
Autonomous and nonautonomous Notch functions for...
10
INTRODUCTION In Drosophila embryos, the neurogenic genes (almondex, big brain, Delta, Enhancer of split, mastermind, neuralized and Notch) govern the development of cells in all germ layers (Lehmann et al., 1983; Hartenstein et al., 1992). The products of these genes are involved in a common signal transduction pathway that regulates neurogenesis (for review, see Campos- Ortega, 1993). During germ band extension, metamerically repeated clusters of ectodermal cells transiently express proneural genes, which promote neural development (Jiménez and Campos-Ortega, 1990). One cell per cluster maintains proneural gene expression and thus becomes a neuroblast; inductive interactions cause the remaining cells shut down proneural gene expression and give rise to epidermal deriva- tives (Skeath and Carroll, 1992). This inductive pathway, called lateral inhibition, is regulated by the neurogenic genes. Embryos lacking function for these genes exhibit a hypertro- phied nervous system and concomitant absence of ventral epidermis (Lehmann et al., 1983). Two of the neurogenic genes, Notch (N) and Delta (Dl), encode transmembrane proteins containing multiple EGF-like repeats in their extracellular domains (Wharton et al., 1985; Vässin et al., 1987). Interactions between the Notch and Delta proteins have been inferred genetically (Brand and Campos- Ortega, 1990; Xu et al., 1990) and demonstrated biochemically (Fehon et al., 1990; Rebay et al., 1991). Studies of epidermal cell development in genetic mosaics suggest that Notch is autonomously required by proneural ectoderm cells for choosing an epidermal fate, consistent with the hypothesis that Notch functions as a signal receptor during lateral inhibition (Hoppe and Greenspan, 1990; Heitzler and Simpson, 1991). Although this hypothesis has been challenged by contradictory observations (Gehring, 1973; Technau and Campos-Ortega, 1987), evidence that the intracellular Notch domain localizes to the nucleus (Lieber et al., 1993; Fortini et al., 1993; Struhl et al., 1993) is consistent with the model that Notch functions as a receptor. Furthermore, cell culture studies show that nuclear localization of the Notch intracellular domain stimu- lates expression of Enhancer of split [E(spl)] proteins (Jennings et al., 1994), which normally act downstream of N during neurogenesis (de la Concha et al., 1988). Poulson (1945) first described the mesodermal defects of N embryos; however, the role of the neurogenic genes in this germ layer has only recently been systematically addressed (Corbin et al., 1991; Hartenstein et al., 1992; Bate et al., 1993). The most striking defect is the absence of differentiated somatic muscles (Poulson, 1945). In mutant embryos, somatic mesoderm expresses β3-tubulin and muscle myosin (Corbin et al., 1991), and apparently fails to generate syncytial myotubes 617 Development 122, 617-626 (1996) Printed in Great Britain © The Company of Biologists Limited 1996 DEV5027 The Notch (N) gene encodes a cell signaling protein that mediates neuronal and epidermal determination in Drosophila embryos. N also regulates several aspects of myogenic development; embryos lacking N function have too many muscle founder cells and fail to properly dif- ferentiate somatic muscle. To identify cell-autonomous requirements for Notch function during muscle develop- ment, we expressed a Notch minigene in the mesoderm, but not in the ectoderm, of amorphic N - embryos. In these embryos, muscle founder hypertrophy is rescued, indicating that Notch is autonomously required by mesoderm cells to regulate the proper number of muscle founders. However, somatic muscle differentiation is only partially normalized, suggesting that Notch is also required in the ectoderm for proper muscle develop- ment. Additionally, mesodermal expression of Notch partially rescues epidermal development in overlying neurogenic ectoderm. This is unexpected, since previous studies suggest that Notch is autonomously required by proneural ectoderm cells for epidermal development. Mesodermal expression of a truncated Notch protein lacking the extracellular domain does not rescue ventral epidermis, suggesting that the extra- cellular domain of Notch can non-autonomously rescue epidermal development across germ layers. Key words: Notch, myogenesis, neurogenesis, cell signaling, Drosophila SUMMARY Autonomous and nonautonomous Notch functions for embryonic muscle and epidermis development in Drosophila Rob Baker 1, * and Gerold Schubiger 2,† 1 Department of Genetics and 2 Department of Zoology, University of Washington, Seattle, WA 98195, USA *Present address: Department of Anatomy, University of Wisconsin, 1300 University Avenue, Madison WI 53706, USA † Author for correspondence
Transcript of Autonomous and nonautonomous Notch functions for...

617Development 122, 617-626 (1996)Printed in Great Britain © The Company of Biologists Limited 1996DEV5027
Autonomous and nonautonomous Notch functions for embryonic muscle and
epidermis development in Drosophila
Rob Baker1,* and Gerold Schubiger2,†
1Department of Genetics and 2Department of Zoology, University of Washington, Seattle, WA 98195, USA
*Present address: Department of Anatomy, University of Wisconsin, 1300 University Avenue, Madison WI 53706, USA†Author for correspondence
The Notch (N) gene encodes a cell signaling protein thatmediates neuronal and epidermal determination inDrosophila embryos. N also regulates several aspects ofmyogenic development; embryos lacking N function havetoo many muscle founder cells and fail to properly dif-ferentiate somatic muscle. To identify cell-autonomousrequirements for Notch function during muscle develop-ment, we expressed a Notch minigene in the mesoderm,but not in the ectoderm, of amorphic N− embryos. In theseembryos, muscle founder hypertrophy is rescued,indicating that Notch is autonomously required bymesoderm cells to regulate the proper number of musclefounders. However, somatic muscle differentiation is onlypartially normalized, suggesting that Notch is
also required in the ectoderm for proper muscle develop-ment.
Additionally, mesodermal expression of Notch partiallyrescues epidermal development in overlying neurogenicectoderm. This is unexpected, since previous studies suggestthat Notch is autonomously required by proneural ectodermcells for epidermal development. Mesodermal expression ofa truncated Notch protein lacking the extracellular domaindoes not rescue ventral epidermis, suggesting that the extra-cellular domain of Notch can non-autonomously rescueepidermal development across germ layers.
Key words: Notch, myogenesis, neurogenesis, cell signaling,Drosophila
SUMMARY
INTRODUCTION
In Drosophila embryos, the neurogenic genes (almondex, bigbrain, Delta, Enhancer of split, mastermind, neuralized andNotch) govern the development of cells in all germ layers(Lehmann et al., 1983; Hartenstein et al., 1992). The productsof these genes are involved in a common signal transductionpathway that regulates neurogenesis (for review, see Campos-Ortega, 1993). During germ band extension, metamericallyrepeated clusters of ectodermal cells transiently expressproneural genes, which promote neural development (Jiménezand Campos-Ortega, 1990). One cell per cluster maintainsproneural gene expression and thus becomes a neuroblast;inductive interactions cause the remaining cells shut downproneural gene expression and give rise to epidermal deriva-tives (Skeath and Carroll, 1992). This inductive pathway,called lateral inhibition, is regulated by the neurogenic genes.Embryos lacking function for these genes exhibit a hypertro-phied nervous system and concomitant absence of ventralepidermis (Lehmann et al., 1983).
Two of the neurogenic genes, Notch (N) and Delta (Dl),encode transmembrane proteins containing multiple EGF-likerepeats in their extracellular domains (Wharton et al., 1985;Vässin et al., 1987). Interactions between the Notch and Deltaproteins have been inferred genetically (Brand and Campos-
Ortega, 1990; Xu et al., 1990) and demonstrated biochemically(Fehon et al., 1990; Rebay et al., 1991). Studies of epidermalcell development in genetic mosaics suggest that Notch isautonomously required by proneural ectoderm cells forchoosing an epidermal fate, consistent with the hypothesis thatNotch functions as a signal receptor during lateral inhibition(Hoppe and Greenspan, 1990; Heitzler and Simpson, 1991).Although this hypothesis has been challenged by contradictoryobservations (Gehring, 1973; Technau and Campos-Ortega,1987), evidence that the intracellular Notch domain localizesto the nucleus (Lieber et al., 1993; Fortini et al., 1993; Struhlet al., 1993) is consistent with the model that Notch functionsas a receptor. Furthermore, cell culture studies show thatnuclear localization of the Notch intracellular domain stimu-lates expression of Enhancer of split [E(spl)] proteins(Jennings et al., 1994), which normally act downstream of Nduring neurogenesis (de la Concha et al., 1988).
Poulson (1945) first described the mesodermal defects of Nembryos; however, the role of the neurogenic genes in thisgerm layer has only recently been systematically addressed(Corbin et al., 1991; Hartenstein et al., 1992; Bate et al., 1993).The most striking defect is the absence of differentiatedsomatic muscles (Poulson, 1945). In mutant embryos, somaticmesoderm expresses β3-tubulin and muscle myosin (Corbin etal., 1991), and apparently fails to generate syncytial myotubes

618 R. Baker and G. Schubiger
(Bate et al., 1993). The number of nautilus-, vestigial- and S59-expressing founder cells is increased in these mutants (Corbinet al., 1991; Bate et al., 1993), presumably at the expense of‘fusion competent’ cells, which normally fuse with musclefounders (Bate, 1990). In the dorsal mesoderm, supernumerarycardioblasts develop at the expense of pericardial cells and theheart tube fails to differentiate (Hartenstein et al., 1992).Neither visceral nor pharyngeal muscles differentiate properly,although visceral muscles development begins normally(Hartenstein et al., 1992).
During myogenesis, Notch is expressed throughout theembryo (Fehon et al., 1990). It is likely that Notch is requiredin the mesoderm to execute most of its myogenic functions,given its apparent cell-autonomous Notch function in theectoderm. This nonetheless does not preclude a myogenic rolefor Notch in the adjacent ectoderm. Since the epidermis playsan integral role in muscle navigation and attachment (Leptinet al., 1989; Bate, 1990; Volk and VijayRaghaven, 1994), it ispossible that the paucity of epidermis in N− embryos augmentsmorphological defects resulting from the loss of mesoderm-specific Notch activity. Moreover, ectoderm inducesexpression of myogenic genes in the underlying mesoderm(Staehling-Hampton et al., 1994; Baker and Schubiger, 1995;Maggert et al., 1995; Frasch, 1995). Given the gross ectoder-mal defects in N− embryos, such inductive interactions willprobably be affected.
Here we investigate cell-autonomous requirements forNotch during myogenesis. We used the GAL4-targeted geneexpression system (Brand and Perrimon, 1994) to generateembryos that express Notch throughout the mesoderm, but notin the ectoderm, during the first half of embryogenesis.Myogenic gene expression and muscle morphogenesis arepartially normalized in these embryos, suggesting that Notchis required in the mesoderm for only some aspects of muscledevelopment. We also show that epidermal development issubstantially rescued in these embryos and demonstrate thatthis phenomenon is not due to expression of Notch in theectoderm. Several possible mechanisms for this novel, nonau-tonomous Notch activity are presented.
MATERIALS AND METHODS
Transgenic fly stocksptwi-GAL4 and pUAS-N (Fig. 1) are P-element transformation vectorsderived from pCaSpeR3 (Thummel et al., 1988). ptwi-GAL4 wasgenerated by replacing the BamHI-XbaI lacZ fragment from pCaSpeR2.9 (Jiang et al., 1991) with the BamHI-SpeI GAL4 fragment frompGaTB (Brand and Perrimon, 1994). pUAS-N was constructed byinserting a NotI/XbaI full-length Notch cDNA fragment (Rebay et al.,1991) into the polylinker of pUAST (Brand and Perrimon, 1994).pUAS-N and ptwi-GAL4 were injected into w1118; P[ry+, ∆2-3],Sb/TM6B embryos by standard methods (Ashburner, 1989). Eclosedadults were crossed with 3-5 w/w or w/Y flies and F1 adults werescored for eye pigmentation and backcrossed to w1118 flies to generatestocks.
Three independent nonlethal X-linked insertions of UAS-N werecrossed with w1118, N54l9 females in order to obtain w1118, N54l9,P[w+,UAS-N] recombinants, which were maintained over the winscychromosome. A nonlethal insertion of P[w+, twi-GAL4] into chromo-some II (twi-GAL41) was maintained both as a homozygous stock andbalanced over CyO. Two independent UAS-∆ECN stocks were
generated by S. Fuerstenberg and E. Giniger, and crossed into a N54l9
background. UAS-∆ECN contains the coding region of the Delta startand membrane transport signal sequence (amino acids 1-27) fused tothe coding region of the Notch transmembrane and intracellular(amino acids 1742-2703), placed within the pUAST expression vector(E. Giniger, pers. comm.). Homozygous transgenic lines UAS-lacZand 69B were generated by Brand and Perrimon (1994). P[UAS-lacZ]flies express the lacZ reporter gene under transcriptional control ofGAL4 upstream activating sequences. Stock 69B expresses GAL4throughout most of the ectoderm and mesoderm during germ bandextension (Castelli-Gair et al., 1994).
Embryo fixation and labelingEmbryos were collected and incubated at 25°C according to Bakerand Schubiger (1995). Embryos were dechorioniated in 50% bleach,rinsed in PBS and fixed in 4% EM-grade paraformaldehyde (ElectronMicroscopy Sciences) in PBS for 30 minutes. The fixative wasreplaced with methanol to devitellinize the embryos. For antibodylabeling, embryos were blocked with PBS + 3% bovine serumalbumin (PBSA) and labeled with antibodies in this medium in thefollowing proportions: 1:50 anti-Notch (specific to the intracellulardomain; S. Artavanis-Tsakonas); 1:40 anti-nuclear lamin (M.Schubiger); 1:200 anti-myosin heavy chain (B. Taylor); 1:500 anti-ELAV (S. Robinow); 1:200 anti-nautilus (B. Paterson); 1:100 anti-vestigial (L. Maves); 1:1000 anti-horseradish peroxidase (USB).Embryos were rinsed in PBSA for an hour and incubated with theappropriate biotin- or fluorochrome-conjugated secondary antibodies(Jackson ImmunoResearch) diluted 1:200 in PBSA overnight at 4°C.Fluorescence-labeled embryos were dehydrated in methanol, andcleared and mounted in a 1:2 mixture of benzyl benzoate and benzylalcohol (Sigma Immunochemicals).
RESULTS
Targeted expression of Notch and ∆ECN in themesodermWe employed the GAL4-targeted gene expression system(Brand and Perrimon, 1994) to activate tissue-specificexpression of Notch proteins in Drosophila embryos. twi-GAL4 (Fig. 1A) expresses the transcriptional activator GAL4under control of the twist promoter (Jiang et al., 1991), whichis active throughout the mesoderm preceding muscle differen-tiation (Thisse et al., 1988; Bate et al., 1991). Two transgenesexpress Notch-specific sequences under control of GAL4upstream activating sequences, or UASs: UAS-N (Fig. 1B),which encodes a complete Notch protein, henceforth called‘N’, and UAS-∆ECN (Fig. 1C), which encodes a truncatedNotch protein lacking the extracellular domain, henceforthcalled ‘∆ECN’.
To assay for the spatial and temporal distribution of twi-GAL4 activity in the embryo, homozygous twi-GAL41 maleswere crossed with homozygous UAS-lacZ females (Brand andPerrimon, 1994) and their progeny probed with antibodyagainst β-galactosidase or stained in X-gal solution at variousdevelopmental stages. Both assays revealed strong signal in themesoderm, endoderm and mesectoderm of embryos aged 4-10hours after egg laying (AEL), but no signal in ectodermalderivatives (not shown). The presence of GAL4 in the midlinecells and midgut is probably due to the transient activity of thetwist promoter in the presumptive mesectoderm and endodermin blastoderm-stage embryos (Thisse et al., 1988), and atteststo the perdurance of GAL4 and/or β-galactosidase inDrosophila.

619Notch functions in the mesoderm
ASNscy
; ++
XY
twiGAL4
twiGAL4; twiGAL4
+N- UASN
Y;
"Ntwi embryo"
X (1/4)
cy; UASICN
UASICN
++
; XY
twiGAL4
twiGAL4;
++
; UASICN+
twiGAL4+Y
N-;;X
"ICNtwi embryo"
(1/4)
GAL4w+ptwist 3'GAL4 twiGAL4
galUASs3'Notchw+ UASN
3'ICNw+galUASs
UASICN
ors and crosses for generating embryos with mosaic Notch expression.atic representations of P-element transformation vectors used in these
. (A) ptwi-GAL4 contains a GAL4 minigene downstream of a 2.9 kbter region. (B) pUAS-N contains a full-length Notch cDNA under theal control of five tandem GAL upstream activating sequences (UASs).ECN encodes a truncated Notch protein lacking the extracellularer control of five GAL UASs. (D,E) Diagrams of genetic crossesgenerate embryos that express Notch (D) or ∆ECN (E) in thef N54l9 embryos (see Materials and Methods for details).
UAS-N and UAS-∆ECN are activated by GAL4 and thesetransgenes are expressed in the mesoderm of transgenic twi-GAL4 embryos. By making appropriate genetic crosses (Fig.1D,E), we express either UAS-N or UAS-∆ECN in themesoderm of N− embryos. An X-linked UAS-N insertion (UAS-N107) was recombined with N54l9, a protein null N allele, andmaintained over the winscy chromosome. One-quarter of theembryos (genotypically N54l9, UAS-N107/Y) from this stock diebefore hatching and are phenotypically indistinguishable fromN54l9 hemizygotes. However, when N54l9, UAS-N107 femalesare crossed with homozygous twi-GAL41 males, one-quarter ofthe progeny lack endogenous N+ function but express N in themesoderm from the UAS-N107 transgene. For convenience, weuse the term ‘Ntwi’ to designate the genotype N54l9, UAS-N107/Y;twi-GAL41/+ (Fig 1D). Between 4 and 10 hours after egglaying (AEL), Ntwi embryos express high levels of N in themesoderm, and somewhat lower amounts in the mesectodermand endoderm (Fig. 2C). In Ntwi embryos N is not detected inthe dorsal or ventral ectoderm. By making analogous crosses(Fig. 1E), we obtained both N+ and N− embryos that express∆ECN in the mesoderm, mesectoderm and endoderm (Fig.2D). N54l9/Y; twi-GAL41/+; UAS-∆ECN3/+ embryos lack Nfunction but express ∆ECN in the mesoderm, and are desig-nated as ‘∆ECNtwi embryos’.
To verify that the UAS-N product is functional, we crossedN54l9,UAS-N107 females to males homozygous for the 69Btransgene, which expresses GAL4 throughout the ectoderm aswell as mesoderm during germ band extension (Castelli-Gairet al., 1994). One-quarter of the progeny from this cross fail tohatch. None of these unhatched embryos exhibitepidermal defects characteristic of N54l9 embryos;although they do not properly involute their headsegments, they secrete a normal ventral cuticle(Fig. 3C). This indicates that the UAS-N productcan function in place of endogenous N duringlateral inhibition.
We performed an analogous assay for UAS-∆ECN function. When the truncated ∆ECNproduct is expressed in neurogenic ectoderm ofN+ embryos, it produces an ‘antineurogenic’phenotype characterized by a reduced number ofneuroblasts (E. Giniger, unpublished results), aswas previously reported for embryos that expressthe intracellular Notch domain under heat shockpromoter control (Lieber et al., 1993; Struhl et al.,1993). N54l9/winscy; UAS-∆ECN/UAS-∆ECNfemales were crossed with 69B males and theirprogeny examined for neurogenic cuticle defects.None of the progeny hatch, but all producenormal ventral cuticle (Fig. 3D), indicating that∆ECN can substitute for the epidermalizingfunction of N in the ectoderm.
Epidermal development in Ntwi and∆ECNtwi embryosPrevious reports indicate that N functions cell-autonomously (Hoppe and Greenspan, 1990;Heitzler and Simpson, 1991). We thereforeanticipated that ventral epidermis would notdevelop in Ntwi embryos. However, Ntwi embryosshow normalized epidermal development relative
N- Uwin
D
wins
N-E
5'A
5'B
5'C
Fig. 1. Vect(A-C) Schemexperimentstwist promotranscription(C) pUAS-∆domain, unddesigned to mesoderm o
to N54l9 mutants. Cuticle preparations of 24-hour-old Ntwi
embryos exhibit significant amounts of ventral epidermis,which is herniated anterior to the first thoracic segment, withadditional hernias in the ventrolateral cuticle of thoracic andabdominal segments (Fig. 3E,F). The most anterior cuticularstructures, such as antennae and cirri, are rarely observed.Ventral denticle belts in Ntwi embryos are often fused, with theanterior complement being reversed in polarity. Epidermaldifferentiation is accompanied with a slight reduction of neuralhypertrophy in the ventral nerve cord (data not shown). Nor-malization of neural hypertrophy is less prominent in the PNSand cephalic region, which secretes little or no cuticle.
One explanation for the cuticle phenotype of Ntwi embryosis that N is expressed, albeit below the threshold of detection,in the ventral ectoderm of these embryos. However, functionalassays for ‘leaky’ expression of UAS-N107 and twi-GAL41
suggest that neither is expressed in the ectoderm. N54l9,UAS-N107 females that are not crossed with any GAL4 line produceembryos that are phenotypically indistinguishable from N54l9
hemizygotes, indicating that UAS-N107 is not expressed in theectoderm. To assay for twi-GAL4 expression in the ectoderm,we analyzed the cuticle of ∆ECN twi embryos. ∆ECN promotesepidermal development when it is expressed in the ventralectoderm (Fig. 3D); since it lacks the EGF-like extracellulardomain, it can only function cell-autonomously. UAS-N107 andUAS-∆ECN3 share identical promoters, and since ∆ECNfunctions as a constitutively activated form of N (E. Giniger,pers. comm.), ∆ECNtwi embryos should generate equivalent orgreater amounts of epidermis as Ntwi embryos, but only if twi-

620 R. Baker and G. Schubiger
GAL41 is expressed in the ectoderm. We find, however, that∆ECN twi embryos generate a typical N− cuticle (Fig. 3G), indi-cating that twi-GAL41 is not expressed in the ectoderm.Therefore, the rescued ventral epidermis in Ntwi embryos mustsomehow be due to expression of N in the mesoderm.
Somatic muscle development in Ntwi and ∆ECNtwi
embryosWe compared muscle morphology and patterns of muscle-specific gene expression in N+, N54l9 and Ntwi embryos todetermine if expression of N in the mesoderm can rescue themyogenic defects characteristic of N− embryos. Stage 16embryos were fluorescently double labeled with antibodiesagainst muscle myosin heavy chain (MHC) and ELAV, whichis expressed in neurons (Robinow and White, 1991). In wild-type embryos, somatic muscle founders fuse with surroundingmesoderm cells to generate 30-33 muscles per thoracic andabdominal hemisegment (Bate, 1993). These muscles areorganized in a repeated pattern of dorsal, lateral and ventralmuscle groups (Fig. 4A). As in other neurogenic mutantembryos (Corbin et al., 1991; Hartenstein et al., 1992; Bate etal., 1993), N54l9 embryos express MHC in irregularly shapedmyocytes that aggregate beneath patches of dorsal epidermisand are rarely observed in the ventral region of the embryo thatis devoid of epidermis (Fig. 4B). By double labeling N54l9
embryos with antibodies to MHC and nuclear lamin, we foundthat most MHC-expressing cells contain between 4 and 8
Fig. 2. Distribution of endogenous and transgenic Notch proteins. Genetan antibody against the Notch intracellular domain, revealing the distribuleft; scale bar = 50 µm. (A) A N+ embryo from the N54l9, UAS-N/winscyNotch is expressed most strongly in the foregut and hindgut, and is stronthe mesoderm exhibits relatively low levels of protein. (B) A hemizygouare phenotypically identical to N54l9 mutant embryos in all respects. A Nconditions. (C) A Ntwi embryo (N54l9,UAS-N107/Y;twi-GAL41/+) expressmesectoderm is not evident in this panel (see Fig. 5D for mesectodermalectoderm, which do not express twist. (D) A N+ embryo that expresses Uthe endogenous protein (in the ectoderm) and the ∆ECN (most of the me
nuclei (see inset to Fig. 4B), indicating that N is not requiredfor myoblast fusion. The degree of fusion in these null mutantembryos was much higher than expected, since Bate et al.(1993) previously reported that myoblasts in strong neurogenicmutants rarely fuse.
Somatic muscle morphology in Ntwi embryos is intermedi-ate between that of wild-type and N54l9 embryos. Ntwi embryosexhibit a significant degree of myotube differentiation, basedon the size, shape and presence of multiple nuclei within MHC-expressing cells (Fig. 4C). The muscle complement is smallerthan in wild-type embryos, and varies between embryos andbetween segments within the same embryo. These musclesoccupy more ventral positions in Ntwi than in N54l9 embryos;this is probably due to the presence of ventral epidermis in Ntwi
embyros, since epidermis provides a substratum for migrationand attachment for developing myotubes (Leptin et al., 1989;Bate, 1990; Volk and VijayRaghaven, 1994). Relative to wild-type embryos, there are few MHC-expressing cells anterior tothe thoracic segments; it is not clear whether this is due tofailed muscle determination or poor migration of mesodermcells into the cephalic region.
Neurogenic mutant embryos exhibit a hypertrophy of cellsthat express nautilus (nau) or vestigial (vg) (Corbin et al.,1991; Bate et al., 1993), which encode transcription factors thatare normally expressed in different sets of muscle founders(Michelson et al., 1990; Paterson et al., 1990; Williams et al.,1991). In wild-type embryos, vg is expressed in two longi-
ically different stage 12 (7.5-9.5 hours AEL) embryos are labeled withtion of the N+, UAS-N and UAS-∆ECN products. Anterior is to thestock exhibits the normal, ubiquitous distribution of Notch protein.gly expressed in the nervous system, epidermis and midgut. Note thats N54l9, UAS-N107 /Y embryo makes no Notch protein. These embryos+ sibling (partly shown in upper left corner) was labeled under identicales Notch in the mesoderm and endoderm. Expression in the expression). Notch is not detected in either the dorsal or ventralAS-∆ECN in the mesoderm, showing the relative levels expression ofsodermal signal).

621Notch functions in the mesoderm
Fig. 3. Epidermal development of N54l9 embryos that express UAS-N or UAS-∆ECN in different germ layers. Anterior is left; scale bars = 50µm. (A) Normal cuticle from a N+ embryo. Ventral denticle belts (black arrows) are produced on the ventral surface of each segment. Theclypeopharyngeal skeleton (white arrow), or CS, and the ‘Filzkörper’ (fk) of the posterior spiracles are indicated. (B) Cuticle from ahemizygous N54l9, UAS-N107 embryo, consisting of reduced dorsal cuticle and posterior spiracles (indicated by the birefringent fk). Ventralepidermis and the CS are absent. (C, D) N54l9 embryos that express UAS-N107 (C) or UAS-∆ECN 3 (D) under control of 69B, which expressesGAL4 throughout the ectoderm and mesoderm. (C) Cuticle from an unhatched embryo produced from the cross N54l9, UAS-N107 /winscy X69B/69B. Approximately one-quarter (22/79) of the progeny from this cross (presumably N54l9 hemizygotes) exhibit head defects (white arrow)and do not hatch. (D) Cuticle from an embryo derived from the cross N54l9/winscy ; UAS-∆ECN 3 / UAS-∆ECN 3 X 69B/69B. Although none ofthe progeny hatch, all generate a normal ventral cuticle. One-quarter of the embryos (17/71), like the one shown here, have an abnormalseparation and extrusion of scleretized CS components (white arrows), suggesting that they are hemizygous for N54l9. (E-G) N54l9 embryos thatexpress UAS-N107 (E,F) or UAS-∆ECN 3 (G) under control of twi-GAL41 exhibit very different cuticle phenotypes. (E,F) Two focal planes of acuticle from a Ntwi embryo derived from the cross N54l9, UAS-N107 /winscy X twi-GAL41 / twi-GAL41. Roughly one-quarter (49/211) of theembryos from this cross lack endogenous N+ function and do not hatch; their cuticle is herniated anterior to the thoracic segments. There issubstantial rescue of ventral cuticle, indicated by the presence of ventral denticle belts (black arrow), which often fuse in reverse orientation(open arrow). (G) A ∆ECNtwi embryo, which represents one-quarter (35/132) of the progeny from the cross indicated in Fig. 1E; three-quartersof the progeny make normal cuticle but fail to hatch due to mesodermal defects. ∆ECNtwi embryo are indistinguishable from N54l9 hemizygotes,indicating that mesodermal expression of ∆ECN does not rescue epidermal development.
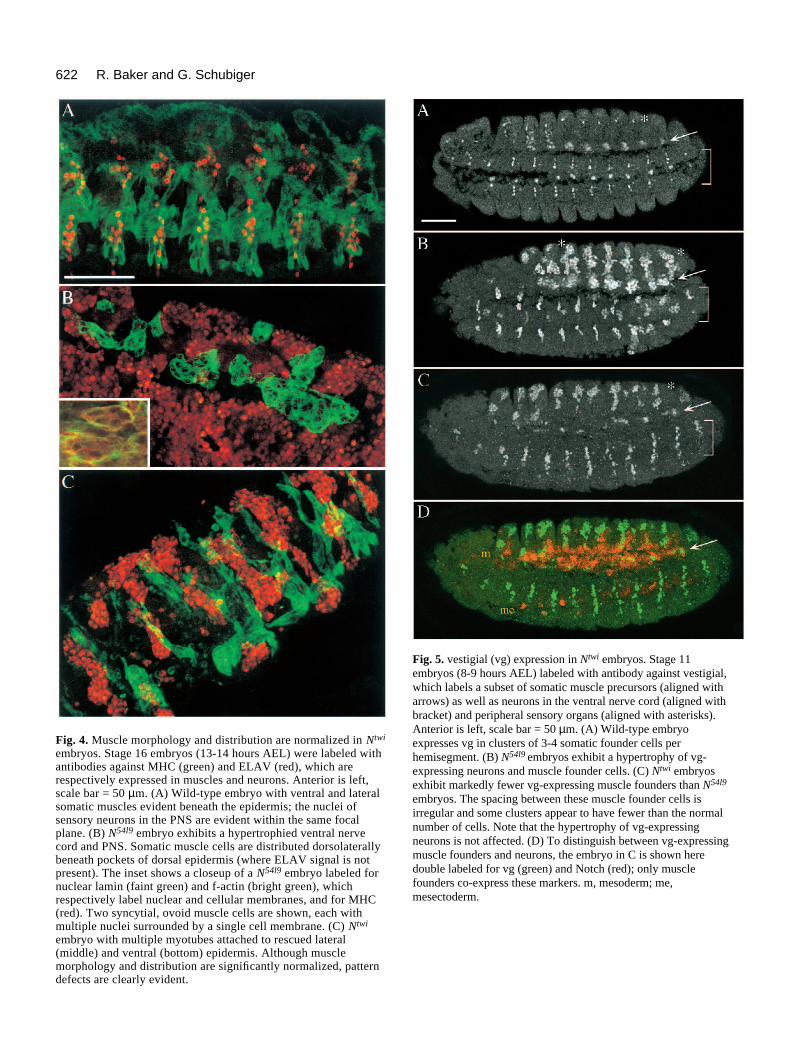
622 R. Baker and G. Schubiger
Fig. 4. Muscle morphology and distribution are normalized in Ntwi
embryos. Stage 16 embryos (13-14 hours AEL) were labeled withantibodies against MHC (green) and ELAV (red), which arerespectively expressed in muscles and neurons. Anterior is left,scale bar = 50 µm. (A) Wild-type embryo with ventral and lateralsomatic muscles evident beneath the epidermis; the nuclei ofsensory neurons in the PNS are evident within the same focalplane. (B) N54l9 embryo exhibits a hypertrophied ventral nervecord and PNS. Somatic muscle cells are distributed dorsolaterallybeneath pockets of dorsal epidermis (where ELAV signal is notpresent). The inset shows a closeup of a N54l9 embryo labeled fornuclear lamin (faint green) and f-actin (bright green), whichrespectively label nuclear and cellular membranes, and for MHC(red). Two syncytial, ovoid muscle cells are shown, each withmultiple nuclei surrounded by a single cell membrane. (C) Ntwi
embryo with multiple myotubes attached to rescued lateral(middle) and ventral (bottom) epidermis. Although musclemorphology and distribution are significantly normalized, patterndefects are clearly evident.
Fig. 5. vestigial (vg) expression in Ntwi embryos. Stage 11embryos (8-9 hours AEL) labeled with antibody against vestigial,which labels a subset of somatic muscle precursors (aligned witharrows) as well as neurons in the ventral nerve cord (aligned withbracket) and peripheral sensory organs (aligned with asterisks).Anterior is left, scale bar = 50 µm. (A) Wild-type embryoexpresses vg in clusters of 3-4 somatic founder cells perhemisegment. (B) N54l9 embryos exhibit a hypertrophy of vg-expressing neurons and muscle founder cells. (C) Ntwi embryosexhibit markedly fewer vg-expressing muscle founders than N54l9
embryos. The spacing between these muscle founder cells isirregular and some clusters appear to have fewer than the normalnumber of cells. Note that the hypertrophy of vg-expressingneurons is not affected. (D) To distinguish between vg-expressingmuscle founders and neurons, the embryo in C is shown heredouble labeled for vg (green) and Notch (red); only musclefounders co-express these markers. m, mesoderm; me,mesectoderm.

623Notch functions in the mesoderm
Fig. 6. nautilus (nau) expression in Ntwi embryos. Parasagittalsections of stage 11 embryos (8-9 hours AEL) labeled with antibodyagainst nautilus, which labels a subset of somatic muscle precursors.Anterior is to the left, scale bar = 50 µm. (A) Wild-type embryo. (B)N54l9 embryo exhibits hypertrophy of nau-expressing cells. Note thatmany clusters are contiguous between segments. (C) Ntwi embryoexhibits distinctive, nonhypertrophied clusters of nau-expressingcells. (D) Same embryo and optical section as in C, labeled with anti-HRP antibody to reveal the hypertrophied nervous systemcharacteristic for Ntwi embryos. None of the embryos assayed (0/83)from the cross indicated in Fig. 1D exhibit the muscle founderhypertrophy characteristic for N54l9 embryos.
Fig. 7. ∆ECN expression in the mesoderm disrupts somaticmuscle development. Embryos shown were obtained from thecross N54l9/winscy; ∆ECN3/∆ECN3 × twiGAL1/twiGAL1. N+
embryos are shown; the muscle phenotype is identical in a N−
background. (A,B) Stage 14 embryo labeled with antibodyagainst MHC. Ventral (A) and dorsal (B) somatic myocytesremain largely unfused at the time muscle fusion is normallycompleted. Unlike myocytes in N− embryos, most MHC-expressing cells are spherical or vaguely spindle-shaped (inset toA). Visceral and cardiac muscle development is partiallydisrupted, although the large pharyngeal muscles appear todevelop normally (B). (C,D) Ventral perspective of a stage 11embryo double labeled for vg (green) and N (red). (C) vg isproperly expressed in the ventral nerve cord; note the elevatedlevels of ∆ECN in the midline cells (mesectoderm). (D) vg is alsoproperly expressed in the PNS but is almost absent from themesoderm (bright red cells in the trunk; compare with wild-typeembryo in Fig. 5A). Arrow indicates two mesoderm cells thatexpress both vg and N and hence appear yellow. Scale bars = 50µm (A-D) and 150 µm (inset to A).

624 R. Baker and G. Schubiger
tudinal rows of ventral myoblasts which flank the ventral nervecord (Fig. 5A). Supernumerary vg-expressing myoblasts arepresent in N54l9 embryos (Fig. 5B), but approximately normalnumbers are observed in Ntwi embryos (Fig. 5C). The hyper-trophy of nau-expressing muscle founders is similarly rescuedin Ntwi embryos (Fig. 6). However, muscle founder cell clustersare irregularly spaced in Ntwi embryos, which frequentlypossess less than the normal complement of vg- or nau-expressing cells. It is unlikely that the apparent paucity ofmuscle founders is due to overexpression of N in themesoderm, since expression of UAS-N in the mesoderm of N+
embryos has no effect on muscle development (not shown).Since a subset of ectodermal cells induces nau expression inthe mesoderm (Baker and Schubiger, 1995), it is possible thatdefects in the ectoderm of both N54l9 and Ntwi embryos con-tribute to the irregular spacing and variable numbers of musclefounders.
It is unclear to what extent improved muscle developmentin Ntwi embryos is due to cell-autonomous Notch functions, orto rescue of ventral epidermis that serves as a substratum formuscle migration and attachment. We attempted to address thisissue by analyzing muscle development in ∆ECNtwi embryos,which possess Notch activity in the mesoderm but do notrescue ventral epidermis. We observe, however, thatexpression of ∆ECN in the mesoderm strongly inhibits somaticmuscle development in either a N+ or N− genetic background(Fig. 7). Most MHC-expressing cells in the somatopleura aremononucleated and generally spherical (Fig 7A). ∆ECN doesnot inhibit muscle development per se, since the pharyngealmuscles are morphologically normal (Fig. 7B). Mesodermal∆ECN expression also appears to inhibit determination ofsomatic muscle founder cells; few or no cells in the somato-pleura express vg, which is properly expressed in the nervoussystem (Fig. 7C,D). Similarly, we were unable to detect nauexpression in the mesoderm of these embryos (data notshown).
DISCUSSION
Notch is required in mesoderm and ectoderm formuscle developmentWe show that expression of a full-length Notch minigene inthe mesoderm of N− embryos normalizes some, but not all,aspects of myogenesis. The hypertrophy of nautilus- andvestigial-expressing cells in the mesoderm is rescued in Ntwi
embryos and a hypotrophy of muscle founders is observedwhen an activated form of N (∆ECN) is expressed in themesoderm. These results suggest that N is requiredautonomously by mesoderm cells to establish the proper ratioof muscle founders. Notch-Delta signaling between mesodermcells may limit the number of muscle founders by a pathwayanalogous to lateral inhibition of neurogenesis in the ectoderm.This seems likely since Dl− embryos also exhibit musclefounder hypertrophy (Corbin et al., 1991; Hartenstein et al.,1992) and Dl is strongly expressed in subsets of mesodermalcells during early muscle differentiation (Kooh et al., 1993).
Several lines of evidence suggest that Notch is required inthe ectoderm for the morphogenesis of elongated somaticmyotubes. Notch is not required for myoblast fusion; confocalanalyses show that the cells that most strongly express MHC
in N54l9 embryos contain between 4 and 8 nuclei; this range issomewhat lower than normal (4-25 nuclei per muscle; Bate,1990), and may be due to having an increased large number ofmuscle founders fuse with a reduced number of fusioncompetent cells. Although myoblasts fuse in N54l9 embryos,very few generate elongated myotubes, which are invariablydistributed beneath the dorsal epidermis. The incompleterescue of myotube morphogenesis in Ntwi embryos is likely dueto the partial restoration of epidermis, and not due to a cell-autonomous function of N in the mesoderm; if it were due toa cell-autonomous function, one might expect that myotubedifferentiation would be more completely rescued, since N isexpressed throughout the mesoderm of Ntwi embryos.Moreover, the somatic muscle morphology of N embryos issimilar to that of l(1)myo embryos, in which the somaticmuscles begin to differentiate normally but ultimately fail toattach to the epidermis (Newman and Wright, 1981).
We therefore propose that Notch is normally required inboth germ layers for proper somatic muscle development: inthe mesoderm, to establish the number of muscle founders and,in the ectoderm, to promote development of the epidermis. Inan attempt to test this hypothesis, we analyzed muscle devel-opment in ∆ECNtwi embryos. Since these embryos possess Nactivity in the mesoderm without rescuing ventral epidermaldevelopment, we anticipated that they would properly expressvg and nau but would not show improvement in myotubedifferentiation. However, we observed that this activatedNotch protein strongly inhibits myoblast fusion and repressestranscription of putative muscle founder determinants; we weretherefore unable to determine by this experiment whethermuscle morphogenesis requires N in the mesoderm, ectoderm,or both germ layers. We have begun further experiments todetermine the extent to which targeted N expression in theectoderm can rescue differentiation of N− mesoderm cells.
Overexpression of Notch in the mesoderm rescuesepidermal developmentWe present evidence that N can nonautonomously induceepidermal development across germ layers. The crux of thisargument rests on the assumption that functioning N protein isnot present in the ectoderm at concentrations below thethreshold of detection. Although this assumption cannot bedirectly tested, indirect functional assays for ‘leaky’ UAS-Nand twi-GAL expression in the ectoderm have already beenmentioned. First, N54l9, UAS-N107 hemizygotes are phenotypi-cally N−, indicating that UAS-N is inactive in the absence ofGAL4. Second, ∆ECNtwi embryos secrete a cuticle indistin-guishable from N54l9 embryos. This strongly suggests that twi-GAL41 is not expressed in the ectoderm because ∆ECN canrescue epidermal development when expressed in the ectoderm(Fig. 3D). Although both UAS-N and UAS-∆ECN have thesame promoter elements, ‘leaky’ low-level transcription oftrangenes may depend on cis-acting enhancers near the site ofinsertion. It is possible, therefore, that, if twi-GAL4 wasexpressed at low levels in the ectoderm, then UAS-N wouldmore readily respond to activation than UAS-∆ECN. Thisexplanation seems unlikely, since we obtained identical resultswith two different transgenes for each allele (ventral epidermisfrom UAS-N67 and UAS-N107, no ventral epidermis from UAS-∆ECN2 and UAS-∆ECN3).
Previous reports have also detected a nonautonomous, epi-

625Notch functions in the mesoderm
dermalizing function for N in the embryonic neurogenicectoderm, although the result has been interpreted differently.Technau and Campos-Ortega (1987) generated mosaic Nembryos by homotopically transplanting ventral ectoderm cellsfrom N− embryos into wild-type hosts. They observed that thetransplanted N− cells differentiate as either neurons orepidermis, suggesting that N is not required autonomously forepidermal development. The opposite conclusion was reachedin the genetic mosaic studies of Hoppe and Greenspan (1990).They observed that, in the ventral ectoderm of gynandro-morphs, N− (XO) cells adjacent to N+ (XX) cells never formedepidermis, suggesting that N is autonomously required forepidermal development. However, Heitzler and Simpson(1991) have shown that, when adjacent proneural cells in thenotum express different amounts of N protein, the cell with thelower amount always adopts a neural fate. This is true even ifboth cells are N+ and thus intrinsically capable of generatingepidermis. If this principle applies to embryonic ectoderm, thenN− cells along the gynandromorph boundary should preferen-tially give rise to neurons, whether or not they are intrinsicallycapable of generating epidermis.
Hoppe and Greenspan (1990) also irradiated heterozygousN− embryos to generate small clones of mutant cells by mitoticrecombination, and observed that between 20 and 40% of theN− clones (depending on the timing of irradiation) differenti-ated epidermis. To reconcile this observation with a cellautonomous N function, the authors postulated that perduranceof N+ product in mutant clones accounts for epidermal devel-opment. Another interpretation of this result, consistent withthose presented here and by Technau and Campos-Ortega(1987), is that N plays a dual function in epidermal develop-ment by acting as both ligand and receptor in different,partially redundant signal transduction pathways.
Possible mechanisms for nonautonomous NotchfunctionIn Ntwi embryos, the mesoderm expresses excessive levels ofN relative to the ectoderm, a condition that is opposite tonormal development. This condition may result in a dispro-portionate amplification of minor Notch functions that previ-ously have been overlooked. For example, N may function asa weak epidermalizing ligand, since its extracellular domain ishighly homologous to that of Delta and Serrate ligands (Vässinet al., 1987; Fleming et al., 1990). If such a pathway normallyplays a minor role in epidermal development, it would explainwhy it rarely manifests itself in genetic mosaics (Technau andCampos-Ortega, 1987; Hoppe and Greenspan, 1990). Such afunction would readily manifest itself in Ntwi embryos,however, since they would have overabundant ligand activitybut no receptor function.
The main argument against this hypothesis is that, in aprevious study, ectopic expression of the Notch extracellulardomain (ECN) did not rescue epidermal development in N−
embryos (Lieber et al., 1993). However, this discrepancy couldbe due to differences in experimental design. Whereas Ntwi
embryos continuously express N in the mesoderm, the ECNconstruct of Lieber et al. (1993) was driven in pulses from aheat-shock promoter; neither the levels of expression nor thestability of their truncated protein was reported. It is thereforepossible that the ECN in these experiments was insufficient (inamount and/or duration) to promote epidermal development.
If Notch does function only as a receptor, several explana-tions may yet account for our observations. One possibility isthat N-Dl interactions between the germ layers of Ntwi embryosallows maternally derived N in the ectoderm to perdure longenough for lateral inhibition to operate during the first wave ofneuroblast delamination. Kooh et al. (1993) have observed sub-cellular vesicles containing either N and Dl, or Dl alone; theypropose that these molecules are endocytosed into N-express-ing cells and subsequently degraded. Since such vesicles arenever observed to contain only N (Kooh et al., 1993), it ispossible that binding with Dl (or Serrate) is required for Ndegradation. In Ntwi embryos, therefore, mesoderm-bound Nmight slow down the rate of maternal N degradation by effec-tively titrating Dl from the ectoderm. It will be instructive todetermine whether or not mesodermal UAS-N expression canrescue epidermal development in N− embryos lacking maternalN transcripts.
A second possibility is that mesodermal N disrupts someuncharacterized neuron-promoting function of Dl. Previousreports have shown that the severity of neurogenic defects inN− embryos is reduced in Dl− backgrounds (Brand andCampos-Ortega, 1990; Xu et al., 1990); conversely, theseverity of the phenotype is increased in N− embryos havingadditional copies of Dl+ (de la Concha et al., 1988). These phe-notypes are not consistent with the hypothesis that during neu-rogenesis, Dl only promotes epidermal development bybinding N. N−, Dp[Dl+] embryos have greater neural hyper-trophy than N− embryos (de la Concha et al., 1988) suggestingthat Dl may normally promote neural development by a N-independent pathway. If so, excessive levels of N in themesoderm may interfere with such a neuralizing function inNtwi embryos, thereby reducing the severity of the neurogenicphenotype.
In summary, we have presented evidence for a novel, non-autonomous Notch function which may or may not play amajor role in normal embryogenesis. If so, there exists a devel-opmental pathway whose elucidation will greatly enhance thepresent understanding of how proneural cells adopt epidermalor neural fates. If not, it is likely that the epidermalizingactivity observed in Ntwi embryos represents an evolutionaryvestige of an ancient Notch function which may yet operate inother species. This novel function could provide valuableinsights into the genetic circuitry that regulates cell determi-nation within all germ layers of Drosophila embryos and, byextension, vertebrate embryos.
We are grateful to Richard Fehon, Jin Jiang, Ed Giniger and AndreaBrand for providing fly stocks and DNA constructs; to SpyrosArtavanis-Tsakonas, Bruce Paterson and Barbara Taylor for anti-bodies and cDNAs; to Jayne Baker and Lisa Stiffler for technicalassistance; and to Raphael Kopan, Margrit Schubiger and Ed Ginigerfor helpful editorial critiques. This work was supported by NIH grantGM33656.
REFERENCES
Ashburner, M. (1989). Drosophila: A Laboratory Manual. Cold SpringHarbor, New York: Cold Spring Harbor Laboratory Press.
Baker, R. and Schubiger, G. (1995). Ectoderm induces muscle-specific geneexpression in Drosophila embryos. Development 121, 1387-1398.
Bate, M. (1990). The embryonic development of larval muscles in Drosophila.Development 110, 791-804.

626 R. Baker and G. Schubiger
Bate, M. (1993). The mesoderm and its derivatives. In The Development ofDrosophila melanogaster (ed. M. Bate and A. Martinez Arias), pp. 1013-1090. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press.
Bate, M., Rushton, E. and Currie, D. (1991).Cells with persistent twistexpression are the embryonic precursors of adult muscles in Drosophila.Development 113, 79-89.
Bate, M., Rushton, E. and Frasch, M. (1993). A dual requirement forneurogenic genes in Drosophila myogenesis. Development 1993Supplement 149-161.
Brand, A. H. and Perrimon, N. (1994). Targeted gene expression as a meansof altering cell fates and generating dominant phenotypes. Development 118,401-415.
Brand, M. and Campos-Ortega, J. A. (1990). Second site modifiers of thesplit mutation of Notch define genes involved in neurogenesis in Drosophilamelanogaster. Roux’s Arch. Dev. Biol. 198, 275-285.
Campos-Ortega, J. A. (1993). Early neurogenesis in Drosophilamelanogaster. In The Development of Drosophila melanogaster. (ed. M.Bate and A. Martinez Arias), pp. 1091-1130. Cold Spring Harbor, New York:Cold Spring Harbor Laboratory Press)
Castelli-Gair, J., Greig, S., Micklem, G. and Akam, M. (1994). Dissectingthe temporal requirements for homeotic gene function. Development 120,1983-1995.
Corbin, V., Michelson, A. M., Abmayr, S. M., Neel, V., Alcomo, E.,Maniatis, T. and Young, M. W. (1991). A role for the Drosophilaneurogenic genes in mesoderm differentiation. Cell 67, 311-323.
de la Concha, A., Dietrich, U., Weigel, D. and Campos-Ortega, J. A. (1988).Functional interactions of neurogenic genes in Drosophila melanogaster.Genetics 118, 499-508.
Fehon, R. G., Kooh, P. J., Rebay, I., Regan, C. L., Xu, T., Muskavitch, M.A. T. and Artavanis-Tsakonas, S. (1990). Molecular interactions of theprotein products of the neurogenic loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61, 525-533.
Fleming, R. J., Scottgale, T. N., Diederich, R. J. and Artavanis-Tsakonas,S. (1990). The gene Serrate encodes a putative EGF-like transmembranedomain essential for proper ectodermal development in Drosophilamelanogaster. Genes Dev. 4, 2188-2201.
Fortini, M. E., Rebay, I., Caron, L. A. and Artavanis-Tsakonas, S. (1993).An activated Notch receptor blocks cell-fate commitment in the developingDrosophila eye. Nature 365, 555-557.
Frasch, M. (1995). Induction of visceral mesoderm by ectodermal Dpp in theearly Drosophila embryo. Nature 374, 464-467.
Gehring, W. (1973). Genetic control of determination in the Drosophilaembryo. In (Genetic Mechanisms of Development: The 31st Symposium ofthe Society for Developmental Biology. New York: Academic Press Inc.
Hartenstein, A. Y., Rugendorff, A., Tepass, U. and Hartenstein, V. (1992).The function of the neurogenic genes during epithelial development in theDrosophila embryo. Development 116, 1203-1220.
Heitzler, P. and Simpson, P. (1991). The choice of cell fate in the epidermis ofDrosophila. Cell 64, 1083-1092.
Hoppe, P. E. and Greenspan, R. J. (1990). The Notch locus of Drosophila isrequired in epidermal cells for epidermal development. Development 109,875-885.
Jennings, B., Preiss, A., Delidakis, C. and Bray, S. (1994). The Notchsignaling pathway is required for Enhancer of split bHLH protein expressionduring neurogenesis in the Drosophila embryo. Development 120, 3537-3548.
Jiang, J., Kosman, D., Ip, Y. T. and Levine, M. (1991). The dorsalmorphogen gradient regulates the mesoderm determinant twist in earlyDrosophila embryos. Genes Dev. 5, 1881-1891.
Jiménez, F. and Campos-Ortega, J. A. (1990). Defective neuroblastcommitment in mutants of the achaete-scute complex and adjacent genes ofD. melanogaster. Neuron 5, 81-89.
Kooh, P. J., Fehon, R. G. and Muskavitch, M. A. T. (1993). Implications ofdynamic patterns of Delta and Notch expression for cellular interactionsduring Drosophila development. Development 117, 493-507.
Kopan, R., Nye, J. S. and Weintraub, H. (1994). The intracellular domain ofmouse Notch: a constitutively activated repressor of myogenesis directed atthe basic helix-loop-helix region of MyoD. Development 120, 2385-2396.
Lehmann, R., Jiménez, F., Dietrich, U. and Campos-Ortega, J. A. (1983).
On the phenotype and development of mutants of early neurogenesis inDrosophila melanogaster. Wilhelm Roux Arch. Dev. Biol. 192, 62-74.
Leptin, M., Bogaert, T., Lehmann, R. and Wilcox, M. (1989). The functionof PS integrins during Drosophila embryogenesis. Cell 56, 401-408.
Lieber, T., Kidd, S., Alcamo, E., Corbin, V. and Young, M. W. (1993).Antineurogenic phenotypes induced by truncated Notch proteins indicate arole in signal transduction and may point to a novel function for Notch innuclei. Genes Dev. 7, 1949-1965.
Maggert, K., Levine, M. and Frasch, M. (1995). The somatic-visceralsubdivision of the embryonic mesoderm is initiated by dorsal gradientthresholds in Drosophila. Development 121, 1207-1216.
Michelson, A. M., Abmayr, S. M., Bate, M., Martinez-Arias, A. andManiatis, T. (1990). Expression of a MyoD family member prefiguresmuscle pattern in Drosophila embryos. Genes Dev. 4, 2086-2097.
Newman, S. M., Jr. and Wright, T. R. F. (1981). Histological andultrastructural analysis of developmental defects produced by the mutationl(1)myospheroid in D. melanogaster. Dev. Biol. 86, 393-402.
Paterson, B. M., Walldorf, U., Eldredge, J., Dübendorfer, A., Frasch, M. andGehring, W. J. (1990). The Drosophila homologue of vertebrate myogenic-determination genes encodes a transiently expressed nuclear protein markingprimary myogenic cells. Proc. Natl. Acad. Sci. USA 88, 3782-3786.
Poulson, D. F. (1945). Chromosomal control of embryogenesis in Drosophila.Am. Nat. 79, 340-363.
Rebay, I., Fleming, R. J., Fehon, R. G., Cherbas, L., Cherbas, P. andArtavanis-Tsakonas, S. (1991). Specific repeats of Notch mediateinteractions with Delta and Serrate: Implications for Notch as amultifunctional receptor. Cell 67, 687-699.
Robinow, S. and White, K. (1991). Characterization and spatial distribution ofthe ELAV protein during Drosophila melanogaster development. J.Neurobiol. 22, 443-461.
Skeath, J. B. and Carroll, S. B. (1992). Regulation of proneural geneexpression and cell fate during neuroblast segregation in the Drosophilaembryo. Development 114, 939-946.
Staehling-Hampton, K., Hoffman, M., Baylies, F. M., Rushton, E. andBate, M. (1994). dpp induces mesodermal gene expression in Drosophila.Nature 373, 783-786.
Struhl, G., Fitzgerald, K. and Greenwald, I. (1993). Intrinsic activity of thelin-12 and Notch intracellular domains in vivo. Cell 74, 331-345.
Technau, G. and Campos-Ortega, J. A. (1987). Cell autonomy of expressionof neurogenic genes of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA84, 4500-4504.
Technau, G., Becker, T. and Campos-Ortega, J. A. (1988). Reversiblecommitment of neural and epidermal precursor cells during embryogenesisof Drosophila melanogaster. Roux’s Arch. Dev. Biol. 197, 413-418.
Thisse, B., Stoetzel, C., Gorostiza, T. C. and Perrin-Schmitt, F. (1988).Sequence of the twist gene and nuclear localization of its protein inendomesodermal cells of early Drosophila embryos. EMBO J. 7, 2175-2183.
Thummel, C. S., Boulet, A. M. and Lipshitz, H. D. (1988). Vectors forDrosophila P-element-mediated transformation and tissue culturetransformation. Gene 74, 445-456.
Vässin, H., Bremer, K. A., Knust, E. and Campos-Ortega, J. A. (1987). Theneurogenic locus Delta of Drosophila melanogaster is expressed inneurogenic territories and encodes a putative transmembrane protein withEGF-like repeats. EMBO J. 6, 3431-3440.
Volk, T. and VijayRaghaven, K. (1994). A central role for epidermal segmentborder cells in the induction of muscle patterning in the Drosophila embryo.Development 120, 59-70.
Wharton, K. A., Johansen, K. M., Xu, T. and Artavanis-Tsakonas, S.(1985). Nucleotide sequence from the neurogenic locus Notch implies a geneproduct that shares homology with proteins containing EGF-like repeats.Cell 43, 567-581.
Williams, J. A., Bell, J. B. and Carroll, S. B. (1991). Control of Drosophilawing and haltere development by the nuclear vestigial gene product. GenesDev. 5, 2481-2495.
Xu, T., Rebay, I., Fleming, R. J., Scottgale, T. N. and Artavanis-Tsakonas,S. (1990). The Notch locus and the genetic circuitry involved in earlyDrosophila neurogenesis. Genes Dev. 4, 464-475
(Accepted 6 November 1995)


















