
Biogeography of Stigmaphyllon (Malpighiaceae) and a Meta ...
20
plants Article Biogeography of Stigmaphyllon (Malpighiaceae) and a Meta-Analysis of Vascular Plant Lineages Diversified in the Brazilian Atlantic Rainforests Point to the Late Eocene Origins of This Megadiverse Biome Rafael Felipe de Almeida 1,2, * and Cássio van den Berg 1 1 Departamento de Ciências Biológicas, Universidade Estadual de Feira de Santana, Molecular Biology of Plants and Fungi Lab (LAMOL), Av. Transnordestina s/n, Novo Horizonte, Feira de Santana 44036-900, Bahia, Brazil; [email protected] 2 Scientifik Consultoria, Petrópolis, Rio de Janeiro 25651-090, Brazil * Correspondence: [email protected] Received: 16 October 2020; Accepted: 11 November 2020; Published: 13 November 2020 Abstract: We investigated the biogeography of Stigmaphyllon, the second-largest lianescent genus of Malpighiaceae, as a model genus to reconstruct the age and biogeographic history of the Brazilian Atlantic Rainforest (BAF). Few studies to date have focused on the tertiary diversification of plant lineages in the BAFs, especially on Stigmaphyllon. Phylogenetic relationships for 24 species of Stigmaphyllon (18 ssp. From the Atlantic forest (out of 31 spp.), three spp. from the Amazon Rainforest, two spp. from the Caatinga biome, and a single species from the Cerrado biome) were inferred based on one nuclear DNA (PHYC) and two ribosomal DNA (ETS, ITS) regions using parsimony and Bayesian methods. A time-calibrated phylogenetic tree for ancestral area reconstructions was additionally generated, coupled with a meta-analysis of vascular plant lineages diversified in the BAFs. Our results show that: (1) Stigmaphyllon is monophyletic, but its subgenera are paraphyletic; (2) the most recent common ancestor of Stigmaphyllon originated in the Brazilian Atlantic Rainforest/Caatinga region in Northeastern Brazil ca. 26.0 Mya; (3) the genus colonized the Amazon Rainforest at two different times (ca. 22.0 and 6.0 Mya), the Caatinga biome at least four other times (ca. 14.0, 9.0, 7.0, and 1.0 Mya), the Cerrado biome a single time (ca. 15.0 Mya), and the Southern Atlantic Rainforests five times (from 26.0 to 9.0 Mya); (4) a history of at least seven expansion events connecting the Brazilian Atlantic Rainforest to other biomes from 26.0 to 9.0 Mya, and (5) a single dispersion event from South America to Southeastern Asia and Oceania at 22.0 Mya via Antarctica was proposed. Compared to a meta-analysis of time-calibrated phylogenies for 64 lineages of vascular plants diversified in the Brazilian Atlantic Rainforests, our results point to a late Eocene origin for this megadiverse biome. Keywords: ancestral area reconstruction; Atlantic rainforest; time-calibrated phylogenies; Malpighiales; neotropical flora 1. Introduction Malpighiaceae Juss. is one of the most diverse plant families of Neotropical shrubs, trees, and lianas [1], with most species confined to this region [2]. Its species are easily recognized by their remarkable floral conservatism, with flowers frequently bearing a pair of oil-secreting glands at the base of sepals, petals clawed at the base, and a posterior petal differentiated from the remaining lateral four [1,2]. This family has received broad phylogenetic attention in the past few years [2–8], including more focused investigations on generic delimitations and the phylogenetic position of Plants 2020, 9, 1569 ; doi:10.3390/plants9111569 www.mdpi.com/journal/plants
Transcript of Biogeography of Stigmaphyllon (Malpighiaceae) and a Meta ...

plants
Article
Biogeography of Stigmaphyllon (Malpighiaceae) anda Meta-Analysis of Vascular Plant LineagesDiversified in the Brazilian Atlantic Rainforests Pointto the Late Eocene Origins of This Megadiverse Biome
Rafael Felipe de Almeida 12 and Caacutessio van den Berg 1
1 Departamento de Ciecircncias Bioloacutegicas Universidade Estadual de Feira de Santana Molecular Biology ofPlants and Fungi Lab (LAMOL) Av Transnordestina sn Novo HorizonteFeira de Santana 44036-900 Bahia Brazil vcassiogmailcom
2 Scientifik Consultoria Petroacutepolis Rio de Janeiro 25651-090 Brazil Correspondence dealmeidarafaelfelipegmailcom
Received 16 October 2020 Accepted 11 November 2020 Published 13 November 2020
Abstract We investigated the biogeography of Stigmaphyllon the second-largest lianescent genus ofMalpighiaceae as a model genus to reconstruct the age and biogeographic history of the BrazilianAtlantic Rainforest (BAF) Few studies to date have focused on the tertiary diversification of plantlineages in the BAFs especially on Stigmaphyllon Phylogenetic relationships for 24 species ofStigmaphyllon (18 ssp From the Atlantic forest (out of 31 spp) three spp from the Amazon Rainforesttwo spp from the Caatinga biome and a single species from the Cerrado biome) were inferredbased on one nuclear DNA (PHYC) and two ribosomal DNA (ETS ITS) regions using parsimonyand Bayesian methods A time-calibrated phylogenetic tree for ancestral area reconstructions wasadditionally generated coupled with a meta-analysis of vascular plant lineages diversified in the BAFsOur results show that (1) Stigmaphyllon is monophyletic but its subgenera are paraphyletic (2) themost recent common ancestor of Stigmaphyllon originated in the Brazilian Atlantic RainforestCaatingaregion in Northeastern Brazil ca 260 Mya (3) the genus colonized the Amazon Rainforest at twodifferent times (ca 220 and 60 Mya) the Caatinga biome at least four other times (ca 140 90 70and 10 Mya) the Cerrado biome a single time (ca 150 Mya) and the Southern Atlantic Rainforestsfive times (from 260 to 90 Mya) (4) a history of at least seven expansion events connecting theBrazilian Atlantic Rainforest to other biomes from 260 to 90 Mya and (5) a single dispersion eventfrom South America to Southeastern Asia and Oceania at 220 Mya via Antarctica was proposedCompared to a meta-analysis of time-calibrated phylogenies for 64 lineages of vascular plantsdiversified in the Brazilian Atlantic Rainforests our results point to a late Eocene origin for thismegadiverse biome
Keywords ancestral area reconstruction Atlantic rainforest time-calibrated phylogeniesMalpighiales neotropical flora
1 Introduction
Malpighiaceae Juss is one of the most diverse plant families of Neotropical shrubs treesand lianas [1] with most species confined to this region [2] Its species are easily recognized by theirremarkable floral conservatism with flowers frequently bearing a pair of oil-secreting glands at thebase of sepals petals clawed at the base and a posterior petal differentiated from the remaininglateral four [12] This family has received broad phylogenetic attention in the past few years [2ndash8]including more focused investigations on generic delimitations and the phylogenetic position of
Plants 2020 9 1569 doi103390plants9111569 wwwmdpicomjournalplants
Plants 2020 9 1569 2 of 20
Old-World clades [26] However there have been few efforts to determine finer-scale patterns ofmolecular analyses and historical biogeography in Neotropical Malpighiaceae [4689]
Stigmaphyllon AJuss is one of the several genera in Malpighiaceae re-circumscribed due torecent molecular phylogenetic studies [26] It is currently the second-largest lianescent genus inMalpighiaceae and the only occurring in forested habitats in the tropics and subtropics of AmericaAfrica Asia and Oceania [10] Most species are woody lianas with long-petioled elliptical to cordateleaves corymbs or umbels of yellow flowers arranged in dichasia styles holding laterally expandedappendages in the apex and schizocarpic fruits splitting into three-winged mericarps bearing a sizeabledorsal wing (Figure 1 andashh) [1011] This genus is currently divided into two subgenera Stigmaphyllonand Ryssopterys treated as separate genera before molecular phylogenies [10] Stigmaphyllon subgenusStigmaphyllon includes ca 90 species restricted to the Neotropics except for Stigmaphyllon bannisterioidesAJuss which is also found in West Africa [11] On the other hand S subgenus Ryssopterys includesca 20 species restricted to Southeast Asia and Oceania [10] The monophyly of Stigmaphyllon and itssubgenera has been suggested by previous phylogenetic studies but never adequately corroboratedbecause its type species was never sampled [2356]
Previous phylogenetic studies for Malpighiaceae suggested a Brazilian Atlantic Rainforest (hereintreated as BAF) origin for Stigmaphyllon with S paralias AJuss being consistently recovered in severalstudies as the first lineage to diverge in the genus [26] Even though the BAFs currently comprisemore than 15000 known vascular plant species [12] they are regarded as one of the most threatenedhotspots for biodiversity worldwide due to being mostly fragmented and disturbed by most of Brazilrsquoshuman population and economic activity [13] Several biogeographic hypotheses have been proposedto explain the origins of the great biodiversity of the BAFs such as 1 Miocene to Pleistocene forestcorridors between BAFs and the Amazon rainforests via Cerradorsquos gallery forests andor via thecoastal region in Northeastern Brazil [14] and 2 Pleistocene refugia hypothesis which suggestedthat Pleistocene climatic fluctuations led to rainforest fragmentation and promoted divergence oflineages or species in isolated forest fragments or refugia [15] Pleistocene diversifications in BAFshave been greatly criticized due to the lack of concordance with empirical phylogenetic data as wellas by the evidence that shifts in forest species distribution rather than fragmentation have been themain consequences of global glaciations in the Neotropics [16] Although most previous studies havefocused on explaining BAFs biodiversity through Miocene to Pleistocene climaticgeological events nostudy to date has focused on timing the age of vascular plant lineages diversification in BAFs
In this study we focus on timing the biogeographic history of BAFs using Stigmaphyllon as amodel genus supplemented by a meta-analysis of vascular plant lineages diversified in this biome toinfer the age of BAFs More specifically we (1) test the monophyly of Stigmaphyllon and its subgenera(2) time-calibrate the phylogenetic tree (3) estimate the ancestral areas of Stigmaphyllon and (4) shedsome light on the age and biogeographic history of BAFs using Stigmaphyllon and a meta-analysis ofvascular plant lineages diversified in this biome
Plants 2020 9 1569 3 of 20
Figure 1 Phylogram from the combined analysis of nuclear and ribosomal markers for StigmaphyllonPosterior probabilities are shown above branches and bootstrap values below branches Black barsrepresent S subg Stigmaphyllon and the white bar represents S subg Ryssopterys (dark blue bar)S paralias group (yellow bar) S ciliatum group (dark green bar) S subg Ryssopterys (orange bar)S finlayanum group (purple bar) S auriculatum group (light green bar) S urenifolium group (lightblue bar) S lalandianum group (red bar) S sinuatum group (pink bar) S blanchetii group (grey bar)S saxicola group (A) adaxial surface of a leaf of S angustilobum AJuss (B) Umbel of S ciliatum (Lam)AJuss in side view evidencing ciliate reduced leaves associated with the inflorescence (C) open flowerof S ciliatum in frontal view (D) androecium and gynoecium of S ciliatum in side view (E) mericarp ofS paralias AJuss in side view (F) winged mericarp of S ciliatum in side view (G) winged mericarp ofS auriculatum (Cav) AJuss in side view (H) winged mericarp of S saxicola CEAnderson in side view(I) winged mericarps of S blanchetii in side view (photographs by RFAlmeida)
2 Results
21 Phylogenetic Analysis
Our combined dataset for ribosomal (ETS ITS) and nuclear (PHYC) markers contains a total of2283 characters of which 1724 characters are constant 257 characters are variable but parsimonyuninformative and 302 characters are parsimony informative The combined analysis of nuclearand ribosomal markers provides higher support for more clades than those based on independentnuclear and ribosomal datasets Overlapping peaks (paralogous copies) for ETS and ITS were notrecorded during electrophoresis and sequencing The heuristic search for the combined dataset found
Plants 2020 9 1569 4 of 20
15 trees (consistency index CI = 080 retention index RI = 070) and the strict consensus tree includes17 moderately supported clades (bootstrap percentage BP gt 75 Figure 1) The Bayesian analysisrecovered 25 well-supported to moderately supported clades (posterior probabilities PP gt 095 andgt080 respectively Figure 1)
The monophyly of Stigmaphyllon (Figure 1) is strongly supported by Maximum Parsimony (MP)and Bayesian Inference (BI) analyses being recovered as sister to Diplopterys The clade Stigmaphyllon+ Diplopterys is well supported (BS100PP10) as sister to Bronwenia (Figure 1) Within StigmaphyllonStigmaphyllon subg Stigmaphyllon is recovered as polyphyletic with its representatives placed inthree separate lineages (Figure 1 black bars) named the early-diverging Stigmaphyllon paralias group(Figure 1 dark blue bar) Stigmaphyllon ciliatum group (Figure 1 yellow bar) and core Stigmaphyllon(here represented by the common ancestor that the lineage S fynlayanum + S puberulum share with theremaining species (Figure 1 black bar)) Stigmaphyllon subg Ryssopterys (Figure 1 dark green bar) isrecovered as monophyletic in our analyses being sister to the Stigmaphyllon ciliatum group (Figure 1white bar)
Within core Stigmaphyllon seven major lineages are recovered here designated as 1 S puberulumgroup 2 S auriculatum group 3 S urenifolium group 4 S lalandianum group 5 S sinuatum group6 S blanchetii group and 7 S gayanum group The S puberulum group (Figure 1 orange bar) iswell-supported (BS100PP10) comprising only two species S finlayanum AJuss and S puberulumGriseb while the S auriculatum group (Figure 1 purple bar) is moderate to well-supported (B77PP10)comprising three species S angustilobum AJuss S arenicola CEAnderson and S auriculatum(Cav) AJuss The S urenifolium group (Figure 1 light green bar) is weak to well-supported(BS70PP10) represented by only its naming species In contrast the S lalandianum group (Figure 1light blue bar) is moderate to well-supported (BS82PP10) comprising four species S alternifoliumAJuss S bonariense (Hook and Arn) CEAnderson S lalandianum AJuss and S vitifolium AJussThe S sinuatum group (Figure 1 red bar) is well-supported (BS100PP10) represented by only twospecies S lindenianum AJuss and S sinuatum (DC) AJuss while the S blanchetii group (Figure 1rose bar) is weakly-supported (BS-PP060) comprising four species S blanchetii CEAndersonS caatingicola RFAlmeida and Amorim S jatrophifolium AJuss and S salzmannii AJuss Finallythe S gayanum group (Figure 1 gray bar) is weakly to well-supported (BS-PP10) and represented byfour species S cavernulosum CEAnderson S gayanum AJuss S macropodum AJuss and S saxicolaCEAnderson
22 Ancestral Area Reconstruction and Divergence Times Estimation
The S-DEC reconstruction suggests that the most recent common ancestor (MRCA) of Stigmaphyllonwas widespread in the northern portion of the BAFs ca 260 Mya (Figures 2 and 3 node 3 Table 1)A dispersal event (node 3) took place then splitting the MRCA of the Stigmaphyllon paralias groupfrom the remaining species which dispersed from Seasonally Dry Tropical Forests (SDTFs) to CamposRupestres in northeastern Brazil ca 140 Mya (Figures 2 and 3 node 4 Table 1) The MRCA of theS ciliatum and S timoriense (DC) CEAnderson clade diverged ca 220 Mya (node 5) and split intothose lineages ca 100 Mya (node 6) colonizing dunes vegetation on the Atlantic and Australasianrainforests (Figures 2 and 3 node 6 Table 1) The MRCA of core Stigmaphyllon dispersed to thesouthern portion of the BAFs ca 190 Mya (Figures 2 and 3 node 7 Table 1) The MRCA of theS puberulum group arose 625 Mya being widespread over both the Amazon and Atlantic rainforestswith a vicariant event giving rise to its current lineages (Figures 2 and 3 node 8 Table 1) The MRCA ofthe remaining species (node 9) remained distributed in the BAFs ca 170 Mya giving rise to the MRCAof the S auriculatum group ca 94 Mya (node 10) A dispersal event followed by the colonizationof dunes vegetation in Southern Brazil gave rise to the S arenicola lineage (Figures 2 and 3 node 10Table 1) In the Pleistocene ca 072 Mya the MRCA of the S auriculatum and S angustilobum arose inthe BAFs from Southeastern Brazil giving rise to those lineages via a dispersal event from rainforests toSDTFs (Figures 2 and 3 node 11 Table 1) The MRCA of S urenifolium (Figures 2 and 3 node 12 Table 1)
Plants 2020 9 1569 5 of 20
arose ca 150 Mya via a dispersal and a vicariant event from the BAFs to the Cerrado The MRCA ofthe remaining species (Figures 2 and 3 node 13 Table 1) arose ca 130 Mya in the BAFs and probablyin the Amazon forest as well The MRCA of node 14 remained distributed within the Southeasternportion of the BAFs ca 70 Mya (Figures 2 and 3 node 14 Table 1) and diversified into its four mainlineages ca 40 Mya (Figures 2 and 3 nodes 15ndash16 Table 1) Both a dispersal and a vicariant eventtook place ca 120 Mya giving rise to the MRCA of node 17 which was distributed within the BAFsCaatinga and the Amazon rainforest The lineage of node 18 colonized the Amazon rainforest for thefirst time but only diversified ca 05 Mya (Figures 2 and 3 node 18 Table 1)
Figure 2 Chronogram and Statistical-Dispersal-Extinction-Cladogenesis (S-DEC) ancestral areareconstructions for Stigmaphyllon Nodes are numbered from 1 to 26 Numbers above branchesrepresent estimated ages (Mya) Branch letters on the left represent the reconstructed ancestralarea(s) (A) Atlantic rainforest (B) Seasonally Dry Tropical Forest (C) Amazon rainforest (D) Cerrado(E) Australasian rainforests Branch letters on the right represent dispersal (d) or vicariant (v) events
Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 2 of 20
Old-World clades [26] However there have been few efforts to determine finer-scale patterns ofmolecular analyses and historical biogeography in Neotropical Malpighiaceae [4689]
Stigmaphyllon AJuss is one of the several genera in Malpighiaceae re-circumscribed due torecent molecular phylogenetic studies [26] It is currently the second-largest lianescent genus inMalpighiaceae and the only occurring in forested habitats in the tropics and subtropics of AmericaAfrica Asia and Oceania [10] Most species are woody lianas with long-petioled elliptical to cordateleaves corymbs or umbels of yellow flowers arranged in dichasia styles holding laterally expandedappendages in the apex and schizocarpic fruits splitting into three-winged mericarps bearing a sizeabledorsal wing (Figure 1 andashh) [1011] This genus is currently divided into two subgenera Stigmaphyllonand Ryssopterys treated as separate genera before molecular phylogenies [10] Stigmaphyllon subgenusStigmaphyllon includes ca 90 species restricted to the Neotropics except for Stigmaphyllon bannisterioidesAJuss which is also found in West Africa [11] On the other hand S subgenus Ryssopterys includesca 20 species restricted to Southeast Asia and Oceania [10] The monophyly of Stigmaphyllon and itssubgenera has been suggested by previous phylogenetic studies but never adequately corroboratedbecause its type species was never sampled [2356]
Previous phylogenetic studies for Malpighiaceae suggested a Brazilian Atlantic Rainforest (hereintreated as BAF) origin for Stigmaphyllon with S paralias AJuss being consistently recovered in severalstudies as the first lineage to diverge in the genus [26] Even though the BAFs currently comprisemore than 15000 known vascular plant species [12] they are regarded as one of the most threatenedhotspots for biodiversity worldwide due to being mostly fragmented and disturbed by most of Brazilrsquoshuman population and economic activity [13] Several biogeographic hypotheses have been proposedto explain the origins of the great biodiversity of the BAFs such as 1 Miocene to Pleistocene forestcorridors between BAFs and the Amazon rainforests via Cerradorsquos gallery forests andor via thecoastal region in Northeastern Brazil [14] and 2 Pleistocene refugia hypothesis which suggestedthat Pleistocene climatic fluctuations led to rainforest fragmentation and promoted divergence oflineages or species in isolated forest fragments or refugia [15] Pleistocene diversifications in BAFshave been greatly criticized due to the lack of concordance with empirical phylogenetic data as wellas by the evidence that shifts in forest species distribution rather than fragmentation have been themain consequences of global glaciations in the Neotropics [16] Although most previous studies havefocused on explaining BAFs biodiversity through Miocene to Pleistocene climaticgeological events nostudy to date has focused on timing the age of vascular plant lineages diversification in BAFs
In this study we focus on timing the biogeographic history of BAFs using Stigmaphyllon as amodel genus supplemented by a meta-analysis of vascular plant lineages diversified in this biome toinfer the age of BAFs More specifically we (1) test the monophyly of Stigmaphyllon and its subgenera(2) time-calibrate the phylogenetic tree (3) estimate the ancestral areas of Stigmaphyllon and (4) shedsome light on the age and biogeographic history of BAFs using Stigmaphyllon and a meta-analysis ofvascular plant lineages diversified in this biome
Plants 2020 9 1569 3 of 20
Figure 1 Phylogram from the combined analysis of nuclear and ribosomal markers for StigmaphyllonPosterior probabilities are shown above branches and bootstrap values below branches Black barsrepresent S subg Stigmaphyllon and the white bar represents S subg Ryssopterys (dark blue bar)S paralias group (yellow bar) S ciliatum group (dark green bar) S subg Ryssopterys (orange bar)S finlayanum group (purple bar) S auriculatum group (light green bar) S urenifolium group (lightblue bar) S lalandianum group (red bar) S sinuatum group (pink bar) S blanchetii group (grey bar)S saxicola group (A) adaxial surface of a leaf of S angustilobum AJuss (B) Umbel of S ciliatum (Lam)AJuss in side view evidencing ciliate reduced leaves associated with the inflorescence (C) open flowerof S ciliatum in frontal view (D) androecium and gynoecium of S ciliatum in side view (E) mericarp ofS paralias AJuss in side view (F) winged mericarp of S ciliatum in side view (G) winged mericarp ofS auriculatum (Cav) AJuss in side view (H) winged mericarp of S saxicola CEAnderson in side view(I) winged mericarps of S blanchetii in side view (photographs by RFAlmeida)
2 Results
21 Phylogenetic Analysis
Our combined dataset for ribosomal (ETS ITS) and nuclear (PHYC) markers contains a total of2283 characters of which 1724 characters are constant 257 characters are variable but parsimonyuninformative and 302 characters are parsimony informative The combined analysis of nuclearand ribosomal markers provides higher support for more clades than those based on independentnuclear and ribosomal datasets Overlapping peaks (paralogous copies) for ETS and ITS were notrecorded during electrophoresis and sequencing The heuristic search for the combined dataset found
Plants 2020 9 1569 4 of 20
15 trees (consistency index CI = 080 retention index RI = 070) and the strict consensus tree includes17 moderately supported clades (bootstrap percentage BP gt 75 Figure 1) The Bayesian analysisrecovered 25 well-supported to moderately supported clades (posterior probabilities PP gt 095 andgt080 respectively Figure 1)
The monophyly of Stigmaphyllon (Figure 1) is strongly supported by Maximum Parsimony (MP)and Bayesian Inference (BI) analyses being recovered as sister to Diplopterys The clade Stigmaphyllon+ Diplopterys is well supported (BS100PP10) as sister to Bronwenia (Figure 1) Within StigmaphyllonStigmaphyllon subg Stigmaphyllon is recovered as polyphyletic with its representatives placed inthree separate lineages (Figure 1 black bars) named the early-diverging Stigmaphyllon paralias group(Figure 1 dark blue bar) Stigmaphyllon ciliatum group (Figure 1 yellow bar) and core Stigmaphyllon(here represented by the common ancestor that the lineage S fynlayanum + S puberulum share with theremaining species (Figure 1 black bar)) Stigmaphyllon subg Ryssopterys (Figure 1 dark green bar) isrecovered as monophyletic in our analyses being sister to the Stigmaphyllon ciliatum group (Figure 1white bar)
Within core Stigmaphyllon seven major lineages are recovered here designated as 1 S puberulumgroup 2 S auriculatum group 3 S urenifolium group 4 S lalandianum group 5 S sinuatum group6 S blanchetii group and 7 S gayanum group The S puberulum group (Figure 1 orange bar) iswell-supported (BS100PP10) comprising only two species S finlayanum AJuss and S puberulumGriseb while the S auriculatum group (Figure 1 purple bar) is moderate to well-supported (B77PP10)comprising three species S angustilobum AJuss S arenicola CEAnderson and S auriculatum(Cav) AJuss The S urenifolium group (Figure 1 light green bar) is weak to well-supported(BS70PP10) represented by only its naming species In contrast the S lalandianum group (Figure 1light blue bar) is moderate to well-supported (BS82PP10) comprising four species S alternifoliumAJuss S bonariense (Hook and Arn) CEAnderson S lalandianum AJuss and S vitifolium AJussThe S sinuatum group (Figure 1 red bar) is well-supported (BS100PP10) represented by only twospecies S lindenianum AJuss and S sinuatum (DC) AJuss while the S blanchetii group (Figure 1rose bar) is weakly-supported (BS-PP060) comprising four species S blanchetii CEAndersonS caatingicola RFAlmeida and Amorim S jatrophifolium AJuss and S salzmannii AJuss Finallythe S gayanum group (Figure 1 gray bar) is weakly to well-supported (BS-PP10) and represented byfour species S cavernulosum CEAnderson S gayanum AJuss S macropodum AJuss and S saxicolaCEAnderson
22 Ancestral Area Reconstruction and Divergence Times Estimation
The S-DEC reconstruction suggests that the most recent common ancestor (MRCA) of Stigmaphyllonwas widespread in the northern portion of the BAFs ca 260 Mya (Figures 2 and 3 node 3 Table 1)A dispersal event (node 3) took place then splitting the MRCA of the Stigmaphyllon paralias groupfrom the remaining species which dispersed from Seasonally Dry Tropical Forests (SDTFs) to CamposRupestres in northeastern Brazil ca 140 Mya (Figures 2 and 3 node 4 Table 1) The MRCA of theS ciliatum and S timoriense (DC) CEAnderson clade diverged ca 220 Mya (node 5) and split intothose lineages ca 100 Mya (node 6) colonizing dunes vegetation on the Atlantic and Australasianrainforests (Figures 2 and 3 node 6 Table 1) The MRCA of core Stigmaphyllon dispersed to thesouthern portion of the BAFs ca 190 Mya (Figures 2 and 3 node 7 Table 1) The MRCA of theS puberulum group arose 625 Mya being widespread over both the Amazon and Atlantic rainforestswith a vicariant event giving rise to its current lineages (Figures 2 and 3 node 8 Table 1) The MRCA ofthe remaining species (node 9) remained distributed in the BAFs ca 170 Mya giving rise to the MRCAof the S auriculatum group ca 94 Mya (node 10) A dispersal event followed by the colonizationof dunes vegetation in Southern Brazil gave rise to the S arenicola lineage (Figures 2 and 3 node 10Table 1) In the Pleistocene ca 072 Mya the MRCA of the S auriculatum and S angustilobum arose inthe BAFs from Southeastern Brazil giving rise to those lineages via a dispersal event from rainforests toSDTFs (Figures 2 and 3 node 11 Table 1) The MRCA of S urenifolium (Figures 2 and 3 node 12 Table 1)
Plants 2020 9 1569 5 of 20
arose ca 150 Mya via a dispersal and a vicariant event from the BAFs to the Cerrado The MRCA ofthe remaining species (Figures 2 and 3 node 13 Table 1) arose ca 130 Mya in the BAFs and probablyin the Amazon forest as well The MRCA of node 14 remained distributed within the Southeasternportion of the BAFs ca 70 Mya (Figures 2 and 3 node 14 Table 1) and diversified into its four mainlineages ca 40 Mya (Figures 2 and 3 nodes 15ndash16 Table 1) Both a dispersal and a vicariant eventtook place ca 120 Mya giving rise to the MRCA of node 17 which was distributed within the BAFsCaatinga and the Amazon rainforest The lineage of node 18 colonized the Amazon rainforest for thefirst time but only diversified ca 05 Mya (Figures 2 and 3 node 18 Table 1)
Figure 2 Chronogram and Statistical-Dispersal-Extinction-Cladogenesis (S-DEC) ancestral areareconstructions for Stigmaphyllon Nodes are numbered from 1 to 26 Numbers above branchesrepresent estimated ages (Mya) Branch letters on the left represent the reconstructed ancestralarea(s) (A) Atlantic rainforest (B) Seasonally Dry Tropical Forest (C) Amazon rainforest (D) Cerrado(E) Australasian rainforests Branch letters on the right represent dispersal (d) or vicariant (v) events
Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 3 of 20
Figure 1 Phylogram from the combined analysis of nuclear and ribosomal markers for StigmaphyllonPosterior probabilities are shown above branches and bootstrap values below branches Black barsrepresent S subg Stigmaphyllon and the white bar represents S subg Ryssopterys (dark blue bar)S paralias group (yellow bar) S ciliatum group (dark green bar) S subg Ryssopterys (orange bar)S finlayanum group (purple bar) S auriculatum group (light green bar) S urenifolium group (lightblue bar) S lalandianum group (red bar) S sinuatum group (pink bar) S blanchetii group (grey bar)S saxicola group (A) adaxial surface of a leaf of S angustilobum AJuss (B) Umbel of S ciliatum (Lam)AJuss in side view evidencing ciliate reduced leaves associated with the inflorescence (C) open flowerof S ciliatum in frontal view (D) androecium and gynoecium of S ciliatum in side view (E) mericarp ofS paralias AJuss in side view (F) winged mericarp of S ciliatum in side view (G) winged mericarp ofS auriculatum (Cav) AJuss in side view (H) winged mericarp of S saxicola CEAnderson in side view(I) winged mericarps of S blanchetii in side view (photographs by RFAlmeida)
2 Results
21 Phylogenetic Analysis
Our combined dataset for ribosomal (ETS ITS) and nuclear (PHYC) markers contains a total of2283 characters of which 1724 characters are constant 257 characters are variable but parsimonyuninformative and 302 characters are parsimony informative The combined analysis of nuclearand ribosomal markers provides higher support for more clades than those based on independentnuclear and ribosomal datasets Overlapping peaks (paralogous copies) for ETS and ITS were notrecorded during electrophoresis and sequencing The heuristic search for the combined dataset found
Plants 2020 9 1569 4 of 20
15 trees (consistency index CI = 080 retention index RI = 070) and the strict consensus tree includes17 moderately supported clades (bootstrap percentage BP gt 75 Figure 1) The Bayesian analysisrecovered 25 well-supported to moderately supported clades (posterior probabilities PP gt 095 andgt080 respectively Figure 1)
The monophyly of Stigmaphyllon (Figure 1) is strongly supported by Maximum Parsimony (MP)and Bayesian Inference (BI) analyses being recovered as sister to Diplopterys The clade Stigmaphyllon+ Diplopterys is well supported (BS100PP10) as sister to Bronwenia (Figure 1) Within StigmaphyllonStigmaphyllon subg Stigmaphyllon is recovered as polyphyletic with its representatives placed inthree separate lineages (Figure 1 black bars) named the early-diverging Stigmaphyllon paralias group(Figure 1 dark blue bar) Stigmaphyllon ciliatum group (Figure 1 yellow bar) and core Stigmaphyllon(here represented by the common ancestor that the lineage S fynlayanum + S puberulum share with theremaining species (Figure 1 black bar)) Stigmaphyllon subg Ryssopterys (Figure 1 dark green bar) isrecovered as monophyletic in our analyses being sister to the Stigmaphyllon ciliatum group (Figure 1white bar)
Within core Stigmaphyllon seven major lineages are recovered here designated as 1 S puberulumgroup 2 S auriculatum group 3 S urenifolium group 4 S lalandianum group 5 S sinuatum group6 S blanchetii group and 7 S gayanum group The S puberulum group (Figure 1 orange bar) iswell-supported (BS100PP10) comprising only two species S finlayanum AJuss and S puberulumGriseb while the S auriculatum group (Figure 1 purple bar) is moderate to well-supported (B77PP10)comprising three species S angustilobum AJuss S arenicola CEAnderson and S auriculatum(Cav) AJuss The S urenifolium group (Figure 1 light green bar) is weak to well-supported(BS70PP10) represented by only its naming species In contrast the S lalandianum group (Figure 1light blue bar) is moderate to well-supported (BS82PP10) comprising four species S alternifoliumAJuss S bonariense (Hook and Arn) CEAnderson S lalandianum AJuss and S vitifolium AJussThe S sinuatum group (Figure 1 red bar) is well-supported (BS100PP10) represented by only twospecies S lindenianum AJuss and S sinuatum (DC) AJuss while the S blanchetii group (Figure 1rose bar) is weakly-supported (BS-PP060) comprising four species S blanchetii CEAndersonS caatingicola RFAlmeida and Amorim S jatrophifolium AJuss and S salzmannii AJuss Finallythe S gayanum group (Figure 1 gray bar) is weakly to well-supported (BS-PP10) and represented byfour species S cavernulosum CEAnderson S gayanum AJuss S macropodum AJuss and S saxicolaCEAnderson
22 Ancestral Area Reconstruction and Divergence Times Estimation
The S-DEC reconstruction suggests that the most recent common ancestor (MRCA) of Stigmaphyllonwas widespread in the northern portion of the BAFs ca 260 Mya (Figures 2 and 3 node 3 Table 1)A dispersal event (node 3) took place then splitting the MRCA of the Stigmaphyllon paralias groupfrom the remaining species which dispersed from Seasonally Dry Tropical Forests (SDTFs) to CamposRupestres in northeastern Brazil ca 140 Mya (Figures 2 and 3 node 4 Table 1) The MRCA of theS ciliatum and S timoriense (DC) CEAnderson clade diverged ca 220 Mya (node 5) and split intothose lineages ca 100 Mya (node 6) colonizing dunes vegetation on the Atlantic and Australasianrainforests (Figures 2 and 3 node 6 Table 1) The MRCA of core Stigmaphyllon dispersed to thesouthern portion of the BAFs ca 190 Mya (Figures 2 and 3 node 7 Table 1) The MRCA of theS puberulum group arose 625 Mya being widespread over both the Amazon and Atlantic rainforestswith a vicariant event giving rise to its current lineages (Figures 2 and 3 node 8 Table 1) The MRCA ofthe remaining species (node 9) remained distributed in the BAFs ca 170 Mya giving rise to the MRCAof the S auriculatum group ca 94 Mya (node 10) A dispersal event followed by the colonizationof dunes vegetation in Southern Brazil gave rise to the S arenicola lineage (Figures 2 and 3 node 10Table 1) In the Pleistocene ca 072 Mya the MRCA of the S auriculatum and S angustilobum arose inthe BAFs from Southeastern Brazil giving rise to those lineages via a dispersal event from rainforests toSDTFs (Figures 2 and 3 node 11 Table 1) The MRCA of S urenifolium (Figures 2 and 3 node 12 Table 1)
Plants 2020 9 1569 5 of 20
arose ca 150 Mya via a dispersal and a vicariant event from the BAFs to the Cerrado The MRCA ofthe remaining species (Figures 2 and 3 node 13 Table 1) arose ca 130 Mya in the BAFs and probablyin the Amazon forest as well The MRCA of node 14 remained distributed within the Southeasternportion of the BAFs ca 70 Mya (Figures 2 and 3 node 14 Table 1) and diversified into its four mainlineages ca 40 Mya (Figures 2 and 3 nodes 15ndash16 Table 1) Both a dispersal and a vicariant eventtook place ca 120 Mya giving rise to the MRCA of node 17 which was distributed within the BAFsCaatinga and the Amazon rainforest The lineage of node 18 colonized the Amazon rainforest for thefirst time but only diversified ca 05 Mya (Figures 2 and 3 node 18 Table 1)
Figure 2 Chronogram and Statistical-Dispersal-Extinction-Cladogenesis (S-DEC) ancestral areareconstructions for Stigmaphyllon Nodes are numbered from 1 to 26 Numbers above branchesrepresent estimated ages (Mya) Branch letters on the left represent the reconstructed ancestralarea(s) (A) Atlantic rainforest (B) Seasonally Dry Tropical Forest (C) Amazon rainforest (D) Cerrado(E) Australasian rainforests Branch letters on the right represent dispersal (d) or vicariant (v) events
Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 4 of 20
15 trees (consistency index CI = 080 retention index RI = 070) and the strict consensus tree includes17 moderately supported clades (bootstrap percentage BP gt 75 Figure 1) The Bayesian analysisrecovered 25 well-supported to moderately supported clades (posterior probabilities PP gt 095 andgt080 respectively Figure 1)
The monophyly of Stigmaphyllon (Figure 1) is strongly supported by Maximum Parsimony (MP)and Bayesian Inference (BI) analyses being recovered as sister to Diplopterys The clade Stigmaphyllon+ Diplopterys is well supported (BS100PP10) as sister to Bronwenia (Figure 1) Within StigmaphyllonStigmaphyllon subg Stigmaphyllon is recovered as polyphyletic with its representatives placed inthree separate lineages (Figure 1 black bars) named the early-diverging Stigmaphyllon paralias group(Figure 1 dark blue bar) Stigmaphyllon ciliatum group (Figure 1 yellow bar) and core Stigmaphyllon(here represented by the common ancestor that the lineage S fynlayanum + S puberulum share with theremaining species (Figure 1 black bar)) Stigmaphyllon subg Ryssopterys (Figure 1 dark green bar) isrecovered as monophyletic in our analyses being sister to the Stigmaphyllon ciliatum group (Figure 1white bar)
Within core Stigmaphyllon seven major lineages are recovered here designated as 1 S puberulumgroup 2 S auriculatum group 3 S urenifolium group 4 S lalandianum group 5 S sinuatum group6 S blanchetii group and 7 S gayanum group The S puberulum group (Figure 1 orange bar) iswell-supported (BS100PP10) comprising only two species S finlayanum AJuss and S puberulumGriseb while the S auriculatum group (Figure 1 purple bar) is moderate to well-supported (B77PP10)comprising three species S angustilobum AJuss S arenicola CEAnderson and S auriculatum(Cav) AJuss The S urenifolium group (Figure 1 light green bar) is weak to well-supported(BS70PP10) represented by only its naming species In contrast the S lalandianum group (Figure 1light blue bar) is moderate to well-supported (BS82PP10) comprising four species S alternifoliumAJuss S bonariense (Hook and Arn) CEAnderson S lalandianum AJuss and S vitifolium AJussThe S sinuatum group (Figure 1 red bar) is well-supported (BS100PP10) represented by only twospecies S lindenianum AJuss and S sinuatum (DC) AJuss while the S blanchetii group (Figure 1rose bar) is weakly-supported (BS-PP060) comprising four species S blanchetii CEAndersonS caatingicola RFAlmeida and Amorim S jatrophifolium AJuss and S salzmannii AJuss Finallythe S gayanum group (Figure 1 gray bar) is weakly to well-supported (BS-PP10) and represented byfour species S cavernulosum CEAnderson S gayanum AJuss S macropodum AJuss and S saxicolaCEAnderson
22 Ancestral Area Reconstruction and Divergence Times Estimation
The S-DEC reconstruction suggests that the most recent common ancestor (MRCA) of Stigmaphyllonwas widespread in the northern portion of the BAFs ca 260 Mya (Figures 2 and 3 node 3 Table 1)A dispersal event (node 3) took place then splitting the MRCA of the Stigmaphyllon paralias groupfrom the remaining species which dispersed from Seasonally Dry Tropical Forests (SDTFs) to CamposRupestres in northeastern Brazil ca 140 Mya (Figures 2 and 3 node 4 Table 1) The MRCA of theS ciliatum and S timoriense (DC) CEAnderson clade diverged ca 220 Mya (node 5) and split intothose lineages ca 100 Mya (node 6) colonizing dunes vegetation on the Atlantic and Australasianrainforests (Figures 2 and 3 node 6 Table 1) The MRCA of core Stigmaphyllon dispersed to thesouthern portion of the BAFs ca 190 Mya (Figures 2 and 3 node 7 Table 1) The MRCA of theS puberulum group arose 625 Mya being widespread over both the Amazon and Atlantic rainforestswith a vicariant event giving rise to its current lineages (Figures 2 and 3 node 8 Table 1) The MRCA ofthe remaining species (node 9) remained distributed in the BAFs ca 170 Mya giving rise to the MRCAof the S auriculatum group ca 94 Mya (node 10) A dispersal event followed by the colonizationof dunes vegetation in Southern Brazil gave rise to the S arenicola lineage (Figures 2 and 3 node 10Table 1) In the Pleistocene ca 072 Mya the MRCA of the S auriculatum and S angustilobum arose inthe BAFs from Southeastern Brazil giving rise to those lineages via a dispersal event from rainforests toSDTFs (Figures 2 and 3 node 11 Table 1) The MRCA of S urenifolium (Figures 2 and 3 node 12 Table 1)
Plants 2020 9 1569 5 of 20
arose ca 150 Mya via a dispersal and a vicariant event from the BAFs to the Cerrado The MRCA ofthe remaining species (Figures 2 and 3 node 13 Table 1) arose ca 130 Mya in the BAFs and probablyin the Amazon forest as well The MRCA of node 14 remained distributed within the Southeasternportion of the BAFs ca 70 Mya (Figures 2 and 3 node 14 Table 1) and diversified into its four mainlineages ca 40 Mya (Figures 2 and 3 nodes 15ndash16 Table 1) Both a dispersal and a vicariant eventtook place ca 120 Mya giving rise to the MRCA of node 17 which was distributed within the BAFsCaatinga and the Amazon rainforest The lineage of node 18 colonized the Amazon rainforest for thefirst time but only diversified ca 05 Mya (Figures 2 and 3 node 18 Table 1)
Figure 2 Chronogram and Statistical-Dispersal-Extinction-Cladogenesis (S-DEC) ancestral areareconstructions for Stigmaphyllon Nodes are numbered from 1 to 26 Numbers above branchesrepresent estimated ages (Mya) Branch letters on the left represent the reconstructed ancestralarea(s) (A) Atlantic rainforest (B) Seasonally Dry Tropical Forest (C) Amazon rainforest (D) Cerrado(E) Australasian rainforests Branch letters on the right represent dispersal (d) or vicariant (v) events
Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 5 of 20
arose ca 150 Mya via a dispersal and a vicariant event from the BAFs to the Cerrado The MRCA ofthe remaining species (Figures 2 and 3 node 13 Table 1) arose ca 130 Mya in the BAFs and probablyin the Amazon forest as well The MRCA of node 14 remained distributed within the Southeasternportion of the BAFs ca 70 Mya (Figures 2 and 3 node 14 Table 1) and diversified into its four mainlineages ca 40 Mya (Figures 2 and 3 nodes 15ndash16 Table 1) Both a dispersal and a vicariant eventtook place ca 120 Mya giving rise to the MRCA of node 17 which was distributed within the BAFsCaatinga and the Amazon rainforest The lineage of node 18 colonized the Amazon rainforest for thefirst time but only diversified ca 05 Mya (Figures 2 and 3 node 18 Table 1)
Figure 2 Chronogram and Statistical-Dispersal-Extinction-Cladogenesis (S-DEC) ancestral areareconstructions for Stigmaphyllon Nodes are numbered from 1 to 26 Numbers above branchesrepresent estimated ages (Mya) Branch letters on the left represent the reconstructed ancestralarea(s) (A) Atlantic rainforest (B) Seasonally Dry Tropical Forest (C) Amazon rainforest (D) Cerrado(E) Australasian rainforests Branch letters on the right represent dispersal (d) or vicariant (v) events
Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 6 of 20
Figure 3 Times of dispersalvicariance events leading to biome shifting in Stigmaphyllon (blue) Atlanticrainforest (purple) Seasonally Dry Tropical Forest (orange) Amazon Rainforest (red) CerradoGreen circles represent the original ancestral populations of Stigmaphyllon Dark circles representpopulations of Stigmaphyllon colonizing new biomes over different time periods
Table 1 Divergence times estimates (maximummeanminimum) ancestral area reconstructions anddispersalvicariance events reconstructed for all nodes in this study
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
1 400 3995 350 AtlanticForestCaatinga Dispersal
2 4129 3639 2992 AtlanticForestCaatinga Dispersal
3 3409 2647 1935 Atlantic Forest Dispersal4 2214 1406 676 Caatinga Dispersal5 3014 2241 1543 Atlantic Forest Dispersal
6 1987 1018 218 Atlantic ForestAsianRainforests Vicariance
7 2614 1919 1271 Atlantic Forest Dispersal
8 1235 625 106AtlanticForestCaatingaAmazonForest
Vicariance
9 2405 1745 1164 Atlantic Forest -10 1622 94 318 Atlantic Forest Dispersal
Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 7 of 20
Table 1 Cont
Nodes Max AgeMean Min Ancestral Area
ReconstructionDispersalVicarianceEvent
11 181 072 003 AtlanticForestCaatinga Dispersal
12 2140 1521 1006 Atlantic Forest DispersalVicariance13 1830 1289 825 Atlantic Forest -14 1216 745 330 Atlantic Forest -15 870 457 142 Atlantic Forest -16 829 439 095 Atlantic Forest -17 1719 1201 732 Atlantic Forest DispersalVicariance18 184 054 00 Amazon Forest -19 160 1109 688 Atlantic Forest Dispersal20 1305 852 443 Atlantic Forest -21 1087 673 304 Atlantic Forest Dispersal22 849 481 128 Atlantic Forest -
23 1349 898 488 AtlanticForestCaatinga Dispersal
24 036 030 010 Caatinga -
25 1035 606 227 AtlanticForestCaatinga Dispersal
26 086 027 00 AtlanticForestCaatinga Dispersal
23 Meta-Analysis
We identified 113 genera of fernslycophytes gymnosperms magnoliids monocots and eudicotscomprising lineages exclusively diversified in the BAF biome (out of a total of 2224 genera currentlyrecorded by the Flora do Brasil Project Table 2) Only 64 of those genera have estimated diversificationages available from 34 phylogenetic studies published from 2004 to 2020 (Figure 4 Table 2) Most ofthose studies were published from 2015 to 2020 when molecular clocks and time-calibrated trees werealready widespread in phylogenetic literature
Table 2 Age of Brazilian Atlantic Rainforests (BAF) lineages of vascular plants based on phylogeneticliterature and distribution data from Flora do Brasil [13] and Plants of the World Online [17] refers toages unavailable from the consulted literature
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Ferns
Cyatheaceae Cyathea Sm 23290 Rainforest 300 [18]
Gymnosperms
Araucariaceae Araucaria Juss 120 Rainforest 100 [19]
Podocarpaceae Podocarpus LrsquoHeacuterex Pers 2115 Rainforest 150 [20]
Magnoliids
Annonaceae Hornschuchia Nees 1010 Rainforest 200 [21]
Lauraceae PhyllostemonodaphneKoesterm 11 Rainforest -
Lauraceae UrbanodendronMez 33 Rainforest -
Monimiaceae MacropeplusPerkins 44 Rainforest 1126 [22]
MonimiaceaeGrazielanthusPeixoto andPer-Moura
11 Rainforest 385 [22]
Monimiaceae HennecartiaJPoiss 11 Rainforest 1558 [22]
Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 8 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Magnoliids
Monimiaceae MacrotorusPerkins 11 Rainforest 1126 [22]
Monimiaceae Mollinedia Ruizand Pav 3255 Rainforest 220 [22]
Monocots
Amaryllidaceae Griffinia Ker Gawl 1722 Grassland -
Araceae Asterostigma Fischand CA Mey 810 Rainforest -
Araceae Dracontioides Engl 22 Rainforest -
Asparagaceae Herreria Ruiz andPav 68 Rainforest -
BromeliaceaeAlcantarea(EMorren ex Mez)Harms
3040 Rainforest 55 33 15 [23]
Bromeliaceae AraeococcusBrongn 69 Rainforest 35 [24]
Bromeliaceae Billbergia Thunb 3563 Rainforest 45 [24]
Bromeliaceae Canistropsis (Mez)Leme 1112 Rainforest 35 [24]
BromeliaceaeStigmatodon LemeGKBr andBarfuss
1818 Rainforest 64 55 28 [23]
Bromeliaceae Vriesea Lindl 167255 Rainforest 68 50 33 [23]Commelinaceae Siderasis Raf 66 Rainforest 1669 857 226 [25]
Commelinaceae DichorisandraJCMikan 4052 Rainforest 638 278 032 [25]
Dioscoreaceae Dioscorea L 81628 Rainforest 300 220 150 [26]
Iridaceae NeomaricaSprague 2727 Grassland 65 [27]
Marantaceae Ctenanthe Eichler 1115 Rainforest -Marantaceae Maranta L 2037 Rainforest -Marantaceae Saranthe Eichler 810 Rainforest -Marantaceae Thalia L 46 Rainforest -Orchidaceae Bifrenaria Lindl 1721 Rainforest 130 [28]
Orchidaceae CapanemiaBarbRodr 69 Rainforest -
Orchidaceae CentroglossaBarbRodr 66 Rainforest -
Orchidaceae Cirrhaea Lindl 77 Rainforest -Orchidaceae Hoehneella Ruschi 22 Rainforest -
Orchidaceae Isabelia BarbRodr 33 Rainforest -
Orchidaceae Lankesterella Ames 711 Rainforest -
Orchidaceae LoefgrenianthusHoehne 11 Rainforest -
Orchidaceae Miltonia Lindl 1919 Rainforest -
Orchidaceae PhymatidiumLindl 99 Rainforest -
Orchidaceae Pseudolaelia Portoand Brade 1015 Rainforest -
Poaceae Chusquea Kunth 45185 Rainforest 90 [29]
Poaceae MerostachysSpreng 4453 Rainforest -
Poaceae Olyra L 915 Rainforest 140 [30]Poaceae Raddia Bertol 912 Rainforest 220 [30]
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-
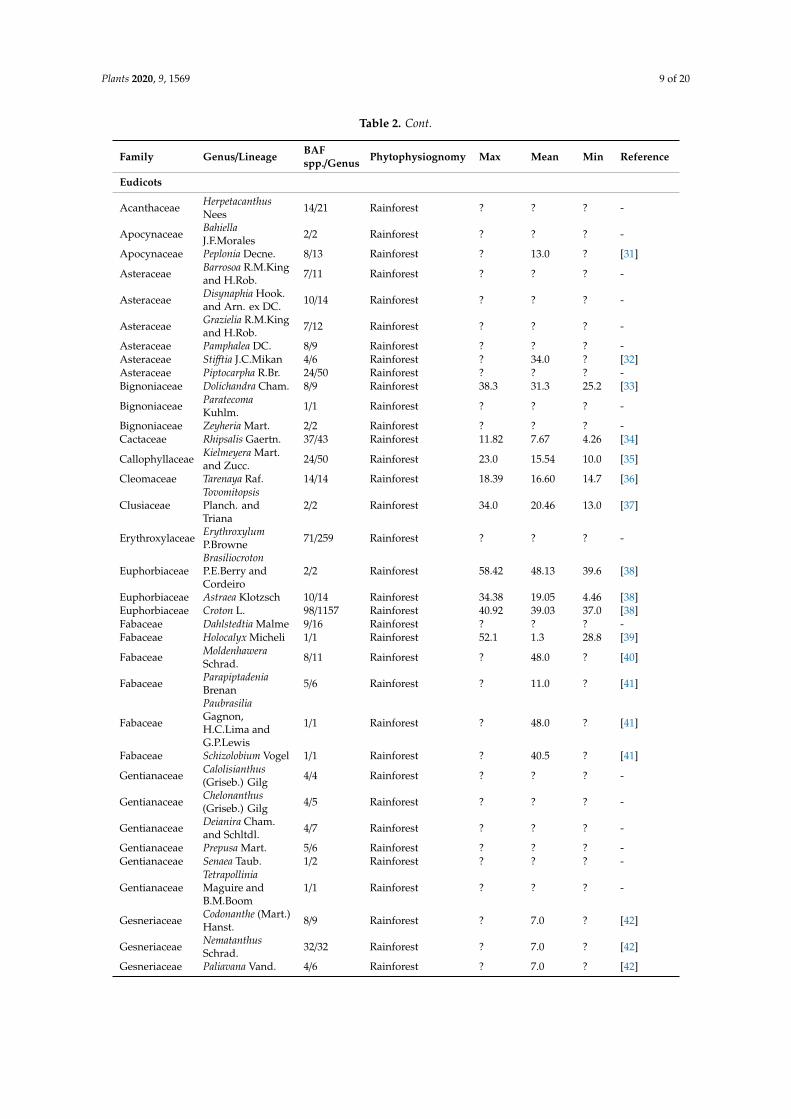
Plants 2020 9 1569 9 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Acanthaceae HerpetacanthusNees 1421 Rainforest -
Apocynaceae BahiellaJFMorales 22 Rainforest -
Apocynaceae Peplonia Decne 813 Rainforest 130 [31]
Asteraceae Barrosoa RMKingand HRob 711 Rainforest -
Asteraceae Disynaphia Hookand Arn ex DC 1014 Rainforest -
Asteraceae Grazielia RMKingand HRob 712 Rainforest -
Asteraceae Pamphalea DC 89 Rainforest -Asteraceae Stifftia JCMikan 46 Rainforest 340 [32]Asteraceae Piptocarpha RBr 2450 Rainforest -Bignoniaceae Dolichandra Cham 89 Rainforest 383 313 252 [33]
Bignoniaceae ParatecomaKuhlm 11 Rainforest -
Bignoniaceae Zeyheria Mart 22 Rainforest -Cactaceae Rhipsalis Gaertn 3743 Rainforest 1182 767 426 [34]
Callophyllaceae Kielmeyera Martand Zucc 2450 Rainforest 230 1554 100 [35]
Cleomaceae Tarenaya Raf 1414 Rainforest 1839 1660 147 [36]
ClusiaceaeTovomitopsisPlanch andTriana
22 Rainforest 340 2046 130 [37]
Erythroxylaceae ErythroxylumPBrowne 71259 Rainforest -
EuphorbiaceaeBrasiliocrotonPEBerry andCordeiro
22 Rainforest 5842 4813 396 [38]
Euphorbiaceae Astraea Klotzsch 1014 Rainforest 3438 1905 446 [38]Euphorbiaceae Croton L 981157 Rainforest 4092 3903 370 [38]Fabaceae Dahlstedtia Malme 916 Rainforest -Fabaceae Holocalyx Micheli 11 Rainforest 521 13 288 [39]
Fabaceae MoldenhaweraSchrad 811 Rainforest 480 [40]
Fabaceae ParapiptadeniaBrenan 56 Rainforest 110 [41]
Fabaceae
PaubrasiliaGagnonHCLima andGPLewis
11 Rainforest 480 [41]
Fabaceae Schizolobium Vogel 11 Rainforest 405 [41]
Gentianaceae Calolisianthus(Griseb) Gilg 44 Rainforest -
Gentianaceae Chelonanthus(Griseb) Gilg 45 Rainforest -
Gentianaceae Deianira Chamand Schltdl 47 Rainforest -
Gentianaceae Prepusa Mart 56 Rainforest -Gentianaceae Senaea Taub 12 Rainforest -
GentianaceaeTetrapolliniaMaguire andBMBoom
11 Rainforest -
Gesneriaceae Codonanthe (Mart)Hanst 89 Rainforest 70 [42]
Gesneriaceae NematanthusSchrad 3232 Rainforest 70 [42]
Gesneriaceae Paliavana Vand 46 Rainforest 70 [42]
Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 10 of 20
Table 2 Cont
Family GenusLineage BAFsppGenus Phytophysiognomy Max Mean Min Reference
Eudicots
Gesneriaceae Sinningia Nees 7075 Rainforest 140 [42]Gesneriaceae Vanhouttea Lem 910 Rainforest 70 [42]
Loasaceae BlumenbachiaSchrad 712 Rainforest 422 275 207 [43]
MalpighiaceaeBarnebya (Griseb)WRAndersonand BGates
12 Rainforest 620 [44]
Melastomataceae Behuria Cham 1717 Rainforest 850 [45]Melastomataceae Bertolonia Raddi 2727 Rainforest 926 [45]Melastomataceae Huberia DC 1317 Rainforest 850 [45]
MelastomataceaePhyseterostemonRGoldenb andAmorim
55 Rainforest 250 [45]
Melastomataceae Pleiochiton Naudinex A Gray 1212 Rainforest 443 [45]
Melastomataceae Pleroma DDon 4685 Rainforest 1226 [45]
Moraceae Clarisia Ruiz andPav 22 Rainforest -
Moraceae Ficus L 38874 Rainforest 250 [46]Myrtaceae Accara Landrum 11 Rainforest 320 [47]
Myrtaceae BlepharocalyxOBerg 34 Rainforest 400 [47]
Myrtaceae Calyptranthes Sw 3535 Rainforest 240 [47]
Myrtaceae Curitiba Salywonand Landrum 11 Rainforest 400 [47]
Myrtaceae Eugenia L 2541149 Rainforest 440 [47]
Myrtaceae MyrceugeniaOBerg 3345 Rainforest 250 [47]
Myrtaceae Myrcia DC 200609 Rainforest 280 [48]Myrtaceae Myrciaria OBerg 1827 Rainforest 260 [48]Myrtaceae Myrrhinium Schott 11 Rainforest 290 [47]
Myrtaceae NeomitranthesDLegrand 1515 Rainforest 120 [47]
Myrtaceae Plinia L 2978 Rainforest 370 [47]Myrtaceae Psidium L 3991 Rainforest 200 [47]Rubiaceae Bathysa C Presl 610 Rainforest -
Rubiaceae Bradea Standl exBrade 66 Rainforest -
Rubiaceae CoccocypselumPBrowne 1422 Rainforest -
Rutaceae ConchocarpusJCMikan 4048 Rainforest -
Rutaceae MetrodoreaASt-Hil 36 Rainforest -
RutaceaeNeoraputiaEmmerich exKallunki
56 Rainforest -
Sapindaceae Cardiospermum L 610 Rainforest 230 [49]
Sapindaceae Thinouia Trianaand Planch 711 Rainforest -
Solanaceae Petunia Juss 1016 Grassland 115 849 55 [50]Vochysiaceae Callisthene Mart 88 Rainforest 300 220 140 [51]
Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 11 of 20
Figure 4 Box plots summarizing the diversification of vascular plants through time in the BrazilianAtlantic Rainforest based on data presented in Table 2 (blue) Eudicots (orange) Monocots(gray)ndashMagnoliids (yellow) Gymnosperms (light blue) Ferns
3 Discussion
31 Phylogenetics of Stigmaphyllon
The topology recovered from the combined dataset (ie ribosomal + nuclear markers) evidencedthat the subgenera of Stigmaphyllon proposed by Anderson [10] are paraphyletic All previousphylogenetic studies of Malpighiaceae sampled mostly Mesoamerican and Amazonian species of thegenus [235944] making it difficult to properly test the monophyly of its subgenera The only BAFspecies of Stigmaphyllon sampled in previous studies were S ciliatum and S paralias However in allthese studies S paralias (a Brazilian Atlantic Rainforest lineage) is consistently recovered as sister to allremaining lineages of Stigmaphyllon Additionally this is the first time S auriculatum the type speciesof the genus is included in a phylogenetic study On the other hand our results highly corroborate theprevious topologies recovered for Stigmaphyllon with the S paralias group recovered as the first lineageto diverge followed by the clade comprising the S ciliatum group + S subg Ryssopterys sister to a largeclade consisting of species from the S tomentosum group (core Stigmaphyllon) [29] Additional speciessampling allied to a thorough morphological study on a phylogenetic perspective in Stigmaphyllon isurgently required to shed some light on its infrageneric classification
32 Divergence Times and Biogeography of Stigmaphyllon
Our divergence times estimation for the MRCA of Stigmaphyllon (ca 265 Mya) is similar to the ageof 225 Mya estimated by Davis et al [44] and of 210 Mya estimated by Willis et al [9] The estimatedages for the MRCAs obtained by us for the S paralias group and the S ciliatum + S subg Ryssopterysclade strongly corroborates those previously reported by Willis et al [9] (170 150 and 100 Myarespectively) The only exception was the age of 190 Mya estimated by us for core-Stigmaphyllon whenWillis et al [9] recovered the estimation of 100 Mya for this clade This apparent difference in ageestimates might be due to the more comprehensive sampling of this clade in the present study
Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 12 of 20
Over these last 260 Mya the lineages of Stigmaphyllon went through 13 dispersals and fourvicariance events (VE) Worth noting is that VEs in this genus were always followed by a biomeshifting event (BSE) from the BAFs to another tropical biome The first VE followed by a BSE inStigmaphyllon occurred ca 150 Mya in the MRCA of S urenifolium from the BAFs to the BrazilianCerrado Several plant lineages present the same diversification pattern with older ancestors arisingin the BAFs and colonizing the Cerrado biome ca 15 Mya such as clade 5 of Amphilophium Kunth(Bignoniaceae) [52] Astraea Klotzsch (Euphorbiaceae) [38] Dolichandra Cham (Bignoniaceae) [33]Fridericia Mart (Bignoniaceae) [52] and Xylophragma Sprague (Bignoniaceae) [53] Dispersal eventsfrom forest to open habitats is one of the main factors explaining richness in Neotropical biodiversity [16]Even though in Stigmaphyllon those dispersal events occurred mostly among forested biomes its singleCerrado lineage seems to point to an older diversification of this biome such as in those fromVochysiaceae (200ndash150 Mya) [51] Few species of Stigmaphyllon successfully colonized the Cerradobiome such as S jobertii S occidentale S tomentosum and S urenifolium Future studies samplingthose species in the molecular phylogeny of Stigmaphyllon are crucial to test if the genus colonizedthe Cerrado biome more than a single time However several studies seem to present the samepattern pointed by us of forest lineages occupying and diversifying in the Cerrado biome [16] with theopposite rarely recorded in biogeographic studies We hypothesize that Cerrado lineages colonizingforested biomes might be rare due to the recent diversification of several plant lineages of Neotropicalsavannas [54]
The second VE followed by a BSE in Stigmaphyllon occurred ca 120 Mya in the MRCA ofS sinuatum group from the BAFs to the Amazon rainforest The same pattern of diversification wasrecorded in other plant lineages with Atlantic rainforest MRCAs colonizing the Amazon rainforest atthis time such as Eugenia L clade G [55] At least 50 dispersal events occurred between the Atlanticand Amazon rainforests [16] The number of lineage exchanges between these biomes fluctuated overthe last 600 Mya with its previous increase starting ca 120 Mya and peaking ca 60ndash30 Mya [16]corroborating our results with Stigmaphyllon The MRCA of S finlayanum group was the fourth VEfollowed by a BSE occurring in the genus ca 630 Mya from the BAFs to the Amazon rainforestThe same pattern of diversification was recorded in other plant lineages with Atlantic rainforestMRCAs colonizing the Amazon rainforest at this time such as the clade 3 of Amphilophium Kunth(Bignoniaceae) [52] and the abovementioned study by Antonelli et al [16]
The third VE followed by a BSE in Stigmaphyllon occurred ca 100 Mya in the MRCA of S subgRyssopterys + S ciliatum group from the BAFs to the Asian rainforests This lineage diversifiedca 220 Mya so we hypothesize that its MRCA was widely distributed among dunes vegetation inthe Atlantic and Asian rainforests via the Antarctic route Antarcticarsquos glaciation process startedca 340 Mya during the EoceneOligocene transition on high altitude regions of this continent [56]From 340ndash120 Mya Antarctica had intermittent ice sheet coverage leaving intact Tertiary pockets ofpre-glaciation fauna and flora that only went completely extinct ca 120 Mya with the formation ofpermanent ice sheets in this continent [57] the same confidence interval recovered in our study for theMRCA of S subg Ryssopterys + S ciliatum group The same pattern of vicariance event is observed inthe Drymophila (Australian) + Luzuriaga (South American) clade (Alstromeriaceae) in which its MRCAsplit ca 230 Mya giving rise to these genera from ca 100 to 40 Mya [58]
On the other hand 14 dispersal events (DE) occurred within Stigmaphyllon mostly within theNorth-South corridors of the BAFs and at least three DEs from the BAFS to the Caatinga biomeFrom these latter three DEs the first occurred ca 140 Mya in the MRCA of S paralias group the secondoccurred ca 90 Mya in the MRCA of S saxicola group and the third occurred ca 10 Mya in theMRCA of the S auriculatum group From the remaining DEs that happened within the BAFs at leastfive of them occurred from North to South ca 220 170 120 110 and 90 Mya The same patternwith Northern BAF lineages colonizing Southern portions of this biome is observed in Aechmea(Bromeliaceae) [59] However the North-South dispersals in this genus started only ca 60 Mya(Bromeliaceae) [59] being much younger than those in Stigmaphyllon
Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 13 of 20
33 Time and Diversification of the Atlantic Rainforest
Several implications for understanding the BAFs historical biogeography might be postulatedfrom the biogeographical study of Stigmaphyllon Our results suggest a late-Eocene origin for theseforests that seems to be partially corroborated by the literature Until the late Paleocene and earlyEocene the Earthrsquos climate was mostly warmer and more humid than today suggesting that SouthAmerica was probably covered by continuous rainforests [6061] When comparing the mean agesrecovered in published studies for several lineages of ferns gymnosperms and angiosperms diversifiedin the BAFs we bring new evidence that vascular plants started colonizing these forests over thelast 600 Mya (Figure 4 Table 2) corroborating the abovementioned authors The oldest lineageto occupy the BAFs might have been the genus Barnebya WRAnderson and BGates ca 60 Mya(Malpighiaceae Eudicots) with the diversification of most Eudicot lineages in these forests occurringfrom 400 to 150 Mya (Figure 4 Table 2) During the late Eocene and Oligocene global episodesof cooling and dryness favored the expansion of grasslands in the southern and central regionsof the continent [6062] which culminated in the formation of a diagonal belt of more open anddrier biomes (also known as ldquodry diagonalrdquo) [63] The formation of the dry diagonal marked theformation of the Atlantic forest in the east and Amazonia in the west [64] On the other handthe colonization and diversification of Magnoliid lineages took place from 180 to 30 Mya (Figure 4Table 2) followed by gymnosperm lineages that diversified from 150 to 110 Mya and finally frommonocot lineages diversifying from 120 to 30 Mya in these rainforests (Figure 4 Table 2) Fossil recordsand paleoclimate studies suggest that the BAFs and the Amazon rainforests were re-connected multipletimes in the Miocene and Pliocene [64] Mean ages regarding the colonization and diversificationof ferns in the BAFs are still incipient with a single study on the family Cyatheaceae evidencing itsinitial diversification in these forests at 300 Mya (Figure 4 Table 2) Additionally from the 113 BAFlineages presented in Table 2 only 64 show mean ages based on time-calibrated phylogenies availablein the literature At least 18 families of angiosperms (Acanthaceae Amaryllidaceae ApocynaceaeAraceae Asparagaceae Asteraceae Bignoniaceae Erythroxylaceae Fabaceae Gentianaceae LauraceaeMarantaceae Moraceae Orchidaceae Poaceae Rubiaceae Rutaceae and Sapindaceae) with lineagesdiversified in the Atlantic forest still lack published time-calibrated phylogenies
Another worth mentioning factor that might have played an important role in the diversificationof the BAFs was the uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges in EasternBrazil Those mountains were previously thought to have uplifted around 1200 Mya but geologicalstudies from the past decade have pointed to an earlier uplifting age for these mountains from 600 to300 Mya [65] The tertiary uplift age of these mountains in Eastern Brazil was a direct result of theAndean uplift coinciding with our results for the colonization of the BAFs by 64 lineages of vascularplants [65]
4 Material and Methods
41 Taxon Sampling and Plant Material
We sampled 24 species of Stigmaphyllon (ca 14 of the Neotropicsrsquo genus diversity 18 spp from the
Brazilian Atlantic Rainforest (out of 31 spp) three spp from the Amazon Rainforest two spp fromthe Caatinga and one spp from the Cerrado biomes) including outgroups Bronwenia WRAndersonand CCDavis and Diplopterys AJuss From this total 23 species represent S subg Stigmaphyllon anda single species [Stigmaphyllon timoriense (DC) CEAnderson] represents S subg Ryssopterys (Table 3)For DNA extraction we used mainly field-prepared silica dried leaves (12ndash80 mg) and herbariumspecimens as necessary (Table 3)
Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 14 of 20
Table 3 Species and DNA regions sampled in this study Genbank accession numbers are presented forcolumns ETS (External Transcribed Spacer) ITS (Internal Transcribed Spacer) and PHYC (PhytochromeC gene) Sequences were obtained from GenBank
SpeciesVoucher(HerbariumAcronym)
ETS ITS PHYC
Bronwenia cinerascens (Benth)WRAnderson and CCDavis Nee 48 658 (SP) KR0545861 HQ2468211 HQ2469871
Diplopterys pubipetala (AJuss)WRAnderson and CCDavis Francener 1126 (SP) KR092986 HQ2468211 HQ2470451
Stigmaphyllon alternifolium AJuss Almeida 501 (SP) KR0546121 - -Stigmaphyllon angustilobum AJuss Almeida 503 (SP) KR0545911 MT559811 -Stigmaphyllon arenicolaCEAnderson Sebastiani 5 (SP) KR0545891 - -
Stigmaphyllon auriculatum (Cav)AJuss
Almeida 584(HUEFS) KR0545921 MT559812 -
Stigmaphyllon blanchetiiCEAnderson Almeida 525 (SP) KR0546151 MT559813 -
Stigmaphyllon bonariense AJuss Queiroz 13 530(HUEFS) KR0546071 - -
Stigmaphyllon caatingicola 1RFAlmeida and Amorim
Almeida 577(HUEFS) KR0545951 MT559814 -
Stigmaphyllon caatingicola 2RFAlmeida and Amorim
Almeida 629(HUEFS) MT490608 MT559815 -
Stigmaphyllon cavernulosumCEAnderson
Cardoso 2083(HUEFS) KR0546091 - -
Stigmaphyllon ciliatum (Lam)AJuss Almeida 541 (SP) KR0545901 - HQ2471511
Stigmaphyllon finlayanum AJuss Pace 457 (SPF) KR0545991 - HQ2471521 Stigmaphyllon gayanum AJuss Almeida 500 (SP) KR0546101 - -Stigmaphyllon harleyiCEAnderson Santos 378 (HUEFS) KR0545941 MT581513 -
Stigmaphyllon jatrophifolium AJuss Filho sn(SP369102) KR0546141 - -
Stigmaphyllon lalandianum AJuss Kollmann 4279(CEPEC) KR0545961 - -
Stigmaphyllon lindenianum AJuss Aguillar 718 (SP) KR0546031 - HQ2471531 Stigmaphyllon macropodum AJuss Almeida 538 (SP) KR0546021 MT559816 -Stigmaphyllon paralias AJuss Almeida 509 (SP) KR0545931 KY4219091 AF5005661 Stigmaphyllon puberulum AJuss Perdiz 732 (HUEFS) KR0546001 - -Stigmaphyllon salzmannii AJuss Almeida 526 (SP) KR0546161 MT559817 -Stigmaphyllon saxicolaCEAnderson
Badini 24261(HUEFS) KR0546051 MT559818 -
Stigmaphyllon sinuatum (DC)AJuss
Amorim 3159(CEPEC) KR0546081 - -
Stigmaphyllon timoriense (DC)CEAnderson Anderson 796 (US) - - AF5005451
Stigmaphyllon urenifolium AJuss Guedes 13932(HUEFS) KR0546041 - -
Stigmaphyllon vitifolium AJuss DalCol 233(HUEFS) KR0545981 - -
42 Molecular Protocols
Genomic DNA was extracted using the 2 times CTAB protocol modified from Doyle and Doyle [66]Three DNA regions (nuclear PHYC gene and the ribosomal external and internal transcribed spacers(ETS and ITS)) were selected based on their variability in previous Malpighiaceae studies [256867]Protocols to amplify and sequence ETS and ITS followed Almeida et al [8] For amplification we usedthe TopTaq (Qiagen) mix following the manufacturerrsquos standard protocol with the addition of betaine(10 M final concentration) and 2 DMSO for the ETS region PCR products were purified using PEG(polyethylene glycol) 11 and sequenced directly with the same primers used for PCR amplification
Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 15 of 20
Sequence electropherograms were produced on an automatic sequencer (ABI 3130XL genetic analyzer)using the Big Dye Terminator 31 kit (Applied Biosystems) Additional sequences for PHYC wereretrieved from GenBank (Table 3) Newly generated sequences were edited using the Geneioussoftware [68] and all datasets were aligned using Muscle [69] with subsequent adjustments in thepreliminary matrices made by eye The complete data matrices are available at TreeBase (accessionnumber S21218) This study was authorized by the Genetic Heritage and Associated TraditionalKnowledge Management National System of Brazil (SISGEN A3B8F19)
43 Phylogenetic Analysis
Analyses were rooted in Bronwenia according to Davis and Anderson [2] Individual analysesfor each marker were performed and since no significant incongruencies were found analyses ofcombined matrices (ie nuclear + ribosomal markers) were performed using maximum parsimony(MP) conducted with PAUP 40b10a [70] A heuristic search was performed using TBR swapping(tree-bisection reconnection) and 1000 random taxon-addition sequence replicates with TBR swappinglimited to 15 trees per replicate to prevent extensive searches (swapping) in suboptimal islandsfollowed by TBR in the resulting trees with a limit of 1000 trees In all analyses the characters wereequally weighted and unordered [71] Relative support for individual nodes was assessed usingnon-parametric bootstrapping [72] with 1000 bootstrap pseudoreplicates TBR swapping simple taxonaddition and a limit of 15 trees per replicate
For the model-based approach we selected the model GTR + I + G using hierarchical likelihoodratio tests (HLRT) on J Modeltest 2 [73] A Bayesian analysis (BA) was conducted with mixed modelsand unlinked parameters using MrBayes 312 [74] The Markov chain Monte Carlo (MCMC) analysiswas performed using two simultaneous independent runs with four chains each (one cold and threeheated) saving one tree every 1000 generations for a total of ten million generations We excluded aslsquoburn-inrsquo trees from the first two million generations and tree distributions were checked for a stationaryphase of likelihood The posterior probabilities (PP) of clades were based on the majority-rule consensusproduced with the remaining trees in MrBayes 312 [74]
44 Calibration
Estimates were conducted based on a simplified ultrametric Bayesian combined tree generatedwith BEAST 184 [75] This analysis used a relaxed uncorrelated lognormal clock and Yule processspeciation prior to inferring trees The calibration parameters were based on previous estimatesderived from a comprehensive fossil-calibrated study of the whole Malpighiaceae [4476] We optedfor calibrating at the root using a normal prior with mean initial values of 400 Mya (representing theage estimated for the MRCA of the Stigmaphylloid clade) and a standard deviation of 10 [446776]Two separate and convergent runs were conducted with 10000000 generations sampling every1000 steps and 2000 trees as burn-in We checked for ESS values higher than 400 for all parameters onTracer 16 [77] Tree topology was assessed using TreeAnnotator and FigTree 140 [78]
45 Ancestral Area Reconstruction
Species distribution data were compiled from the taxonomic revision of Bronwenia [79]Diplopterys [80] and Stigmaphyllon [1011] (Figure 3) Occurrences were categorized according toa modified version of the biome domains adopted by IBGE [81] and WWF [82] which reflectsdistribution patterns in Stigmaphyllon namely (A) Brazilian Atlantic Rainforest (B) Seasonally DryTropical Forest [Caatinga] (C) Amazon Rainforest (D) Cerrado and (E) Australasian Rainforests(Figure 2) Ancestral areas of Stigmaphyllon and its relatives were estimated using a maximum likelihoodanalysis of geographic range evolution using software Rasp 32 [83] Estimates were conducted usingthe statistical dispersal-extinction-cladogenesis (S-DEC) [84] according to the parameters proposed byRee and Sanmartiacuten [85]
Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 16 of 20
46 Meta-Analysis
Ages of BAF lineages of vascular plants (fernslycophytes gymnosperms magnoliids monocotsand eudicots) were compiled based on the phylogenetic literature and distribution data from Florado Brasil [12] and Plants of the World Online [17] Online repositories such as GBIF BIEN andSpecies Link were not used since specimen identification is not usually updated or are not identifiedby a Malpighiaceae specialist Additionally the main problem in using all the above-mentionedrepositories is that only Flora do Brasil present reliable information on the biome distribution of speciessampled in our study We considered a genus or lineage within a genus diversified in the BAF when atleast 50 of its total number of species occurred in this biome Data from BAF lineages of vascularplant species are presented in Table 2 alongside estimated ages and references A boxplot graphicis also presented in Figure 4 showing the diversification of vascular plants through time in the BAFbased on data presented in Table 2
5 Conclusions
Even though dispersal events from forested to open habitats have been recently identified asone of the main factors explaining richness in Neotropical biodiversity [16] the same pattern wasnot recovered for Stigmaphyllon A late-Eocene origin for this genus is suggested with its MRCAoriginating in the Northeastern BAFs with several dispersal events taking place to other Neo- andPaleotropical biomes from 220 to 10 Mya alongside several dispersals from Northern to Southernportions of the BAFs When comparing our results with published divergence times for BAFsrsquo vascularplant lineages a late-Eocene origin for these forests was evidenced The immense gap in time-calibratedphylogenies focusing on BAFsrsquo vascular plant lineages is still the most significant impediment for a morecomprehensive understanding of the plant diversification timeframe in these forests Additionallythe recent evidenced tertiary uplift of Serra do Mar and Serra da Mantiqueira Mountain Ranges mightalso have played an important role in the diversification of the megadiverse BAFs
Author Contributions Conceptualization RFdA methodology RFdA and CvdB software RFdAvalidation RFdA and CvdB formal analysis RFdA and CvdB investigation RFdA and CvdBresources RFdA and CvdB data curation RFdA writingmdashoriginal draft preparation RFdA and CvdBwritingmdashreview and editing RFdA and CvdB visualization RFdA and CvdB supervision CvdBproject administration CvdB funding acquisition CvdB All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by CNPq grant number 3098802013-0 Smithsonian Institution Cuatrecasasaward and FAPESB grant number PNX00142009
Acknowledgments We thank the staff and curators of all herbaria for their assistance with loans and DNA samplesMarco Octaacutevio de Oliveira Pellegrini for valuable suggestions on an early version of the manuscript and review ofthe English language and three anonymous reviewers for valuable comments during the reviewing process
Conflicts of Interest The authors declare no conflict of interest The funders had no role in the design of thestudy in the collection analyses or interpretation of data in the writing of the manuscript or in the decision topublish the results
References
1 Xi Z Ruhfel BR Schaefer H Amorim AMA Sugumaran M Wurdack KJ Endress PKMatthews ML Stevens PF Mathews S et al Phylogenomics and a posteriori data partitioningresolve Cretaceous angiosperm radiation Malpighiales Proc Natl Acad Sci USA 2012 109 17519ndash17524[CrossRef] [PubMed]
2 Davis CC Anderson WR A complete generic phylogeny of Malpighiaceae inferred from nucleotidesequence data and morphology Am J Bot 2010 97 2031ndash2048 [CrossRef] [PubMed]
3 Cameron KM Chase MW Anderson WR Hills HG Molecular systematics of Malpighiaceae Evidencefrom plastid rbcL and matK sequences Am J Bot 2001 88 1847ndash1862 [CrossRef] [PubMed]
4 Davis CC Madagasikaria (Malpighiaceae) A new genus from Madagascar with implications for floralevolution in Malpighiaceae Am J Bot 2002 89 699ndash706 [CrossRef]
Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 17 of 20
5 Davis CC Anderson WR Donoghue MJ Phylogeny of Malpighiaceae Evidence from chloroplast ndhFand trnL-F nucleotide sequences Am J Bot 2001 88 1830ndash1846 [CrossRef]
6 Davis CC Bell CD Fritsch PW Mathews S Phylogeny of Acridocarpus-Brachylophon (Malpighiaceae)Implications for tertiary tropical floras and Afroasian biogeography Evolution 2002 56 2395ndash2405 [CrossRef]
7 Davis CC Fritsch PW Bell CD Mathews S High-latitude tertiary migrations of an exclusively tropicalclade Evidence from Malpighiaceae Int J Plant Sci 2004 165 S107ndashS121 [CrossRef]
8 Almeida RF Amorim AMA Correa AMS van den Berg C A new infrageneric classification forAmorimia (Malpighiaceae) based on morphological phytochemical and molecular evidence Phytotaxa 2017313 231ndash248 [CrossRef]
9 Willis CG Franzone BF Xi Z Davis CC The establishment of Central American migratory corridorsand the biogeographic origins of seasonally dry tropical forests in Mexico Front Genet 2014 5 433[CrossRef]
10 Anderson CE Revision of Ryssopterys and transfer to Stigmaphyllon (Malpighiaceae) Blumea 2011 56 73ndash104[CrossRef]
11 Anderson CE Monograph of Stigmaphyllon (Malpighiaceae) Syst Bot Monographs 1997 51 1ndash313[CrossRef]
12 Flora do Brasil 2020 Available online httpfloradobrasiljbrjgovbrrefloralistaBrasilConsultaPublicaUC
ResultadoDaConsultaNovaConsultadoCondicaoTaxonCP (accessed on 10 November 2020)13 Scarano FR Ceotto P Brazilian Atlantic forest Impact vulnerability and adaptation to climate change
Biodivers Conserv 2015 24 2319ndash2331 [CrossRef]14 Ledo R Colli GR The historical connections between the Amazon and the Atlantic forest revisited
J Biogeogr 2017 44 2551ndash2563 [CrossRef]15 Vanzolini PE Williams EF The vanishing refuge A mechanism for ecogeographic speciation
Pap Avulsos Zool 1981 34 251ndash25516 Antonelli A Zizka A Carvalho FA Scharm R Bacon CD Silvestro D Condamine FL Amazonia is
the primary source of Neotropical biodiversity Proc Natl Acad Sci USA 2018 115 6034ndash6039 [CrossRef][PubMed]
17 POWO-Plants of the World Online 2020 Available online httpwwwplantsoftheworldonlineorg
(accessed on 10 November 2020)18 Korall P Pryer KM Global biogeography of scaly tree ferns (Cyatheaceae) Evidence for Gondwanan
vicariance and limited transoceanic dispersal J Biogeogr 2014 41 402ndash413 [CrossRef]19 Kranitz ML Biffin E Clark A Hollingsworth ML Ruhsam M Gardner MF Thomas P Mill RR
Ennos RA Gaudeul M et al Evolutionary diversification of New Caledonian Araucaria PLoS ONE 20149 e110308 [CrossRef]
20 Quiroga MP Mathiasen P Iglesias A Mill RR Premoli AC Molecular and fossil evidence disentanglethe biogeographical history of Podocarpus a key genus in plant geography J Biogeogr 2016 43 372ndash383[CrossRef]
21 Richardson JE Chatrou LW Mols JB Erkens RHJ Prie MD Historical biogeography of twocosmopolitan families of flowering plants Annonaceae and Rhamnaceae Phil Trans R Soc Lond B 2004359 1495ndash1508 [CrossRef]
22 Renner SS Strijk JS Strasberg D Theacutebaud C Biogeography of the Monimiaceae (Laurales) A rolefor East Gondwana and long-distance dispersal but not West Gondwana J Biogeogr 2010 37 1227ndash1238[CrossRef]
23 Kessous IM Neves B Couto DR Paixatildeo-Souza B Pederneiras LC Moura RL Barfuss MHJSalgueiro F Costa AF Historical biogeography of a Brazilian lineage of Tillandsioideae (subtribe VrieseinaeBromeliaceae) The Paranaean Sea hypothesized as the main vicariant event Bot J Linn Soc 2020192 625ndash641 [CrossRef]
24 Givnish TJ Barfuss MHJ Van Ee B Riina R Schulte K Horres R Gonsiska PA Jabaily RSCrayn DM Smith JAC et al Phylogeny adaptative radiation and historical biogeography inBromeliaceae Insights from an eight-locus plastid phylogeny Am J Bot 2011 98 872ndash895 [CrossRef][PubMed]
25 Pellegrini MOO Systematics of Commelinales Focusing on Neotropical Lineages PhD Thesis Instituto deBiociecircncias Satildeo Paulo Brazil 2019 650p
Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 18 of 20
26 Couto RS Martins AC Bolson M Lopes RC Smidt EC Braga JMA Time calibrated tree of Dioscorea(Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene Bot J Linn Soc 2018188 144ndash160 [CrossRef]
27 Vasconcelos TNC Alcantara S Andrino CO Forest F Reginato M Simon MF Pirani JRFast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancientNeotropical mountains Proc R Soc B 2020 287 20192933 [CrossRef]
28 Gustafsson ALS Verola CF Antonelli A Reassessing the temporal evolution of orchids with new fossilsand a Bayesian relaxed clock with implications for the diversification of the rare South American genusHoffmannseggella (Orchidaceae Epidendroideae) BMC Evol Biol 2010 10 177 [CrossRef]
29 Ruiz-Sanchez E Biogeography and divergence time estimates of woody bamboos Insights in the evolutionof Neotropical bamboos Bol Soc Bot Meacutex 2011 88 67ndash75 [CrossRef]
30 Zhang XZ Zeng CX Ma PF Haevermans T Zhang YX Zhang LN Guo ZH Li DZ Multi-locusplastid phylogenetic biogeography supports the Asian hypothesis of the temperate woody bamboos (PoaceaeBambusoideae) Mol Phylogenet Evol 2016 96 118ndash129 [CrossRef]
31 Rapini A van den Berg C Liede-Schumann S Diversification of Asclapiadoideae (Apocynaceae) in theNew World Ann Missouri Bot Gard 2007 94 407ndash422 [CrossRef]
32 Mandel JR Dikow RB Siniscalchi CM Thapa R Watson LE Funk VA A fully resolved backbonephylogeny reveals numerous dispersals and explosive diversifications throughout the history of AsteraceaeProc Natl Acad Sci USA 2019 116 14083ndash14088 [CrossRef]
33 Fonseca LHM Lohmann LG Biogeography and evolution of Dolichandra (Bignonieae Bignoniaceae)Bot J Linn Soc 2015 179 403ndash420 [CrossRef]
34 Hernaacutendez-Hernaacutendez T Brown JW Schlumpberger BO Eguiarte LE Magalloacuten S Beyond aridificationMultiple explanations for the elevated diversification of cacti in the New World Succulent Biome New Phytol2014 202 1382ndash1397 [CrossRef] [PubMed]
35 Meseguer AS Lobo JM Cornuault J Beerling D Ruhfel BR Davis CC Jousselin E Sanmartiacuten IReconstructing deep-time palaeoclimate legacies in the clusioid Malpighiales unveil their role in the evolutionand extinction of the boreotropical flora Glob Ecol Biogeogr 2018 27 616ndash628 [CrossRef]
36 Soares-Neto RL Thomas WW Barbosa MRV Roalson EH Diversification of New World Cleomaceaewith emphasis on Tarenaya and the description of Iltisiella a new genus Taxon 2020 69 321ndash336 [CrossRef]
37 Ruhfel BR Bove CP Philbrick CT Davis CC Dispersal largely explains the Gondwanan distribution ofthe ancient tropical clusioid plant clade Am J Bot 2016 103 1117ndash1128 [CrossRef]
38 Silva OLM Riina R Cordeiro I Phylogeny and biogeography of Astraea with new insights into theevolutionary history of Crotoneae (Euphorbiaceae) Mol Phylogenet Evol 2020 145 106738 [CrossRef][PubMed]
39 Lavin M Herendeen PS Wojciechowski MF Evolutionary rates analysis of Leguminosae implicatesa rapid diversification of lineages during the Tertiary Syst Biol 2005 54 575ndash594 [CrossRef]
40 Bruneau A Mercure M Lewis GP Herendeen PS Phylogenetic patterns and diversification in thecaesalpinioid legumes Botany 2008 86 697ndash718 [CrossRef]
41 Gagnon E Ringelberg JJ Bruneau A Lewis GP Hughes CE Global Succulent Biome phylogeneticconservatism across the pantropical Caesalpinia group (Leguminosae) New Phytol 2019 222 1994ndash2008[CrossRef]
42 Roalson EH Roberts WR Distinct processes drive diversification in different clades of GesneriaceaeSyst Biol 2016 65 662ndash684 [CrossRef]
43 Castillo RA Luebert F Henning T Weigend M Major lineages of Loasaceae subfam Loasoideaediversified during the Andean uplift Mol Phylogenet Evol 2019 141 106616 [CrossRef]
44 Davis CC Schaefer H Xi Z Baum DA Donoghue MJ Harmon LJ Long-term morphological stasismaintained by a plant-pollinator mutualism Proc Natl Acad Sci USA 2014 111 5914ndash5919 [CrossRef][PubMed]
45 Reginato M Vasconcelos TNC Kriebel R Simotildees AO Is dispersal mode a driver of diversificationand geographical distribution in the tropical plant family Melastomataceae Mol Phylogenet Evol 2020148 106815 [CrossRef] [PubMed]
Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 19 of 20
46 Machado AFP Ronsted N Bruun-Lund S Pereira RAS Queiroz LP Atlantic forests to the all AmericasBiogeographical history and divergence times of Neotropical Ficus (Moraceae) Mol Phylogenet Evol 2018122 46ndash58 [CrossRef] [PubMed]
47 Vasconcelos TNC Proenccedila CEB Ahmad B Aguilar DS Aguilar R Amorim BS Campbell KCosta IR De-Carvalho PS Faria JEQ et al Myrteae phylogeny calibration biogeographyand diversification patterns Increased understanding in the most species-rich tribe of MyrtaceaeMol Phylogenet Evol 2017 109 113ndash137 [CrossRef] [PubMed]
48 Santos MF Lucas E Sano PT Buerki S Staggemeier VG Forest F Biogeographical patterns of Myrcia sl(Myrtaceae) and their correlation with geological and climatic history in the Neotropics Mol Phylogenet Evol2017 108 34ndash48 [CrossRef] [PubMed]
49 Buerki S Forest F Alvarez N Nylander JAA Arrigo N Sanmartiacuten I An evaluation of newparsimony-based versus parametric inference methods in biogeography A case study using the globallydistributed plant family Sapindaceae J Biogeogr 2011 38 531ndash550 [CrossRef]
50 Saumlrkinen T Bohs L Olmstead RG Knapp S A phylogenetic framework for evolutionary study of thenightshades (Solanaceae) A dated 1000-tip tree BMC Evol Biol 2013 13 214 [CrossRef]
51 Gonccedilalves DJP Shimizu GH Ortiz EM Jansen RK Simpson BB Historical biogeography ofVochysiaceae reveals an unexpected perspective of plant evolution in the Neotropics Am J Bot 2020107 1ndash17 [CrossRef]
52 Thode VA Sanmartiacuten I Lohmann LG Contrasting patterns of diversification between Amazonian andAtlantic forest clades of Neotropical lianas (Amphilophium Bignonieae) inferred from plastid genomic dataMol Phylogenet Evol 2019 133 92ndash106 [CrossRef]
53 Lohmann LG Bell CD Calioacute MF Winkworth RC Pattern and timing of biogeographical history in theNeotropical tribe Bignonieae (Bignoniaceae) Bot J Linn Soc 2013 171 154ndash170 [CrossRef]
54 Simon MF Grether R Queiroz LP Skema C Pennington RT Hughes CE Recent assemblyof the Cerrado a neotropical plant diversity hotspot by in situ evolution of adaptations to fireProc Natl Acad Sci USA 2009 106 20359ndash20364 [CrossRef] [PubMed]
55 Mazine FF Faria JEQ Giaretta A Vasconcelos T Forest F Lucas E Phylogeny and biogeographyof the hyper-diverse genus Eugenia (Myrtaceae Myrteae) with emphasis on E sect Umbellatae the mostunmanageable clade Taxon 2018 67 752ndash769 [CrossRef]
56 Davies BJ Hambrey MJ Smelle JL Carrivick JL Glasser NF Antarctic Peninsula ice sheet evolutionduring the Cenozoic era Quat Sci Rev 2012 31 30ndash66 [CrossRef]
57 Lewis AR Marchant DR Ashworth AC Hedenaumls L Hemming SR Johnson JV Leng MLMachlus ML Newton AE Raine JI et al Mid-Miocene cooling and the extinction of tundra incontinental Antarctica Proc Natl Acad Sci USA 2008 105 10676ndash10680 [CrossRef]
58 Chacoacuten J Assis MC Meerow AW Renner SS From East Gondwana to Central America Historicalbiogeography of the Alstroemeriaceae J Biogeogr 2012 39 1806ndash1818 [CrossRef]
59 Maciel JR Estudos Taxonocircmicos Filogeneacuteticos e Biogeograacuteficos em Aechmea (Bromeliaceae) PhD ThesisUniversidade Federal de Pernambuco Recife Brazil 2017 213p
60 Morley RJ Origin and Evolution of Tropical Rain Forests Wiley Chichester UK 2000 p 36261 Zachos J Pagani M Sloan L Thomas E Billups K Trends rhythms and aberrations in global climate
65 Ma to present Science 2001 292 686ndash693 [CrossRef]62 Flower BP Kennett JP The middle Miocene climate transition East Antarctic ice sheet development
deep ocean circulation and global carbon cycling Palaeogeogr Palaeoclimatol Palaeoecol 1994 108 537ndash555[CrossRef]
63 Prado DE Gibbs PE Patterns of species distributions in the dry seasonal forests of South AmicaAnn Mo Bot Gard 1993 80 902ndash927 [CrossRef]
64 Peres EA Pinto-da-Rocha R Lohmann LG Michelangeli FA Miyaki CY Carnaval AC Patterns ofspecies and lineage diversity in the Atlantic Rainforest of Brazil In Neotropical Diversification Patterns andProcesses Rull V Carnaval AC Eds Springer Cham Switzerland 2020 pp 415ndash520
65 Karl M Glasmacher UA Kollenz S Franco-Magalhatildees AOB Stockli DF Hackspacher PC Evolutionof the South Atlantic passive continental margin in southern Brazil derived from zircon and apatite(UndashThndashSm)He and fission-track data Tectonophysics 2013 604 224ndash244 [CrossRef]
Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-

Plants 2020 9 1569 20 of 20
66 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissue Phytochem Bull1987 19 11ndash15
67 Almeida RF Amorim AMA van den Berg C Timing the origin and past connections between Andeanand Atlantic Seasonally Dry Tropical Forests in South America Insights from the biogeographical history ofAmorimia (Malpighiaceae) Taxon 2018 67 739ndash751 [CrossRef]
68 Kearse M Moir R Wilson A Stones-Havas S Cheung M Sturrock S Buxton S Cooper AMarkowitz S Duran C et al Geneious Basic An integrated and extendable desktop software platform forthe organization and analysis of sequence data Bioinformatics 2012 28 1647ndash1649 [CrossRef] [PubMed]
69 Edgar RC MUSCLE A multiple sequence alignment method with reduced time and space complexityBMC Inform 2004 5 113
70 Swofford DL PAUP Phylogenetic Analysis Using Parsimony and Other Methods Version 40b10 SinauerSunderland UK 2002
71 Fitch WM Towards defining the course of evolution Minimum change for a specific tree topology Syst Zool1971 20 406ndash416 [CrossRef]
72 Felsenstein J Confidence limits on phylogenies An approach using the bootstrap Evolution 1985 39 783ndash791[CrossRef]
73 Darriba D Taboada GL Doallo R Posada D jModelTest 2 More models new heuristics and parallelcomputing Nat Methods 2012 9 772 [CrossRef]
74 Ronquist F Huelsenbeck JP MrBayes 3 Bayesian phylogenetic inference under mixed modelsBioinformatics 2003 19 1572ndash1574 [CrossRef]
75 Drummond AJ Suchard MA Xie D Rambaut A Bayesian phylogenetics with BEAUti and the BEAST17 Mol Biol Evol 2012 29 1969ndash1973 [CrossRef]
76 Cai L Xi Z Peterson K Rushworth C Beaulieu J Davis CC Phylogeny of Elatinaceae and the tropicalGondwanan origin of the Centroplacaceae (Malpighiaceae Elatinaceae) clade PLoS ONE 2016 11 e0161881[CrossRef]
77 Rambaut A Suchard MA Xie D Drummond AJ Tracer v16 2014 Available online httpbeastbioedacukTracer (accessed on 10 November 2020)
78 FigTree 2020 Available online httptreebioedacuksoftwarefigtree (accessed on 10 November 2020)79 Anderson WR Davis CC Generic adjustments in Neotropical Malpighiaceae Contr Univ Michigan Herb
2007 25 137ndash16680 Gates B Banisteriopsis Diplopterys (Malpighiaceae) Flora Neotrop 1982 30 1ndash23881 IBGE-Instituto Brasileiro de Geografia e Estatiacutestica Mapa de vegetaccedilatildeo do Brasil Rio de Janeiro Brasil 2012
Available online httpwwwibgegovbrhomepresidencianoticias21052004biomashtmlshtm (accessed on10 November 2020)
82 WWF-World Wildlife Fund Tropical and Subtropical Dry Broadleaf Forests 2020 Available online httpwwwworldwildlifeorgbiomestropical-and-subtropical-dry-broadleaf-forests (accessed on 10 November2020)
83 Yu Y Harris AJ Blair C He XJ RASP (Reconstruct Ancestral State in Phylogenies) A tool for historicalbiogeography Mol Phylogenet Evol 2015 87 46ndash49 [CrossRef] [PubMed]
84 Ree RH Smith SA Maximum likelihood inference of geographic range evolution by dispersallocal extinction and cladogenesis Syst Biol 2008 57 4ndash14 [CrossRef] [PubMed]
85 Ree RH Sanmartiacuten I Conceptual and statistical problems with the DEC+J model of founder-eventspeciation and its comparison with DEC via model selection J Biogeogr 2018 45 741ndash749 [CrossRef]
Publisherrsquos Note MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations
copy 2020 by the authors Licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
- Introduction
- Results
-
- Phylogenetic Analysis
- Ancestral Area Reconstruction and Divergence Times Estimation
- Meta-Analysis
-
- Discussion
-
- Phylogenetics of Stigmaphyllon
- Divergence Times and Biogeography of Stigmaphyllon
- Time and Diversification of the Atlantic Rainforest
-
- Material and Methods
-
- Taxon Sampling and Plant Material
- Molecular Protocols
- Phylogenetic Analysis
- Calibration
- Ancestral Area Reconstruction
- Meta-Analysis
-
- Conclusions
- References
-


















