Correlation-Induced Oscillations in Spatio-Temporal Excitable Systems Andre Longtin Physics...
31
Correlation-Induced Oscillations in Spatio-Temporal Excitable Systems Andre Longtin Physics Department, University of Ottawa Ottawa, Canada
-
Upload
sylvia-strickland -
Category
Documents
-
view
214 -
download
0
Transcript of Correlation-Induced Oscillations in Spatio-Temporal Excitable Systems Andre Longtin Physics...

Correlation-Induced Oscillations in Spatio-Temporal Excitable Systems
Andre LongtinPhysics Department, University of Ottawa
Ottawa, Canada

Co-Workers
Brent Doiron
Benjamin Lindner
Maurice Chacron Physics Department, University of Ottawa
Leonard MalerDepartment of Cellular and Molecular Medicine,
University of Ottawa
Joseph BastianDeparment of Zoology, University of Oklahoma

Synopsis
Introduction to weakly electric fish Oscillatory activity for communication but not Prey StimuliModeling I: Feedback is required Experimental verificationModeling II: stochastic oscillatory dynamics in a spatially extended neural system
Doiron, Chacron, Maler, Longtin and Bastian, Nature 421 (Jan 30, 2003)

Weakly Electric Fish

Why study weakly electric fish? (Biology)
from molecular to behavioral studies of neural codingperipheral ↔ centralfeedforward ↔ feedbackin vivo ↔ in vitroStimuli: simple (sines etc…) ↔ naturalbehaviors: simple ↔ evolved
(electrolocation) (electrocommunication)

Why study weakly electric fish?(Mathematical Biology/Biophysics)
Single cell dynamics: simple ↔ complex
Linear, nonlinear, stochastic (get ready for noise!)
Information processing:
black box ↔ detailed biophysics
Math, Physics, Neuroscience, Computation
Applications: signal detection, novel circuitry, prosthetic design (e.g. with feedback)

Sensory Neurons
ELL Pyramidal Cell
Sensory Input
Higher Brain
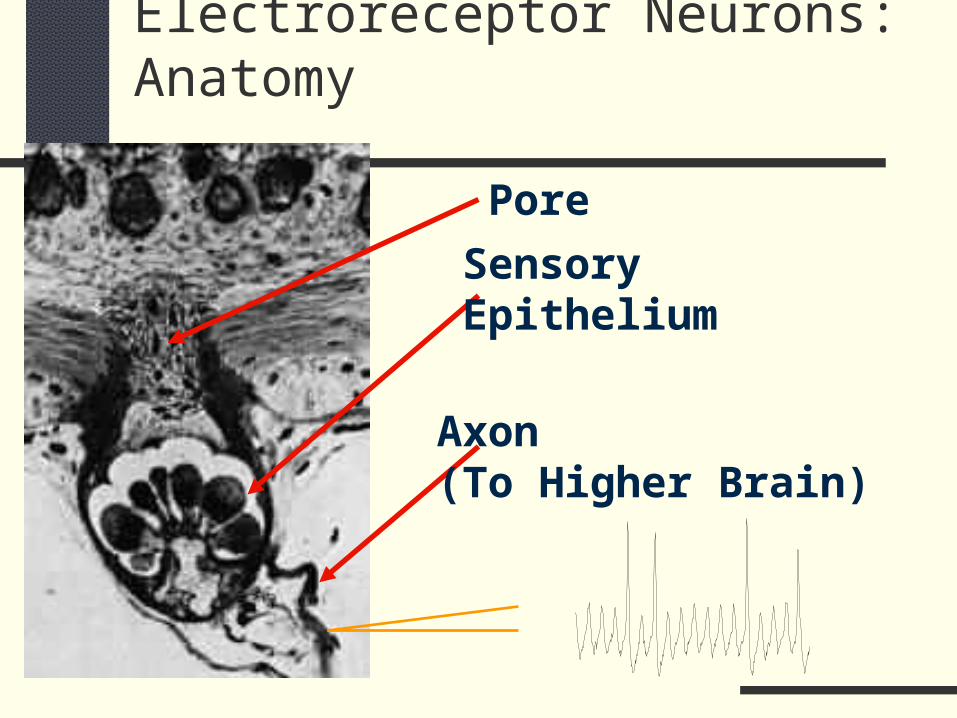
Electroreceptor Neurons: Anatomy
Pore
SensoryEpithelium
Axon(To Higher Brain)

Biology: Weakly electric fish
90 100 110 120-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
mV
time (EOD cycles)
transepidermal voltage amplitude
90 95 100 105 110 115 120
0.7
0.8
0.9
1.0
1.1
1.2
1.3
mV
time (EOD cycles)
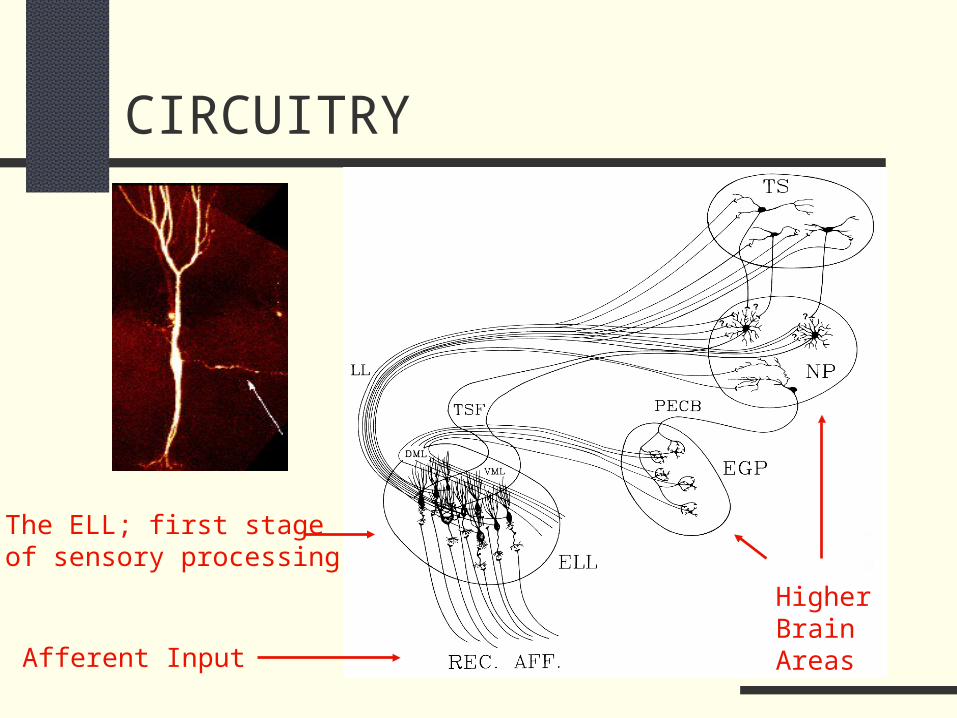
CIRCUITRY
Afferent Input
Higher Brain Areas
The ELL; first stage of sensory processing

Prey Stimuli
Electric fish prey on small insects (water fleas). These prey excite only a fraction of the electroreceptors that line the fish’s skin. We label this stimulation geometry local

Communication Stimuli
Electric fish communicate by modulating their own electric field, giving a specific input to other fish. These communication calls stimulate the entire surface of the receiving fish’s skin. We label this stimulation geometry global
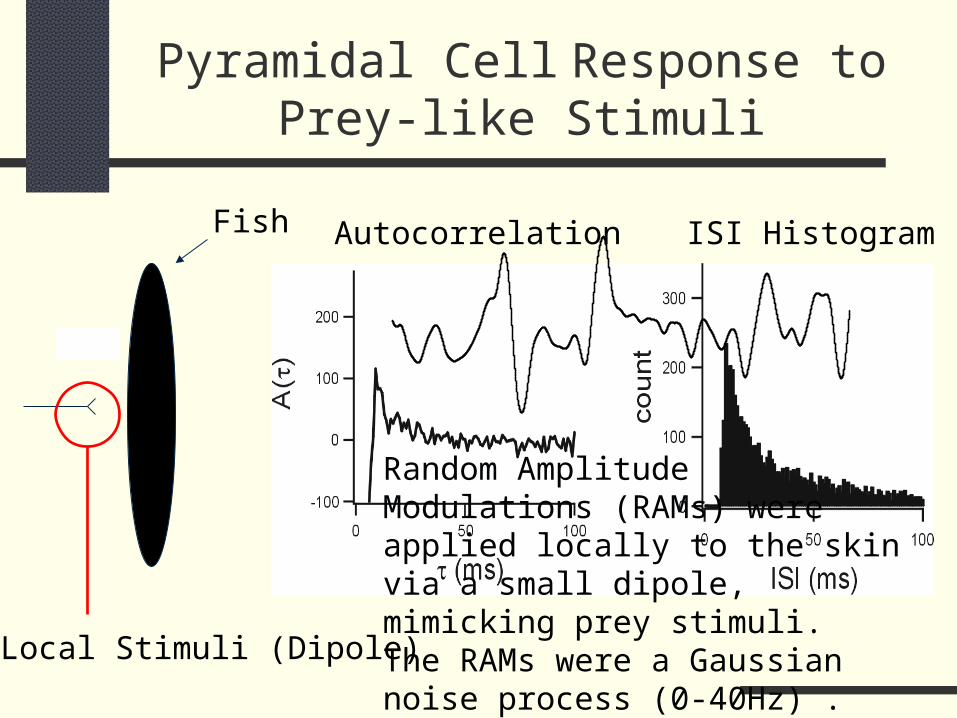
Pyramidal Cell Response to Prey-like Stimuli
Autocorrelation ISI Histogram
Local Stimuli (Dipole)
Fish
Random Amplitude Modulations (RAMs) were applied locally to the skin via a small dipole, mimicking prey stimuli. The RAMs were a Gaussian noise process (0-40Hz) .

Pyramidal Cell Response to Communication-like Stimuli
Global Stimuli
Autocorrelation ISI HistogramFish
RAMs were applied globally to the skin via a large dipole, mimicking communication stimuli. The RAMs were generated by a Gaussian noise process (0-40Hz) .

Model Pyramidal Cell
We model the ELL pyramidal cell network as an network of Leaky Integrate and Fire (LIF) neurons. The membrane potential of the ith neuron obeys the following dynamics
),()( diii tVGtIV
dt
dV
Ii(t) – inputG(V,t-d) – interactions V
i
time
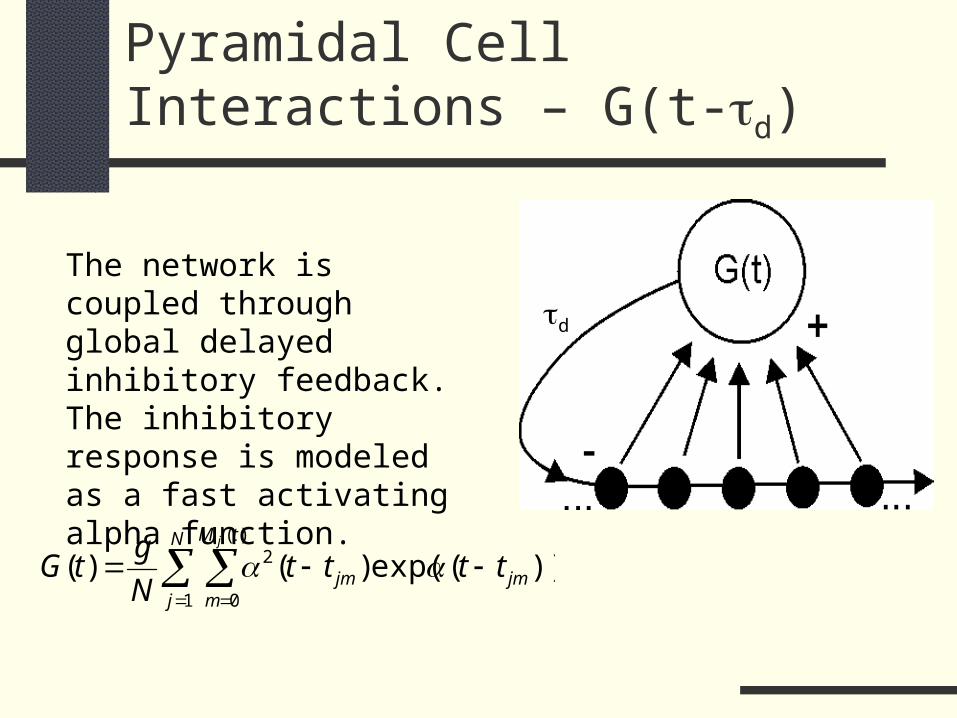
Pyramidal Cell Interactions – G(t-d)
))(exp()()(1
)(
0
2jm
N
j
tM
mjm tttt
N
gtG
j
The network is coupled through global delayed inhibitory feedback. The inhibitory response is modeled as a fast activating alpha function.
d

Pyramidal Cell Input - I(t)
Intrinsic Noise(biased Ornstein-Uhlenbeck process; =15 ms). Uncorrelated between neurons.
External Stimuli (Zero mean band passed Gaussian noise, 0-40Hz). Identical to experiments.
Pyramidal Cell
Ii(t) is composed of two types of “stimuli”

Network Model – Local Input
Autocorrelation Histogram
To mimic prey stimuli we apply the external stimulus to only one neuron

Network Model – Global Stimuli
To mimic communication stimuli we apply the external stimulus to all neurons equally.
Autocorrelation Histogram

Oscillation Mechanism
Local StimuliExternal Stimulus is applied heterogeneously across the network. No stimulus induced correlations.
Global StimuliExternal Stimulus is applied homogenously across the network. Significant stimulus induced correlations. Correlated activity cause a “wave” of inhibition after a delay. This wave carves out the oscillation.
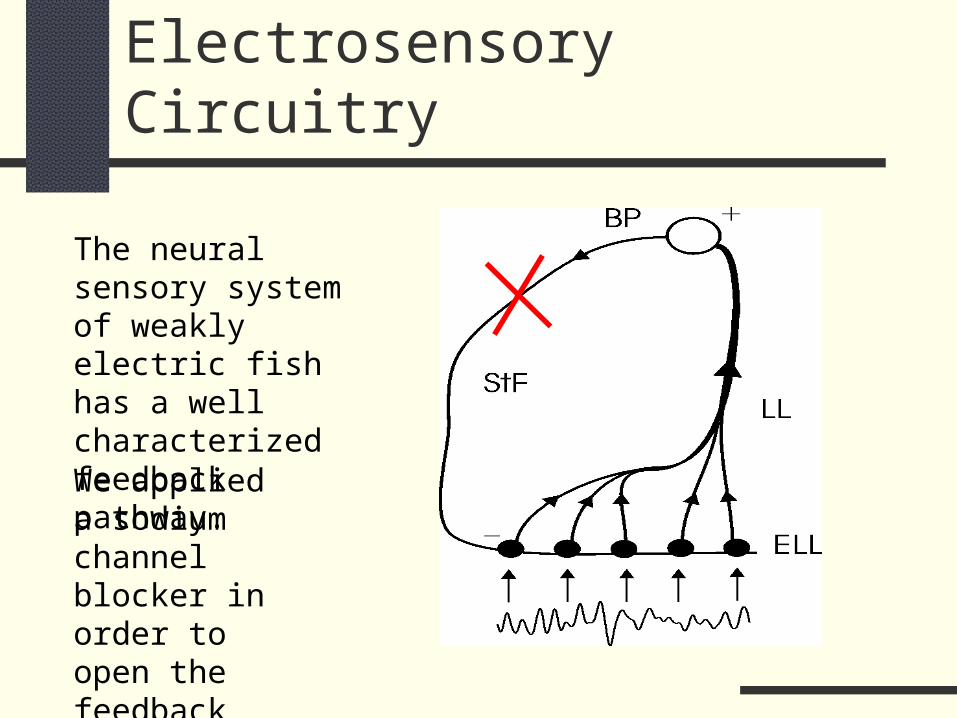
Electrosensory Circuitry
The neural sensory system of weakly electric fish has a well characterized feedback pathway.
We applied a sodium channel blocker in order to open the feedback loop.

Experimental Verification
ISI Histogram Autocorrelation
control
block
recover

Feedback: Open vs Closed Loop Architecture
Higher Brain
Higher Brain
Loop time d

Correlated Stimuli in Experiments
Dipole 1
Dipole 2
Dipole 3
Dipole 4
)()(1)( tctct Gii
The random signal emitted from each dipole was composed of an intrinsic, i(t), and global source, G(t). The relative strength of these two sources was parameterized by c, representing the covariance between dipoles.
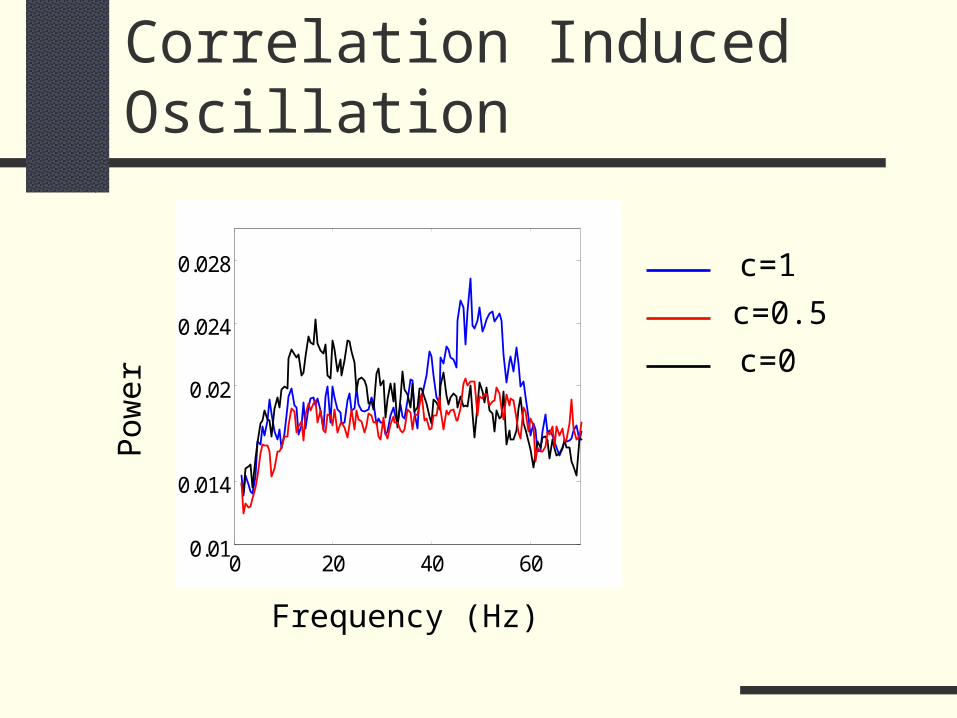
Correlation Induced Oscillation
0 20 40 600.01
0.014
0.02
0.024
0.028 c=1
c=0.5
c=0
Frequency (Hz)
Pow
er

Linear Response
Consider the spike train from the ith neuron in our network, . Assuming weak inputs we have that the Fourier transform of the spike train is
)()( iji ttt
)()()(~)(~ ibgi XA
where A(is the susceptibility of the neuron determined by the intrinsic properties of the cell. The Fourier transform of the input (external + feedback) is given by Xi().
(1)

Feedback Input
)(~)(~
)(~
)( jii dKN
gIX
Now consider the globally delay coupled LIF network used earlier. Let the “input” into neuron i be
Then it can be shown that for an infinite network we have
2
~2
22
)(~
1
)(~
)(~
Re2
)()()(
AKg
AKgAKg
AcSS
d
dd
bg

Fokker-Planck analysis on noisy Leaky Integrate-and-fire neurons
30
25
20
15
S (
spik
es2 /s
)
140120100806040200
frequency (Hz)
simulation c=1 theory c=1 simulations c=0 theory c=0

Shift in Coding Strategies
)()(
)()(
2
fPfP
fPfC
RRSS
SR
The spike train/stimulus coherence shifts from lowpass to highpass as we transition from local to global stimuli
Chacron, Doiron, Maler, Longtin, and Bastian,
Nature 423, 77-81 (May 1st 2003).

Spike Time Reliability
When high-frequency stimuli (40-60 Hz) are given, spike time reliability is increased dramatically when the stimulus is applied globally.

Conclusion
Electric fish use delayed inhibitory feedback to differentially respond to communication vs. prey stimuli.
Our work shows how a sensory system adapts its processing to its environment (local vs. global).