
CopyrightC Munksgaard 2001 2: 395–405 Munksgaard ...€¦ · the molecular basis for the role of...
11
Copyright C Munksgaard 2001 Traffic 2001 2: 395–405 Munksgaard International Publishers ISSN 1398-9219 Endocytosis of NBD-Sphingolipids in Neurons: Exclusion from Degradative Compartments and Transport to the Golgi Complex Teresa Babia ` 1 , Maria Dolores Ledesma 2,3 , Rainer Saffrich 2 , Jan Willem Kok 4 , Carlos G. Dotti 2,3 and Gustavo Egea 1, * 1 Dept. Biologia Cel.lular i Anatomia Patolo ` gica, Facultat de Medicina, Universitat de Barcelona and Institut d’Investigacions Biome ` diques August Pi i Sunyer (IDIBAPS), 08036 Barcelona, Spain 2 Cell Biology and Biophysics Program, European Molecular Biology Laboratory (EMBL), 69117 Heidelberg, Germany 3 Universita degli Studi di Torino, Cavalieri Ottolenghi Scientific Institute, A.O. San Luigi Gonzaga, Orbassano, Turin, Italy 4 Membrane Cell Biology Department, University of Groningen Medical School, 9713 AV Groningen, The Netherlands *Corresponding author: Gustavo Egea, [email protected] Sphingolipids are abundant constituents of neuronal membranes that have been implicated in intracellular signaling, neurite outgrowth and differentiation. Differ- ential localization and trafficking of lipids to membrane domains contribute to the specialized functions. In non-neuronal cultured cell lines, plasma membrane short-chain sphingomyelin and glucosylceramide are recycled via endosomes or sorted to degradative com- partments. However, depending on cell type and lipid membrane composition, short-chain glucosylceramide can also be diverted to the Golgi complex. Here, we show that NBD-labeled glucosylceramide and sphingo- myelin are transported from the plasma membrane to the Golgi complex in cultured rat hippocampal neurons irrespective of the stage of neuronal differentiation. Golgi complex localization was confirmed by colocaliz- ation and Golgi disruption studies, and importantly did not result from conversion of NBD-glucosylceramide or NBD-sphingomyelin to NBD-ceramide. Double-labeling experiments with transferrin or wheat-germ agglutinin showed that NBD-sphingolipids are first internalized to early/recycling endosomes, and subsequently trans- ported to the Golgi complex. The internalization of these two sphingolipid analogs was energy and tem- perature dependent, and their intracellular transport was insensitive to the NBD fluorescence quencher so- dium dithionite. These results indicate that vesicles mediate the transport of internalized NBD-glucosylcer- amide and NBD-sphingomyelin to the Golgi complex. Key words: endocytosis, Golgi complex, lipid traffick- ing, NBD-sphingolipids, neurons 395 Received 25 January 2001, revised and accepted for publication 23 March 2001 Lipid membrane segregation in epithelial cells has been studied by examining the trafficking of fluorescent, short acyl chain lipid analogs (C 6 -NBD-lipids) (1). Newly synthesized analogs of (glyco)sphingolipids (SLs), such as glucosylcera- mide (GlcCer) and sphingomyelin (SM), reach the epithelial cell surface with a different apical/basolateral polarity, re- flecting the surface polarity of endogenous GlcCer and SM (2). GlcCer is preferentially transported to the apical surface (3), and its transport to the plasma membrane involves ves- icular (4–7) as well as nonvesicular trafficking (8). Upon nonvesicular transport to the cytosolic leaflet of the plasma membrane, NBD-GlcCer can appear on the noncytosolic leaflet by transbilayer movement involving ATP-binding cas- sette proteins, such as P-glycoprotein (9,10). The ratio of SLs in the apical/basolateral plasma membrane domains de- pends on the nature of SLs and the epithelial tissues studied (9). Neurons are another example of polarized cells (11–14). In contrast to protein transport, little information is available on lipid trafficking in neuronal cells. In cultured rat hippocampal neurons, the ganglioside GM1 is internalized to late endocyt- ic compartments, where only the axonal turnover is depend- ent on the stage of neuronal development (15). Detailed knowledge of lipid trafficking and metabolism could provide the molecular basis for the role of SLs in the neurite out- growth and differentiation (16,17). With this aim we have in- vestigated the trafficking of SLs in cultured neuronal cells at different developmental stages by using short-chain analogs of glucosylceramide (NBD-aminohexanoyl-glucosylcerami- de, C 6 -NBD-GlcCer or NBD-GlcCer) and sphingomyelin (NBD-aminohexanoyl-sphingomyelin, C 6 -NBD-SM or NBD- SM). We show that in cultured rat hippocampal neurons NBD-GlcCer and NBD-SM are internalized to the Golgi com- plex via early/recycling endosomes. This result is particularly significant for NBD-SM since, to our knowledge, its transport to the Golgi complex has not been previously reported in cultured cells. Finally, both NBD-GlcCer and NBD-SM are transported to the Golgi complex by a vesicle-mediated mechanism. The functional relevance of the transport to the Golgi complex of internalized SLs for the biosynthetic and recycling pathways and neurites differentiation and out- growth is discussed.
Transcript of CopyrightC Munksgaard 2001 2: 395–405 Munksgaard ...€¦ · the molecular basis for the role of...

Copyright C Munksgaard 2001Traffic 2001 2: 395–405
Munksgaard International Publishers ISSN 1398-9219
Endocytosis of NBD-Sphingolipids in Neurons:Exclusion from Degradative Compartments andTransport to the Golgi Complex
Teresa Babia1, Maria Dolores Ledesma2,3,Rainer Saffrich2, Jan Willem Kok4,Carlos G. Dotti2,3 and Gustavo Egea1,*
1Dept. Biologia Cel.lular i Anatomia Patologica, Facultat de
Medicina, Universitat de Barcelona and Institut
d’Investigacions Biomediques August Pi i Sunyer
(IDIBAPS), 08036 Barcelona, Spain2Cell Biology and Biophysics Program, European
Molecular Biology Laboratory (EMBL), 69117 Heidelberg,
Germany3Universita degli Studi di Torino, Cavalieri Ottolenghi
Scientific Institute, A.O. San Luigi Gonzaga, Orbassano,
Turin, Italy4Membrane Cell Biology Department, University of
Groningen Medical School, 9713 AV Groningen, The
Netherlands
*Corresponding author: Gustavo Egea,
Sphingolipids are abundant constituents of neuronalmembranes that have been implicated in intracellularsignaling, neurite outgrowth and differentiation. Differ-ential localization and trafficking of lipids to membranedomains contribute to the specialized functions. Innon-neuronal cultured cell lines, plasma membraneshort-chain sphingomyelin and glucosylceramide arerecycled via endosomes or sorted to degradative com-partments. However, depending on cell type and lipidmembrane composition, short-chain glucosylceramidecan also be diverted to the Golgi complex. Here, weshow that NBD-labeled glucosylceramide and sphingo-myelin are transported from the plasma membrane tothe Golgi complex in cultured rat hippocampal neuronsirrespective of the stage of neuronal differentiation.Golgi complex localization was confirmed by colocaliz-ation and Golgi disruption studies, and importantly didnot result from conversion of NBD-glucosylceramide orNBD-sphingomyelin to NBD-ceramide. Double-labelingexperiments with transferrin or wheat-germ agglutininshowed that NBD-sphingolipids are first internalized toearly/recycling endosomes, and subsequently trans-ported to the Golgi complex. The internalization ofthese two sphingolipid analogs was energy and tem-perature dependent, and their intracellular transportwas insensitive to the NBD fluorescence quencher so-dium dithionite. These results indicate that vesiclesmediate the transport of internalized NBD-glucosylcer-amide and NBD-sphingomyelin to the Golgi complex.
Key words: endocytosis, Golgi complex, lipid traffick-ing, NBD-sphingolipids, neurons
395
Received 25 January 2001, revised and accepted forpublication 23 March 2001
Lipid membrane segregation in epithelial cells has beenstudied by examining the trafficking of fluorescent, short acylchain lipid analogs (C6-NBD-lipids) (1). Newly synthesizedanalogs of (glyco)sphingolipids (SLs), such as glucosylcera-mide (GlcCer) and sphingomyelin (SM), reach the epithelialcell surface with a different apical/basolateral polarity, re-flecting the surface polarity of endogenous GlcCer and SM(2). GlcCer is preferentially transported to the apical surface(3), and its transport to the plasma membrane involves ves-icular (4–7) as well as nonvesicular trafficking (8). Uponnonvesicular transport to the cytosolic leaflet of the plasmamembrane, NBD-GlcCer can appear on the noncytosolicleaflet by transbilayer movement involving ATP-binding cas-sette proteins, such as P-glycoprotein (9,10). The ratio of SLsin the apical/basolateral plasma membrane domains de-pends on the nature of SLs and the epithelial tissues studied(9).
Neurons are another example of polarized cells (11–14). Incontrast to protein transport, little information is available onlipid trafficking in neuronal cells. In cultured rat hippocampalneurons, the ganglioside GM1 is internalized to late endocyt-ic compartments, where only the axonal turnover is depend-ent on the stage of neuronal development (15). Detailedknowledge of lipid trafficking and metabolism could providethe molecular basis for the role of SLs in the neurite out-growth and differentiation (16,17). With this aim we have in-vestigated the trafficking of SLs in cultured neuronal cells atdifferent developmental stages by using short-chain analogsof glucosylceramide (NBD-aminohexanoyl-glucosylcerami-de, C6-NBD-GlcCer or NBD-GlcCer) and sphingomyelin(NBD-aminohexanoyl-sphingomyelin, C6-NBD-SM or NBD-SM). We show that in cultured rat hippocampal neuronsNBD-GlcCer and NBD-SM are internalized to the Golgi com-plex via early/recycling endosomes. This result is particularlysignificant for NBD-SM since, to our knowledge, its transportto the Golgi complex has not been previously reported incultured cells. Finally, both NBD-GlcCer and NBD-SM aretransported to the Golgi complex by a vesicle-mediatedmechanism. The functional relevance of the transport to theGolgi complex of internalized SLs for the biosynthetic andrecycling pathways and neurites differentiation and out-growth is discussed.
Babia et al.
Results
NBD-GlcCer and NBD-SM are internalized and
transported to the Golgi complex in rat hippocampal
neurons irrespective of their differentiation stage
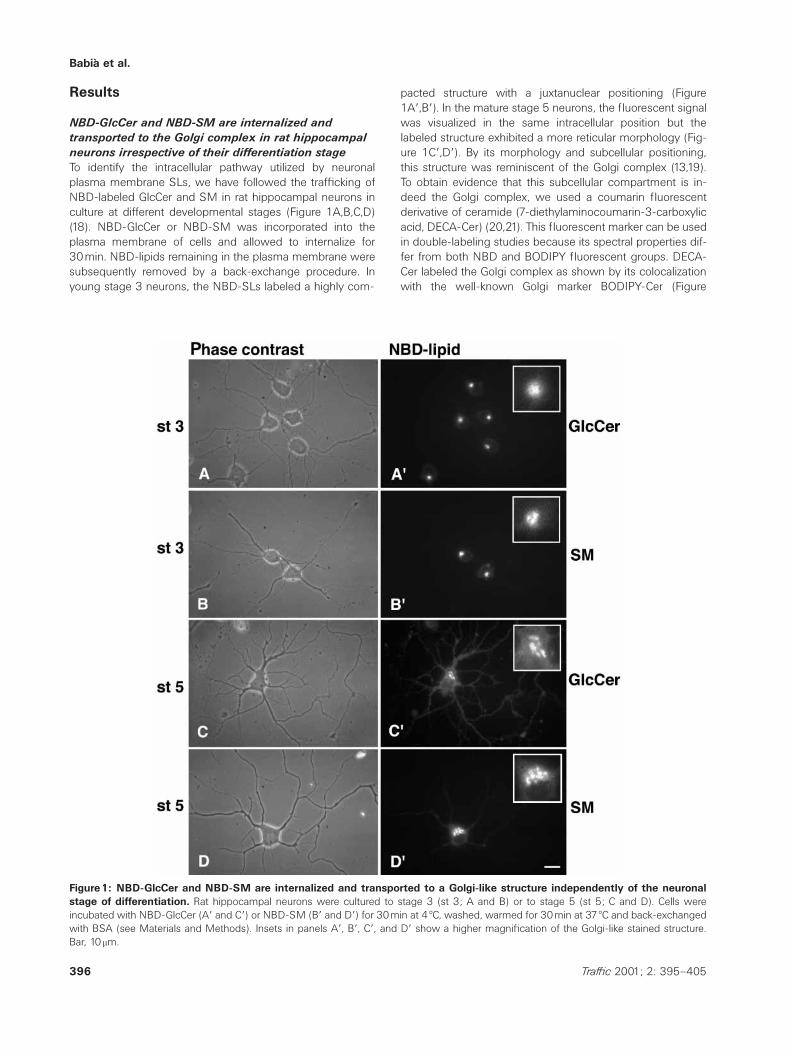
To identify the intracellular pathway utilized by neuronalplasma membrane SLs, we have followed the trafficking ofNBD-labeled GlcCer and SM in rat hippocampal neurons inculture at different developmental stages (Figure 1A,B,C,D)(18). NBD-GlcCer or NBD-SM was incorporated into theplasma membrane of cells and allowed to internalize for30min. NBD-lipids remaining in the plasma membrane weresubsequently removed by a back-exchange procedure. Inyoung stage 3 neurons, the NBD-SLs labeled a highly com-
Figure1: NBD-GlcCer and NBD-SM are internalized and transported to a Golgi-like structure independently of the neuronalstage of differentiation. Rat hippocampal neurons were cultured to stage 3 (st 3; A and B) or to stage 5 (st 5; C and D). Cells wereincubated with NBD-GlcCer (Aƒ and Cƒ) or NBD-SM (Bƒ and Dƒ) for 30min at 4 æC, washed, warmed for 30min at 37 æC and back-exchangedwith BSA (see Materials and Methods). Insets in panels Aƒ, Bƒ, Cƒ, and Dƒ show a higher magnification of the Golgi-like stained structure.Bar, 10mm.
396 Traffic 2001; 2: 395–405
pacted structure with a juxtanuclear positioning (Figure1Aƒ,Bƒ). In the mature stage 5 neurons, the fluorescent signalwas visualized in the same intracellular position but thelabeled structure exhibited a more reticular morphology (Fig-ure 1Cƒ,Dƒ). By its morphology and subcellular positioning,this structure was reminiscent of the Golgi complex (13,19).To obtain evidence that this subcellular compartment is in-deed the Golgi complex, we used a coumarin fluorescentderivative of ceramide (7-diethylaminocoumarin-3-carboxylicacid, DECA-Cer) (20,21). This fluorescent marker can be usedin double-labeling studies because its spectral properties dif-fer from both NBD and BODIPY fluorescent groups. DECA-Cer labeled the Golgi complex as shown by its colocalizationwith the well-known Golgi marker BODIPY-Cer (Figure
Lipid endocytosis to the Golgi in neurons
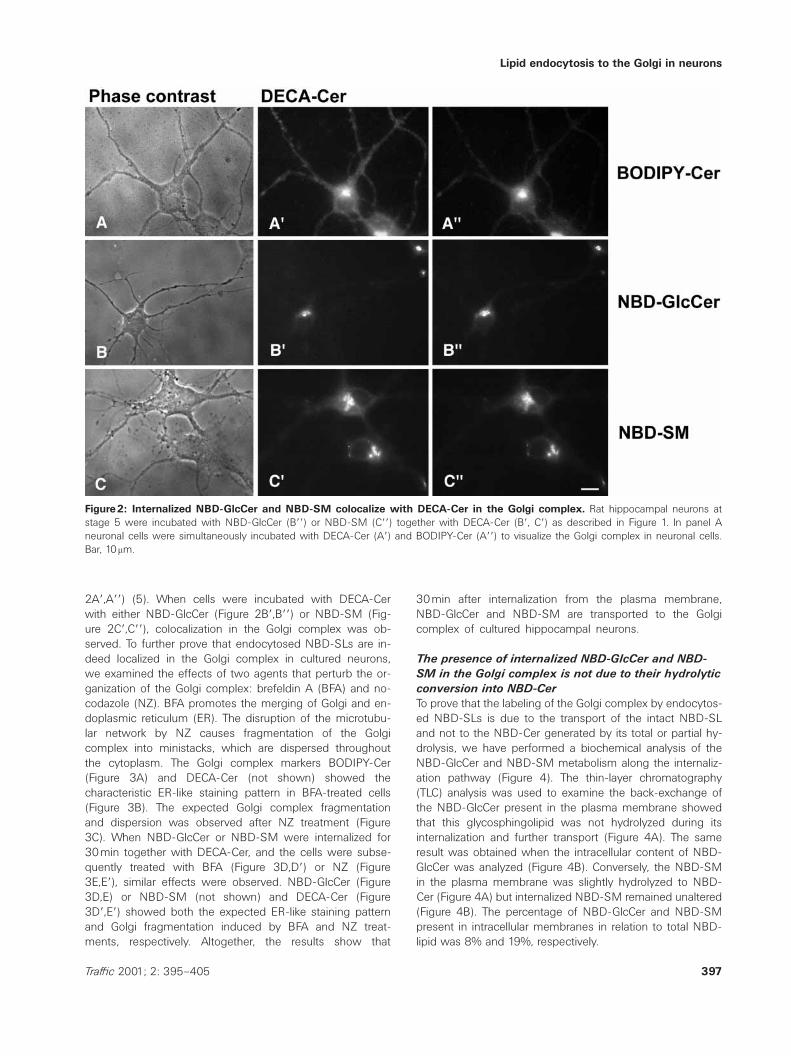
Figure2: Internalized NBD-GlcCer and NBD-SM colocalize with DECA-Cer in the Golgi complex. Rat hippocampal neurons atstage 5 were incubated with NBD-GlcCer (Bƒƒ) or NBD-SM (Cƒƒ) together with DECA-Cer (Bƒ, Cƒ) as described in Figure 1. In panel Aneuronal cells were simultaneously incubated with DECA-Cer (Aƒ) and BODIPY-Cer (Aƒƒ) to visualize the Golgi complex in neuronal cells.Bar, 10mm.
2Aƒ,Aƒƒ) (5). When cells were incubated with DECA-Cerwith either NBD-GlcCer (Figure 2Bƒ,Bƒƒ) or NBD-SM (Fig-ure 2Cƒ,Cƒƒ), colocalization in the Golgi complex was ob-served. To further prove that endocytosed NBD-SLs are in-deed localized in the Golgi complex in cultured neurons,we examined the effects of two agents that perturb the or-ganization of the Golgi complex: brefeldin A (BFA) and no-codazole (NZ). BFA promotes the merging of Golgi and en-doplasmic reticulum (ER). The disruption of the microtubu-lar network by NZ causes fragmentation of the Golgicomplex into ministacks, which are dispersed throughoutthe cytoplasm. The Golgi complex markers BODIPY-Cer(Figure 3A) and DECA-Cer (not shown) showed thecharacteristic ER-like staining pattern in BFA-treated cells(Figure 3B). The expected Golgi complex fragmentationand dispersion was observed after NZ treatment (Figure3C). When NBD-GlcCer or NBD-SM were internalized for30min together with DECA-Cer, and the cells were subse-quently treated with BFA (Figure 3D,Dƒ) or NZ (Figure3E,Eƒ), similar effects were observed. NBD-GlcCer (Figure3D,E) or NBD-SM (not shown) and DECA-Cer (Figure3Dƒ,Eƒ) showed both the expected ER-like staining patternand Golgi fragmentation induced by BFA and NZ treat-ments, respectively. Altogether, the results show that
397Traffic 2001; 2: 395–405
30min after internalization from the plasma membrane,NBD-GlcCer and NBD-SM are transported to the Golgicomplex of cultured hippocampal neurons.
The presence of internalized NBD-GlcCer and NBD-
SM in the Golgi complex is not due to their hydrolytic
conversion into NBD-Cer
To prove that the labeling of the Golgi complex by endocytos-ed NBD-SLs is due to the transport of the intact NBD-SLand not to the NBD-Cer generated by its total or partial hy-drolysis, we have performed a biochemical analysis of theNBD-GlcCer and NBD-SM metabolism along the internaliz-ation pathway (Figure 4). The thin-layer chromatography(TLC) analysis was used to examine the back-exchange ofthe NBD-GlcCer present in the plasma membrane showedthat this glycosphingolipid was not hydrolyzed during itsinternalization and further transport (Figure 4A). The sameresult was obtained when the intracellular content of NBD-GlcCer was analyzed (Figure 4B). Conversely, the NBD-SMin the plasma membrane was slightly hydrolyzed to NBD-Cer (Figure 4A) but internalized NBD-SM remained unaltered(Figure 4B). The percentage of NBD-GlcCer and NBD-SMpresent in intracellular membranes in relation to total NBD-lipid was 8% and 19%, respectively.

Babia et al.
Figure3: The Golgi complex localization of internalized NBD-GlcCer and NBD-SM is indicated by its sensitivity to brefeldin A(BFA) and nocodazole (NZ) treatments. Cells were incubated first with BODIPY-Cer for 20min at 4 æC, washed, warmed to 37 æC forlabeling of the neuronal Golgi complex, and then treated with BFA (B) or NZ (C) (see Materials and Methods). Next, neuronal cells (stage5) were first processed for the NBD-GlcCer (D and E) and DECA-Cer (Dƒ and Eƒ) as described in the legend of Figure 2. Thereafter, neuronswere treated with BFA (D and Dƒ) or NZ (E and Eƒ). As expected, treatments with BFA and NZ showed the respective ER-like stainingpattern and Golgi fragmentation and cytoplasmic dispersion for DECA-Cer and BODIPY-Cer (B and C) and, most important, for NBD-GlcCeras well (D and E). Bar, 10mm.
Figure4: NBD-GlcCer and NBD-SM are not hydrolyzed to NBD-Cer during their internalization and further transport to theGolgi complex. The NBD-lipid content of the plasma membrane and the intracellular pools were monitored by thin-layer chromatographyanalysis at different times of the internalization process as detailed in Materials and Methods. A representative result is shown here. Noticethat NBD-GlcCer is maintained unaltered in both the plasma membrane (A) and intracellular (B) pools. A small conversion of NBD-SM toNBD-Cer in the plasma membrane at 10 and 30min of incubation was observed (A). However, the NBD-GlcCer and the NBD-SM presentin the intracellular pool remain unaltered (B).
398 Traffic 2001; 2: 395–405

Lipid endocytosis to the Golgi in neurons
Internalized NBD-GlcCer and NBD-SM are diverted to
the Golgi complex between the early/recycling
endosomes and lysosomes
To determine whether the trafficking to the Golgi complex ofthe internalized SLs involved endocytic organelles, we per-formed double-fluorescence experiments with well-estab-lished endocytic markers. First, NBD-GlcCer or NBD-SM wasinternalized in the presence of TRITC-transferrin (TRITC-Tf) tomonitor the initial phases of the endocytic pathway by label-ing early/recycling endosomes (Figure 5). At early times(10min), NBD-GlcCer (Figure 5Aƒ) or NBD-SM (Figure 5Bƒ)appeared in characteristic punctated structures mostlylocated in the neuronal processes. These structures mostlycolocalized (80%) with TRITC-Tf (Figure 5Aƒƒ,Bƒƒ, small ar-rows). Thus, the results suggest that after their incorporationinto the plasma membrane NBD-SLs are transported toearly/recycling endosomes.
To determine whether after early/recycling endosomes NBD-glycolipids travel to late endosomes and lysosomes, we nextused the TRITC-WGA. This lectin accumulates in late endo-somes and lysosomes of hippocampal neurons (22). Neur-onal cells were first incubated with TRITC-WGA for 30min,
Figure5: NBD-GlcCer and NBD-SM partially colocalize with TRITC-transferrin (Tf) in the early/recycling endosomes. Culturedhippocampal neurons were first incubated with TRITC-Tf to trace the early phases of the neuronal endocytosis, and then with NBD-GlcCeror NBD-SM in the continuous presence of TRITC-Tf (see Materials and Methods for details). Note the partial colocalization (arrows) of NBD-GlcCer (Aƒ) or NBD-SM (Bƒ) in cytoplasmic punctate structures which are also labeled by TRITC-Tf (Aƒƒ and Bƒƒ).
399Traffic 2001; 2: 395–405
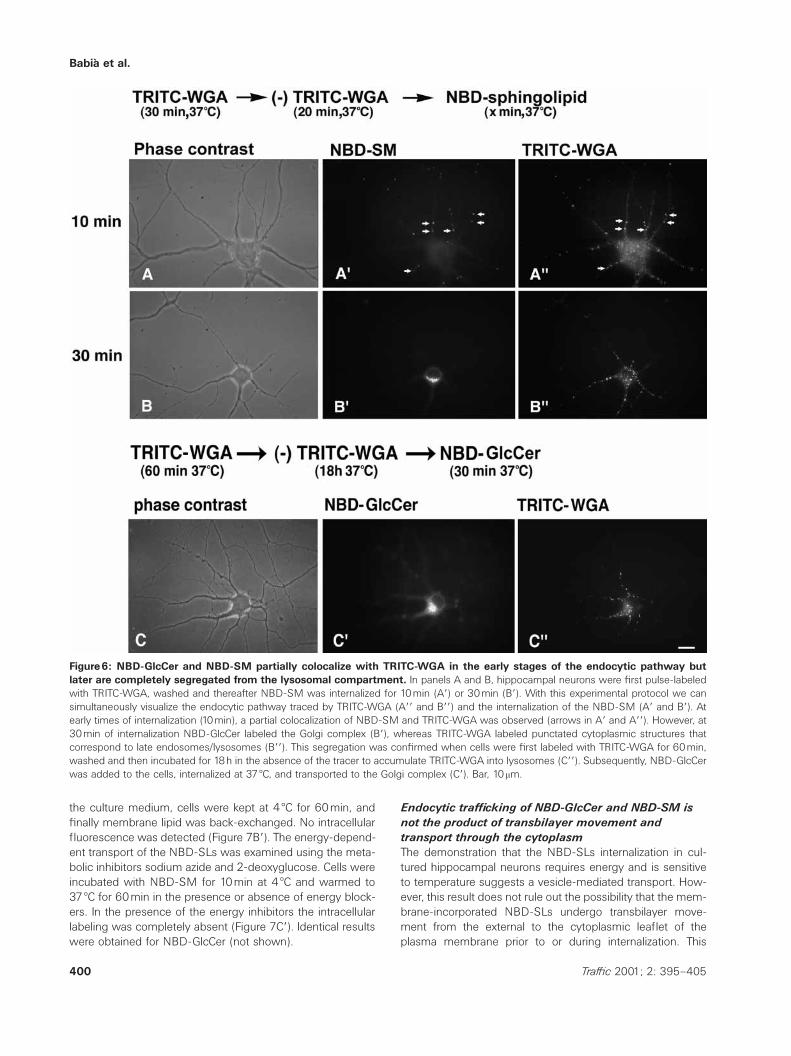
the lectin was then removed from the culture medium, andsubsequently NBD-SM (Figure 6) or NBD-GlcCer (notshown) was internalized for 10min (Figure 6, series A) or30min (Figure 6, series B). At 10min, NBD-SM partially co-localized with TRITC-WGA (Figure 6Aƒ,Aƒƒ, small arrows).However, at 30min, the fluorescent labeling was completelysegregated. NBD-SM was seen in the Golgi complex (Figure6Bƒ), whereas TRITC-WGA labeled the expected cytoplasmicpunctate structures that mainly corresponded to late endo-somes/lysosomes (Figure 6Bƒƒ). The same result was ob-tained when TRITC-WGA was accumulated in lysosomesafter a chase of 18h (Figure 6C,Cƒ,Cƒƒ).
The internalization of NBD-GlcCer and NBD-SM in
cultured neuronal cells is temperature and energy
dependent
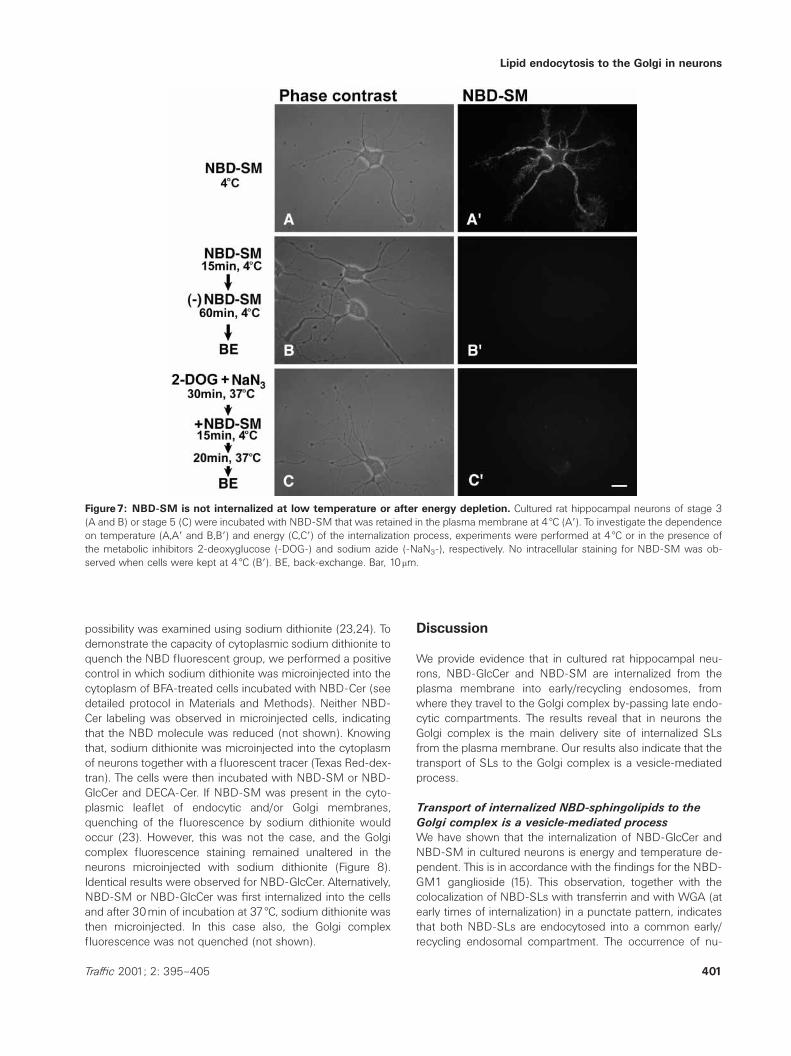
To determine whether the internalization of NBD-GlcCer andNBD-SM in cultured neuronal cells was an active or a passiveprocess, we next examined NBD-SL internalization underconditions of temperature and energy block. Addition ofNBD-SM to cells at low temperature (4 æC) resulted in theuniform distribution of the sphingolipid in the plasma mem-brane (Figure 7Aƒ). Thereafter, NBD-SM was removed from
Babia et al.
Figure6: NBD-GlcCer and NBD-SM partially colocalize with TRITC-WGA in the early stages of the endocytic pathway butlater are completely segregated from the lysosomal compartment. In panels A and B, hippocampal neurons were first pulse-labeledwith TRITC-WGA, washed and thereafter NBD-SM was internalized for 10min (Aƒ) or 30min (Bƒ). With this experimental protocol we cansimultaneously visualize the endocytic pathway traced by TRITC-WGA (Aƒƒ and Bƒƒ) and the internalization of the NBD-SM (Aƒ and Bƒ). Atearly times of internalization (10min), a partial colocalization of NBD-SM and TRITC-WGA was observed (arrows in Aƒ and Aƒƒ). However, at30min of internalization NBD-GlcCer labeled the Golgi complex (Bƒ), whereas TRITC-WGA labeled punctated cytoplasmic structures thatcorrespond to late endosomes/lysosomes (Bƒƒ). This segregation was confirmed when cells were first labeled with TRITC-WGA for 60min,washed and then incubated for 18h in the absence of the tracer to accumulate TRITC-WGA into lysosomes (Cƒƒ). Subsequently, NBD-GlcCerwas added to the cells, internalized at 37 æC, and transported to the Golgi complex (Cƒ). Bar, 10mm.
the culture medium, cells were kept at 4 æC for 60min, andfinally membrane lipid was back-exchanged. No intracellularfluorescence was detected (Figure 7Bƒ). The energy-depend-ent transport of the NBD-SLs was examined using the meta-bolic inhibitors sodium azide and 2-deoxyglucose. Cells wereincubated with NBD-SM for 10min at 4 æC and warmed to37 æC for 60min in the presence or absence of energy block-ers. In the presence of the energy inhibitors the intracellularlabeling was completely absent (Figure 7Cƒ). Identical resultswere obtained for NBD-GlcCer (not shown).
400 Traffic 2001; 2: 395–405
Endocytic trafficking of NBD-GlcCer and NBD-SM is
not the product of transbilayer movement and
transport through the cytoplasm
The demonstration that the NBD-SLs internalization in cul-tured hippocampal neurons requires energy and is sensitiveto temperature suggests a vesicle-mediated transport. How-ever, this result does not rule out the possibility that the mem-brane-incorporated NBD-SLs undergo transbilayer move-ment from the external to the cytoplasmic leaflet of theplasma membrane prior to or during internalization. This
Lipid endocytosis to the Golgi in neurons
Figure7: NBD-SM is not internalized at low temperature or after energy depletion. Cultured rat hippocampal neurons of stage 3(A and B) or stage 5 (C) were incubated with NBD-SM that was retained in the plasma membrane at 4 æC (Aƒ). To investigate the dependenceon temperature (A,Aƒ and B,Bƒ) and energy (C,Cƒ) of the internalization process, experiments were performed at 4 æC or in the presence ofthe metabolic inhibitors 2-deoxyglucose (-DOG-) and sodium azide (-NaN3-), respectively. No intracellular staining for NBD-SM was ob-served when cells were kept at 4 æC (Bƒ). BE, back-exchange. Bar, 10mm.
possibility was examined using sodium dithionite (23,24). Todemonstrate the capacity of cytoplasmic sodium dithionite toquench the NBD fluorescent group, we performed a positivecontrol in which sodium dithionite was microinjected into thecytoplasm of BFA-treated cells incubated with NBD-Cer (seedetailed protocol in Materials and Methods). Neither NBD-Cer labeling was observed in microinjected cells, indicatingthat the NBD molecule was reduced (not shown). Knowingthat, sodium dithionite was microinjected into the cytoplasmof neurons together with a fluorescent tracer (Texas Red-dex-tran). The cells were then incubated with NBD-SM or NBD-GlcCer and DECA-Cer. If NBD-SM was present in the cyto-plasmic leaflet of endocytic and/or Golgi membranes,quenching of the fluorescence by sodium dithionite wouldoccur (23). However, this was not the case, and the Golgicomplex fluorescence staining remained unaltered in theneurons microinjected with sodium dithionite (Figure 8).Identical results were observed for NBD-GlcCer. Alternatively,NBD-SM or NBD-GlcCer was first internalized into the cellsand after 30min of incubation at 37 æC, sodium dithionite wasthen microinjected. In this case also, the Golgi complexfluorescence was not quenched (not shown).
401Traffic 2001; 2: 395–405
Discussion
We provide evidence that in cultured rat hippocampal neu-rons, NBD-GlcCer and NBD-SM are internalized from theplasma membrane into early/recycling endosomes, fromwhere they travel to the Golgi complex by-passing late endo-cytic compartments. The results reveal that in neurons theGolgi complex is the main delivery site of internalized SLsfrom the plasma membrane. Our results also indicate that thetransport of SLs to the Golgi complex is a vesicle-mediatedprocess.
Transport of internalized NBD-sphingolipids to the
Golgi complex is a vesicle-mediated process
We have shown that the internalization of NBD-GlcCer andNBD-SM in cultured neurons is energy and temperature de-pendent. This is in accordance with the findings for the NBD-GM1 ganglioside (15). This observation, together with thecolocalization of NBD-SLs with transferrin and with WGA (atearly times of internalization) in a punctate pattern, indicatesthat both NBD-SLs are endocytosed into a common early/recycling endosomal compartment. The occurrence of nu-

Babia et al.
Figure8: The Golgi complex fluorescent labeling by the internalized NBD-SM is not abolished by sodium dithionite microin-jected into the cytoplasm. Cultured rat hippocampal neurons were first microinjected with the NBD-fluorescence quencher sodiumdithionite, and then NBD-SM and DECA-Cer were internalized and subsequently accumulated into the Golgi complex. Notice that in microin-jected cells (A, arrows), the Golgi fluorescence by NBD-SM remained virtually unaltered compared to nonmicroinjected cells where theGolgi labeling by DECA-Cer (B) and NBD-SM (C) was also visualized.
merous fluorescent punctate cytoplasmic structures and theenergy and temperature dependence of the internalization ofNBD-GlcCer and NBD-SM argue that these SLs are trans-ported to the Golgi complex by a vesicle-mediated mechan-ism. The results of the sodium dithionite microinjection ex-periments further support this idea. The fluorescent stainingof the plasma membrane, the Golgi complex and the punc-tate cytoplasmic structures remains unaltered despite thepresence of the NBD-fluorescence quencher in the cyto-plasm. This also indicates that there is no transbilayer move-ment of NBD-GlcCer and NBD-SM from the external to thecytoplasmic leaflet followed by passive monomer transportthrough the cytoplasm to the Golgi complex. Such a mechan-ism has been previously shown for NBD-GlcCer in humanskin fibroblasts (25). Moreover, the relatively long period re-quired for the transport of NBD-SLs to the Golgi is compat-ible with a vesicle-mediated mechanism. Additionally, thebiochemical analysis revealed that NBD-Cer is not generatedduring the internalization process. This result is interestingsince NBD-Cer is transported to the Golgi complex throughthe cytoplasm via a nonvesicular process (5).
The functional significance of internalized
sphingolipids in the Golgi complex of neuronal cells
The Golgi complex localization of both NBD-SLs after theirinternalization from the plasma membrane is independent ofthe state of neuronal differentiation. This finding is interestingbecause in HT29 human colon carcinoma cells, endocytosedNBD-GlcCer accumulates in the Golgi complex only in undif-ferentiated cells (26,27). Although we do not know thephysiological significance of internalizing lipids to the Golgicomplex in neuronal cells, it is tempting to speculate thatthis could be a good source to supply the high demands of
402 Traffic 2001; 2: 395–405
membrane transport in these highly dynamic cells. Thus, inearly stages of neuronal development, the lipids internalizedfrom the growth cones and transported to the Golgi complexmay be used for the synthesis of new membrane carriers,which in turn are transported to the cell surface to supportaxonal elongation (28). In mature neurons, endocytosis ofmembrane lipids to the Golgi complex could also serve as arapid source of new membrane needed to replenish plasmamembrane loss due to synaptic-mediated membrane inter-nalization. This view is supported by: (a) the requirement ofboth SLs in the development and differentiation of neuronalprocesses (16,29), and (b) the developmental upregulationof SLs in the maturation of the neuronal membrane-sortingpathway (30).
In most of the clonal cell lines, endocytosed SLs follow therecycling (via early/recycling endosomes) or the degradative(late endosomes/lysosomes) pathways (31–34). The endo-cytic sorting to the Golgi complex of glycosphingolipids suchas globosides (through shiga toxin trafficking) (35), ganglios-ides (36,37), NBD-glycosphingolipids analogs such asGlcCer, and lactosylceramide (25,26,38) and the glycosyl-ation of nondegradable GlcCer analogs (39) have also beendescribed. However, to our knowledge this is the first timethat internalized NBD-GlcCer and NBD-SM are both divertedto the Golgi complex. Although the differential sorting of in-ternalized SLs may depend on the specific cellular demands,cell-specific molecular mechanisms appear to govern thefinal destination of the lipids. One of these mechanismswould be the concentration of cholesterol in the plasmamembrane. It has recently been reported that the Golgi com-plex localization of lactosylceramide in some sphingolipid-storage diseases depends on the levels of cholesterol in the

Lipid endocytosis to the Golgi in neurons
plasma membrane (40,41). Hence, neuronal cells could sortendocytosed SLs to the Golgi complex because they containlower levels of plasma membrane cholesterol (42). Anothermechanism for the Golgi trafficking seen in neurons may berelated to the interaction of SLs with specific proteins, forexample those with lectin-binding properties such as shigaand cholera toxins that bind to globotriaosylceramide (43)and GM1 (44), respectively. These toxins are first transportedto the Golgi complex and subsequently to the ER (45–47).Strikingly, in cultured hippocampal neurons NBD-GM1 is in-ternalized to the lysosomal compartment (15), except whencholera toxin is bound to this ganglioside, in which case thetoxin and the ganglioside are transported to the Golgi com-plex (48). Presumably, GM1 is delivered to the Golgi complexpassively accompanying the toxin that is actively transportedto the Golgi complex and subsequently to the ER (49). In asimilar way, NBD-GlcCer and NBD-SM could be diverted tothe Golgi complex from early/recycling endosomes by inter-action with some endogenous proteins that follow a toxin-like pathway. There is evidence that proteins can pass directlyfrom early endosomes to the trans-Golgi network (TGN)without passing through late endosomes (50). However, themolecular mechanism of this transport step is less character-ized than that of transport from late endosomes to the TGN,which is regulated by the small GTPase Rab 9 (51,52).
Materials and Methods
Materials
1b-D-glucosylsphingosine, sphingosylphosphorylcholine, D-sphingosine, iron-free transferrin (Tf), brefeldin A and noco-dazole were purchased from Sigma Chemical Co. (St.Louis, MO, USA). 6-[N-(7-nitro-2,1,3-benzoxadiazol-4-yl)a-mino]hexanoic acid, BODIPY-Ceramide and wheat-germagglutinin were from Molecular Probes (Eugene, OR, USA).Sodium dithionite was from Merck (Darmstadt, Germany).Iron saturation and TRITC conjugation of transferrin wascarried out as described previously (34).
Hippocampal neurons culture
Primary cultures of hippocampal neurons were prepared fromrat brains of 18-day-old embryos as described (53). Neuronswere kept in culture for 48–72h (Stage 3) or 5–10days(Stage 5).
Synthesis of fluorescence-labeled sphingolipids
C6-NBD-glucosylceramide (NBD-GlcCer), C6-NBD-sphingo-myelin (NBD-SM) and DECA-Ceramide (DECA-Cer) weresynthesized as described (20,21).
Plasma membrane insertion of fluorescent lipids
NBD-SLs, BODIPY-Cer and DECA-Cer were stored in chloro-form/methanol(2 :1 [v/v]), dried under a nitrogen atmos-phere, and solubilized in absolute ethanol (0.5%, final con-centration). Thereafter, this ethanolic solution was dissolvedinto N2 culture medium containing taxol (0.5mM, final con-
403Traffic 2001; 2: 395–405
centration), vigorously mixed, and added to cultured neuronsat 4 æC.
Back-exchange of membrane-inserted fluorescent
lipid
Back-exchange was carried out by incubating the cells at 4 æCfor 15min with 3% BSA (wt/vol) in Hank’s solution, followedby extensive rinses with this solution. Thereafter, cells wereincubated at 20 æC for 2min with Hank’s solution containingsodium dithionite (80mM, final concentration).
Lipid extraction and TLC analysis
After the NBD-SLs internalization experiments, neurons wereback-exchanged at 4 æC with 3% BSA in Hank’s solution toseparate NBD-SLs present in the plasma membrane fromthose localized in the intracellular membranes. Extracted NBD-SLs were subsequently analyzed by thin-layer chromatogra-phy on silica gel 60 HPTLC plates using chloroform/methanol/20% (wt/vol) NH4OH [70 :30 :5] as the running solvent sys-tem. For quantitative analysis, individual spots were scrapedfrom the HPTLC plates and eluted from the silica with 1% (v/v)Triton-X100 solution by vigorous shaking and incubated at37 æC for 60min. Silica particles were centrifuged, and the flu-orescence in the supernatant was measured in a fluorimeter(Kontron Instruments Fluorimeter, mod. SFM 25, Kontron In-struments, Ltd, Southampton, UK) at excitation and emissionwavelengths of 465nm and 530nm, respectively.
Brefeldin A and nocodazole treatments
Cultured neurons were first incubated with BFA (10mg/mL)and NZ (20mM) for 60min at 37 æC, and then labeled with theNBD-SL as described above.
Cytochemical double-labeling experiments
Double-labeling experiments with DECA-Cer and NBD-GlcCer or NBD-SM were performed by incubating the neu-rons with NBD-SL (4mM) and DECA-Cer (1mM) at 4 æC, rinsedin Hank’s solution, warmed to 37 æC, and back-exchanged at4 æC. Colabeling experiments with TRITC-Tf (Figure 5) wereperformed as follows: cells were first labeled with TRITC-Tf(0.8mM) for 30min at 37 æC. Thereafter, cells were labeledwith the corresponding NBD-SL at 4 æC, and subsequentlywarmed to 37 æC for internalization at different times with thepresence in the culture medium of TRITC-Tf. Finally, a back-exchange procedure was performed.
TRITC-WGA experiments (Figure 6) were performed incubat-ing the neurons with TRITC-WGA (15mg/mL) for 30 or 60minat 37 æC, washed with Hank’s solution, and then incubated at37 æC (without WGA) for 20min or 18h. Thereafter, cells wereincubated with NBD-SL at 4 æC for its insertion in the plasmamembrane and subsequently warmed to 37 æC for internaliz-ation. Finally, a back-exchange procedure was performed.
Energy dependence
Cultured neurons were preincubated for 30min at 37 æC withsodium azide (5mM)/2-deoxyglucose (50mM), followed bythe incubation with NBD-SL at 4 æC, rinsing with PBS,

Babia et al.
warmed to 37 æC for 20min in the presence of the drugs, andback-exchanged at 4 æC.
Sodium dithionite microinjection experiments
Neurons were microinjected first with sodium dithionite (80–800mM), and subsequently labeled with NBD-SM or NBD-GlcCer at 4 æC. Thereafter, cells were incubated at 37 æC fordifferent times and back-exchanged at 4 æC. Alternatively,NBD-SM or NBD-GlcCer was internalized first into the cellsfollowed by sodium dithionite microinjection. A positive con-trol experiment with sodium dithionite present in the cyto-plasm was performed as follows: cells were treated with BFA(5mg/mL) for 15min at 37 æC, subsequently incubated withNBD-Cer (4mM, final concentration) for 30min at 4 æC andthen for 10min at 37 æC. Finally, a back-exchange procedurewith BSA (3%) for 30min at 4 æC was performed. Some cellswere then microinjected with sodium dithionite (700mM) to-gether with Texas-Red-dextran for their identification.
Finally, all images were captured at nonsaturating integrationlevels with a charge-coupled device camera (Cohu 4193,Cohu, San Diego, CA, USA) connected to a Macintosh com-puter (Power Macintosh 7300/166).
Acknowledgments
This article is dedicated to the memory of Eladio Vinuela and FernandoJimenez. We thank Bianca Hellias and Etienne Cassin for the culture andmaintenance of rat hippocampal neurons, Robin Rycroft (SAL, UB) forlinguistic assistance, and Maite Munoz for technical support with themicroinjection equipment. This work was supported by grants to C.G.D.(Deutsche Forschungsgemeinshaft, SFB 317), and to G.E. (CICYTSAF20000–0042 and Accion Integrada .HA1997–0020).
References
1. Pagano RE, Sleight RG. Defining lipid transport pathways in animalcells. Science 1985;229:1051–1057.
2. van’t Hof W, van Meer G. Generation of lipid polarity in intestinal epi-thelial (Caco-2) cells: sphingolipid synthesis in the Golgi complex andsorting before vesicular traffic to the plasma membrane. J Cell Biol1990;111:977–986.
3. van Meer G, Stelzer EH, Wijnaendts-van-Resandt RW, Simons K. Sort-ing of sphingolipids in epithelial Madin-Darby canine kidney cells. JCell Biol 1987;105:1623–1635.
4. Lipsky NG, Pagano RE. Sphingolipid metabolism in cultured fibro-blasts: microscopic and biochemical studies employing a fluorescentceramide analogue. Proc Natl Acad Sci USA 1983;80:2608–2612.
5. Lipsky LW, Pagano RE. A vital stain for the Golgi apparatus. Science1985;228:745–747.
6. Kobayashi T, Pagano RE. Lipid transport during mitosis. Alternativepathways for delivery of newly synthesized lipids to the cell surface.J Biol Chem 1989;264:5966–5973.
7. Babia T, Kok JW, van der Haar M, Kalicharan R, Hoekstra D. Transportof biosynthetic sphingolipids from Golgi to plasma membrane in HT29cells: involvement of different carrier vesicle populations. Eur J CellBiol 1994;172:172–181.
404 Traffic 2001; 2: 395–405
8. Warnock DE, Lutz MS, Blackburn WA, Young WW Jr, Baenziger JU.Transport of newly synthesized glucosylceramide to the plasma mem-brane by a non-Golgi pathway. Proc Natl Acad Sci USA1994;91:2708–2712.
9. van der Bijl P, L. Lopes-Cardoso M, van Meer G. Sorting of newlysynthesized galactosphingolipids to the two surface domains of epi-thelial cells. J Cell Biol 1996;132:813–821.
10. Raggers RJ, van Helvoort A, Evers R, van Meer G. The human multid-rug resistance protein MRP1 translocates sphingolipid analogs acrossthe plasma membrane. J Cell Sci 1999;112:415–422.
11. Dotti CG, Simons K. Polarized sorting of viral glycoproteins to the axonand dendrites of hippocampal neurons in culture. Cell 1990;62:63–70.
12. Dotti CG, Kartenbeck J, Parton R, Simons K. Polarized distribution ofviral glycoproteins of vesicular stomatitis, fowl plague and Semliki For-est viruses in hippocampal neurons in culture: a light and electronmicroscopy study. Brain Res 1993;610:141–147.
13. Dotti CG, Banker G. Intracellular organization of hippocampal neuronsduring the development of neuronal polarity. J Cell Sci Suppl1991;15:75–84.
14. Pietrini G, Suh Y, Edelman L, Rudnik G, Caplan M. The axonal g-amino-butyric acid transporter GAT-1 is sorted to the apical membranes ofpolarized epithelial cells. J Biol Chem 1994;269:4668–4674.
15. Sofer A, Schwarzmann G, Futerman AH. The internalization of a shortacyl chain analogue of ganglioside GM1 in polarized neurons. J CellSci 1996;109:2111–2119.
16. Futerman AH, Banker GA. The economics of neurite outgrowth – theaddition of new membrane to growing axons. Trends Neurosci1996;19:144–149.
17. Schwartz A, Futerman AH. Distinct roles for ceramide and glucosyl-ceramide at different stages of neuronal growth. J Neurosci1997;17:2929–2938.
18. Craig AM, Banker G. Neuronal polarity. Ann Rev Neurosci1994;17:267–310.
19. Krijnse-Locker J, Parton RG, Fuller SD, Griffiths G, Dotti CG. The organ-ization of the endoplasmic reticulum and the intermediate compart-ment in cultured rat hippocampal neurons. Mol Biol Cell 1995;6:1315–1332.
20. van’t Hof W, Silvius J, Wieland F, van Meer G. Epithelial sphingolipidsorting allows for extensive variation of the fatty chain and the sphin-gosine backbone. Biochem J 1992;283:913–917.
21. Kok JW, Hoekstra D. Fluorescent lipid analogues: applications in celland membrane biology. In: Mason WT, editors. Fluorescent and Lumi-nescent Probes for Biological Activity. London: Academic Press; 1999.p. 136–155.
22. Matteoli M, Takei K, Perin MS, Südhof TC, De Camilli P. Exo-endocyticrecycling of synaptic vesicles in developing processes of culturedhippocampal neurons. J Cell Biol 1992;117:849–861.
23. McIntyre JC, Sleight RG. Fluorescent assay for phospholipid mem-brane asymmetry. Biochemistry 1991;30:11819–11827.
24. Kok JW, Babia T, Klappe K, Hoekstra D. Fluorescent, short-chain C6-NBD-sphingomyelin, but not C6-NBD-glucosylceramide, is subject toextensive degradation in the plasma membrane: implications for sig-nal transduction related to cell differentiation. Biochem J1995;309:905–912.
25. Martin OC, Pagano RE. Internalization and sorting of a fluorescentanalogue of glucosylceramide to the Golgi apparatus of human skinfibroblasts: utilization of endocytic and non-endocytic mechanisms. JCell Biol 1994;125:769–781.
26. Kok JW, Babia T, Hoekstra D. Sorting of sphingolipids in the endocyticpathway of HT29 cells. J Cell Biol 1991;114:231–239.
27. Babia T, Kok JW, Hulstaert C, de Weerd H, Hoekstra D. Differentialmetabolism and trafficking of sphingolipids in differentiated versusundifferentiated HT29 cells. Int J Cancer 1993;5:839–845.

Lipid endocytosis to the Golgi in neurons
28. Bradke T, Dotti CG. Membrane traffic in polarized neurons. BiochimBiophys Acta 1998;1404:245–258.
29. Yamashita T, Wada R, Sasaki T, Deng C, Bierfreund U, Sandhoff K,Proia RL. A vital role for glycosphingolipid synthesis during develop-ment and differentiation. Proc Natl Acad Sci USA 1999;96:9142–9147.
30. Ledesma MD, Brugger B, Bunning C, Wieland FT, Dotti CG. Maturationof the axonal plasma membrane requires upregulation of sphingo-myelin synthesis and formation of protein–lipid complexes. EMBO J1999;18:1761–1771.
31. Koval M, Pagano R. Lipid recycling between the plasma membraneand intracellular compartments: transport and metabolism ofsphingomyelin analogs in cultured fibroblasts. J. Cell Biol1989;108:2169–2181.
32. Koval M, Pagano RE. Sorting of an internalized plasma membranelipid between recycling and degradative pathways in normal and Nie-mann-Pick, type A fibroblast. J Cell Biol 1990;111:429–442.
33. Hoekstra D, Kok JW. Trafficking of glycosphingolipids in eukaryoticcells – sorting and recycling lipids. Biochem Biophys Acta1992;1113:277–294.
34. Kok JW, Hoekstra K, Eskelinen S, Hoekstra D. Recycling pathways ofglucosylceramide in BHK cells: distinct involvement of early and lateendosomes. J Cell Sci 1992;103:1139–1152.
35. Johannes L, Goud B. Surfing on a retrograde wave: how does Shigatoxin reach the endoplasmic reticulum? Trends Cell Biol 1998;8:158–162.
36. Schwarzmann G, Sandhoff K. Metabolism and intracellular transportof glycosphingolipids. Biochemistry 1990;29:10865–10871.
37. Sillence DJ, Allan D. Repair of BHK cell surface ganglioside GM3 afterits degradation by extracellular sialidase. Mol Membr Biol1998;15:229–235.
38. Chen CS, Patterson MC, Wheatley CL, O’Brien JF, Pagano RE. Broadscreening test for sphingolipid-storage diseases. Lancet1999;354:901–905.
39. Schwarzmann G, Hofmann P, Putz U, Albrecht B. Demonstration ofdirect glycosylation of nondegradable glucosylceramide analogs incultured cells. J Biol Chem 1995;270:21271–21276.
40. Puri V, Watanabe R, Domınguez M, Sun X, Wheatley CL, Marks DL,Pagano RE. Cholesterol modulates membrane traffic along the endo-cytic pathway in sphingolipid-storage diseases. Nat Cell Biol1999;1:386–388.
405Traffic 2001; 2: 395–405
41. Pagano RE, Puri V, Dominguez M, Marks DL. Membrane traffic insphingolipid storage diseases. Traffic 2000;1:807–815.
42. Saito M, Benson EP, Saito M, Rosenberg A. Metabolism of cholesteroland triacylglycerol in cultured chick neuronal cells, glial cells, andfibroblasts: accumulation of esterified cholesterol in serum-free cul-ture. J Neurosci Res 1987;18:319–325.
43. Lingwood CA. Verotoxins and their glycolipid receptors. Adv Lipid Res1993;25:189–211.
44. Fishman PH, Pacuczka T, Orlandi PA. Gangliosides as receptors forbacterial enterotoxins. Adv Lipid Res 1993;25:165–167.
45. Sandvig K, Garred O, Prydz K, Koslov JV, Hansen SH, van Deurs B.Retrograde transport of endocytosed Shiga toxin to the endoplasmicreticulum. Nature 1992;358:510–512.
46. Nambiar M, Oda T, Chen C, Kuwazuru Y, Wu HC. Involvement of theGolgi region in the intracellular trafficking of cholera toxin. J Cell Physi-ol 1993;154:222–228.
47. Mallard F, Antony C, Tenza D, Salamero J, Goud B, Johannes L. Directpathway from early/recycling endosomes to the Golgi apparatus re-vealed through the study of shiga toxin B-fragment transport. J CellBiol 1998;143:973–990.
48. Sofer A, Futerman AH. Rate of retrograde transport of cholera toxinfrom the plasma membrane to the Golgi apparatus and endoplasmicreticulum decreases during neuronal development. J Neurochem1996;67:2134–2140.
49. Majoul IV, Bastiaens PI, Söling H-D. Transport of an external Lys-Asp-Glu-Leu (KDEL) protein from the plasma membrane to the endoplas-mic reticulum: studies with cholera toxin in Vero cells. J Cell Biol1996;133:777–789.
50. Mellman I. Endocytosis and molecular sorting. Ann Rev Cell Dev Biol1996;12:575–625.
51. Lombardi D, Soldati T, Riederer MA, Goda Y, Zerial M, Pfeffer SR. Rab9 functions in transport between late endosomes and the trans Golginetwork. EMBO J 1993;12:677–682.
52. Riederer MA, Soldati AD, Shapiro AD, Lin J, Pfeffer SR. Lysosomesbiogenesis requires Rab9 function and receptor recycling from endo-somes to the trans-Golgi network. J Cell Biol 1994;125:573–582.
53. Goslin K, Banker G. Rat hippocampal neurons in low-density culture.In: Banker G, Goslin K, editors. Culturing Nerve Cells. Cambridge(USA): MIT; 1991. p. 251–281.









![ReactiveBalanceControlinResponseto ... · responses intheSLR[16,17]. In viewofreflexive musclecompensationin response tosurface translation,distinctionis drawnbetweenperturbation](https://static.fdocuments.us/doc/165x107/5d4e3c6888c993a43e8bb8c1/reactivebalancecontrolinresponseto-responses-intheslr1617-in-viewofreflexive.jpg)








