Appendix A Isolation of Parasitic Protozoa from Infected ...978-1-349-15809-6/1.pdf · Appendix A...
24
Appendix A Isolation of Parasitic Protozoa from Infected Animals A.l Genus Trypanosoma (except T. cruz;,i) Before 1968, blood trypomastigotes for biochemical studies were usually isolated by simple differential centrifugation. This procedure often can yield preparations uncontaminated by host red and white blood cells but large numbers of platelets (up to 30 per cent by number) will be present. These cell fragments contain many enzyme activities such as the entire glycolytic sequence including lactate dehydrogenase (see chapter 4). When the platelets are removed using the technique of defibrination (which essentially involves allowing the blood to clot), the lactate dehydrogenase activities reported previously to be present in trypanosomes such as T. rhodesiense can no longer be detected. The consequence of this discovery is that the literature prior to 1968 on enzymatic studies of trypanosomes is treated with suspicion, unless the ·infected blood was defibrinated before centrifugation. Protozoan biochemists now take much more notice of the importance. of working with pure preparations of parasites. The current method of choice involves the use of columns of DEAE cellulose swollen in either a phosphate-saline-glucose (PSG) or Tris-saline-glucose (TSG) buffer of suitable pH and ionic strength (for exam- ple, PSG of pH 8.0, I = 0.217 for T. bruce!). The swollen cellulose is poured into a scintered glass funnel and the column of cellulose thus formed is thoroughly washed in PSG or TSG. Infected blood, collected with heparin as anticoagulant, is poured on to the top of the column. When the blood has com- pletely penetrated into it, the column is eluted with PSG or TSG. Red and white blood cells and platelets are adsorbed on to the cellulose; trypanosomes come through in the eluate essentially uncontaminated by host blood cells and can be collected by centrifugation. The factors which most influence the degree of separation achieved are the buffer (PSG or TSG) and its pH and ionic strength. The surface charges of both trypanosomes and mammalian host blood cells vary and so the optimum conditions for separation have had to be determined for each host-parasite combination. Recoveries of up to 100 per cent have been recorded with some species of trypanosomes and values rarely fall below 50 per cent. Highest recoveries are obtained when there is the greatest difference in surface charge between host blood cells (especially red cells) and parasites. Species isolated by
Transcript of Appendix A Isolation of Parasitic Protozoa from Infected ...978-1-349-15809-6/1.pdf · Appendix A...

Appendix A Isolation of Parasitic Protozoa from Infected Animals
A.l Genus Trypanosoma (except T. cruz;,i)
Before 1968, blood trypomastigotes for biochemical studies were usually isolated by simple differential centrifugation. This procedure often can yield preparations uncontaminated by host red and white blood cells but large numbers of platelets (up to 30 per cent by number) will be present. These cell fragments contain many enzyme activities such as the entire glycolytic sequence including lactate dehydrogenase (see chapter 4). When the platelets are removed using the technique of defibrination (which essentially involves allowing the blood to clot), the lactate dehydrogenase activities reported previously to be present in trypanosomes such as T. rhodesiense can no longer be detected. The consequence of this discovery is that the literature prior to 1968 on enzymatic studies of trypanosomes is treated with suspicion, unless the ·infected blood was defibrinated before centrifugation. Protozoan biochemists now take much more notice of the importance. of working with pure preparations of parasites.
The current method of choice involves the use of columns of DEAE cellulose swollen in either a phosphate-saline-glucose (PSG) or Tris-saline-glucose (TSG) buffer of suitable pH and ionic strength (for example, PSG of pH 8.0, I = 0.217 for T. bruce!). The swollen cellulose is poured into a scintered glass funnel and the column of cellulose thus formed is thoroughly washed in PSG or TSG. Infected blood, collected with heparin as anticoagulant, is poured on to the top of the column. When the blood has completely penetrated into it, the column is eluted with PSG or TSG. Red and white blood cells and platelets are adsorbed on to the cellulose; trypanosomes come through in the eluate essentially uncontaminated by host blood cells and can be collected by centrifugation.
The factors which most influence the degree of separation achieved are the buffer (PSG or TSG) and its pH and ionic strength. The surface charges of both trypanosomes and mammalian host blood cells vary and so the optimum conditions for separation have had to be determined for each host-parasite combination. Recoveries of up to 100 per cent have been recorded with some species of trypanosomes and values rarely fall below 50 per cent. Highest recoveries are obtained when there is the greatest difference in surface charge between host blood cells (especially red cells) and parasites. Species isolated by

APPENDIX A 149
this method are (in order of ease of separation): T. simiae (a parasite of pigs), T. evansi, T. gambiense, T. rhodesiense, T. brucei, T. congolense, T. vivax and T. lewisi. Trypanosomes isolated from DEAE cellulose columns are motile, normal in appearance and have lost none of their infectivity for their hosts.
None of the stages parasitic in the insect vector has yet been isolated in quantities sufficient for biochemical study although some preliminary work has been carried out with small numbers of mid-gut forms.
A.2 Trypanosoma crud
Blood trypomastigotes can be isolated by a DEAE cellulose column method similar to that used for other trypanosomes provided most of the red cells are first removed by differential centrifugation (500g for 5 min). The pH of the eluting buffer is critical (PSG, pH 7.5, I= 0.206) and organisms need to be collected in serum since they are very friable. Recoveries are about 50 per cent.
Preparation of the intracellular amastigote stages is more difficult. These have now been isolated from the hind skeletal muscle of rats, mice and chinchillas. The tissue is disrupted by homogenisation to release the parasites. The homogenate is then filtered through gauze to remove large debris and incubated with DNase, to destroy muscle cell nuclei, collagenase, to break down collagen-like proteins, and trypsin, to degrade other proteins. Parasites are then collected from the homogenate by differential centrifugation and further purified, if necessary, in a low-speed (225g for 5 min) linear sucrose gradient (0.25-{).70 M sucrose). Recoveries range from 30 to 70 per cent, contamination from 0.2 to 2.0 per cent, depending on whether a sucrose gradient step is used. Stages parasitic in the reduviid bugs have not yet been isolated.
A.3 Genus Leishmania
Leishmania donovani amastigotes have been isolated from spleen macrophages, though detailed methodology has yet to be published. The spleen is removed, homogenised in a phosphate-buffered saline solution and the amastigotes purified by differential centrifugation. L. mexicana amastigotes have been isolated from mouse lesions by coarse homogenisation, passage through tissue paper and washing in tissue culture medium 199. Promastigotes have not been isolated from sand flies.
A.4 Genus Eimeria
No-one has yet isolated trophozoites and schizonts from chicken intestinal epithelial cells. Methods for preparation from tissue cells have been published but have yet to be used in biochemical studies. In contrast, oocysts can be easily extracted and purified from the faeces of infected chickens. This is why most of the biochemical studies of Eimeria have been carried out with these stages. Merozoites and sporozoites also have been isolated.

150 BIOCHEMISTRY OF PARASITIC PROTOZOA
A.S Genus Toxoplasma
Trophozoites have been isolated from peritoneal macrophages. The macrophages are collected by centrifugation from peritoneal washings and shaken with glass beads to release the parasites. White cells are removed by passage of the homogenate through a sintered glass funnel or by density gradient centrifugation and red cells by immunolysis. After a thorough washing parasites are ready for use. None of the other stages has yet been isolated in quantities sufficient for biochemical study.
A.6 Genus Plasmodium
Intraerythrocytic stages can be isolated readily from infected blood. Since infection in many species develops synchronously, individual stages within the intraerythrocytic cycle can be purified by collecting the blood at the appropriate
Sucrose __ gradient
BEFORE 225g
5 min
0.25M
Linear sucrose gradient
0.70M
AFTER
-Platelets
Infected red ISO(SOO(<;x<l------ cell bond (brown)
- Red blood cell bond (red)
Figure A. I Isolation of malaria-infected red blood cells using a linear sucrose gradient.
time. The methodology used involves first removing white cells, platelets and as many uninfected red cells as possible and then releasing the parasites by disruption of the infected red cells. White cells are often removed by passage of the blood through columns of dry cellulose powder. Another method is to use a low speed (225g for 5 min) linear sucrose gradient (0.25-0. 70 M) which separates the infected red cells not only from the white cells and platelets but also from most of the non-infected red cells (figure A.l). The infected red cell band can be distinguished easily by its brown colour (due to malaria pigment-see section 1.10) and can readily be removed. Parasites can be

APPENDIX A 151
released from red cells by a number of methods including saponin lysis, nitrogen cavitation and disruption in the French press. The most gentle method involves immunolysis with anti-red cell serum in the presence of an excess of complement. It must be remembered, however, that none of these methods removes adequately the erythrocyte-derived plasma membrane by which the parasite becomes surrounded during penetration into the red cell. After immunolysis, parasites are collected and washed thoroughly by centrifugation. Recoveries are greater than 50 per cent and contamination by white cells is less than 0.1 per cent.
Merozoites have been obtained by incubating infected blood in a culture chamber fitted with a polycarbonate sieve of 2 p,m pore size. This allows merozoites (but not infected blood cells) to pass out of the chamber as they are released. Exoerythrocytic liver schizonts have not yet been isolated, but oocysts from mosquito gut and sporozoites from mosquito salivary glands have been obtained in small quantities.
A. 7 Further Reading
Lanham, S. H. and Godfrey, D. G. (1970). Isolation of salivarian trypanosomes from man and other mammals using DEAE cellulose. Expl Parasit., 28, 521-534
Williamson, J. and Cover, B. (1975). The rapid isolation from human blood of concentrated, white cell free preparations of Plasmodiumfalciparum. Trans. R. Soc. trop. Med. Hyg., 69, 78-87

Appendix 8 Culture of Parasitic Protozoa
B.l Genus Trypanosoma (except T. cruzi)
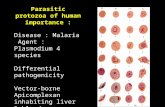
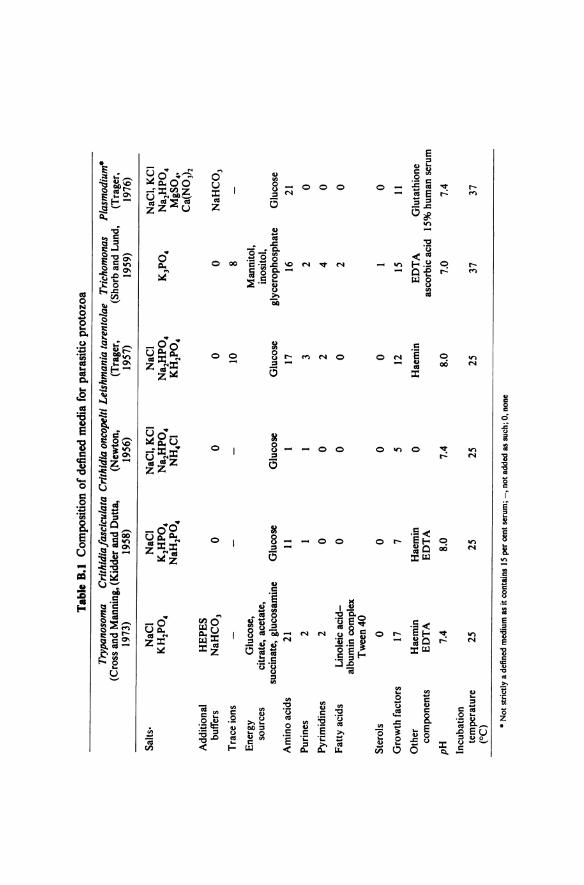
In the past, culture forms have been grown in both diphasic (for example, blood agar base with a buffered salts-glucose overlay in which the organisms grow) and monophasic (for example, Pittam's medium: tryptose-caesin hydrolysate-liver digest-buffered salts-glucose-blood extract) media. A defined medium is, however, now available and being used extensively (Cross and Manning's medium-table B.l). It contains haemin since a characteristic feature of trypanosomatids is an inability to synthesise the porphyrin ring. Note that cultures are incubated at vector temperatures (25 °C) not mammalian temperatures (3 7 °C).
Blood trypomastigote stages have been cultured for up to five generations in tissue culture media in the presence of a monolayer of mammalian tissue cells. Recently, the use of tissue culture medium RPMI 1640 over a layer of fibroblasts has been reported to support serial culture of blood-stream T. brucei and T. congo/ense at 37°C, although only at relatively low cell densities.
8.2 Trypanosoma crud
Culture epimastigote forms can be grown at 25 °C in a number of media including LIT (tryptose-liver infusion-buffered saline-glucose-haemin-calf serum). It has been reported also that they will grow in Cross and Manning's defined medium (table B.1). Blood trypomastigote stages do not grow and divide in vivo without prior transformation to intracellular amastigote stages so that, not surprisingly, these cannot be cultured serially. They have, however, been produced in large numbers in tissue culture. Amastigote stages have been cultured serially in Pan's medium (tissue culture medium 199-trypticase-haemin-bovine foetal serum-chicken plasma) at 35 °C but it is not clear how similar they are to intracellular amastigote stages. The latter can be grown in tissue culture at 37 °C.
B.3 Genus Crithidia (except C. oncopelti)
These model trypanosomatids will grow readily at 25 °C in undefined (for example, peptone-liver infusion-NaCl-glucose-haemin-adenine-folic acid)
Tabl
e B
. I
Com
posi
tion
of d
efin
ed m
edia
for
par
asiti
c pr
otoz
oa
Tryp
anos
oma
Cri
thid
iafa
scic
ulat
a C
rith
idia
onc
opel
ti Le
ishm
ania
tare
ntol
ae
Tric
hom
onas
Pl
asm
odiu
m•
(Cro
ss a
nd M
anni
ng, (
Kid
der
and
Dut
ta,
(New
ton,
(T
rage
r,
(Sho
rb a
nd L
und,
(T
rage
r,
I973
) I9
58)
I956
) I9
57)
I959
) I9
76)
Salts
· N
aCI
NaC
I N
aCI,
KC
I N
aCI
NaC
I,K
CI
KH
2P0
4 K
2HP
0 4
Na 2
HP
04
Na 2
HP
04
K3P
04
Na 2
HP
04
NaH
2P0
4 N
H4C
I K
H2P
04
MgS
0 4,
Ca(
N0
3)2
Add
ition
al
HE
PE
S
buff
ers
NaH
C0
3 0
0 0
0 N
aHC
03
Tra
ce io
ns
--
-IO
8
Ene
rgy
Glu
cose
, M
anni
tol,
sour
ces
citr
ate,
ace
tate
, in
osito
l, su
ccin
ate,
glu
cosa
min
e G
luco
se
Glu
cose
G
luco
se
glyc
erop
hosp
hate
G
luco
se
Am
ino
acid
s 2
I II
I
I7
I6
21
Puri
nes
2 I
I 3
2 0
Pyri
mid
ines
2
0 0
2 4
0 F
atty
aci
ds
Lin
olei
c ac
id-
0 0
0 2
0 al
bum
in c
ompl
ex
Tw
een4
0
Ster
ols
0 0
0 0
I 0
Gro
wth
fac
tors
I7
7
5 I2
I5
II
Oth
er
Hae
min
H
aem
in
0 H
aem
in
ED
TA
G
luta
thio
ne
com
pone
nts
ED
TA
E
DT
A
asco
rbic
aci
d I5
% h
uman
ser
um
pH
7.
4 8.
0 7.
4 8.
0 7.
0 7.
4
Incu
batio
n te
mpe
ratu
re
25
25
25
25
37
37
(OC)
• N
ot s
tric
tly
a de
fine
d m
ediu
m a
s it
cont
ains
IS
per
cen
t ser
um
;-, n
ot a
dded
as
such
; 0, n
one

154 BIOCHEMISTRY OF PARASITIC PROTOZOA
and defined (for example, Kidder and Dutta's medium-table B. I) media. The high levels of folic acid in these media can be replaced partially by addition of an unconjugated pteridine such as biopterin. A nutritional requirement for an unconjugated pteridine is unusual among microorganisms and has led to the use of C. fasciculata as a biological assay for these compounds.
8.4 Crithidia oncopelti
This trypanosomatid will grow at 25 °C in very simple undefined (peptone-NaCl-glucose) and defined (table B. I) media without haemin. The use of C. oncopelti in biochemical studies of trypanosomes, however, is best avoided (see appendix C).
B.S Genus Leishmania
Promastigote stages can be cultured at 25 °C in biphasic (for example blood agar base with an overlay of glucose-containing buffered salts) or monophasic (for example brain heart infusion with 10 per cent citrated, lysed, filtered human blood) media. Defined media (for example, Trager's medium-table 8.1) are also available. Amastigote stages can readily be grown at 3 7 °C in tissue culture.
8.6 Genus Trichomonas
Most parasitic protozoa are aerobic. Two exceptions are Trichomonas and Entamoeba which prefer anaerobic conditions. These are obtained by the addition of reducing compounds such as ascorbic acid to the medium. The metabolism of these organisms will also tend to produce an anaerobic environment. Both undefined (for example, Diamond's medium: trypticase-yeast extract-maltose-cysteine-ascorbic acid-agar-heat inactivated sheep serum) and defined (for example, Shorb and Lund's medium, table B. I) media can be used for Trichomonas. Cultures are incubated at 37 °C.
B. 7 Genus Entamoeba
Only undefined media (for example, Carter, Levy and Diamond's medium: peptone-liver digest-glucose-vitamins-serum) are available for axenic culture.
8.8 Genera Eimeria, Toxoplasma and Theileria
Trophozoites of Eimeria and Toxoplasma can be grown in tissue culture but these have been little used so far in biochemical studies. The exoerythrocytic stages of Theileria can be grown in transformed lymphoblasts and are used in experimental vaccine production. Recently, methods have been developed which support growth of all the mammalian stages.
Gro
wth
fac
tor
Thi
amin
e (v
itam
in 8
1)
Rib
ofla
vin
(vita
min
82)
Pyri
doxi
ne (v
itam
in 8
6)
or r
elat
ed c
ompo
unds
Pan
toth
enat
e
Co A
Bio
pter
in
Folic
aci
d
p-A
min
oben
zoic
aci
d
Bio
tin
Nic
otin
ic a
cid
Hae
min
Tab
le 8
.2 G
row
th f
acto
rs f
or p
aras
itic
pro
tozo
a
Req
uire
d by
Mos
t par
asit
ic p
roto
zoa
(not
Tri
chom
onas
)
Cri
thid
ia fa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Cri
thid
ia fa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Tric
hom
onas
Cri
thid
ia fa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Tric
hom
onas
Pla
smod
ium
loph
urae
Cri
thid
ia fa
scic
ulat
a
Cri
thid
iafa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Tric
hom
onas
Pla
smod
ium
Cri
thid
ia fa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Cri
thid
ia fa
scic
ulat
a Le
ishm
ania
tare
ntol
ae
Tric
hom
onas
gal
linae
Try
pano
som
atia
s
Pro
babl
e fu
nctio
n
Pre
curs
or o
f thi
amin
e py
roph
osph
ate,
a c
oenz
yme
in m
any
deca
rbox
ylat
ion
reac
tion
s
Pre
curs
or o
f FA
D a
nd F
MN
coe
nzym
es in
ele
ctro
n tr
ansf
er r
eact
ions
Pre
curs
or o
f pyr
idox
al p
hosp
hate
, a c
oenz
yme
in m
any
enzy
me
reac
tion
s, e
spec
ially
th
ose
invo
lvin
g tr
ansa
min
atio
n
Pre
curs
or o
f Co
A, i
mpo
rtan
t in
man
y re
acti
ons
as a
car
rier
of a
cety
l, m
alon
yl
and
succ
inyl
gro
ups
As
abov
e
In h
ydro
xyla
se r
eact
ions
incl
udin
g th
e de
satu
rati
on o
f fat
ty a
cids
and
the
sy
nthe
sis
of o
rota
te
Pre
curs
or o
ftet
rahy
drof
olat
e, im
port
ant i
n ac
tive
one
-car
bon
tran
sfer
re
acti
ons
In th
e sy
nthe
sis
of t
etra
hydr
ofol
ate
Coe
nzym
e in
car
boxy
lati
on r
eact
ions
Pre
curs
or o
f NA
D(P
)
Pro
sthe
tic
grou
p o
f cyt
ochr
omes
, cat
alas
e an
d pe
roxi
dase

156 BIOCHEMISTRY OF PARASITIC PROTOZOA
8.9 Genus Plasmodium
After many years of effort, serial culture of the intraerythrocytic stages was first obtained in 1976. Best results to date have been with P.falciparum, one of the human malaria parasites. One medium used is a tissue culture medium designated RPMI 1640 (developed originally for human leukocytes) supplemented with HE PES buffer and 1 S per cent human serum. A gas phase with 7 per cent C02, and a low oxygen tension (around S per cent) is provided. Human red blood cells infected at a low level with parasites (approximately 0.1 per cent) are held in a stationary layer and the medium is changed either daily by hand, or continuously by a slow flow method. The blood cells are diluted with fresh uninfected cells every third or fourth day. In the initial period of SO days, the original infected cell suspension was diluted more than 107 times. Parasites lost their synchrony and gametocytes were not seen but some infectivity to Aotus monkeys was retained. It is not clear which of the particular features of this system are critical in obtaining success; most likely this is a result of selection of the correct permutation of all the possible combinations. No other stages can be cultured serially.
8.10 General Conclusions
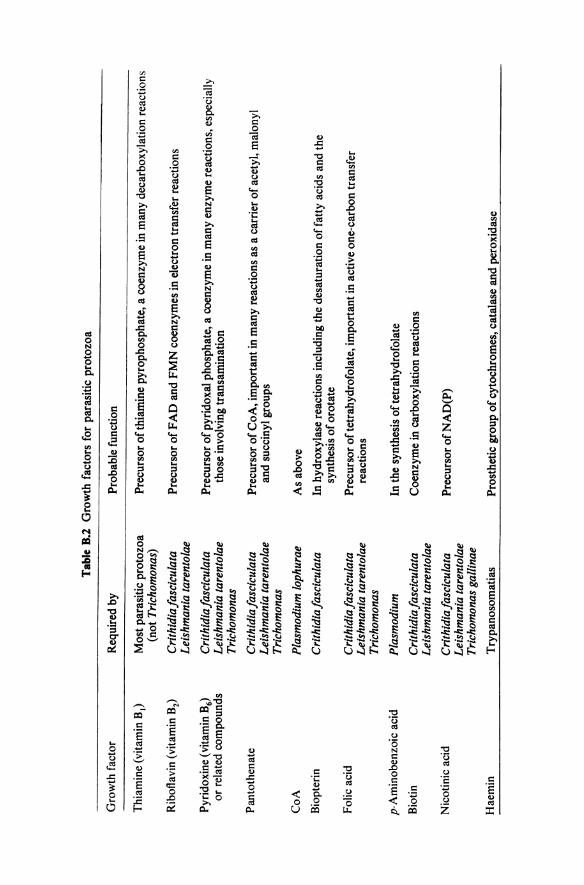
It is clear from the previous sections that although a number of undefined media are available for some genera of parasitic protozoa, few defined media have so far been produced. The composition of those that have are summarised in table B. I. With the exception of those for Crithidia species they are not minimal media so that their constituents do not allow conclusions to be drawn about the overall biosynthetic capabilities of the parasites. However, all seem to require at least some growth factors. A list of the more common ones and their likely biochemical role is given in table B.2.
8.11 Further Reading
Cross, G. A.M. and Manning, J. C. (1973). Cultivation of Trypanosoma brucei sspp.
in semidefined and defined media. Parasitology, 67, 315-331 Kidder, G. W. and Dutta, B. N. (1958). The growth and nutrition of Crithidia
fasciculata. J. gen. Microbial., 18, 621-638 Newton, B. A. (1956). A synthetic growth medium for the trypanosomatid flagellate
Strigononas (Herpetomonas) oncopelti. Nature, 177, 279-280
Shorb, M. S. and Lund, P. G. (1959). Requirements of trichomonads for unidentified
growth factors, saturated and unsaturated fatty acids. J. Protozoal., 6, 122-130
Taylor, A. E. R. and Baker, J. R. (1968). The cultivation of parasites in vitro. Blackwell Scientific Publications, Oxford
Trager, W. (1957). Nutrition of a hemoflagellate (Leishmania tarentolae) having an in
terchangeable requirement for choline or pyridoxal. J. Protozoal., 4, 269-276

APPENDIX B 157
Trager, W. (1976). Prolonged cultivation of malaria parasites Plasmodium coatneyi and P. falciparum. In Biochemistry of Parasites and Host-Parasite Relationships (ed. H. Van den Bossche), Elsevier-North Holland Biomedical Press, Amsterdam, pp.427-434

Appendix C The Enigma of Crithidia oncopelti
C.l The Presence of Bipolar Bodies
Investigations of the biochemistry of trypanosomes are, as was indicated in section 3.2, still seriously hampered by problems of culturing these organisms in vitro. One way round these difficulties is to use some of the readily cultured insect trypanosomatids (especially members of the genus Crithidia) as model systems. By far the easiest of these to handle is C. oncopelti, which has thus been used in a number of investigations.
Figure C.l Light photomicrograph of a fixed and stained preparation of Crithidia oncopelti showing the presence of bipolar bodies (1000). N, nucleus; K, kinetoplast; B, bipolar bodies.
Reference has already been made in appendix B to the fact that C. oncope/ti will grow in a simple, completely defined medium, consisting of salts, glucose, methionine, adenine and five growth factors (see table B.l). This situation contrasts with the complex media required for other trypanosomatids. A lack of requirement for haemin, from which, for example, the prosthetic groups of cytochromes and other enzymes are synthesised, is a further feature unique to C. oncope/ti among the trypanosomatids. Another unusual feature of the organism is the presence of basophilic, rod-shaped particles in the cytoplasm (figure C.l). C. oncopelti contains one or two of these structures which are located at the posterior end of the cell. When they were discovered in 19 57, they were named bipolar bodies because of their characteristic bipolar appearance in Giemsa-stained preparations. They were shown to be composed mainly of ribonucleoprotein but no suggestion was made at the time as to what they represented.

APPENDIX C 159
C.l Initial Conclusions
Two distinct pathways are known to occur in nature for the biosynthesis of the amino acid lysine. a-e-Diaminopimelic acid is a key intermediate in one of the pathways (present in bacteria, algae, plants and a few fungi) and a-aminoadipic acid in the other (present in most fungi and euglenids). Animal cells and most protozoa are unable to synthesise lysine. However, in 1962, it was reported that C. oncopelti synthesises lysine by the diaminopimelic acid pathway, which, as indicated above, is characteristic of bacteria. Diaminopimelic acid decarboxylase, the terminal enzyme in this pathway, was concentrated in that fraction of the broken cell preparation of the organism which contained the bipolar body. If C. oncopelti was grown in a complex medium containing penicillin G (6 mg/ml), the bipolar bodies disappeared. It was suggested therefore that bipolar bodies are endosymbiotic bacteria which synthesise metabolites such as lysine for the host cell. Such a phenomenon would explain the simple nutritional requirements of C. oncopelti. This view. was strengthened by a finding in 1963 that a DNA component, which could be distinguished, by means of its density in CsC 1 gradients, from those in the nucleus and kinetoplast of the organism, is associated with the bipolar bodies. Further support came from a report that the bipolar bodies in C. oncopelti disappeared after repeated subculture in a complex medium containing haemin. They could not, however, be cultured free from the protozoan.
C.3 Doubts
The conclusion that the bipolar bodies are endosymbiotic bacteria was questioned, however, when further evidence became available. First, it was shown that diaminopimelic acid decarboxylase was present in C. fasciculata, which does not contain bipolar bodies. Secondly, it became apparent that penicillin was not causing the elimination of the bipolar bodies but merely their apparent disappearance by reduction of their staining intensity. When organisms were transferred back to drug-free media, they were soon detectable again. Thirdly, it became increasingly clear that unique varieties of DNA exist in other cell organelles such as chloroplasts and mitochondria. Fourth and last, other workers failed to eliminate the bipolar bodies from C. oncopelti using the original technique. Thus, when a comprehensive literature review was made in 1968, it was concluded that there was no unequivocal evidence that the bipolar bodies were of bacterial origin.
C.4 The Resolution of the Problem
However, in 1971, a re-examination of the ultrastructure of the bipolar body by electron microscopy showed that it was limited by two unit membranes mutually separated by a lucent zone and that focal outpouchings into the cytoplasm occurred (figure C.2). This outer structure is similar to that of

160 BIOCHEMISTRY OF PARASITIC PROTOZOA
protoplasts (cell wall material removed by enzyme digestion) of Gram-negative bacteria showing everted mesosomes.
Then, in 1974, it was reported first that the antibacterial antibiotic, chloramphenicol, affected the ultrastructure of the bipolar body and subsequently that bipolar bodies could be eliminated from cultures by a single onemonth period of incubation with the drug. The bipolar-body-free strain grew more slowly than the original strain, suggesting that the bipolar bodies did indeed supply nutrients to the protozoa. Also at this time, methods were developed to prepare relatively clean preparations of bipolar bodies from cells disrupted by immune lysis or sonication.
Figure C.2 Electron micrograph ofthe bipolar body in Crithidia oncopelti(x 30 ()()()).
The bipolar-body-free strain was shown to require haemin for growth. The requirement could be replaced by protoporphyrin IX, indicating that the flagellate contains ferrochelatase, the terminal enzyme in the haem biosynthetic pathway, but not by d-aminolevulinic acid or porphobilinogen (see figure C.3). Microassay of uroporphyrinogen synthetase revealed that the specific activity was high in the strain containing bipolar bodies, higher still in isolated bipolar bodies but negligible in the bipolar-body-free strain. It was concluded therefore that the bipolar body augments a very limited (or even non-existent) haem biosynthetic capacity of host flagellates by supplying uroporphyrinogen synthetase and perhaps other enzymes preceding ferrochelatase in the haem biosynthetic chain.
Further work showed that the DNA isolated from purified bipolar body preparations had renaturation kinetics indicative of a genome complexity similar to that in the bacterium Escherichia coli and far higher than that of any organelle DNAs, including kinetoplast DNA, which have so far been examined. It was also found that \Pe bipolar bodies contain 67S ribonucleoprotein particles (compared with 87S'for cytoplasmic ribosomes) and RNA components of identical molecular weights to those in E. coli and quite distinct from cytoplasmic rRNA (see table 6.2).
Thus, it is now generally agreed that the bipolar bodies in C. oncopelti do in

APPENDIX C 161
Glycine + Succinyl CoA
8 - Aminolevulinic acid
l Porphobilinogen
11 Uroporphyrinogen
1 Coproporphyrinogen
l Protoporphyrin lX
Fe-'i2
Haem
Figure C.3 Pathway for the biosynthesis of haem. Enzymes: 1, Uroporphyrinogen synthetase; 2, ferroche1atase.
fact represent endosymbiotic bacteria, their lack of sensitivity to penicillin being due to the absence of cell wall material since they are present as protoplasts. Their use in biochemical studies of parasitic trypanosomes is thus best avoided and care should be taken in the interpretation of experiments in which they have been used.
C.S Further Reading
Chang, K-P. (1974). Ultrastructure of symbiotic bacteria in normal and antibiotic treated Blastocrithidia culicis and Crithidia oncope/ti. J. Protozool., 21, 699-707
Chang, K-P. (1975). Reduced growth of Blastocrithidia culicis arid Crithidia oncopelti freed of intracellular symbiotes by chloramphenicol. J. Protozoo/., 22, 271-2 76
Chang, K-P., Chang, C. S. and Sassa, S. (1975). Heme biosynthesis in bacteriumprotozoon symbioses: enzymic defect in host hemofiagellates and complementary role of the intracellular symbiotes. Proc. natn. A cad. Sci. U.S A., 12, 2979-2983
Newton, B. A. (1968). Biochemical peculiarities of trypanosomatid flagellates. A. Rev. Microbiol., 22, 109-130
Spencer, R. and Cross, G. A. M. (1975). Purification and properties of nucleic acids

162 BIOCHEMISTRY OF PARASITIC PROTOZOA
from an unusual cytoplasmic organelle in the flagellate protozoan Crithidia oncopelti. Biochim. Biophys. Acta, 390, 141-154
Tuan, R. S. and Chang, K-P. (1975). Isolation of intracellular symbionts by immune lysis of flagellate protozoa and characterisation of their DNA. J. Cell Bioi., 65, 309-323
Vogel, H. J. (1965). Lysine biosynthesis and evolution. In Evolving Genes and Proteins (ed. V. Bryson and H. J. Vogel), Academic Press, New York, pp. 25-40

Appendix D Biochemical Protozoology Literature
D.l Literature Searches Now that you have read this book, you might be interested in following some of the topics further. The suggestions for further reading given at the end of each chapter have been selected with this in mind since most contain a detailed list of references. Consultation of these and the literature that they cite makes it possible to build up quickly a comprehensive list of papers in any given area.
This process can of course be short-circuited, if funds are available, by the commission of a search by one of the commercial organisations which specialises in this field. Alternatively, the searching can be done personally, using either Chemical Abstracts (Chemical Abstracts Service, Columbus, Ohio) which covers most of the biochemical protozoology literature or Protozoological Abstracts (Commonwealth Agricultural Bureaux, Slough, England) Personal searches can be cross checked against advanced texts (see section D.3) and reviews that appear from time to time in proceedings oflnternational Protozoology and Parasitology Congresses, Annual Reviews of Biochemistry, Annual Reviews of Microbiology (both Academic Press, New York) and some of the parasitology journals (especially Experimental Parasitology-Academic Press, New York).
0.2 Current Awareness
A limitation of all literature search procedures is that there is a lag of up to twelve months between the publishing of a paper and its appearance in these systems. Current awareness searches are available commercially but tend to be expensive. Most people therefore resort to either scanning the title pages of journals or consulting Current Contents (Life Sciences section) (Institute for Scientific Information, Philadelphia). An indication of the journals in which you are most likely to find papers in the area of biochemical protozoology is given in table D.l. Note the wide spread of biochemical, pharmacological, cytological, parasitological and microbiological journals involved. Top of the list is the Journal of Protozoology, the official journal published by the International Society of Protozoologists. This society is based in the U.S.A. but sections exist in many other countries (for example, United Kingdom, France,

164 BIOCHEMISTRY OF PARASITIC PROTOZOA
Table D.l Source of references for the biochemical protozoology literature•
Position Journal %of total references
1 Journal of Protozoology 9.7 2 Biochimica Biophysica Acta 6.6 3 Experimental Parasitology 5.9 4 Nature 5.2 5 Annals of Tropical Medicine and Parasitology 4.1 6= Comparative Biochemistry and Physiology 3.8 6= Journal of Parasitology 3.8 8 American Journal of Tropical Medicine and Hygiene 3.5 9 Parasitology 3.2
10 Biochemical and Biophysical Research Communications 3.0 11= Archives of Biochemistry and Biophysics 2.3 11= Biochemical Pharmacology 2.3 11= Zeitschriftfur Tropenmedizin un Parasitologie 2.3 14 Transactions of the Royal Society of Tropical Medicine 2.2
and Hygiene 15 Biochemistry 2.1 16 Proceedings of the National Academy of Sciences of the U.S.A. 2.0 17 Science 1.9 18 Journal of Biological Chemistry 1.7 19 European Journal of Biochemistry 1.6 20= Journal of Cellular Biology 1.5 20= Journal of Medicinal Chemistry 1.5 20= Revista do Instituto de Medicina Tropical de Sao Paulo 1.5 23= International Journal of Parasitology 1.4 23= Journal of Bacteriology 1.4 25= BiochemicalJournal 1.3 25= Molecular Pharmacology 1.3 27= F.E.B.S. Letters 1.0 27= International Journal of Biochemistry 1.0 27= Journal of General Microbiology 1.0 27= Journal of Molecular Biology 1.0 27= Antimicrobial Agents and Chemotherapy 1.0
All other journals(< 1% each) 17.9
• Based on a survey of954 references indexed as being of interest to biochemical protozoologists (though not necessarily directly on protozoa) during the period January 1970 to July 1976
Italy, Czechoslovakia). Membership of the society is to be encouraged since not only does it make it possible to obtain theJournal of Protozoology at a reduced rate but attendance at its meetings enables one to be aware of the progress being made in our understanding of biochemical protozoology long before information is published in journals.
0.3 Advanced Texts
Kidder, G. W. (ed.) (1967). Chemical Zoology, vol. I, Protozoa. Academic Press, New York

APPENDIX D 165
Lehninger, A. L. (1975). Biochemistry. Second edition, Worth Publishers, Inc., New York
Van den Bossche, H. (ed.) (1972). Comparative Biochemistry of Parasites. Academic Press, New York
Van den Bossche, H. (ed.) (1976). Biochemistry of Parasites and Host-Parasite Relationships. Elsevier-North Holland Biomedical Press, Amsterdam
Von Brand, T. (1973). Biochemistry of Parasites. Second edition, Academic Press, New York

Index No attempt has been made to produce a comprehensive index since we regard this more as a book to read than as a work of reference. However, there is a full list of contents at the beginning of the book and the text is cross-referenced extensively. Only information which cannot be found readily from these sources has been included in this index.
Acetate 40, 45, 46, 48, 49, 50, 51, 55, 60, 65, 117, 119
Acetate kinase 51 Acetyl carnitine 51, 62 Acetyl carnitine hydrolase 51 Acetylcholine 114 Acetyl CoA 33, 35, 37, 46, 50, 51, 97,
116, 117, 118, 119 Acetyl CoA synthetase 51 Acetyl phosphate 50, 51 Acetyl thiokinase 46, 51 Aconitase 3 7 Acylglycerols 108, 109-10 Adenine 72, 73 Adenosine 72, 73 Adenosine deaminase 72, 73, 74 Adenosine diphosphate (ADP) 32, 72 Adenosine kinase 73, 74 Adenosine monophosphate (AMP) 32,
70, 72 Adenosine triphosphate (ATP) 32, 35,
38, 119 Adenylate cyclase 73, 106 Adenylosuccinate lyase 74 Adenylosuccinate synthetase 74 Alanine 60, 61, 62, 66, 93, 94, 97 Alanine aminotransferase 60, 61, 62,
63 Alcohol dehydrogenase 50 Amastigote 9 Amino acids 34, 89, 90, 91, 93, 97 Amino acid oxidase 97, 99, 104 Amino acid tRNA synthetase 95, 96 p-Aminobenzoic acid 130, 133, 155 Aminonucleoside of puromycin 75 Aminoquinaldines 22, 139, 144 4-Aminoquinolines 23, 135, 144 8-Aminoquinolines 21, 23, 122, 126 Aminotransferases 94, 97, 98, 103 Ammonia 74, 90, 93, 97, 98, 99 Amoebae 3 Amphotericin B 21, 22, 127, 128, 142,
143
Amprolium 22, 23, 129, 142, 143 Amylases 45, 48 Amylopectin 54, 66 Amylopectin phosphorylase 66 Amytal 38, 63 Anopheles 5, 15 Antigenic variation 100 Antimonials 22, 122 Antimycin A 38 Antrycide 20, 22, 24, 95, 139, 142, 143 Arsenicals 20, 22, 122 Aspartate 60, 61, 69, 74, 76, 93, 94, 97 Aspartate aminotransferase 60, 61 Aspartate carbamoyltransferase 76 ATPase 63, 65, 125
Base composition 81 Berenil 20, 22, 23, 24, 137, 142, 143 Bicarbonate 74, 75 Biochemical taxonomy 82, 106-7 Biopterin 117, 154, 155 Biotin 155 Bipolar bodies 73, 94, 158-62 Buoyant density of DNA 81, 82 Buquinolate 22, 125, 142, 143
Carbamoyl phosphate 76 Carbanilide 22, 139, 140, 142, 143 Carbon dioxide 34, 40, 45, 56, 60, 65,
69 Cardiolipin 47, Ill Carnitine 51, 119 Carnitine acyltransferase 51, 62, 63 Catalase 103, 104, 105 Cell cycle 81 Chagas' disease 5, 10, 20, 22 Chemotherapeutic index 20 Chloramphenicol 95, 96, 121, 122, 160 m-Chlorobenzhydroxamic acid 42 Chloroquine 21, 23, 24, 135, 136, 138,
143, 144

168
Cholesterol 113,114, 115,116 Choline 110, 118 Citrate 37 Citrate synthase 37, 64 Classification 1, 3 Clindamycin 23, 24, 95, 139, 140, 142,
143 Cloning 29 Clopidol 22, 139, 140, 143 Coccidia 12 Cordycepin 75 Cristae 39 Culture forms 9, 26, 59, 152 Cyanide 38, 62, 105 3' ,5'-Cyclic AMP 73, 105-6 Cyclic AMP phosphodiesterase 73, 106 Cycloheximide 95, 96 Cyclopropane 109, 117 Cysts 5, 12, 13, 14, 66, 96 Cytidine 77 Cytidine deaminase 77 Cytidine triphosphate synthetase 77 Cytochromes 37, 38, 62, 64, 66, 75, 78,
106, 125 Cytochrome oxidase 38, 55 Cytosine 77
Dapsone 21, 23, 130, 142, 143 DEAE cellulose chromatography 29,
148 Decoquinate 22, 125, 142, 143 Defined media 29, 153, 156 Deoxycytidine 77, 79 Deoxycytidine kinase 77, 79 Deoxycytidylate aminohydrolase 77 Deoxyribonuclease 87 Deoxyribose 69 Deoxythymidine monophosphate
(dTMP) 71, 79, 80 Deoxythymidylate synthase see
thymidylate synthase Deoxyuridine monophosphate
(dUMP) 71 Diamidines 22, 23, 137, 138, 144 a-&-Diaminopimelic acid 159 a-&-Diaminopimelic acid decarboxylase
159 2,4-Diaminopyrimidines 22, 23, 79,
129, 131, 133, 134 Diaveridine 22, 130, 133, 142, 143 Differential centrifugation 29, 78, 148 Dihydrofolate 79, 80, 131, 132, 133
INDEX
Dihydrofolate reductase 79, 80, 88, 124, 131, 132, 133, 134, 144
Dihydroorotase 76, 78 Dihydroorotate 75, 76 Dihydroorotate dehydrogenase 75, 76,
78, 88, 125, 126 Dihydroorotate hydroxylase 78, 88 Dihydropteroate 130, 131, 132 Dihydropteroate synthetase 130, 131,
132 Dihydroxyacetone phosphate 41 Dimidium 20, 22, 135, 142, 143 Diseases 5 DNA 47, 69, 81, 82, 83, 94
base composition 81, 83 kinetoplast 81,84-6,88, 138 melting temperature 81-2 mitochondria 82, 83 polymerase 81, 135 repair 81
Dyskinetoplasty 86
East coast fever 5, 15 Ehrlich 20 Epimastigote 9 Ergosterol 113, 114, 115, 119 Ethanol 49, 50, 52 Ethanolamine 110 Ethidium bromide 20, 22, 24, 135, 142,
143 Ethopabate 22, 23, 129, 130, 142, 143 Exoerythrocytic schizonts 14, 15
Fa tty acids 34, 65, 108-9, 116, 119 Fatty acid synthesis 116 Ferredoxin 46, 53, 138 Ferrochelatase 160, 161 Flagellates 3, 4 Flavodoxin 47, 53 Flavoproteins 38, 46, 79 5-Fluorouracil 78 Folate 80, 93, 131, 133, 155 Folate reductase 80, 133 Formate 55, 69, 72, 154 F ormylmethioninesulphone-methyl-
phosphate (FMSMP) 101, 102 Fructose 40 Fructose-1,6-diphosphate 49 Fructose-6-phosphate 42, 49 Fumarate 37, 46, 62, 97 Fumarate hydratase 37, 46 Fumarate reductase 46, 62

INDEX
Galactose 48, 102 P-Galactosidase 45, 54, 66 Gametes 13, 14 Gametocytes 13, 14, 15 Glossina 4 Glucose 32, 33, 39, 40 Glucose-1-phosphate 33 Glucose-6-phosphate 32, 35, 39 Glutamate 61, 93, 94, 97, 98, 133 Glutamate dehydrogenase 97, 99 Glyceraldehyde-3-phosphate 35, 42 Glycerol 40, 109, 118 L-a-Glycerophosphate 41, 117 L-a-Giycerophosphate dehydrogenase
41 L-a-Giycerophosphate oxidase 41, 43,
62, 64, 123, 124 Glycine 60, 69, 72, 73, 80, 93, 97, 117,
161 Glycogen 32, 33, 45, 48 Glycogen phosphorylase 34 Glycolysis 34, 35, 36, 45 Glycoproteins 89, 102-3 Growth factors 155, 156 Guanine 72 Guanosine 72 Guanosine triphosphate (GTP) 37, 46,
50, 80, 131, 132, 133
Haemin 105, 152, 155, 158 Haemoglobin 91, 92, 98, 105, 135 Hexokinase 49 Histidine 97, 105 Histidine-rich protein 105 Histones 82, 106 Hydrogen 45, 46, 48, 51 Hydrogen peroxide 50, 97, 103, 104,
105 Hydrogenase 46,53 Hydrogenosomes 12, 47, 83, 104, 105,
112 Hydroxamic acids 42 Hypoxanthine 72, 73 Hypoxanthine
phosphoribosyltransferase 74 p-Hydroxyphenyllactate 98 5-Hydroxytryptamine 100 Hydroxyurea 79
Imidocarb 24 Indole acetate 98, 99 Indole ethanol 98, 99
Indole lactate 98, 99 Indole pyruvate 98, 99 Inosine 72, 73 Inosine phosphorylase 73, 74 Intraerythrocytic schizonts 15 Iron-sulphur proteins, 46, 50, 53 Isocitrate 37 Isocitrate dehydrogenase 37, 103
Kala-azar 5, 11 a-Ketoglutarate 37, 60, 61, 97, 98 a-Ketoglutarate dehydrogenase 37 a-Keto-y-hydroxybutyrate 93, 97 Kinetoplast 4, 84 Kinetoplastida 4
169
Lactate 28, 34, 35, 40, 45, 55, 67, 103, 104
Lactate dehydrogenase 43, 45, 56, 99, 103, 104, 148
Lampit 20, 21, 22, 24, 137, 142, 143 Lincomycin 23, 24, 121, 122, 139, 140 Linoleic acid 108, 115 Linolenic acid 115 Lipase 33, 34, 115, 116, 119 Lysine 97, 159 Lysosomes 1, 34, 45, 87, 91, 92, 113,
116
Malaria 5, 14, 15, 23 Malate 37, 43, 45, 61, 66 Malate dehydrogenase 37, 46, 50 Malic enzyme 45, 50, 60, 61, 63 Mannose 40, 102 Mel B 20, 22, 24, 122 Melting temperature of DNA 81 Membrane proteins 100 Menoctone 23, 24, 56, 126, 142, 143 Merozoite 12, 13, 14, 15, 16, 105 Methionine 93, 97, 117, 119, 131 Methylation reactions 131, 134 Methylbenzoquate 23, 125, 142, 143 N5, N'0-Methylene tetrahydrofolate 79 Metronidazole 21, 22, 137, 138, 142,
143 Microbodies 41, 47, 103-4 Minocycline 23, 24, 95, 139, 140, 142,
143 Mitochondria 4, 34, 39, 95, 119 Monensin 22, 127, 128, 142, 143

170 NADH oxidase 47, 53 NADPH oxidase 53, 54 Nagana 4, 20 Nalidixic acid 121, 122, 125 Naphthoquinones 23, 56, 122, 126 Nicotinamide adenine dinucleotide
(NAD) 35,38 reduced (NADH) 35, 38, 104, 119
Nicotinamide adenine dinucleotide phosphate (NADP) 35, 38
reduced (NADPH) 35, 37, 99 Nicotinic acid 155 5-Nitrofurans 22, 136, 139 2-Nitroimidazoles 22, 136, 139 5-Nitroimidazoles 21, 22, 47, 51, 54,
137, 138 Norepinephrine 100 Nucleases 34, 87 Nucleosides 69, 106 Nucleoside hydrolase 72, 73, 74, 87 Nucleoside kinase 72, 77 Nucleoside phosphorylase 72, 77 Nucleotides 69, 70, 71 Nucleotide diphosphate kinase 72, 77 Nucleotide monophosphate kinase 72,
77 Nucleotide phosphatase 72, 77
Oleic acid 108, ll6, ll7 Oriental sore 5, II Ormetoprim 23, 130, 131, 133, 142,
143 Orotate 75, 76, 77, 78 Orotidine-5-phosphate
decarboxylase 76 Orotidine-5 -phosphate
pyrophosphorylase 76 Oxaloacetate 37, 43, 45, 61, 66, 97 Oxygen 36, 37, 77, 104
Palmitic acid 108, ll6 Pantothenate 155 Penicillins 20, 29, 121, 122, 159 Pentamidine 20, 21, 22, 24, 137, 138,
142, 143, 144 Pentoses 35, 37, 60 Pentostam 21, 22, 123, 142, 143 Peroxidase 105 Peroxisomes 97, 103, 105 Phagocytosis 91 Phenanthridines 22, 135, 138, 144 Phenylalanine 97, 98, I 00
INDEX
Phlorrhizin 48 Phenylpyruvate 98 Phosphatases 34, 87 Phosphate acyltransferase 51 Phosphatidylcholine Ill, 118 Phosphatidylethanolamine Ill, 118 Phosphatidylinositol Ill Phosphatidylserine Ill, 118 Phosphoenolpyruvate 43, 61, 66, 93, 94 Phosphoenolpyruvate carboxykinase
(GTP) 45, 61, 63 Phosphoenolpyruvate carboxylase 46 Phosphoenolpyruvate
carboxytransphosphorylase 50 Phosphoenolpyruvate tetrahydrofolate
hydroxymethyltransferase 93, 94 Phosphofructokinase (ATP) 49, 123 Phosphofructokinase (P~) 50 Phosphogluconate pathway 35, 37 Phosphoglycerate kinase 50 Phosphoglycerides 108, 109, llQ-12,
118 Phosphoribosyltransferase 72, 77 Pigment 15, 105 Pinocytosis 91, 92 Piroplasms 3 Plasmalogens 111, 118 Polyenes 22, 121 Polyhistidine 105 Polyphosphate 39 Polyphosphoinositides 111 Polysaccharides 32 Primaquine 21, 23, 126-7, 142, 143 Proline 60, 61, 65, 66, 94, 97 Proline oxidase 61, 64 Prophylaxis 18 Promastigote 9 Prostaglandins 108, 114 Proteases 34, 91 Protein 89-107 Protein kinases 106 Protein, membranes 100, 106
microbodies 103--4 surface coats lOQ-3 synthesis 94--6 taxonomy 106-7
Protoporphyrin IX 160, 161 Pteridines 75, 77, 130, 132, 154 Purines 69, 70, 72 Pyridoxine 155 Pyrimethamine 21, 23, 130, 131, 134,
142, 143, 144 Pyrimidines 69, 71 Pyrophosphate 39, 50, 51, 52, 81

INDEX
Pyrophosphate phosphatase 81 Pyruvate 34, 35, 61, 66, 97 Pyruvate carboxylase 45, 60, 61, 63 Pyruvate kinase 50, 123 Pyruvate oxidase 50, 53 Pyruvate phosphate dikinase 50 Pyruvate synthase 46 Pyruvate: X oxidoreductase 46
Quinine 21, 23, 135, 136, 142, 143 Quinolones 22, 23, 54, 66, 122, 125
Radanil 21,22, 137,142,143 Reduviid bugs 5, 10 Regulation 73, 105-6 Respiratory chain 36, 37, 38, 63, 64,
65, 66, 75, 78, 125, 126, 127 Respiratory chain phosphorylation 36,
37, 38, 63, 66, 125 Resistance to drugs 24, 121-4 Ribonucleotide reductase 72, 77, 79 Riboflavin 155 Ribose 69 Ribosomes 86, 94, 95, 139, 160 Rifampicin 121, 122 RNA 69, 81, 86
messenger 86, 94 ribosomal 82, 86, 87, 88, 95, 96,
160 transfer 82, 86, 95
RNA polymerase 81, 85, 86, 94, 135 Robenidine 22, 122, 124, 142, 143 Rotenone 38, 63
Salicylhydroxamic acid (SHAM) 42, 124
S-adenosyl methionine 93, 131, 134 Sand flies 5, 11 Schizogony 12, 15 Sedimentation coefficient (S value) 84 Serine 80, 93, 94, 97 Serine hydroxymethyltransferase 80 Serotonin 99, 100 Serum 115 Sleeping sickness 4, 20 Sphingolipids 108, 112, 113 Sporozoa 3, 4 Sporozoites 15 Stabilate 28 Stearic acid 108, 116 Steroids 108, 113
171
Sterols 113, 118, 127 Succinate 37, 40, 45, 46, 60, 62, 65 Succinate dehydrogenase 37, 64 Succinyl acetyl CoA transferase 46, 51 SuccinylCoA 46, 51, 97,161 Succinyl CoA synthetase 37 Succinyl thiokinase 46, 51 Sulphadimethoxine 23, 129, 130, 142,
143 Sulphaquinoxaline 22, 129, 130, 142,
143 Sulphonamide 22, 23, 80, 121, 122,
129, 130, 134 Sulphone 23, 129 Superoxide 104-5, 164 Superoxide dismutase 47, 104-5 Suramin 20, 22, 24, 79, 123, 124, 142,
143 Surface coat proteins 100-3 Synergism 134 Syringe passage 27
Taxonomy 3, 82, 106-7 Terpenes 108, 113 Tetracycline 23, 24, 121, 122, 139, 140 Tetraethylthiuram disulphide 117 Tetrahydrofolate 80, 93, 94, 130, 131,
132, 133 Tetrahydrobiopterin 77, 78 Thiamine 129, 155 Thioredoxin 79 Threonine 60, 65, 93, 98, 117 Threonine dehydrogenase 117 Thymidine 77, 80, 81, 85 Thymidine phosphorylase 87 Thymidylate phosphatase 87 Thymidylate synthase 77, 79, 80, 88,
124, 134 Thymine 77, 80, 87, 100 Transhydrogenase 53 Tricarboxylic acid cycle 34, 35, 36, 37,
39, 97 Tryparsamide 20, 22, 122, 142, 143 Trypomastigote 9 Tryptophan 97, 98, 100 Tryptophol 98 Tyrosine 97, 98, 100
Ubiquinone 38, 56, 63, 75, 78, 126, 127 Uracil 77 Urea cycle 97, 98, 100 Uridine 77

172 INDEX
Uridine monophosphate (UMP) 71, 74, Waxes 108, 113 76 WR 30090 23, 24, 136, 142, 143
Uroporphyrinogen synthetase 160, 161 WR 33063 23, 24, 136, 142, 143
Vaginitis 5, 12 Zoalene 23, 139, 140, 142, 143