
Differential role of T regulatory and Th17 in Swiss mice infected with Plasmodium berghei ANKA and...
11
Differential role of T regulatory and Th17 in Swiss mice infected with Plasmodium berghei ANKA and Plasmodium yoelii Tarun Keswani, Arindam Bhattacharyya ⇑ Immunology Laboratory, Department of Zoology, University of Calcutta, 35, Ballygunge Circular Road, Kolkata 700019, West Bengal, India highlights Low levels of Treg and increased Th17 cells expression during lethal PbA infection. Increased Treg and decreased Th17 cells expression during P. yoelii infections. Increased expressions of TGF-b and IL-10 high during P. yoelii infection. Whereas, TNF-a, IL-6, IFN-c and IL-23 expression is high during PbA infection. Possible role of imbalance Treg cells and Th17 cells during malaria infection. graphical abstract article info Article history: Received 27 September 2013 Received in revised form 21 January 2014 Accepted 4 March 2014 Available online 24 March 2014 Keywords: Malaria Plasmodium berghei ANKA Plasmodium yoelii T regulatory cell Spleen Th17 cell abstract The outcome of malaria infection is determined, in part, by the balance of pro-inflammatory and regula- tory immune responses. Host immune responses in disease including malaria are finely regulated by the opposing effects of Th17 and T regulatory (Treg) cells. Here we have examined the role of Treg cells and Th17 cells during malaria infection and find that low levels of Treg cells possibly influence the outcome of infections with the lethal strain of Plasmodium berghei ANKA (PbA). In contrast, high level of Treg cells may influence the outcome of nonlethal Plasmodium yoelii NXL (P. yoelii) infections. We observed decreased expressions of key regulators of Treg inductions-TGF-b, CD4IL-2 and IL-10 during PbA infection, whereas their expression remains high during P. yoelii infection. On the other hand TNF-a, IL-6, IFN-c and IL-23 expression is high during PbA infection and lower during P. yoelii infection. Thus, results from this study suggest that the differential expression of Treg and Th17 might have a key role on host pathogenesis dur- ing malaria infection. The high level of IL-6 and low level of TGF-b may composite of the advantaged local microenvironment for the production of Th17 cells in the spleen of the PBA infected mice and vice verse during nonlethal P. yoelii. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction Malaria is now being appreciated in India as an emerging public health problem. Approximately 1,000,000 new disease cases are detected every year with inflammatory relation along with its masterful ability to avoid clearance by the innate and adaptive host immune responses. Failure to develop an effective balance of pro- inflammatory and regulatory immune responses can lead to unre- stricted parasite replication and severe immunopathology (Couper et al., 2008). Several groups of T cells, including Th1, Th2, NKT cells and Treg cells, are involved in early immune mechanisms control- ling Plasmodium infection (Chen et al., 2009; Rouse and Suvas, http://dx.doi.org/10.1016/j.exppara.2014.03.003 0014-4894/Ó 2014 Elsevier Inc. All rights reserved. ⇑ Corresponding author. Fax: +91 33 2461 4849. E-mail addresses: [email protected] (T. Keswani), [email protected] (A. Bhattacharyya). Experimental Parasitology 141 (2014) 82–92 Contents lists available at ScienceDirect Experimental Parasitology journal homepage: www.elsevier.com/locate/yexpr
Transcript of Differential role of T regulatory and Th17 in Swiss mice infected with Plasmodium berghei ANKA and...

Experimental Parasitology 141 (2014) 82–92
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Differential role of T regulatory and Th17 in Swiss mice infectedwith Plasmodium berghei ANKA and Plasmodium yoelii
http://dx.doi.org/10.1016/j.exppara.2014.03.0030014-4894/� 2014 Elsevier Inc. All rights reserved.
⇑ Corresponding author. Fax: +91 33 2461 4849.E-mail addresses: [email protected] (T. Keswani), [email protected]
(A. Bhattacharyya).
Tarun Keswani, Arindam Bhattacharyya ⇑Immunology Laboratory, Department of Zoology, University of Calcutta, 35, Ballygunge Circular Road, Kolkata 700019, West Bengal, India
h i g h l i g h t s
� Low levels of Treg and increased Th17cells expression during lethal PbAinfection.� Increased Treg and decreased Th17
cells expression during P. yoeliiinfections.� Increased expressions of TGF-b and
IL-10 high during P. yoelii infection.� Whereas, TNF-a, IL-6, IFN-c and IL-23
expression is high during PbAinfection.� Possible role of imbalance Treg cells
and Th17 cells during malariainfection.
g r a p h i c a l a b s t r a c t
a r t i c l e i n f o
Article history:Received 27 September 2013Received in revised form 21 January 2014Accepted 4 March 2014Available online 24 March 2014
Keywords:MalariaPlasmodium berghei ANKAPlasmodium yoeliiT regulatory cellSpleenTh17 cell
a b s t r a c t
The outcome of malaria infection is determined, in part, by the balance of pro-inflammatory and regula-tory immune responses. Host immune responses in disease including malaria are finely regulated by theopposing effects of Th17 and T regulatory (Treg) cells. Here we have examined the role of Treg cells andTh17 cells during malaria infection and find that low levels of Treg cells possibly influence the outcome ofinfections with the lethal strain of Plasmodium berghei ANKA (PbA). In contrast, high level of Treg cells mayinfluence the outcome of nonlethal Plasmodium yoelii NXL (P. yoelii) infections. We observed decreasedexpressions of key regulators of Treg inductions-TGF-b, CD4IL-2 and IL-10 during PbA infection, whereastheir expression remains high during P. yoelii infection. On the other hand TNF-a, IL-6, IFN-c and IL-23expression is high during PbA infection and lower during P. yoelii infection. Thus, results from this studysuggest that the differential expression of Treg and Th17 might have a key role on host pathogenesis dur-ing malaria infection. The high level of IL-6 and low level of TGF-b may composite of the advantaged localmicroenvironment for the production of Th17 cells in the spleen of the PBA infected mice and vice verseduring nonlethal P. yoelii.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
Malaria is now being appreciated in India as an emerging publichealth problem. Approximately 1,000,000 new disease cases are
detected every year with inflammatory relation along with itsmasterful ability to avoid clearance by the innate and adaptive hostimmune responses. Failure to develop an effective balance of pro-inflammatory and regulatory immune responses can lead to unre-stricted parasite replication and severe immunopathology (Couperet al., 2008). Several groups of T cells, including Th1, Th2, NKT cellsand Treg cells, are involved in early immune mechanisms control-ling Plasmodium infection (Chen et al., 2009; Rouse and Suvas,

T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92 83
2004). Tregs have been reported to contribute to immune evasionduring malaria infection in mice and humans, suggesting that acti-vating Tregs is one of the mechanisms by which malaria parasitessubvert host immune systems (Todryk et al., 2008; Walther et al.,2009). However, these studies provided limited mechanistic in-sight into the role of Treg cells in severe malarial disease. Numbersof studies have also showed that Treg cells may facilitate parasiteclearance (Chen et al., 2009; Feng et al., 2012), limit malaria-in-duced immune pathology (Haque et al., 2010), enhance parasiteburden (Cambos et al., 2008) and help parasite to escape host im-mune defense (Hisaeda et al., 2004). These observations suggest apossibility that Treg cells are able to control different types of theimmune response by tailoring their suppressive function to a par-ticular inflammatory environment.
Recently however, specific depletion of FoxP3+ Treg cells didnot protect against experimental cerebral malaria (ECM), bringinginto question the role for these cells in mediating disease (Steeget al., 2009). Although given that ECM is mediated by pathogenicT cells that promote parasite tissue sequestration (Amante et al.,2010), we hypothesized that under certain conditions, Treg cellscan suppress deleterious T cell responses and protect againstECM. Studies in an experimental model of ECM caused by infectionof C57BL/6 mice with Plasmodium berghei ANKA (PbA) have high-lighted the important role played by various immune cells indisease pathogenesis, including CD4+ T cells, CD8+ T cells, conven-tional dendritic cells and Natural Killer (NK) cells (deWalick et al.,2007; Hansen et al., 2007, 2005, 2003; Nitcheu et al., 2003;Belnoue et al., 2002; Yanez et al., 1996). Until recently, the delete-rious role proposed for Treg cells in studies of human malaria hasbeen difficult to test in mice, due to the lack of appropriate re-agents (Amante et al., 2007; Vigario et al., 2007). Likewise ablationof Treg cells in adult healthy mice leads to augmented generationof Th1, Th2, and Th17 cells and death within 2 weeks from highlyaggressive inflammatory lesions in a variety of organs (Kim et al.,2007).
Furthermore, the shared requirement for TGF-b in the develop-ment Tregs and Th17 cells has provided an elegant solution forlinking the potentially pathogenic Th17 pathway with a potentcounter-regulatory pathway that can control it (Veldhoen et al.,2006). In the absence of IL-6, TGF-b promotes the differentiationof Tregs. Interestingly, however, naïve T cells stimulated withTGF-b alone were found to upregulate both Foxp3 and RORct,although they failed to express an appreciable level of IL17 andprogressively extinguished RORct as they differentiated into Tregcells (Yang et al., 2008; Zhou et al., 2007). Similarly, early Th17 dif-ferentiation induced by TGF-b and IL-6 was accompanied by tran-sient co-expression of RORct and Foxp3, with Foxp3 extinguishedas Th17 development progressed (Zhou et al., 2008; Yang et al.,2008). High doses of TGF-b repressed RORct function via increasedFoxp3; but at lower doses, TGF-b cooperated with signals initiatedby IL-6 to overcome Foxp3-mediated repression of RORct. IL-23 isstrongly associated with activation of APC and NK cells and alsowith development of Th17 that is capable BBB disruption (Sunet al., 2004). During infection, these cells activate and cooperatewith each other to allow T cell migration into brain. This resultsin blood–brain barrier (BBB) disruption and is a key pathologicalfeature of ECM (Ishida et al., 2010). We questioned whether thebalance between anti-inflammatory Tregs and pro-inflammatoryTh17 cells may play a role in the persistence of Plasmodium inthe spleen.
In our previous work we have evaluated the outcome of TGF-band TNF-a production in context of splenocyte apoptosis duringlethal (PbA) infection (Keswani and Bhattacharyya, 2013). Wehypothesize that TGF-b and IL-6 might play an important role inmodulating the balance between the differentiation of Treg andTh17 cells in malaria.
2. Materials and methods
2.1. Reagents
Giemsa’s azur eosin methylene blue solution (#109204), hema-toxylin and eosin stains for microscopy were purchased from MerckChemicals (Mumbai, India). A phosphate buffer saline (PBS), sodiumbicarbonate, sodium azide, RNase A, NBT and BCIP (Cat# RM 578,RM 2577) were procured from Himedia, Mumbai, India. Fetal Bo-vine Serum from Axxora and Invitrogen, Darmstadt, Germany. Anti-bodies against TNF-a (#3707), TGF-b (#3711), NFAT (#4389), Stat-3(#4904) and b-actin (#4967), AP-linked anti-rabbit (#7054), AP-linked anti-mouse (#7056) secondary antibodies were purchasedfrom the Cell signaling Technology (Cell Signaling Technology,Inc, Denver, MA, USA). Anti-IL6 antibody (#ab6672) procured fromabcam. Pre stain molecular weight protein marker, goat anti-mouse-HRP and anti-rabbit-HRP conjugated antibody and TMBdeveloping reagents for enzyme-linked immunoabsorbant assay(ELISA) and DAB developing system for immunohistochemistrywere purchased from Bangalore Genei (Bangalore, India). Perme-abilization buffer (#421002), Fixation buffer (#420801), PE-anti-CD25 (#102007), Alexa Fluor-anti-Foxp3 (#320013), PE Anti-IL-2(#503807), purified anti-CD16/32 (#101301), PE Anti-IL-17(#506903), purified anti mouse-IL-17 (#506905), purified Foxp3(#320001), purified IL-23 (#513801) and purified IL-6 (#504505)were purchased from Bio legend (New Delhi, India). Isotype con-trols for FoxP3: Alexa Fluor� 647 Mouse IgG1j (#400130), CD25:PE Rat IgG1k (#401905) IL-2: PE Rat IgG2b j (#400607), andIL17: PE Rat IgG1j (400407) were also procured from Biolegend.PE anti mouse CD-8(H35-17.2) and FITC anti-mouse CD4 (RM4-4)procured from BD Biosciences (San Jose, CA, USA). APC Anti–mouseRORct (#17-6988-80) and respective APC Rat IgG2aj Isotype Con-trol (#17-4321) were procured from e bioscience, Inc.g, (San Diego,CA, USA). All solutions were prepared with commercial reagents ofat least pro-analysis quality and with sterilized 18 MX milliQwater. When necessary, the specific origins of reagents are listedin the text.
2.2. In vivo malaria infection
Male Swiss albino mice (�25 g each; aged 6–8 weeks) weremaintained in sterilized cages and absorbent media; food andwater were provided ad libitum. Animal experiments were car-ried out as per the guidelines of the Committee for the Purposeof Control and Supervision of Experimental Animals (CPCSEA),Government of India (Registration No. 885/ac/05/ CPCSEA) andas approved by the Institutional Animal Ethics Committee (IAEC)University of Calcutta, and conforms with the Guide for theCare and Use of Laboratory Animals published by the US Na-tional Institutes of Health (NIH Publication No. 85-23, revised1985).
Parasite strain Plasmodium berghei ANKA and Plasmodium yoelii17XNL were obtained from National Institute Malaria ResearchCenter, New Delhi. Parasitized mouse red blood cells (pRBC) froma liquid N2 preserved stabilize were injected into mice of the samebackground. After amplification, mice were infected with PbA andP. yoelii, respectively with 1 � 106 pRBC, in 100 ll of phosphatebuffer solution (PBS) by intraperitoneal injection. Control mice re-ceived an equal number of uninfected erythrocytes. Parasitemiawas monitored daily in all experimental groups by Giemsa-stainedthin blood smears made from tail snips. Survivability and weightchange of mice (n = 60) were also observed daily. The percentageof parasitemia was calculated as follows: Parasitemia (%) = ((num-ber of infected erythrocytes)/(total number of erythrocytescounted)) � 100.

84 T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92
2.3. Immunohistochemical analysis of Foxp3 and IL-17
Sections (5 lm) were cut from paraffin embedded spleen tissuesand mounted on positively charged super frost slides (Export Men-gel CF). Tissues were deparaffinized, rehydrated through gradedalcohols, and then blocked for endogenous peroxidase in 3% hydro-gen peroxide in methanol. All tissues were pre blocked in Tris-buffered saline containing 0.3% Triton, and 0.5% blocking agent(BSA, SRL, India, and batch No. 832095) and incubated with Foxp3and IL-17 primary antibody (1:30) overnight at 4 �C for positive con-trol. Antisera specific for Foxp3 and IL-17 were diluted 1:30 in Tris-buffered saline containing 0.3% Triton, and 0.5% blocking agent.Immunoreactive complexes were detected using DAB system (Ban-galore GeNei DAB system, Cat #SFE5). Slides were counterstainedbriefly in haematoxylin (MERCK), mounted in DPX (MERCK).
2.4. Spleen cell culture
Spleen was aseptically removed from each mouse and pressedthrough a sterile fine-wire mesh with 10 ml of RPMI-1640 (LifeTechnologies) supplemented with 5% heat-inactivated fetal calfserum (FCS; Hyclone Laboratories, Inc.), 25 mM Hepes (Life Tech-nologies), 0.12% gentamicin (Schering), and 2 mM glutamine (LifeTechnologies). Cell suspensions were centrifuged at 350g for10 min at room temperature (RT). Erythrocytes were lysed withcold 0.17 M NH4Cl and the cells were washed twice with freshmedium. The viability of the spleen cells was determined by trypanblue exclusion and was always N90%. Spleen cells were adjusted toa final concentration of 107 cells/ml in RPMI-1640 supplementedwith 10% heat-inactivated FCS. Aliquots (500 ll/well) of the cellsuspension were incubated in 24-well flat bottom culture plates(FALCON) in triplicate for 48 h at 37 �C in a humidified 5% CO2
incubator. At different dpi cells were stimulated with PMA and ion-omycin for 2 h at 37 �C and then Golgi Stop (BD Bioscience) wasadded to each reaction (1:500 (v/v)). After continued co-culturefor 4 h at 37 �C, cells were washed with 3% FBS and resuspendedin 100 ll of 3% FBS and 0.1% NaN3(CSB). The 24-well plates werethen centrifuged at 350g for 10 min at RT, and the supernatantswere collected and stored at �80 �C until assayed for cytokinelevels.
2.5. Flow cytometric analyses of CD4+ and CD8+ T lymphocytepopulation in spleen
Aliquots (each containing 5 � 106 cells/ml) of isolated spleno-cytes were then removed, centrifuged, and suspended in 1 mlPBS. The T-lymphocyte phenotyping was conducted to analyzethe effect of PbA and P. yoelii on CD4 and CD8 surface molecules.Positioning of quadrants on FITC/PE dot plots was performed andCD4+, CD8+, CD4+CD8+ (Double positive; DP) and CD4�, CD8� (Dou-ble negative; DN) subpopulations were distinguished. Briefly, afterincubation time (30 min), the harvested cells were suspended in1 ml cell staining buffer. An aliquot of 100 ll was incubated with5 ll FITC-conjugated anti-CD4 monoclonal antibody and 5 ll PE-conjugated anti-CD8 monoclonal antibody for 30 min in dark at37 �C, after which 400 ll PBS was added to each sample. The FITCand PE fluorescence were measured through FL-1 filter (530 nm)and FL-2 filter (585 nm), respectively, and 10,000 events were ac-quired. The flow cytometric (FACS Aria III; Becton Dickinson) datawas analyzed using FACS diva software (Becton Dickinson).
2.6. FACS analysis of CD4+CD25+ Foxp3+, CD4+IL-17+ RORct+ andCD4+Il-2+ expression
Spleen cells collected from mice at different time points afterPbA and P. yoelii infection were previously stimulated with PMA
and ionomycin for 2 h at 37 �C and then Golgi Stop (BD Bioscience)was added to each reaction (1:500 (v/v)). After continued co-cul-ture for 4 h at 37 �C, cells were washed with (CSB). For surfacestaining FITC-anti-CD4 and PE-anti-CD25 (Biolegend) were addedand incubated for 30 min at 4 �C. These cells were then incubatedwith anti-CD16/32 (Fc-block) and stained with surface markers,followed by fixation with 4% paraformaldehyde and permeabiliza-tion with 0.1% saponin. The cells were then fixed washed twicewith Cell staining buffer and intracytoplasmic staining was per-formed using APC-conjugated anti-Foxp3 for Treg cells and PE-IL-17, APC-RORct for Th17 cells and PE-IL2, respectively along withrespective isotype controls. The cells were then analyzed in a FACSAria III with FACS diva software (BD bioscience). Viable cells weregated by forward and side scattering.
2.7. Western blot analysis
To perform Western blot analysis of TGF-b, TNF-a, Stat-3, IL-6,Foxp3 and NFAT expression, cell lysates (Mitra et al., 2012) fromeach spleen were loaded into dedicated wells in a 10–12% poly-acrylamide gel. After resolution of the sample contents, the gel pro-teins were transferred to a nitrocellulose membrane and the latterblocked for 30 min at 4 �C with non-fat dry milk in TBS containing0.1% Tween-20. Each primary antibody was diluted to 1:1000 in 5%BSA and then applied to the membrane. After overnight incubationat 4 �C, the membrane was rinsed free of unbound primary anti-body and again blocked with non-fat dry milk in TBS/Tween-20.Secondary antibodies were then diluted to 1:1000 ratios in 5%BSA and applied to the membrane. After 2 h incubation at 4 �C, un-bound antibody was rinsed away and the membrane developedusing NBT/BCIP (nitroblue tetrazolium chloride/5-bromo-4-chloro-3-indolyl-phosphate; Hi-Media, Mumbai, India). b-Actinwas also analyzed on each membrane for confirmation of gel sam-ple loading (i.e., based on constitutive expression).
2.8. Cytokine analysis
IFN-c, TNF-a, IL-6, IL-10, and IL-17 levels in splenocyte superna-tants were measured using commercial enzyme-linked immuno-sorbent assay (ELISA) kits according to the manufacturer’sprotocol (R&D Systems, Minneapolis, MN, USA). The OD valueswere read in a microplate reader at 450 nm. The concentration ofIFN-c, TNF-a, IL-6, IL-10, and IL-17 in samples was calculatedagainst the standard curve generated using recombinant cytokines.
2.9. RNA isolation, reverse transcription, and RT-PCR
After treatment, total RNA extraction was performed using Tri-Zol reagent (Bangalore Genei, India) followed by DNase treatment,phenol–chloroform extraction, and ethanol precipitation. RNA pur-ity was checked by spectrophotometry, next, 1 lg of RNA was tran-scribed using the reverse transcription system (MMLV HighPerformance Reverse Transcriptage, Epicentre Biotechnologies)(Random hexmer – Promega # C1181). GAPDH was used as consti-tutive expression:
TGF-b; Forward primer: ACTGCTATGCTGCCTGCTCTTACT,Reverse primer: TGGCCTTGTAGACACCTTGGTCTT.
IL-6; Forward primer: CACAGCAGTGGCATCGG-GGG,Reverse primer: CTGGCCGCTCCTCCGAGTCT30.
IL-10 Forward primer: ACTGC –TATGCTGCCTGCTCTTACT,Reverse primer: TGGCCTTGTAGACACCTTGGTCTT.
Foxp3; Forward primer: GCAACAGCACTGGAACCTTCACAA,Reverse primer: AGTTG CTGCTTTAGGTGGAGTGGA.
GAPDH; Forward primer: CCATGGAGAAGGCTGGGG,Reverse primer: CAAAGTTGTCATGGATGAC.

T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92 85
2.10. Statistical analysis
Values between groups on same or different dpi were analyzedusing one or two way ANOVA. All values are shown as mean ± SEM,except where otherwise indicated. Data were analyzed and whenappropriate, significance of the differences between mean valueswas determined using Student’s test. Survival curve analyses wereperformed using the Graph Pad Prism software version 3.03. Signif-icant differences between survivals were assessed by Log rank test.p values < 0.05 were considered significant for all statistical analy-sis, otherwise stated.
Fig. 1. (a) Parasitemia, (b) survival rate and (c) weight change of Swiss albino mice afterwith 1 � 106 infected RBCs intraperitonially obtained from a homologous passage mouGiemsa-stained thin smear was made daily as shown in (a). Values represent the meaexperiments. (d) Representative immunohisto-chemistry of Foxp3+ cells in the 8 dpi splespleen tissue sections. (e) Representative immunohisto-chemistry of IL-17+ cells in theexpression in spleen tissue sections. Magnification = 40�. (For interpretation of the referarticle.)
3. Results
3.1. Characteristics of PbA and P. yoelii infection in Swiss albino mice
The clinical course of infection in the experimental mice is sum-marized in Fig. 1. Mice age matched and similar body weight wereinfected with 1 � 106 PbA and P. yoelii infected erythrocytes, respec-tively and the infection was monitored daily. Parasitemia rose grad-ually till 3 dpi in both PbA infected and P. yoelii infected set. CM wasinduced in mice after injection of virulent PbA blood-stage parasites.The mice died after paralysis and coma ensued between days 8 and
infection with P. berghei ANKA and Plasmodium yoelii. Mice were separately infectedse that had been infected with PbA and P. yoelii. For monitoring of parasitemia, a
n with SEM (n = 5 mice per group). Results are representative of five independenten of PbA infected and uninfected mice, red arrows represent expression of Foxp3 in8 dpi spleen of P. yoelii infected and uninfected mice, green arrows represent IL-17ences to colour in this figure legend, the reader is referred to the web version of this
86 T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92
12 after infectious challenge, with parasitemia (�10–20%). In con-trary P. yoelii infected mice developed high parasitemia but all micesurvived until natural clearances of parasites (Fig. 1a). During PbAinfection parasitemia at the point of mean median survival wasaround 4–9 dpi and most of the mice succumbed to CM during thisperiod, whereas all mice survived after P. yoelii infection (Fig. 1b).Significant differences between weights changes were observedduring PbA infection compared to P. yoelii infection and respectivecontrol mice (Fig. 1c). Statistically significant differences in thedevelopment of parasitemia, percentage of survival and weightchange was observed (Fig. 1a–c). Further the immuno-histochemi-cal staining of Foxp3 for Treg and IL-17 for Th17 cells is documentedin the nucleus in the spleen of 8 dpi mice infected with PbA and P.yoelii along with respective control mice. Focusing in T effector cellrelevant distribution, Foxp3 cells were predominantly distributedin the spleen tissue in P. yoelii infected mice (Fig. 1d and c), followedby lower expression of Foxp3+ cells during PbA infection (Fig. 1d anda) with respect to control (Fig. 1d, a and b). The PbA infected miceacquired an increased accumulation of IL-17+ cells in the spleen tis-sue (Fig. 1e and b), while IL-17+ cells expression was lower in thespleen of P. yoelii mice (Fig. 1e and d) with respect to control(Fig. 1e, a and b).
3.2. Alterations in the frequencies of CD4+ and CD8+ T cells in thespleen during blood-stage PbA and P. yoelii infection
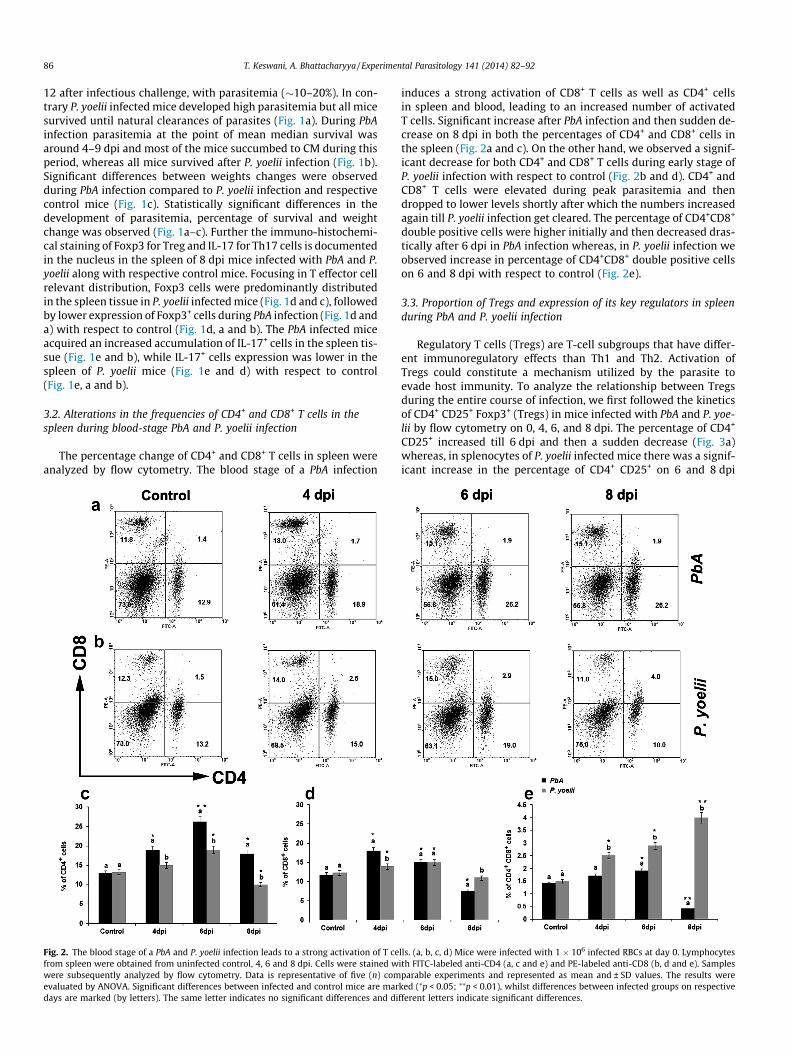
The percentage change of CD4+ and CD8+ T cells in spleen wereanalyzed by flow cytometry. The blood stage of a PbA infection
Fig. 2. The blood stage of a PbA and P. yoelii infection leads to a strong activation of T cefrom spleen were obtained from uninfected control, 4, 6 and 8 dpi. Cells were stained wwere subsequently analyzed by flow cytometry. Data is representative of five (n) comevaluated by ANOVA. Significant differences between infected and control mice are mardays are marked (by letters). The same letter indicates no significant differences and di
induces a strong activation of CD8+ T cells as well as CD4+ cellsin spleen and blood, leading to an increased number of activatedT cells. Significant increase after PbA infection and then sudden de-crease on 8 dpi in both the percentages of CD4+ and CD8+ cells inthe spleen (Fig. 2a and c). On the other hand, we observed a signif-icant decrease for both CD4+ and CD8+ T cells during early stage ofP. yoelii infection with respect to control (Fig. 2b and d). CD4+ andCD8+ T cells were elevated during peak parasitemia and thendropped to lower levels shortly after which the numbers increasedagain till P. yoelii infection get cleared. The percentage of CD4+CD8+
double positive cells were higher initially and then decreased dras-tically after 6 dpi in PbA infection whereas, in P. yoelii infection weobserved increase in percentage of CD4+CD8+ double positive cellson 6 and 8 dpi with respect to control (Fig. 2e).
3.3. Proportion of Tregs and expression of its key regulators in spleenduring PbA and P. yoelii infection
Regulatory T cells (Tregs) are T-cell subgroups that have differ-ent immunoregulatory effects than Th1 and Th2. Activation ofTregs could constitute a mechanism utilized by the parasite toevade host immunity. To analyze the relationship between Tregsduring the entire course of infection, we first followed the kineticsof CD4+ CD25+ Foxp3+ (Tregs) in mice infected with PbA and P. yoe-lii by flow cytometry on 0, 4, 6, and 8 dpi. The percentage of CD4+
CD25+ increased till 6 dpi and then a sudden decrease (Fig. 3a)whereas, in splenocytes of P. yoelii infected mice there was a signif-icant increase in the percentage of CD4+ CD25+ on 6 and 8 dpi
lls. (a, b, c, d) Mice were infected with 1 � 106 infected RBCs at day 0. Lymphocytesith FITC-labeled anti-CD4 (a, c and e) and PE-labeled anti-CD8 (b, d and e). Samplesparable experiments and represented as mean and ± SD values. The results were
ked (⁄p < 0.05; ⁄⁄p < 0.01), whilst differences between infected groups on respectivefferent letters indicate significant differences.

T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92 87
(Fig. 3b) w.r.t control mice. During PbA infection CD4+ Foxp3+ washigh at 4 dpi followed by a decrease in percentage on 6 and 8 dpi(Fig. 3a). In contrary CD4+ Foxp3+ T cells increased considerablyafter 4 dpi in P. yoelii infection (Fig. 3b). Tregs in the spleen CD4+
T-cell population increased after infection, with peaks on 4 dpiand gradually declined from 5 to 8 dpi during PbA infection(Fig. 3a and c). In addition, the percentage and of Tregs in P. yoeliiinfected mice were significantly lower than PbA on 4 dpi and afterthat a gradual increase till the mice recovered from infection(Fig. 3a–c).
Next we observed the expression pattern of TGF-b in both PbAand P. yoelii infected mice. During PbA infection we observeddownregulation of TGF-b expression with respect to control,(Fig. 3d and e) whereas, upregulation of TGF-b in P. yoelii infection(Fig. 3d and f). We also observed downregulation of NFAT after
Fig. 3. Proportions of Treg in CD4+ T cells. (a, b, c) Flowcytometric analysis demonstratedMice were infected with 1 � 106 infected RBCs. At day 4, 6 and 8 dpi, along with controlstained intracellularly for Foxp3 using allophycocyanin-labeled anti-Foxp3. Quadrant mMouse IgG1k (FoxP3) isotype controls on FITC CD4 gated on splenocytes, respectively ananalysis of Treg cells. Representative dot plots (upper) and column diagram (lower) of Foxc) PbA and (b, c) P. yoelii infected mice, as indicated, were shown at various time pointslysate from control 4, 6 and 8 dpi mice infected with PbA and P. yoelii were subjected to WTGF-b, anti-NFAT, and anti-FOXP3 antibodies. Bars represent quantitative densitometriccontrol. Data shown is representative of five (n) comparable experiments. Data is represeThe results were evaluated by ANOVA. Significant differences between infected and corespective days are marked (by letters). The same letter indicates no significant differen
initial PbA infection (Fig. 3d and e) whereas, upregulation of NFATduring P. yoelii infection (Fig. 3d and f). We further observed theexpression of Foxp3 during the course of malaria infection. Inter-estingly, supporting our previous results we observed upregulationof Foxp3 by Western blot during initial stage and sudden decreaseon 6 and 8 dpi in lethal PbA infection (Fig. 3d and e) whereas, de-layed upregulation during resolving P. yoelii infection, suggestingprotective role of TGF-b induced Foxp3 (Fig. 3d and f).
3.4. Proportion of Th17 and expression of its key regulators in spleenduring PbA and P. yoelii infection
To further analyze the relationship between Th-17 cells duringthe entire course of infection, we first followed the kinetics of CD4+
IL-17+ RORct+ (Tregs) in mice infected with PbA and P. yoelii by
Tregs in CD4+ T cell population in control, (a, c) PbA and (b, c) P. yoelii infected mice.spleen cells were stained with FITC-labeled anti-CD4 and PE-labeled anti-CD25, andarkers were set based on staining with PE Rat IgG1 k (CD25) and Alexa Fluor� 647d Alexa Fluor� 647 Mouse IgG1k isotype control gated on the CD4+ population forp3 and CD25 expression on splenocytes gated on CD4+ T cells from control mice, (a,(days). PbA and P. yoelii induced expression of TGF-b, NFAT and FOXP3. Spleen cell
estern blot analysis. The cell lysates were analyzed by immunoblotting with, anti-values of the expressed proteins, respectively (d, e and f). b-Actin used as loading
ntative of five (n) comparable experiments and represented as mean and ±SD values.ntrol mice are marked (⁄p < 0.05), whilst differences between infected groups onces and different letters indicate significant differences.
88 T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92
flow cytometry on days 0, 4, 6, and 8 dpi. The percentage of CD4+
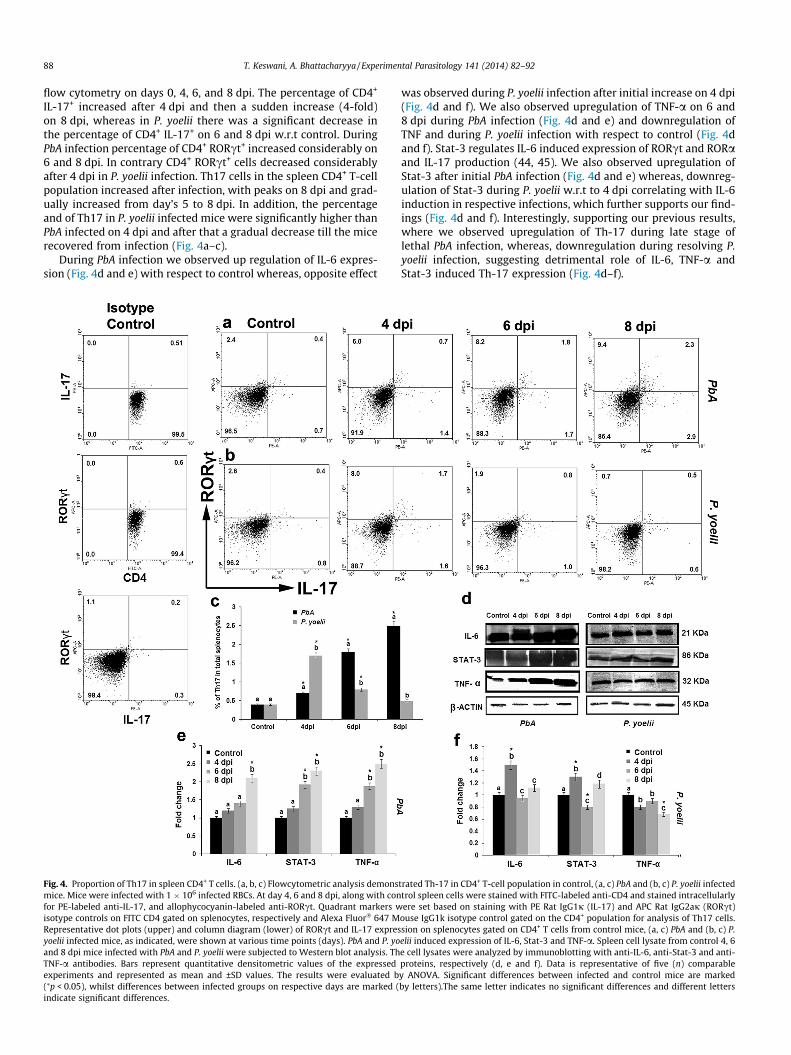
IL-17+ increased after 4 dpi and then a sudden increase (4-fold)on 8 dpi, whereas in P. yoelii there was a significant decrease inthe percentage of CD4+ IL-17+ on 6 and 8 dpi w.r.t control. DuringPbA infection percentage of CD4+ RORct+ increased considerably on6 and 8 dpi. In contrary CD4+ RORct+ cells decreased considerablyafter 4 dpi in P. yoelii infection. Th17 cells in the spleen CD4+ T-cellpopulation increased after infection, with peaks on 8 dpi and grad-ually increased from day’s 5 to 8 dpi. In addition, the percentageand of Th17 in P. yoelii infected mice were significantly higher thanPbA infected on 4 dpi and after that a gradual decrease till the micerecovered from infection (Fig. 4a–c).
During PbA infection we observed up regulation of IL-6 expres-sion (Fig. 4d and e) with respect to control whereas, opposite effect
Fig. 4. Proportion of Th17 in spleen CD4+ T cells. (a, b, c) Flowcytometric analysis demonsmice. Mice were infected with 1 � 106 infected RBCs. At day 4, 6 and 8 dpi, along with confor PE-labeled anti-IL-17, and allophycocyanin-labeled anti-RORct. Quadrant markers wisotype controls on FITC CD4 gated on splenocytes, respectively and Alexa Fluor� 647 MRepresentative dot plots (upper) and column diagram (lower) of RORct and IL-17 expresyoelii infected mice, as indicated, were shown at various time points (days). PbA and P. yoand 8 dpi mice infected with PbA and P. yoelii were subjected to Western blot analysis. ThTNF-a antibodies. Bars represent quantitative densitometric values of the expressedexperiments and represented as mean and ±SD values. The results were evaluated b(⁄p < 0.05), whilst differences between infected groups on respective days are marked (indicate significant differences.
was observed during P. yoelii infection after initial increase on 4 dpi(Fig. 4d and f). We also observed upregulation of TNF-a on 6 and8 dpi during PbA infection (Fig. 4d and e) and downregulation ofTNF and during P. yoelii infection with respect to control (Fig. 4dand f). Stat-3 regulates IL-6 induced expression of RORct and RORaand IL-17 production (44, 45). We also observed upregulation ofStat-3 after initial PbA infection (Fig. 4d and e) whereas, downreg-ulation of Stat-3 during P. yoelii w.r.t to 4 dpi correlating with IL-6induction in respective infections, which further supports our find-ings (Fig. 4d and f). Interestingly, supporting our previous results,where we observed upregulation of Th-17 during late stage oflethal PbA infection, whereas, downregulation during resolving P.yoelii infection, suggesting detrimental role of IL-6, TNF-a andStat-3 induced Th-17 expression (Fig. 4d–f).
trated Th-17 in CD4+ T-cell population in control, (a, c) PbA and (b, c) P. yoelii infectedtrol spleen cells were stained with FITC-labeled anti-CD4 and stained intracellularlyere set based on staining with PE Rat IgG1j (IL-17) and APC Rat IgG2aj (RORct)
ouse IgG1k isotype control gated on the CD4+ population for analysis of Th17 cells.sion on splenocytes gated on CD4+ T cells from control mice, (a, c) PbA and (b, c) P.elii induced expression of IL-6, Stat-3 and TNF-a. Spleen cell lysate from control 4, 6e cell lysates were analyzed by immunoblotting with anti-IL-6, anti-Stat-3 and anti-proteins, respectively (d, e and f). Data is representative of five (n) comparabley ANOVA. Significant differences between infected and control mice are markedby letters).The same letter indicates no significant differences and different letters

T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92 89
3.5. Expression of activation marker IL-2 by splenic CD4+ T cells and IL-6, IL-10, TGF-b and FoxP3 mRNA levels during malaria infection
The cytokine IL-2 is produced by T cells early after activationand has been shown to inhibit the generation of Th17 cells. Thepercentages of IL-2 secreting CD4+ T cells progressively increasedduring initial PbA infection and then decreased till the mice dies(Fig. 5a and c). Interestingly in P. yoelii infection CD4IL-2 expres-sion decrease after initial infection, then as the infection pro-gresses the percentage of CD4 IL2 increase till the infectionresolves (Fig. 5b and c). As expected, when Foxp3+ Treg cellfrequencies were low then IL-2 expression was also low confirm-ing the known importance of IL-2 for Treg cell homeostasis(Fig. 5a–c). The partial restoration of IL-17 production when IL-2was low showed that regulation of IL-2 is one of the previously
Fig. 5. IL-2 secretion was analyzed in Fitc-CD4+ T cells from total splenocytes obtained fmice (a, b and c). Quadrant markers were set based on staining with PE Rat IgG2b j isotypuninfected control mice and mice infected with 1 � 106 infected RBC of PbA and P. yoelii,on control, 4, 6 and 8 dpi. The RNA samples were obtained from five mice per time poinagarose gels containing 0.5 lg of ethidium bromide per ml and observed with a UV trproteins, respectively (d, e and f). Molecular size marker (100-bp ladder) was used. Data±SD values. The results were evaluated by ANOVA. Significant differences between infegroups on respective days are marked (by letters).The same letter indicates no significa
unappreciated mechanisms of action of Foxp3+ Treg cells in pro-moting Th17 cells in vivo.
The levels of IL-6, IL-10, TGF-b and FoxP3 mRNAs were inducedor elevated in the spleen during the course of infection. IL-6 andFoxP3 mRNAs were detectable in uninfected controls. CytokinemRNAs for IL-10, and TGF-b were not detectable in uninfected con-trols. During PbA infection IL-6 mRNA level expression increasedon 6 and 8 dpi with respect to control, and in P. yoelii IL-6 mRNAlevel decrease on 6 and 8 dpi, with increase on 4 dpi. During initialstage of PbA infection increase in TGF-b and IL-10 mRNA expres-sion was observed, which then subsequently downregulated. Incontrary, TGF-b and IL-10 mrna expression increased after 4 dpiwith respect to control mice during P. yoelii infection. Supportingour previous observations, we observed increased Foxp3 mRNA le-vel during initial stage of lethal PbA infection whereas, in P. yoelii
rom mice on days 4, 6, and 8 dpi spleen during PbA and P. yoelii along with controle controls. Temporal expression of IL-6, IL-10, TGF-b and FoxP3 mRNAs in spleens of
respectively. Total RNA was extracted from each spleen from all experimental groupt. RT-PCR was performed, and the PCR products were electrophoresed on 1.5–2.5%ansilluminator. Bars represent quantitative densitometric values of the expressedis representative of five (n) comparable experiments and represented as mean andcted and control mice are marked (⁄p < 0.05), whilst differences between infected
nt differences and different letters indicate significant differences.

90 T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92
infection Foxp3 mRNA expression increased after the initial stageof infection (Fig. 5d–f).
3.6. Dynamics of pro and anti inflammatory cytokines in splenocytesduring PbA and P. yoelii infection
To evaluate the relationship between the levels of pro and antiinflammatory cytokines and the outcomes of mice infected withPbA and P. yoelii, pro-inflammatory cytokines (IL-23, TNF-a, IL-6,and IL-17) an anti-inflammatory cytokine (TGF-b and IL-10) inthe supernatants of cultured splenocytes were assayed by ELISA.The proinflammatory cytokines IL-23, TNF-a, IL-6 and IL-17 beganto increase after infection with a peak on day 4 dpi and continuedto increase during the course of PbA infection. During P. yoelii infec-tion the same proinflammatory cytokines IL-23, TNF-a, IL-6, andIL-17 showed profound increased on 4 dpi followed by significantdecrease on 6 and 8 dpi. The anti-inflammatory cytokine IL-10 alsoreached a peak on day 5 dpi following a significant decrease,whereas TGF-b level decrease continuously with respect to control
Fig. 6. Concentration of pro and anti inflammatory cytokines in the splenocytes of mice ifive (n) comparable experiments and represented as mean and ±SD values. The results winfected and control mice are marked (⁄p < 0.05; ⁄⁄p < 0.01), whilst differences between ino significant differences and different letters indicate significant differences.
during PbA infection. The anti-inflammatory cytokine TGF-b and IL-10 showed marked increase after 4 dpi and continued to increaseduring P. yoelii infection. The levels of pro-inflammatory cytokinesin PbA infected mice were higher than in P. yoelii infected mice;TGF-b and IL-10 exhibited changes opposite to the changes in pro-inflammatory cytokines. Overall, the levels of TGF-b and IL-10 pro-duction in PbA mice were lower than that those of P. yoelii infectedmice suggesting that early death in PbA infected mice is caused bylower immune response rather than cytokine storm. Furthermore,it is suggested that elevated TGF-b and IL-10 levels in P. yoelii in-fected mice may protect from early death by promoting Treg pro-duction (Fig. 6).
4. Discussion
The spleen is a complex organ with several functions, includingthe removal of senescent or aberrant red blood cells (RBC) from cir-culation and removal of circulating pathogenic organisms (Krücken
nfected with PbA and P. yoelii along with matched controls. Data is representative ofere evaluated by ANOVA, with Bonferroni post-test. Significant differences betweennfected groups on respective days are marked (by letters).The same letter indicates

T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92 91
et al., 2005) with reports also demonstrating an immunopathologicinvolvement of this organ in human (Faucher et al., 2006), rodentmodels (Oster et al., 1980) and in experimental malaria models (El-ing, 1980). The spleen is the major site of antigen presentation andT cell priming during blood-borne infections (Banchereau et al.,2000). Animal models and particularly rodents are just one wayto examine the initiation of immune responses in the T-cell areasand germinal centers of the spleen (del Portillo et al., 2012). BothCD4+ and CD8+ T cells are involved in the pathogenesis of ECM(Hermnsen et al., 1997) but are also required for the effective con-trol of malaria parasites (Weidanz et al., 1990) emphasizing thedelicate balance that exists between host-mediated control ofinfection. Although the physiopathology of CM has been exten-sively investigated, the exact cellular and molecular basis of thespleen pathology during CM is still unclear (Keswani and Bhatta-charyya, 2013).
CD4+ T cells produce various cytokines and play pivotal roles inimmune responses (Grau et al., 1986; Cruz Cubas et al., 1994).CD4+ T cells are now classified into four subsets: Th1, Th2, Th17,and Treg cells according to the cytokines they produce and thefunctions they perform (Weaver et al., 2007). Th17 cells, whichhave been characterized recently, play important roles in inflam-matory and autoimmune responses (Weaver et al., 2007). In thepresent study; we have observed a possible role of Treg andTh17 in the context of pro inflammatory and anti-inflammatorycytokine milieu during PbA infection. These findings have beenconfirmed by treatment of mice separately with nonlethal P. yoeliiinfection. Survival and weight change exhibited by PbA and P yoeliiinfected mice animals of all experimental groups of mice, showedan indirect relationship with parasitemia level. However theseobservations are still only associations and further evidence willconfirm the pathways responsible for the disease outcomes.
At present, there are no reports concerning Th17 responsesagainst malaria parasite infection. Treg suppress host immune re-sponses and promote the proliferation of pathogens leading to per-sistent infection (Sakaguchi et al., 1995; Belkaid and Rouse, 2005).There are contradictory reports on the roles of Treg cells in malaria,depending on the host and parasite species, interestingly we ob-served increased expression of Th17 in spleen during nonresolvingPbA infected mice at 8 dpi whereas, increased Foxp3 in the spleenof resolving P. yoelii infected mice at 8 dpi. Several researchers re-ported that Treg cells suppress the host immune system and aggra-vate malaria both in human and mice (Belkaid and Rouse, 2005;Hisaeda et al., 2004) and also it has been reported that Treg cellsprotect hosts through suppression of severe inflammation (Seixasand Ostler, 2005). Other subsets such as Th17 cells have howevernot been described during PbA infection. The ability of proinflam-matory cytokines such as IL-6 and regulatory cytokines like TGF-b to either promote or protect from severe malaria are likely to de-pend on many factors, including host and parasite genetics and theimmune status of an infected individual. Given the delicate balanc-ing act to be achieved between control of overwhelming infectionand prevention of immune-pathology (Artavanis-Tsakonas et al.,2003), it is very tempting to invoke a potential role for regulatoryT cells and Th17 cells in malaria.
We propose that host immune responses in malaria are mayberegulated by the differential effects of Th17 and Treg cells. We ob-served increase TGF-b expression when Tregs expression were ele-vated, and both TGF-b and IL-6 when Th17 is high. Previous studieshave reported that IL-1b and IL-6, but not TGF-b, are responsiblefor Th17 differentiation in humans (Acosta-Rodriguez et al.,2007), but others have suggested that TGF-b dosage plays an essen-tial role in the differentiation of naïve human CD4+ T cells towardthe Th17 lineage (O’Garra et al., 2008). This has also raised impor-tant questions concerning how TGF-b might act at apparentlycrossed purposes to do this. Several recent studies have built on
earlier observations to begin to provide a mechanistic basis bywhich these two lineages, although linked by a common require-ment for TGF-b, go their separate ways (Zhou et al., 2008; Ichiyamaet al., 2008).
IL-2 is crucial for the maintenance and expansion of naturalFoxp3+ Treg cell populations (Setoguchi et al., 2005) and inhibitsTh17 development, an effect that is dependent on STAT5, TNF-a,interferon-c (IFN-c), interleukin-17, and IL-23 (Laurence et al.,2007; Chen et al., 2011), further supporting our findings. On theother hand, Foxp3 regulatory T cells control inflammation throughmultiple mechanisms including production of the cytokine IL-10and TGF-b (Chen et al., 2011). The current consensus is that IL-6 in-duces Th17 differentiation together with lower dose of TGF-b. Re-cently, it was revealed that IL-23 is also critical for thedevelopment of Th17 and for immune cell activation (Ishidaet al., 2010).
However, in the case of Treg differentiation, TGF-b inducedtranscription factor Smad3 possibly cooperates with NFAT to in-duce the expression of Foxp3 (Tone et al., 2008). In contrast,TGF-b and IL-6 trigger the coordinated activation of Smad3 andSTAT3 to induce the transcription factor RORct necessary forTh17 differentiation (Veldhoen et al., 2006). We hypothesize thatTGF-b plays an important role in modulating the balance betweenthe differentiation of Treg and Th17 cells in malaria. In this study,we also examined the expression of Foxp3, IL-17A, and RORct inPbA and P. yoelii induced mice splenic CD4 T cells. Mechanistically,the activation status of STAT3 and NFATc2 following TGF-b or IL-6plus TGF-b will be evaluated in more details after in vitro study andusing specific inhibitors. These studies indicate that possibly TGF-bis a central cytokine to the differentiation of both cells and IL-6 isthe key cytokine regulating Treg/Th17 balance.
Acknowledgments
Authors want to thank ICMR, India, Santion No. 80/790/2013-ECD-I, Dated 12/07/2013 for fellowship support, Board of Researchin Nuclear Science, DAE, India, (Santion No. 2010/37B/41/BRNS,Dated 06/09/2010) and Department of Science and Technology,(DST SR/SO/HS/0133/2009 SERB/F/1762, Dated – 06.02.2012) Govt.of India for all financial support. We like to thank Centre for Re-search in Nanoscience & Nanotechnology, University of Calcutta,for their Flow cytometry instrument facility. Authors also wantto give special thanks to Soham Mitra and Samir Jana for theirassistance during the experiments. Also we like to thank Depart-ment of Science and Technology, Govt. of India FIST Program.
References
Acosta-Rodriguez, E.V., Napolitani, G., Lanzavecchia, A., Sallusto, F., 2007.Interleukins 1beta and 6 but not transforming growth factor-beta areessential for the differentiation of interleukin 17-producing human T helpercells. Nature Immunology 8, 942–949.
Amante, F.H., Haque, A., Stanley, A.C., Rivera, L.F., Randall, L.M., Wilsonm, Y.A., Yeo,G., Pieper, C., Crabb, B.S., de Koning-Ward, T.F., Lundie, R.J., Good, M.F., Pinzon-Charry, A., Pearson, M.S., Duke, M.G., McManus, D.P., Loukas, A., Hill, G.R.,Engwerda, C.R., 2010. Immune-mediated mechanisms of parasite tissuesequestration during experimental cerebral malaria. Journal of Immunology185, 3632–3642.
Amante, F.H., Stanley, A.C., Randall, L.M., Zhou, Y., Haque, A., McSweeney, K., Waters,A.P., Janse, C.J., Good, M.F., Hill, G.R., Engwerda, C.R., 2007. A role for naturalregulatory T cells in the pathogenesis of experimental cerebral malaria.American Journal of Pathology 171, 548–559.
Artavanis-Tsakonas, K., Tongren, E., Riley, E., 2003. The war between the malariaparasite and the immune system: immunity, immunoregulation andimmunopathology. Clinical and Experimental Immunology 133, 145–152.
Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S., Liu, Y.J., Pulendran, B.,Palucka, K., 2000. Immunobiology of dendritic cells. Annual ReviewImmunology 18, 767–811.
Belkaid, Y., Rouse, B.T., 2005. Natural regulatory T cells in infectious disease. NatureImmunology 6, 353–360.

92 T. Keswani, A. Bhattacharyya / Experimental Parasitology 141 (2014) 82–92
Belnoue, E., Kayibanda, M., Vigario, A.M., Deschemin, J.C., van Rooijen, N., Viguier,M., Snounou, G., Rénia, L., 2002. On the pathogenic role of brain-sequesteredalphabeta CD8+ T cells in experimental cerebral malaria. Journal of Immunology169, 6369–6375.
Cambos, M., Belanger, B., Jacques, A., Roulet, A., Scorza, T., 2008. Natural regulatory(CD4+CD25+FOXP+) T cells control the production of pro-inflammatorycytokines during Plasmodium chabaudi adami infection and do not contributeto immune evasion. International Journal of Parasitology 38, 229–238.
Chen, G., Liu, J., Wang, Q.H., Wu, Y., Feng, H., Zheng, W., Guo, S.Y., Li, D.M., Wang, J.C.,Cao, Y.M., 2009. Effects of CD4(+) CD25(+) Foxp3(+) regulatory T cells on earlyPlasmodium yoelii 17XL infection in BALB/c mice. Parasitology 136, 1107–1120.
Chen, Yi., Haines, C.J., Gutcher, I., Hochweller, K., Blumenschein, W.M., McClanahan,T., Hämmerling, G., Li, M.O., Cua, D.J., McGeachy, M.J., 2011. Foxp3+ regulatory Tcells promote T helper 17 cell development in vivo through regulation ofinterleukin-2. Immunity 34, 409–421.
Couper, K.N., Blount, D.G., Wilson, M.S., Hafalla, J.C., Belkaid, Y., Kamanaka, M.,Flavell, R.A., de Souza, J.B., Riley, E.M., 2008. IL-10 from CD4+ CD25� Foxp3�
CD127� adaptive regulatory T cells modulates parasite clearance and pathologyduring malaria infection. PLoS Pathogens 4 (2), e1000004.
Cruz Cubas, A.B., Gentilini, M., Monjour, L., 1994. Cytokines and T-cell response inmalaria. Biomedicine & Pharmacotherapy 48, 27–33.
Del Portillo, H.A., Ferrer, M., Brugat, T., Martin-Jaular, L., Langhorne, J., Lacerda,M.V.G., 2012. The role of the spleen in malaria. Cellular Microbiology 14, 343–355.
deWalick, S., Amante, F.H., McSweeney, K.A., Randall, L.M., Stanley, A.C., Haque, A.,Kuns, R.D., Kelli, P.A., MacDonald Hill, G.R., Engwerda, C.R., 2007. Cutting edge:conventional dendritic cells are the critical APC required for the induction ofexperimental cerebral malaria. Journal of Immunology 178, 6033–6037.
Eling, W.M., 1980. Role of spleen in morbidity and mortality of Plasmodium bergheiinfection in mice. Infection and Immunity 30, 635–641.
Faucher, J.F., Creantor, C., Hustache-Mathieu, L., Chirouze, C., Millon, L., Hoen, B.,2006. Atypical course of falciparum malaria in an asplenic patient. PresseMedicale 35, 793–795.
Feng, H., Zhu, X.T., Qi, Z.M., Wang, Q.H., Wang, G.G., Pan, Y.Y., Li, Y., Zheng, L., Jiang,Y.J., Shang, H., Cui, L., Cao, Y.M., 2012. Transient attenuated Foxp3 expression onCD4(+) T cells treated with 7D4 mAb contributes to the control of parasiteburden in DBA/2 mice infected with lethal Plasmodium chabaudi chabaudi AS.Scandinavian Journal of Immunology 75, 46–53.
Grau, G.E., Piguet, P.F., Engers, H.D., Louis, J.A., Vassalli, P., Lambert, P.H., 1986. L3T4+T lymphocytes play a major role in the pathogenesis of murine cerebral malaria.Journal of Immunology 137, 2348–2354.
Hansen, D.S., Bernard, N.J., Nie, C.Q., Schofield, L., 2007. NK cells stimulaterecruitment of CXCR3+ T cells to the brain during Plasmodium bergheimediated cerebral malaria. Journal of Immunology 178, 5779–5788.
Hansen, D.S., Evans, K.J., D’Ombrain, M.C., Bernard, N.J., Sexton, A.C., Buckingham, L.,Scalzo, A.A., Schofield, L., 2005. The natural killer complex regulates severemalarial pathogenesis and influences acquired immune responses toPlasmodium berghei ANKA. Infection and Immunity 73, 2288–2297.
Hansen, D.S., Siomos, M.A., Buckingham, L., Scalzo, A.A., Schofield, L., 2003.Regulation of murine cerebral malaria pathogenesis by CD1d-restricted NKTcells and the natural killer complex. Immunity 18, 391–402.
Haque, A., Best, S.E., Amante, F.H., Mustafah, S., Desbarrieres, L., de Labastida, F.,Sparwasser, T., Hill, G.R., Engwerda, C.R., 2010. CD4+ natural regulatory T cellsprevent experimental cerebral malaria via CTLA-4 when expanded in vivo. PLoSPathogens 6, e1001221.
Hermsen, C., van de Wiel, T., Mommers, E., Sauerwein, R., Eling, W., 1997. Depletionof CD4+ or CD8+ T-cells prevents Plasmodium berghei induced cerebral malaria inend-stage disease. Parasitology 114, 7–12.
Hisaeda, H., Maekawa, Y., Iwakawa, D., Okada, H., Himeno, K., Kishihara, K.,Tsukumo, S., Yasutomo, K., 2004. Escape of malaria parasites from hostimmunity requires CD4+CD25+ regulatory T cells. Nature Medicine 10, 29–30.
Ichiyama, K., Yoshida, H., Wakabayashi, Y., Chinen, T., Saeki, K., Nakaya, M., Takaesu,G., Hori, S., Yoshimura, A., Kobayashi, T., 2008. Foxp3 inhibits RORgammat-mediated IL-17A mRNA transcription through direct interaction withRORgammat. Journal of Biological Chemistry 283, 17003–17008.
Ishida, H., Matsuzaki-Moriya, C., Imai, T., Yanagisawa, K., Nojima, Y., Suzue, K., Hirai,M., Iwakura, Y., Yoshimura, A., Hamano, S., Shimokawa, C., Hisaeda, H., 2010.Development of experimental cerebral malaria is independent of IL-23 and IL-17. Biochemical and Biophysical Research Communications 402, 790–795.
Keswani, T., Bhattacharyya, A., 2013. Splenocyte apoptosis in Plasmodium bergheiANKA infection: possible role of TNF-a and TGF-b. Parasite Immunology 35, 73–90.
Kim, J.M., Rasmussen, J.P., Rudensky, A.Y., 2007. Regulatory T cells preventcatastrophic autoimmunity throughout the lifespan of mice. NatureImmunology 8, 191–197.
Krücken, J., Mehnert, L.I., Dkhil, M.A., El-Khadragy, M., Benten, W.P.M., Mossmann,H., 2005. Massive destruction of malaria-parasited red blood cells despitespleen closure. Infection and Immunity 73, 6390–6398.
Laurence, A., Tato, C.M., Davidson, T.S., Kanno, Y., Chen, Z., Yao, Z., Blank, R.B.,Meylan, F., Siegel, R., Hennighausen, L., Shevach, E.M., O’shea, J.J., 2007.Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation.Immunity 26, 371–381.
Mitra, S., Keswani, T., Dey, M., Bhattacharya, S., Sarkar, S., Goswami, S., Ghosh, N.,Dutta, A., Bhattacharyya, A., 2012. Copper induced immunotoxicity involves cellcycle arrest and cell death in the spleen and thymus. Toxicology 293, 78–88.
Nitcheu, J., Bonduelle, O., Combadiere, C., Tefit, M., Seilhean, D., Mazier, D.,Combadiere, B., 2003. Perforin-dependent brain-infiltrating cytotoxic CD8+ Tlymphocytes mediate experimental cerebral malaria pathogenesis. Journal ofImmunology 170, 2221–2228.
O’Garra, A., Stockinger, B., Veldhoen, M., 2008. Differentiation of human T(H)-17cells does require TGF-beta! Nature Immunology 9, 588–590.
Oster, C.N., Koontz, L.C., Wyler, D.J., 1980. Malaria in asplenic mice: effects ofsplenectomy, congenital asplenia, and splenic reconstitution on the course ofinfection. American Journal of Tropical Medicine and Hygiene 29, 1138–1142.
Rouse, B.T., Suvas, S., 2004. Regulatory cells and infectious agents: detentes cordialeand contraire. Journal of Immunology 173, 2211–2215.
Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M., Toda, M., 1995. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor a-chains(CD25): breakdown of a single mechanism of self-tolerance causes variousautoimmune diseases. Journal of Immunology 155, 1151–1164.
Seixas, E., Ostler, D., 2005. Plasmodium chabaudi chabaudi (AS): differential cellularresponses to infection in resistant and susceptible mice. ExperimentalParasitology 110, 394–405.
Setoguchi, R., Hori, S., Takahashi, T., Sakaguchi, S., 2005. Homeostatic maintenanceof natural Foxp3(+) CD25(+) CD4(+) regulatory T cells by interleukin (IL)-2 andinduction of autoimmune disease by IL-2 neutralization. Journal ofExperimental Medicine 201, 723–735.
Steeg, C., Adler, G., Sparwasser, T., Fleischer, B., Jacobs, T., 2009. Limited role ofCD4+Foxp3+ regulatory T cells in the control of experimental cerebral malaria.Journal of Immunology 183, 7014–7022.
Sun, W., He, X., Guo, Z., Wang, Q., Li, X., Rayner, J., Zhang, L., Wang, J., Cao, X., 2004.IL12p40-overexpressing immature dendritic cells induce T cellhyporesponsiveness in vitro but accelerate allograft rejection in vivo: role ofNK cell activation and interferon-gamma production. Immunology Letters 94,191–199.
Todryk, S.M., Bejon, P., Mwangi, T., Plebanski, M., Urban, B., Marsh, K., Hill, A.V.S.,Flanagan, K.L., 2008. Correlation of memory T cell responses against TRAP withprotection from clinical malaria, and CD4 CD25 high T cells with susceptibilityin Kenyans. PLoS ONE 3, e2027.
Tone, Y., Furuuchi, K., Kojima, Y., Tykocinski, M.L., Greene, M.I., Tone, M., 2008.Smad3 and NFAT cooperate to induce Foxp3 expression through its enhancer.Nature Immunology 9, 194–202.
Veldhoen, M., Hocking, R.J., Atkins, C.J., Locksley, R.M., Stockinger, B., 2006. TGF betain the context of an inflammatory cytokine milieu supports de novodifferentiation of IL-17-producing T cells. Immunity 24, 179–189.
Vigario, A.M., Gorgette, O., Dujardin, H.C., Cruz, T., Cazenave, P.A., Six, A., Bandeira,A., Pied, S., 2007. Regulatory CD4+ CD25+ Foxp3+ T cells expand duringexperimental Plasmodium infection but do not prevent cerebral malaria.International Journal of Parasitology 37, 963–973.
Walther, M., Jeffries, D., Finney, O.C., Njie, M., Ebonyi, A., Deininger, S., 2009. Distinctroles for FOXP3 and FOXP3 CD4 T cells in regulating cellular immunity touncomplicated and severe Plasmodium falciparum malaria. Public Library ofScience Pathogens 5, e1000364.
Weaver, C.T., Hatton, R.D., Mangan, P.R., Harrington, L.E., 2007. IL-17 familycytokines and the expanding diversity of effector T cell lineage. Annual Reviewof Immunology 25, 821–852.
Weidanz, W.P., Melancon-Kaplan, J., Cavacini, L.A., 1990. Cell-mediated immunityto the asexual blood stages of malarial parasites: animal models. ImmunologyLetters 25, 87–95.
Yanez, D.M., Manning, D.D., Cooley, A.J., Weidanz, W.P., van der Heyde, H.C., 1996.Participation of lymphocyte subpopulations in the pathogenesis ofexperimental murine cerebral malaria. Journal of Immunology 157, 1620–1624.
Yang, X.O., Pappu, B.P., Nurieva, R., Akimzhanov, A., Kang, H.S., Chung, Y., Ma, L.,Shah, B., Panopoulos, A.D., Schluns, K.S., Watowich, S.S., Tian, Q., Jetten, A.M.,Dong, C., 2008. T helper 17 lineage differentiation is programmed by orphannuclear receptors ROR alpha and ROR gamma. Immunity 28, 29–39.
Zhou, L., Ivanov, I.I., Spolski, R., Min, R., Shenderov, K., Egawa, T., Levy, D.E., Leonard,W.J., Littman, D.R., 2007. IL-6 programs T(H)-17 cell differentiation bypromoting sequential engagement of the IL-21 and IL-23 pathways. NatureImmunology 8, 967–974.
Zhou, L., Lopes, J.E., Chong, M.M.W., Ivanov, I.I., Min, R., Victora, G.D., Shen, Y., Du, J.,Rubtsov, Y.P., Rudensky, A.Y., Ziegler, S.F., Littman, D.R., 2008. TGF-beta inducedFoxp3 inhibits TH17 cell differentiation by antagonizing RORgammat function.Nature 453, 236–240.


















