
Plasmodium berghei-Infected Primary Hepatocytes … · and Giampietro Corradin2* ... phase F-moc...
11
of August 29, 2018. This information is current as Cells In Vitro T + Circumsporozoite Protein to Specific CD8 Hepatocytes Process and Present the -Infected Primary Plasmodium berghei Laurent Renia and Giampietro Corradin Silayuv E. Bongfen, Ralph Torgler, Jackeline F. Romero, http://www.jimmunol.org/content/178/11/7054 doi: 10.4049/jimmunol.178.11.7054 2007; 178:7054-7063; ; J Immunol References http://www.jimmunol.org/content/178/11/7054.full#ref-list-1 , 19 of which you can access for free at: cites 53 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2007 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on August 29, 2018 http://www.jimmunol.org/ Downloaded from by guest on August 29, 2018 http://www.jimmunol.org/ Downloaded from
-
Upload
truongkhue -
Category
Documents
-
view
214 -
download
0
Transcript of Plasmodium berghei-Infected Primary Hepatocytes … · and Giampietro Corradin2* ... phase F-moc...

of August 29, 2018.This information is current as
Cells In Vitro T+Circumsporozoite Protein to Specific CD8
Hepatocytes Process and Present the -Infected PrimaryPlasmodium berghei
Laurent Renia and Giampietro CorradinSilayuv E. Bongfen, Ralph Torgler, Jackeline F. Romero,
http://www.jimmunol.org/content/178/11/7054doi: 10.4049/jimmunol.178.11.7054
2007; 178:7054-7063; ;J Immunol
Referenceshttp://www.jimmunol.org/content/178/11/7054.full#ref-list-1
, 19 of which you can access for free at: cites 53 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Plasmodium berghei-Infected Primary Hepatocytes Process andPresent the Circumsporozoite Protein to Specific CD8� T CellsIn Vitro1
Silayuv E. Bongfen,* Ralph Torgler,* Jackeline F. Romero,* Laurent Renia,†‡§¶
and Giampietro Corradin2*
A substantial and protective response against malaria liver stages is directed against the circumsporozoite protein (CSP) andinvolves induction of CD8� T cells and production of IFN-�. CSP-derived peptides have been shown to be presented on thesurface of infected hepatocytes in the context of MHC class I molecules. However, little is known about how the CSP andother sporozoite Ags are processed and presented to CD8� T cells. We investigated how primary hepatocytes from BALB/cmice process the CSP of Plasmodium berghei after live sporozoite infection and present CSP-derived peptides to specificH-2Kd-restricted CD8� T cells in vitro. Using both wild-type and spect�/� P. berghei sporozoites, we show that both infectedand traversed primary hepatocytes process and present the CSP. The processing and presentation pathway was found toinvolve the proteasome, Ag transport through a postendoplasmic reticulum compartment, and aspartic proteases. Thus,it can be hypothesized that infected hepatocytes can contribute in vivo to the elicitation and expansion of a T cellresponse. The Journal of Immunology, 2007, 178: 7054 –7063.
T he feasibility of vaccine development against malarialiver stages is indicated by studies (1, 2) which show thatsterile immunity against the liver stage can be obtained by
immunization of animals and humans with irradiated and, recently,genetically attenuated sporozoites (3). Irradiated sporozoite-in-duced protective immunity has been shown to involve IFN-�,CD4�, and CD8� T cells directed against liver stage parasites, andin vivo depletion of these effectors has led to blood infection upona live sporozoite challenge (4–10).
Even though sporozoites pass through Kupffer cells before in-fecting hepatocytes (11), hepatocytes are the only cells that sustaincomplete development of malaria liver stages after infection withsporozoites. Maintenance of the anti-sporozoite protective CD8�
T cell response requires hepatic stages, since protection againstsporozoite challenge is abrogated if these hepatic stages are elim-inated (12). Furthermore, immunization with irradiated Plasmo-dium berghei-infected hepatocytes (but not infected liver non-parenchymal cells) conferred protection against a sporozoitechallenge (13). It has been suggested that in malaria, hepatocytesexpress MHC class I-peptide complexes on their surface and thatthe recognition of these complexes by CD8� T cells is necessaryfor protection (reviewed in Ref. 14). This notion is supported bystudies which show that irradiated sporozoite-immunized �2m�/�
mice are not protected against sporozoite challenge, despite induc-tion of IL-12, IFN-�, and proliferating T cells (15).
It was widely believed that only endogenous Ags could be pre-sented on MHC class I molecules to CD8� T cells. A large bodyof evidence now shows that peptides derived from exogenous Agsare also capable of binding to MHC class I molecules and stimu-lating CD8� T cells, a process known as cross-presentation. Sev-eral models of cross-presentation have been described (16–21),especially for professional APCs. In addition, the use of exogenousAgs such as long synthetic peptides to immunize mice (22, 23) andhumans (24) to obtain MHC class I-restricted CTLs also supportsthe concept of cross-presentation. Besides professional APCs, non-professional APCs have also been shown to be capable of cross-presenting long polypeptides to CD8� T cells (25, 26).
The mechanisms involved in malaria Ag presentation to T cellsremain unclear, because the low number of Plasmodium-infectedhepatocytes in the liver has prevented the development of directAg presentation studies. The question is further complicated by thefact that sporozoites do not infect the first hepatocyte they encoun-ter, but traverse through many hepatocytes before infecting a finalone (27). As they traverse, they leave behind trails of the circum-sporozoite protein (CSP)3 in the traversed cells, some of whichreseal their membranes and survive and are, in principle, capableof presenting the Ag. This suggests that sporozoite Ags could ei-ther be presented by the infected cell, the traversed cell, or both. Ithas been suggested (28) that migration of sporozoites throughhepatocytes is essential for final infection. However, a recentstudy (29) has identified a sporozoite microneme protein essen-tial for cell traversal (SPECT) and has shown that sporozoites
*Department of Biochemistry, University of Lausanne, Epalinges, Switzerland; †In-stitut Cochin, Departement d’Immunologie, Hopital Cochin, Paris, France; ‡InstitutNational de la Sante et de la Recherche Medicale, Paris, France; §Centre National dela Recherche Scientifique Unite Mixte de Recherche, Paris, France; and ¶UniversiteRene Descartes, Paris, France
Received for publication June 28, 2006. Accepted for publication March 19, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by European Community Grant QLK 2-CT-2002-00700.2 Address correspondence and reprint requests to Dr. Giampietro Corradin, Depart-ment of Biochemistry, Chemin des Boveresses 155, Epalinges, Switzerland. E-mailaddress: [email protected]
3 Abbreviations used in this paper: CSP, circumsporozoite; SPECT, sporozoite mi-croneme protein essential for cell traversal; wt, wild type; BFA, brefeldin A; LLnL,N-acetyl-L-leucyl-L-leucyl-L-norleucinal; Cbz-LLL, carbobenzoxyl-leucinyl-leucinyl-leucinal; ICS, intracellular cytokine staining; RT, room temperature; DAPI, 4�,6�-diamidino-2-phenylindole; EEF, exoerythrocytic form; GdCl, gadolinium chloride;DC, dendritic cell; HSP, heat shock protein.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

deficient in the spect gene are incapable of cell traversal, butcapable of infection and normal development. spect�/� para-sites are therefore an essential tool in determining the ability ofthe infected hepatocyte to process and present sporozoite Ags toCD8� T cells.
Despite evidence for the importance of hepatic stages to maintaina protective immune response and suggestions that hepatocytespresent MHC class I-peptide complexes on their surface, very fewstudies (30, 31) have attempted to address the question of direct Agpresentation by hepatocytes in an in vitro or in vivo assay.
In this study, using mouse primary hepatocytes, wild-type (wt)and spect�/� P. berghei sporozoites, we investigated the cell (tra-versed or infected hepatocytes or both) involved in processing andpresentation of the CSP of P. berghei to specific CD8� T cells. Wealso investigated the pathway of this processing and presentationby the primary hepatocytes after infection with wt sporozoites. Weformally show that both infected and traversed hepatocytes are able toprocess the native CSP present on live parasites for presentation tospecific CD8� T cells. Our data show that this processing is predom-inantly dependent on the proteasome and also on aspartic proteases.
Materials and MethodsPeptides
The peptide PbCS245–253, representing the H-2Kd-restricted CTL epitope ofthe CSP of P. berghei and the peptide PfCS327–335, an HLA-A2.1-restrictedpeptide from the CSP of P. falciparum, were synthesized by standard solid-phase F-moc chemistry. Peptide stock solutions (2 mg/ml) were preparedin 1� PBS and stored at �20°C.
Cells
The APCs were primary hepatocytes isolated from BALB/c mice by atwo-step perfusion of liver lobules with collagenase as previously de-scribed (32), with slight modifications. Briefly, mice were sacrificed byCO2 inhalation, dissected, and a liver lobule was cut out. The lobule wasperfused for 10 min with Ca2�-free HEPES buffer (pH 7.6) at 37°C and ata rate of 5 ml/min. The lobule was then perfused with type IV collagenase(Sigma-Aldrich) solution (Ca2�-free HEPES buffer containing 0.04% typeIV collagenase and 0.075% CaCl2�2H2O) for 5 min at 37°C. The perfusedlobule was incubated for 10 min at 37°C in the collagenase solution. Usingsterile pipettes, the tissue was gently teased apart to release cells. Cellswere washed once with Ca2�-free HEPES buffer at 800 rpm for 30 s at4°C. The pellet was gently resuspended in DMEM, layered on 60% Percoll,and centrifuged at 2000 rpm for 2 min at 4°C. The resulting pellet wasresuspended in complete culture medium (DMEM supplemented with 10%FCS, 1% penicillin-streptomycin, 1% HEPES, and 0.05 mM 2-ME), andcentrifuged again at 800 rpm for 30 s at 4°C. The pellet was resuspendedin complete culture medium, 105 cells seeded per well in a 48-well gelatin-coated culture plate and incubated at 37°C. Two to 4 h later, the mediumwas replaced by fresh culture medium and the cells were incubated over-night at 37°C.
To ensure the purity of the hepatocyte preparation, the isolated hepato-cytes were stained with a panel of Abs to exclude the presence of contam-inating cells (anti-F4/80 Ab conjugated to FITC to exclude macrophages,anti-CD11c Ab conjugated to PE to exclude dendritic cells (DCs), andanti-MHC class II Ab conjugated to PE to exclude both macrophages andDCs) and also positively stained with anti-rabbit ferritin Ab, using rabbitanti-mouse IgG Alexa Ab conjugated to Alexa Fluor 488 as a second Ab.As a positive control for the anti-F4/80 Ab, peritoneal macrophages wereused, whereas DCs were used as a positive control for the anti-CD11c andanti-MHC class II Abs. The cells were then analyzed by FACS after stain-ing (data not shown).
The human hepatoma cell line HepG2 was used in certain experiments.The H-2Kd-restricted CD8� T cell clone PbCS-C7, specific for the P.berghei CSP CTL epitope PbCS245–253 (5), was used as effector cells.
Mice and sporozoites
Six- to 8-wk-old BALB/c and C57BL/6 mice were obtained from HarlanBreeders. All mice were housed under pathogen-free conditions in the an-imal facility of the Swiss Institute for Experimental Cancer Research (Ep-alinges, Switzerland). Animals were handled according to institutionalguidelines.
Anopheles stephensi mosquitoes containing either wt or spect�/� (29) P.berghei sporozoites were maintained under the same conditions. Sporozo-ites (wt and spect�/�) were obtained by dissection of infected female A.stephensi mosquitoes to obtain salivary glands, followed by homogeniza-tion of the salivary glands to release sporozoites. Wild-type midgut sporo-zoites were also used.
Inhibitors
The inhibitors used were brefeldin A (BFA), N-acetyl-L-leucyl-L-leucyl-L-norleucinal (LLnL), carbobenzoxyl-leucinyl-leucinyl-leucinal (Cbz-LLL),lactacystin, pepstatin, leupeptin, chloroquine, and ammonium chloride(NH4Cl). All inhibitors were obtained from Sigma-Aldrich. In preliminaryexperiments, we showed that none of these inhibitors were toxic for theprimary hepatocytes at the concentrations used (data not shown).
IFN-� ELISPOT assay
The IFN-� ELISPOT assay was performed as previously described (33).Briefly, 105 primary hepatocytes were seeded per well in 1% gelatin-coated48-well culture plates and incubated overnight at 37°C to enable them toadhere. Hepatocytes were washed with culture medium and infected fordifferent lengths of time with P. berghei sporozoites (25,000/well). Unin-fected cells were used as a negative control. All cells were washed twicewith culture medium and cocultured with 5000 PbCS-C7 CD8� T cells perwell for 4 h at 37°C. T cells were then transferred to ELISPOT platespreviously coated with anti-mouse IFN-� Ab and blocked with 1% BSA.The ELISPOT plates were incubated for 20–24 h at 37°C, after which theplates were washed and developed. Spots were visualized and counted withan automated ELISPOT reader.
Intracellular cytokine staining (ICS)
Primary hepatocytes (105) were seeded per well in 1% gelatin-coated 48-well culture plates and incubated overnight at 37°C. Cells were washed andinfected with 25.000 wt, spect�/�, or midgut sporozoites per well for 8 hat 37°C. As controls, heat-killed wt sporozoites or material from salivaryglands of uninfected mosquitoes were incubated with hepatocytes for 8 hbefore coculture with specific CD8� T cells. As a positive control, cellswere pulsed with 1 �g/ml of the PbCS245–253 peptide for 2 h at 37°C.Unpulsed/uninfected cells were used as a negative control. After pulsingand infection, cells were washed and cocultured with 5000 PbCS-C7 CD8�
T cells per well for 4 h at 37°C in the presence of 10 �g/ml of the intra-cellular transport inhibitor BFA. T cells were then harvested, washed, andstained with FITC-conjugated anti-mouse CD8 Ab (BD Biosciences) for30 min at 4°C. Cells were washed twice with PBS-3% FCS and fixed for10 min at room temperature with prewarmed 4% paraformaldehyde. Aftertwo washes, with 0.5% saponin-1� PBS, cells were stained overnight at4°C with PE-conjugated anti-mouse IFN-� Ab (BD Biosciences) diluted inthe saponin solution. Cells were washed with PBS-3% FCS and analyzedfor CD8� and IFN-�� T cells by flow cytometry.
FIGURE 1. Primary hepatocytes optimally process and present CSP-derived peptides 8–20 h after infection with sporozoites. Primary hepato-cytes were infected with P. berghei sporozoites for different lengths oftime. Secretion of IFN-� by specific CD8� T cells after coculture withinfected hepatocytes was determined by an IFN-� ELISPOT assay. Errorbars, Mean � SD of duplicate wells.
7055The Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In separate experiments, primary hepatocytes were prepared from aBALB/c mouse 24 h after i.v. injection of 10 mg/kg body weight of gad-olinium chloride (GdCl) to depress Kupffer cell function (34). These hepa-tocytes were then plated on 1% gelatin-coated 48-well plates and ICS wasconducted as described above.
Inhibition assay
In the inhibition assays, the inhibitors were used at the following concen-trations: LLnL (50 �M), Cbz-LLL (10 �M), lactacystin (20 �M), pepstatin(36 �M), leupeptin (110 �M), BFA (35 �M), chloroquine (10 �M), andNH4Cl (10 mM). Hepatocytes were incubated for 2 h at 37°C with thedifferent inhibitors before pulsing with peptide or infection with sporozo-
ites. Pulsing or infection of inhibitor-treated cells was performed eitherwithout washing off the inhibitor or after washing. Cells were then washedand cocultured with specific CD8� T cells. The inhibition assays werecompleted as described for ICS.
Chromium release assay
Primary hepatocytes (105) were plated per well in a 1% gelatin-coated96-well plate and incubated overnight at 37°C. They were washed andeither pulsed with 1 �g/ml of the PbCS245–253 peptide (using the HLA-A2.1-restricted CTL epitope from the CSP of P. falciparum, PfCS327–335,as a negative control) for 1 h at 37°C, or infected with 25,000 wt sporo-zoites for 16 h. Seventy-five microcuries of chromium was then added and
FIGURE 2. Primary hepatocytes specifically process and present the CSP of P. berghei. H-2Kd hepatocytes were prepared from wt mice and from micetreated with 10 mg/kg body weight of GdCl. Hepatocytes (105/well) were either infected with P. berghei sporozoites (a) or pulsed with the PbCS245–253
peptide (b), then cocultured for 4 h with 5 � 103 CD8� T cells. c, H-2Kd hepatocytes (105) were either infected with sporozoites or incubated with materialfrom uninfected mosquito salivary glands (sgm) before coculture with 5 � 103 CD8� T cells. d, Hepatocytes were either infected with P. bergheisporozoites or incubated with heat-inactivated (HK spz) or midgut sporozoites, then cocultured with CD8� T cells. e, Hepatocytes were prepared from eitherBALB/c (H-2Kd) or C57BL/6 (H-2Kb) mice. They were infected with P. berghei sporozoites and cocultured with CD8� T cells. Activation of the T cellswas determined by ICS and flow cytometry. Error bars, Mean � SD of duplicate wells. Negative, Uninfected and unpulsed cells.
7056 PRESENTATION OF SPOROZOITE Ags BY HEPATOCYTES
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

incubation was continued for another hour. Cells were washed and cocul-tured with PbCS-C7 CTLs at different E:T ratios in the presence or absenceof 10 �g/ml Fas-Fc for 4 h at 37°C. The supernatants were collected andread with a gamma counter.
Detection of apoptotic hepatocytes
Primary hepatocytes (8 � 104) were seeded per well in 8-well LabTekchamber slides and incubated overnight at 37°C. Cells were washed andeither infected with 2 � 104 P. berghei sporozoites for 8 h or pulsed withthe PbCS245–253 peptide for 2 h at 37°C. Hepatocytes were washed andcocultured with PbCS-C7 CTLs at a ratio of 1:1 for 2 h at 37°C. The CTLswere washed off with 1� PBS and the hepatocytes were fixed for 10 minwith ice-cold methanol. After washing, cells were blocked with 1%BSA-1� PBS for 1 h at room temperature (RT). Hepatocytes infected withsporozoites were stained for the intracellular parasite by incubation withanti-heat shock protein (HSP) 70 Ab (2E6) (35) for 1 h at RT, followed bywashing and staining for 1 h at RT with a 1/100 dilution of anti-mouse IgG Abconjugated to Alexa Fluor 488. Cells were washed and all cells were stainedwith a 1/100 dilution of 4�,6�-diamidino-2-phenylindole (DAPI) to detect ap-optotic nuclei. The cells were then viewed under a fluorescence microscope.
Quantification of intracellular sporozoites
Primary hepatocytes (8 � 104) were seeded per well in 8-well LabTekchamber slides and incubated overnight at 37°C. Cells were pretreated withdifferent inhibitors for 2 h at 37°C. Wild-type sporozoites (2 � 104) wereadded without washing off inhibitors and incubated for 2 h at 37°C. Cellswere washed with 1� PBS and either fixed with cold methanol (for de-
tection of both extracellular and intracellular sporozoites) or with cold 1%glutaraldehyde (for detection of extracellular sporozoites only) for 10 min.All cells were washed and blocked for 1 h at RT with 1% BSA-1� PBS,followed by incubation with an anti-PbCS repeat Ab for 1 h at RT. Afterwashing, cells were stained for 1 h at RT with a 1/100 dilution of anti-mouseIgG Ab conjugated to Alexa Fluor 488. Cells were then washed, mounted in50% glycerol, and observed under a fluorescence microscope. Sporozoiteswere counted in duplicate wells. The number of extracellular sporozoites (glu-taraldehyde fixation) was subtracted from the number of sporozoites in wellswith methanol fixation to yield the number of intracellular sporozoites.
Quantification of exoerythrocytic forms (EEFs)
Primary hepatocytes (8 � 104) were seeded per well in 8-well LabTekchamber slides and incubated overnight at 37°C. Cells were infected with2 � 104 wt or spect�/� P. berghei sporozoites per well for 20 h at 37°C.Cells were washed with 1� PBS and fixed for 10 min with ice-cold meth-anol. Cells were blocked with 1% BSA-1� PBS for 1 h at RT followingwashing with 1� PBS. Infected hepatocytes were then stained with anti-HSP 70 Ab for 1 h at RT. After washing, cells were stained for 1 h at RTwith a 1/100 dilution of anti-mouse IgG Ab conjugated to Alexa Fluor 488.Cells were then washed, mounted in 50% glycerol, and observed under afluorescence microscope. EEFs were counted over the entire well.
Quantification of the migration capacity of sporozoites
Sporozoites migrate through several hepatocytes breaching their plasmamembranes, before infecting a final one by formation of a parasitopho-rous vacuole (27). The wounded cells release cytosolic contents into the
FIGURE 3. Both infected and uninfected (traversed) hepatocytes process and present the CSP. a, HepG2 cells were treated with chromium and infected witheither wt or spect�/� sporozoites. Migration of sporozoites through cells was determined by quantifying the amount of chromium released. b, Primary hepatocyteswere infected with either wt or spect�/� sporozoites. The number of EEFs was determined by staining intracellular parasites with an anti-HSP70 Ab. c, Primaryhepatocytes were infected with either wt or spect�/� sporozoites. After coculture with specific CD8� T cells, activation of T cells was determined by ICS and flowcytometry. Error bars, Mean � SD of duplicate wells. Negative, Uninfected cells. d, Primary hepatocytes were infected with either wt or spect�/� sporozoites,then cocultured with T cells at different E:T ratios for 20 h. The number of EEFs was determined for each condition. Error bars, Mean � SD of duplicate wells.
7057The Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

microenvironment (R. Torgler, S. E. Bongfen, J. F. Romero, A. Tardivel,M. Thome, and G. Corradin, submitted for publication). We exploited thisability of sporozoites to wound cells and release cytosolic contents to quan-tify their migration capacity.
HepG2 cells (105) were seeded per well in a 96-well plate and incubatedfor 48 h at 37°C to enable them to adhere. Seventy-five micorcuries ofchromium was then added and incubated for 1 h at 37°C. Cells werewashed several times to eliminate all residual chromium, then infected with5 � 104 wt or spect�/� sporozoites for 3 h at 37°C. The resulting super-natant was collected and read with a gamma counter. The amount of chro-mium released was proportional to the number of sporozoites added andhence to hepatocytes wounded due to the migratory action of sporozoites.
Statistical analysis
The one-way ANOVA and two-tailed Student t test were used to determinethe significance of variation between test samples. All p values �0.05 wereconsidered to be significant.
ResultsPrimary hepatocytes can process and present the P. bergheiCSP, and do so optimally 8–20 h after infection withsporozoites
Our preliminary in vitro data showed that primary hepatocytes arecapable of processing and presenting sporozoite Ags after infec-tion (data not shown). However, the level of CD8� T cell activa-tion varied with the length of time of hepatocyte infection. To findout the optimal time necessary for processing and presentation ofthe CSP after infection with sporozoites, hepatocytes were infectedwith sporozoites for different time periods over a total period of 24 h.Secretion of IFN-� by the PbCS-C7 CD8� T cells after coculture withinfected hepatocytes was determined by an ELISPOT assay. Optimalactivation of the T cells occurred when hepatocytes were infectedwith the sporozoites for a time period between 8 and 20 h. Acti-vation was lower when T cells were cocultured with hepatocytesinfected with sporozoites for �8 h or �20 h. (Fig. 1). This sug-gests that the optimal time necessary for hepatocytes to processand present the CSP to CD8� T cells after infection with sporo-zoites is between 8 and 20 h.
To establish the purity of the hepatocyte preparation, primaryhepatocytes were stained with different Abs (see Materials andMethods) to exclude the presence of contaminating cells, and thepreparation was found to contain essentially hepatocytes (data notshown). To further prove that the Ag presentation was performedby hepatocytes and not by Kupffer cells that could contaminate thehepatocyte preparation, primary hepatocytes were prepared from aBALB/c mouse 24 h after i.v. injection of 10 mg/kg body weightof GdCl. GdCl is known to deplete Kupffer cells in vivo (34).Hepatocytes from both a wt and GdCl-treated mouse were thenused in ICS. As shown in Fig. 2a, hepatocytes depleted of Kupffercells presented the CSP of P. berghei just as well as hepatocytesfrom wt mice. Similarly, presentation of the PbCS245–253 peptideby both preparations of hepatocytes was comparable (Fig. 2b).
Taking into consideration that T cells could be activated bycontaminants of parasite preparations, hepatocytes were incubatedwith material from uninfected mosquito salivary glands as a con-trol. Fig. 2c shows that the activation of T cells observed is specificfor sporozoites, because T cells were not activated in the presenceof uninfected salivary gland material. To eliminate the possibilityof Ag from extracellular degradation being presented, hepatocyteswere incubated with heat-killed and with midgut sporozoites(which contain the mature CSP, but neither migrate through norinfect hepatocytes). T cells were not activated in either case, implyingthat migration and/or infection of sporozoites are necessary for theCSP to be processed and presented by hepatocytes (Fig. 2d).
To further demonstrate that hepatocytes are indeed presentingthe peptide, ICS was performed with hepatocytes from C57BL/6
mice (H-2Kb). As can be seen in Fig. 2e, T cells were only acti-vated when they were cocultured with hepatocytes from BALB/cmice, which express the correct MHC molecules (H-2Kd).
Both infected and wounded (traversed) hepatocytes process andpresent the CSP
Sporozoites migrate through several hepatocytes before infecting afinal one by formation of a parasitophorous vacuole (27). Duringmigration, they leave behind a trail of the CSP. Some of the tra-versed cells reseal their membrane and survive, and thus can, inprinciple, present the CSP to T cells.
To find out which cells were presenting the CSP (traversed orinfected cells or both), an ICS assay was done with both wt andspect�/� sporozoites, whose migratory ability through hepatocytesis totally or profoundly inhibited (Fig. 3a) (29), but which infectand develop normally in hepatocytes (Fig. 3b). As shown in Fig.3c, T cells are activated when hepatocytes are infected with eitherwt or spect�/� sporozoites. The level of activation of T cells by wtsporozoites is about 5-fold that observed with spect�/� sporozo-ites. Given that spect�/� parasites are capable of infection but notmigration, and that migration and/or infection are necessary forCSP processing and presentation (Fig. 2d), these results suggestthat both infected and uninfected (traversed) hepatocytes present
FIGURE 4. Proteasome inhibitors and BFA inhibit processing and pre-sentation of the CSP. Primary hepatocytes were treated with the differentinhibitors 2 h before infection with sporozoites (spz; a) or pulsing with thePbCS245–253 peptide (b). Infected and pulsed cells were cocultured with5 � 103 CD8� T cells for 4 h at 37°C. Activation of CD8� T cells wasdetermined by ICS and flow cytometry. Negative, Untreated cells. Errorbars, Mean � SD of duplicate wells.
7058 PRESENTATION OF SPOROZOITE Ags BY HEPATOCYTES
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
the CSP to T cells, with most of the presentation being done by thetraversed cells. To further show that infected cells present the CSP,hepatocytes were infected with either wt or spect�/� sporozoites
and cocultured with specific CD8� T cells at different E:T ratiosfor 20 h. The number of EEFs in each case was then determined.Fig. 3di shows that EEFs from hepatocytes infected with wt andspect�/� sporozoites are equally eliminated, indicating that in-fected cells do present the CSP. The perfect correlation (r � 0.997)between the elimination of EEFs from wt or spect�/� sporozoite-infected cells as a function of E:T ratio (Fig. 3dii) indicates a nullor limited contribution of parasite killing due to activation ofCD8� T cells by traversed cells.
Processing and presentation of the CSP by primary hepatocytesare dependent on the proteasome and on vesicular transportthrough the Golgi apparatus
The presentation of proteasome-generated cytosolic peptides byMHC class I molecules requires vesicular transport from the en-doplasmic reticulum through the Golgi complex to the cell surface.The dependence of the processing of the CSP on the proteasomewas tested using the proteasome inhibitors LLnL, Cbz-LLL, andlactacystin. Processing and presentation of the CSP after infectionof hepatocytes with sporozoites was strongly inhibited ( p � 0.05)by the proteasome inhibitors (Fig. 4a). In contrast, presentation ofthe PbCS245–253 peptide was unaffected (Fig. 4b). Likewise, sporo-zoite-infected hepatocytes lost their capacity to present the CSP whenthey were treated with the intracellular transport inhibitor BFA,whereas presentation of the PbCS245–253 peptide was not affected.
These results indicate that after sporozoite infection, processing andpresentation of the CSP follow a proteasome-dependent pathway.
To establish that the inhibitors added did not affect the biosyn-thetic machinery of the parasite, inhibitor-treated cells werewashed before infection, and ICS was conducted as before. Nodifference in activation of T cells was observed, whether or not theinhibitors were washed before infection (data not shown), indicat-ing that the inhibitors do not have an effect on the parasite, butinhibit a process in the hepatocyte.
Effect of lysosomal protease inhibitors and inhibitors oflysosomal acidification on processing of the CSP
Sporozoites are a source of exogenous Ags and, as shown in othersystems (26), it is possible that processing of sporozoite proteinscould depend, at least in part, on the vacuolar pathway. To test theinvolvement of endolysosomal proteases in CSP processing, hepa-tocytes were treated with leupeptin and pepstatin before infection
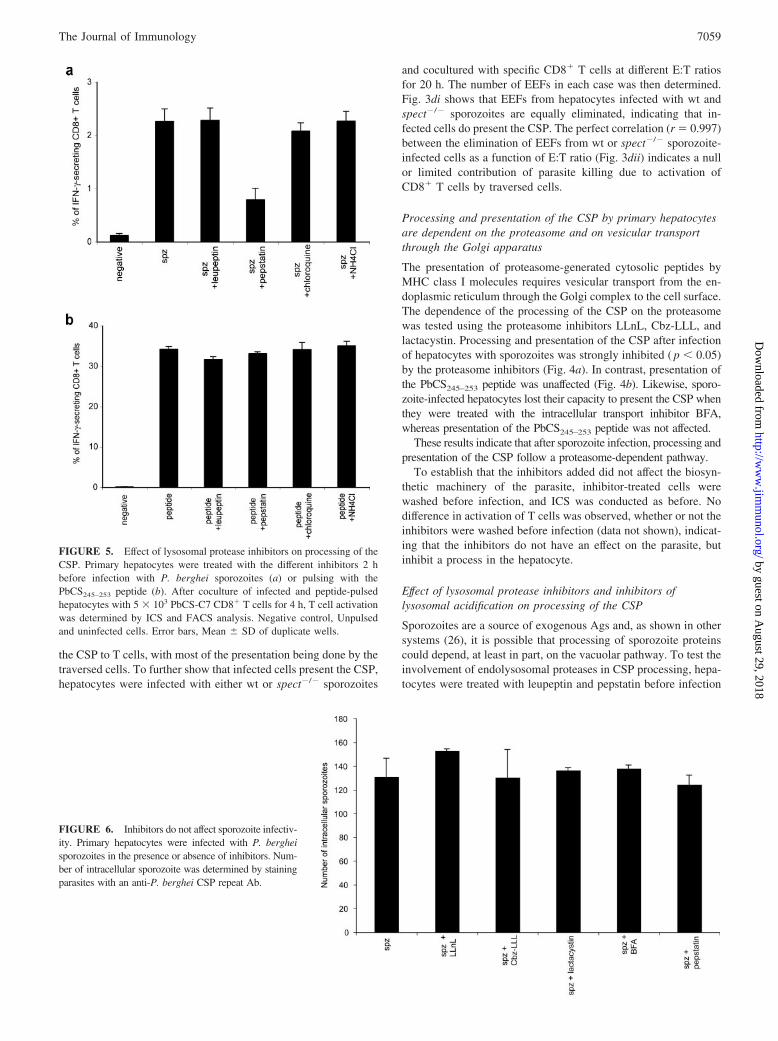
FIGURE 5. Effect of lysosomal protease inhibitors on processing of theCSP. Primary hepatocytes were treated with the different inhibitors 2 hbefore infection with P. berghei sporozoites (a) or pulsing with thePbCS245–253 peptide (b). After coculture of infected and peptide-pulsedhepatocytes with 5 � 103 PbCS-C7 CD8� T cells for 4 h, T cell activationwas determined by ICS and FACS analysis. Negative control, Unpulsedand uninfected cells. Error bars, Mean � SD of duplicate wells.
FIGURE 6. Inhibitors do not affect sporozoite infectiv-ity. Primary hepatocytes were infected with P. bergheisporozoites in the presence or absence of inhibitors. Num-ber of intracellular sporozoite was determined by stainingparasites with an anti-P. berghei CSP repeat Ab.
7059The Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
with sporozoites or pulsing with the PbCS245–253 peptide. Process-ing and presentation of the CSP were not significantly affected byleupeptin ( p � 0.05). However, activation of T cells was signifi-cantly reduced ( p � 0.05) when hepatocytes were treated with theaspartic protease inhibitor pepstatin. To further establish the role ofthe endosomal compartment, the hepatocytes were treated withchloroquine and ammonium chloride, which alter the acidificationof lysosomes and endolysosomes. No significant effect on process-ing and presentation was observed (Fig. 5a). As expected, presen-
tation of the PbCS245–253 peptide was not affected by any of theinhibitors (Fig. 5b).
The different inhibitors used have no effect on sporozoiteinfection of hepatocytes
To show that the inhibitors which had an effect on processing andpresentation did not affect the rate of hepatocyte infection bysporozoites, hepatocytes were infected with sporozoites in thepresence or absence of inhibitors for 2 h. Infected cells were
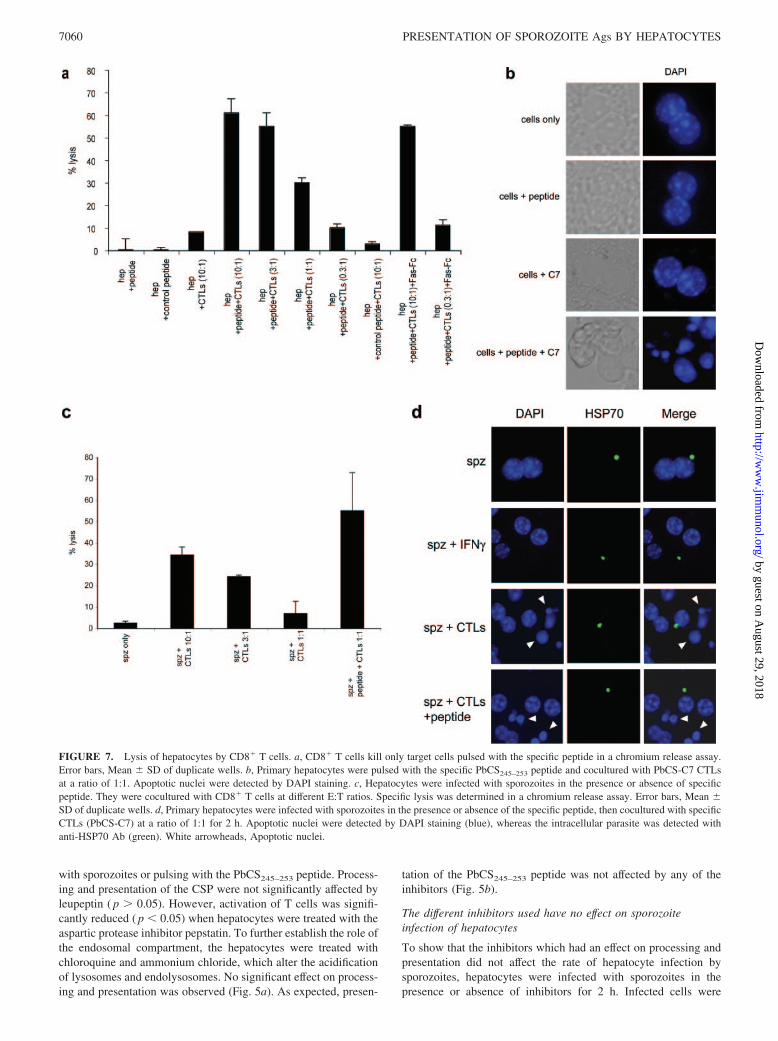
FIGURE 7. Lysis of hepatocytes by CD8� T cells. a, CD8� T cells kill only target cells pulsed with the specific peptide in a chromium release assay.Error bars, Mean � SD of duplicate wells. b, Primary hepatocytes were pulsed with the specific PbCS245–253 peptide and cocultured with PbCS-C7 CTLsat a ratio of 1:1. Apoptotic nuclei were detected by DAPI staining. c, Hepatocytes were infected with sporozoites in the presence or absence of specificpeptide. They were cocultured with CD8� T cells at different E:T ratios. Specific lysis was determined in a chromium release assay. Error bars, Mean �SD of duplicate wells. d, Primary hepatocytes were infected with sporozoites in the presence or absence of the specific peptide, then cocultured with specificCTLs (PbCS-C7) at a ratio of 1:1 for 2 h. Apoptotic nuclei were detected by DAPI staining (blue), whereas the intracellular parasite was detected withanti-HSP70 Ab (green). White arrowheads, Apoptotic nuclei.
7060 PRESENTATION OF SPOROZOITE Ags BY HEPATOCYTES
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

stained for intracellular sporozoites using a mAb against the re-peats of the P. berghei CSP. As shown in Fig. 6, the number ofintracellular sporozoites did not vary significantly ( p � 0.05)whether or not infection was done in the presence of inhibitors.
Hepatocytes are lysed by CD8� T cells after pulsing with thePbCS CTL epitope or after incubation with sporozoites
In addition to production of cytokines such as IFN-�, CD8� Tcells also induce target cell death via a perforin/granzyme B- orFas-dependent mechanism. To further prove that hepatocytes areable to process and present the CSP after infection and to show thatthe CD8� T cells used were functional, chromium release assayswere conducted to find out whether hepatocytes could be lysed byCD8� T cells after pulsing with the optimal CTL epitope or afterinfection with sporozoites.
Hepatocytes pulsed with the specific PbCS245–253 peptide werefound to be efficiently lysed by specific CD8� T cells, as opposedto hepatocytes pulsed with a CTL epitope of P. falciparum,PfCS327–335, even at high E:T ratios (Fig. 7a). The killing of pep-tide-pulsed hepatocytes observed here seemed to be primarily dueto perforin/granzyme B, and not via Fas-Fas ligand, because ad-dition of soluble Fas-Fc did not inhibit killing. This killing wasfurther shown by staining the peptide-pulsed hepatocytes for ap-optotic nuclei 2 h after coculture with specific CTLs. In this case,massive apoptosis was observed, as shown by the presence of frag-mented nuclei (Fig. 7b, fourth panel). No apoptosis was observedwhen hepatocytes were cocultured with CTLs in the absence ofpeptide (Fig. 7b, third panel) or when peptide alone was added tohepatocytes (Fig. 7b, second panel).
Hepatocytes were also lysed by specific CTLs after infectionwith sporozoites, as shown by a chromium release assay (Fig. 7c).The lysis was greatly increased when the specific peptide wasadded to infected hepatocytes. However, by microscopically ex-amining cocultures of CD8� T cells and hepatocytes after sporo-zoite infection, we found that infected hepatocytes did not appearto undergo apoptosis, as shown by the absence of apoptotic nucleiin infected cells 2 h after coculture with specific CTLs in the pres-ence (Fig. 7d, fourth panel) or absence (Fig. 7d, third panel) ofpeptide. Interestingly, the uninfected cells bordering infected cellswere apoptotic whether or not peptide was added (Fig. 7d, thirdand fourth panels, arrowheads). Importantly, no apoptosis was ob-served when infected cells were cultured alone (Fig. 7d, firstpanel) or in the presence of IFN-� for 2 h (Fig. 7d, second panel).
DiscussionThe aim of this study was to provide evidence that primary hepa-tocytes are able to process the P. berghei CSP after infection withsporozoites and to present the optimal P. berghei CSP CTL epitopeto specific CD8� T cells. In this light, we show that both infectedand uninfected hepatocytes (traversed by sporozoites) process andpresent the CSP, and that processing is largely dependent on pro-teasomes and, to a lesser extent, aspartic proteases.
There is a continuous discussion about the cell type involvedin the processing and presentation of the CSP after sporozoite in-fection. Three cell types may be implicated in this process: Kupffercells, DCs, and hepatocytes.
Although little is known about the role of Kupffer cells, there isno doubt that DCs can present sporozoite-derived Ags, and a pro-tective status is attained in mouse experimental models in which arelatively high number of sporozoites are injected (36–38). How-ever, in the field, where the number of sporozoites injected viamosquito bites is low, and with a high proportion of these sporo-zoites going directly to the hepatocyte within minutes (11), DCpresentation may not be the relevant pathway for priming. Thus,
the crucial, still unresolved question is: can infected hepatocytespresent Ags to T cells in a primary and secondary response? Ourdata formally prove that at least in vitro, both infected and tra-versed hepatocytes can present sporozoite-derived Ags to clonedCD8� T cells. spect�/� P. berghei sporozoites have been shown tobe incapable of cell traversal, but capable of infection and normaldevelopment in hepatocytes (29). Our finding that CD8� T cellscould be activated when hepatocytes were infected with spect�/�
sporozoites (Fig. 3c) shows that infected hepatocytes present the P.berghei CSP CTL epitope to T cells, because in this case we havea situation of infection only and no or minimal traversal. This isfurther proven by the fact that in the presence of T cells, EEFsfrom spect�/� parasites could be eliminated as well as those fromwt parasites (Fig. 3d). The activation observed is unlikely to be dueto uptake and processing of extracellular CSP by hepatocytes,because no activation of T cells was observed when hepatocyteswere incubated with heat-killed or midgut sporozoites (Fig. 2d).The long-lasting protection observed with the irradiated sporozoitemodel is also consistent with continuous presentation of sporozoite-derived Ags by infected hepatocytes to primed T cells. It has been,in fact, shown that as early as 12 h after infection, no liver mono-nuclear cells contain parasite Ags (38), suggesting that the onlycells that contain parasite Ags for a long period of time are hepa-tocytes. In addition, elimination of hepatic stages after irradiatedsporozoite immunization abrogates protection (12). More recently,long-lasting protection has been obtained by immunization withlive sporozoites of mice under chloroquine treatment (39). Thisprotection was abrogated by eliminating hepatic forms with pri-maquine. Although the role of primaquine in Ag presentation invivo has not been elucidated, these data suggest that activation ofprimary responses may be mediated by infected hepatocytes. It isknown that priming of immune responses requires expression ofcostimulatory molecules by the APCs. In fact, costimulatory mol-ecules have been shown to be expressed in the cytoplasm of hepa-tocytes during hepatitis C viral infection (40) and in biliary atresiapatients with liver dysfunction (41). It is, therefore, possible thatsporozoite infection of hepatocytes induces expression of costimu-latory molecules necessary for priming T cell responses.
Although not formally proven, our in vitro microscopy datashow that sporozoite-infected hepatocytes do not undergo nuclearfragmentation, suggesting that they are resistant to CTL-inducedapoptosis. This is in conformity with studies which have shownthat infected cells are protected from apoptosis (42, 43). In addi-tion, the finding that Fas- and perforin-deficient mice could still beprotected from P. berghei (44) and P. yoelii (45) infection suggeststhat other mechanisms are implicated in the destruction of hepaticforms. It is interesting to note that even though infected cells didnot undergo apoptosis, neighboring noninfected cells (probablytraversed by sporozoites) readily underwent apoptosis upon addi-tion of specific CTLs (Fig. 7d, third panel). This suggests that theCS protein left behind by sporozoites traveling through or betweenhepatocytes (27, 46) is presented by traversed hepatocytes. Thisfinding is further substantiated by the fact that hepatocytes infectedwith spect�/� sporozoites do not activate T cells as much as thoseinfected with wt sporozoites, suggesting that traversed hepatocytesprocess and present the CSP to T cells. This presentation by tra-versed hepatocytes could represent an escape mechanism elabo-rated by the parasite to evade the host immune system, by directingspecific T cells to traversed hepatocytes which present malaria-derived peptides. However, further studies are needed to determinewhether this mechanism is operative in vivo. Nevertheless, the factthat protection could still be observed in Fas- and perforin-defi-cient mice (44) suggests that infected cells do not undergo apo-ptosis in vivo.
7061The Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

We found that optimum processing and presentation occur 8–20h after infection of hepatocytes with sporozoites (Fig. 1). This is inagreement with previous studies conducted in vivo (47). In thatstudy, it was shown that maximum CD8� T cell activation oc-curred within the first 8 h after immunization with normal or ir-radiated P. yoelii sporozoites, and lasted for 48 h.
The dependence of processing on the proteasome is consistentwith studies which showed that sporozoites deposit the CSP in thecytoplasm of cells, beginning from the time of their attachment tothe cell surface and peaking 4–6 h after invasion (48). In addition,parasite Ags have also been shown to be inserted into the parasi-tophorous vacuole membrane and into the hepatocyte cytoplasm(49). Recently, a pentameric motif, the Plasmodium export ele-ment (termed Pexel), was discovered and shown to be necessaryfor trafficking of Plasmodium falciparum proteins in the infectederythrocyte (50, 51). This motif is present in the P. berghei CSPand could be used to traffic the CSP from the parasitophorousvacuole to the cytoplasm in infected hepatocytes.
After infection of human hepatoma cells with sporozoites, theparasites were found not to be localized in an acidified vacuole,and the released CSP did not enter the endocytic compartment ofthe host cell (48). In agreement with this, we found that CD8� Tcell activation was not affected by chloroquine and NH4Cl, sug-gesting that an acidic compartment may not be involved in CSPprocessing. It has, however, been shown that cross-presentation ofAgs by DCs occurs through a phagolysosome, which is resistant toacidification (52). Whether this mechanism also occurs in hepato-cytes has not been determined. Interestingly, pepstatin inhibitedprocessing, suggesting that cellular or parasitic aspartic proteasesplay a role in processing the CSP. These proteases could possiblycleave the CSP in the parasitophorous vacuole before its exit intothe hepatocyte cytoplasm.
On the other hand, additional studies using spect�/� parasitesand inhibitors of Ag processing and presentation as shown here areneeded to clearly distinguish the requirements of processing andpresentation of live sporozoites from those of proteins left behindby the traveling parasite.
In summary, our results clearly show that primary hepatocytesprocess and present the CSP of P. berghei live sporozoites via a path-way involving the proteasome and aspartic proteases. The biologicalsignificance of hepatocyte processing and presentation is very likelyassociated with the maintenance of the protective immune responseinduced by irradiated sporozoites and in subsequent protection againsta parasite challenge, both of which have to be formally proven in vivo.On the other hand, in our opinion, the role of infected hepatocytes inthe elicitation of a primary immune response still remains elusive.
The finding that infected hepatocytes also present parasite Agshas profound consequences for vaccine development. The CSP isthe most widely studied pre-erythrocytic stage vaccine candidateand is an immunodominant protective Ag in irradiated sporozoites(53). However, the developing parasite also expresses other Agsinside the infected cell. If these Ags are presented on the surfaceof the infected hepatocyte, they could be potential candidates for apre-erythrocytic stage vaccine.
Finally, a clear understanding of the mechanisms involved inprocessing and presentation of sporozoite Ags could assist in thedesign of effective vaccines capable of stimulating CD4� andCD8� T cell responses.
AcknowledgmentsWe thank Drs. Peter van Endert, Joerg Reimann, and Robert Menard fortheir helpful discussion and Robert Menard for providing spect�/� para-sites. We also thank Masao Yuda for generation of the SPECT-deficientparasites.
DisclosuresThe authors have no financial conflict of interest.
References1. Clyde, D. F. 1975. Immunization of man against falciparum and vivax malaria by
use of attenuated sporozoites. Am. J. Trop. Med. Hyg. 24: 397–401.2. Nussenzweig, R. S., J. Vanderberg, H. Most, and C. Orton. 1967. Protective
immunity produced by the injection of x-irradiated sporozoites of plasmodiumberghei. Nature 216: 160–162.
3. Mueller, A. K., M. Labaied, S. H. Kappe, and K. Matuschewski. 2005. Geneti-cally modified Plasmodium parasites as a protective experimental malaria vac-cine. Nature 433: 164–167.
4. Schofield, L., J. Villaquiran, A. Ferreira, H. Schellekens, R. Nussenzweig, andV. Nussenzweig. 1987. � Interferon, CD8� T cells and antibodies required forimmunity to malaria sporozoites. Nature 330: 664–666.
5. Romero, P., J. L. Maryanski, G. Corradin, R. S. Nussenzweig, V. Nussenzweig,and F. Zavala. 1989. Cloned cytotoxic T cells recognize an epitope in the cir-cumsporozoite protein and protect against malaria. Nature 341: 323–326.
6. Sedegah, M., F. Finkelman, and S. L. Hoffman. 1994. Interleukin 12 induction ofinterferon �-dependent protection against malaria. Proc. Natl. Acad. Sci. USA 91:10700–10702.
7. Seguin, M. C., F. W. Klotz, I. Schneider, J. P. Weir, M. Goodbary, M. Slayter,J. J. Raney, J. U. Aniagolu, and S. J. Green. 1994. Induction of nitric oxidesynthase protects against malaria in mice exposed to irradiated Plasmodiumberghei infected mosquitoes: involvement of interferon � and CD8� T cells.J. Exp. Med. 180: 353–358.
8. Wang, R., Y. Charoenvit, G. Corradin, P. De La Vega, E. D. Franke, andS. L. Hoffman. 1996. Protection against malaria by Plasmodium yoelii sporozoitesurface protein 2 linear peptide induction of CD4� T cell- and IFN-�-dependentelimination of infected hepatocytes. J. Immunol. 157: 4061–4067.
9. Scheller, L. F., S. J. Green, and A. F. Azad. 1997. Inhibition of nitric oxideinterrupts the accumulation of CD8� T cells surrounding Plasmodium berghei-infected hepatocytes. Infect. Immun. 65: 3882–3888.
10. Rodrigues, E. G., J. Claassen, S. Lee, J. M. Wilson, R. S. Nussenzweig, andM. Tsuji. 2000. Interferon-�-independent CD8� T cell-mediated protective anti-malaria immunity elicited by recombinant adenovirus. Parasite Immunol. 22:157–160.
11. Frevert, U., S. Engelmann, S. Zougbede, J. Stange, B. Ng, K. Matuschewski,L. Liebes, and H. Yee. 2005. Intravital observation of Plasmodium berghei sporo-zoite infection of the liver. PLoS Biol. 3: E192.
12. Scheller, L. F., and A. F. Azad. 1995. Maintenance of protective immunityagainst malaria by persistent hepatic parasites derived from irradiated sporozo-ites. Proc. Natl. Acad. Sci. USA 92: 4066–4068.
13. Renia, L., M. M. Rodrigues, and V. Nussenzweig. 1994. Intrasplenic immuni-zation with infected hepatocytes: a mouse model for studying protective immu-nity against malaria pre-erythrocytic stage. Immunology 82: 164–168.
14. Good, M. F., and D. L. Doolan. 1999. Immune effector mechanisms in malaria.Curr. Opin. Immunol. 11: 412–419.
15. White, K. L., H. L. Snyder, and U. Krzych. 1996. MHC class I-dependent pre-sentation of exoerythrocytic antigens to CD8� T lymphocytes is required forprotective immunity against Plasmodium berghei. J. Immunol. 156: 3374–3381.
16. Yewdell, J. W., C. C. Norbury, and J. R. Bennink. 1999. Mechanisms of exog-enous antigen presentation by MHC class I molecules in vitro and in vivo: im-plications for generating CD8� T cell responses to infectious agents, tumors,transplants, and vaccines. Adv. Immunol. 73: 1–77.
17. Guermonprez, P., L. Saveanu, M. Kleijmeer, J. Davoust, P. Van Endert, andS. Amigorena. 2003. ER-phagosome fusion defines an MHC class I cross-pre-sentation compartment in dendritic cells. Nature 425: 397–402.
18. Houde, M., S. Bertholet, E. Gagnon, S. Brunet, G. Goyette, A. Laplante,M. F. Princiotta, P. Thibault, D. Sacks, and M. Desjardins. 2003. Phagosomes arecompetent organelles for antigen cross-presentation. Nature 425: 402–406.
19. Rock, K. L. 2003. The ins and outs of cross-presentation. Nat. Immunol. 4:941–943.
20. Ackerman, A. L., C. Kyritsis, R. Tampe, and P. Cresswell. 2005. Access ofsoluble antigens to the endoplasmic reticulum can explain cross-presentation bydendritic cells. Nat. Immunol. 6: 107–113.
21. Bevan, M. J. 1976. Cross-priming for a secondary cytotoxic response to minor Hantigens with H-2 congenic cells which do not cross-react in the cytotoxic assay.J. Exp. Med. 143: 1283–1288.
22. Blum-Tirouvanziam, U., C. Beghdadi-Rais, M. A. Roggero, D. Valmori,S. Bertholet, C. Bron, N. Fasel, and G. Corradin. 1994. Elicitation of specificcytotoxic T cells by immunization with malaria soluble synthetic polypeptides.J. Immunol. 153: 4134–4141.
23. Roggero, M. A., V. Meraldi, J. A. Lopez, G. Eberl, J. C. Romero, H. Matile,B. Betschart, G. Corradin, and J. Renggli. 2000. The synthetic, oxidized C-ter-minal fragment of the Plasmodium berghei circumsporozoite protein elicits ahigh protective response. Eur. J. Immunol. 30: 2679–2685.
24. Lopez, J. A., C. Weilenman, R. Audran, M. A. Roggero, A. Bonelo, J. M. Tiercy,F. Spertini, and G. Corradin. 2001. A synthetic malaria vaccine elicits a potentCD8� and CD4� T lymphocyte immune response in humans: implications forvaccination strategies. Eur. J. Immunol. 31: 1989–1998.
25. Gnjatic, S., D. Atanackovic, M. Matsuo, E. Jager, S. Y. Lee, D. Valmori,Y. T. Chen, G. Ritter, A. Knuth, and L. J. Old. 2003. Cross-presentation of HLAclass I epitopes from exogenous NY-ESO-1 polypeptides by nonprofessionalAPCs. J. Immunol. 170: 1191–1196.
7062 PRESENTATION OF SPOROZOITE Ags BY HEPATOCYTES
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

26. Prato, S., T. Maxwell, A. Pinzon-Charry, C. W. Schmidt, G. Corradin, andJ. A. Lopez. 2005. MHC class I-restricted exogenous presentation of a synthetic102-mer malaria vaccine polypeptide. Eur. J. Immunol. 35: 681–689.
27. Mota, M. M., G. Pradel, J. P. Vanderberg, J. C. Hafalla, U. Frevert,R. S. Nussenzweig, V. Nussenzweig, and A. Rodriguez. 2001. Migration of Plas-modium sporozoites through cells before infection. Science 291: 141–144.
28. Mota, M. M., J. C. Hafalla, and A. Rodriguez. 2002. Migration through host cellsactivates Plasmodium sporozoites for infection. Nat. Med. 8: 1318–1322.
29. Ishino, T., K. Yano, Y. Chinzei, and M. Yuda. 2004. Cell-passage activity isrequired for the malarial parasite to cross the liver sinusoidal cell layer.PLoS. Biol. 2: E4.
30. Weiss, W. R., S. Mellouk, R. A. Houghten, M. Sedegah, S. Kumar, M. F. Good,J. A. Berzofsky, L. H. Miller, and S. L. Hoffman. 1990. Cytotoxic T cells rec-ognize a peptide from the circumsporozoite protein on malaria-infected hepato-cytes. J. Exp. Med. 171: 763–773.
31. Renia, L., M. S. Marussig, D. Grillot, S. Pied, G. Corradin, F. Miltgen,G. Del Giudice, and D. Mazier. 1991. In vitro activity of CD4� and CD8� Tlymphocytes from mice immunized with a synthetic malaria peptide. Proc. Natl.Acad. Sci. USA 88: 7963–7967.
32. Seglen, P. O. 1976. Preparation of isolated rat liver cells. Methods Cell Biol. 13:29–83.
33. Bonelo, A., D. Valmori, F. Triponez, J. M. Tiercy, G. Mentha, J. Oberholzer,P. Champagne, J. F. Romero, F. Esposito, I. Nebie, et al. 2000. Generation andcharacterization of malaria-specific human CD8� lymphocyte clones: effect ofnatural polymorphism on T cell recognition and endogenous cognate antigenpresentation by liver cells. Eur. J. Immunol. 30: 3079–3088.
34. Rai, R. M., S. Q. Yang, C. McClain, C. L. Karp, A. S. Klein, and A. M. Diehl.1996. Kupffer cell depletion by gadolinium chloride enhances liver regenerationafter partial hepatectomy in rats. Am. J. Physiol. 270: G909–G918.
35. Tsuji, M., D. Mattei, R. S. Nussenzweig, D. Eichinger, and F. Zavala. 1994.Demonstration of heat-shock protein 70 in the sporozoite stage of malaria para-sites. Parasitol. Res. 80: 16–21.
36. Jung, S., D. Unutmaz, P. Wong, G. Sano, K. De los Santos, T. Sparwasser, S. Wu,S. Vuthoori, K. Ko, F. Zavala, et al. 2002. In vivo depletion of CD11c� dendriticcells abrogates priming of CD8� T cells by exogenous cell-associated antigens.Immunity 17: 211–220.
37. Bruna-Romero, O., and A. Rodriguez. 2001. Dendritic cells can initiate protectiveimmune responses against malaria. Infect. Immun. 69: 5173–5176.
38. Leiriao, P., M. M. Mota, and A. Rodriguez. 2005. Apoptotic Plasmodium-in-fected hepatocytes provide antigens to liver dendritic cells. J. Infect. Dis. 191:1576–1581.
39. Belnoue, E., F. T. Costa, T. Frankenberg, A. M. Vigario, T. Voza, N. Leroy,M. M. Rodrigues, I. Landau, G. Snounou, and L. Renia. 2004. Protective T cellimmunity against malaria liver stage after vaccination with live sporozoites underchloroquine treatment. J. Immunol. 172: 2487–2495.
40. Mochizuki, K., N. Hayashi, K. Katayama, N. Hiramatsu, T. Kanto, E. Mita,T. Tatsumi, N. Kuzushita, A. Kasahara, H. Fusamoto, et al. 1997. B7/BB-1 ex-pression and hepatitis activity in liver tissues of patients with chronic hepatitis C.Hepatology 25: 713–718.
41. Kobayashi, H., Z. Li, A. Yamataka, G. J. Lane, and T. Miyano. 2003. Role ofimmunologic costimulatory factors in the pathogenesis of biliary atresia. J. Pe-diatr. Surg. 38: 892–896.
42. Leiriao, P., S. S. Albuquerque, S. Corso, G. J. van Gemert, R. W. Sauerwein,A. Rodriguez, S. Giordano, and M. M. Mota. 2005. HGF/MET signalling protectsPlasmodium-infected host cells from apoptosis. Cell Microbiol. 7: 603–609.
43. van de Sand, C., S. Horstmann, A. Schmidt, A. Sturm, S. Bolte, A. Krueger,M. Lutgehetmann, J. M. Pollok, C. Libert, and V. T. Heussler. 2005. The liverstage of Plasmodium berghei inhibits host cell apoptosis. Mol. Microbiol. 58:731–742.
44. Renggli, J., M. Hahne, H. Matile, B. Betschart, J. Tschopp, and G. Corradin.1997. Elimination of P. berghei liver stages is independent of Fas (CD95/Apo-I)or perforin-mediated cytotoxicity. Parasite Immunol. 19: 145–148.
45. Doolan, D. L., and S. L. Hoffman. 2000. The complexity of protective immunityagainst liver-stage malaria. J. Immunol. 165: 1453–1462.
46. Mota, M. M., and A. Rodriguez. 2004. Migration through host cells: the first stepsof Plasmodium sporozoites in the mammalian host. Cell Microbiol. 6:1113–1118.
47. Hafalla, J. C., G. Sano, L. H. Carvalho, A. Morrot, and F. Zavala. 2002. Short-term antigen presentation and single clonal burst limit the magnitude of the CD8�
T cell responses to malaria liver stages. Proc. Natl. Acad. Sci. USA 99:11819–11824.
48. Hugel, F. U., G. Pradel, and U. Frevert. 1996. Release of malaria circumsporo-zoite protein into the host cell cytoplasm and interaction with ribosomes. Mol.Biochem. Parasitol. 81: 151–170.
49. Atkinson, C. T., M. Aikawa, S. B. Aley, and M. R. Hollingdale. 1989. Expressionof Plasmodium berghei circumsporozoite antigen on the surface of exoerythro-cytic schizonts and merozoites. Am. J. Trop. Med. Hyg. 41: 9–17.
50. Marti, M., R. T. Good, M. Rug, E. Knuepfer, and A. F. Cowman. 2004. Targetingmalaria virulence and remodeling proteins to the host erythrocyte. Science 306:1930–1933.
51. Hiller, N. L., S. Bhattacharjee, C. van Ooij, K. Liolios, T. Harrison,C. Lopez-Estrano, and K. Haldar. 2004. A host-targeting signal in virulence pro-teins reveals a secretome in malarial infection. Science 306: 1934–1937.
52. Savina, A., C. Jancic, S. Hugues, P. Guermonprez, P. Vargas, I. C. Moura,A. M. Lennon-Dumenil, M. C. Seabra, G. Raposo, and S. Amigorena. 2006.NOX2 controls phagosomal pH to regulate antigen processing during crosspre-sentation by dendritic cells. Cell 126: 205–218.
53. Kumar, K. A., G. Sano, S. Boscardin, R. S. Nussenzweig, M. C. Nussenzweig,F. Zavala, and V. Nussenzweig. 2006. The circumsporozoite protein is animmunodominant protective antigen in irradiated sporozoites. Nature 444:937–940.
7063The Journal of Immunology
by guest on August 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















