a c t c o r r e s p o n d i n g to 5 u p p e r gastric regions/ ml, • ~- ex t r ac t co r r e spond ing to 5 heads /ml .
T A B L E I
D i s t r i bu t ion of the foo t - i nh ib i t i ng act iv i ty in d i f f e ren t b o d y regions of hydra .
Region Mass/ C o n c e n t r a t i o n Specif ic region o f e x t r a c t for ac t iv i ty (OD2s0) 1 BU (no. of ( B U / O D 2 s 0 )
p ieces /ml)
F o o t i n h i b i t o r / region (% of to ta l )
Head 0 .020 5 1.00 Uppe r gastric 0 .022 5 0.91
region Lower gastric 0 .023 4 1.09
region F o o t 0 .014 1 7.14
12.1 12.1
15.2
60.6
_>
-6 - - r - i
-/+ c~ -2 y , ~
\ \ \ \ \
6 ' 2b ' ~ o ' ~ o ' 8 ' o ' 1 d o Oisfribution of mass in different regions (% 0Dz~/t0fat 00.28o)
Fig. 7. D i s t r i bu t ion o f f oo t - i nh i b i t o r y act iv i ty in hydra .

158
lower gastric regions, and 1 foo t per ml, respectively. In Fig. 6 the concentra t ion corresponding to 1 BU was assayed simul-
taneously (same day of experiment) for the different regions. From this it was calculated (Table I, Fig. 7) that most of the foot-inhibiting activity (61%) is concentra ted in the foo t region which represents only 17.7% of the total mass of the animal. The remaining regions contain approximately equal amounts of the foot-inhibiting activity in a 7--8 fold lower concentrat ion.
DISCUSSION
Evidence is presented in this paper that crude extracts of hydra contain a substance or substances probably of low-molecular weight which specifically inhibit foot , but not head regeneration. Inhibition of foot formation was ob- tained at a concentra t ion of crude extract corresponding to 1.5 hydra per ml. Since one hydra or a regenerate on which the crude extract is assayed, has a volume of approximately 1 pl, the effect is obtained with a concen- t rat ion almost 700-fold lower than that present in the assay animal. This re- sult is interpreted as meaning that within the animal the foot inhibitor(s) is present in an inactive, s t ructure-bound form. The same was found for the head activator (Schaller et al., 1973) and for the 'head' inhibitor (Berking, 1974).
The foot inhibitor is present in hydra as a gradient from foot to head. The foot region contains 60% of the total activity in a 7--8 fold higher concentra- tion than the other regions. This result is in accordance with the findings of MacWilliams et al. (1974) that a foot-inhibiting principle is most concen- t ra ted in the basal disk.
The inhibitor described by Berking (1974) principally inhibits bud forma- tion, but it also inhibits head and foot regeneration in hydra. This inhibitor is most concentra ted in the head region (Berking, 1974). This indicates that the two substances are not identical. It is, however, expected that part or all of the inhibitory effect of crude extracts from heads or foot formation is caused by the 'head' inhibitor. Consequently, the gradient in the distribution of the foot inhibitor in the different regions may be steeper than we ob- served.
The extract from 1.5 hydra had the same inhibiting effect as the extract from 1 foot . From this it is concluded that a whole hydra contains possible antagonists at a higher relative ratio than the foot.
The foot inhibitor was purified chemically from the 'head' inhibitor (Ber- king, 1974) and from the head activator (Schaller, 1973). Its distribution in hydra and the specific effect on foot regeneration indicate that the foot in- hibitor is a new morphogen which may be responsible for the inhibition of foo t format ion in hydra.

159
ACKNOWLEDGEMENT
We wish to thank A. Wegmann for excellent technical assistance and the Deutsche Forschungsgemeinschaft for support.
REFERENCES
Berking, S. : Dissertation, 1974. Burnett, A.L.: In: The stability of the Differentiated State, ed. H. Ursprung (Springer
Verlag, Berlin) (1968). Gierer, A. and H. Meinhardt: Kybernetik 12, 30--39 (1972). Lenhoff, H.M. and R. Dubois Brown: Lab. Animals 4, 139--154 (1970). MacWilliams, H.K. and F.C. Kafatos: Science 159, 1246--1247 (1968). MacWilliams, H.K., F.C. Kafatos and W.H. Bossert: Develop. Biol. 23, 380--398 (1970). MacWilliams, H.K., K. Harry and F.C. Kafatos: Amer. Zool. 14,633--645 (1974). Schaller, H.: J. Embryol. Exp. Morph. 29, 27--38 (1973). Schaller, H. and A. Gierer: J. Embryol. Exp. Morph. 29, 39--52 (1973). Webster, G. : Biol. Rev. 46, 1--46 (1971). Wolpert, L. : Curt. Top. Develop. Biol. 6, 183--224 (1971).


















