
Colonization of cashew plants by Lasiodiplodia...
10
Micron 42 (2011) 419–428 Contents lists available at ScienceDirect Micron journal homepage: www.elsevier.com/locate/micron Colonization of cashew plants by Lasiodiplodia theobromae: Microscopical features C.R. Muniz a , F.C.O. Freire a , F.M.P. Viana a , J.E. Cardoso a , P. Cooke b , D. Wood c , M.I.F. Guedes d,∗ a Centro Nacional de Pesquisa da Agroindústria Tropical, Empresa Brasileira de Pesquisa Agropecuária, Embrapa, Fortaleza, Ceará, Brazil b Eastern Regional Research Center, Agricultural Research Service, U.S. Department of Agriculture, 600 East Mermaid Lane, Wyndmoor, PA, USA c Western Regional Research Center, Agricultural Research Service, U.S. Department of Agriculture, 800 Buchanan Street, Albany, CA 94710, USA d Renorbio, Rede Nordeste de Biotecnologia, Universidade Estadual do Ceará, Centro de Ciência da Saúde, Av. Paranjana, 1700 Fortaleza, Ceará, Brazil article info Article history: Received 17 November 2010 Accepted 8 December 2010 Key-words: Anacardium occidentale L. Immunofluorescence Cell wall degradation Endophytic fungi abstract Lasiodiplodia theobromae is a phytopathogenic fungus causing gummosis, a threatening disease for cashew plants in Brazil. In an attempt to investigate the ultrastructural features of the pathogen coloniza- tion and its response to immunofluorescence labeling, light, confocal and electron microscope studies were conducted on different severity scale patterns of diseased plants. Lasiodiplodia-antisera was checked for cross reactivity against common cashew plants fungi. Optical microscopy analysis revealed a longitu- dinally sectioned hyphae located within the xylem vessels, showing an extensive hyphal development in the secondary xylem tissue. SEM images demonstrated that the fungus was found in some asymptomatic samples, particularly within the xylem vessels as confirmed by the optical images. Symptomatic sample images showed an extensive distribution of the fungus along the secondary xylem, within the vessels, infecting xylem parenchyma. A closer look in the secondary xylem parenchyma reveals a heavy and pro- fuse invasion of the cells with a distinguishable cell wall disintegration and fully hyphae dispersal. There was no reactivity of Lasiodiplodia-antisera against mycelial extracts of Colletotrichum gloeosporioides, Pho- mopsis anardii and Pestalotiopsis guepinii. Following incubation of sections with the polyclonal antisera, the hyphae were intensely and regularly labeled. Rays, vessels and parenchyma cells were the preferred pathway for L. theobromae colonization. Artificial infection provides the information that the vascular cylinder is undoubtedly employed and used by the fungus for hyphae distribution. Immunofluorescence assay employed in situ was applied and the polyclonal antisera produced was able to recognize the fungus and proved to be a sensitive technique to detect it. © 2010 Elsevier Ltd. All rights reserved. 1. Introduction The cashew (Anacardium occidentale L.) produces a nutritious and tasteful kernel that is highly valued as an export commodity, widely consumed in Europe and USA. In 2005, 2.3 million tonnes of cashew nuts were produced in the world and Brazil, which is the third producer, contributed with 11% of the whole trade (FAO, 2008). The high production observed is mainly due to the use of dwarf cashew clones that help to increase the plantation area and also the nut size. However, clonal orchards may contribute to reduce genetic variability, which increases the vulnerability to pathogens (Paiva et al., 2002). Species of Botryosphaeriaceae family are cosmopolitan and occur on a variety of plant hosts causing dieback and canker diseases (von Arx, 1987). This type of species of the genus Lasiodiplodia, L. theo- ∗ Corresponding author. Tel.: +55 85 31019822. E-mail address: fl[email protected] (M.I.F. Guedes). bromae (Pat.) Griffon & Maubl., formerly referred as Botryodiplodia theobromae Pat., is an important pathogenic fungus to both trop- ical and subtropical plants. It has been associated with leaf spots, necrosis, gummosis and even death of many host plants (Encinas, 1996; Encinas and Ahmad, 1999), including herbaceous plants such as cassava (Onyeka et al., 2005) and eggplant (Woodward et al., 2005), and woody plants such as mango (Khanzada et al., 2004), coconut palms (Lakshmanan and Jagadeesan, 2004), eucalyptus (Roux et al., 2001), apricot and peaches (Li et al., 1995), Spondias (Tavares, 2002) and more recently, highbush blueberries (Wright and Harmon, 2009), causing several losses all over the world. It also occurs as an endophyte (Rubini et al., 2005). Less frequently it has also been associated with phaeohyphomycosis (Summerbell et al., 2004) and pneumonia (Woo et al., 2008) in humans. In cashew plants, gummosis is one of the most important dis- eases in Brazil. In fact, under semi-arid conditions in north-eastern Brazil, severe epidemics have occurred in recent years (Freire et al., 2002). Although the disease shows as early as at the first year of cropping, it is only after the second year that severe damage to 0968-4328/$ – see front matter © 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.micron.2010.12.003
Transcript of Colonization of cashew plants by Lasiodiplodia...

Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
Cf
Ca
b
c
d
a
ARA
KAICE
1
awot2oatp
oA
0d
Micron 42 (2011) 419–428
Contents lists available at ScienceDirect
Micron
journa l homepage: www.e lsev ier .com/ locate /micron
olonization of cashew plants by Lasiodiplodia theobromae: Microscopicaleatures
.R. Muniza, F.C.O. Freirea, F.M.P. Vianaa, J.E. Cardosoa, P. Cookeb, D. Woodc, M.I.F. Guedesd,∗
Centro Nacional de Pesquisa da Agroindústria Tropical, Empresa Brasileira de Pesquisa Agropecuária, Embrapa, Fortaleza, Ceará, BrazilEastern Regional Research Center, Agricultural Research Service, U.S. Department of Agriculture, 600 East Mermaid Lane, Wyndmoor, PA, USAWestern Regional Research Center, Agricultural Research Service, U.S. Department of Agriculture, 800 Buchanan Street, Albany, CA 94710, USARenorbio, Rede Nordeste de Biotecnologia, Universidade Estadual do Ceará, Centro de Ciência da Saúde, Av. Paranjana, 1700 Fortaleza, Ceará, Brazil
r t i c l e i n f o
rticle history:eceived 17 November 2010ccepted 8 December 2010
ey-words:nacardium occidentale L.
mmunofluorescenceell wall degradationndophytic fungi
a b s t r a c t
Lasiodiplodia theobromae is a phytopathogenic fungus causing gummosis, a threatening disease forcashew plants in Brazil. In an attempt to investigate the ultrastructural features of the pathogen coloniza-tion and its response to immunofluorescence labeling, light, confocal and electron microscope studieswere conducted on different severity scale patterns of diseased plants. Lasiodiplodia-antisera was checkedfor cross reactivity against common cashew plants fungi. Optical microscopy analysis revealed a longitu-dinally sectioned hyphae located within the xylem vessels, showing an extensive hyphal development inthe secondary xylem tissue. SEM images demonstrated that the fungus was found in some asymptomaticsamples, particularly within the xylem vessels as confirmed by the optical images. Symptomatic sampleimages showed an extensive distribution of the fungus along the secondary xylem, within the vessels,infecting xylem parenchyma. A closer look in the secondary xylem parenchyma reveals a heavy and pro-fuse invasion of the cells with a distinguishable cell wall disintegration and fully hyphae dispersal. There
was no reactivity of Lasiodiplodia-antisera against mycelial extracts of Colletotrichum gloeosporioides, Pho-mopsis anardii and Pestalotiopsis guepinii. Following incubation of sections with the polyclonal antisera,the hyphae were intensely and regularly labeled. Rays, vessels and parenchyma cells were the preferredpathway for L. theobromae colonization. Artificial infection provides the information that the vascularcylinder is undoubtedly employed and used by the fungus for hyphae distribution. Immunofluorescenceassay employed in situ was applied and the polyclonal antisera produced was able to recognize the fungusive te
and proved to be a sensit. Introduction
The cashew (Anacardium occidentale L.) produces a nutritiousnd tasteful kernel that is highly valued as an export commodity,idely consumed in Europe and USA. In 2005, 2.3 million tonnes
f cashew nuts were produced in the world and Brazil, which ishe third producer, contributed with 11% of the whole trade (FAO,008). The high production observed is mainly due to the usef dwarf cashew clones that help to increase the plantation areand also the nut size. However, clonal orchards may contributeo reduce genetic variability, which increases the vulnerability to
athogens (Paiva et al., 2002).Species of Botryosphaeriaceae family are cosmopolitan and occurn a variety of plant hosts causing dieback and canker diseases (vonrx, 1987). This type of species of the genus Lasiodiplodia, L. theo-
∗ Corresponding author. Tel.: +55 85 31019822.E-mail address: [email protected] (M.I.F. Guedes).
968-4328/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.micron.2010.12.003
chnique to detect it.© 2010 Elsevier Ltd. All rights reserved.
bromae (Pat.) Griffon & Maubl., formerly referred as Botryodiplodiatheobromae Pat., is an important pathogenic fungus to both trop-ical and subtropical plants. It has been associated with leaf spots,necrosis, gummosis and even death of many host plants (Encinas,1996; Encinas and Ahmad, 1999), including herbaceous plants suchas cassava (Onyeka et al., 2005) and eggplant (Woodward et al.,2005), and woody plants such as mango (Khanzada et al., 2004),coconut palms (Lakshmanan and Jagadeesan, 2004), eucalyptus(Roux et al., 2001), apricot and peaches (Li et al., 1995), Spondias(Tavares, 2002) and more recently, highbush blueberries (Wrightand Harmon, 2009), causing several losses all over the world. It alsooccurs as an endophyte (Rubini et al., 2005). Less frequently it hasalso been associated with phaeohyphomycosis (Summerbell et al.,2004) and pneumonia (Woo et al., 2008) in humans.
In cashew plants, gummosis is one of the most important dis-eases in Brazil. In fact, under semi-arid conditions in north-easternBrazil, severe epidemics have occurred in recent years (Freire et al.,2002). Although the disease shows as early as at the first year ofcropping, it is only after the second year that severe damage to

Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
4 icron
tsenawtiticseotni
cUcab(opdutmi
2
2
toPCbsfdca
eosntk5
apalkgiffi
20 C.R. Muniz et al. / M
he cashew tree occurs. Gummosis symptoms are characterized bywollen cankers in the trunk or in woody branches, which mayventually crack and ooze a transparent resin-like gum; hence theame of the disease (Freire et al., 2002). It is presumed that dam-ge to the plant by gummosis is brought about by reduction inater and nutrient transport and destruction of branches, leading
o reduced photosynthesis; and plant death. Gummosis importances enhanced by the fact that no practical measures for controllinghe disease have proved efficient under epidemic situation. Whennfected tissue is located in the trunk, the surgical removal of theanker, followed by a copper fungicide treatment to protect the cuturface until healing, has been recommended (Freire, 1991). How-ver, the prevalence of branch infections and observed reinfectionf the trunk after 2 or 3 months (Cardoso et al., 1995) contribute tohe low effectiveness of this method. Pathogen dissemination wasot totally revealed, although evidence of seed and propagule borne
nfection was obtained (Cardoso et al., 1998; Freire et al., 1999).Microscopical observations are extremely useful for a more
omprehensive understanding of pathogen colonization pathways.ltrastructural studies of the attack of L. theobromae was previouslyarried out by Encinas (1996) in Caribbean and Scot pine woodnd birch wood, by Encinas and Daniell (1997) in aspen and rub-erwood and more recently by Sato et al. (2008) in Jew’s marrownalta jute, Corchorus olitorius L.). Results showed a high efficiencyf this fungus to decay secondary cell walls and lignin. In cashewlants, ultrastructural studies of gummosis have so far not beenone and the complete characterization of the disease remainednexplained. The present studies were undertaken to investigatehe ultrastructural features of the infection and pathogen develop-
ent within cashew tissues and to explore the feasibility of usingmmunofluorescence labeling for detection of L. theobromae.
. Materials and methods
.1. Naturally infected plant material and fungus isolates
In total, 32 samples of nuts, shells and branches from plants ofhe commercial clone CCP-76 were collected from different areasf northeastern Brazil, including the municipal district of Pio IX,I (6◦31′30S, 40◦47′19W, 600–605 m asl) and Pacajus, State ofeará. This cashew orchard area has been studied and proved toe infested by L. theobromae (Cysne et al., 2010), showing differenteverity scale patterns of diseased plants: no symptoms; small andew cankers on trunk and branches, small cracks without gum exu-ation; cracked cankers on trunks and branches, cracked cankersompletely girdling trunk or branches, foliage yellowing, diebacknd gum exudation.
Small blocks of samples collected from plants showing differ-nt patterns of symptoms were obtained and fixed with a mixturef 1% (v/v) glutaraldehyde and 4% (w/v) formaldehyde in 100 mModium cacodylate buffer (pH 7.2). L. theobromae isolates origi-ally obtained from gummosis infected tissue were obtained fromhe collection maintained at EMBRAPA-Agroindústria Tropical andept on Potato Dextrose Agar (PDA) (Difco Laboratories, Detroit) at◦C.
The fungal isolates Colletotrichum gloeosporioides, Phomopsisnardii, Pestalotiopsis guepinii, usually found also within cashewlants were used to test specificity of Lasiodiplodia-antisera as wells a strain of Neofusiccocum sp. and were obtained from the col-ection maintained at EMBRAPA-Agroindústria Tropical and also
ept on PDA at 5 ◦C. These fungi and Lasiodiplodia isolates wererown in Potato Dextrose Broth (PDB) (Difco Laboratories, Detroit)n flasks on a rotary shaker at 140 rpm for 10 days at 25 ◦C. Theungal material was then macerated in liquid nitrogen and thene powder obtained was resuspended in general extraction buffer42 (2011) 419–428
(GEB), which consisted of 0.01 M phosphate buffer (pH 7.4), 0.01 Msodium sulfite, 2% polyvinylpyrrollidone, 0.2% powdered bovineserum albumin and 2% tween-20. Saline (PBS), constituting themycelial extracts for ELISA protocol.
2.2. Artificially infected plant material
A total of two-month-old 45 seedlings originated from seed ger-mination of the cashew commercial clone (CCP 76) were grownin a growth chamber at 25 ◦C, 60–70% relative humidity witha photoperiod of 12/12 h light/dark, and provided an approxi-mate 180 �mol/m2/s radiation (PAR). Plants were inoculated withmycelium of L. theobromae in the stems of plants using a small elec-trical drill to make a hole, in which the mycelium was put inside andcovered with a tape. Experimental design was completely random-ized designed with 3 treatments (plants with no inoculation, plantsinoculated with L. theobromae and plants inoculated only with PBS(also through the hole in the stem) and five repetitions with por-tions of three pots containing one plant per pot. After every week,until 45 days, samples from leaves, petioles, branches and stemswere collected for light microscopy.
2.3. Light microscopy
Samples fragments were washed in 0.1 M phosphate buffer andthen dehydrated in a graded ethanol series before being embed-ded in a tissue freezing medium overnight. Sections (40 �m) werecut with an IEC cryotome (Minotome Plus, Needham Heights, MA),collected on glass slides and stained with toluidine blue. Imageswere collected with a TCS-SP confocal system with an invertedmicroscope (Leica Microsystems, Wetzlar, Germany).
2.4. Fluorescence microscopy
Stereofluorescence microscopy was performed with a LeicaMZFLIII stereofluorescent dissecting microscope, using a reflectedwhite light and a blue-induced fluorescence. Images were capturedby using a charge-coupled device (CCD) camera and formatted inPhotoshop (Adobe Systems, Mountain View, CA).
2.5. Immunization and antibody generation
L. theobromae was grown on PDA and agar disks containing fun-gus mycelia were transferred into 250 ml flasks containing 150 mlof 2% Sabouraud Dextrose Broth and incubated in a rotary shakerat 150 rpm at 28 ◦C for 14 days. Mycelia were harvested onto cellu-lose filters in a Buchner funnel under vacuum, washed with sterilewater, minced in liquid nitrogen, lyophilized and stored at −20 ◦C.Antiserum was raised in New Zealand white rabbit following thestandard immunization protocol (Harlow and Lane, 1988). A femaleNew Zealand adult rabbit was immunized subcutaneously with100 �g of L. theobromae-cultured mycelial antigen emulsified in2 mL of complete Freund’s adjuvant (Sigma, St. Louis, MO, USA). Theanimal was bled and serum was used for titration and immunos-taining.
2.6. Titre and specificity of antisera
For determining the titre and specificity of the antisera, anindirect enzyme-linked immunosorbent assay (ELISA) was used.Working volume of 100 �L per well in ninety-six-well microtitre
plates and incubation at 4 ◦C overnight was employed betweeneach step. Between each step the plates were washed four timeswith PBS–Tween (PBS containing 0.05% Tween 20). The plates werepreviously treated with 0.005% poly-l-lysine (PLL), to enhance theadhesion of the mycelial extracts to the wells (Banks and Cox,
Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
C.R. Muniz et al. / Micron 42 (2011) 419–428 421
Table 1Reactivity of Lasiodiplodia theobromae isolates, other Botryosphaeriaceae fungi and other fungal genera from cashew plants in L. theobromae enzyme-linked immunosorbentassay.
Fungus species, isolate number Host, place of origin OD at 405 nma
Lasiodiplodia theobromae #21 Anacardium occidentale-gummosis symptoms, São Raimundo Nonato, Piauí, Brasil 2.7528Lasiodiplodia theobromae #23 Anacardium occidentale-gummosis symptoms, Pacajus, Ceará, Brasil 2.7121Neofusiccocum sp. #11 Euterpe oleraceae, Ceará, Brasil 1.8345Colletotrichum gloeosporioides #1 Anacardium occidentale-symptomless, Ceará, Brasil 0.4011Colletotrichum gloeosporioides #1b Anacardium occidentale-symptomless, Ceará, Brasil 0.3415
ptomlptoml
30 m
11eogwct0muo1a
2
ptoobwgwcs
2
(tetatoM
2
gtFwbQmnt
Pestalotiopsis guepinii #2 Anacardium occidentale-symPhomopsis anacardii #20 Anacardium occidentale-sym
a OD = optical density measurements, made after the substrate was incubated for
992). To determine the titre of antibodies, dilutions of 1:100 to:18,000 were used against five isolates of Lasiodiplodia-mycelialxtracts. Cross reactivity was tested by employing mycelial extractsf Neofusiccocum sp. (also from Botryosphaeriaceae family), C.loeosporioides, P. anardii and P. guepinii. The mycelial extractsere added into the wells and after the incubation, plates were
oated with the rabbit antisera at dilutions of 1:750 in Extrac-ion buffer (0.05% Tween-20, 2% polyvinylpyrrolidone (PVP-25),.2% ovalbumine, grade VI, 0.01 M PBS). Anti rabbit IgG (wholeolecule) alkaline phosphatase conjugate (Sigma, A-8025) was
sed at 0.7 units/mL diluted in Extraction buffer. Plates were devel-ped with p-nitrophenyl phosphate substrate at 1.0 mg/mL inM diethanolamine buffer (pH 9.8) and measured at 405 nm ofbsorbance on a SpectraFluor programme (Tecan).
.7. Immunostaining
The sections obtained in the cryotome were quenched withhosphate-buffered saline (PBS), pH 7.2 containing 0.1 M glycineo inactivate any residual aldehyde groups and then was carriedut as follows: the primary antibodies were used at a dilution ratef 5 �g/mL in phosphate-buffered saline (PBS). Cells and the anti-ody were incubated at room temperature for 1.5 h. After the cellsere washed, the second antibody, Alexa Fluor 546-conjugated
oat anti-rabbit IgG (Molecular Probe), diluted 1:100, was reactedith the samples at room temperature for 1 h. After washing, the
ells were observed by fluorescent microscopy or confocal lightcanning microscopy.
.8. Scanning electron microscope
For SEM analysis, the fixed material was rinsed three times10 min each) in phosphate buffer. After three 10 min rinses in dis-illed water, samples were dehydrated through 70, 80, 90, 100%thanol (5 min in each stage) and three changes in 100% at roomemperature. Ethanol was then replaced by liquid carbon dioxidend samples dried using EMS 850 critical point drying appara-us. They were then mounted on stubs, coated with gold andbserved in a Quanta 200 FEG Environmental Scanning Electronicroscope.
.9. Transmission electron microscope
Fixed pieces (3–4 mm) confirmed for the presence of the fun-us using light microscopy were post-fixed with 1% (w/v) osmiumetroxide in phosphate-buffered saline (PBS), pH 7.2 for 1 h at 4 ◦C.ollowing serial dehydration steps including 20 min in each changeith 80, 95 and 100% ethyl alcohol and with propylene oxide,
locks were embedded with LR White (J.B. EM Services Inc., Dorval,uebec, Canada). Ultrathin sections were cut from using a dia-ond knife and then mounted on Formvar/carbon 100 mesh coated
ickel grids (Canemco-Marivac, St. Laurent, Quebec, Canada). Sec-ions were subsequently stained with 5% aqueous uranyl acetate for
ess, Ceará, Brasil 0.1860ess, Ceará, Brasil 0.2173
in. Values are means of four replicate wells per isolate.
15 min, prior to viewing with a Philips EM301 transmission electronmicroscope.
3. Results
3.1. Response of mycelium extracts of cashew fungi toanti-Lasiodiplodia mycelium sera
The anti-Lasiodiplodia mycelium sera used at a dilutionof 1:12,000 (v/v) as the detection antibody provided optimalreactivity with Lasiodiplodia isolates originated from gummosissymptomatic cashew plants in the ELISA (Table 1). It was also usedto test the specificity of the antisera against mycelial extracts of C.gloeosporioides, P. anardii and P. guepinii. Highest absorbances wereobserved in the mycelial extracts of L. theobromae isolates. Reac-tivity against the isolate Neofusiccocum sp. was higher than othergenera, but lower than Lasiodiplodia.
3.2. Microscopy
Optical microscopy analysis of the branches revealed a longitu-dinally sectioned hyphae located within the xylem vessels, showingan extensive hyphal development in the secondary xylem tissue(Fig. 1a–d). A high hyphal diameter and cell wall thickness wereobserved with the direction growth along the vessels (Fig. 1c andd). For some asymptomatic samples (3 of 5 samples), the funguswas found to occur only inside the vessels. Upon examination ofbranches from infected and severe symptomatic plants sections,it was apparent that spread of the fungus was initially throughthe secondary xylem, within the xylem vessels, moving then indirection to adjacent parenchyma cells and rays (Fig. 1e and f).
Scanning electron microscopy (SEM) demonstrated that thefungus was found in some asymptomatic samples, particularlywithin the xylem vessels as confirmed by the optical images. Forthese samples, fungus was not seen along the parenchyma cells(Fig. 2a–e). Transverse section of others asymptomatic samples didnot show the presence of the fungus (Fig. 2f). Symptomatic sampleimages showed an extensive distribution of the fungus along thesecondary xylem, within the vessels, infecting cells of the xylemparenchyma, especially near to the cankers aggravated during dis-ease process (Fig. 3a–c). The fungal hyphae was found crossingthe cell walls, presenting great diameter variability and growingboth transversely and longitudinally, occupying extensive areas ofthe parenchyma cells interior (Fig. 3c). Some interior cells wereseen with large masses of homogeneous groups of hyphae showingboth regular and irregular septal arrangements (Figs. 3f and 4a–e).A closer look in the cells from the secondary xylem parenchymareveals a heavy and profuse invasion of the cells with a distinguish-
able cell wall disintegration and fully hyphae dispersal (Fig. 3d–f).The fungus was never seen in the phloem or in the medulla, indicat-ing that these tissues were not used by the fungus to spread withinthe plant. The inter- and intracellular growth of the pathogen in thehost tissues was associated with pronounced alterations including
Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
422 C.R. Muniz et al. / Micron 42 (2011) 419–428
F broms issue( arrow
dgdmwa
cHIrfiivF
ig. 1. Microphotographs of cashew plants cells infected with Lasiodiplodia theoize = 30 �m. (b) Hyphae of Lasiodiplodia theobromae within xylem parenchyma td–f) Hyphae spreading from xylem vessel in direction to parenchyma tissue (black
isintegration and digestion of the host cell walls, which sug-ests that the pathogen may produce cell-wall-degrading enzymesuring infection and spreading in the host tissues. Under higheragnifications, penetration of the fungus hyphae into the cell wallsas clearly visible, indicating that cell walls complex carbohydrates
re easily broken by its enzymatic apparatus.Profuse distribution of the fungus hyphae within parenchyma
ells is widely seen as well as many holes on the cell wall surfaces.igher magnification shows large hyphae distribution (Fig. 4b).
t possibly means that penetration of the fungus into the cells iselated to the growth and expansion of mycelium. At the same timeungi spreads along the tissue, cell walls are being broken down by
ts enzymes. This idea is strongly supported by images providedn Fig. 4d–f. Attachment of the hyphae to the cell walls are clearlyisible, specially the distribution of fungus between neighbor cells.reeze fracture of samples allowed the rupture of cells exactly atae. (a) Hyphae of Lasiodiplodia theobromae growing along longitudinal rays. Bar(black arrow). Bar size = 30 �m. (c) A high diameter hyphae within xylem vessel.
s).
the intersection between them, evidencing the sites of penetrationand invasion (Fig. 4f).
Following incubation of sections with polyclonal antisera, thefungus hyphae were intensely and regularly labeled. Host tis-sues structures were free of significant labeling. Control sectionsincubated with secondary antibody probe alone exhibited no label-ing. Again, inter and intracellular hyphae of different diameterswere observed in the parenchyma, showing an extensive spreadwith hyphae growth occurring both transversely and longitudinally(Fig. 5a and b). Numerous hyphal strands were observed spreadingnear to the xylem vessels (Fig. 5b).
Artificially infected samples were followed in time and 7 days
after inoculation, hyphae are found between adjacent epidermalcells (Fig. 6a). Transversal sections of leaves provide the observa-tion of several globose and strongly stained structures, resemblingpicnidiospores that evolves to hyphae segments, specially near to
Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
C.R. Muniz et al. / Micron 42 (2011) 419–428 423
F f cash( withit
xtol
whtSTt
4
cCtmnd
w
ig. 2. SEM micrographs showing aspects of Lasiodiplodia theobromae colonization oH). Natural entrances and perforations are widely used by the fungus to spreadheobromae. (c and d) Hyphae spreading along xylem vessels.
ylem and phloem (Fig. 6b and c), and at least, 21 days of inocula-ion. Hyphae was found near to the infection hole at the beginningf the experiment, and after 45 days of inoculation, spread along theeaves surface (Fig. 6d), and also presenting gum exudation (Fig. 6e).
Cell wall penetration is clearly shown by means of TEM (Fig. 7),here degradation of cell wall layers takes place after fungalyphae invasion. It starts from the attachment of the hyphae tohe innermost layer of the secondary wall (Fig. 7a), delamination of2 layer and cell wall penetration occurs afterwards (Fig. 7b and c).he lumen is plentiful occupied by hyphae with evident damage tohe cell wall (Fig. 7d).
. Discussion
Gumosis is a serious and threatening disease, affecting theashew agri-industry in the Brazil Northeast regions. Expansion ofCP-76 cultivar, which is highly susceptible to its causative agent,he fungus L. theobromae, has allowed the emergence of epidemics,
ainly due to the destructive character of the infection, dissemi-ation through rootstocks and the lack of studies involving earlyetection.
The whole comprehension of L. theobromae pathogenicityithin cashew plants, including adhesion, colonization and infec-
ew plants branches. Vessels (V) and rays (R) intensively colonized by fungal hyphaen vessels system (arrows). (a and b) Transverse ray colonization by Lasiodiplodia
tion processes as well as the plant reaction are essential to givesupport to gummosis resistance studies. Little information existson the latency of this pathogen. This research is the first full reportdescribing the microscopical details of cashew tissues colonized byL. theobromae.
L. theobromae mycelial extracts were recognized as antigensby the Lasiodiplodia-antisera, which did not react with mycelialextracts of others related cashew fungi. Cross-reaction with Neofus-iccocum sp. demonstrates that these close taxonomic related generashare common antigens.
Symptomatic and asymptomatic plants presented different fea-tures. Although some asymptomatic plants did not present thefungus, some xylem vessels from other asymptomatic sampleswere found to contain abundant Lasiodiplodia hyphae. It is pos-sibly due to the fact that the fungus is able to inhabit the plantwithout causing apparent disease symptoms. After some externalsignal, which may be an abiotic or a biotic stress, symptoms beginto appear and disease becomes established. Müllen et al. (1991)
reported L. theobromae colonization of healthy plant tissues with-out exhibiting symptoms. They carried out pathogenicity tests ondogwood stems with and without drought stress and managed toisolate L. theobromae from all inoculated plants, but cankers devel-oped only on stressed plants.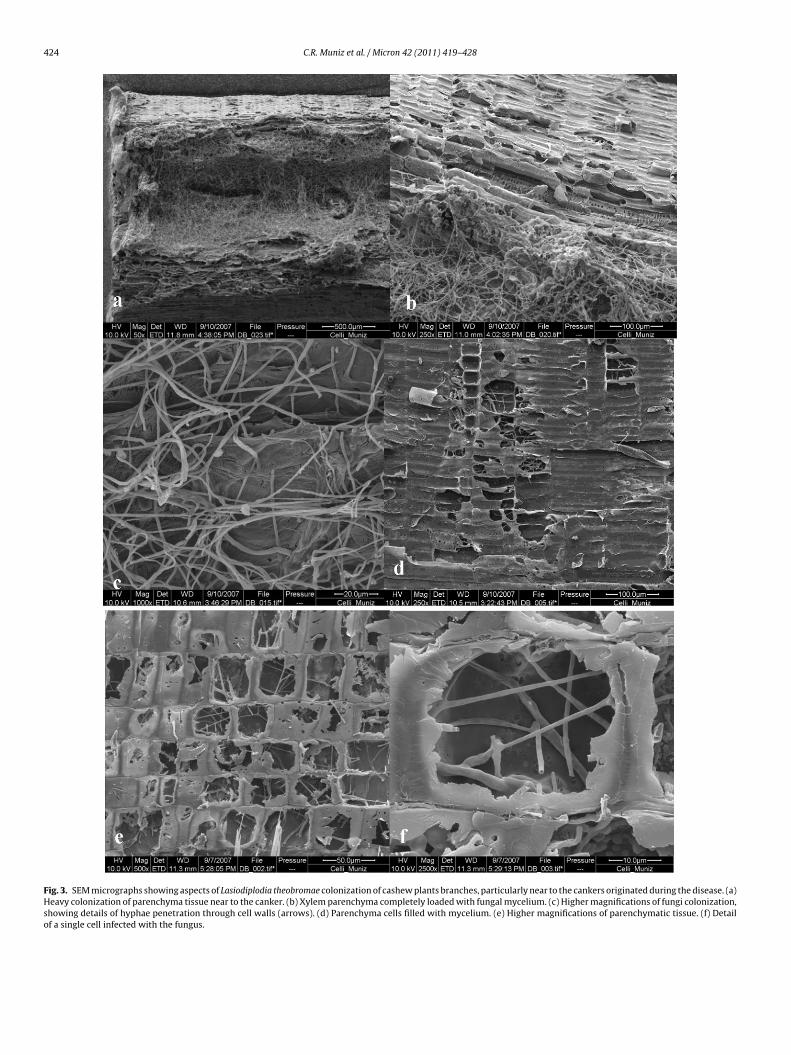
Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
424 C.R. Muniz et al. / Micron 42 (2011) 419–428
Fig. 3. SEM micrographs showing aspects of Lasiodiplodia theobromae colonization of cashew plants branches, particularly near to the cankers originated during the disease. (a)Heavy colonization of parenchyma tissue near to the canker. (b) Xylem parenchyma completely loaded with fungal mycelium. (c) Higher magnifications of fungi colonization,showing details of hyphae penetration through cell walls (arrows). (d) Parenchyma cells filled with mycelium. (e) Higher magnifications of parenchymatic tissue. (f) Detailof a single cell infected with the fungus.

Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
C.R. Muniz et al. / Micron 42 (2011) 419–428 425
F diplo( th hypc ation
fehsm(otn
ig. 4. SEM micrographs of longitudinal sections of cashew plants infected with Lasioa and b) Crescent magnifications of the same region, showing a cell fully filled wirossing cell walls. (e) Hyphae strongly attached to the cell walls. (f) Hyphae penetr
Studies on various biological aspects of the Botryosphaeriaceaeamily have begun to provide efficient tools to characterize theirndophytic nature. Based on the wide taxonomic distribution andigh frequency of endophytic infection on various hosts for thosepecies examined to date for endophytism, it is now thought thatost, if not all Botrysphaeriaceae, might have an endophytic phase
Slippers and Wingfield, 2007). As defined by Petrini (1991), anyrganism that establishes a symptomless infection of the internalissues of a plant is considered an endophyte. Plant pathologistsame these infections as latent and visual field infections fail as
dia theobromae, showing aspects of invasion sites with pits and roles (white arrows).hae. (c) Hyphae spreading within and outside the cell (black arrows). (d) Hyphaewithin the intercellular space of two adjacent cells (white arrow).
a detection method. Microbiological, microscopic, molecular andgenetic tools are extremely useful to characterize these interac-tions (Bacon and White, 2000). Although the need for expensiveequipment for sample preparation and visualization, the extent anddetermination of colonization sites are only possible to be reachedusing microscopical techniques, mainly electron microscopy. In a
study with cashew vegetative propagules carried out by Cardosoet al. (2009), transplanting materials were produced by using root-stocks from seeds originated from healthy and gummosis-infectedplants. Soon after, L. theobromae was isolated from up to 80 cm dis-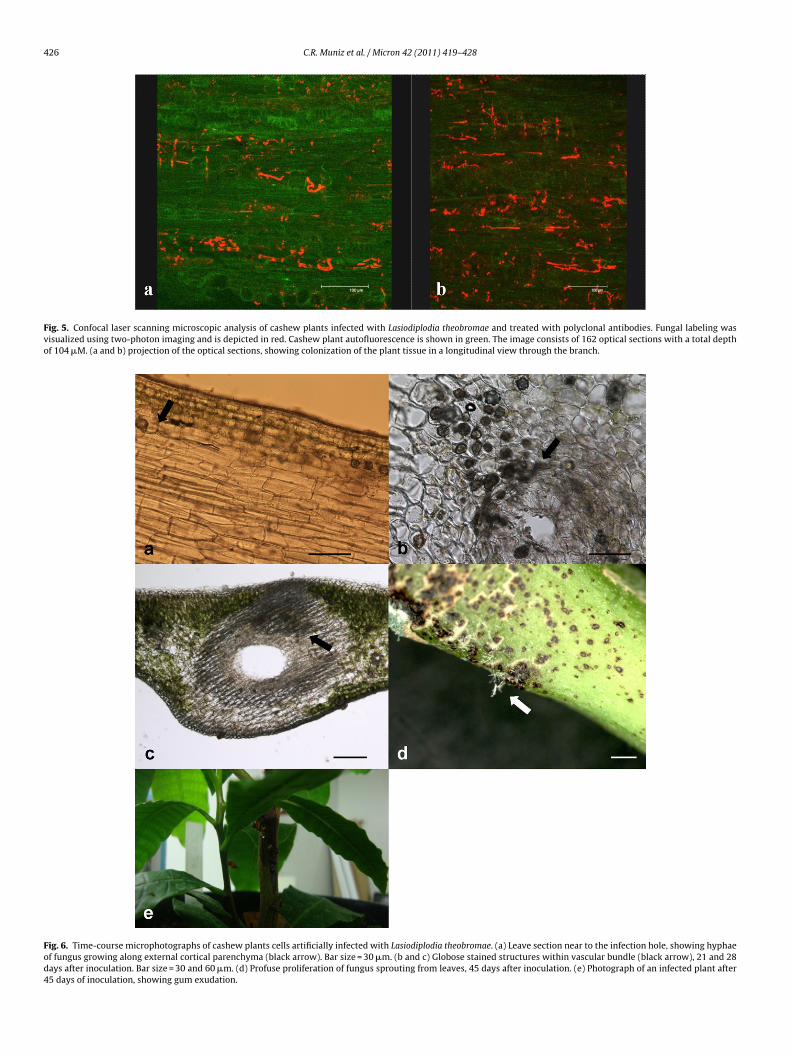
Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
426 C.R. Muniz et al. / Micron 42 (2011) 419–428
Fig. 5. Confocal laser scanning microscopic analysis of cashew plants infected with Lasiodiplodia theobromae and treated with polyclonal antibodies. Fungal labeling wasvisualized using two-photon imaging and is depicted in red. Cashew plant autofluorescence is shown in green. The image consists of 162 optical sections with a total depthof 104 �M. (a and b) projection of the optical sections, showing colonization of the plant tissue in a longitudinal view through the branch.
Fig. 6. Time-course microphotographs of cashew plants cells artificially infected with Lasiodiplodia theobromae. (a) Leave section near to the infection hole, showing hyphaeof fungus growing along external cortical parenchyma (black arrow). Bar size = 30 �m. (b and c) Globose stained structures within vascular bundle (black arrow), 21 and 28days after inoculation. Bar size = 30 and 60 �m. (d) Profuse proliferation of fungus sprouting from leaves, 45 days after inoculation. (e) Photograph of an infected plant after45 days of inoculation, showing gum exudation.

Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
C.R. Muniz et al. / Micron 42 (2011) 419–428 427
F . theob( ow) (
tbr
ncgaScsiolssoetUbwh2tptoawsibhs
LVgmTnbi
ig. 7. Transmission electron micrographs of cashew plants sections colonized by Lb) Cell wall penetration by hyphae, showing total dissolution of S2 layer (white arr
ance from canker in both directions, suggesting the endophyticehavior of L. theobromae in cashew tissues and the importance ofootstock seed as a source of primary inoculum.
Pit fields are used by the fungus, deriving benefits from theseatural entrances. When this mechanism finishes or is limited,ell wall degradation takes place. Encinas and Daniell (1996) sug-ested an initial stage of L. theobromae where it uses proteinaceousnd other substances such as starch, pectins and free sugars.ubsequent degradation of ray parenchyma, cell walls and resinhannels occur after assimilable substances have been used. Con-idering the extensive degradation of the cell walls by the fungi,t is possibly that the attack results from the combined actionf different enzymes. According to Joseleau and Ruel (1994), cel-ulose and hemicellulose breakdown by fungi is related to theynergistic action of several hydrolytic and oxidative enzymes,uch as glucanases, cellobiohydrolases, glucosidases (cellulases)r xylanases, xylosidases and glucuronidases (hemicellulolyticnzymes). However, the magnitude and intensity of the degrada-ive ability depends on the aggressiveness of L. theobromae strain.ltrastructural investigations of bamboo culm exposed to L. theo-romae presented penetration patterns via outer and inner culmall and the metaxylem was the main route of access for itsyphae, fully able to spread to adjacent culm tissue (Kleist et al.,002). Florence et al. (2002) clearly demonstrated through elec-ron microscopy evidences that B. theobromae has the ability toroliferate and colonize rubberwood tissues from cell to cell pene-ration. Encinas and Daniell (1997) have shown that several strainsf L. theobromae lead to G-layer and true cell wall degradation inspen and rubberwood, with pronounced erosion attack. In thisork, images also provide evidences that cell wall is fully degraded,
uggesting that enzymes rather than hydrostatic pressure may benvolved in the breakdown of pit membranes. As observed in rub-erwood, cell wall invasion and disruption of inner layers alsoappen within cashew plants infected and showing gummosisymptoms.
Cedeno et al. (1996) carried out microscopical observations of. theobromae, infecting Caribbean pine in the Eastern region ofenezuela. A high variability in hyphal diameter of this fungus andreat extension of colonization sites were also reported. Further-
ore, it was observed that thicker hyphae form the thinner ones.he direction of the hyphae expansion seems to be driven by autrient requirement, thus vessels and cellular content are possi-ly a source for this supply. Some endophytes are only able to assess
ntercellular spaces as they do not develop specialized structures or
romae. (a) Attachment of the hyphae to the innermost layer of the secondary wall.c) hyphae occupying the cell lumen. H, hyphae; CW, cell wall.
enzymes. Our observations confirm prior studies made on woodyspecies, where rays, vessels and parenchyma cells become the pre-ferred pathway for L. theobromae colonization. Artificial infectionfollowed in time is also a clue that the vascular bundle is undoubt-edly employed and used by the fungus for hyphae distribution.Initial samples collected after 7 days of inoculation show fungalcolonization near to the epidermis rather than xylem. This is dueto the fact that samples endophytically and naturally colonized byLasiodiplodia, already present the fungus deeply attached to theinner tissues. It occurs when samples are collected from an infestedorchard in which the plants have been infected during the graftingtechniques, for example. Oppositely, in artificially infected samples,the fungus flux happens from outside to inside. However, in bothmechanisms, the fungus will reach the xylem anyway.
L. theobromae is a serious threatening for cashew crop areas inBrazil. Its capacity to colonize vessels, rays and parenchyma cells,leading to the disruption of cell walls in cashew plants is a proofof its versatility and aggressiveness. The pattern of infection forcashew plants shown in this work is very similar to data found forothers woody samples, such as aspen, rubberwood and pine. Thesebroad colonization sites coupled with its endophytic nature high-light the importance to develop methods to detect it in seedlings,seeds or any other asymptomatic propagative material as an effortto manage and control this pathogen. Immunofluorescence assayemployed in situ was applied and the polyclonal antisera producedwas able to recognize the fungus and proved to be a sensitive tech-nique to detect it. Fungi in situ recognition by antiserum and ELISAresults encourage further steps for the production of serologicaldetection methods.
Acknowledgment
We wish to thank Embrapa Labex USA for the support receivedduring the experiments, especially to Luiz H.C. Mattoso and PedroAntonio Arraes Pereira.
References
Bacon, C.W., White, J.F., 2000. Microbial Endophytes. Marcel Dekker Inc., New York,
USA.Banks, J.N., Cox, S.J., 1992. The solid phase attachment of fungal hyphae in na ELISAto screen for antifungal antibodies. Mycopathologia 120, 79–85.
Cardoso, J.E., Aragão, M.L., Bleicher, E., Cavalcante, M.J.B., 1995. Efeito de práticasagronômicas na ocorrência da resinose do cajueiro. Fitopatologia Brasileira 20,242.

Journal Identification = JMIC Article Identification = 1594 Date: March 10, 2011 Time: 5:4 pm
4 icron
C
C
C
C
E
E
E
E
F
F
F
F
F
H
J
K
K
L
28 C.R. Muniz et al. / M
ardoso, J.E., Freire, F.C.O., Sá, F.T., 1998. Disseminacão e controle da resinose emtroncos de cajueiro decepados para substituicão de copa. Fitopatologia Brasileira23, 48–50.
ardoso, J.E., Bezerra, M.A., Viana, F.M.P., Sousa, T.R.M., Cysne, A.Q., Farias, F.C.,2009. Endophyte occurrence of Lasiodiplodia theobromae in cashew tissues andits transmission by vegetative propagules. Summa Phytopathologica 35 (4),262–266.
edeno, L., Mohali, S., Palacios-Prü, E., 1996. Ultrastructure of Lasiodiplodia theo-bromae causal agent of caribbean pine blue stain in Venezuela. Interciencia 21,264–265.
ysne, A.Q., Cardoso, J.E., Maia, A.H.N., Farias, F.C., 2010. Spatial-temporal analysis ofgummosis in three cashew clones at Northeastern Brazil. Journal of Phytopathol-ogy 158, 676–682.
ncinas, O., 1996. Development and Significance of Attack by Lasiodiplodia theobro-mae (Pat.) Griff. & Maubl. in Caribbean Pine Wood and Some Other Wood Species.Uppsala, Sweden: Swedish University of Agricultural Sciences, PhD Thesis.
ncinas, O., Daniel, G., 1996. Wood cell wall biodegradation by the blue stain fungusBotryodiplodia theobromae. Material und organismen 29, 256–272.
ncinas, O., Daniell, D., 1997. Degradation of the gelatinous layer in aspen and rub-berwood by the blue stain fungus Lasiodiplodia theobromae. IAWA Journal l8,107–115.
ncinas, O., Ahmad, T., 1999. Mannitol and arabitol, metabolic products producedby Lasiodiplodia theobromae in wood. Interciencia 24, 267–268.
AO (Food and Agriculture Organization of the United Nations). FAOSTAT, 2008[accessed 24 november 2008]. Available: http://faostat.fao.org/.
lorence, E.J.M., Gnanaharan, R., Singh, P.A., Sharma, J.K., 2002. Weight loss andcell wall degradation in rubberwood caused by sapstain fungus Botryodiplodiatheobromae. Holzforschung 56, 225–228.
reire, F.C.O., 1991. A Resinose do Cajueiro. Caju Informativo (EMBRAPA/CNPCa) 4,2.
reire, F.C.O., Kozakiewicz, Z., Paterson, R.R.M., 1999. Mycoflora and mycotoxins ofBrazilian cashew kernels. Mycopathology 145, 95–103.
reire, F.C.O., Cardoso, J.E., Santos, A.A., Viana, F.M.P., 2002. Diseases of cashew (Anac-ardium occidentale L.) in Brazil. Crop Protection 21, 489–494.
arlow, E., Lane, D., 1988. Antibodies, a Laboratory Manual. Cold Spring HarborLaboratory Press, New York, USA.
oseleau, J.P., Ruel, K., 1994. Wood polysaccharides and their degradation by fungi.In: Petrini, O., Ouellette, G.B. (Eds.), Host Wall Alterations by Parasitic Fungi. APSPress, The American Phytopathological Society, St. Paul, USA, pp. 45–54.
leist, G., Morris, I., Murphy, R., 2002. Invasion and colonisation of bamboo culmmaterial by stain and decay fungi. Proceedings of the 33rd Annual MeetingCardiff, United Kingdom, 2002. London, United Kingdom: The International
Research Group on Wood Preservation, IRG/WP 02-10453.hanzada, M.A., Lodhi, A.M., Shahzad, S., 2004. Pathogenicity of Lasiodiplodia theo-bromae and Fusarium solani on mango. Pakistan Journal of Botany 36, 181–189.
akshmanan, P., Jagadeesan, R., 2004. Malformation and cracking of nuts in coconutpalms (Cocos nucifera) due to the interaction of the eriophyid mite Aceria
42 (2011) 419–428
guerreronis and Botryodiplodia theobromae in Tamil Nadu, India. Zeitschrift furPflanzenkrankheiten und Pflanzenschutz. Journal of Plant Diseases and Protec-tion 111, 206–207.
Li, H.Y., Cao, R.B., Mu, Y.T., 1995. In-vitro inhibition of Botryosphaeria-dothideaand Lasiodiplodia-theobromae, and chemical control of gummosis disease ofJapanese apricot and peach-trees in Zhejiang province, China. Crop Protection14, 187–191.
Müllen, J.M., Gilliam, C.H., Hagen, A.K., Morgan Jones, G., 1991. Lasiodiplodia theo-bromae cancer of dogwood, a disease influenced by drought stress or cultivarselection. Plant Disease 75, 886–889.
Onyeka, T.J., Ekpo, E.J.A., Dixon, A.G.O., 2005. Virulence and host-pathogen interac-tion of Botryodiplodia theobromae isolates of cassava root rot disease. Journal ofPhytopathology 153, 726–729.
Paiva, J.R., Cardoso, J.E., Crisóstomo, J.R., Cavalcanti, J.J.V., Alencar, E.S., 2002. Clonede Cajueiro-Anão Precoce BRS 226 ou Planalto: Nova Alternativa para o Plantiona Região Semi-árida do Nordeste. Comunicado Técnico (EMBRAPA/CNPAT) 78,4–14.
Petrini, O., 1991. Fungal endophytic of tree leaves. In: Andrews, J., Hirano, S.S. (Eds.),Microbial Ecology of Leaves. Spring Verlag, New York, USA, pp. 179–197.
Roux, J., Wingfield, M.J., Byabashaija, M.D., Wingfield, M.J., 2001. Diseases of planta-tion Eucalyptus in Uganda. South African Journal of Science 97, 16–18.
Rubini, M.R., Silva-Ribeiro, R.T., Pomella, A.W., Maki, C.S., Araujo, W.L., dos Santos,D.R., Azevedo, J.L., 2005. Diversity of endophytic fungal community of cacao(Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agentof Witches’ Broom Disease. International Journal of Biological Sciences 1, 24–33.
Sato, T., Iwamoto, Y., Tomioka, K., Taba, S., Ooshiro, A., Takaesu, K., 2008. Black bandof Jew’s marrow caused by Lasiodiplodia theobromae. Journal of General PlantPathology 74, 91–93.
Slippers, B., Wingfield, M.J., 2007. Botryosphaeriaceae as endophytes and latentpathogens of woody plants: diversity, ecology and impact. Fungal BiologyReviews 21, 90–106.
Summerbell, R.C., Krajden, S., Levine, R., Fuksa, M., 2004. Subcutaneous phaeo-hyphomycosis caused by Lasiodiplodia theobromae and successfully treatedsurgically. Medical Mycology 42, 543–547.
Tavares, S.C.C.H., 2002. Epidemiologia e manejo integrado de Botryodiplodiatheobromae—Situacão atual no Brasil e no mundo. Fitopatologia Brasileira 27,46–52.
von Arx, J.A., 1987. Plant Pathogenic Fungi. Beheifte zur Nova Hedwigia. 87, J. Cramer,Berlin.
Woo, P.C., Lau, S.K., Ngan, A.H., Tse, H., Tung, E.T., Yuen, K.Y., 2008. Lasiodiplodiatheobromae pneumonia in a liver transplant recipient. Journal of Clinical Micro-biology 46 (1), 380–384.
Woodward, J.E., Langston, D.B., Brock, J.H., Kemerait, C.J., Brenneman, T.B., Beard,G.H., 2005. First demonstration of Koch’s postulates for Lasiodiplodia theobromaefruit spot on eggplant (Solanum melongena). Plant Disease 89, 687–1687.
Wright, A.F., Harmon, P.F., 2009. First report of Lasiodiplodia theobromae associatedwith stem blight of southern highbush blueberries in Florida. Plant Disease 93(9), 962.


















