
Development Immunity Epstein-Barr Herpesvirus …iai.asm.org/content/17/1/28.full.pdf · 49 8,700...
8
INFECTION AND IMMUNITY, JUlY 1977, P. 28-35 Copyright © 1977 American Society for Microbiology Vol. 17, No. 1 Printed in U.S.A. Development of Cell-Mediated Immunity to Epstein-Barr Herpesvirus in Infectious Mononucleosis as Shown by Leukocyte Migration Inhibition P. K. LAI,I* M. P. ALPERS,2 AND E. M. MAcKAY-SCOLLAY Department of Microbiology, Perth Medical Centre, Shenton Park 6009, Western Australia Received for publication 20 September 1976 Eight patients with infectious mononucleosis, aged between 8 and 24, were studied for cell-mediated immunity by the in vitro leukocyte migration inhibi- tion test at acute and convalescent stages. Follow-up studies were also carried out at up to 4 months after clinical illness. Cell-mediated immunity to Epstein- Barr virus (EBV) in the peripheral leukocytes from these patients was absent or incipient in all cases during the acute phase, although it was present in lymphocytes from a biopsied lymph node obtained from one of the patients. In contrast, cell-mediated immunity to EBV was detected readily in peripheral leukocytes obtained during convalescence and in the follow-up studies. A block- ing factor that abrogated leukocyte migration inhibition induced by EBV anti- gen was detected in acute and convalescent sera obtained from six of eight patients, whereas serum antinuclear autoantibodies were detected in the two patients whose sera failed to block leukocyte migration inhibition. When sera were fractionated, this blocking effect was observed only in the serum im- munoglobulin G fractions. In follow-up studies, neither the blocking factor nor the antinuclear autoantibodies were found in the sera collected. It has been established that the Epstein-Barr herpesvirus (EBV) causes infectious mononu- cleosis, a benign lymphoproliferative illness (13). Epidemiological evidence suggests that the occurrence of clinical infectious mononucle- osis at the individual level and its prevalence at the population level are related inversely to the age of EBV infection (6, 7, 14, 18) and that serological status is an index of protection (6, 27), since the disease arises only in individuals who have no serological evidence of previous exposure to the virus. Cell-mediated immunity (CMI) is considered a primary factor in defense against infection with most viruses (1, 23). However,-transient anergy as a manifestation of generalized de- pression of CMI associated with a variety of viral diseases and administration of viral vac- cines has been reported (3, 25, 26, 28). In an earlier study (19) we were able to demonstrate the presence of CMI to EBV in nornal subjects who had serum antibody to EBV and showed that CMI to EBV in five patients with acute infectious mononucleosis was not detected, de- spite the presence of antibody to EBV capsid ' Present address: National Cancer Institute, Bethesda, MD 20014. 2 Present address: Papua New Guinea Institute of Medical Research, P.O. Box 60, Goroka, Papua, New Guinea. antigen, but appeared in the convalescent phase of the illness. A blocking factor to EBV was also found in the acute and convalescent sera from these patients but not in normal sub- jects with serum EBV antibody (19). The pres- ence of a blocking factor and the absence of CMI specific to EBV in acute infectious mononucleo- sis are of special interest, because the virus has also been incriminated in the pathogenesis of Burkitt's lymphoma and nasopharyngeal carci- noma (12, 15, 30). In this communication, we report on results obtained from studies carried out on another series of eight patients and confirm our findings reported earlier on CMI to EBV in patients with infectious mononucleosis (19). MATERIALS AND METHODS Subjects. Eight patients, aged between 8 and 24, with clinically diagnosed and hematologically con- firmed infectious mononucleosis (International Classification of Diseases 075) were studied for CMI to EBV by the leukocyte migration inhibition test. None of the subjects was receiving medication known to cause depression of CMI, and all had en- joyed prior good health. Hyperbilirubinemia was detected in three patients, and elevated serum transaminase levels were detected in four of them during the acute phase of illness. Heterophile anti- bodies were detected in five, and hepatosplenomeg- aly was detected in six, whereas lymphadenopathy 28 on August 27, 2018 by guest http://iai.asm.org/ Downloaded from
Transcript of Development Immunity Epstein-Barr Herpesvirus …iai.asm.org/content/17/1/28.full.pdf · 49 8,700...

INFECTION AND IMMUNITY, JUlY 1977, P. 28-35Copyright © 1977 American Society for Microbiology
Vol. 17, No. 1Printed in U.S.A.
Development of Cell-Mediated Immunity toEpstein-Barr Herpesvirus in Infectious Mononucleosis as
Shown by Leukocyte Migration InhibitionP. K. LAI,I* M. P. ALPERS,2 AND E. M. MAcKAY-SCOLLAY
Department of Microbiology, Perth Medical Centre, Shenton Park 6009, Western Australia
Received for publication 20 September 1976
Eight patients with infectious mononucleosis, aged between 8 and 24, werestudied for cell-mediated immunity by the in vitro leukocyte migration inhibi-tion test at acute and convalescent stages. Follow-up studies were also carriedout at up to 4 months after clinical illness. Cell-mediated immunity to Epstein-Barr virus (EBV) in the peripheral leukocytes from these patients was absent orincipient in all cases during the acute phase, although it was present inlymphocytes from a biopsied lymph node obtained from one of the patients. Incontrast, cell-mediated immunity to EBV was detected readily in peripheralleukocytes obtained during convalescence and in the follow-up studies. A block-ing factor that abrogated leukocyte migration inhibition induced by EBV anti-gen was detected in acute and convalescent sera obtained from six of eightpatients, whereas serum antinuclear autoantibodies were detected in the twopatients whose sera failed to block leukocyte migration inhibition. When serawere fractionated, this blocking effect was observed only in the serum im-munoglobulin G fractions. In follow-up studies, neither the blocking factor northe antinuclear autoantibodies were found in the sera collected.
It has been established that the Epstein-Barrherpesvirus (EBV) causes infectious mononu-cleosis, a benign lymphoproliferative illness(13). Epidemiological evidence suggests thatthe occurrence of clinical infectious mononucle-osis at the individual level and its prevalence atthe population level are related inversely to theage of EBV infection (6, 7, 14, 18) and thatserological status is an index of protection (6,27), since the disease arises only in individualswho have no serological evidence of previousexposure to the virus.Cell-mediated immunity (CMI) is considered
a primary factor in defense against infectionwith most viruses (1, 23). However,-transientanergy as a manifestation of generalized de-pression of CMI associated with a variety ofviral diseases and administration of viral vac-cines has been reported (3, 25, 26, 28). In anearlier study (19) we were able to demonstratethe presence of CMI to EBV in nornal subjectswho had serum antibody to EBV and showedthat CMI to EBV in five patients with acuteinfectious mononucleosis was not detected, de-spite the presence of antibody to EBV capsid
' Present address: National Cancer Institute, Bethesda,MD 20014.
2 Present address: Papua New Guinea Institute ofMedical Research, P.O. Box 60, Goroka, Papua, NewGuinea.
antigen, but appeared in the convalescentphase of the illness. A blocking factor to EBVwas also found in the acute and convalescentsera from these patients but not in normal sub-jects with serum EBV antibody (19). The pres-ence ofa blocking factor and the absence ofCMIspecific to EBV in acute infectious mononucleo-sis are of special interest, because the virus hasalso been incriminated in the pathogenesis ofBurkitt's lymphoma and nasopharyngeal carci-noma (12, 15, 30).
In this communication, we report on resultsobtained from studies carried out on anotherseries ofeight patients and confirm our findingsreported earlier on CMI to EBV in patientswith infectious mononucleosis (19).
MATERIALS AND METHODSSubjects. Eight patients, aged between 8 and 24,
with clinically diagnosed and hematologically con-firmed infectious mononucleosis (InternationalClassification of Diseases 075) were studied for CMIto EBV by the leukocyte migration inhibition test.None of the subjects was receiving medicationknown to cause depression of CMI, and all had en-joyed prior good health. Hyperbilirubinemia wasdetected in three patients, and elevated serumtransaminase levels were detected in four of themduring the acute phase of illness. Heterophile anti-bodies were detected in five, and hepatosplenomeg-aly was detected in six, whereas lymphadenopathy
28
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CMI TO EBV IN INFECTIOUS MONONUCLEOSIS 29
and general malaise were found in all patients. Mul-tiple heparinized blood samples were collected at theacute stage and at convalescence, when all clinicalsymptoms and hematological abnormalities had dis-appeared. In one case, leukocytes derived from a
cervical lymph node biopsy were available at theacute stage of the disease. All patients were testedfor CMI to EBV repeatedly in follow-up studies forup to 4 months after the clinical illness. In addition,six normal subjects, aged between 18 and 26, werestudied. Four were randomly chosen from a group ofmedical students volunteering for this study,whereas those remaining were selected because theydid not have antibody (seronegative) to EBV. At thetime of the study, all were enjoying sound healthand none had a history of drug addiction.
Leukocyte migration inhibition test. Freshly col-lected, heparinized peripheral blood samples wereallowed to sediment by gravity for 1 h at 37°C.Dextran was used to assist sedimentation when sep-aration of erythrocytes from plasma was insignifi-cant after 30 min. The leukocyte-rich plasma frac-tions were collected and centrifuged at 800 x g for 5min. After washing three times in Hanks balancedsalt solution containing heparin (100 IU/ml), theleukocytes were suspended in growth medium(RPMI-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, standard amountsof antibiotics [penicillin, 100 IU/ml; streptomycin,100 ,ug/ml], and 2% of a 1.4 M N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid solution) at a
concentration of 107 cells per ml. Leukocyte migra-tion inhibition was performed according to Lai et al.(16), using a crude EBV antigen prepared fromP3HR-1 cells (16), and the area of leukocyte migra-tion for each tube was measured (19) without know-ing its origin. The percentage of inhibition wascalculated according to Lai et al. (19), and its sig-nificance was determined statistically.
Test for serum blocking factor. Sera collected atthe acute stage, at convalescence, and in the follow-up studies were tested for the presence of blockingfactor. Twofold dilutions ofeach serum sample weremade, beginning with 1:5, and each serum dilutionwas mixed with EBV antigen or control antigen.The mixtures were used to challenge heterologousand autologous leukocytes after a briefincubation at37°C, as described by Lai et al. (19).
Fractionation of serum. Three of the eight seracollected from patients with acute infectious mono-
nucleosis were fractionated into immunoglobulin G(IgG) and IgM fractions in a column (2.5 by 100 cm;Pharmacia, Uppsala, Sweden), using Sephadex G200 (Pharmacia), at a flow rate of 20 ml/h. Thefractions were then examined at 280 nm in a spectro-photometer (Beckman Instruments, Purley, UnitedKingdom) before being used in immunodiffusion andimmunofluorescent and immunoelectroosmophoreticstudies. Presence of blocking factor in each fractionwas tested for as described.
Immunodiffusion. Immunodiffusion of serumfractions was carried out by using commerciallyavailable immunoplates (Hyland Laboratories, Inc.,Costa Mesa, Calif.). In addition, each fraction wastested against rabbit antihuman IgG and rabbit
antihuman IgM (Behringwerke, A.G., Marburg/Lahn, W. Germany) according to the method ofOuchterlony (24) in 0.8% agarose (Seakem agarose,Marine Colloids, Inc., Springfield, N.J.) buffered atpH 7.6.
Detection of serum EBV antibody by immuno-fluorescence. The presence of IgM and IgG anti-bodies to EBV in serum samples and in serum frac-tions was determined by indirect immunofluores-cence (11, 18), using SH-RP cells (20) previouslyultraviolet irradiated for the enhancement of viruscapsid antigen (17).
Immunoelectroosmophoresis. Immunoelectroos-mophoresis of serum IgG fractions was carried outaccording to Ouchterlony (24) with rabbit antihu-man serum and antihuman IgG (Behringwerke) in0.8% agarose buffered at pH 7.3, using a Vokampower pack (Shendon Scientific, United Kingdom)and a Kohn tank (Shendon Scientific) with constantcurrent at 8 A.Serum ANF. The presence of antinuclear antibod-
ies (ANF) in serum samples was tested for by indi-rect immunofluorescence, using acetone-fixedsmears of EBV-nonproducing Raji cells and humanembryonic lung fibroblasts grown in tissue culture.
RESULTS
Migration inhibition response to antigens.Peripheral leukocytes from normal subjects ser-
opositive to EBV responded to EBV antigen bymarked inhibition (.20%), as determined sta-tistically in leukocyte migration inhibitiontests, whereas peripheral leukocytes fromEBV-seronegative adults failed to respond (Ta-ble 1). In contrast, peripheral leukocytes frompatients with infectious mononucleosis col-lected during the acute phase of the disease didnot respond to EBV antigen (Tables 2 and 3),
TABLE 1. Leukocyte migration-inhibitory response
of circulating leukocytes from seropositive andseronegative normal subjects to EBV antigen
SeuIGt- .Lekocyte migrationNormal subjects Steru"mtoEBV inhibition (mean %Normal tsrs6 to EBV ± SE)b
11 10 72.3 ± 2.8 (+)12 40 37.4 ± 1.7 (+)13 160 46.5 ± 1.4 (+)14 10 31.2 ± 2.1 (+)15 0 -3.7 ± 2.1 (-)16 0 9.8 ± 2.3 (-)
a Assayed by indirect immunofluorescence (11,18), using SH-RP cells (17, 20).
b Freshly collected leukocytes were used in theleukocyte migration inhibition test. Mean percentinhibition ± standard error (SE) was determinedfrom a minimum of four replicate cultures. Twentypercent was considered positive, and <15% was con-sidered negative. Peripheral leukocytes from allsubjects failed to respond (<15%) to control antigenfrom Raji cells.
VOL. 17, 1977
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
30 LAI, ALPERS, AND MAcKAY-SCOLLAY
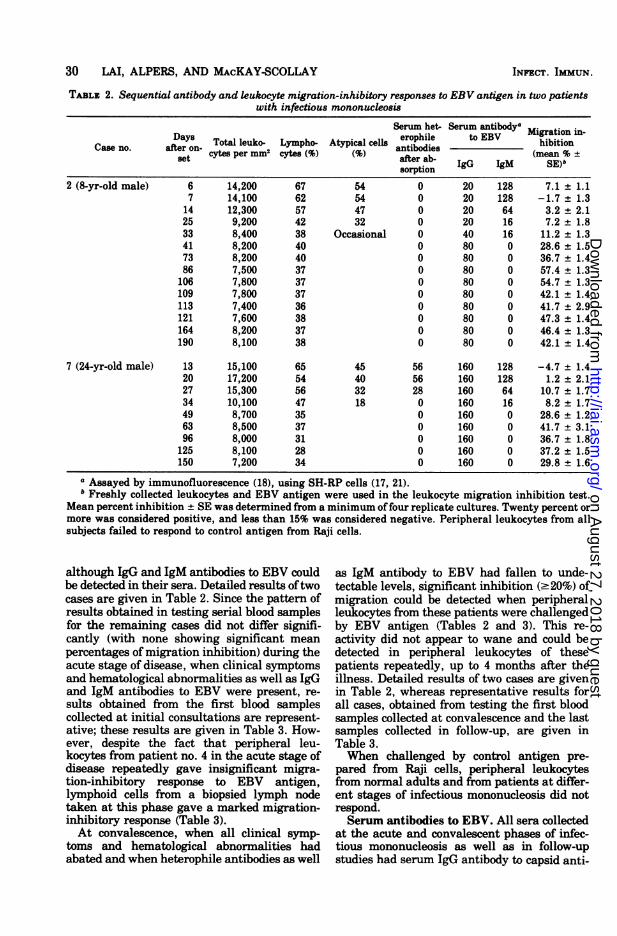
TABLE 2. Sequential antibody and leukocyte migration-inhibitory responses to EBV antigen in two patientswith infectious mononucleosis
Serum het- Serum antibodya
Da.ay Total leuko- Lympho- Atypical cells eropile to EBV_hibitionCase no. after on-ye
antibodies(m %±t cytes per mm cytes ( after ab- (mean %S
sorption2 (8-yr-old male) 6 14,200 67 54 0 20 128 7.1 ± 1.1
7 14,100 62 54 0 20 128 -1.7 ± 1.314 12,300 57 47 0 20 64 3.2 + 2.125 9,200 42 32 0 20 16 7.2 1.833 8,400 38 Occasional 0 40 16 11.2 ± 1.341 8,200 40 0 80 0 28.6 ± 1.573 8,200 40 0 80 0 36.7 ± 1.486 7,500 37 0 80 0 57.4 ± 1.3106 7,800 37 0 80 0 54.7 ± 1.3109 7,800 37 0 80 0 42.1 ± 1.4113 7,400 36 0 80 0 41.7 ± 2.9121 7,600 38 0 80 0 47.3 ± 1.4164 8,200 37 0 80 0 46.4 ± 1.3190 8,100 38 0 80 0 42.1 ± 1.4
7 (24-yr-old male) 13 15,100 65 45 56 160 128 -4.7 ± 1.420 17,200 54 40 56 160 128 1.2 ± 2.127 15,300 56 32 28 160 64 10.7 ± 1.734 10,100 47 18 0 160 16 8.2 ± 1.749 8,700 35 0 160 0 28.6 ± 1.263 8,500 37 0 160 0 41.7 ± 3.196 8,000 31 0 160 0 36.7 ± 1.8125 8,100 28 0 160 0 37.2 ± 1.5150 7,200 34 0 160 0 29.8 ± 1.6
a Assayed by immunofluorescence (18), using SH-RP cells (17, 21).b Freshly collected leukocytes and EBV antigen were used in the leukocyte migration inhibition test.
Mean percent inhibition ± SE was determined from a minimum offour replicate cultures. Twenty percent ormore was considered positive, and less than 15% was considered negative. Peripheral leukocytes from allsubjects failed to respond to control antigen from Raji cells.
although IgG and IgM antibodies to EBV couldbe detected in their sera. Detailed results oftwocases are given in Table 2. Since the pattern ofresults obtained in testing serial blood samplesfor the remaining cases did not differ signifi-cantly (with none showing significant meanpercentages of migration inhibition) during theacute stage of disease, when clinical symptomsand hematological abnormalities as well as IgGand IgM antibodies to EBV were present, re-sults obtained from the first blood samplescollected at initial consultations are represent-ative; these results are given in Table 3. How-ever, despite the fact that peripheral leu-kocytes from patient no. 4 in the acute stage ofdisease repeatedly gave insignificant migra-tion-inhibitory response to EBV antigen,lymphoid cells from a biopsied lymph nodetaken at this phase gave a marked migration-inhibitory response (Table 3).At convalescence, when all clinical symp-
toms and hematological abnormalities hadabated and when heterophile antibodies as well
as IgM antibody to EBV had fallen to unde-tectable levels, significant inhibition (.20%) ofmigration could be detected when peripheralleukocytes from these patients were challengedby EBV antigen (Tables 2 and 3). This re-activity did not appear to wane and could bedetected in peripheral leukocytes of thesepatients repeatedly, up to 4 months after theillness. Detailed results of two cases are givenin Table 2, whereas representative results forall cases, obtained from testing the first bloodsamples collected at convalescence and the lastsamples collected in follow-up, are given inTable 3.When challenged by control antigen pre-
pared from Raji cells, peripheral leukocytesfrom normal adults and from patients at differ-ent stages of infectious mononucleosis did notrespond.Serum antibodies to EBV. All sera collected
at the acute and convalescent phases of infec-tious mononucleosis as well as in follow-upstudies had serum IgG antibody to capsid anti-
INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CMI TO EBV IN INFECTIOUS MONONUCLEOSIS
TABLE 3. Antibody and leukocyte migration-inhibitory response to EBV antigen in patients with infectiousmononucleosis
Serum antibody to Paul-BunnellCase of infec- EBVa heterophile an- Migration inhibi-tious mononu- Stage Days after on- tibody titers tionb (mean % +set after absorption SE)
IgG IgM with guinea pigkidney
1 Acutec 24 80 64 0 4.2 ± 1.3Convalescentd 64 80 0 0 32.7 ± 2.1Follow-upe 179 80 0 0 31.4 ± 1.7
2 Acute 6 20 128 0 7.1 ± 1.1Convalescent 4i 80 0 0 28.6 ± 1.5Follow-up 190 80 0 0 42.1 ± 1.4
3 Acute 15 40 32 84 16.2 ± 2.3Convalescent 84 40 0 0 37.2 ± 2.1Follow-up 110 40 0 0 39.8 ± 1.7
4f Acute 11 320 32 84 4.3 ± 1.5Convalescent 92 160 0 0 24.1 ± 2.4Follow-up 222 160 0 0 32.7 ± 1.6
5 Acute 16 320 16 280 6.7 ± 1.9Convalescent 65 320 0 0 47.4 ± 1.4Follow-up 145 320 0 0 46.4 ± 2.0
6 Acute 10 10 128 0 2.1 ± 1.1Convalescent 69 640 0 0 33.1 ± 1.7Follow-up 140 320 0 0 30.6 ± 1.9
7 Acute 13 160 128 56 -4.7 ± 1.4Convalescent 49 160 0 0 28.6 ± 1.2Follow-up 150 160 0 0 29.8 ± 1.6
8 Acute 18 40 64 56 6.7 ± 2.4Convalescent 60 40 0 0 52.7 ± 3.1Follow-up 285 40 0 0 38.7 ± 2.8
a Assayed by immunofluorescence (18), using SH-RP cells (17, 21).b Freshly collected leukocytes and EBV antigen were used in the leukocyte migration inhibition test.
Mean percent inhibition ± SE was determined from a minimum of four replicate cultures. Twenty percentwas considered positive, 15% was considered negative, and in-between was considered equivocal. Peripheralleukocytes from all subject failed to respond to control antigen from Raji cells.
c Acute infectious mononucleosis is defined clinically and hematologically in the presence of specific IgMto EBV and/or Paul-Bunnell heterophile antibodies. Results are those obtained from the first blood samplescollected at initial consultations.
d Convalescence is defined by the absence of clinical, hematological, and serological evidence of infectiousmononucleosis.
e Follow-up studies were carried out up to 4 months after clinical illness. Results are those obtained fromthe last blood samples.
f Leukocytes from a lymph node biopsy of this patient at the acute phase of infectious mononucleosistested against EBV antigen and control antigen gave mean percentages of inhibition of 24.3 ± 1.8 and 7.2 ±1.3, respectively.
gen of EBV. In two of the patients, a signifi-cant rise in IgG titers could also be detected.Serum IgM antibody to EBV was detected inall acute sera; this varied in individuals andranged from 1:16 to 1:128 (Table 3). However,there is no correlation between the levels ofspecific antibodies (IgG and IgM) to EBV and
the level of leukocyte migration-inhibitory re-sponse (Tables 2 and 3).ANF. ANF was detected in two of the eight
patients with acute infectious mononucleosis(cases 4 and 5, Table 4). Nuclear staining waspatchy or speckled, and, occasionally, periph-eral or rim nuclear staining was observed. Se-
31VOL. 17, 1977
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
32 LAI, ALPERS, AND MACKAY-SCOLLAY
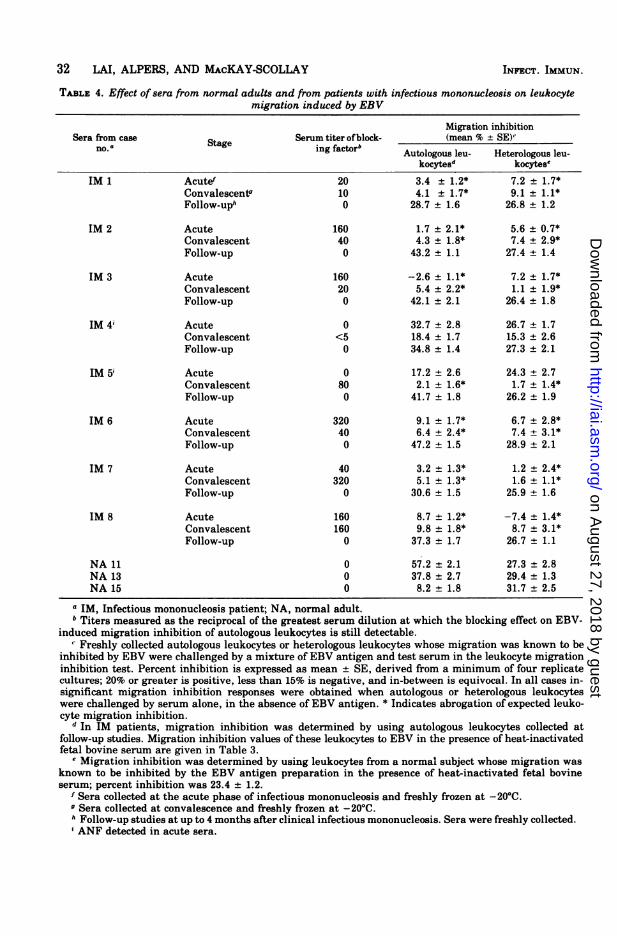
TABLE 4. Effect of sera from normal adults and from patients with infectious mononucleosis on leukocytemigration induced by EBV
Migration inhibitionSera from case Serum titer of block- (mean % ± SE)'no.naStage ing factorb Autologous leu- Heterologous leu-
kocytesd kocytese
IM 1 Acutef 20 3.4 ± 1.2* 7.2 ± 1.7*Convalescentg 10 4.1 ± 1.7* 9.1 + 1.1*Follow-uph 0 28.7 ± 1.6 26.8 ± 1.2
IM 2 Acute 160 1.7 ± 2.1* 5.6 ± 0.7*Convalescent 40 4.3 ± 1.8* 7.4 ± 2.9*Follow-up 0 43.2 + 1.1 27.4 ± 1.4
IM 3 Acute 160 -2.6 ± 1.1* 7.2 ± 1.7*Convalescent 20 5.4 ± 2.2* 1.1 ± 1.9*Follow-up 0 42.1 ± 2.1 26.4 ± 1.8
IM 4i Acute 0 32.7 ± 2.8 26.7 + 1.7Convalescent <5 18.4 ± 1.7 15.3 ± 2.6Follow-up 0 34.8 ± 1.4 27.3 ± 2.1
IM 5i Acute 0 17.2 ± 2.6 24.3 + 2.7Convalescent 80 2.1 ± 1.6* 1.7 ± 1.4*Follow-up 0 41.7 ± 1.8 26.2 ± 1.9
IM 6 Acute 320 9.1 ± 1.7* 6.7 ± 2.8*Convalescent 40 6.4 ± 2.4* 7.4 ± 3.1*Follow-up 0 47.2 ± 1.5 28.9 ± 2.1
IM 7 Acute 40 3.2 + 1.3* 1.2 ± 2.4*Convalescent 320 5.1 ± 1.3* 1.6 ± 1.1*Follow-up 0 30.6 ± 1.5 25.9 ± 1.6
IM 8 Acute 160 8.7 ± 1.2* -7.4 + 1.4*Convalescent 160 9.8 ± 1.8* 8.7 + 3.1*Follow-up 0 37.3 ± 1.7 26.7 ± 1.1
NA 11 0 57.2 ± 2.1 27.3 ± 2.8NA 13 0 37.8 ± 2.7 29.4 ± 1.3NA 15 0 8.2 + 1.8 31.7 2.5
a IM, Infectious mononucleosis patient; NA, normal adult.b Titers measured as the reciprocal of the greatest serum dilution at which the blocking effect on EBV-
induced migration inhibition of autologous leukocytes is still detectable." Freshly collected autologous leukocytes or heterologous leukocytes whose migration was known to be
inhibited by EBV were challenged by a mixture of EBV antigen and test serum in the leukocyte migrationinhibition test. Percent inhibition is expressed as mean ± SE, derived from a minimum of four replicatecultures; 20% or greater is positive, less than 15% is negative, and in-between is equivocal. In all cases in-significant migration inhibition responses were obtained when autologous or heterologous leukocyteswere challenged by serum alone, in the absence of EBV antigen. * Indicates abrogation of expected leuko-cyte migration inhibition.
d In IM patients, migration inhibition was determined by using autologous leukocytes collected atfollow-up studies. Migration inhibition values of these leukocytes to EBV in the presence of heat-inactivatedfetal bovine serum are given in Table 3.
e Migration inhibition was determined by using leukocytes from a normal subject whose migration wasknown to be inhibited by the EBV antigen preparation in the presence of heat-inactivated fetal bovineserum; percent inhibition was 23.4 ± 1.2.
f Sera collected at the acute phase of infectious mononucleosis and freshly frozen at -20°C.o Sera collected at convalescence and freshly frozen at -20°C.h Follow-up studies at up to 4 months after clinical infectious mononucleosis. Sera were freshly collected.i ANF detected in acute sera.
INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CMI TO EBV IN INFECTIOUS MONONUCLEOSIS 33
rum titers of ANF (IgG and IgM) in the twopatients were low, at 1:8 (case 5) and 1:16 (case4); treatment of the sera with mercaptoethanolresulted in decreases in serum titers but failedto abolish nuclear staining.When tested at convalescence and again in
follow-up studies up to 4 months after clinicalillness, ANF was not detected by indirect im-munofluorescence in sera from any patient.Serum blocking factor. Ofthe eight patients
tested, a blocking effect on EBV-induced migra-tion inhibition of autologous and heterologousleukocytes was found in six sera collected at theacute phase, in seven collected at convales-cence, and in none of the sera collected in thefollow-up studies. Detailed results are given inTable 4. The titer of blocking factor in eachserum collected from the eight patients withinfectious mononucleosis is given in Table 4.The titers of blocking factor do not correlatewith serum antibody titers to EBV (Tables 3and 4). Sera from normal adults did not showany blocking effect on migration inhibition ofleukocytes whose migration was known to beinhibited by EBV antigen in the presence offetal bovine serum (Table 4).When IgG and IgM fractions of acute sera
collected from three of the eight patients weretested, a blocking effect on autologous leuko-cyte migration inhibition induced by the EBVantigen was observed in the IgG fractions, butnot in the IgM fractions.Immunodiffusion and immunoelectroosmo-
phoresis. Immunodiffusion of IgG and IgMfractions in immunoplates or against antihu-man IgG and antihuman IgM standard serashowed that the IgG fractions reacted only withantihuman IgG and not with antihuman IgM.Similarly, the IgM fractions reacted with anti-human IgM only. However, when the IgG frac-tions were tested with antihuman IgG and anti-
human serum by immunoelectroosmophoresis,multiple components were observed (Fig. 1).
DISCUSSIONThe evidence that infectious mononucleosis
is caused by EBV is now extensive, but thebasis for the self-limiting course of the illness isstill unknown.
In an earlier study, we found that CMI toEBV in the peripheral leukocytes of five pa-tients with acute infectious mononucleosis wasnot detected by the leukocyte migration inhibi-tion test, but became detectable in convales-cence (19). In this study, we confirm and extendthis finding in another series of eight patients.The results suggest that CMI to EBV persistsfor a long period, at least up to 4 months afterclinical illness, and probably for life, since itcan be detected consistently in EBV-seroposi-tive adults, as shown in Table 1 and in previ-ous reports (8, 16; P. K. Lai, E. M. MacKay-Scollay, and M. P. Alpers, Proc. Aust. Soc.Med. Res., p. 151, 1973).In the present study, the absence of migra-
tion-inhibitory response to EBV in peripheralleukocytes, together with the temporaryanergy reported by others (2, 10, 21) duringacute infectious mononucleosis, suggests thepossibility of generalized immunosuppression.This, however, is difficult to reconcile with thehistopathology of lymphoid tissues (4) duringthe acute phase of infectious mononucleosis.The temporary anergy in these patients may, ofcourse, be explained by antigenic competitionin the presence of excessive EBV antigen, andthe absence ofCMI to EBV in peripheral leuko-cytes does not preclude its presence in otherhost tissues. In fact, we have demonstrated amigration-inhibitory response to EBV in lymphnode leukocytes obtained from one of the pa-tients (case 4, Table 3) at a time when it was
. '4 ..s....
0 I gG frac"t io:4n
Ant uman serum A
FIG. 1. Immunoelectroosmophoresis ofserum IgG fraction against antihuman IgG and antihuman serumin 0.8% agarose atpH 7.3. The slide was photographed after washing in saline and staining with a2% amidoblack solution.
VOL. 17, 1977
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

34 LAI, ALPERS, AND MAcKAY-SCOLLAY
undetected in circulating leukocytes. This find-ing is consistent with the morphologicalchanges in lymph nodes in infectious mononu-cleosis (4), in which the paracortex, a thymus-dependent zone not primarily involved in anti-body formation, is found to be hyperactive.Although the migration inhibition test has
been reported to correlate well with delayed-type hypersensitivity (9) and involves the pro-duction ofa migration inhibition factor (16, 29),it is still an open question whether cellularimmune reaction detected by this assay is ex-clusively a T cell fumction. We are aware of theshortcomings in using a single in vitro correlateto detect CMI and are, therefore, developing anantigen-stimulated blastogenic test as well asan interferon release assay to complement ourstudy on cellular immunity to EBV in patientswith infectious mononucleosis.
Intense antibody fornation in infectiousmononucleosis has long been recognized. Hu-moral antibodies include heterophile antibod-ies, EBV antibodies, and a battery of hetero-,iso-, and autoantibodies. This intense antibodyresponse is unique among infectious diseasesand is equalled only occasionally in Sjogren'sdisease and systemic lupus erythematosus (5).However, the role of these antibodies, espe-cially EBV-associated antibodies, in the controlof infectious mononucleosis is obscure.
Recently, a blocking factor that abrogatesCMI to EBV detected by the leukocyte migra-tion inhibition test has been described in seracollected from patients with infectious mononu-cleosis (19). This serum blocking effect is prob-ably specific to EBV, since it fails to negateCMI response to an unrelated antigen such asone prepared from Escherichia coli (19). In thepresent study, we have demonstrated thatacute and convalescent sera collected from pa-tients with infectious mononucleosis were ableto abrogate in vitro leukocyte migration-inhibi-tory response to EBV, whereas sera collectedfrom the same patients in follow-up studies upto 4 months after the illness failed to blockEBV-induced leukocyte migration inhibition. Itis impossible to say whether this blocking fac-tor is simply an IgG antibody or, more gen-erally, whether it is a product of infected ornormal B cells, since although it is found in theIgG fraction, immunoelectroosmophoretic studyof the fraction gave multiple precipitationlines.The precise function ofthis blocking factor in
vivo is unknown, although it interferes withleukocyte migration inhibition in vitro, asshown in the present study. It is thus not clearwhether this blocking action demonstrated invitro contributes to the intensive but self-limit-
ing lymphoproliferation seen in infectiousmononucleosis in vivo.In acute viral hepatitis, it has been postu-
lated that a hyperactive T cell response is re-sponsible for the development offulminant hep-atitis in 1% of patients and results in severeliver cell necrosis, whereas impaired T cellfunction is responsible for the development of ahepatitis B antigen carrier state in 10% of pa-tients and gives rise to slow, progressive,chronic liver disease (22). It is possible that theblocking factor detected in this study is part ofthe normal negative feedback mechanism in-volved in immunity to virus infection. Destruc-tion of EBV-carrying cells in vivo may result inthe release of autoantigen, and this processmay have been enhanced by the absence ofblocking-factor production as well as by a re-duced efficiency of normal control mechanismagainst autoantibody formation (cases 4 and 5,Table 4).The interrelationship between the humoral
and cell-mediated components of immunity isobviously complex, and further information onthis delicate immunological balance in EBVinfection may help elucidate the pathogenesisof EBV-related diseases and other persistentviral infections.
ACKNOWLEDGMENTSWe thank F. Deinhardt and L. Wolfe for their discus-
sions and advice and P. Lowder for assistance in prepara-tion of the manuscript.
This work was supported by a collaborative agreementbetween the Biological Carcinogenesis Unit, InternationalAgency for Research on Cancer, World Health Organiza-tion, and the Department of Microbiology, University ofWestern Australia, and by Public Health Service Contractno. N01 CP 43296 from the Division of Cancer Cause andPrevention within the Virus-Cancer Program of the Na-tional Cancer Institute. At the time of writing, P. K. L. wasin receipt of a Research Fellowship from the InternationalAgency for Research on Cancer, World Health Organiza-tion, Lyons, France. M. P. A. was a Senior Research Fel-low of the National Health and Medical Research Councilof Australia. E. M. M.-S. would like to thank the Com-missioner of Health of Western Australia for permissionto publish.
LITERATURE CITED1. Allison, A. C. 1967. Cell-mediated immune responses to
virus infections and virus-induced tumours. Br. Med.Bull. 23:60-65.
2. Bentzon, J. W. 1953. The effect of certain infectiousdiseases on tuberculin allergy. Tubercle 34:34-41.
3. Brody, J. A., T. Overfield, and L. M. Hammes. 1964.Depression of the tuberculin reaction by viral vac-cines. N. Engl. J. Med. 271:1294-1296.
4. Carter, R. L. 1975. Infectious mononucleosis: model forself-limiting lymphoproliferation. Lancet i:846-849.
5. Dameshek, W. 1969. Speculations on the nature of in-fectious mononucleosis, p. 225-250. In R. L. Carterand H. G. Penman (ed.), Infectious mononucleosis.Blackwell Scientific Inc., Oxford.
6. Evans, A. S., and J. C. Niederman. 1972. Epidemiologyof infectious mononucleosis-a review, p. 351-356. In
INFECT. IMMUN.
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

CMI TO EBV IN INFECTIOUS MONONUCLEOSIS 35
P. M. Biggs, G. de-The, and L. N. Payne (ed.), Onco-genesis and herpesviruses. International Agency forResearch on Cancer, Lyons, France.
7. Evans, A. S., J. C. Niederman, and R. W. McCollum.1968. Seroepidemiologic studies ofinfectious mononu-cleosis with EB virus. N. Engl. J. Med. 274:1121-1127.
8. Fimmel, P. J., P. K. Lai, D. Keast, E. M. MacKay-Scollay, and M. P. Alpers. 1974. In vitro cellularimmunity to Epstein-Barr virus in normal humansubjects. Int. J. Cancer 13:599-05.
9. George, M., and J. H. Vaughn. 1962. In vitro cell migra-tion as a model for delayed hypersensitivity. Proc.Soc. Exp. Biol. Med. 111:514-51.
10. Haider, S., M. D. Continho, and R. T. D. Emond. 1973.Tuberculin anergy and infectious mononudleosis.Lancet ii:74.
11. Henle, G., and W. Henle. 1966. Immunofluorescence incells derived from Burkitt's lymphoma. J. Bacteriol.91:1248-1256.
12. Henle, G., W. Henle, and V. Diehl. 1968. Relation ofBurkitt's tumour associated herpes-type virus to in-fectious mononudleosis. Proc. Natl. Acad. Sci. U.S.A.59:94-101.
13. Henle, W. and G. Henle. 1972. Epstein-Barr virus: thecause of infectious mononucleosis-a review, p. 269-274. In P. M. Biggs, G. de-The, and L. N. Payne (ed.),Oncogenesis and herpesviruses. InternationalAgency for Research on Cancer, Lyons, France.
14. Hubble, M. P., P. K. Lai, E. M. MacKay-Scollay, andM. P. Alpers. 1974. Epidemiology of infectious mono-nucleosis in Western Australia. A hospital survey,1966-1972. Med. J. Aust. 2:863-66.
15. Klein, G. 1973. The Epstein-Barr virus, p. 521-550. InA. Kaplan (ed.), The herpeaviruses. Academic PressInc., New York.
16. Lai, P. K., M. P. Alpers, and E. M. MacKay-Scollay.1975. In vitro evaluation of cell-mediated immunity toEpstein-Barr herpesvirms by cell migration inhibitiontests. J. Natl. Cancer Inst. 55:1319-1322.
17. Lai, P. K., E. M. MacKay-Scollay, and M. P. Alpers.1973. Synthesis of virus-capsid antigen (VCA) en-hanced by ultraviolet irradiation of a lymphoblastoidcell line carrying Epstein-Barr virus. J. Gen. Virol.21:135-143.
18. Lai, P. K., E. M. MacKay-Scollay, and M. P. Alpers.
1975. Epidemiological studies of Epstein-Barr herpes-virus infection in Western Australia. J. Hyg. 74:329-338.
19. Lai, P. K., E. M. MacKay-Scollay, P. J. Fimmel, M. P.Alpers, and D. Keast. 1974. Cell-mediated immunityto Epstein-Barr virus and a blocking factor in pa-tients with infectious mononucleosis. Nature (Lon-don) 252:608-610.
20. Lai, P. K., J. M. Papadimitriou, D. W. G. Kennett, E.M. MacKay-Scollay, and M. P. Alpers. 1973. A lym-phoblastoid cell line derived from cells of myeloidleukaemia by infection with Epstein-Barr herpesvi-rus. Cytobios 8:125-138.
21. Mangi, R. J., J. C. Niederman, J. E. Kelleher, Jr., J. M.Dwyer, A. S. Evans, and F. S. Kantor. 1974. Depres-sion of cell-mediated immunity during acute infec-tious mononucleosb. N. Engl. J. Med. 291:1149-1153.
22. Nielsen, J. 0. 1976. Clinical course and prognosis ofacute hepatitis. Ann. Clin. Res. 8:151-157.
23. Notkins, A. L. 1974. Immune mechanisms by which thespread of viral infections is stopped. Cell. Immunol.11:478-483.
24. Ouchterlony, 0. 1968. Handbook of immunodiffusionand immunoelectrophoresis. Ann Arbor Science Pub-lishers, London.
25. Reed, C. P., J. W. Olds, and A. L. Kisch. 1972. De-creased skin-test reactivity associated with influ-enza. J. Infect. Dis. 125:398-402.
26. Salaman, M. H. 1970. Immunodepression by mamma-lian viruses and plasmodia. Proc. R. Soc. Med. 63:11-15.
27. Sawyer, R. N., A. S. Evans, J. C. Niederman, and R. W.McCollum. 1971. Prospective studies of a group ofYale University freshmen. I. Occurrence of infectiousmononucleosis. J. Infect. Dis. 123:263-270.
28. Starr, S., and S. Berkovich. 1964. The depression oftuberculin reactivity during chickenpox. Pediatrics33:769-772.
29. Thor, D. E., R. E. Jureziz, S. R. Veach, et al. 1968. Cellmigration inhibition factor released by antigen fromhuman peripheral lymphocytes. Nature (London)219:755-757.
30. Wolf, H., H. ZurHausen, and V. Becker. 1973. EB viralgenomes in epithelial nasopharyngeal carcinomacells. Nature (London) New Biol. 244:245-247.
VOL. 17, 1977
on August 27, 2018 by guest
http://iai.asm.org/
Dow
nloaded from


















