
CHAPTER 10 Lipids - Minificciones · PDF file142 CHAPTER 10 Lipids 10.1 Fatty Acids Are a Main...
12
142 CHAPTER 10 Lipids 10.1 Fatty Acids Are a Main Source of Fuel 10.2 Triacylglycerols Are the Storage Form of Fatty Acids 10.3 There Are Three Common Types of Membrane Lipids L ipids are defined as water-insoluble molecules that are highly soluble in organic solvents. This important class of molecules has a variety of biochemical roles. For instance, lipids are widely used to store energy. They are also key components of membranes and play a variety of roles in signal-transduction pathways. Unlike the three other classes of biomolecules (carbohydrates, amino acids, and nucleic acids), lipids do not form polymers. Their individual and collective properties— as noncovalent assemblies—make them extraordinarily important. We will examine five classes of lipids here. 1. Free Fatty Acids (Nonesterified Fatty Acids). This simplest type of lipid is most commonly used as a fuel. 2. Triacylglycerols. This class of lipid is the storage form of fatty acids. 3. Phospholipids. These membrane lipids consist of fatty acids attached to a scaf- fold that also bears a charged phosphoryl group, creating a macromolecule with a polar head and nonpolar tail. 4. Glycolipids. These lipids are bound to carbohydrates and are important mem- brane constituents. 5. Steroids. These lipids differ from the other lipids in that they are polycyclic hydrocarbons. Steroids function as hormones that control a variety of physio- logical functions. The most common steroid is cholesterol, another vital mem- brane component. Olive trees. The fruit of a small tree native to the eastern areas of the Mediterranean, olives are unusual in that they are an inedible fruit unless processed. Olive oil, which has been used in cooking since antiquity, is rich in monounsaturated fatty acids as well as antioxidants. These antioxidants may be one of the reasons for the positive health effects of the Mediterranean diet. [David Pick/Alamy.]
Transcript of CHAPTER 10 Lipids - Minificciones · PDF file142 CHAPTER 10 Lipids 10.1 Fatty Acids Are a Main...

142
CHAPTER
10 Lipids10.1 Fatty Acids Are a Main Source
of Fuel
10.2 Triacylglycerols Are the StorageForm of Fatty Acids
10.3 There Are Three Common Types of Membrane Lipids
L ipids are defined as water-insoluble molecules that are highly soluble in organicsolvents. This important class of molecules has a variety of biochemical roles.
For instance, lipids are widely used to store energy. They are also key componentsof membranes and play a variety of roles in signal-transduction pathways. Unlikethe three other classes of biomolecules (carbohydrates, amino acids, and nucleicacids), lipids do not form polymers. Their individual and collective properties—as noncovalent assemblies—make them extraordinarily important. We willexamine five classes of lipids here.
1. Free Fatty Acids (Nonesterified Fatty Acids). This simplest type of lipid is mostcommonly used as a fuel.
2. Triacylglycerols. This class of lipid is the storage form of fatty acids.
3. Phospholipids. These membrane lipids consist of fatty acids attached to a scaf-fold that also bears a charged phosphoryl group, creating a macromoleculewith a polar head and nonpolar tail.
4. Glycolipids. These lipids are bound to carbohydrates and are important mem-brane constituents.
5. Steroids. These lipids differ from the other lipids in that they are polycyclichydrocarbons. Steroids function as hormones that control a variety of physio-logical functions. The most common steroid is cholesterol, another vital mem-brane component.
Olive trees. The fruit of a small tree native to the eastern areas of the Mediterranean, olivesare unusual in that they are an inedible fruit unless processed. Olive oil, which has beenused in cooking since antiquity, is rich in monounsaturated fatty acids as well asantioxidants. These antioxidants may be one of the reasons for the positive health effects ofthe Mediterranean diet. [David Pick/Alamy.]

14310.1 Fatty Acids
10.1 Fatty Acids Are a Main Source of FuelFats, or fatty acids, are chains of hydrogen-bearing carbon atoms,called hydrocarbons,that terminate with carboxylic acid groups. These hydrocarbon chains are ofvarious lengths and may have one or more double bonds, depending on the fat.Two key roles for fatty acids are as fuels and as building blocks for membranelipids. Fats are good fuels because they are more reduced than carbohydrates;that is, the carbon atoms are bonded to hydrogen atoms and other carbonatoms rather than to oxygen atoms, as is the case for carbohydrates. Because ofthis greater reduction, fats yield more energy than carbohydrates when under-going combustion to carbon dioxide and water. We will return to this discussionin Chapter 14.
Fats have common names and systematic names. Although we will use thecommon names for fatty acids, familiarity with the systematic names is importantbecause their use is sometimes required to prevent confusion. A fatty acid’s sys-tematic name is derived from the name of its parent hydrocarbon by the substitu-tion of oic for the final e. For example, the C18 saturated fatty acid known familiarlyas stearic acid, is called octadecanoic acid because the parent hydrocarbon is octade-cane: octadec- for the 18 carbon atoms, and –ane because it is composed completelyof single bonds. Fatty acids composed of single bonds only are called saturated fattyacids, because every carbon atom is attached to four other atoms. Fatty acids withone or more double or triple bonds are called unsaturated fatty acids. Oleic acid, aC18 fatty acid with one double bond, is called octadecenoic acid; whereas linoleicacid, with two double bonds, is octadecadienoic acid; and linolenic acid, with threedouble bonds, is octadecatrienoic acid. The composition and structure of a fattyacid can also be designated by numbers. The notation 18:0 denotes a C18 fatty acidwith no double bonds, whereas 18:2 signifies that there are two double bonds. Thestructure of the ionized forms of two common fatty acids—palmitic acid (16:0)and oleic acid (18:1)—are shown in Figure 10.1.
When considering fatty acids, we often need to distinguish the individual car-bon atoms. Fatty acid carbon atoms are numbered starting at the carboxyl termi-nus, as shown in the margin. Carbon atoms 2 and 3 are often referred to as � and�, respectively. The last carbon atom in the chain, which is almost always a methylcarbon atom, is called the v-carbon atom (� is the Greek symbol for omega, thelast letter of the Greek alphabet). The position of a double bond is represented bythe symbol � followed by a superscript number. For example, cis-�9 means thatthere is a cis double bond between carbon atoms 9 and 10; trans-�2 means thatthere is a trans double bond between carbon atoms 2 and 3. Just as in proteins, cisand trans designate the relative positions of substituents on either side of the dou-ble bond. Just like amino acids, fatty acids are ionized at physiological pH, and so
C
OPalmitate
(ionized form of palmitic acid)
Oleate(ionized form of oleic acid)
O
C
O
O
–
–
Figure 10.1 Structures of two fatty acids.Palmitate is a 16-carbon, saturated fattyacid, and oleate is an 18-carbon fatty acidwith a single cis double bond.
CH2
CH2C
CH2
O
OH3C
–
n
12
3
α
β
ω
H
H
CH2
H3CCω-Carbon
atomω-Carbon
atom
ω-3double bond
C
C
CH2
COO–
(CH2)n
An �-3 fatty acid

Table 10.1 Some naturally occurring fatty acids in animals
Number of Number of carbon atoms double bonds Common name Systematic name Formula
12 0 Laurate n-Dodecanoate CH3(CH2)10COO�
14 0 Myristate n-Tetradecanoate CH3(CH2)12COO�
16 0 Palmitate n-Hexadecanoate CH3(CH2)14COO�
18 0 Stearate n-Octadecanoate CH3(CH2)16COO�
20 0 Arachidate n-Eicosanoate CH3(CH2)18COO�
22 0 Behenate n-Docosanoate CH3(CH2)20COO�
24 0 Lignocerate n-Tetracosanoate CH3(CH2)22COO�
16 1 Palmitoleate cis-�9-Hexadecenoate CH3(CH2)5CH CH(CH2)7COO�
18 1 Oleate cis-�9-Octadecenoate CH3(CH2)7CH CH(CH2)7COO�
18 2 Linoleate cis, cis-�9, �12- CH3(CH2)4(CH CHCH2)2(CH)6COO�
Octadecadienoate
18 3 Linolenate all-cis-�9, �12, �15- CH3CH2(CH CHCH2)3(CH2)6COO�
Octadecatrienoate
20 4 Arachidonate all-cis �5, �8, �11, �14- CH3(CH2)4(CH CHCH2)4(CH2)2COO�
Eicosatetraenoate“
“
“““
it is preferable to refer to them according to their carboxylate form: for example,palmitate rather than palmitic acid.
¬CH2¬
C
OPalmitate
(ionized form of palmitic acid)
O
–
C
OHPalmitic acid
O
Fatty Acids Vary in Chain Length and Degree of UnsaturationFatty acids in biological systems usually contain an even number of carbonatoms, typically between 14 and 24 (Table 10.1), with the 16- and 18-carbon fattyacids being the most common. As heretofore described, the hydrocarbon chain maybe saturated or it may contain one or more double bonds. The configuration ofthe double bonds in most unsaturated fatty acids is cis. The double bonds inpolyunsaturated fatty acids are separated by at least one methylene group.
The properties of fatty acids and of lipids derived from them are markedlydependent on chain length and degree of saturation. Unsaturated fatty acids havelower melting points than those of saturated fatty acids of the same length. Forexample, the melting point of stearic acid is 69.6°C, whereas that of oleic acid(which contains one cis double bond) is 13.4°C. The melting points of polyunsat-urated fatty acids of the C18 series are even lower. The presence of a cis doublebond introduces a kink in the fatty acid and makes tight packing between thechains impossible. The lack of tight packing limits the van der Waals interactionsbetween chains, lowering the melting temperature.
Stearate
C
O
O
–
Methylene group
14410 Lipids

Chain length also affects the melting point, as illustrated by the fact that the melt-ing temperature of palmitic acid (C16) is 6.5 degrees lower than that of stearic acid(C18). Thus, short chain length and cis unsaturation enhance the fluidity of fatty acidsand of their derivatives. The fat that accumulates in the pan as bacon is fried is com-posed primarily of saturated fatty acids and solidifies soon after the burner is turnedoff. Olive oil, on the other hand, is composed of high concentrations of oleic acid andsome polyunsaturated fatty acids and remains liquid at room temperature. The vari-ability of melting points is not merely an arcane chemical insight. Melting tempera-tures of fatty acids are key elements in the control of the fluidity of cell membranes,and the proper degree of fluidity is essential for membrane function (Chapter 11).
The Degree and Type of Unsaturation Are Important to HealthAlthough fats are crucial biochemicals, too much saturated and trans-unsaturatedfats in the diet are correlated with high blood levels of cholesterol and cardiovas-cular disease. The biochemical basis for this correlation remains to be determined,although trans-unsaturated fats appear to cause inflammation. In contrast, certaincis-polyunsaturated fatty acids are essential in our diets because we cannot synthe-size them. Such fatty acids include the �-3 fatty acids—polyunsaturated fatty acidscommon in cold-water fish such as salmon, which have been suggested to play arole in protection from cardiovascular disease. The important �-3 fatty acids are�-linolenic acid, found in vegetable oils, eicosapentaenoic acid (EPA) and docosa-hexaenoic acid (DHA), both of which are found in fatty fish and shellfish.
QUICK QUIZ 1 What factorsdetermine the melting point of fatty
acids?
cis-Oleate
C
O
O
–
trans-Oleate
C
O
O
–
C
O
O
–
�-Linolenate
C
O
O
–
Eicosapentaenoate (EPA)
C
O
O
–
Docosahexaenoate (DHA)
14510.1 Fatty Acids

These fatty acids are precursors to important hormones. However, the natureof their protective effect against heart disease is not known. Studies show that �-3fatty acids, especially those derived from marine organisms, can prevent suddendeath from a heart attack, reduce triacylglycerides in the blood, lower blood pres-sure, and prevent thrombosis by slightly inhibiting blood clotting. Revealing themechanisms of action of these molecules is an active area of biochemical research.
10.2 Triacylglycerols Are the Storage Form of Fatty Acids
Despite the fact that fatty acids are our principal energy source, the con-centration of free fatty acids in cells or the blood is low because free fattyacids are strong acids. High concentrations of free fatty acids would dis-rupt the pH balance of the cells. Fatty acids required for energy genera-tion are stored in the form triacylglycerols, which are formed by theattachment of three fatty acid chains to a glycerol molecule. The fatty acidsare attached to the glycerol through ester linkages, in a process known asesterification. Common soaps are fatty acids generated by treating triacyl-glyerols with strong bases (Figure 10.2).
Three fattyacid chains
Glycerolbackbone
H2C CH2 CH3(CH2)NO C
O
HC CH2 CH3(CH2)NO C
O
H2C CH2 CH3(CH2)NO C
O
14610 Lipids
When energy is required during a fast (for instance, while sleeping),the fatty acids are cleaved from the triacyglycerol and carried to the cells.The ingestion of food replenishes the triacylglycerol stores.
As stated in Chapter 9, fatty acids are richer in energy than carbohy-drates. Additionally, compared with carbohydrates, fatty acids storeenergy more efficiently in the form of triacylglycerols, which arehydrophobic and so are stored in a nearly anhydrous form. Polar car-bohydrates, in contrast, bind to water molecules. Glycogen, for instance,binds to water. In fact, 1 g of dry glycogen binds about 2 g of water.Consequently, a gram of nearly anhydrous fat stores more than six timesas much energy as a gram of hydrated glycogen. For this reason, triacyl-glycerols rather than glycogen were selected in evolution as the majorenergy reservoir. Consider a typical 70-kg man. Triacylglycerols consti-tute about 11 kg of his total body weight. If this amount of energy werestored in glycogen, his total body weight would be 55 kg greater. Theglycogen and glucose stores provide enough energy to sustain biologi-cal function for about 18 to 24 hours, whereas the triacylglycerol storesallow survival for several weeks.
In mammals, the major site of accumulation of triacylglycerols isadipose tissue, which is distributed under the skin and elsewherethroughout the body. In adipose cells (fat cells), droplets of triacylglyc-erol coalesce to form a large globule in the cytoplasm, which may occupymost of the cell volume (Figure 10.3). Adipose cells are specialized forthe synthesis and storage of triacylglycerols and for their mobilization
Mitochondrion
Mitochondria
Lipiddroplet
Nucleus
Figure 10.3 Electron micrograph of an adipocyte. Asmall band of cytoplasm surrounds the large deposit oftriacylglycerols. [Biophoto Associates/Photo Researchers.]
Figure 10.2 Soaps are the sodium and potassiumsalts of long chain fatty acids. They are derived fromthe treatment of triacylglycerides with a strong base, aprocess called saponification. Common sources oftriacylglycerides are the animal lipids lard (from hogs)or tallow (from beef or sheep). [Photonica/Getty Images.]

14710.3 Types of Membrane Lipids
Fattyacid
Fattyacid
Glycerol
Phosphate Alcohol
Figure 10.5 Schematic structure of aphospholipid. The platform is glycerol inthis case.
into fuel molecules that are transported to other tissues by the blood. Adipose tis-sue also serves as a thermal insulator to help maintain body temperature. Polarbears provide an example of the insulating properties of adipose tissue. Duringfierce arctic storms, polar bears curl up into tight balls and are so well insulatedthat they cannot be detected by heat-sensing instruments.
The utility of triacylglycerols as an energy source is dramatically illustratedby the abilities of migratory birds, which can fly great distances without eating.Examples are the American golden plover and the ruby-throated hummingbird(Figure 10.4). The golden plover flies from Alaska to the southern tip of SouthAmerica; a large segment of the flight (3800 km, or 2400 miles) is over openocean, where the birds cannot feed. The ruby-throated hummingbird can flynonstop across the Gulf of Mexico. Fatty acids derived from triacylglycerols pro-vide the energy source for these prodigious feats.
10.3 There Are Three Common Types of Membrane Lipids
Thus far, we have considered only one type of complex lipid—triacylglycerols, thestorage form of fatty acids. However, lipids can be used for more than just fuel andenergy storage: they serve as hormones, messengers in signal-transduction path-ways, and components of membranes. The first two of these functions of lipidswill be considered in later chapters. We now consider the three major kinds ofmembrane lipids: phospholipids, glycolipids, and cholesterol.
Phospholipids Are the Major Class of Membrane LipidsPhospholipids are abundant in all biological membranes. A phospholipid mole-cule is constructed from four components: one or more fatty acids, a platform towhich the fatty acids are attached, a phosphate, and an alcohol attached to thephosphate (Figure 10.5).
The platform on which phospholipids are built may be glycerol, a three-carbon alcohol, or sphingosine, a more complex alcohol. Phospholipidsderived from glycerol are called phosphoglycerides and are composed of a glyc-erol backbone to which are attached two fatty acid chains and a phosphory-lated alcohol.
In phosphoglycerides, the hydroxyl groups at C-1 and C-2 of glycerol are ester-ified to the carboxyl groups of the two fatty acid chains. The C-3 hydroxyl group ofthe glycerol backbone is esterified to phosphoric acid. When no further additionsare made, the resulting compound is phosphatidate (diacylglycerol 3-phosphate),
Figure 10.4 Ruby-throated hummingbird. [William Leaman/Alamy.]

CC
OOCC
OO
CO H
CH2
H2C
ORR11
RR22
OP
O
O
OHO
OH
OH
HH
H
HH
OHOH
H–
CC
OOCC
OO
CO H
CH2
H2C
ORR11
RR22
OP
O
OO
H2C
CH2
NH3+–
CO H
CH2
H2C
OCC
RR11
OOCC
OO
RR22
OP
O
OO
H2C
CNH3
+
HCOO–
–
CC
OOCC
OO
CO H
CH2
H2C
ORR11
RR22
OP
O
OO
H2C
CH2
N
CH3
CH3CH3
– +
Phosphatidylserine Phosphatidylcholine
Phosphatidylethanolamine Phosphatidylinositol
Diphosphatidylglycerol (cardiolipin)
CC
OOCC
OO
CC
OO
CC
OO
CO H
CH
H2C
ORR11
RR22
OP
O
OO
H2C
C
H2C
C OH
H2C
CH2
O RR33
RR44
OP
O
OO
OHH
– –
2
Figure 10.7 Some common phosphoglycerides found in membranes.
the simplest phosphoglyceride (Figure 10.6). Only small amounts of phosphati-date are present in membranes. However, the molecule is a key intermediate inthe biosynthesis of the other phosphoglycerides as well as triacylglycerides(Chapter 28).
The major phosphoglycerides are derived from phosphatidate by the forma-tion of an ester linkage between the phosphoryl group of phosphatidate and thehydroxyl group of one of several alcohols. The common alcohol moieties ofphosphoglycerides are the amino acid serine, ethanolamine, choline, glycerol,and inositol. The structural formulas of phosphatidylcholine and the other prin-cipal phosphoglycerides are given in Figure 10.7.
CO H
CH2
H2C
OCC
R1R1
22
33
11
OOCC
OO
R2R2
OP
O
OO
2–
Phosphatidate(Diacylglycerol 3-phosphate)
Acyl groupswith fatty acidhydrocarbon chains
Acyl groupswith fatty acidhydrocarbon chains
Figure 10.6 Structure of phosphatidate(diacylglycerol 3-phosphate). The absoluteconfiguration of the central carbon atom(C-2) is shown.
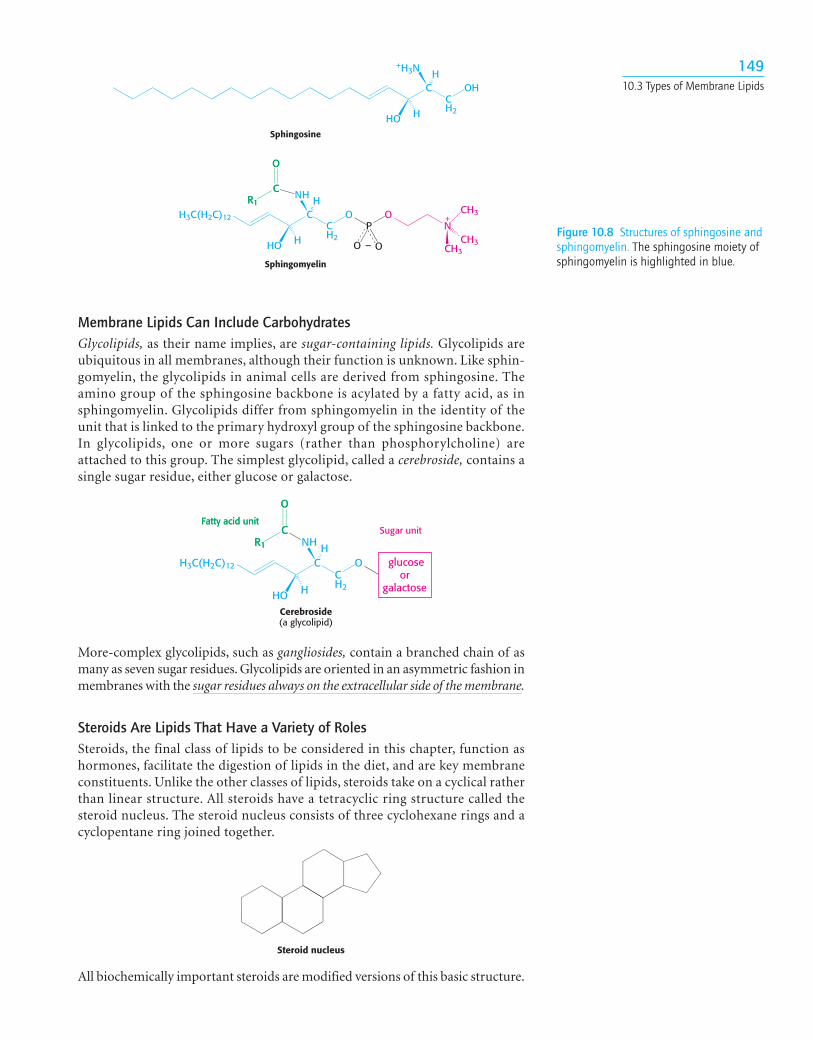
Phospholipids built on a sphingosine backbone are called sphingolipids.Sphingosine is an amino alcohol that contains a long, unsaturated hydrocarbonchain (Figure 10.8). Sphingomyelin is a common sphingolipid found in mem-branes. In sphingomyelin, the amino group of the sphingosine backbone islinked to a fatty acid by an amide bond. In addition, the primary hydroxylgroup of sphingosine is attached to phosphorylcholine through an ester link-age. Sphingomyelin is found in the plasma membrane of many cells but is espe-cially rich in the myelin sheath of nerve cells.
Niemann–Pick disease can result from anaccumulation of sphingomyelin owing tothe lack of sphingomyelinase, an enzymethat degrades sphingomyelin. Symptomsof Niemann–Pick disease include mentalretardation, seizures, eye paralysis,ataxia, and retarded growth.
14810 Lipids
14910.3 Types of Membrane Lipids
Figure 10.8 Structures of sphingosine andsphingomyelin. The sphingosine moiety ofsphingomyelin is highlighted in blue.
CCH2
OH
HO H
+H3N H
H3C(H2C)12 CCH2
O
HO H
NH HCC
OO
RR11
PO
O O
N
CH3
CH3CH3
+
–
Sphingosine
Sphingomyelin
Membrane Lipids Can Include CarbohydratesGlycolipids, as their name implies, are sugar-containing lipids. Glycolipids areubiquitous in all membranes, although their function is unknown. Like sphin-gomyelin, the glycolipids in animal cells are derived from sphingosine. Theamino group of the sphingosine backbone is acylated by a fatty acid, as insphingomyelin. Glycolipids differ from sphingomyelin in the identity of theunit that is linked to the primary hydroxyl group of the sphingosine backbone.In glycolipids, one or more sugars (rather than phosphorylcholine) areattached to this group. The simplest glycolipid, called a cerebroside, contains asingle sugar residue, either glucose or galactose.
Steroid nucleus
H3C(H2C)12 CCH2
O
HO H
NH H
CC
OO
RR11
glucoseor
galactose
Cerebroside(a glycolipid)
Sugar unitFatty acid unitFatty acid unit
More-complex glycolipids, such as gangliosides, contain a branched chain of asmany as seven sugar residues. Glycolipids are oriented in an asymmetric fashion inmembranes with the sugar residues always on the extracellular side of the membrane.
Steroids Are Lipids That Have a Variety of RolesSteroids, the final class of lipids to be considered in this chapter, function ashormones, facilitate the digestion of lipids in the diet, and are key membraneconstituents. Unlike the other classes of lipids, steroids take on a cyclical ratherthan linear structure. All steroids have a tetracyclic ring structure called thesteroid nucleus. The steroid nucleus consists of three cyclohexane rings and acyclopentane ring joined together.
All biochemically important steroids are modified versions of this basic structure.

A hydrocarbon tail is linked to the steroid at one end, and a hydroxyl group isattached at the other end. As we will see in Chapter 11, cholesterol is important inmaintaining proper membrane fluidity. Cholesterol is absent from prokaryotesbut is found to varying degrees in virtually all animal membranes. It constitutesalmost 25% of the membrane lipids in certain nerve cells but is essentially absentfrom some intracellular membranes. Free cholesterol does not exist outside ofmembranes. Rather, it is esterified to a fatty acid for storage and transport.
Biological Insight
Membranes of Extremeophiles Are Built from Ether Lipids with Branched ChainsThe membranes of archaea differ in composition from those of eukaryotes or bac-teria in two important ways. These differences clearly relate to the hostile livingconditions of many archaea. First, the fatty acid chains are joined to a glycerolbackbone by ether rather than ester linkages. The ether linkage is more resistantto hydrolysis than the ester linkage is. Second, the alkyl chains are branched ratherthan linear. They are built up from repeats of a fully saturated five-carbon frag-ment. These branched, saturated hydrocarbons are more resistant to oxidation.
HO
CH
Cholesterol
3
CH3
CH3
CH3
H3C
HH33CC O
HH33CC
C H
H2C
CH2O
OP
OCH2
H2C
NH3+
O
Membrane lipid from the archaeon Methanococcus jannaschii
O
HH33CC
HH33CC
HH33CC HH33CC HH33CCHH HH HH
CHCH33 CHCH33 CHCH33HH HH HH
–
The ability of these lipids to resist hydrolysis and oxidation may help theseorganisms to withstand the extreme conditions, such as high temperature, lowpH, or high salt concentration, under which some of these archaea grow. ■
Membrane Lipids Contain a Hydrophilic and a Hydrophobic MoietyThe repertoire of membrane lipids is extensive. However, these lipids possess acritical common structural theme: membrane lipids are amphipathic molecules(amphiphilic molecules) containing both a hydrophilic and a hydrophobic moiety.
Let us look at a model of a phosphoglyceride, such as phosphatidylcholine. Itsoverall shape is roughly rectangular (Figure 10.9). The two hydrophobic fatty acidchains are approximately parallel to each other, whereas the hydrophilic phospho-rylcholine moiety points in the opposite direction. Sphingomyelin has a similarconformation, as does the archaeal lipid depicted. Therefore, the following short-hand has been adopted to represent these membrane lipids: the hydrophilic unit,
The most common steroid, and the precursor to many biochemically activesteroids, is cholesterol.
15010 Lipids

15110.3 Types of Membrane Lipids
Phosphoglyceride
Sphingomyelin
Archaeal lipid Shorthand depiction
(A)
(B)
Figure 10.9 Representations of membranelipids. (A) Space-filling models of aphosphoglyceride, sphingomyelin, and anarchaeal lipid show their shapes anddistribution of hydrophilic andhydrophobic moieties. (B) A shorthanddepiction of a membrane lipid.
also called the polar head group, is represented by a circle, whereas the hydrocar-bon tails are depicted by straight or wavy lines (see Figure 10.9B).
Some Proteins Are Modified by the Covalent Attachment of Hydrophobic GroupsJust as carbohydrates are attached to proteins to modify the properties of proteins,lipids also can be attached to proteins to provide them with additional biochemi-cal properties. Often, such attachments are necessary for a protein to associate witha hydrophobic environment such as a membrane. These hydrophobic attachmentscan insert into the hydrophobic interior of the membrane and thus affix, or local-ize, the protein to the membrane surface. Such localization is required for proteinfunction. Three such attachments are shown in Figure 10.10: (1) a palmitoyl groupattached to a cysteine residue by a thioester bond, (2) a farnesyl group attached toa cysteine residue at the carboxyl terminus, and (3) a glycolipid structure termed aglycosylphosphatidylinositol (GPI) anchor attached to the carboxyl terminus.
HN
H S
OO
O
OCH3
NH
HN
H
O
OP
O O H2C
O
O
OHOR
OHOHO
OO
OO
OO
OO
O
RR
R
R
R
–
P
O O–
Cys
Carboxyl terminus
S-Palmitoylcysteine C-terminal S-farnesylcysteine methyl ester
Glycosyl phosphatidylinositol (GPI) anchor
HN
H SO
Cys
Figure 10.10 Membrane anchors.Membrane anchors arehydrophobic groups that arecovalently attached to proteins (inblue) and tether the proteins tothe membrane. The green circlesand blue square correspond tomannose and GlcNAc,respectively. R groups representpoints of additional modification.
QUICK QUIZ 2 What are the threemajor types of membrane lipids?

Clinical Insight
Premature Aging Can Result from the Improper Attachmentof a Hydrophobic Group to a Protein
Farnesyl is a hydrophobic group that is oftenattached to proteins, usually so that the protein isable to associate with a membrane (see Figure10.10). Inappropriate farnesylation has been shownto result in Hutchinson–Gilford progeria syndrome(HGPS), a rare disease of premature aging. Earlypostnatal development is normal, but the childrenfail to thrive, develop bone abnormalities, and havea small beaked nose, a receding jaw, and a completeloss of hair (Figure 10.11). Affected children usuallydie at an average age of 13 years of severe atheroscle-rosis, a cause of death more commonly seen in theelderly.
The cause of HGPS appears to be a mutation inthe gene for the nuclear protein lamin, a proteinthat forms a scaffold for the nucleus and may takepart in the regulation of gene expression. The foldedpolypeptide that will eventually become lamin ismodified and processed many times before the
mature protein is produced. One key processing event is the removal of a farne-syl group that had been added to the nascent protein earlier in processing. InHGPS patients, the farnesyl group is not removed, owing to a mutation in thelamin. The incorrectly processed lamin results in a deformed nucleus (see Figure10.11) and aberrant nuclear function that results in HGPS. Much researchremains to determine precisely how the failure to remove the farnesyl group leadsto such dramatic consequences. ■
SUMMARY
10.1 Fatty Acids Are a Main Source of FuelLipids are defined as water-insoluble molecules that are soluble in organicsolvents. Fatty acids are an important lipid in biochemistry. Fatty acids arehydrocarbon chains of various lengths and degrees of unsaturation that ter-minate with a carboxylic acid group. The fatty acid chains in membranesusually contain between 14 and 24 carbon atoms; they may be saturated orunsaturated. Short chain length and unsaturation enhance the fluidity offatty acids and their derivatives by lowering the melting temperature.
10.2 Triacylglycerols Are the Storage Form of Fatty AcidsFatty acids are stored as triacylglycerol molecules in adipose cells. Triacyl-glycerols are composed of three fatty acids esterified to a glycerol backbone.Triacylglycerols are stored in an anhydrous form.
10.3 There Are Three Common Types of Membrane LipidsThe major classes of membrane lipids are phospholipids, glycolipids, andcholesterol. Phosphoglycerides, a type of phospholipid, consist of a glycerolbackbone, two fatty acid chains, and a phosphorylated alcohol. Phos-phatidylcholine, phosphatidylserine, and phosphatidylethanolamine aremajor phosphoglycerides. Sphingomyelin, a different type of phospholipid,contains a sphingosine backbone instead of glycerol. Glycolipids are sugar-containing lipids derived from sphingosine. Cholesterol, which modulates
15210 Lipids
Figure 10.11 Huthchinson–Gilfordprogeria syndrome (HGPS). (A) A 15-year-old boy suffering from HGPS. (B) Anormal nucleus. (C) A nucleus from aHGPS patient. [(A) AP Photo/GeraldHerbert; (B and C) from P. Scaffidi, L. Gordon,and T. Misteli, PLoS Biol.3(11):e395/doi:10.1371/journal.pbio.0030395, 2005.
(C)(A)
(B)

153Problems
Key Terms
fatty acid (p. 143)triacylglycerol (p. 146)phospholipid (p. 147)sphingosine (p. 147)
phosphoglyceride (p. 147)sphingomyelin (p. 148)glycolipid (p. 149)cerebroside (p. 149)
ganglioside (p. 149)cholesterol (p. 150)amphipathic molecule (p. 150)
membrane fluidity, is constructed from a steroid nucleus. A common fea-ture of these membrane lipids is that they are amphipathic molecules, hav-ing one hydrophobic and one hydrophilic end.
CH3
CH3(CH2)15
+N(CH3)3
O
O
PO O
O O–
O
H
Answers to QUICK QUIZZES
1. Chain length and the degree of cis unsaturation. 2. Phospholipids, sphingolipids (of which glycolipids area subclass), and cholesterol.
1. Structure and name. Draw the structure of each of the fol-lowing fatty acids and give the structure its common name:
(a) n-Dodecanate(b) cis-�9-Hexadecenoate(c) cis, cis-�9, �12-Octadecadienoate
2. Fluidity matters. Triacylglycerols are used for fuel storagein both plants and animals. The triacylglycerols from plantsare often liquid at room temperature, whereas those fromanimals are solid. Suggest some reasons for this difference.
3. Contrast. Distinguish between phosphoglycerides andtriacylglycerols.
4. Compare. What structural features differentiate sphin-golipids from phosphoglycerides?
5. Depict a lipid. Draw the structure of a triacylglycerolcomposed of equal amounts of palmitic acid, stearic acid,and oleic acid.
6. Like finds like 1. What structural characteristic of lipidsaccounts for their solubility in organic solvents?
7. Like finds like 2. Suppose that a small amount of phospho-lipid were exposed to an aqueous solution. What structurewould the phospholipid molecules assume? What would bethe driving force for the formation of this structure?
8. Linkages. Platelet-activating factor (PAF) is a phospho-lipid that plays a role in allergic and inflammatoryresponses, as well as in toxic shock syndrome. The structureof PAF is shown here. How does it differ from the structuresof the phospholipids discussed in this chapter?
9. Coming clean. Draw the structure of the sodium salt ofstearic acid. How might it function to remove grease fromyour clothes or your hands?
10. Hard-water problems. Some metal salts of fatty acids arenot as soluble as the sodium or potassium salts. Forinstance, magnesium or calcium salts of fatty acids arepoorly soluble. What would be the effect of taking a bath inwater that is rich in magnesium or calcium?
11. Melting point 1. Explain why oleic acid (18 carbons, onecis bond) has a lower melting point than stearic acid, whichhas the same number of carbon atoms but is saturated. Howwould you expect the melting point of trans-oleic acid tocompare with that of cis-oleic acid?
12. Melting point 2. Explain why the melting point of palmiticacid (C16) is 6.5 degrees lower than that of stearic acid (C18).
13. A sound diet. Small mammalian hibernators can withstandbody temperatures of 0° to 5°C without injury. However, thebody fats of most mammals have melting temperatures ofapproximately 25°C. Predict how the composition of thebody fat of hibernators might differ from that of their non-hibernating cousins.
Problems
Platelet-activating factor (PAF)
Selected readings for this chapter can be found online at www.whfreeman.com/Tymoczko


















