
Chapter 8 Energy - University of Tokyocsls-text.c.u-tokyo.ac.jp/pdf/Chap_08.pdfChapter 8 Energy CSLS...
17
Chapter 8 Energy 08 CSLS / THE UNIVERSITY OF TOKYO 149 Part II Principles of Individual Cell Function Chapter 8 Energy This chapter discusses the mechanisms of mitochondrial respiration and photosynthesis by chloroplasts, which are processes involved in the production of biological energy. ATP synthesis by respiration is known as oxidative phosphorylation, and that carried out by photosynthesis is called photophosphorylation. These processes, combined with the substrate-level phosphorylation discussed in the previous chapter, represent almost all the energy production pathways found in cells. This chapter also outlines the production of organic compounds by photosynthesis. Reactions in which organisms use energy to perform activities are diverse, but the production methods for energy and organic compounds are universal and very simple. Energy production systems have been studied as a central pillar of biology since the early 20th century. The energy production system mediated by oxidation-reduction reactions in the biological membrane is particularly well understood from the atomic and molecular levels to the cellular level, and is an active area in which cutting-edge research remains ongoing. I. Biological Energy Biological energy is produced by the breakdown of various organic compounds through fermentation and respiration*1, by respiration using inorganic compounds, and by photosynthesis using light energy. It is controlled by the high- energy phosphate bonds of ATP and other substances and by the concentration gradient of H + for use by organisms in various activities. As outlined in Chapter 7, free energy change (ΔG) resulting from the hydrolysis of ATP’s terminal phosphate bond is expressed by Equation 8-1. Interestingly, ATP has two phosphate bonds, and only the terminal one is formed by the processes of respiration and photosynthesis discussed in this chapter; the inner bond is formed by other enzymes*2. The concentration gradient of H + across the membrane can be expressed as the electrochemical potential of H + (ΔμH+) (Equation 8-2). This is the same as the free energy change that occurs when H + is transported across the membrane. Here, log10[H + in] and log10[H + out] can be converted to the pH *2 ATP’s terminal high-energy phosphate bond is formed by ATP synthase. High-energy phosphate bonds in ADP are formed by adenylate kinase from AMP and ATP: AMP + ATP ⇄ 2ADP *1 Fermentation and respiration: In energy metabolism, fermentation refers to reactions that do not accompany oxidation and reduction, while respiration refers to reactions that accompany oxidation by oxygen or other substances. Sulfate respiration and nitrate respiration, which respectively use SO4 2- and NO3 - instead of oxygen, are also known.
Transcript of Chapter 8 Energy - University of Tokyocsls-text.c.u-tokyo.ac.jp/pdf/Chap_08.pdfChapter 8 Energy CSLS...

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 149
Part II
Principles of Individual Cell Function
Chapter 8Energy
This chapter discusses the mechanisms of mitochondrial respiration and photosynthesis
by chloroplasts, which are processes involved in the production of biological
energy. ATP synthesis by respiration is known as oxidative phosphorylation, and
that carried out by photosynthesis is called photophosphorylation. These processes,
combined with the substrate-level phosphorylation discussed in the previous chapter,
represent almost all the energy production pathways found in cells. This chapter also
outlines the production of organic compounds by photosynthesis. Reactions in which
organisms use energy to perform activities are diverse, but the production methods
for energy and organic compounds are universal and very simple. Energy production
systems have been studied as a central pillar of biology since the early 20th century.
The energy production system mediated by oxidation-reduction reactions in the
biological membrane is particularly well understood from the atomic and molecular
levels to the cellular level, and is an active area in which cutting-edge research
remains ongoing.
I . Biological Energy
Biological energy is produced by the breakdown of various organic compounds
through fermentation and respiration*1, by respiration using inorganic
compounds, and by photosynthesis using light energy. It is controlled by the high-
energy phosphate bonds of ATP and other substances and by the concentration
gradient of H+ for use by organisms in various activities. As outlined in Chapter
7, free energy change (ΔG) resulting from the hydrolysis of ATP’s terminal
phosphate bond is expressed by Equation 8-1. Interestingly, ATP has two
phosphate bonds, and only the terminal one is formed by the processes of
respiration and photosynthesis discussed in this chapter; the inner bond is formed
by other enzymes*2. The concentration gradient of H+ across the membrane can
be expressed as the electrochemical potential of H+ (ΔμH+) (Equation 8-2). This
is the same as the free energy change that occurs when H+ is transported across
the membrane. Here, log10[H+in] and log10[H+
out] can be converted to the pH
*2ATP’s terminal high-energy phosphate bond is formed by ATP synthase. High-energy phosphate bonds in ADP are formed by adenylate kinase from AMP and ATP: AMP + ATP ⇄ 2ADP
*1Fermentation and respiration: In energy metabolism, fermentation refers to reactions that do not accompany oxidation and reduction, while respiration refers to reactions that accompany oxidation by oxygen or other substances. Sulfate respiration and nitrate respiration, which respectively use SO4
2- and NO3
- instead of oxygen, are also known.

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 150
inside and outside the membrane, and ΔΦ is the membrane potential. In other
words, even membrane potential formed by the transport of ions other than H+
contributes to the high-energy state of H+.
Free energy change by ATP hydrolysis:
Equation 8-1
ΔG = ΔG˚’ + 2.3RTlog10[ADP][H3PO4] / [ATP]
Electrochemical potential of H+ across the membrane:
Equation 8-2
ΔμH+ = 2.3RTlog10 [H+in] / [H+
out] + FΔΦ
where R is the gas constant, T is the absolute temperature, ΔG˚’ is the free
energy change under standard conditions (pH7) and F is the Faraday constant.
Organisms have many enzymes that catalyze a range of chemical reactions in
their cells. Cells working in normal conditions constantly incorporate nutrients and
energy from outside and discard waste materials, thereby keeping the intracellular
conditions constant (i.e., in a state of dynamic equilibrium). This state is achieved
through a series of metabolic reactions that proceed almost constantly through the
catalytic action of enzymes. If this is the case, do all chemical reactions catalyzed
by enzymes in living cells reach dynamic equilibrium? The answer is no. If all
such reactions reached this state, new reactions would not occur without the entry
and exit of new materials, although existing conditions might be maintained.
Such cells could no longer be called autonomously living cells.
In performing new reactions, actual organisms change equilibrium by putting
energy into particular reactions. For this purpose, a high-energy state that deviates
from equilibrium needs to be maintained. Such a state involves the concentration
gradient of ATP and H+, and the mechanism of putting in energy as appropriate
involves the activity regulation of enzymes that use the energy (e.g., the ATPase
of myosins that is used for cell movement). The main function of organisms is to
convert the energy produced by the fermentation of various materials, as well as
by respiration and photosynthesis, into the standardized forms, namely ATP and
concentration gradient of H+, and maintain it.
The organic compounds and energy that support the activities of almost all
organisms on earth are produced and supplied through photosynthesis by plants
and other organisms. All oxygen is also produced and released by this process.
The mechanism of photosynthesis is completely different from that of fermentation

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 151
and respiration in that it uses physical energy in the form of light, but the
mechanism of ATP synthesis using the physical energy obtained from the
concentration gradient of H+ created by electron transport is very similar among
the three mechanisms. In processes other than photosynthesis, organic compound
energy is supplied through autotrophic chemical synthesis by bacteria*3.
I I . Outline of the Respiratory Chain and Oxidative Phosphorylation
As outlined in Chapter 7, glucose is fully broken down into carbon dioxide and
hydrogen (NADH and FADH2), and only four ATP molecules per glucose are
synthesized during the process. However, coupling with a series of reactions
through which NADH and FADH2 (generated during the process) are fully
oxidized by oxygen to become water molecules generates 38 molecules of
ATP*4. The series of oxidation-reduction reactions beginning from NADH and
FADH2 are known as electron transport reactions or the respiration chain, and the
mechanism of synthesizing ATP coupling with the reactions is called oxidative
phosphorylation.
In the 1950s, researchers initially looked for metabolic intermediates with high-
energy phosphate bonds on the assumption that, as in glycolysis, kinases
(enzymes that transfer the phosphate group to ATP) were also involved in reactions
that synthesize large amounts of ATP, but these could not be found. Soon
thereafter, the mystery surrounding ATP synthesis was solved by the chemiosmotic
theory proposed in 1961 by P. Mitchell, who was involved in the investigation
of active transport. This is the process that transports H+ and other substances
across the membrane against the concentration gradient using ATP energy. In
Mitchell’s hypothesis, ATP is synthesized using the concentration gradient of
transported H+ coupled with electron transport reactions. In other words, the
concentration gradient of H+ is a high-energy state that is interconvertible with
ATP, and the conversion is catalyzed by F-ATP synthase (discussed later).
I I I . Oxidation-Reduction Reactions and the Respiration Chain
In terms of the utilization of biological energy, the ability to convert energies
*3The sulfur bacteria (Thiomicrospira spp.) found in organisms such as Calyptogena spp. that live in hydrothermal vents on the ocean floor (also known as black smokers) are an example of this.
*4Calculation example: To obtain the standard free energy change of the two-electron reaction NADH + H+ + 1/2 O2 ⇄ NAD+ + H2O, with O2 (E˚’ = +0.815 V) and NAD (E˚’ = -0.315 V), n = 2 is assigned to Equation 8-3: ΔG˚’ = -2 x 96.5 x [0.815 – (-0.315)] = -218 kJ mol-1
where the Faraday constant is 96.5 kJV -1 mol-1

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 152
generated by various oxidation-reduction reactions to a single form, H+
electrochemical potential, is a significant advantage. Oxidation-reduction
reactions involve the transfer of electrons between two materials, and are
mediated mainly by cofactors (i.e., coenzymes). Since the same cofactors (e.g.,
NAD+) tend to be used in different enzymatic oxidation-reduction reactions in
cells, energy can be efficiently produced through a common mechanism by
converging enzymatic oxidation-reduction reactions into the respiration chain
through the mediation of cofactors. Among the common cofactors, NADH and
FADH2 are often utilized, and quinones (such as ubiquinone) and cytochrome c
are also used (Fig. 8-1). Most of these cofactors are common to all organisms,
and photosynthesis (discussed in V in this chapter) also uses very similar cofactors.
The free energy change of oxidation-reduction reactions has a linear relationship
with the difference in oxidation-reduction potential between two reactants, A
and B (Equation 8-3).
Equation 8-3
ΔG˚’ = -nF(E˚’A – E˚’B)
where n is the number of electrons transferred and F is the Faraday constant.
The hydrogen (E˚’ = -0.315 V) in NADH releases 218 kJmol-1 of energy by
reacting directly with oxygen (E˚’ = +0.815 V)*4, but the respiration chain is
separated into electrons and H+ . These electrons, which have a high reducing
ability (i.e., low oxidation-reduction potential), gradually release energy via
around 20 types of electron carrier and finally react with oxygen in mild
conditions. Among these electron carriers, ubiquinone (a low molecular weight
compound) (Fig. 8-1) and cytochrome c (a small protein) act as mobile electron
carriers, and the rest are contained in four types of protein complex (Complexes
I – IV) as cofactors. These cofactors, which strongly bind to proteins, are known
as the prosthetic group. In terms of substrate oxidation and reduction, these
protein complexes are also called NADH dehydrogenase (Complex I), succinate
dehydrogenase (Complex II), cytochrome bc1 complex (Complex III) (also known
as ubiquinone-cytochrome c oxidoreductase) and cytochrome c oxidase (Complex
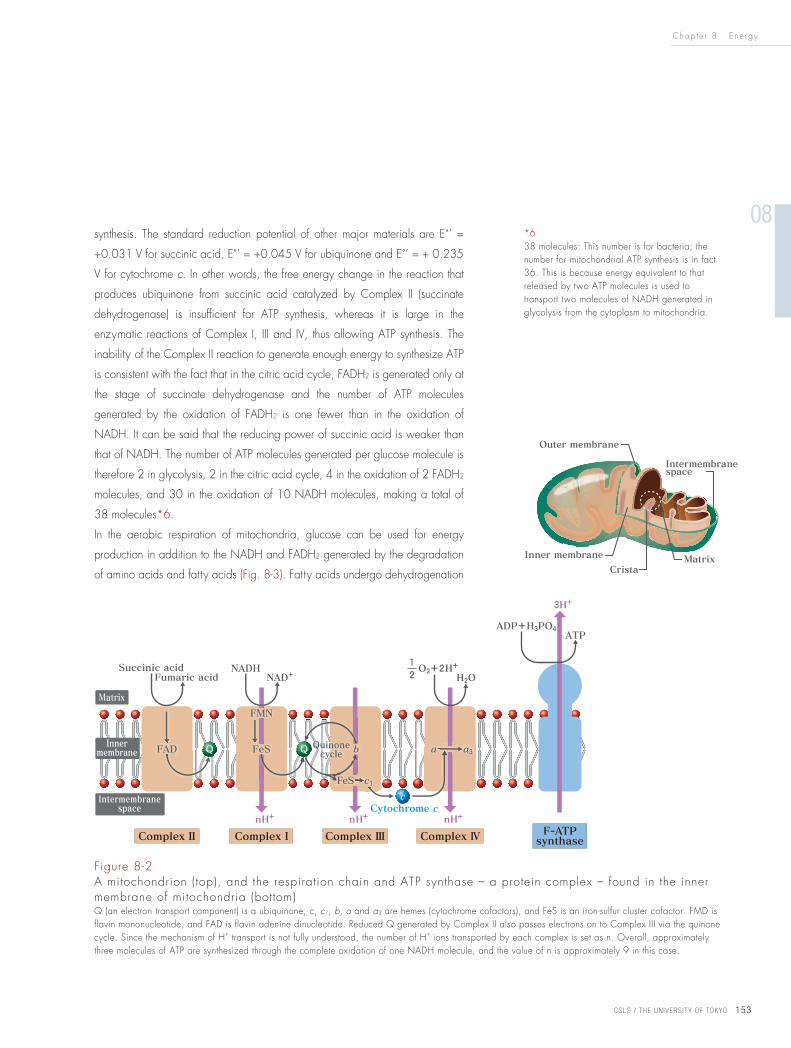
IV) (Fig. 8-2*5).
The free energy change during the complete oxidation of NADH (ΔG o’ = -218
kJmol-1) is equivalent to that of around seven ATP molecules (ΔG o’ = -30.5 kJmol-1
per molecule); however, only three ATP molecules are actually generated, and
the remaining free energy change is used to tilt the equilibrium toward ATP
*5NADH is a water-soluble coenzyme that is shared by many enzymes. FADH2, on the other hand, binds to certain enzymes, and is a coenzyme (or cofactor) that facilitates the transfer of electrons in enzymatic reactions. Therefore, in Figure 8-2, NADH is in the matrix, and FADH2 (FAD in the figure – the oxidized form that receives electrons) binds to the inside of Complex II.
Figure 8-1Oxidized and reduced forms of ubiquinoneAlso known as coenzyme Q, this is a lipophilic compound that can transfer electrons and H+ separately, and is involved in the creation of the H+ concentration gradient.
Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 153
synthesis. The standard reduction potential of other major materials are E˚’ =
+0.031 V for succinic acid, E˚’ = +0.045 V for ubiquinone and E˚’ = + 0.235
V for cytochrome c. In other words, the free energy change in the reaction that
produces ubiquinone from succinic acid catalyzed by Complex II (succinate
dehydrogenase) is insufficient for ATP synthesis, whereas it is large in the
enzymatic reactions of Complex I, III and IV, thus allowing ATP synthesis. The
inability of the Complex II reaction to generate enough energy to synthesize ATP
is consistent with the fact that in the citric acid cycle, FADH2 is generated only at
the stage of succinate dehydrogenase and the number of ATP molecules
generated by the oxidation of FADH2 is one fewer than in the oxidation of
NADH. It can be said that the reducing power of succinic acid is weaker than
that of NADH. The number of ATP molecules generated per glucose molecule is
therefore 2 in glycolysis, 2 in the citric acid cycle, 4 in the oxidation of 2 FADH2
molecules, and 30 in the oxidation of 10 NADH molecules, making a total of
38 molecules*6.
In the aerobic respiration of mitochondria, glucose can be used for energy
production in addition to the NADH and FADH2 generated by the degradation
of amino acids and fatty acids (Fig. 8-3). Fatty acids undergo dehydrogenation
*638 molecules: This number is for bacteria; the number for mitochondrial ATP synthesis is in fact 36. This is because energy equivalent to that released by two ATP molecules is used to transport two molecules of NADH generated in glycolysis from the cytoplasm to mitochondria.
Figure 8-2A mitochondrion (top), and the respiration chain and ATP synthase – a protein complex – found in the inner membrane of mitochondria (bottom)Q (an electron transport component) is a ubiquinone, c, c1, b, a and a3 are hemes (cytochrome cofactors), and FeS is an iron-sulfur cluster cofactor. FMD is flavin mononucleotide, and FAD is flavin adenine dinucleotide. Reduced Q generated by Complex II also passes electrons on to Complex III via the quinone cycle. Since the mechanism of H+ transport is not fully understood, the number of H+ ions transported by each complex is set as n. Overall, approximately three molecules of ATP are synthesized through the complete oxidation of one NADH molecule, and the value of n is approximately 9 in this case.

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 154
by two-carbon-atom units (β-oxidation) and enter the citric acid cycle as acetyl-
CoA, where they are completely oxidized. Amino groups that constitute amino
acids are converted to urea, and the remaining carbon skeletons are completely
oxidized in glycolysis and the citric acid cycle for use in ATP production. The
mitochondrial respiration chain is thus a key component of energy production.
The main role of the respiration chain is to efficiently transport H+ across the inner
membrane to the outside by coupling with the stepwise oxidation and reduction
of electron carriers, and the details of the mechanism, including the quinone cycle
proposed by Mitchell, have been gradually revealed. Quinones, such as
ubiquinone, are lipophilic materials that transfer H+ through oxidation and
reduction (Fig. 8-1), and are therefore widely used by various organisms in
reactions that create high-energy states by transporting H+ across the membrane.
The quinone cycle is a system that transports two H+ ions by the transfer of one
electron when electrons are transferred from a ubiquinone to a cytochrome bc1
complex. Such complexes have separate sites for quinone oxidation and
reduction, and a reduced ubiquinone bound to an oxidation site releases two H+
ions and two electrons. Of these, one electron is transferred to cytochrome c1 via
iron-sulfur clusters, and the other reduces another ubiquinone at the quinone
reduction site via cytochrome b. Both H+ ions are subsequently released to the
outside of the inner membrane. With the progress of spectroscopy and structural
biology in recent years, the structure of these protein complexes and the behavior
of each electron carrier have gradually been clarified, and Japan leads the world
in the field of research into cytochrome c oxidase.
Figure 8-3Mitochondrial respiration chainGlycolysis takes place in the cytoplasm.

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 155
IV. ATP Synthase
F-ATP synthase synthesizes ATP by coupling with the H+ transport that follows the
concentration gradient of H+ (Fig. 8-4). This enzyme consists of F0 (the membrane
intrinsic site) and F1 (the membrane superficial site). F0 consists of a rotor and a
fixed part, between which channel-like paths for H+ transport are found. F1
involves ATP synthetic and degradative activities, and is also known as F-ATPase.
If F1 is removed from the membrane, electron transport does not lead to ATP
synthesis. For this reason, F1 was initially known as the coupling factor for electron
transport and ATP synthesis. It was subsequently revealed that an enzyme of the
same type as mitochondrial ATP synthase exists in chloroplasts and the cell
membrane of eubacteria, and that it is widely involved in ATP synthesis; this
enzyme is called the F-type (F stands for “factor”).
The important point in the coupling of H+ transport and ATP synthesis reactions is
that, coupling with H+ transport, the rotor and the stalk rotate in a clockwise
direction, thus supplying energy to F1. F1 does not rotate, and synthesizes ATP
using the energy generated by the rotation of the stalk. The phenomenon of ATP
synthase rotating like a motor as a reverse reaction has been demonstrated in an
elegantly designed experiment (see the Column on p.157). The details regarding
the coupling of H+ transport and ATP synthesis are not covered here, but the
stoichiometry is 3H+/ATP.
Figure 8-4F-ATP synthaseThis consists of F1 and F0, with the F0 further divided into rotor and stator parts.

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 156
V. Outline of Photosynthesis
Photosynthesis consists of reactions in which light energy is absorbed and
converted to chemical energy to produce ATP and reducing power (light
reactions), and reactions in which carbon dioxide is fixed as organic compounds
using the ATP and reducing power (dark reactions). Photosynthesis proceeds in
chloroplasts (organelles unique to plant cells), light reactions occur locally in the
thylakoid membrane, and dark reactions occur locally in the stroma (an aqueous
space in chloroplasts). In light reactions, light energy is absorbed and transported
by antenna pigment*7, and electrons are released in the photochemical reaction
center*8, thus driving electron transport. The electron transport reaction of
photosynthesis is different from the respiration chain in that NADPH as well as
ATP is synthesized through H+ transport. The reducing power necessary in the
series of reactions is obtained from the oxidation of water molecules, and oxygen
that is no longer needed is disposed of. As an example, the atmosphere did not
contain any oxygen when the earth was created; all the oxygen that currently
exists in the air is derived from photosynthesis. In dark reactions, the carbon
dioxide fixation reaction and the Calvin cycle (or saccharometabolic cycle) are
driven by ATP and NADPH. A carbon dioxide fixation similar to that in dark
reactions of photosynthesis also takes place in chemoautotrophic bacteria.
VI. Absorption of Light Energy
Antenna pigments that absorb light energy include chlorophyll a, chlorophyll b
and carotenoid*9. Blue and red pigments, such as phycobilin, are also known
in red algae and cyanobacteria. When visible light excites these pigments,
nearly 100% of the excitation energy moves between the pigments and is
conveyed to the photosynthetic reaction center. For this purpose, the antenna
pigments need to be located close to each other, and are incorporated into
proteins and accumulated within the thylakoid membrane. Although plant leaves
may appear evenly green, only the thylakoid membrane of chloroplasts, which
contain chlorophyll a and b, is in fact green. Other organelles, such as those
found in animal cells, have no color. While most terrestrial plants have green
leaves, pigments found in algae vary in color (e.g., phycocyanin is blue,
phycoerythrin is red and fucoxanthin is brown) because the spectrum of sunlight
deviates in water depending on the conditions of absorption and scattering. On
*7Antenna pigment: A pigment that, following the absorption of light energy, does not perform photosynthesis and conveys energy to the reaction center.
*8Photosynthetic reaction center: The core site containing the chlorophyll pigment that performs photosynthetic reactions on being excited by light energy.
*9Carotenoid: A fat-soluble material with a long-chain polyene. The material functions as an antenna pigment, and also known as a vitamin A precursor.

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 157
the other hand, anthocyan – a water-soluble colorful pigment found in plant
tissues such as flowers – is located in vacuoles, and cannot use absorbed light
energy for photosynthesis.
A Japanese research group demonstrated the rotation of ATP synthase
through an elegantly designed experiment. In the setup, F1 of ATP synthase
was fixed to a glass slide, and an actin filament treated with a fluorescent
dye was bound to the stalk, thus making one complex visible under a
fluorescence microscope. When ATP was applied to the glass, the complex
started rotating as ATP hydrolysis proceeded, and this action was video-
observed. Interestingly, the rotation took place in an anticlockwise direction
in three 120-degree steps, indicating that ATP is degraded via three stable
intermediates and the energy is transferred to the stalk. It can be considered
that ATP synthesis proceeds in the opposite direction. It is believed that,
through H+ transport, the energy of H+ causes ATP synthesis through the
rotation of the stalk. Crystal structure analysis of F1 had already shown that
an asymmetric stalk-like γ-subunit is located on the bottom surface of the
threefold symmetrical sphere containing three α-subunits with ATP synthetic
activity, and that these three subunits are in different states. This indicated
that the structural change of the α-subunits, which are involved in ATP
synthesis, is linked to the positional relationship with the stalk, thus rotating
the complex in three 120-degree steps.
Column Demonstration of ATP Synthase Rotation
Column Figure 8-1Schematic diagram of the video-recorded rotationA modified version of the original diagram created by Toru Hisabori, Associate Professor at Tokyo Institute of Technology.

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 158
VII. Photochemical Reaction and Electron Transport
In the photosynthetic reaction center, a photochemical reaction occurs in which
certain chlorophyll a receives excitation energy and releases electrons (similar to
the process seen in photoelectric reactions), thus generating chlorophyll a+. This
drives electron transport, creating a concentration gradient of H+ (as seen in
mitochondrial electron transport reactions) based on which ATP is synthesized
and NADP+ is reduced to produce NADPH. The pigments and electron carriers
involved in these reactions are buried in the thylakoid membrane as three types
of protein complex, and the mobile electron carriers that connect these complexes
are plastoquinone and plastocyanin (Fig. 8-5).
Although not described in detail here, these two photochemical complexes are
akin to photoelectric semiconductors, and are designed so that electron transport
steadily proceeds without the recombination of electrons and positive charges
separated in photochemical reactions. The cytochrome b6f complex, on the other
hand, transports H+, and has a structure and functionality similar to those of the
mitochondrial cytochrome bc1 complex. The basic skeleton of plastoquinone is
the same as that of mitochondrial ubiquinone, and the cytochrome b6f complex
Figure 8-5Light reaction system of photosynthesisQ is plastoquinone, and b6 and f are hemes (i.e., cytochrome cofactors). The cytochrome b6f complex and the quinone cycle have the same basic structure as mitochondrial complex III.

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 159
also transports H+ using the similar quinone cycle. Plastocyanin is a small, copper-
binding protein, and is one of the major reasons why plants require copper in
particular. When copper is deficient, some algae instead use c - type cytochrome,
which bears a close resemblance to mitochondrial cytochrome c. In this way,
respiration and photosynthesis use very similar electron transport mechanisms,
and the systems are therefore believed to have evolved from the same energy
metabolic system.
When photosynthetic electron transport is considered in terms of oxidation-
reduction potential, the chlorophyll a+ (E˚’ = +1.1 – 1.2 V) generated in
photosystem II deprives electrons from stable water molecules using its high
oxidizing power, thereby emitting oxygen. On the other hand, the highly
reducing electrons (E˚’ = -1.4 V) produced in photosystem I reduce NADP+ to
produce NADPH. In this way, a series of electron transport from water molecules
to NADPH is driven by light energy through the tandem connection of the two
photosystems. The reason for these systems being connected in tandem is that the
two reactions cannot take place simultaneously with the energy of visible light.
Indeed, photosynthetic bacteria which have only one photosystem cannot break
down water molecules. Besides the electron transport pathway in which the two
photosystems are connected in tandem, a cyclic electron transport pathway also
exists, in which electrons flow from photosystem I to near the cytochrome b6f
complex. This pathway transports only H+ for ATP synthesis.
One of these two pathways is used in accordance with the environment and the
needs at hand. Also in chloroplasts, F-ATP synthase located in the thylakoid
membrane works in the coupling of H+ transport and ATP synthesis. This enzyme
is essentially the same as those found in mitochondria and eubacteria (Fig. 8-4).
Equation 8-4
Photosynthetic light reaction can therefore be summarized as follows:
2H2O + 2NADP+ + Light energy → O2 + 2NADPH + 2H+ (+ nATP)
The number of ATP molecules synthesized here is n, because it changes depending
on the combination of the two electron transport pathways mentioned above.
The value of n is 3 (NADPH:ATP = 2:3) in the general photosynthetic dark
reactions discussed later (Equation 8-5), and is 4 or 5 in C4 photosynthesis
(discussed in IX), which requires more ATP molecules.
Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 160
Reactions regulated by light (→) proceed in one direction. All
reactions other than these and the carbon dioxide fixation
reactions catalyzed by RuBisCO are reversible (↔).
VIII. Dark Reaction: Carbon Dioxide Fixation
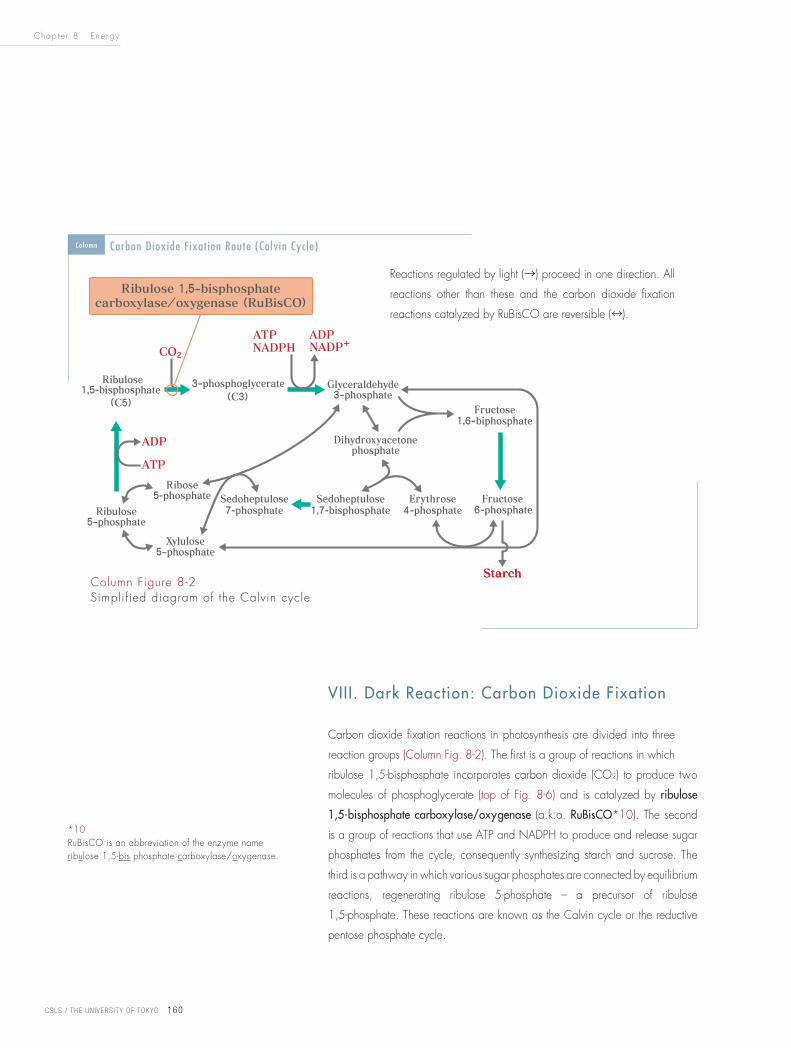
Carbon dioxide fixation reactions in photosynthesis are divided into three
reaction groups (Column Fig. 8-2). The first is a group of reactions in which
ribulose 1,5-bisphosphate incorporates carbon dioxide (CO2) to produce two
molecules of phosphoglycerate (top of Fig. 8-6) and is catalyzed by ribulose
1,5-bisphosphate carboxylase/oxygenase (a.k.a. RuBisCO*10). The second
is a group of reactions that use ATP and NADPH to produce and release sugar
phosphates from the cycle, consequently synthesizing starch and sucrose. The
third is a pathway in which various sugar phosphates are connected by equilibrium
reactions, regenerating ribulose 5-phosphate – a precursor of ribulose
1,5-phosphate. These reactions are known as the Calvin cycle or the reductive
pentose phosphate cycle.
*10RuBisCO is an abbreviation of the enzyme name ribulose 1,5-bis phosphate carboxylase/oxygenase.
Column Figure 8-2Simplified diagram of the Calvin cycle
Column Carbon Dioxide Fixation Route (Calvin Cycle)

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 161
Since ATP synthesis reactions involving kinase (phosphotransferase) are coupled with
metabolic reactions at a ratio of 1:1, the ATP synthesis efficiency is 100%. On the
other hand, ATP synthesis reactions involving F-ATP synthase are indirectly coupled
with electron transport via H+ transport, making the story a lit t le more complex. As
an example, the appearance of chloroplasts differs between daytime and nighttime.
During the day, when light is available, chloroplasts are in a state of high energy
with the thylakoid membrane maintaining an H+ gradient by electron transport, and
are ready to supply ATP when it is used. However, at nighttime, it is difficult to
maintain this high-energy state. Although the thylakoid membrane has low
permeability for H+ and is therefore suited to maintaining the concentration gradient,
it cannot stop H+ from gradually leaking through the membrane during the night.
Generally, enzymatic reactions are reversible, and when the concentration gradient
of H+ is decreased, the gradient is restored by breaking down ATP, which is a waste
of energy. In the F-ATP synthase of chloroplasts, therefore, an activity regulation
mechanism involving thioredoxin (an oxidation-reduction protein) has evolved. When
thioredoxin with cyctein residue is reduced by photosynthetic electron transport, the
cyctein residue of a protein in the stalk of ATP synthase is reduced, thereby activating
the enzyme; in dark places, on the other hand, the activity of the enzyme is inhibited
through the oxidation of the cyctein residue, thereby reducing the wasteful use of
energy. The sum of dark reactions after removing the substances regenerated in the
reactions outlined above is expressed as follows:
Equation 8-5
CO2 + 3ATP + 2NADPH + 2H+ →
(CH2O) + H2O + 2NADP+ + 3ADP + 2H3PO4
Figure 8-6 Two reactions catalyzed by RuBisCOThe carbon dioxide fixation reaction (top) and the reaction with oxygen (bottom). It should be noted that two molecules of phosphoglycerate are generated in the above reaction.
Column Regulation by Coupling and Light

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 162
(CH2O) corresponds to a sugar. This equation indicates that three ATP molecules
and two NADPH molecules are needed to fix one molecule of CO2.
A distinct characteristic of photosynthetic carbon dioxide fixation reactions is the
unique nature of RuBisCO. As its full name suggests, RuBisCO involves
carboxylase and oxygenase activity; when the oxygen concentration is high
and the carbon dioxide concentration is low, it incorporates O2 instead of CO2
and wastefully uses ribulose 1,5-bisphosphate (bottom of Fig. 8-6). The reactivity
of RuBisCO to CO2 is more than 100 times higher than its reactivity to O2;
however, since the oxygen concentration in the atmosphere is 500 times higher
than that of CO2, the oxygenase activity in chloroplasts cannot be ignored. A
byproduct generated in the oxygenase reaction – phosphoglycolate – is
converted back to phosphoglycerate using ATP and NADPH; this pathway is
called photorespiration, since CO2 is released during the process. Additionally,
since the rate of reactions catalyzed by RuBisCO is much slower than that of
other metabolic enzymes, the amount of RuBisCO required is several hundred
times more than that of other enzymes that catalyze the reactions seen before
and after those catalyzed by RuBisCO. As a result, RuBisCO proteins account for
over half the water-soluble proteins found in chloroplasts, making them the most
abundant proteins on earth. These intriguing characteristics of RuBisCO reflect the
high CO2 concentration and absence of O2 in the atmosphere of primitive earth
when phototrophs first appeared.
IX. C4 Photosynthesis
For many plants, the initial carbon dioxide fixation product in the Calvin cycle is
phosphoglycerate (with three carbon atoms), whereas in other plants, carbon
dioxide is fixed as malic acid or aspartic acid (with four carbon atoms). Those in
the former group, which includes rice, spinach and trees, are called C3 plants,
and those in the latter, which includes corn, are known as C4 plants.
In C4 plants, photosynthesis occurs in a roundabout way in which
phosphoenolpyruvate carboxylase with a bicarbonate ion (HCO3-) – against
which oxygen does not compete – as a substrate fixes carbon dioxide, other
enzymes are then released in the cell, and finally RuBisCO refixes carbon
dioxide. Although this C4 photosynthetic reaction uses extra energy (ATP),
oxygenase activity is not induced under conditions of low extracellular carbon
dioxide concentration, thus allowing efficient photosynthesis. These C4 plants

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 163
have thrived over tens of millions of years, reducing the carbon dioxide
concentration in the atmosphere from higher levels to the current 0.037%.
Practical application of C4 photosynthesis has been investigated with the aim of
increasing the productivity of crops such as rice.
When the first organisms appeared on earth 3.8 billion years ago, it is
believed that the carbon dioxide concentration as a ratio of atmospheric
pressure was much higher than today. However, this level was greatly
reduced by the emergence of phototrophs to just a few percent in the
Paleozoic era, and the growth of terrestrial plants along with the emergence
of C4 plants further reduced it to today’s levels. Additionally, significant
short-term changes have occurred in connection with human activity and
climate change. As an example, the carbon dioxide level, which stood at
0.027% in the 17th century, rose sharply after the Industrial Revolution to
the current figure of 0.037%.
X. Topology of Mitochondria and Chloroplasts
As outlined in Chapter 5, mitochondria and chloroplasts originate from certain
eubacteria*11 that lived symbiotically in eukaryotic cells (endosymbiotic theory:
see the Column on p. 104 in Chapter 5). Unlike other organelles, mitochondria
and chloroplasts are therefore surrounded by double membranes – one inner and
one outer *12. The inner membrane of mitochondria forms cristae, through which
H+ is transported outside the membrane by electron transport in the respiration
chain. In chloroplasts, on the other hand, the photosynthetic system (including
chlorophyll) does not exist in the inner envelope. Rather, it is found in the thylakoid
membrane further inside, and transports H+ into this membrane by photosynthetic
electron transport. The directions of this transport seem opposite to each other;
however, since the matrix of mitochondria and the stroma of chloroplasts are
homologous compartments, the two directions are in fact the same when it is
Column Photosynthesis and Changes in Carbon Dioxide Concentration in the Earth’s Atmosphere
*11 Eubacteria: A type of cyanobacteria (a blue-green algae), which are ancestors of chloroplasts. The major candidate ancestors of mitochondria are rickettsiae (intracellular parasitic pathogens and a type of α-proteobacterium), but due to the marked specialization of mitochondria, their ancestral relationship has not been determined.
*12 Inner and outer membranes: In chloroplasts, these are also referred to as the inner and outer envelopes. In both chloroplasts and mitochondria, the outer membrane has non-specific carriers and therefore does not block material transport. Endosymbiotic theory suggests that the outer membrane is derived from a host cell, but another well-accepted theory is that the outer membrane is homologous to a special outer membrane of cyanobacteria, which are ancestors of chloroplasts.

Chap t e r 8 E n e r g y
CSLS / THE UN IVERS ITY OF TOKYO 164
considered that H+ is emitted from them (i.e., they are topologically homologous).
The matrix of mitochondria (in which citric-acid-cycle enzymes work) and the
stroma of chloroplasts (in which Calvin-cycle enzymes work) are homologous, and
H+ movement in electron transport is directed outward from these compartments.
Coupling with H+ transport based on this H+ concentration gradient, F-ATP synthase
therefore synthesizes ATP in the same compartment as the citric acid cycle and the
Calvin cycle.
• The production patterns of biological energy are divided into substrate-
level phosphorylation (see Chapter 7), oxidative phosphorylation
(respiration) and photophosphorylation (photosynthesis) as discussed in
this chapter.
• In respiration and photosynthesis, through the convergence of a series of
oxidation-reduction reactions into a common electron transport chain, all
energy is replaced with the concentration gradient of H+ (electrochemical
potential), and common F-ATP synthase synthesizes ATP using the
electrochemical potential of H+.
• The respiration chain, the electron transport chain in photosynthesis and
ATP synthase act as protein complexes incorporated into the biological
membrane. These complexes react differently from many enzymes in
aqueous solution, and have been studied in detail as typical examples of
life’s complex systems.
• Energy changes in various reactions (free energy change (ΔG˚’), changes
in oxidation-reduction potential (ΔE˚’), changes in the electrochemical
potential of H+ (ΔμH+) and the formation and dissociation of ATP high-
energy phosphate bonds) can be understood uniformly as being
interconvertible.
• In respiration and photosynthesis, a mechanism exists to maintain a state
of high energy by regulating ATP production and supply in response to
ATP utilization and environmental changes. This mechanism is known as
homeostasis.
• Biological systems can function autonomously because a high-energy state
is maintained within cells.
Summary Chapter 8

Chap t e r 8 E n e r g y
08
CSLS / THE UN IVERS ITY OF TOKYO 165
[1]
ATP is known as “the currency of biological energy.” Explain
the reasons for this in connection with the overall processes
of intracellular metabolism.
[2]
In eukaryotic cells, H+ released by glycolysis and the citric
acid cycle produces energy via oxidative phosphorylation
in the electron transport chain. Briefly explain the mechanism
of ATP synthesis in mitochondria using the terms below (and
drawings if desired).
Terms: inner membrane, intermembrane space, matrix, F-ATP
synthase
[3]
The oxidation of sugar molecules in cells follows the equation
below:
C6H12O6 (glucose) + 6O2 → 6CO2 + 6H2O + energy
Indicate whether the following statements are correct, and
provide reason for your answers.
A) Energy is produced by the oxidation of carbon atoms.
B) The water required by cells is largely supplied by this
reaction.
C) In cells, this reaction is a several-step process.
D) Reaction with oxygen occurs in many of the steps of sugar
molecule oxidation.
E) In some organisms, the reverse reaction occurs.
F) Some cells produce CO2 while growing in an O2-free
environment.
[4]
Explain how material transport in the mitochondrial inner
membrane occurs, and how this mechanism contributes to
mitochondrial functions.
[5]
In chloroplasts, the pH of the external solution is increased
by photosynthetic electron transport, whereas in mitochondria
it is reduced by electron transport. However, in both cases,
ATP synthase is arranged facing the stroma or matrix. Explain
the topological differences regarding this H+ movement.
[6]
The energy stored in the mitochondrial inner membrane (H+
motive force, pmf) is expressed by the equation below:
ΔG = 2.3 RT [pH (inside) – pH (outside)] + ZFΔψ
where R is the gas constant, T is the absolute temperature, Z
is the electric charge (e.g., H+ = 1), F is the Faraday constant
and ψ is the membrane potential.
1) Calculate the value of ΔG when the membrane potential
is 0.168 V (inside - negative), the pH difference between
the inside and outside of the membrane is 0.75 and the
temperature is 37 ˚C.
2) The free energy change required to synthesize ATP from
ADP in vivo is slightly larger than that required in standard
conditions. Assuming that the value of this change is 45
kJmol-1, give the minimum number of H+ ions that need to
be transported to synthesize one ATP molecule.
Problems
(Answers on p.256)


![NEW BOUNDS FOR SOLUTIONS OF SECOND ORDER ......NEW BOUNDS FOR SOLUTIONS OF SECOND ORDER ELLIPTIC 555 (3.3) Δφ + λψ = 0 in D φ = 0 on B. It is well known [1] that since u = 0 on](https://static.fdocuments.us/doc/165x107/5fdac98d8272d344e53bfbcb/new-bounds-for-solutions-of-second-order-new-bounds-for-solutions-of-second.jpg)













