
Enzymes in Replication
16
Enzymes of DNA Replication Helicase: Unwounds a portion of the DNA Double Helix RNA Primase: Attaches RNA pr imers to the replicating strands. DNA Polymerase delta (ä): Binds to the 5 ! " strand in order to bring nucleotides and create the daughter leading strand. DNA Polymerase epsilon (å): Binds to the " ! 5 strand in order to create discontinuous segments starting from different RNA primers. Exonuclease (DNA Polymerase I): #inds and remo$es the RNA %rimers DNA i!ase: Adds phosphate in the remaining gaps of the phosphate ! sugar bac&bone Nucleases: Remo$e wrong nucleotides from the daughter strand. Wa tson and Crick immediately saw the relationship of the double helix to genetic replication. They proposed that each strand of the chromosome serves as a template to specify a new, complementary DN strand. template is a pattern for making something! DN acts as a template because each strand specifies the newdaughter strand by base‐ pairing. This template feature makes DN replication semiconservative " after replication, each daughter chromosome has one strand of newly synthesi#ed DN and one strand of DN from the parental chromosome. $ee %igure & .
-
Upload
rawatpooran05 -
Category
Documents
-
view
11 -
download
0
description
Enzymes involved in DNA replication
Transcript of Enzymes in Replication

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 1/16
Enzymes of DNA ReplicationHelicase: Unwounds a portion of the DNA Double Helix
RNA Primase: Attaches RNA primers to the replicating strands.
DNA Polymerase delta (ä): Binds to the 5 ! " strand in order to bring nucleotides and create thedaughter leading strand.
DNA Polymerase epsilon (å): Binds to the " ! 5 strand in order to create discontinuous segmentsstarting from different RNA primers.
Exonuclease (DNA Polymerase I): #inds and remo$es the RNA %rimers
DNA i!ase: Adds phosphate in the remaining gaps of the phosphate ! sugar bac&bone
Nucleases: Remo$e wrong nucleotides from the daughter strand.
Watson and Crick immediately saw the relationship of the double helix to genetic replication.
They proposed that each strand of the chromosome serves as a template to specify a new,complementary DN strand. template is a pattern for making something! DN acts as a
template because each strand specifies the newdaughter strand by base‐ pairing. This template
feature makes DN replication semiconservative" after replication, each daughter
chromosome has one strand of newly synthesi#ed DN and one strand of DN from the
parental chromosome. $ee %igure& .

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 2/16
%igure &
DNA polymerases
These en#ymes copy DN se'uences by using one strand as a template. The reaction cataly#ed by DN polymerases is the addition of deoxyribonucleotides to a DN chain by using dNT(s as
substrates, as shown in %igure ).

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 3/16
%igure )
ll DN polymerases re'uire a template strand, which is copied. DN polymerases also re'uire
a primer, which is complementary to the template. The reaction of DN polymerases is thus
better understood as the addition of nucleotides to a primer to make a se'uence complementary
to a template. The re'uirement for template and primer are exactly what would be expected ofa replication en#yme. *ecause DN is the information store of the cell, any ability of DN
polymerases to make DN se'uences from nothing would lead to the degradation of the cell+s
information copy.
ore than one DN polymerase exists in each cell. The key distinction among the en#yme formsis their processitivity -how long a chain they synthesi#e before falling off the template. DN
polymerase used in replication is more processive than a repair en#yme. The replication en#ymeneeds to make a long enough chain to replicate the entire chromosome. The repair en#yme needs
only to make a long enough strand to replace the damaged se'uences in the chromosome. The
best‐studied bacterium, E. coli, has three DN polymerase types.
DNA polymerase I (Pol I) is primarily a repair en#yme, although it also has a function in
replication. bout // (ol 0 molecules exist in a single bacterium. DN polymerase 0 onlymakes an average of )/ phosphodiester bonds before dissociating from the template. These
properties make good sense for an en#yme that is going to replace damaged DN. Damage
occurs at separate locations so the large number of (ol 0 molecules means that a repair en#yme is
always close at hand.DN polymerase 0 has nucleolytic 1depolymeri#ing2 activities, which are an intimate part of their
function. The 34 to 54 exonuclease activity removes base‐ paired se'uences ahead of the
polymeri#ing activity. During replication, this can remove primers ahead of the polymeri#ingfunction of the polymerase. $ee %igure 5 .

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 4/16
%igure 5
nother intimate function of DN polymerase 0 1and of the other forms of DN polymerasefound in 6. coli2 is the 54 to 34 exonuclease activity. This activity can de‐ polymeri#e DN
starting from the newly synthesi#ed end. 0magining why DN polymerase would have an
activity that opposes the action of the en#yme is a little difficult. The 54 to 34 exonuclease activity
serves an editing function to ensure the fidelity of replication. $uppose DN polymerase were tomake a mistake and add a T opposite a 7 in the template strand. When the en#yme begins the
next step of polymeri#ation, the T is not properly paired with the template. The 54 to 34
exonucleolytic activity of DN polymerase then removes the unpaired nucleotide, releasing
T(, until a properly paired stretch is detected. Then polymeri#ation can resume. This cyclecosts two high‐energy phosphate bonds because TT( is converted to T(. While this may seem
wasteful of energy, the editing process does keep the information store of the cell intact, as
shown in %igure .
DNA polymerase II is a speciali#ed repair en#yme. 8ike (ol 0, a large number of (ol 00molecules reside in the cell 1about &//2. The en#yme is more processive than (ol 0. (ol 00 has the
same editing 154 to 342 activity as (ol 0, but not the 34 to 54 exonuclease activity.
The actual replication en#yme in 6. coli is DNA polymerase III. 0ts properties contrast with (ol
0 and (ol 00 in several respects. (ol 000 is much more processive than the other en#ymes, makingabout 3//,/// phosphodiester bonds on the average. 0n other words, it is about 3,/// times more
processive than (ol 0 and 3/ times more processive than (ol 00. (ol 000 is a multisubunit en#yme.
0t lacks a 34 to 54 exonucleolytic activity, although a subunit of the en#yme carries out the editing154 to 342 function during replication. %inally, only about &/ molecules of (ol 000 reside in each
cell. This remains consistent with the function of (ol 000 in replication, because the chromosome
only needs to be copied once per generation. Therefore, the cell only re'uires a few molecules ofthe en#yme. (ol 000 synthesi#es DN at least a hundred times more rapidly than the other
polymerases. 0t can synthesi#e half of the bacterial chromosome in a little more than )/ minutes,
which is the fastest that the bacterium can replicate.

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 5/16
%igure
Chromosomal replication
The process of chromosomal replication in bacteria is complex. *acterial chromosomes are
double‐stranded DN and almost always circular. DN replication starts at a specific se'uence,
the origin, on the chromosome and proceeds in two directions towards another specific region,
the terminus, as shown in %igure 3 .

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 6/16
%igure 3
Initiation
t an origin, the replication process first involves DN strand opening so that each strand of theDN molecule is available as a template. 0nitiation is the rate‐limiting step for replication of the
chromosome. 8ike other metabolic pathways, the control of replication is exerted at the first
committed step.
0nitiation se'uences contain a set of repeated se'uences, which bind the essential initiator protein, Dna. The Dna protein opens the helix to make a short region of separated strands.
Then a speciali#ed single‐strand binding protein binds to the DN strands to keep them apart.
This process makes a template, but replication can+t happen because no primer yet exists. $ee
%igure 9.
%igure 9

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 7/16
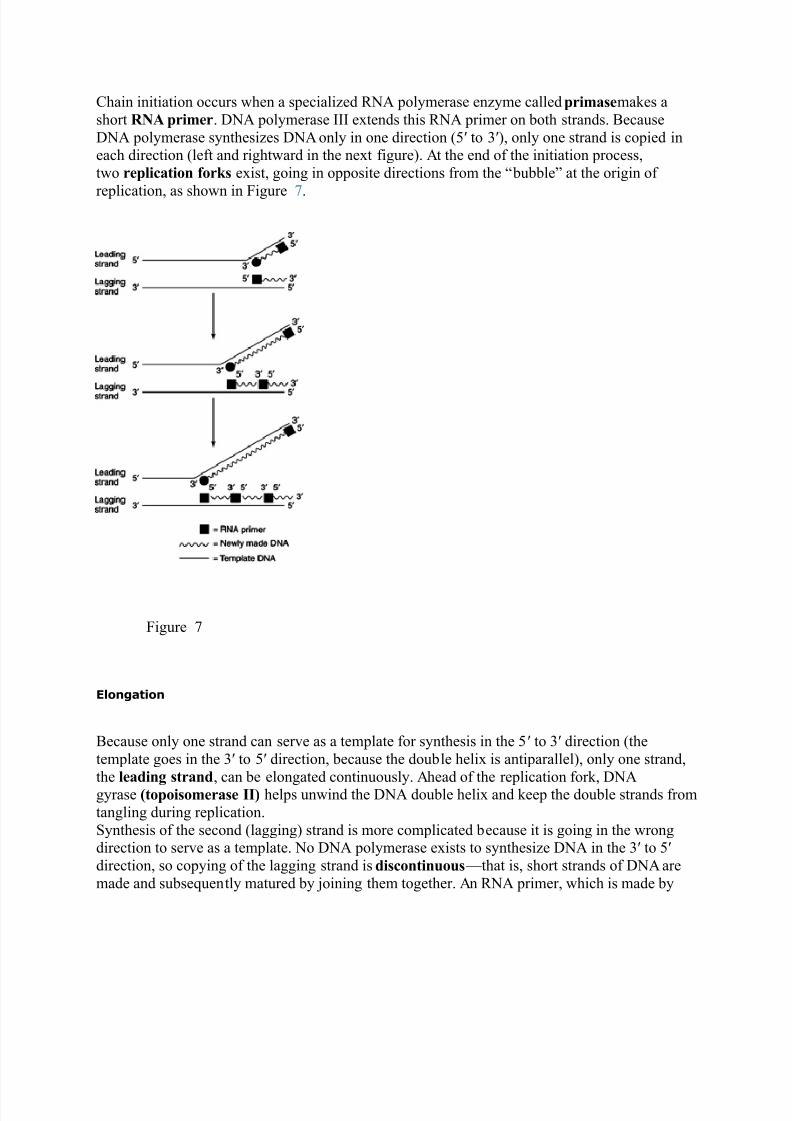
Chain initiation occurs when a speciali#ed :N polymerase en#yme called primasemakes a
short RNA primer. DN polymerase 000 extends this :N primer on both strands. *ecause
DN polymerase synthesi#es DN only in one direction 134 to 542, only one strand is copied ineach direction 1left and rightward in the next figure2. t the end of the initiation process,
two replication forks exist, going in opposite directions from the ;bubble< at the origin of
replication, as shown in %igure =.
%igure =
Elongation
*ecause only one strand can serve as a template for synthesis in the 34 to 54 direction 1the
template goes in the 54 to 34 direction, because the double helix is antiparallel2, only one strand,
the leading strand, can be elongated continuously. head of the replication fork, DNgyrase (topoisomerase II) helps unwind the DN double helix and keep the double strands from
tangling during replication.
$ynthesis of the second 1lagging2 strand is more complicated because it is going in the wrongdirection to serve as a template. No DN polymerase exists to synthesi#e DN in the 54 to 34
direction, so copying of the lagging strand is discontinuous -that is, short strands of DN are
made and subse'uently matured by >oining them together. n :N primer, which is made by

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 8/16
primase, initiates each of these small pieces of DN. Then DN polymerase 000 elongates the
primer until it butts up against the 34 end of the next primer molecule.
DN polymerase 0 then uses its polymeri#ing and 34 to 54 exonuclease activities to remove the:N primer and fill in this se'uence with DN. *ecause (ol 0 is not very processive, it falls off
the lagging strand after a relatively short‐length synthesis. DN polymerases can+t seal up the
nicks that result from the replacement of :N primers with DN. 0nstead, anotheren#yme, DNA ligase, seals off the nicks by using high energy phosphodiester bonds in T( or ND to >oin a free 54 hydroxyl with an ad>acent 34 phosphate.
multien#yme complex simultaneously carries out both leading and lagging strand replication.
?ou can see the best model of the process in the next figure! the lagging strand may curl aroundso it presents the correct face to the en#yme. The two replication forks proceed around the
chromosome, until they meet at the terminus. Termination is poorly defined biochemically, but it
is known to re'uire some form of DN gyrase activity. $ee %igure @.
%igure @

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 9/16
atson and Crick immediately saw the relationship of the double helix to genetic replication. They
proposed that each strand of the chromosome serves as a template to specify a new,
complementary DN strand. template is a pattern for making something! DN acts as a
template because each strand specifies the newdaughter strand by base‐ pairing. This template
feature makes DN replication semiconservative" after replication, each daughter
chromosome has one strand of newly synthesi#ed DN and one strand of DN from the
parental chromosome. $ee %igure& .
%igure &

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 10/16
DNA polymerases
These en#ymes copy DN se'uences by using one strand as a template. The reaction cataly#ed
by DN polymerases is the addition of deoxyribonucleotides to a DN chain by using dNT(s as
substrates, as shown in %igure ).
%igure )
ll DN polymerases re'uire a template strand, which is copied. DN polymerases also re'uirea primer, which is complementary to the template. The reaction of DN polymerases is thus
better understood as the addition of nucleotides to a primer to make a se'uence complementary
to a template. The re'uirement for template and primer are exactly what would be expected ofa replication en#yme. *ecause DN is the information store of the cell, any ability of DN
polymerases to make DN se'uences from nothing would lead to the degradation of the cell+s
information copy.ore than one DN polymerase exists in each cell. The key distinction among the en#yme forms
is their processitivity -how long a chain they synthesi#e before falling off the template. DN
polymerase used in replication is more processive than a repair en#yme. The replication en#ymeneeds to make a long enough chain to replicate the entire chromosome. The repair en#yme needsonly to make a long enough strand to replace the damaged se'uences in the chromosome. The
best‐studied bacterium, E. coli, has three DN polymerase types.
DNA polymerase I (Pol I) is primarily a repair en#yme, although it also has a function in
replication. bout // (ol 0 molecules exist in a single bacterium. DN polymerase 0 onlymakes an average of )/ phosphodiester bonds before dissociating from the template. These
properties make good sense for an en#yme that is going to replace damaged DN. Damage

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 11/16
occurs at separate locations so the large number of (ol 0 molecules means that a repair en#yme is
always close at hand.
DN polymerase 0 has nucleolytic 1depolymeri#ing2 activities, which are an intimate part of their function. The 34 to 54 exonuclease activity removes base‐ paired se'uences ahead of the
polymeri#ing activity. During replication, this can remove primers ahead of the polymeri#ing
function of the polymerase. $ee %igure 5 .
%igure 5
nother intimate function of DN polymerase 0 1and of the other forms of DN polymerasefound in 6. coli2 is the 54 to 34 exonuclease activity. This activity can de‐ polymeri#e DN
starting from the newly synthesi#ed end. 0magining why DN polymerase would have an
activity that opposes the action of the en#yme is a little difficult. The 54 to 34 exonuclease activityserves an editing function to ensure the fidelity of replication. $uppose DN polymerase were to
make a mistake and add a T opposite a 7 in the template strand. When the en#yme begins the
next step of polymeri#ation, the T is not properly paired with the template. The 54 to 34exonucleolytic activity of DN polymerase then removes the unpaired nucleotide, releasing
T(, until a properly paired stretch is detected. Then polymeri#ation can resume. This cycle
costs two high‐energy phosphate bonds because TT( is converted to T(. While this may seem
wasteful of energy, the editing process does keep the information store of the cell intact, as
shown in %igure .DNA polymerase II is a speciali#ed repair en#yme. 8ike (ol 0, a large number of (ol 00
molecules reside in the cell 1about &//2. The en#yme is more processive than (ol 0. (ol 00 has the
same editing 154 to 342 activity as (ol 0, but not the 34 to 54 exonuclease activity.The actual replication en#yme in 6. coli is DNA polymerase III. 0ts properties contrast with (ol
0 and (ol 00 in several respects. (ol 000 is much more processive than the other en#ymes, making
about 3//,/// phosphodiester bonds on the average. 0n other words, it is about 3,/// times more processive than (ol 0 and 3/ times more processive than (ol 00. (ol 000 is a multisubunit en#yme.

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 12/16
0t lacks a 34 to 54 exonucleolytic activity, although a subunit of the en#yme carries out the editing
154 to 342 function during replication. %inally, only about &/ molecules of (ol 000 reside in each
cell. This remains consistent with the function of (ol 000 in replication, because the chromosomeonly needs to be copied once per generation. Therefore, the cell only re'uires a few molecules of
the en#yme. (ol 000 synthesi#es DN at least a hundred times more rapidly than the other
polymerases. 0t can synthesi#e half of the bacterial chromosome in a little more than )/ minutes,which is the fastest that the bacterium can replicate.
%igure
Chromosomal replication
The process of chromosomal replication in bacteria is complex. *acterial chromosomes are
double‐stranded DN and almost always circular. DN replication starts at a specific se'uence,
the origin, on the chromosome and proceeds in two directions towards another specific region,the terminus, as shown in %igure 3 .

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 13/16
%igure 3
Initiation
t an origin, the replication process first involves DN strand opening so that each strand of theDN molecule is available as a template. 0nitiation is the rate‐limiting step for replication of the
chromosome. 8ike other metabolic pathways, the control of replication is exerted at the first
committed step.
0nitiation se'uences contain a set of repeated se'uences, which bind the essential initiator protein, Dna. The Dna protein opens the helix to make a short region of separated strands.
Then a speciali#ed single‐strand binding protein binds to the DN strands to keep them apart.
This process makes a template, but replication can+t happen because no primer yet exists. $ee
%igure 9.
%igure 9
7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 14/16
Chain initiation occurs when a speciali#ed :N polymerase en#yme called primasemakes a
short RNA primer. DN polymerase 000 extends this :N primer on both strands. *ecause
DN polymerase synthesi#es DN only in one direction 134 to 542, only one strand is copied ineach direction 1left and rightward in the next figure2. t the end of the initiation process,
two replication forks exist, going in opposite directions from the ;bubble< at the origin of
replication, as shown in %igure =.
%igure =
Elongation
*ecause only one strand can serve as a template for synthesis in the 34 to 54 direction 1the
template goes in the 54 to 34 direction, because the double helix is antiparallel2, only one strand,
the leading strand, can be elongated continuously. head of the replication fork, DNgyrase (topoisomerase II) helps unwind the DN double helix and keep the double strands from
tangling during replication.
$ynthesis of the second 1lagging2 strand is more complicated because it is going in the wrongdirection to serve as a template. No DN polymerase exists to synthesi#e DN in the 54 to 34
direction, so copying of the lagging strand is discontinuous -that is, short strands of DN are
made and subse'uently matured by >oining them together. n :N primer, which is made by

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 15/16
primase, initiates each of these small pieces of DN. Then DN polymerase 000 elongates the
primer until it butts up against the 34 end of the next primer molecule.
DN polymerase 0 then uses its polymeri#ing and 34 to 54 exonuclease activities to remove the:N primer and fill in this se'uence with DN. *ecause (ol 0 is not very processive, it falls off
the lagging strand after a relatively short‐length synthesis. DN polymerases can+t seal up the
nicks that result from the replacement of :N primers with DN. 0nstead, anotheren#yme, DNA ligase, seals off the nicks by using high energy phosphodiester bonds in T( or ND to >oin a free 54 hydroxyl with an ad>acent 34 phosphate.
multien#yme complex simultaneously carries out both leading and lagging strand replication.
?ou can see the best model of the process in the next figure! the lagging strand may curl aroundso it presents the correct face to the en#yme. The two replication forks proceed around the
chromosome, until they meet at the terminus. Termination is poorly defined biochemically, but it
is known to re'uire some form of DN gyrase activity. $ee %igure @.
%igure @

7/18/2019 Enzymes in Replication
http://slidepdf.com/reader/full/enzymes-in-replication 16/16


















