Dorsal-ventral patterning in the mammalian telencephalon
7
Dorsal-ventral patterning in the mammalian telencephalon Kenneth Campbell The telencephalon is the most diverse region of the brain with respect to both morphology and neuronal subtypes. This fact makes the task of unraveling the mechanisms underlying the development of this brain region rather daunting. Recent attempts to subdivide the embryonic telencephalon into distinct progenitor domains along the dorsal–ventral axis have provided an important framework on which to begin this process. These progenitor domains are defined by the restricted expression of transcriptional regulators and are proposed to give rise to specific subtypes of neurons. Work over recent years has provided important insights into the establishment and maintenance of these progenitor domains in the developing telencephalon. Addresses Division of Developmental Biology, Children’s Hospital Research Foundation, 3333 Burnet Avenue, Cincinnati, OH 45229, USA e-mail: [email protected] Current Opinion in Neurobiology 2003, 13:50–56 This review comes from a themed issue on Development Edited by Magdalena Go ¨ tz and Samuel L Pfaff 0959-4388/03/$ – see front matter ß 2003 Elsevier Science Ltd. All rights reserved. DOI 10.1016/S0959-4388(03)00009-6 Abbreviations BMP bone morphogenetic protein CGE caudal ganglionic eminence DV dorsal–ventral LGE lateral ganglionic eminence MGE medial ganglionic eminence RA retinoic acid SHH sonic hedgehog SVZ subventricular zone VZ ventricular zone Introduction The vertebrate telencephalon derives from the most anterior portion of the developing CNS. This brain region is instrumental in organizing intellectual functions and voluntary movements. Two of the principal telencephalic structures that regulate these processes are the cerebral cortex and the basal ganglia (including both the striatum and the globus pallidus). Although the cortex is located in the dorsal telencephalon, its neuronal constituents derive from both the dorsal and ventral halves of the embryonic telencephalon: whereas cortical projection neurons arise in the dorsal telencephalon from a thin neuroepithelium [1], called the pallium (Figure 1), most of the interneur- ons that populate cortical structures originate in the ventral telencephalon (i.e. subpallium) and subsequently migrate tangentially into the cortex [2,3,4 ]. Neurons of the basal ganglia, by contrast, derive exclu- sively from the ganglionic eminences of the ventral tele- ncephalon (Figure 1). At rostral levels, the ganglionic eminences form separate medial (MGE) and lateral (LGE) elevations, whereas at caudal levels there is a single eminence, known as the caudal ganglionic emi- nence (CGE). The MGE is a very heterogeneous struc- ture, giving rise to projection neurons of the pallidum and basal forebrain [5,6], as well as to interneurons that populate the ventral telencephalon and cortical structures [3,4 ,6,7]. The LGE produces the projection neurons of the striatum [4 ,5,6,8] and interneurons that migrate rostrally to the olfactory bulb [4 ,9]. Although the LGE and CGE share many similarities, a recent study has shown that the CGE gives rise to both projection neurons and interneurons that are distinct from those produced in either the LGE or the MGE [10 ]. Thus, the neuronal diversity present in many telencephalic structures arises through the generation of neuronal sub- types from localized subregions of the embryonic tele- ncephalon and the subsequent distribution of these neurons through radial or tangential migration. An important issue in CNS development is what regulates the regional production of specific neuronal subtypes. Work on the developing spinal cord has shown that distinct neuronal subtypes are generated from discrete progenitor domains located along the dorsal–ventral (DV) axis [11,12]. These progenitor domains are characterized by the restricted expression of transcription factor genes and seem to be established by the coordinated signaling of bone morphogenetic proteins (BMPs) and sonic hedge- hog (SHH) from the dorsal midline and ventral midline, respectively [11,12]. Gene expression studies have estab- lished that similar progenitor domains can be identified along the DV axis of the developing telencephalon. This review will cover recent work on the molecular mechan- isms underlying the establishment and maintenance of these telencephalic progenitor domains. Regional subdivisions along the dorsal– ventral axis of the telencephalon The morphological boundaries in the embryonic telence- phalon do not always correlate with the progenitor domains defined by gene expression. Notably, the bound- ary between the dorsal and ventral halves of the tele- ncephalon (i.e. the pallio-subpallial boundary) does not lie at the angle of the pallium and LGE but is slightly more ventral in the dorsal-most portion of the LGE 50 Current Opinion in Neurobiology 2003, 13:50–56 www.current-opinion.com
-
Upload
kenneth-campbell -
Category
Documents
-
view
218 -
download
1
Transcript of Dorsal-ventral patterning in the mammalian telencephalon

Dorsal-ventral patterning in the mammalian telencephalonKenneth Campbell
The telencephalon is the most diverse region of the brain with
respect to both morphology and neuronal subtypes. This fact
makes the task of unraveling the mechanisms underlying the
development of this brain region rather daunting. Recent
attempts to subdivide the embryonic telencephalon into distinct
progenitor domains along the dorsal–ventral axis have provided
an important framework on which to begin this process. These
progenitor domains are defined by the restricted expression of
transcriptional regulators and are proposed to give rise to
specific subtypes of neurons. Work over recent years has
provided important insights into the establishment and
maintenance of these progenitor domains in the developing
telencephalon.
AddressesDivision of Developmental Biology, Children’s Hospital Research
Foundation, 3333 Burnet Avenue, Cincinnati, OH 45229, USA
e-mail: [email protected]
Current Opinion in Neurobiology 2003, 13:50–56
This review comes from a themed issue on
Development
Edited by Magdalena Gotz and Samuel L Pfaff
0959-4388/03/$ – see front matter
� 2003 Elsevier Science Ltd. All rights reserved.
DOI 10.1016/S0959-4388(03)00009-6
AbbreviationsBMP bone morphogenetic proteinCGE caudal ganglionic eminence
DV dorsal–ventral
LGE lateral ganglionic eminence
MGE medial ganglionic eminence
RA retinoic acid
SHH sonic hedgehog
SVZ subventricular zone
VZ ventricular zone
IntroductionThe vertebrate telencephalon derives from the most
anterior portion of the developing CNS. This brain region
is instrumental in organizing intellectual functions and
voluntary movements. Two of the principal telencephalic
structures that regulate these processes are the cerebral
cortex and the basal ganglia (including both the striatum
and the globus pallidus). Although the cortex is located in
the dorsal telencephalon, its neuronal constituents derive
from both the dorsal and ventral halves of the embryonic
telencephalon: whereas cortical projection neurons arise
in the dorsal telencephalon from a thin neuroepithelium
[1], called the pallium (Figure 1), most of the interneur-
ons that populate cortical structures originate in the
ventral telencephalon (i.e. subpallium) and subsequently
migrate tangentially into the cortex [2,3,4��].
Neurons of the basal ganglia, by contrast, derive exclu-
sively from the ganglionic eminences of the ventral tele-
ncephalon (Figure 1). At rostral levels, the ganglionic
eminences form separate medial (MGE) and lateral
(LGE) elevations, whereas at caudal levels there is a
single eminence, known as the caudal ganglionic emi-
nence (CGE). The MGE is a very heterogeneous struc-
ture, giving rise to projection neurons of the pallidum and
basal forebrain [5,6], as well as to interneurons that
populate the ventral telencephalon and cortical structures
[3,4��,6,7]. The LGE produces the projection neurons of
the striatum [4��,5,6,8] and interneurons that migrate
rostrally to the olfactory bulb [4��,9]. Although the
LGE and CGE share many similarities, a recent study
has shown that the CGE gives rise to both projection
neurons and interneurons that are distinct from those
produced in either the LGE or the MGE [10�]. Thus,
the neuronal diversity present in many telencephalic
structures arises through the generation of neuronal sub-
types from localized subregions of the embryonic tele-
ncephalon and the subsequent distribution of these
neurons through radial or tangential migration.
An important issue in CNS development is what regulates
the regional production of specific neuronal subtypes.
Work on the developing spinal cord has shown that
distinct neuronal subtypes are generated from discrete
progenitor domains located along the dorsal–ventral (DV)
axis [11,12]. These progenitor domains are characterized
by the restricted expression of transcription factor genes
and seem to be established by the coordinated signaling
of bone morphogenetic proteins (BMPs) and sonic hedge-
hog (SHH) from the dorsal midline and ventral midline,
respectively [11,12]. Gene expression studies have estab-
lished that similar progenitor domains can be identified
along the DV axis of the developing telencephalon. This
review will cover recent work on the molecular mechan-
isms underlying the establishment and maintenance of
these telencephalic progenitor domains.
Regional subdivisions along the dorsal–ventral axis of the telencephalonThe morphological boundaries in the embryonic telence-
phalon do not always correlate with the progenitor
domains defined by gene expression. Notably, the bound-
ary between the dorsal and ventral halves of the tele-
ncephalon (i.e. the pallio-subpallial boundary) does not
lie at the angle of the pallium and LGE but is slightly
more ventral in the dorsal-most portion of the LGE
50
Current Opinion in Neurobiology 2003, 13:50–56 www.current-opinion.com

(Figure 1). This is evident from the restricted expression
of numerous transcription factors, including Pax6 on the
pallial side and Gsh2 on the subpallial side [13,14]. In the
subventricular zone (SVZ) and the mantle regions, the
pallio-subpallial boundary is marked by a stream of Pax6-
positive cells originating from the region where Pax6 and
Gsh2 meet in the ventricular zone (VZ) and continuing
down to the pial surface (Figure 1; [13–15]).
Recent studies [13,15–17] have suggested that the pal-
lium can been subdivided into separate ventral, lateral,
dorsal and medial compartments (Figure 1) that give rise
to projection neurons of the clastroamygdaloid complex,
lateral cortex, neocortex and hippocampus, respectively.
The ventral pallium was initially defined both by its
expression of Pax6 in the VZ and Tbr1 in the mantle
region and by its lack of Emx1 expression [15,16]. Recent
work has shown that the VZ of the ventral pallium
uniquely expresses the homeobox gene Dbx [13] and
the putative Wnt antagonist SFRP2 [18].
Unlike the ventral pallium, the specific boundaries of the
other pallial domains are not as easily marked by discrete
gene expression. In these regions, graded gene expression
is more common. For example, Emx1, Emx2 [19,20] and
Lhx2 [21��] all show high expression in the medial pal-
lium, with a progressive reduction of expression in more
ventral regions. Conversely, Pax6 and Tbr2 show the
opposite profile with their highest expression in the VZ
of the ventral and lateral pallium [13,14,17].
In the subpallium, many genes are expressed in both the
LGE and MGE, including Gsh2 in the VZ [22] and
members of the Dlx gene family (i.e. Dlx1, Dlx2, Dlx5and Dlx6) in the VZ and the SVZ [23]. The ventromedial
telencephalon, including the MGE, steptum and the
preoptic area, expresses Nkx2.1 [24,25]. Given the con-
siderable heterogeneity of the ventromedial telencepha-
lon with respect to the oligodendrocytes [26�,27�] and the
different interneurons that it generates [3,7], it seems
likely that future studies will identify separate progenitor
subdomains in this telencephalic region as well.
Recently, the LGE has been divided into a small dorsal
domain (dorsal LGE) and a larger ventral domain (ventral
LGE) (Figure 1; [13]). The dorsal LGE expresses the Ets
gene Er81 [13]. Although this gene is expressed in the
VZ, protein expression is observed only in the SVZ of the
dorsal LGE. This Er81-positive SVZ domain has been
suggested to give rise to interneurons that migrate in the
rostral migratory stream to populate the olfactory bulb
[28�]. Progenitors in the SVZ of the ventral LGE uniquely
express the LIM homeobox protein Islet-1 and generate
striatal projection neurons [14,28�].
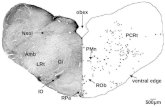
Figure 1
MGE
dLGE
vLGE
VP
LP
DPMP
MGE
LGE
pallium
VZSVZ
Pallio-subpallialboundary
Dorsal
Ventral
Current Opinion in Neurobiology
Coronal hemisections of the mouse telencephalon at embryonic day 12.5, showing morphologically defined structures and the progenitor
subdomains identified by restricted expression of transcription factor genes. The ventricular zone (VZ) extends along the whole DV axis and contains
precursor cells. The subventricular zone (SVZ, indicated by broken blue lines) also contains precursor cells, which is a unique feature of the
telencephalon. Note the placement of the DV boundary (i.e. pallio-subpallial boundary) in the dorsal portion of the LGE. Broken red lines indicate the
approximate boundaries between distinct telencephalic progenitor domains (see text for details). dLGE, dorsal LGE; DP, dorsal pallium; LP, lateralpallium; MP, medial pallium; vLGE, ventral LGE; VP, ventral pallium.
Dorsal-ventral patterning in the mammalian telencephalon Campbell 51
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:50–56

Dorsal telencephalic patterningThe roof plate is thought to have an essential role in
patterning the dorsal spinal cord, primarily through the
expression of BMPs [11,12]. A recent study has assessed
the role of the roof plate in dorsal telencephalic patterning
[21��]. In this study, genetic ablation of the telencephalic
roof plate resulted in a severely reduced expression of
Lhx2 and a severe reduction in cortical size. A require-
ment for BMPs was implied from these results because
the dorsal midline of the telencephalon expresses numer-
ous BMPs [29]. Indeed, explant studies support a role for
BMPs in dorsal telencephalic development [29] and, in
particular, in the expression of Lhx2 [21��].
The requirement for BMP signaling in dorsal telence-
phalic patterning has recently been assessed by condi-
tionally inactivating the BMP receptor 1a specifically in
the mouse telencephalon [30�]. Although the choroid
plexus (a dorsal midline derivative) does not form prop-
erly in these conditional mutants, the telencephalon
seems to be patterned correctly along the DV axis.
Compensation by the BMP receptor 1b in the dorsal-
most telencephalon (i.e. medial and dorsal pallium) was
not observed, indicating that BMP signaling is required
absolutely for midline development but other factors can
organize dorsal patterning in the telencephalon.
The dorsal midline of the telencephalon is also known to
express Wnt molecules [31], and Wnt signaling is
required for hippocampal development (i.e. medial pal-
lium) [32,33]. Furthermore, certain Wnts are also
expressed throughout the pallial VZ at early stages of
telencephalic development [31,34] and thus may have a
broader role in patterning the dorsal telencephalon. Inter-
estingly, ablation of the roof plate in the telencephalon
also resulted in the loss of the Wnt2b expression from the
dorsal midline [21��], suggesting that the observed phe-
notype may result from the combined loss of BMP and
Wnt signaling.
A recent study has identified a telencephalic enhancer of
the Emx2 gene that is expressed at its highest levels in the
medial and dorsal pallium (similar to Lhx2) [35�]. This
enhancer contains requisite binding sites for both Smad
and Tcf proteins, which are transcriptional mediators of
BMP and Wnt signaling, respectively. In addition, both
BMPs and Wnts can activate this DNA enhancer in
midbrain explants. Thus, it may be that BMP signaling
from the dorsal midline is functionally redundant with
Wnt signaling in specifying certain aspects of dorsal
telencephalic fates.
The downstream effects of BMP and/or Wnt signaling in
the dorsal telencephalon ultimately involves the combi-
natorial actions of transcription factors such as Emx1,
Emx2 and Lhx2, which function to specify and to expand
the medial and dorsal pallium. This is most evident in the
Lhx2 mutants, which lack most of the hippocampus and
neocortex [21��,36,37]. The Emx mutants do not show as
marked phenotypes in the dorsal telencephalon as do the
Lhx2 mutants, although hippocampal development is
impaired in Emx2 mutants [38,39]. This is not thought
to result from altered specification, however, but rather
from reduced growth and maturation [40].
Another transcription factor that is crucially involved in
dorsal patterning is the zinc-finger gene Gli3[41,42,43��]. Loss of Gli3 function results in a loss of
Emx gene expression as well as the ectopic expression of
certain genes characteristic of ventral telencephalic pro-
genitors, such as Gsh2. With respect to the latter pheno-
type, a recent study has shown that Gli3 is required to
antagonize the ventralizing signal SHH in the dorsal
telencephalon [43��]. It is currently unclear whether
Gli3 lies upstream, downstream or in a parallel pathway
with BMP and Wnt signaling.
Ventral telencephalic patterningThe secreted glycoprotein SHH is required for ventral
development at caudal levels of the developing CNS
(reviewed in [11,12]), and many studies have shown that
SHH is also involved in patterning the ventral telence-
phalon [44–48]. Results obtained from the analysis of
Shh-null mice [49] also support a role for SHH in tele-
ncephalic patterning, but it does not seem to be required
for all aspects of ventral telencephalic development.
Although these mutants lack any sign of MGE develop-
ment, such as expression of Nkx2.1 [43��,50], many of
them express genes normally found in both the MGE and
LGE, such as Gsh2 and Dlx2, as well as the LGE-specific
cellular retinol-binding protein 1 ([43��]; and H Toresson,
K Campbell, unpublished data). Thus, SHH is required
for ventromedial (i.e. MGE) telencephalic development,
but aspects of ventrolateral (i.e. LGE) telencephalic
patterning can occur in its absence. Although it is cur-
rently unclear whether dorsal LGE characteristics are
present in the Shh-null mice, Islet-1 cells (i.e. ventral
LGE) can be found (H Toresson, K Campbell, unpub-
lished data).
As mentioned above, a role of Gli3 is to antagonize SHH
signaling. Recent work has shown that this is a mutually
antagonistic interaction with SHH repressing Gli3 func-
tion both in the ventral spinal cord [51] and in the tele-
ncephalon [43��]. In the spinal cord of Shh�/�;Gli3�/�
mutants, ventral patterning is greatly improved over that
observed in single Shh mutants, however, the ventral-
most cell types (i.e. the floor plate and V3 neurons) do not
form [51]. This may be different in the telencephalon,
because Shh�/�;Gli3�/� and even Shh�/�;Gli3þ/� mutants
show Nkx2.1 expression and a morphologically distinct
MGE [43��]. It remains unclear, however, whether
the ventral-most telencephalic regions (e.g. preoptic area)
are rescued in the Shh�/�;Gli3�/� mutants. Notably,
52 Development
Current Opinion in Neurobiology 2003, 13:50–56 www.current-opinion.com

patterning in the dorsal telencephalon of Shh;Gli3 double
mutants is improved over that in Gli3 single mutants,
indicating that excessive SHH signaling is involved in the
manifestation of the Gli3 mutant phenotype.
It is currently unknown what patterns the telencephalon
in the absence of SHH and Gli3. Other hedgehog proteins
are not likely candidates because double mutants of
Smoothened (an essential component of SHH signaling
[52]) and Gli3 also show improved DV patterning similar
to that observed in the Shh�/�;Gli3�/� embryos [43��]. As
suggested by Rallu et al. [43��] other (potentially novel)
signaling pathways may direct DV patterning in the
absence of SHH and Gli3. If this is true, it will be
interesting to determine whether these pathways nor-
mally have a role in DV patterning or if they do so only as
a compensatory mechanism.
The source of SHH for telencephalic DV patterning
remains unclear. Unlike the developing spinal cord, the
telencephalon lacks both a floor plate and the underlying
mesoderm expressing SHH [44]. In addition, the expres-
sion of Shh in the ventral telencephalon is an unlikely
source for patterning because it coincides with, rather than
precedes, ventral specification (e.g. see [24]). Previously,
the ventral midline of the diencephalon was suggested to
be the source of SHH for telencephalic ventralization [44];
however, a more recent study has proposed that SHH
from the node controls this process [46]. If this is true,
then the initiation of ventral patterning in the telence-
phalon occurs much earlier than was previously thought.
Patterning the intermediate telencephalonThe intermediate region of the telencephalon includes
the LGE, as well as the ventral and lateral pallium, and is
divided by the pallio-subpallial boundary. Although sig-
nals from both the dorsal and the ventral aspects of the
telencephalon probably have an impact on the patterning
of this region, it is also likely that distinct lateral signals are
involved (Figure 2). Indeed, these putative lateral signals
may contribute to the expression of ventrolateral genes in
the ventral midline of the Shh mutant (see above).
The expression profiles of certain genes in the intermedi-
ate telencephalon also support the notion of distinct
lateral signals. For example, Pax6 and Gsh2 show graded
expression patterns in the dorsal and ventral portions,
respectively, with their highest expression occurring at
the pallio-subpallial boundary [13,14]. Previous work has
shown that a SHH-independent retinoic acid (RA) path-
way can pattern the intermediate region of the developing
spinal cord [53]. Interestingly, at early stages of telence-
phalic patterning, mesenchymal cells in the olfactory
placodes that produce RA are in close proximity to the
ventral telencephalon and induce the expression of reti-
noid reporter genes in the ventrolateral region [54]. Thus,
RA may participate in patterning the telencephalon.
As mentioned above, the pallio-subpallial boundary
divides the telencephalon into a dorsal and ventral por-
tion. Pax6 and Gsh2 are crucial for the positioning of
this boundary through the regulation of mutually re-
pressive genetic programs [13,14]. The telencephalic
subdomains lying directly on each side of the pallio-
subpallial boundary (i.e. the dorsal LGE and the ventral
pallium) are most dependent on correct positioning of
this boundary. In the absence of Pax6, genes character-
istic of the dorsal LGE become ectopically expressed
early in the ventral pallium and subsequently spread as
far as the dorsal pallium [13,28�]. The opposite is true
in Gsh2 mutants, in which ventral pallial markers are
ectopically expressed in the LGE [13]. These altera-
tions in gene expression do not represent a transforma-
tion of the dorsal LGE into the ventral pallium in Gsh2mutants or vice versa in Pax6 mutants, because other
markers of DV identity are expressed normally in these
mutants [14].
Figure 2
Wnts
BMPs
RA??
SHH?
MGE
vLGE
dLGE
VP
LP
DPMPrp
Current Opinion in Neurobiology
The mouse telencephalon at embryonic day 10, showing the extrinsic
signals that establish different progenitor domains along the DV axis.BMPs are most highly expressed in dorsal–medial portions of the
telencephalon, where they are required to act locally for midline
development. Wnts are expressed throughout the pallium and may
function more broadly in dorsal specification than BMPs. Although SHH
is required for ventromedial specification, some aspects of ventrolateral
patterning can occur in its absence. DV patterning can occur in the
absence of both Shh and Gli3, suggesting the existence of alternative
signals and/or pathways in ventralizing the telencephalon. Lateral
signals are proposed to participate in patterning the intermediate
telencephalon. RA may be involved in this process. dLGE, dorsal LGE;
DP, dorsal pallium; LP, lateral pallium; MP, medial pallium; rp, roof
plate; vLGE, ventral LGE; VP, ventral pallium.
Dorsal-ventral patterning in the mammalian telencephalon Campbell 53
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:50–56

The fact that Pax6 and Gsh2 do not control all aspects of
the DV identity in the intermediate telencephalon indi-
cates that parallel and overlapping genetic pathways are
also involved. Indeed, Gli3 mutants show ectopic ventral
gene expression in the pallium similar to that seen in Pax6mutants, despite the fact that Pax6 seems to be normally
expressed [41,43��]. In addition, a recent study [55��] has
shown that in Emx2�/�;Pax6�/� mutants dorsal patterning
is considerably more perturbed than in single Pax6mutants. Although Emx2 mutants do not show overt
patterning defects in the intermediate telencephalon,
removal of Pax6 gene function from the Emx2 mutant
background causes the whole pallium to lose its dorsal
identity [55��]. In the absence of pallial gene expression,
markers of the ventral LGE (e.g. Islet-1) are expressed in
the dorsal telencephalon of these double mutants, sug-
gesting that there is a transformation of the pallium into
subpallium. Thus, correct DV patterning in the telence-
phalon relies on the superimposition of several genetic
pathways.
ConclusionsWork carried out to date has provided a substantial
framework on which to uncover the mechanisms that
control DV identity in the developing telencephalon.
The identification and refinement of progenitor domains
along the DV axis has been and will continue to be central
to this endeavor. Signals from each of the dorsal, ventral
and possibly lateral aspects of the developing telence-
phalon participate in the establishment and maintenance
of these progenitor domains, by regulating both separate
and overlapping genetic pathways (Figure 2). In future
studies it will be important to define further the roles of
BMPs, RA, SHH and Wnts, as well as other as yet
unknown signals, in DV patterning of the telencephalon.
Many of the mechanisms involved in patterning the DV
axis of the spinal cord are also at work in the telence-
phalon. What is it, then, that makes the mature telence-
phalon so much more complex than the spinal cord? A
recent study has provided evidence that, in addition to
spatial (e.g. DV) patterning, temporal patterning events
contribute extensively to generating neuronal diversity in
the Drosophila CNS [56]. Because the telencephalon
shows a very protracted period of neurogenesis as com-
pared with the spinal cord, it may be that different
neuronal subtypes are generated from each telencephalic
progenitor domain at specific time points during devel-
opment. Thus, it will be important in future work to
establish a connection between DV patterning and tem-
poral patterning to account for the generation of all of the
subtypes of neurons present in the telencephalon.
UpdateA recent study has demonstrated a role for the orphan
nuclear receptor TLX in the establishment of the pallio-
subpallial boundary and maintenance of ventral pallial
identity [57�]. Interestingly, a genetic interaction between
TLX and Pax6 was found to be crucial for this process.
AcknowledgementsI thank Doug Epstein, Magdalena Gotz, Mike Matise and Jan Stenman forcritically reading the manuscript, and Gord Fishell for providing preprints ofpapers discussed in the review.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest��of outstanding interest
1. Gorski JA, Talley T, Qiu M, Puelles L, Rubenstein JL, Jones KR:Cortical excitatory neurons and glia, but not GABAergicneurons, are produced in the Emx1-expressing lineage.J Neurosci 2002, 22:6309-6314.
2. Stuhmer T, Puelles L, Ekker M, Rubenstein JL: Expression from aDlx gene enhancer marks adult mouse cortical GABAergicneurons. Cereb Cortex 2002, 12:75-85.
3. Marin O, Rubenstein JL: A long, remarkable journey: tangentialmigration in the telencephalon. Nat Rev Neurosci 2001, 2:780-790.
4.��
Wichterle H, Turnbull DH, Nery S, Fishell G, Alvarez-Buylla A: Inutero fate mapping reveals distinct migratory pathways andfates of neurons born in the mammalian basal forebrain.Development 2001, 128:3759-3771.
Although many slice culture studies have shown tangential migration ofneurons from the medial ganglionic eminence to the dorsal telencepha-lon, this study was the first in vivo demonstration of this importantphenomenon.
5. Olsson M, Campbell K, Turnbull DH: Specification of mousetelencephalic and mid-hindbrain progenitors followingheterotopic ultrasound-guided embryonic transplantation.Neuron 1997, 19:761-772.
6. Olsson M, Bjorklund A, Campbell K: Early specification of striatalprojection neurons and interneuronal subtypes in the lateral andmedial ganglionic eminence. Neuroscience 1998, 84:867-876.
7. Marin O, Anderson SA, Rubenstein JL: Origin and molecularspecification of striatal interneurons. J Neurosci 2000,20:6063-6076.
8. Deacon TW, Pakzaban P, Isacson O: The lateral ganglioniceminence is the origin of cells committed to striatalphenotypes: neural transplantation and developmentalevidence. Brain Res 1994, 668:211-219.
9. Wichterle H, Garcia-Verdugo JM, Herrera DG, Alvarez-Buylla A:Young neurons from medial ganglionic eminence disperse inadult and embryonic brain. Nat Neurosci 1999, 2:461-466.
10.�
Nery S, Fishell G, Corbin JG: The caudal ganglionic eminence is anovel source of distinct cortical and subcortical cellpopulations. Nat Neurosci 2002, 5:1279-1282.
The CGE has been proposed to represent a distinct structure in thedeveloping telencephalon. This study provides the first functional evi-dence of this by using ultrasound-guided embryonic transplantation tomap the fates of cells derived from this region.
11. Jessell TM: Neuronal specification in the spinal cord: inductivesignals and transcriptional codes. Nat Rev Genet 2000, 1:20-29.
12. Briscoe J, Ericson J: Specification of neuronal fates in theventral neural tube. Curr Opin Neurobiol 2001, 11:43-49.
13. Yun K, Potter S, Rubenstein JL: Gsh2 and Pax6 playcomplementary roles in dorsoventral patterning of themammalian telencephalon. Development 2001, 128:193-205.
14. Toresson H, Potter SS, Campbell K: Genetic control of dorsal–ventral identity in the telencephalon: opposing roles for Pax6and Gsh2. Development 2000, 127:4361-4371.
15. Puelles L, Kuwana E, Puelles E, Rubenstein JL: Comparison of themammalian and avian telencephalon from the perspective ofgene expression data. Eur J Morphol 1999, 37:139-150.
54 Development
Current Opinion in Neurobiology 2003, 13:50–56 www.current-opinion.com

16. Puelles L, Kuwana E, Puelles E, Bulfone A, Shimamura K, Keleher J,Smiga S, Rubenstein JL: Pallial and subpallial derivatives in theembryonic chick and mouse telencephalon, traced by theexpression of the genes Dlx-2, Emx-1, Nkx- 2.1, Pax-6, andTbr-1. J Comp Neurol 2000, 424:409-438.
17. Stoykova A, Treichel D, Hallonet M, Gruss P: Pax6 modulates thedorsoventral patterning of the mammalian telencephalon.J Neurosci 2000, 20:8042-8050.
18. Kim AS, Anderson SA, Rubenstein JL, Lowenstein DH, Pleasure SJ:Pax-6 regulates expression of SFRP-2 and Wnt-7b in thedeveloping CNS. J Neurosci 2001, 21:RC132(1-5).
19. Simeone A, Gulisano M, Acampora D, Stornaiuolo A, Rambaldi M,Boncinelli E: Two vertebrate homeobox genes related to theDrosophila empty spiracles gene are expressed in theembryonic cerebral cortex. EMBO J 1992, 11:2541-2550.
20. Gulisano M, Broccoli V, Pardini C, Boncinelli E: Emx1 and Emx2show different patterns of expression during proliferation anddifferentiation of the developing cerebral cortex in the mouse.Eur J Neurosci 1996, 8:1037-1050.
21.��
Monuki ES, Porter FD, Walsh CA: Patterning of the dorsaltelencephalon and cerebral cortex by a roof plate–Lhx2pathway. Neuron 2001, 32:591-604.
This study provides evidence that the roof plate is an important signalingcenter for dorsal telencephalic development. The results from this studyimplicate BMPs and possibly also Wnts in patterning the dorsal tele-ncephalon.
22. Hsieh-Li HM, Witte DP, Szucsik JC, Weinstein M, Li H, Potter SS:Gsh-2, a murine homeobox gene expressed in the developingbrain. Mech Dev 1995, 50:177-186.
23. Eisenstat DD, Liu JK, Mione M, Zhong W, Yu G, Anderson SA,Ghattas I, Puelles L, Rubenstein JL: DLX-1, DLX-2, and DLX-5expression define distinct stages of basal forebraindifferentiation. J Comp Neurol 1999, 414:217-237.
24. Shimamura K, Hartigan DJ, Martinez S, Puelles L, Rubenstein JL:Longitudinal organization of the anterior neural plate andneural tube. Development 1995, 121:3923-3933.
25. Sussel L, Marin O, Kimura S, Rubenstein JL: Loss of Nkx2.1homeobox gene function results in a ventral to dorsalmolecular respecification within the basal telencephalon:evidence for a transformation of the pallidum into the striatum.Development 1999, 126:3359-3370.
26.�
Nery S, Wichterle H, Fishell G: Sonic hedgehog contributes tooligodendrocyte specification in the mammalian forebrain.Development 2001, 128:527-540.
See annotation [27�].
27.�
Tekki-Kessaris N, Woodruff R, Hall AC, Gaffield W, Kimura S, StilesCD, Rowitch DH, Richardson WD: Hedgehog-dependentoligodendrocyte lineage specification in the telencephalon.Development 2001, 128:2545-2554.
Together with [26�], this study shows that the ventromedial telencephalonis a principal source of oligodendrocytes for this brain region and thatSHH is involved in generating this cell type.
28.�
Stenman J, Toresson H, Campbell K: Identification of two distinctprogenitor populations in the lateral ganglionic eminence:implications for striatal and olfactory bulb neurogenesis.J Neurosci 2003, 23:167-174.
This study shows that the Dlx-expressing subventricular zone ofthe LGE can be divided into at least two distinct progenitor domains:a dorsal domain marked by Er81, which is proposed to generate inter-neurons that migrate rostrally to the olfactory bulb; and a more ventraldomain expressing Islet-1, which gives rise to the striatal projectionneurons.
29. Furuta Y, Piston DW, Hogan BL: Bone morphogenetic proteins(BMPs) as regulators of dorsal forebrain development.Development 1997, 124:2203-2212.
30.�
Hebert J, Mishina Y, McConnell S: BMP signaling is requiredlocally to pattern the dorsal telencephalic midline. Neuron 2002,35:1029.
This study addresses the role of BMPs in patterning the dorsal telence-phalon by conditional inactivation of the BMP receptor 1a. The resultsindicate that loss of BMP signaling in the dorsal telencephalon results in anormal dorsal–ventral pattern but a lack of normal development of the
choroid plexus, suggesting a local role for BMPs only in the dorsaltelencephalic midline.
31. Grove EA, Tole S, Limon J, Yip L, Ragsdale CW: The hem of theembryonic cerebral cortex is defined by the expression ofmultiple Wnt genes and is compromised in Gli3-deficient mice.Development 1998, 125:2315-2325.
32. Lee SM, Tole S, Grove E, McMahon AP: A local Wnt-3a signal isrequired for development of the mammalian hippocampus.Development 2000, 127:457-467.
33. Galceran J, Miyashita-Lin EM, Devaney E, Rubenstein JL,Grosschedl R: Hippocampus development and generation ofdentate gyrus granule cells is regulated by LEF1. Development2000, 127:469-482.
34. Parr BA, Shea MJ, Vassileva G, McMahon AP: Mouse Wnt genesexhibit discrete domains of expression in the early embryonicCNS and limb buds. Development 1993, 119:247-261.
35.�
Theil T, Aydin S, Koch S, Grotewold L, Ruther U: Wnt and Bmpsignalling cooperatively regulate graded Emx2 expressionin the dorsal telencephalon. Development 2002,129:3045-3054.
In this paper, a telencephalic enhancer for the Emx2 gene is identified andshown to contain requisite Smad- and Tcf-binding sites, implicating bothBMPs and Wnts in the regulation of this dorsal telencephalic gene.
36. Porter FD, Drago J, Xu Y, Cheema SS, Wassif C, Huang SP, Lee E,Grinberg A, Massalas JS, Bodine D et al.: Lhx2, a LIM homeoboxgene, is required for eye, forebrain, and definitive erythrocytedevelopment. Development 1997, 124:2935-2944.
37. Bulchand S, Grove EA, Porter FD, Tole S: LIM-homeodomaingene Lhx2 regulates the formation of the cortical hem.Mech Dev 2001, 100:165-175.
38. Pellegrini M, Mansouri A, Simeone A, Boncinelli E, Gruss P:Dentate gyrus formation requires Emx2. Development 1996,122:3893-3898.
39. Yoshida M, Suda Y, Matsuo I, Miyamoto N, Takeda N, Kuratani S,Aizawa S: Emx1 and Emx2 functions in development of dorsaltelencephalon. Development 1997, 124:101-111.
40. Tole S, Goudreau G, Assimacopoulos S: Grove EA: Emx2 isrequired for growth of the hippocampus but not forhippocampal field specification. J Neurosci 2000, 20:2618-2625.
41. Theil T, Alvarez-Bolado G, Walter A, Ruther U: Gli3 is required forEmx gene expression during dorsal telencephalondevelopment. Development 1999, 126:3561-3571.
42. Tole S, Ragsdale CW, Grove EA: Dorsoventral patterning of thetelencephalon is disrupted in the mouse mutant extra-toes(J).Dev Biol 2000, 217:254-265.
43.��
Rallu M, Machold RP, Gaiano N, Corbin JG, McMahon AP, FishellG: Dorsoventral patterning is established in the telencephalonof mutants lacking both Gli3 and Hedgehog signaling.Development 2002, 129:4963-4974.
The results of this paper provide evidence for the cross-repressiveactivities of SHH and Gli3 in the telencephalon. In the absence ofboth Shh and Gli3 gene function, dorsal–ventral patterning is markedlybetter than in either single mutant. These findings suggests that anothersignal or signals can pattern the telencephalon in the absence of SHHand Gli3.
44. Ericson J, Muhr J, Placzek M, Lints T, Jessell TM, Edlund T: Sonichedgehog induces the differentiation of ventral forebrainneurons: a common signal for ventral patterning within theneural tube. Cell 1995, 81:747-756.
45. Kohtz JD, Baker DP, Corte G, Fishell G: Regionalization within themammalian telencephalon is mediated by changes inresponsiveness to Sonic Hedgehog. Development 1998,125:5079-5089.
46. Gunhaga L, Jessell TM, Edlund T: Sonic hedgehog signaling atgastrula stages specifies ventral telencephalic cells in thechick embryo. Development 2000, 127:3283-3293.
47. Gaiano N, Kohtz JD, Turnbull DH, Fishell G: A method for rapidgain-of-function studies in the mouse embryonic nervoussystem. Nat Neurosci 1999, 2:812-819.
Dorsal-ventral patterning in the mammalian telencephalon Campbell 55
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:50–56

48. Corbin JG, Gaiano N, Machold RP, Langston A, Fishell G: The Gsh2homeodomain gene controls multiple aspects of telencephalicdevelopment. Development 2000, 127:5007-5020.
49. Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H,Beachy PA: Cyclopia and defective axial patterning in micelacking Sonichedgehoggene function. Nature 1996,383:407-413.
50. Pabst O, Herbrand H, Takuma N, Arnold HH: NKX2 geneexpression in neuroectoderm but not in mesendodermallyderived structures depends on sonic hedgehog in mouseembryos. Dev Genes E 2000, 210:47-50.
51. Litingtung Y, Chiang C: Specification of ventral neuron types ismediated by an antagonistic interaction between Shh and Gli3.Nat Neurosci 2000, 3:979-985.
52. Ingham PW, McMahon AP: Hedgehog signaling in animaldevelopment: paradigms and principles. Genes Dev 2001,15:3059-3087.
53. Pierani A, Brenner-Morton S, Chiang C, Jessell TM: A sonichedgehog-independent, retinoid-activated pathway ofneurogenesis in the ventral spinal cord. Cell 1999,97:903-915.
54. LaMantia AS, Colbert MC, Linney E: Retinoic acid induction andregional differentiation prefigure olfactory pathway formationin the mammalian forebrain. Neuron 1993, 10:1035-1048.
55.��
Muzio L, DiBenedetto B, Stoykova A, Boncinelli E, Gruss P,Mallamaci A: Conversion of cerebral cortex into basal ganglia inEmx2�/� Pax6Sey/Sey double-mutant mice. Nat Neurosci 2002,15:15.
In this study, loss of function for both Pax6 and Emx2 is shown to result ina respecification of the pallium towards subpallial fates typical of theventral LGE (i.e. striatal specification). Because Emx2 mutants do notshow overt patterning defects, these findings reveal a role for Emx2 in theabsence of Pax6 gene function.
56. Isshiki T, Pearson B, Holbrook S, Doe CQ: Drosophila neuroblastssequentially express transcription factors which specify thetemporal identity of their neuronal progeny. Cell 2001,106:511-521.
57.�
Stenman J, Yu RT, Evans RM, Campbell K: TLX and Pax6cooperate genetically to establish the pallio-subpallialboundary in the embryonic mouse telencephalon. Development2003, in press.
See update.
56 Development
Current Opinion in Neurobiology 2003, 13:50–56 www.current-opinion.com