
Distribution of Myrosinase in Rapeseed Tissues1
9
Plant Physiol. (1991) 95, 213-221 0032-0889/91 /95/0000/09/$01 .00/0 Received for publication July 23, 1990 Accepted September 13, 1990 Distribution of Myrosinase in Rapeseed Tissues1 Anna-Stina Hoglund, Marit Lenman, Anders Falk, and Lars Rask* Department of Cell Research, Uppsala Biomedical Center, Swedish University of Agricultural Sciences, Box 596, S-751 24 Uppsala, Sweden ABSTRACT Immunocytochemical studies on Brassica napus (rapeseed) tissues using a monoclonal antibody against myrosinase (thiog- lucoside glucohydrolase) showed that the enzyme was only pres- ent in a small number of cells. In the developing embryo, scattered myrosinase-containing cells were present in both cotyledons and axis. The enzyme accumulated in these cells during the later stages of seed development, approximately from day 20 until day 40 after pollination. Parallel staining with the immunocytochemical technique and a histochemical method identified these cells as myrosin cells. Myrosinase appeared to be located outside the myrosin grains, although the occasional association with the membrane of the grains also was noted. In leaves, petals, and siliques, scattered parenchyma cells were stained in the meso- phyll as well as in the vascular tissue. In young leaves, guard cells also contained myrosinase. The enzyme was also present in xylem cells of the stem. The glucosinolates are a group of about 100 naturally occurring low mol wt compounds found mainly in members of the Cruciferae. They consist of a thioglucoside linked to a variety of side chains which are usually amino acid derivatives. Coexisting with the glucosinolates in the plant tissue is the enzyme myrosinase or thioglucoside glucohydrolase (EC 3.2.3.1). This enzyme catalyzes the hydrolysis of glucosino- lates to yield glucose, sulfate and, depending on the glucosi- nolate species, any one of the following products; isothiocya- nates, thiocyanates, nitriles, cyanoepithioalkanes, or oxazoli- dine-2-thiones (2, 8, 14). Myrosinase from rapeseed has been purified and characterized. It consists of several isoforms. Various degrees of glycosylation account for at least part of the charge heterogeneity seen in this enzyme (10). By the use of monoclonal and polyclonal antibodies, two main types of myrosinase were found in rapeseed protein extract: 75 kD subunits occurred as free dimers, whereas 70 and 65 kD myrosinases existed in complexes with two 52 and 50 kD myrosinase-binding proteins (9). The function of the latter proteins is not known. Hitherto there has not been a method for direct identifica- tion of myrosinase in tissue sections. On the basis of mor- phology and by certain staining techniques like Millon's re- agent and Saffranin-fast green, myrosin cells or protein-ac- cumulating idioblasts have been identified (4, 5, 15). In these This work was supported by grants from The Swedish Natural Science Research Council and from the Swedish Research Council for Forestry and Agriculture. 213 cells, certain globular structures, myrosin grains, were stained (13, 18) and the "myrosin" was assumed to be myrosinase (19). To localize the enzyme in situ by electron microscopy, some investigators have taken advantage of the possibility of forming an electron-dense precipitate of barium- or lead- sulfate during the hydrolysis of glucosinolates at the sites of myrosinase activity. The distribution of the precipitate thus formed was then assumed to represent the location of the myrosinase (6). The enzyme was thereby detected in the majority of the cells in the embryo where it appeared to be localized to the plasma membrane (12). Later observations using subcellular fractionation suggested that myrosinase is present in the cytoplasm but has a remark- able tendency to adhere to membrane surfaces (1 1), while the glucosinolates are found inside the myrosin grains/vacuoles. This distribution of myrosinase as a cytosolic enzyme agrees well with the hypothesis of a different subcellular localization of the enzyme and of the glucosinolates. In the present study, we have used antibodies made against myrosinase to detect the enzyme in the embryo during seed development and germination, as well as in adult plants of Brassica napus. MATERIALS AND METHODS Plant Material A dihaploid line of rapeseed (Brassica napus), Svalofs Karat (Svalof AB, Svalov, Sweden), was used in -this study. Seeds were germinated at 25°C in the dark for 2 to 8 d in a constant- environment cabinet. Plants were grown in soil in a green- house with 18 h light and 6 h darkness. Monoclonal and Polyclonal Antimyrosinase Antibodies Myrosinase was purified from rapeseed by a combination of ammonium sulfate precipitation and ion exchange chro- matography. The resulting enzyme preparation consisted of a dimer of 75 kD chains. This material was used as antigen for the production of monoclonal and polyclonal antibodies. The monoclonal antibodies were selected for their ability to pre- cipitate myrosinase activity or to react with myrosinase in Western analysis. One of these monoclonal antibodies, 3D7, as well as a rabbit polyclonal antiserum, K505, reacted with myrosinase species with subunits of molecular masses 75, 70, and 65 kD. Quantitative immunoprecipitations with K505 showed that this antiserum precipitated at least 80% of the total myrosinase activity in a rapeseed protein extract. 3D7 and K505 were selected for use in immunocytochemical studies because they reacted well with rapeseed tissues after www.plantphysiol.org on April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
Transcript of Distribution of Myrosinase in Rapeseed Tissues1

Plant Physiol. (1991) 95, 213-2210032-0889/91 /95/0000/09/$01 .00/0
Received for publication July 23, 1990Accepted September 13, 1990
Distribution of Myrosinase in Rapeseed Tissues1
Anna-Stina Hoglund, Marit Lenman, Anders Falk, and Lars Rask*Department of Cell Research, Uppsala Biomedical Center, Swedish University of Agricultural Sciences, Box 596,
S-751 24 Uppsala, Sweden
ABSTRACT
Immunocytochemical studies on Brassica napus (rapeseed)tissues using a monoclonal antibody against myrosinase (thiog-lucoside glucohydrolase) showed that the enzyme was only pres-ent in a small number of cells. In the developing embryo, scatteredmyrosinase-containing cells were present in both cotyledons andaxis. The enzyme accumulated in these cells during the laterstages of seed development, approximately from day 20 until day40 after pollination. Parallel staining with the immunocytochemicaltechnique and a histochemical method identified these cells asmyrosin cells. Myrosinase appeared to be located outside themyrosin grains, although the occasional association with themembrane of the grains also was noted. In leaves, petals, andsiliques, scattered parenchyma cells were stained in the meso-phyll as well as in the vascular tissue. In young leaves, guardcells also contained myrosinase. The enzyme was also presentin xylem cells of the stem.
The glucosinolates are a group of about 100 naturallyoccurring low mol wt compounds found mainly in membersof the Cruciferae. They consist of a thioglucoside linked to a
variety ofside chains which are usually amino acid derivatives.Coexisting with the glucosinolates in the plant tissue is theenzyme myrosinase or thioglucoside glucohydrolase (EC3.2.3.1). This enzyme catalyzes the hydrolysis of glucosino-lates to yield glucose, sulfate and, depending on the glucosi-nolate species, any one of the following products; isothiocya-nates, thiocyanates, nitriles, cyanoepithioalkanes, or oxazoli-dine-2-thiones (2, 8, 14). Myrosinase from rapeseed has beenpurified and characterized. It consists of several isoforms.Various degrees of glycosylation account for at least part ofthe charge heterogeneity seen in this enzyme (10). By the use
of monoclonal and polyclonal antibodies, two main types ofmyrosinase were found in rapeseed protein extract: 75 kDsubunits occurred as free dimers, whereas 70 and 65 kDmyrosinases existed in complexes with two 52 and 50 kDmyrosinase-binding proteins (9). The function of the latterproteins is not known.
Hitherto there has not been a method for direct identifica-tion of myrosinase in tissue sections. On the basis of mor-phology and by certain staining techniques like Millon's re-
agent and Saffranin-fast green, myrosin cells or protein-ac-cumulating idioblasts have been identified (4, 5, 15). In these
This work was supported by grants from The Swedish NaturalScience Research Council and from the Swedish Research Councilfor Forestry and Agriculture.
213
cells, certain globular structures, myrosin grains, were stained(13, 18) and the "myrosin" was assumed to be myrosinase(19). To localize the enzyme in situ by electron microscopy,some investigators have taken advantage of the possibility offorming an electron-dense precipitate of barium- or lead-sulfate during the hydrolysis of glucosinolates at the sites ofmyrosinase activity. The distribution of the precipitate thusformed was then assumed to represent the location of themyrosinase (6). The enzyme was thereby detected in themajority of the cells in the embryo where it appeared to belocalized to the plasma membrane (12).
Later observations using subcellular fractionation suggestedthat myrosinase is present in the cytoplasm but has a remark-able tendency to adhere to membrane surfaces (1 1), while theglucosinolates are found inside the myrosin grains/vacuoles.This distribution of myrosinase as a cytosolic enzyme agreeswell with the hypothesis of a different subcellular localizationof the enzyme and of the glucosinolates.
In the present study, we have used antibodies made againstmyrosinase to detect the enzyme in the embryo during seeddevelopment and germination, as well as in adult plants ofBrassica napus.
MATERIALS AND METHODS
Plant Material
A dihaploid line ofrapeseed (Brassica napus), Svalofs Karat(Svalof AB, Svalov, Sweden), was used in -this study. Seedswere germinated at 25°C in the dark for 2 to 8 d in a constant-environment cabinet. Plants were grown in soil in a green-house with 18 h light and 6 h darkness.
Monoclonal and Polyclonal Antimyrosinase Antibodies
Myrosinase was purified from rapeseed by a combinationof ammonium sulfate precipitation and ion exchange chro-matography. The resulting enzyme preparation consisted of adimer of 75 kD chains. This material was used as antigen forthe production of monoclonal and polyclonal antibodies. Themonoclonal antibodies were selected for their ability to pre-cipitate myrosinase activity or to react with myrosinase inWestern analysis. One of these monoclonal antibodies, 3D7,as well as a rabbit polyclonal antiserum, K505, reacted withmyrosinase species with subunits of molecular masses 75, 70,and 65 kD. Quantitative immunoprecipitations with K505showed that this antiserum precipitated at least 80% of thetotal myrosinase activity in a rapeseed protein extract. 3D7and K505 were selected for use in immunocytochemicalstudies because they reacted well with rapeseed tissues after
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 95, 1991
Figure 1. The presence of myrosinase in cells of rapeseed embryos 35 to 40 d after pollination, as detected by immunohistochemical staining.In (a) a rabbit antiserum (K505), in (b) the preimmune rabbit serum, in (c) a monoclonal antibody (3D7) against myrosinase, and in (d) a monoclonalantibody against human #2-microglobulin (as a control) were used. Scale bar equals 10 ,um.
fixation. A detailed characterization of these antibodies hasbeen presented elsewhere (9).
Immunocytochemistry
Whole seeds or tissues cut into small pieces (1 x 2 mm)were fixed in 10% (w/v) formaldehyde in PBS for 2 h to 7 dat 4°C. Samples were then dehydrated through a graded seriesof alcohol followed by incubation in xylene before infiltrationand embedding in histowax (tissue embedding mediumn, pel-lets m.p. 56-58°C, Histolab, Gothenburg, Sweden). Sections
(4,um thick) were cut with a Reichert microtome (Nussloch,FRG) and placed on chrome-gelatin-coated coverslips. Beforestaining, sections were rehydrated in xylene followed by de-creasing concentrations of ethanol and finally distilled water,after which they were treated with 1% H202 in H20 for 20min. To prevent nonspecific binding of the antibodies, thesections were incubated in normal goat serum diluted 1/10in PBS containing 4% (w/v) BSA for 15 min. Myrosinase wasdetected by 1 h incubation at room temperature with themonoclonal antibody 3D7, diluted 1/400 in PBS, followedby rabbit anti-mouse IgG antiserum, diluted 1/40 (DAKO-
214 HOGLUND ET AL.
oi
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

DISTRIBUTION OF MYROSINASE IN RAPESEED TISSUES
C
4.'1
Figure 2. Visualization of myrosinase in cells of a 39-d-old embryo of rapeseed. Consecutive sections were used for staining with a monoclonalantibody, 3D7, directed against myrosinase (a) and with Millon's reagent (b). The same cells were stained by both methods (arrows). The stainingof the cytoplasm with the antibodies and the myrosin grains with Millon's reagent is illustrated at higher magnification in (c) and (d), respectively.For (a) and (b) scale bar equals 10 Am, for (c) and (d) scale bar equals 1 ;m.
215
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 95, 1991
PATTS a/s, Copenhagen) in PBS containing 4% BSA, for 30min. Finally, the sections were incubated with a complex ofhorseradish peroxidase and mouse antiperoxidase antibodies,diluted 1/250 in PBS with 4% BSA, for 30 min followed bydevelopment in 0.8 mM 3-amino-9-ethyl carbazole, 5%DMSO, 0.02 M sodium acetate (pH 5.0), to which 30% H202was added to a final concentration of 0.05% (v/v). When thepolyclonal antiserum K505 was used, it was diluted 1/12,000in PBS. Goat anti-rabbit IgG antiserum, diluted 1/40 in PBSwith 4% BSA, was employed as the secondary antibody. Theimmunocomplexes were detected by horse radish peroxidaseand rabbit antiperoxidase antibodies followed by developmentas described above. For counter staining, Mayers HTX wasused followed by a 10 min rinse in water. Specimens weremounted in glycine-gelatin and micrographs were taken in aNikon optiphot.
In control experiments, the staining procedure was carriedout as above but with the 3D7 antibody either replaced witha monoclonal antibody against an irrelevant antigen (human32-microglobulin) or pre-absorbed with excessive amounts ofpurified myrosinase prior to being applied to the specimens.For the polyclonal rabbit antiserum (K505), pre-immuneserum was used.
Histochemical Staining of Myrosin Cells
Myrosin cells were identified in rapeseed embryos by stain-ing with Millon's reagent as described (7).
Western Analysis
Proteins were separated by electrophoresis in 10 to 15%polyacrylamide gradient gel containing SDS (1) and subse-quently electrotransferred to nitrocellulose membranes (17).The filter was stained according to the protocols for theProtoblot system (Promega Biotec, Madison, WI). 3D7 wasused as the primary antibody.
RESULTS
Immunolocalization of myrosinase in sections of Brassicanapus embryos using either the polyclonal antiserum K505or the monoclonal antibody 3D7 showed that the enzymewas present in only a small number of cells (Fig. la, c). Noreactivity was seen when K505 was replaced with normalrabbit serum (Fig. lb), or when 3D7 had been exchanged fora monoclonal antibody towards a human membrane protein(Fig. ld) or preabsorbed with purified myrosinase (notshown). Since the staining with 3D7 gave a higher reproduc-ibility, this reagent was used throughout this study.The distribution of myrosinase as seen by immunohisto-
chemistry was compared to that observed when the traditionaltechnique for detecting myrosin cells, the Millon's reagent,was used on consecutive sections of a rapeseed embryo taken39 d after pollination. Evidently, the two procedures identifiedthe same set of cells (Fig. 2). The antibody detected theenzyme either uniformly distributed in the cytoplasm ororganized as particulate bodies. In addition, marked accu-mulation could be seen at the periphery of the myrosin grains(Fig. 2c).
To investigate the expression of myrosinase during embry-ogenesis, immature seeds were sampled at different times afterpollination and stained for the enzyme. The results obtainedby the immunolocalization technique showed that the enzymeaccumulates during a relatively well defined period of theembryogenesis. The first myrosinase-containing cells ap-peared at approximately d 20, and they increased in numberuntil approximately d 30, when 2 to 5% of the cells in eachsection were stained. At later stages of embryogenesis, thenumber of myrosin cells remained constant even though thestaining became increasingly more intense (data not shown).The synthesis of myrosinase during embryo growth was
also analyzed by immunodetection with the 3D7 antibody onWestern blots of protein extracts from seeds. According tothese data, myrosinase appeared at d 30 after pollination andincreased in amount to approximately d 40, after which itremained constant until the end of the ripening process (Fig.3). A difference in sensitivity between the two methods mayaccount for the somewhat later appearance of the enzymewhen detected by immunoblotting as compared to the lightmicroscopy.
In the seed, myrosinase was found solely in embryonic cellswith a uniform distribution and no preference for either thecotyledons or the axis. The cells in the epidermis, meriste-matic cells in the axis, and the provascular cells in the coty-ledon (not shown), however, did not appear to possess theenzyme. This distribution of myrosin cells in the embryoremained unchanged for the first 2 d after germination. Theintracellular localization of the enzyme, however, changedtoward a more pronounced accumulation around the vacu-oles/myrosin grains during this period (Fig. 4a). Eight daysafter initiation of germination, the staining of the cotyledonrevealed that myrosinase was expressed in the newly formedguard cells as well as in a small number of parenchyma cells(Fig. 4c). In the axis, a few elongated parenchyma cells were
:3
CD lr EOO N.OCNM
kD-94
43
Figure 3. Western immunoblot analysis of myrosinase in total proteinextracts from developing seeds sampled at different time points afterpollination. Protein (50 ,Ag) was loaded in each well. A 10 to 15%SDS-polyacrylamide gradient gel was used to separate the proteins.The figures above the lanes denote days after pollination. Molecularmass markers indicated are phosphorylase B (94 kD), bovine serum
albumin (67 kD), and ovalbumin (43 kD).
HOGLUND ET AL.216
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

:*nF A .. .~~0,Z t o - s . v+ ' ..,, ,. 4.b, -
7sX , a
? _'t #~
40
I.
Figure 4. Presence of myrosinase in seeds during germination as visualized by staining with the monoclonal antibody 3D7. In panel (a) a highmagnification of a cotyledon cell 2 d after initiation of germination, myrosinase appeared to be accumulated around the vacuoles. Panels (b) and(c) show the embryo 8 d after initiation of germination: in (b) part of axis with two positive parenchyma cells and some positive vascular cells, in(c) part of a cotyledon with stained guard cells and a few stained parenchyma cells. Insets show some of these cells at higher magnification(arrows in low magnification). Bar in low magnifications equals 10 Am, in high magnifications 1 gim.
217
YF.7E
IfI.A
Fb
I.I. '
.,If-
4
0
.i
1
9'
trIs ) ;
.4.i*e
? t .
X, -.,
+ -_. , i 5
v ^ \ i
* 8 8 8 t 4X 2,ts X 's. A.;:
.
*S....*_s _
.:t..-
_t?. /.+ +_'' j., u |
.@ a
8 ''"i..
I
iA?
.' "f ...It2t
412MIL.
.4.1
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

b
.4
Figure 5. Localization of myrosinase in leaf, petal, and silique of the plant using the monoclonal antibody 307. Panel (a) shows part of a foliageleaf with stained guard cells and parenchyma cells in the vascular tissue. Panels (b) and (c) are parts of a petal and a silique, respectively, withstained parenchyma cells in the vascular tissue. Bar in low magnifications equals 10 Am, in high magnifications 1 Am.
218
...;
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.
DISTRIBUTION OF MYROSINASE IN RAPESEED TISSUES 219
4s .X'J~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~'A A
44
I I
A ., jv, .~r44 r< J
>~~~~~~~~~I;-~ a .
j. X FttS
4, 4 _, * * 9 -, -t
tf,/t~~~~~~~A,*,,^ s '?~~*~~~~~h-KCttwv*J N.
A-'
4. 4~~~~~~~~N 0
I ~~*
~~trr-A- Aw ~
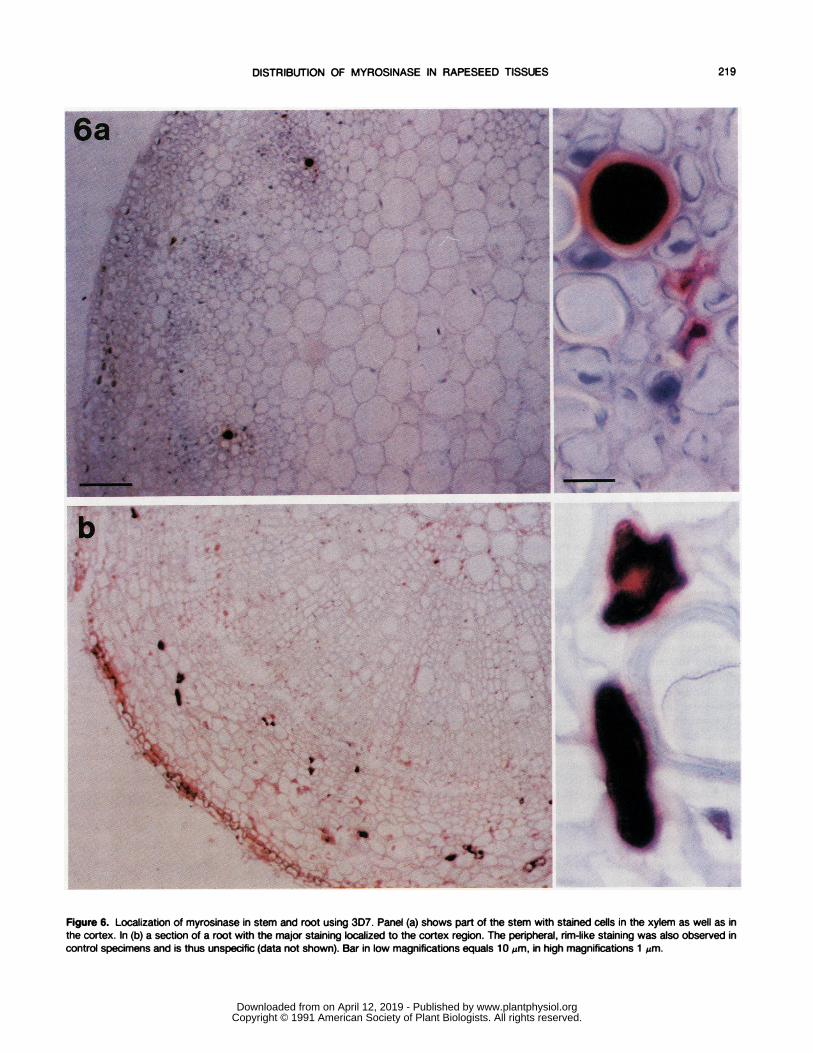
Figure 6. Localization of myrosinase in stem and root using 3D7. Panel (a) shows part of the stem with stained cells in the xylem as well as inthe cortex. In (b) a section of a root with the major staining localized to the cortex region. The peripheral, rim-like staining was also observed incontrol specimens and is thus unspecific (data not shown). Bar in low magnifications equals 10 pm, in high magnifications 1 pm.
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 95, 1991
stained, and the enzyme also began to appear in the differ-entiating vascular tissue (Fig. 4b).The staining pattern of young foliage leaves resembled that
of the cotyledon with myrosinase being present in the guardcells and parenchyma. In addition, stained cells appeared inthe vascular tissue close to the phloem (Fig. 5a). In olderleaves, however, the guard cells no longer contained myrosi-nase. In petals and siliques, the localization of myrosinasewas similar to that in the older leaves, i.e. scattered paren-chyma cells were stained in the mesophyll as well as in thevascular tissue (Fig. 5, b and c). In the stem, a major locationfor the enzyme was the xylem cells, although some cells inthe cortex also contained myrosinase (Fig. 6a). In the root themyrosin cells were mainly located in the cortex (Fig. 6b).
DISCUSSION
This paper presents the distribution of myrosinase studiedby immunohistochemical staining of tissue sections. In pre-liminary studies, a polyclonal antiserum (K505) and a mono-clonal antibody (3D7) were compared. Western blot analysesand immunoprecipitations had shown that these antibodiesreacted with the same myrosinase species (9). The two reagentsreacted in a similar way also when used in immunohisto-chemistry. Only a small number of cells were stained. SinceK505 in quantitative immunoprecipitations reacted withmore than 80% of all myrosinase activity present in a rapeseedprotein extract (9), it is likely that very little ifany myrosinaseis located outside these cells. These results are in completeagreement with those of a recent study in which myrosinasewas identified in Brassica napus seeds with a different mono-clonal antibody (16). In a previous study, myrosinase ap-peared to be expressed in the majority of rapeseed embryoniccells (12), a suggestion in conflict with that of the presentstudy. It seems reasonable that diffusion of the enzyme and/or the salts in the unfixed tissue of the earlier study mighthave caused an erroneous result.Approximately 2 to 5% of the cells in the mature seed
appeared to contain myrosinase. The first myrosinase-con-taining cells emerged at approximately d 20 after pollination.The number increased until approximately d 30. The moreintense staining of these cells during the later stages of seeddevelopment suggested that the cells continued to accumulatemyrosinase after this period. Indeed, when protein extractsfrom developing seeds were investigated for myrosinase byWestern analysis, increasing amounts of the enzyme wererecorded up to d 38 after pollination.The confinement of myrosinase to only a few cells, and to
different cell types in the various organs ofthe plant, indicatesthat a sophisticated regulatory mechanism governs the expres-sion of the myrosinase genes. It follows that the promoterregions of these genes will be interesting to investigate withrespect to the regulation of their cellular specificity.
In a comparative study on consecutive sections of seeds,we showed that Millon's reagent and the antibodies identifiedthe same set of cells. Thus, the immunocytochemical tech-nique is a reliable method for the identification of myrosincells. However, the two different techniques stain differentcompartments of the myrosin cells; the antibodies localize theenzyme in the cytoplasm, whereas Millon's reagent stained
the interior of the myrosin grains. The Millon's reagent reactswith tyrosine and tryptophan residues, i.e. compounds whichcontain an aromatic ring structure (7). Some of the glucosi-nolates, the methylindolyls, have structures similar to trypto-phan and, hence, are probably the reactants detected by theMillon's reagent. The localization of glucosinolates in theinterior of myrosin grains is also in agreement with earlierfindings that glucosinolates are present only in the myrosincells where they are stored in the myrosin grains/vacuoles (3).
There are many conflicting reports concerning the intracel-lular localization of myrosinase; in early studies mainly basedon morphological criteria, myrosinase was suggested to belocalized to the interior of the myrosin grains (13, 18, 19),while results of later reports suggested a cytoplasmic distri-bution, sometimes in association with membranes (6, 12).The latter alternative is in accordance with our findings, whichinclude an occasional association with the outside of themyrosin grains. The nature of the interaction of the enzymewith membranous structures is unclear, and elucidation ofthis question has to await further biochemical characterizationof the myrosinase molecule.The formation of a series of potentially toxic cleavage
products upon the hydrolysis of glucosinolates by myrosinasehas led to a proposal that the myrosinase-glucosinolate systemmight have a role as a nonspecific defense system in the plant(11). Our findings support this proposal, because we haveshown that there is specific expression of myrosinase in guardcells and in xylem, which are both prime locations for path-ogen invasion. It remains to be determined whether myrosi-nase synthesis might be induced in higher amounts by theaddition of different agents that damage plant tissue.
ACKNOWLEDGMENT
Ms. Maria Philgren is gratefully acknowledged for providing experttechnical assistance.
LITERATURE CITED
1. Dobberstein B, Garrof H, Warren G (1979) Cell-free synthesisand membrane insertion of mouse H-2Dd histocompatibilityantigen and 32-microglobulin. Cell 17: 759-769
2. Ettlinger MG, Kjaer A (1968) Sulfur compounds in plants.Recent Adv Phytochem 1: 59-144
3. Grob K, Matile P (1979) Vacuolar location of glucosinolates inhorseradish root cells. Plant Sci Lett 14: 327-335
4. Guignard L (1890) Recherches sur la localisation des principesactifs des crucifers. J Bot Paris 4: 385-394, 412-430, 435-455
5. Heinricher E (1884) Uber Eiweisstoffe fuhrende idioblasten beieinigen cruciferen. Ber Dtsch Bot Ges 2: 463-466
6. Iversen TH (1970) Cytochemical localization of myrosinase (f-thioglucosidase) in root cells of Sinapis alba L. Protoplasma71: 451-466
7. Jensen WA (1962) Botanical Histochemistry. WH Freeman andCo, San Francisco, pp 72-73, 228
8. Larsen PO (1981) Glucosinolates. In EE Conn, ed, The Biochem-istry of Plants, Vol 7. Academic Press, New York, pp 501-525
9. Lenman M, R6din J, Josefsson LG, Rask L (1991) Immunolog-ical characterization of rapeseed myrosinase. Eur J Biochem(in press)
10. Lonnerdahl B, Jansson JC (1973) Purification and characteriza-tion of a myrosinase from rapeseeds (Brassica napus L.).Biochim Biophys Acta 315: 421-429
11. Luthy B, Matile P (1984) The mustard oil bomb; Rectifiedanalysis of the subcellular organization of the myrosinase sys-tem. Biochem Physiol Pflanzen 179: 5-12
HOGLUND ET AL.220
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.

DISTRIBUTION OF MYROSINASE IN RAPESEED TISSUES
12. Maheshwari PN, Stanley DW, Beveridge TJ, van de Vort FR(1981) Localization of myrosinase (thioglucoside glucohydro-lase, EC 3.2.3.1) in cotelydon cells ofrapeseed. J Food Biochem5: 39-61
13. Rest JA, Vaughan JG (1972) The development of protein andoilbodies in the seed of Sinapis alba L. Planta 105: 245-262
14. Rodman JE (1981) Divergence, convergence, and parallelism inphytochemical characters: the glucosinolate-myrosinase sys-tem. In DA Young, DA Siegler, eds, Phytochemistry andAngiosperm Phylogeny. Praeger, New York, pp 43-79
15. Sharma M (1971) Ontogenic studies of the myrosin idioblasts inBrassica napus and Brassica montana. Bot Tidskrift 66: 51-59
16. Thangstad OP, Iversen TH, Slupphaug G, Bones A (1990) Im-munocytochemical localization of myrosinase in Brassica na-pus L. Planta 180: 245-248
17. Towbin HT, Staethlin T, Gordon J (1979) Electrophoretic trans-fer of proteins from polyacrylamide gels to nitrocellulosesheets: Procedure and some applications. Proc Natl Acad SciUSA 76: 4350-4354
18. Werker E, Vaughan JG (1974) Anatomical and ultrastructuralchanges in aleurone and myrosin cells of Sinapis alba duringgermination. Planta 116: 243-255
19. Werker E, Vaughan JG (1976) Ontogeny and distribution ofmyrosin cells in the shoot of Sinapis alba L. A light- andelectron microscope study. Israel J Bot 25: 140-151
221
www.plantphysiol.orgon April 12, 2019 - Published by Downloaded from Copyright © 1991 American Society of Plant Biologists. All rights reserved.


















