
Antimicrobial resistance patterns and characterization of integrons in clinical isolates of ...
6
ARTICLE Antimicrobial resistance patterns and characterization of integrons in clinical isolates of Shigella from China Haifei Yang, Yachao Pan, Lifen Hu, Yanyan Liu, Ying Ye, Jun Cheng, and Jiabin Li Abstract: One hundred fifty-three Shigella isolates were examined for multiple antibiotic resistance phenotypes and prevalence of class 1 and class 2 integron sequences. The gene cassettes dfrA17-aadA5, dfrA12-orfF-aadA2, and arr-3-aacA4 were found in typical class 1 integrons. The gene cassettes bla OXA-1 -aadA1 and dfrA1-sat1-aadA1 were detected in atypical class 1 integrons and in class 2 integrons, respectively. This is the first report of arr-3-aacA4 cassette detected in typical class 1 integrons among Shigella isolates. Rates of antibiotic resistance were different between integron-positive and integron-negative strains (P < 0.05), and all integron- positive isolates were resistant to at least 3 different antimicrobial agents. Typical class 1 integron-positive isolates showed higher resistance rates to cefotaxime and ciprofloxacin than did integron-negative ones (P < 0.05). Typical class 1 integrons and -lactamase genes were found in conjugative plasmids, otherwise class 2 and atypical class 1 integrons were located on chro- mosome. This study demonstrated the wide distribution of class 1 integrons in Shigella spp., which may lead resistance to cefotaxime and ciprofloxacin in China. Key words: Shigella spp., antimicrobial susceptibility, integron, -lactamase. Résumé : On a examiné les phénotypes d’antibiorésistance multiple et la prévalence de séquences d’intégrons de classe 1 et 2 chez 153 isolats de Shigella. Les cassettes géniques de dfrA17-aadA5, dfrA12-orfF-aadA2 et arr-3-aacA4 ont été retrouvées dans des intégrons de classe 1 typiques. Les segments bla OXA-1 -aadA1 et dfrA1-sat1-aadA1 ont été détectés dans des intégrons de classe 1 atypiques et de classe 2, respectivement. Il s’agit du premier rapport signalant une cassette arr-3-aacA4 dans un intégron de classe 1 typique chez des isolats de Shigella. Les taux d’antibiorésistance étaient différents chez les souches intégron positives et négatives (P < 0,05), et tous les isolats recelant un intégron résistaient a ` au moins 3 antimicrobiens distincts. Les isolats a ` intégron typique de classe 1 étaient liés a ` des taux de résistance au cefotaxime et a ` la ciprofloxacine supérieurs aux isolats dépourvus de cet type d’intégron (P < 0,05). On a retrouvé les intégrons typiques de classe 1 et les gènes de la -lactamase dans des plasmides de conjugaison; a ` l’opposé, les intégrons de classe 2 et de classe 1 atypiques étaient situés dans le chromosome. Cette étude a fait état d’une distribution élargie d’intégrons de classe 1 chez Shigella spp., ce qui pourrait donner lieu a ` une résistance au cefotaxime et a ` la ciprofloxacine en Chine. [Traduit par la Rédaction] Mots-clés : Shigella spp., susceptibilité aux antimicrobiens, intégron, -lactamase. Introduction Shigellosis is a common diarrhoeal disease of both developed and developing countries. Global studies suggest there are 164.7 million episodes of shigellosis per year, of which 163.2 million are in developing countries and 1.5 million in developed countries. These episodes result in 1.1 million deaths, particularly among children in developing countries (Kotloff et al. 1999). In China, 0.8–1.7 million episodes of shigellosis were reported in 2000 and the predominant species was Shigella flexneri (Wang et al. 2006). Treatment with antimicrobial agents has been effective in allevi- ating the dysenteric syndrome of shigellosis, in reducing the du- ration of pathogen excretion to prevent disease transmission, and in lowering the risk of potential complications, for the past sev- eral decades. But the problem of antimicrobial resistance, espe- cially multidrug resistance (resistance to 3 or more different kinds of antimicrobial agents), in Shigella continues to be alarming. It is of great concern that integrons with resistance gene cassettes have been identified in plasmids, transposons, and chro- mosome (Bennish et al. 1992; Rajakumar et al. 1997). Mobile ge- netic elements may facilitate the dissemination of resistance determinants among species, even genera. Integrons are gene- capture systems that harbour antibiotic resistance genes and may provide a flexible approach for bacteria to adapt to the pressure caused by antibiotics. The resistance to some antibi- otics in Shigella species is associated with the presence of class 1 and class 2 integrons that contain resistance gene cassettes. The class 1 and class 2 integrons remain the most common integrons associated with resistance in clinical isolates (Goldstein et al. 2001; Sabaté and Prats 2002). Integrons and -lactamase-coding genes are responsible for the horizontal transfer of antimicrobial resis- tance among Gram-negative bacilli (Hall and Collis 1995; White et al. 2001). Received 16 December 2013. Revision received 18 February 2014. Accepted 5 March 2014. H. Yang* and Y. Pan.* Department of Infectious Diseases, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China. L. Hu.* Department of Center Laboratory, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; Institute of Bacterial Resistance, Anhui Medical University, Hefei, Anhui, People’s Republic of China; Anhui Center for Surveillance of Bacterial Resistance, Hefei, Anhui, People’s Republic of China. Y. Liu, Y. Ye, J. Cheng, and J. Li. Department of Infectious Diseases, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; Institute of Bacterial Resistance, Anhui Medical University, Hefei, Anhui, People’s Republic of China; Anhui Center for Surveillance of Bacterial Resistance, Hefei, Anhui, People’s Republic of China. Corresponding authors: Jun Cheng (e-mail: [email protected]) and Jiabin Li (e-mail: [email protected]). *These authors contributed equally to this work. 237 Can. J. Microbiol. 60: 237–242 (2014) dx.doi.org/10.1139/cjm-2013-0893 Published at www.nrcresearchpress.com/cjm on 7 March 2014. Can. J. Microbiol. Downloaded from www.nrcresearchpress.com by UNIV OF MANCHESTER on 10/11/14 For personal use only.
Transcript of Antimicrobial resistance patterns and characterization of integrons in clinical isolates of ...

ARTICLE
Antimicrobial resistance patterns and characterization ofintegrons in clinical isolates of Shigella from ChinaHaifei Yang, Yachao Pan, Lifen Hu, Yanyan Liu, Ying Ye, Jun Cheng, and Jiabin Li
Abstract: One hundred fifty-three Shigella isolates were examined for multiple antibiotic resistance phenotypes and prevalenceof class 1 and class 2 integron sequences. The gene cassettes dfrA17-aadA5, dfrA12-orfF-aadA2, and arr-3-aacA4 were found in typicalclass 1 integrons. The gene cassettes blaOXA-1-aadA1 and dfrA1-sat1-aadA1 were detected in atypical class 1 integrons and in class 2integrons, respectively. This is the first report of arr-3-aacA4 cassette detected in typical class 1 integrons among Shigella isolates.Rates of antibiotic resistance were different between integron-positive and integron-negative strains (P < 0.05), and all integron-positive isolates were resistant to at least 3 different antimicrobial agents. Typical class 1 integron-positive isolates showedhigher resistance rates to cefotaxime and ciprofloxacin than did integron-negative ones (P < 0.05). Typical class 1 integrons and�-lactamase genes were found in conjugative plasmids, otherwise class 2 and atypical class 1 integrons were located on chro-mosome. This study demonstrated the wide distribution of class 1 integrons in Shigella spp., which may lead resistance tocefotaxime and ciprofloxacin in China.
Key words: Shigella spp., antimicrobial susceptibility, integron, �-lactamase.
Résumé : On a examiné les phénotypes d’antibiorésistance multiple et la prévalence de séquences d’intégrons de classe 1 et 2chez 153 isolats de Shigella. Les cassettes géniques de dfrA17-aadA5, dfrA12-orfF-aadA2 et arr-3-aacA4 ont été retrouvées dans desintégrons de classe 1 typiques. Les segments blaOXA-1-aadA1 et dfrA1-sat1-aadA1 ont été détectés dans des intégrons de classe 1atypiques et de classe 2, respectivement. Il s’agit du premier rapport signalant une cassette arr-3-aacA4 dans un intégron declasse 1 typique chez des isolats de Shigella. Les taux d’antibiorésistance étaient différents chez les souches intégron positives etnégatives (P < 0,05), et tous les isolats recelant un intégron résistaient a au moins 3 antimicrobiens distincts. Les isolats a intégrontypique de classe 1 étaient liés a des taux de résistance au cefotaxime et a la ciprofloxacine supérieurs aux isolats dépourvus decet type d’intégron (P < 0,05). On a retrouvé les intégrons typiques de classe 1 et les gènes de la �-lactamase dans des plasmidesde conjugaison; a l’opposé, les intégrons de classe 2 et de classe 1 atypiques étaient situés dans le chromosome. Cette étude a faitétat d’une distribution élargie d’intégrons de classe 1 chez Shigella spp., ce qui pourrait donner lieu a une résistance au cefotaximeet a la ciprofloxacine en Chine. [Traduit par la Rédaction]
Mots-clés : Shigella spp., susceptibilité aux antimicrobiens, intégron, �-lactamase.
IntroductionShigellosis is a common diarrhoeal disease of both developed and
developing countries. Global studies suggest there are 164.7 millionepisodes of shigellosis per year, of which 163.2 million are indeveloping countries and 1.5 million in developed countries.These episodes result in 1.1 million deaths, particularly amongchildren in developing countries (Kotloff et al. 1999). In China,0.8–1.7 million episodes of shigellosis were reported in 2000 andthe predominant species was Shigella flexneri (Wang et al. 2006).Treatment with antimicrobial agents has been effective in allevi-ating the dysenteric syndrome of shigellosis, in reducing the du-ration of pathogen excretion to prevent disease transmission, andin lowering the risk of potential complications, for the past sev-eral decades. But the problem of antimicrobial resistance, espe-cially multidrug resistance (resistance to 3 or more different kindsof antimicrobial agents), in Shigella continues to be alarming.
It is of great concern that integrons with resistance genecassettes have been identified in plasmids, transposons, and chro-mosome (Bennish et al. 1992; Rajakumar et al. 1997). Mobile ge-netic elements may facilitate the dissemination of resistancedeterminants among species, even genera. Integrons are gene-capture systems that harbour antibiotic resistance genes andmay provide a flexible approach for bacteria to adapt to thepressure caused by antibiotics. The resistance to some antibi-otics in Shigella species is associated with the presence of class 1and class 2 integrons that contain resistance gene cassettes. Theclass 1 and class 2 integrons remain the most common integronsassociated with resistance in clinical isolates (Goldstein et al. 2001;Sabaté and Prats 2002). Integrons and �-lactamase-coding genesare responsible for the horizontal transfer of antimicrobial resis-tance among Gram-negative bacilli (Hall and Collis 1995; Whiteet al. 2001).
Received 16 December 2013. Revision received 18 February 2014. Accepted 5 March 2014.
H. Yang* and Y. Pan.* Department of Infectious Diseases, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic ofChina.L. Hu.* Department of Center Laboratory, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’s Republic of China; Instituteof Bacterial Resistance, Anhui Medical University, Hefei, Anhui, People’s Republic of China; Anhui Center for Surveillance of Bacterial Resistance, Hefei,Anhui, People’s Republic of China.Y. Liu, Y. Ye, J. Cheng, and J. Li. Department of Infectious Diseases, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, People’sRepublic of China; Institute of Bacterial Resistance, Anhui Medical University, Hefei, Anhui, People’s Republic of China; Anhui Center for Surveillanceof Bacterial Resistance, Hefei, Anhui, People’s Republic of China.Corresponding authors: Jun Cheng (e-mail: [email protected]) and Jiabin Li (e-mail: [email protected]).*These authors contributed equally to this work.
237
Can. J. Microbiol. 60: 237–242 (2014) dx.doi.org/10.1139/cjm-2013-0893 Published at www.nrcresearchpress.com/cjm on 7 March 2014.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.

The objectives of our study were to examine the molecularcharacteristics of class 1 and class 2 integrons, including theirdistribution and locations in the genome, and the correlationbetween gene cassettes and antibiotic resistance in Shigella iso-lates collected from Anhui, China, during a period of 5 years(2005–2009).
Materials and methods
Bacterial isolatesA total of 153 nonduplicate Shigella isolates were collected from
34 secondary level hospitals located in Anhui Province of Chinabetween September 2005 and September 2009. The patients in-cluded in this study were from 18 different cities in Anhui Province:Hefei, Wuhu, Bengbu, Huainan, Ma’anshan, Huaibei, Tongling,Anqing, Huangshan, Chuzhou, Fuyang, Suzhou, Lu’an, Bozhou,Chizhou, Chaohu, and Xuancheng, which distributed across theprovince. The ages of our patients ranged from 1 to 76.
Stool specimens from patients with either diarrhea or dysen-tery were collected before the patients received antibiotics ther-apy and were then screened for Shigella spp. by conventionalbiochemical methods in local hospitals. All isolates were con-firmed with API 20E (bioMérieux, Marcy l’Étoile, France) again inour laboratory. Of these Shigella isolates, 132 (86.3%) were S. flexneri,19 (12.4%) Shigella sonnei, and 2 (0.7%) Shigella boydii.
Escherichia coli ATCC 25922, E. coli ATCC 35218, E. coli V517, andsodium-azide-resistant E. coli J53 were stored at the Anhui Centerfor Surveillance of Bacterial Resistance (Hefei, Anhui, China).
Antimicrobial susceptibility testingThe minimum inhibitory concentrations (MICs) of ampicillin (AMP),
cefotaxime (CTX), cefoxitin (FOX), ceftazidime (CAZ), cefepime (FEP),nalidixic acid (NA), ciprofloxacin (CIP), levofloxacin (LEV), norfloxacin(NOR), gatifloxacin (GAT), gentamicin (GM), amikacin (AMK),chloramphenicol (CHL), trimethoprim–sulfamethoxazole (SXT),tetracycline (TET), and imipenem (IMP) were determined by theagar dilution method, according to the guidelines of the ClinicalLaboratory Standards Institute (2012). Escherichia coli ATCC 25922and E. coli ATCC 35218 were used as a quality control strain. Thesusceptibility data of Shigella isolates tested were accepted onlywhen the MIC for quality control strains tested in parallel waswithin the acceptable ranges given in the Clinical LaboratoryStandards Institute guidelines. Multiple resistance was defined asresistance to 3 or more antimicrobials.
Polymerase chain reaction (PCR) amplificationAll Shigella isolates were screened for the class 1, class 2, and
class 3 integrase genes (intI1, intI2, and intI3, respectively) and theextended-spectrum �-lactamases genes (blaCTX-M, blaOXA, blaTEM,and blaSHV) using the primer pairs described in Table 1.
All integron-positive Shigella isolates were screened for the pres-ence of plasmid-mediated quinolone resistance (PMQR) genes(qnrA, qnrB, qnrS, qnrC, qnrD, aac(6=)-Ib-cr, and qepA) using methodsdescribed previously (Yamane et al. 2007; Xiong et al. 2008; Cavacoet al. 2009; Wang et al. 2009). The mutations in the quinolone-resistance-determining regions of the gyrA, gyrB, parC, and parEgenes, in 16S rRNA methylase genes (armA, rmtB, rmtC, and npmA),and in plasmid-mediated ampC were determined by PCR with themethods described previously for the PMQR-positive isolates(Dutta et al. 2005; Cattoir et al. 2007; Doi and Arakawa 2007;Dallenne et al. 2010; Hu et al. 2011).
All the purified PCR products were sequenced on an ABI Prism3730 sequencer (Applied Biosystems, Foster City, California, USA).Sequence alignment was compared with the GenBank nucleotidedatabase using the nucleotide BLAST program. The sequencesof blaCTX-M-3, blaCTX-M-15, and blaCTX-M-55 have been submitted toGenBank under accession Nos. JN646780, JN627489, and JN627490,respectively.
Conjugation experimentsConjugation experiments were carried out for all isolates
positive for integrons and the �-lactamase gene, with sodium-azide-resistant E. coli J53 as the recipient. Transconjugants wereselected on Luria–Bertani agar plates supplemented with sodiumazide (200 mg/L) (Sigma Chemical Co., St. Louis, Missouri, USA)and AMP (16 mg/L). Transconjugants were tested by biochemicalmethod and confirmed using API 20E (bioMérieux) as E. coli. Plas-mid DNA extraction from donors and transconjugants was per-formed using Qiagen Plasmid Purification kit (QIAGEN, Hilden,Germany). The transconjugants were examined by PCR for thepresence of integrons and the �-lactamase gene using plasmidDNA as the template and were tested for susceptibility as de-scribed above.
Southern blot hybridizationThe plasmid sizes of donors and transconjugants were esti-
mated by agarose gel electrophoresis as previously described(Wang et al. 2003). The presence of integron sequences were con-
Table 1. Primers used for polymerase chain reaction.
Primer Sequence (5=¡3=) Location Product (bp) Reference
intI1-F ACATGTGATGGCGACGCACGA intI1 569 —intI1-R ATTTCTGTCCTGGCTGGCGA intI1intI2-F GTAGCAAACGAGTGACGAAATG intI2 789 —intI2-R CACGGATATGCGACAAAAAGGT intI2intI3-F AGTGGGTGGCGAATGAGTG intI3 600 —intI3-R TGTTCTTGTATCGGCAGGTG intI3In1-R AAGCAGACTTGACCTGA 5= conserved sequence of class 1 integron Variable —In1-F GGCATCCAAGCAGCAAG 3= conserved sequence of class 1 integronIn2-F GACGGCATGCACGATTTGTA 5= conserved sequence of class 2 integron Variable White et al. 2001In2-R GATGCCATCGCAAGTACGAG 3= conserved sequence of class 2 integronintI1ca CGTAGA AGA ACAGCAAGG intI1 2453 —IS1 AGTGAGAGCAGAGATAGC IS1CTX-M-F ATGGTTAAAAAATCACTGCGCC blaCTX-M 833 —CTX-M-R TCCCGACGGCTTTCCGCCTT blaCTX-M
TEM-F TTAGACGTCAGGTGGCACTT blaTEM 1009 —TEM-R GGACCGGAGTTACCAATGCT blaTEM
OXA-F AAGAAACGCTACTCGCCTGC blaOXA 478 —OXA-R CCACTCAACCCATCCTACCC blaOXA
SHV-F TCTTTCCGATGCCGCCGCCAGTCA blaSHV 1115 —SHV-R GCCGGGTTATTCTTATTTGTCGC blaSHV
238 Can. J. Microbiol. Vol. 60, 2014
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.
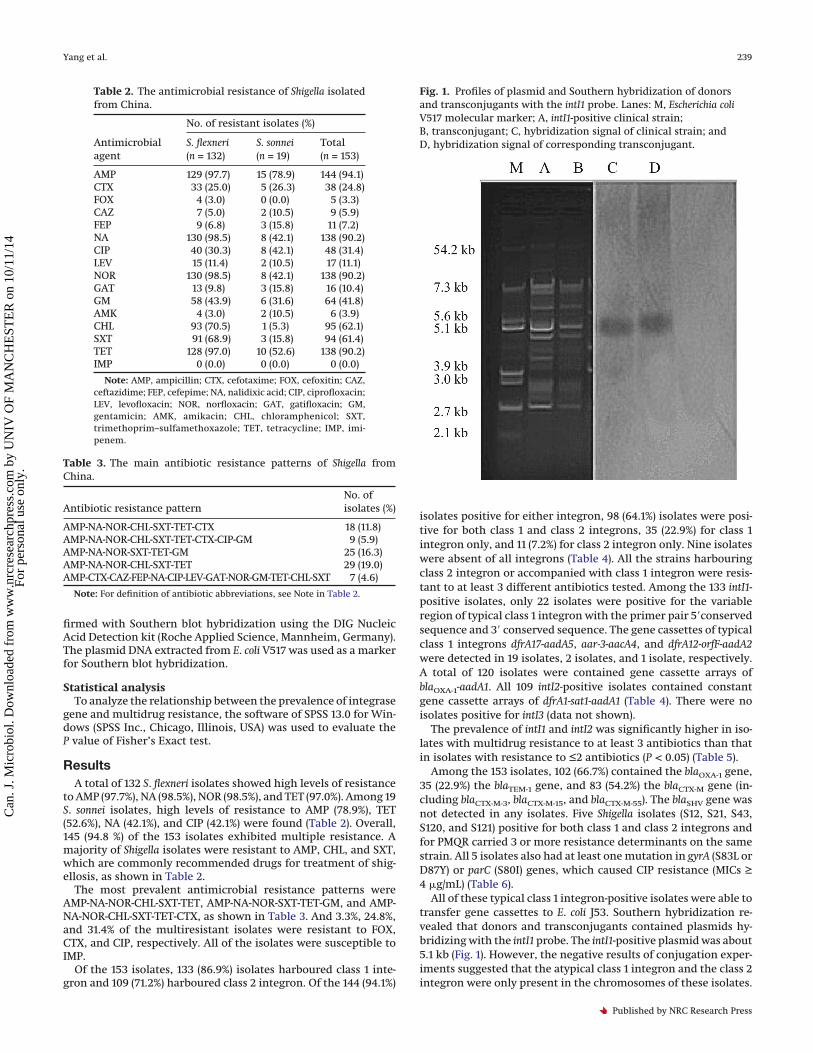
firmed with Southern blot hybridization using the DIG NucleicAcid Detection kit (Roche Applied Science, Mannheim, Germany).The plasmid DNA extracted from E. coli V517 was used as a markerfor Southern blot hybridization.
Statistical analysisTo analyze the relationship between the prevalence of integrase
gene and multidrug resistance, the software of SPSS 13.0 for Win-dows (SPSS Inc., Chicago, Illinois, USA) was used to evaluate theP value of Fisher’s Exact test.
ResultsA total of 132 S. flexneri isolates showed high levels of resistance
to AMP (97.7%), NA (98.5%), NOR (98.5%), and TET (97.0%). Among 19S. sonnei isolates, high levels of resistance to AMP (78.9%), TET(52.6%), NA (42.1%), and CIP (42.1%) were found (Table 2). Overall,145 (94.8 %) of the 153 isolates exhibited multiple resistance. Amajority of Shigella isolates were resistant to AMP, CHL, and SXT,which are commonly recommended drugs for treatment of shig-ellosis, as shown in Table 2.
The most prevalent antimicrobial resistance patterns wereAMP-NA-NOR-CHL-SXT-TET, AMP-NA-NOR-SXT-TET-GM, and AMP-NA-NOR-CHL-SXT-TET-CTX, as shown in Table 3. And 3.3%, 24.8%,and 31.4% of the multiresistant isolates were resistant to FOX,CTX, and CIP, respectively. All of the isolates were susceptible toIMP.
Of the 153 isolates, 133 (86.9%) isolates harboured class 1 inte-gron and 109 (71.2%) harboured class 2 integron. Of the 144 (94.1%)
isolates positive for either integron, 98 (64.1%) isolates were posi-tive for both class 1 and class 2 integrons, 35 (22.9%) for class 1integron only, and 11 (7.2%) for class 2 integron only. Nine isolateswere absent of all integrons (Table 4). All the strains harbouringclass 2 integron or accompanied with class 1 integron were resis-tant to at least 3 different antibiotics tested. Among the 133 intI1-positive isolates, only 22 isolates were positive for the variableregion of typical class 1 integron with the primer pair 5=conservedsequence and 3= conserved sequence. The gene cassettes of typicalclass 1 integrons dfrA17-aadA5, aar-3-aacA4, and dfrA12-orfF-aadA2were detected in 19 isolates, 2 isolates, and 1 isolate, respectively.A total of 120 isolates were contained gene cassette arrays ofblaOXA-1-aadA1. All 109 intI2-positive isolates contained constantgene cassette arrays of dfrA1-sat1-aadA1 (Table 4). There were noisolates positive for intI3 (data not shown).
The prevalence of intI1 and intI2 was significantly higher in iso-lates with multidrug resistance to at least 3 antibiotics than thatin isolates with resistance to ≤2 antibiotics (P < 0.05) (Table 5).
Among the 153 isolates, 102 (66.7%) contained the blaOXA-1 gene,35 (22.9%) the blaTEM-1 gene, and 83 (54.2%) the blaCTX-M gene (in-cluding blaCTX-M-3, blaCTX-M-15, and blaCTX-M-55). The blaSHV gene wasnot detected in any isolates. Five Shigella isolates (S12, S21, S43,S120, and S121) positive for both class 1 and class 2 integrons andfor PMQR carried 3 or more resistance determinants on the samestrain. All 5 isolates also had at least one mutation in gyrA (S83L orD87Y) or parC (S80I) genes, which caused CIP resistance (MICs ≥4 �g/mL) (Table 6).
All of these typical class 1 integron-positive isolates were able totransfer gene cassettes to E. coli J53. Southern hybridization re-vealed that donors and transconjugants contained plasmids hy-bridizing with the intI1 probe. The intI1-positive plasmid was about5.1 kb (Fig. 1). However, the negative results of conjugation exper-iments suggested that the atypical class 1 integron and the class 2integron were only present in the chromosomes of these isolates.
Table 2. The antimicrobial resistance of Shigella isolatedfrom China.
Antimicrobialagent
No. of resistant isolates (%)
S. flexneri(n = 132)
S. sonnei(n = 19)
Total(n = 153)
AMP 129 (97.7) 15 (78.9) 144 (94.1)CTX 33 (25.0) 5 (26.3) 38 (24.8)FOX 4 (3.0) 0 (0.0) 5 (3.3)CAZ 7 (5.0) 2 (10.5) 9 (5.9)FEP 9 (6.8) 3 (15.8) 11 (7.2)NA 130 (98.5) 8 (42.1) 138 (90.2)CIP 40 (30.3) 8 (42.1) 48 (31.4)LEV 15 (11.4) 2 (10.5) 17 (11.1)NOR 130 (98.5) 8 (42.1) 138 (90.2)GAT 13 (9.8) 3 (15.8) 16 (10.4)GM 58 (43.9) 6 (31.6) 64 (41.8)AMK 4 (3.0) 2 (10.5) 6 (3.9)CHL 93 (70.5) 1 (5.3) 95 (62.1)SXT 91 (68.9) 3 (15.8) 94 (61.4)TET 128 (97.0) 10 (52.6) 138 (90.2)IMP 0 (0.0) 0 (0.0) 0 (0.0)
Note: AMP, ampicillin; CTX, cefotaxime; FOX, cefoxitin; CAZ,ceftazidime; FEP, cefepime; NA, nalidixic acid; CIP, ciprofloxacin;LEV, levofloxacin; NOR, norfloxacin; GAT, gatifloxacin; GM,gentamicin; AMK, amikacin; CHL, chloramphenicol; SXT,trimethoprim–sulfamethoxazole; TET, tetracycline; IMP, imi-penem.
Table 3. The main antibiotic resistance patterns of Shigella fromChina.
Antibiotic resistance patternNo. ofisolates (%)
AMP-NA-NOR-CHL-SXT-TET-CTX 18 (11.8)AMP-NA-NOR-CHL-SXT-TET-CTX-CIP-GM 9 (5.9)AMP-NA-NOR-SXT-TET-GM 25 (16.3)AMP-NA-NOR-CHL-SXT-TET 29 (19.0)AMP-CTX-CAZ-FEP-NA-CIP-LEV-GAT-NOR-GM-TET-CHL-SXT 7 (4.6)
Note: For definition of antibiotic abbreviations, see Note in Table 2.
Fig. 1. Profiles of plasmid and Southern hybridization of donorsand transconjugants with the intI1 probe. Lanes: M, Escherichia coliV517 molecular marker; A, intI1-positive clinical strain;B, transconjugant; C, hybridization signal of clinical strain; andD, hybridization signal of corresponding transconjugant.
Yang et al. 239
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.
DiscussionAmong Shigella isolates, S. flexneri is the major agent that causes
bacterial diarrhea in most Asian countries (Kotloff et al. 1999; Choiet al. 2007). Our study also showed that S. flexneri remains thepredominant serotype among Shigella strains in Anhui Province.Antimicrobial resistance in Shigella is common nowadays; it isparticularly resistant to TET and sulfonamides–trimethoprim(Peirano et al. 2005). However, the resistance rates to AMP, CHL,and NA vary greatly (Jafari et al. 2009; Rosewell et al. 2010). In ourstudy, Shigella isolates had a high prevalence of resistance to all ofthese commonly used antimicrobial agents. The most frequentantimicrobial resistance pattern was AMP-NA-NOR-CHL-SXT-TET.Our findings illustrated that more than two thirds of Shigella iso-lates were resistant to at least 6 antimicrobials, which confirmedthat these antimicrobial agents are now generally inefficient inthe empirical treatment of shigellosis. Shigella had good sensitivityto CIP and CTX in vitro, but the resistance rates were still high inour study in contrast with studies for some other countries (Ohet al. 2003; Rosewell et al. 2010). Although AMP and SXT have beenrecommended for treating shigellosis in the past, they are cur-rently considered ineffective for empirical therapy (Niyogi 2005).In our study, we found a high prevalence of Shigella isolates withresistance to many kinds of antimicrobials. In China, patientswith diarrhea have commonly been treated with antimicrobialsby doctors without the result of bacterial culture. In many cases,patients with diarrhea took antimicrobials by themselves beforethey saw a doctor, regardless of whether the diarrhea was causedby bacteria or virus. Therefore, antibiotic abuse and misuse is veryserious.
According to the current study, the observed susceptibility ofShigella isolates to FOX, AMK, third-generation cephalosporins,and IMP indicated that these antibiotics may be the preferredempirical treatment options in Anhui, China. From our findingsand that of others (Oh et al. 2003; Peirano et al. 2005; Jafari et al.2009; Rosewell et al. 2010), we concluded that antimicrobial resis-tance in Shigella isolates may change over time and along withregions, so continuous surveillance of resistance is necessary, es-pecially in developing countries.
The prevalence of integrons found in Shigella varies from coun-try to country. In our study, most Shigella isolates harbouredclasses 1 or 2 integrons, and class 1 integron was the most fre-quently detected, which is in keeping with the results of otherstudies (Dubois et al. 2007; Gassama Sow et al. 2010). However,some reports also revealed that class 2 integron was predominantin some countries (Peirano et al. 2005; Ahmed et al. 2006). Theprevalence of class 1 and class 2 integrons in Shigella isolates inAnhui Province was higher than that in previous studies (Duboiset al. 2007). The resistance to antimicrobials in Shigella isolateswith class 1 integron and (or) class 2 integron was far more com-mon than that in Shigella isolates without them (P < 0.05). Thisdemonstrates that horizontal gene transfer mediated by inte-grons contributes to the dissemination of antimicrobial drug re-sistance in Shigella isolates in Anhui, China.
Our results proved that the majority of typical class 1 integronscontained the gene cassette dfrA17-aadA5. In addition, dfr12-orfF-aadA2was detected in 1 Shigella isolate while arr-3-aacA4 was detected in2 Shigella isolates. These cassettes have also been reported in dif-ferent bacterial species worldwide (Chang et al. 2009; Krauland
Table 4. Profiles of integrons and gene cassettes in 153 Shigella isolates.
Gene cassette
Integrase gene Typical class 1 integron Atypical class 1 integron Class 2 integron
Shigellaserotype
No. ofisolates intI1 intI2 Both* None dfrA17-aadA5 aar-3-aacA4 dfrA12-orfF-aadA2 blaOXA-1-aadA1 dfrA1-sat1-aadA1
S. flexneri 132 119 91 85 7 17 2 1 107 91S. sonnei 19 13 16 12 2 2 0 0 12 16S. boydii 2 1 2 1 0 0 0 0 1 2
*“Both” means isolates contain intI1 and intI2 simultaneously.
Table 5. The profiles of integron-positive and multiresistant Shigella from China.
Group No.
No. of isolates (%)
P*Multiresistantto ≥3 antibiotics
Susceptive to all orresistant to ≤2 antibiotics
Positive for either integron 144 140 (97.2) 4 (2.8) 0.000Negative for all integrons 9 5 (55.6) 4 (44.4)Positive for intI1 gene 133 131 (98.5) 2 (1.5) 0.000Negative for intI1 gene 20 14 (70.0) 6 (30.0)Positive for intI2 gene 109 106 (97.2) 3 (2.8) 0.044Negative for intI2 gene 44 39 (88.6) 5 (11.4)
*Fisher's Exact test.
Table 6. Profile of 5 Shigella isolates positive for both class 1 and class 2 integrons.
�-LactamaseMutationin QRDRs
Isolate blaCTX-M15 blaOXA-1 blaTEM-1 blaSHV
PMQRdeterminants gyrA parC Methylase Antibiotic resistance pattern
S12 + + – – aac(6=)-Ib-cr S83L AMP-CTX-NA-LEV-GAT-GM-TET-CHL-SXTS21 – + – – qepA D87Y S80I rmtB AMP-NA-CIP-LEV-NOR-GM, AMK-TET-CHL-SXTS43 + – – – aac(6=)-Ib-cr S83L AMP-CTX-NA-TET-SXTS120 + + – – qnrS2 S83L S80I AMP-CTX-FEP-NA-CIP-LEV-GAT-NOR-GM-TET-CHL-SXTS121 + + + – qnrS2 S83L S80I rmtB AMP-CTX-CAZ-FEP-NA-CIP-LEV-GAT-NOR-GM-AMK-TET-CHL-SXT
Note: PMQR, plasmid-mediated quinolone resistance; QRDRs, quinolone-resistance-determining regions. For definition of antibiotic abbreviations, see Note inTable 2.
240 Can. J. Microbiol. Vol. 60, 2014
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.

et al. 2009; Zhang et al. 2009). The typical class 1 integron-positiveisolates showed higher resistance to CTX and CIP in contrast withclass 1 integron-negative ones (P < 0.05). The typical class 1integron-positive Shigella isolates had a higher incidence of blaTEM
compared with atypical class 1 integron- or class 2 integron-positive isolates (P < 0.05). But the reason for this phenomenon isnot clear, and thus needs further investigation. Conjugation ex-periments and Southern hybridization indicated the horizontaltransfer of typical class 1 integron through conjugative plasmids.
The present study also identified an atypical class 1 integronwith an unusual 3= conserved sequence linked to an insertionsequence called IS1, which was first discovered on the chromo-some of S. flexneri 2a strain YSH6000 isolated in Japan (Rajakumaret al. 1997). According to the previous report (Rajakumar et al.1997), the atypical class 1 integron together with CHL and TETresistance determinants were located in the Shigella resistancelocus. Our result showed that atypical class 1 integron-positiveShigella isolates had a higher resistance rates to AMP, CHL, and TET(P < 0.05), which also confirmed that the atypical class 1 integronwas linked to CHL and TET resistance determinants.
Previous studies have proved that class 2 integron was oftenassociated with S. sonnei isolates (Chang et al. 2011). In our study,class 2 integrons were detected in a large quantity of S. flexneri andS. sonnei isolates indiscriminately. The only gene cassette arrayfound in the class 2 integron was dfrA1-sat1-aadA1. Both the class 2integron and atypical class 1 integron can not be transferred byconjugation. Moreover, the failure of hybridization confirmedtheir chromosome location, which indicated the transmission ofclones carrying class 2 integron and atypical class 1 integronamong isolates.
The bla genes for TEM, CTX-M, and OXA were also detected in153 isolates, and all the blaOXA-positive isolates were found tocontain gene cassette arrays of blaOXA-1-aadA1. The presence ofthese resistance genes in combination with integrons may haveimportant clinical implications, which will make the multidrugresistance more serious, as the resistance genes can widely andrapidly distribute within and between species. In our study,5 Shigella isolates positive for both class 1 and class 2 integrons andfor PMQR carried 3 or more resistance determinants on the samestrain All 5 isolates had at least 1 mutation in gyrA (S83L or D87Y)or parC (S80I) genes, which caused CIP resistance (MICs ≥ 4 �g/mL).So these mechanisms should be explored further to monitor andcontrol antimicrobial resistance.
In conclusion, this study demonstrated that multiple antibioticresistance is common in clinical isolates of Shigella in Anhui,China, especially against widely used therapeutic agents for shig-ellosis. Multiple antibiotic resistance in the isolates was conferredby class 1 and class 2 integrons and other antimicrobial resistancegenes. Therefore, further strengthening of surveillance of anti-microbial resistance in China is urgently needed.
AcknowledgementsThis study was supported by the Natural Science Foundation
of China (Nos. 30972631 and 81172737) and Provincial Natural Sci-ence Foundation Key Program of Higher Education of China(Nos. KJ2011A180, KJ2012A177, and KJ2010A344). We are grateful toHuimin Ma, Xue Zhou, Liguang Liu and Xiuying Miao for assis-tance in antimicrobial susceptibility testing. Authors declare noconflict of interest.
ReferencesAhmed, A.M., Furuta, K., Shimomura, K., Kasama, Y., and Shimamoto, T. 2006.
Genetic characterization of multidrug resistance in Shigella spp. from Japan.J. Med. Microbiol. 55: 1685–1691. doi:10.1099/jmm.0.46725-0. PMID:17108272.
Bennish, M.L., Salam, M.A., Hossain, M.A., Myaux, J., Khan, E.H., Chakraborty, J.,et al. 1992. Antimicrobial resistance of Shigella isolates in Bangladesh, 1983-1990: increasing frequency of strains multiply resistant to ampicillin,trimethoprim–sulfamethoxazole, and nalidixic acid. J. Infect. Dis. 14: 1055–1060. doi:10.1093/clinids/14.5.1055. PMID:1600006.
Cattoir, V., Poirel, L., Rotimi, V., Soussy, C.J., and Nordmann, P. 2007. MultiplexPCR for detection of plasmid-mediated quinolone resistance qnr genes inESBL-producing enterobacterial isolates. J. Antimicrob. Chemother. 60: 394–397. doi:10.1093/jac/dkm204. PMID:17561500.
Cavaco, L.M., Hasman, H., Xia, S., and Aarestrup, F.M. 2009. qnrD, a novel geneconferring transferable quinolone resistance in Salmonella enterica serovarKentucky and Bovismorbificans strains of human origin. Antimicrob. AgentsChemother. 53: 603–608. doi:10.1128/AAC.00997-08. PMID:19029321.
Chang, C.Y., Fang, Y.T., Tsai, S.M., Chang, L.L., and Yu, W.L. 2009. Characteriza-tion of class 1 integrons and gene cassettes in clinical isolates of Klebsiellapneumoniae from Taiwan. Diagn. Microbiol. Infect. Dis. 65: 214–216. doi:10.1016/j.diagmicrobio.2009.06.005. PMID:19748438.
Chang, C.Y., Lu, P.L., Lin, C.C., Lee, T.M., Tsai, M.Y., and Chang, L.L. 2011. Integrontypes, gene cassettes, antimicrobial resistance genes and plasmids of Shigellasonnei isolates from outbreaks and sporadic cases in Taiwan. J. Med. Micro-biol. 60: 197–204. doi:10.1099/jmm.0.022517-0. PMID:20947666.
Choi, S.Y., Jeon, Y.S., Lee, J.H., Choi, B., Moon, S.H., von Seidlein, L., et al. 2007.Multilocus sequence typing analysis of Shigella flexneri isolates collected inAsian countries. J. Med. Microbiol. 56: 1460–1466. doi:10.1099/jmm.0.47322-0.PMID:17965345.
Clinical and Laboratory Standards Institute. 2012. Performance Standards forAntimicrobial Susceptibility Testing; Nineteenth Information Supplement.M100-S22. CLSI, Wayne, Penn., USA.
Dallenne, C., Da Costa, A., Decré, D., Favier, C., and Arlet, G. 2010. Developmentof a set of multiplex PCR assays for the detection of genes encoding impor-tant �-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 65: 490–495. doi:10.1093/jac/dkp498. PMID:20071363.
Doi, Y., and Arakawa, Y. 2007. 16S ribosomal RNA methylation: emerging resis-tance mechanism against aminoglycosides. Clin. Infect. Dis. 45: 88–94. doi:10.1086/518605. PMID:17554708.
Dubois, V., Parizano, M.P., Arpin, C., Coulange, L., Bezian, M.C., and Quentin, C.2007. High genetic stability of integrons in clinical isolates of Shigella spp. ofworldwide origin. Antimicrob. Agents Chemother. 51: 1333–1340. doi:10.1128/AAC.01109-06. PMID:17242143.
Dutta, S., Kawamura, Y., Ezaki, T., Nair, G.B., Lida, K., and Yoshida, S. 2005.Alteration in the GyrA subunit of DNA gyrase and the ParC subunit of topo-isomerase IV in quinolone-resistant Shigella dysenteriae serotype I clinical iso-lates from Kolkata, India. Antimicrob. Agents Chemother. 49: 1660–1661.doi:10.1128/AAC.49.4.1660-1661.2005. PMID:15793166.
Gassama Sow, A., Aïdara-Kane, A., Barraud, O., Gatet, M., Denis, F., and Ploy, M.C.2010. High prevalence of trimethoprim-resistance cassettes in class 1 and 2integrons in Senegalese Shigella isolates. J. Infect. Dev. Counttries, 4: 207–212.doi:10.3855/jidc.583. PMID:20440057.
Goldstein, C., Lee, M.D., Sanchez, S., Hudson, C., Phillips, B., Register, B., et al.2001. Incidence of class 1 and 2 integrases in clinical and commensal bacteriafrom livestock, companion animals and exotics. Antimicrob. Agents Che-mother. 45: 723–726. doi:10.1128/AAC.45.3.723-726.2001. PMID:11181350.
Hall, R.M., and Collis, C.M. 1995. Mobile gene cassettes and integrons: captureand spread of genes by site-specific recombination. Mol. Microbiol. 15: 593–600. doi:10.1111/j.1365-2958.1995.tb02368.x. PMID:7783631.
Hu, L.F., Chang, X., Ye, Y., Wang, Z.X., Shao, Y.B., Shi, W., et al. 2011. Stenotroph-omonas maltophilia resistance to trimethoprim/sulfamethoxazole mediated byacquisition of sul and dfrA genes in a plasmid-mediated class 1 integron. Int. J.Antimicrob. Agents, 37: 230–234. doi:10.1016/j.ijantimicag.2010.10.025. PMID:21296557.
Jafari, F., Hamidian, M., Rezadehbashi, M., Doyle, M., Salmanzadeh-Ahrabi, S.,Derakhshan, F., and Reza Zali, M. 2009. Prevalence and antimicrobial resis-tance of diarrheagenic Escherichia coli and Shigella species associated withacute diarrhea in Tehran, Iran. Can. J. Infect. Dis. Med. Microbiol. 20: e56–e62. PMID:20808457.
Kotloff, K.L., Winickoff, J.P., Ivanoff, B., Clemens, J.D., Swerdlow, D.L.,Sansonetti, P.J., et al. 1999. Global burden of Shigella infections: implicationsfor vaccine development and implementation of control strategies. Bull. W.H. O. 77: 651–666. PMID:10516787.
Krauland, M.G., Marsh, J.W., Paterson, D.L., and Harrison, L.H. 2009. Integron-mediated multidrug resistance in a global collection of nontyphoidalSalmonella enterica isolates.Emerg. Infect.Dis.15: 388–396.doi:10.3201/eid1503.081131. PMID:19239750.
Niyogi, S.K. 2005. Shigellosis. J. Microbiol. 43: 133–143. PMID:15880088.Oh, J.Y., Yu, H.S., Kim, S.K., Seol, S.Y., Cho, D.T., and Lee, J.C. 2003. Changes in
patterns of antimicrobial susceptibility and integron carriage among Shigellasonnei isolates from southwestern Korea during epidemic periods. J. Clin.Microbiol. 41: 421–423. doi:10.1128/JCM.41.1.421-423.2003. PMID:12517883.
Peirano, G., Agersø, Y., Aarestrup, F.M., and dos Prazeres Rodrigues, D. 2005.Occurrence of integrons and resistance genes among sulphonamide-resistant Shigella spp. from Brazil. J. Antimicrob. Chemother. 55: 301–305. doi:10.1093/jac/dki012. PMID:15681578.
Rajakumar, K., Bulach, D., Davies, J., Ambrose, L., Sasakawa, C., and Adler, B.1997. Identification of a chromosomal Shigella flexneri multi-antibiotic resis-tance locus which shares sequence and organizational similarity with theresistance region of the plasmid NR1. Plasmid, 37: 159–168. doi:10.1006/plas.1997.1280. PMID:9200219.
Rosewell, A., Ropa, B., Posanai, E., Dutta, S.R., Mola, G., Zwi, A., and
Yang et al. 241
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.

MacIntyre, C.R. 2010. Shigella spp. antimicrobial drug resistance, Papua NewGuinea, 2000-2009. Emerg. Infect. Dis. 16: 1797–1799. doi:10.3201/eid1611.101025. PMID:21029550.
Sabaté, M., and Prats, G. 2002. Structure and function of integrons. Enferm.Infecc. Microbiol. Clin. 20: 341–345. doi:10.1016/S0213-005X(02)72813-9. PMID:12237001.
Wang, M., Tran, J.H., Jacoby, G.A., Zhang, Y.Y., Wang, F., and Hooper, D.C. 2003.Plasmid-mediated quinolone resistance in clinical isolates of Escherichia colifrom Shanghai, China. Antimicrob. Agents Chemother. 47: 2242–2248. doi:10.1128/AAC.47.7.2242-2248.2003. PMID:12821475.
Wang, M., Guo, Q., Xu, X., Wang, X., Ye, X., Wu, S., et al. 2009. New plasmid-mediated quinolone resistance gene, qnrC, found in a clinical isolate ofProteus mirabilis. Antimicrob. Agents Chemother. 53:1892–1897. doi:10.1128/AAC.01400-08. PMID:19258263.
Wang, X.Y., Tao, F, Xiao, D., Lee, H., Deen, J., Gong, J., et al. 2006. Trend and
disease burden of bacillary dysentery in China (1991-2000). Bull. W.H.O. 84:561–568. doi:10.2471/BLT.05.023853. PMID:16878230.
White, P.A., McIver, C.J., and Rawlinson, W.D. 2001. Integrons and gene cassettesin the Enterobacteriaceae. Antimicrob. Agents Chemother. 45: 2658–2661. doi:10.1128/AAC.45.9.2658-2661.2001. PMID:11502548.
Xiong, Z., Wang, P., Wei, Y., Wang, H., Cao, H., Huang, H., and Li, J. 2008.Investigation of qnr and aac(6=)-Ib-cr in Enterobacter cloacae isolates from AnhuiProvince, China. Diagn. Microbiol. Infect. Dis. 62: 457–459. doi:10.1016/j.diagmicrobio.2008.07.010. PMID:18814992.
Yamane, K., Wachino, J., Suzuki, S., Kimura, K., Shibata, N., Kato, H., et al. 2007.New plasmid-mediated fluoroquinolone efflux pump, QepA, found in anEscherichia coli clinical isolate. Antimicrob. Agents Chemother. 51: 3354–3360.doi:10.1128/AAC.00339-07. PMID:17548499.
Zhang, X.Y., Ding, L.J., and Yue, J. 2009. Occurrence and characteristics of class 1and class 2 integrons in resistant Escherichia coli isolates from animals andfarm workers in northeastern China. Microb. Drug Resist. 15: 323–328. doi:10.1089/mdr.2009.0020. PMID:19857140.
242 Can. J. Microbiol. Vol. 60, 2014
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IV O
F M
AN
CH
EST
ER
on
10/1
1/14
For
pers
onal
use
onl
y.


















