
Anatomical analysis of afferent projections to the medial ...vertes/pfc-afferents.pdf · Anatomical...
31
ORIGINAL ARTICLE Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat Walter B. Hoover Robert P. Vertes Received: 27 February 2007 / Accepted: 4 June 2007 / Published online: 27 July 2007 ȑ Springer-Verlag 2007 Abstract The medial prefrontal cortex (mPFC) has been associated with diverse functions including attentional processes, visceromotor activity, decision making, goal directed behavior, and working memory. Using retrograde tracing techniques, we examined, compared, and contrasted afferent projections to the four divisions of the mPFC in the rat: the medial (frontal) agranular (AGm), anterior cingu- late (AC), prelimbic (PL), and infralimbic (IL) cortices. Each division of the mPFC receives a unique set of afferent projections. There is a shift dorsoventrally along the mPFC from predominantly sensorimotor input to the dorsal mPFC (AGm and dorsal AC) to primarily ‘limbic’ input to the ventral mPFC (PL and IL). The AGm and dorsal AC re- ceive afferent projections from widespread areas of the cortex (and associated thalamic nuclei) representing all sensory modalities. This information is presumably inte- grated at, and utilized by, the dorsal mPFC in goal directed actions. In contrast with the dorsal mPFC, the ventral mPFC receives significantly less cortical input overall and afferents from limbic as opposed to sensorimotor regions of cortex. The main sources of afferent projections to PL/IL are from the orbitomedial prefrontal, agranular insular, perirhinal and entorhinal cortices, the hippocampus, the claustrum, the medial basal forebrain, the basal nuclei of amygdala, the midline thalamus and monoaminergic nuclei of the brainstem. With a few exceptions, there are few projections from the hypothalamus to the dorsal or ventral mPFC. Accordingly, subcortical limbic information mainly reaches the mPFC via the midline thalamus and basal nu- clei of amygdala. As discussed herein, based on patterns of afferent (as well as efferent) projections, PL is positioned to serve a direct role in cognitive functions homologous to dorsolateral PFC of primates, whereas IL appears to rep- resent a visceromotor center homologous to the orbitome- dial PFC of primates. Keywords Claustrum Á Nucleus reuniens Á Memory Á Mediodorsal nucleus of thalamus Á Insular cortex Abbreviations AA Anterior area of amygdala AC Anterior cingulate cortex ACC Nucleus accumbens AD Anterodorsal nucleus of thalamus AGm Medial agranular (frontal) cortex AGl Lateral agranular (frontal) cortex AH Anterior nucleus of hypothalamus AI,d,p,v Agranular insular cortex, dorsal, posterior, ventral divisions AM,d Anteromedial nucleus of thalamus, dorsal division AON, m,v Anterior olfactory nucleus, medial, ventral parts AQ Cerebral aqueduct APN Anterior pretectal nucleus AUD Auditory cortex AV Anteroventral nucleus of thalamus BF Basal forebrain BLA Basolateral nucleus of amygdala BMA,p Basomedial nucleus of amygdala, posterior part BST Bed nucleus of stria terminalis W. B. Hoover Á R. P. Vertes (&) Center for Complex Systems and Brain Sciences, Florida Atlantic University, Boca Raton, FL 33431, USA e-mail: [email protected] 123 Brain Struct Funct (2007) 212:149–179 DOI 10.1007/s00429-007-0150-4
-
Upload
truongtruc -
Category
Documents
-
view
217 -
download
0
Transcript of Anatomical analysis of afferent projections to the medial ...vertes/pfc-afferents.pdf · Anatomical...

ORIGINAL ARTICLE
Anatomical analysis of afferent projections to the medialprefrontal cortex in the rat
Walter B. Hoover Æ Robert P. Vertes
Received: 27 February 2007 / Accepted: 4 June 2007 / Published online: 27 July 2007
� Springer-Verlag 2007
Abstract The medial prefrontal cortex (mPFC) has been
associated with diverse functions including attentional
processes, visceromotor activity, decision making, goal
directed behavior, and working memory. Using retrograde
tracing techniques, we examined, compared, and contrasted
afferent projections to the four divisions of the mPFC in the
rat: the medial (frontal) agranular (AGm), anterior cingu-
late (AC), prelimbic (PL), and infralimbic (IL) cortices.
Each division of the mPFC receives a unique set of afferent
projections. There is a shift dorsoventrally along the mPFC
from predominantly sensorimotor input to the dorsal mPFC
(AGm and dorsal AC) to primarily ‘limbic’ input to the
ventral mPFC (PL and IL). The AGm and dorsal AC re-
ceive afferent projections from widespread areas of the
cortex (and associated thalamic nuclei) representing all
sensory modalities. This information is presumably inte-
grated at, and utilized by, the dorsal mPFC in goal directed
actions. In contrast with the dorsal mPFC, the ventral
mPFC receives significantly less cortical input overall and
afferents from limbic as opposed to sensorimotor regions of
cortex. The main sources of afferent projections to PL/IL
are from the orbitomedial prefrontal, agranular insular,
perirhinal and entorhinal cortices, the hippocampus, the
claustrum, the medial basal forebrain, the basal nuclei of
amygdala, the midline thalamus and monoaminergic nuclei
of the brainstem. With a few exceptions, there are few
projections from the hypothalamus to the dorsal or ventral
mPFC. Accordingly, subcortical limbic information mainly
reaches the mPFC via the midline thalamus and basal nu-
clei of amygdala. As discussed herein, based on patterns of
afferent (as well as efferent) projections, PL is positioned
to serve a direct role in cognitive functions homologous to
dorsolateral PFC of primates, whereas IL appears to rep-
resent a visceromotor center homologous to the orbitome-
dial PFC of primates.
Keywords Claustrum � Nucleus reuniens � Memory �Mediodorsal nucleus of thalamus � Insular cortex
Abbreviations
AA Anterior area of amygdala
AC Anterior cingulate cortex
ACC Nucleus accumbens
AD Anterodorsal nucleus of thalamus
AGm Medial agranular (frontal) cortex
AGl Lateral agranular (frontal) cortex
AH Anterior nucleus of hypothalamus
AI,d,p,v Agranular insular cortex, dorsal, posterior,
ventral divisions
AM,d Anteromedial nucleus of thalamus, dorsal
division
AON, m,v Anterior olfactory nucleus, medial, ventral
parts
AQ Cerebral aqueduct
APN Anterior pretectal nucleus
AUD Auditory cortex
AV Anteroventral nucleus of thalamus
BF Basal forebrain
BLA Basolateral nucleus of amygdala
BMA,p Basomedial nucleus of amygdala, posterior
part
BST Bed nucleus of stria terminalis
W. B. Hoover � R. P. Vertes (&)
Center for Complex Systems and Brain Sciences,
Florida Atlantic University,
Boca Raton, FL 33431, USA
e-mail: [email protected]
123
Brain Struct Funct (2007) 212:149–179
DOI 10.1007/s00429-007-0150-4

CA1,3 Field CA1 and CA3 of Ammon’s horn
CB Cinguum bundle
CC Corpus callosum
CEA Central nucleus of amygdala
CL Central lateral nucleus of the thalamus
CLA Claustrum
CLi Central linear nucleus
CM Central medial nucleus of thalamus
COA Cortical nucleus of amygdala
CP Caudate-putamen
CUN Cuneiform nucleus
DBh Nucleus of diagonal band, horizontal limb
DG Dentate gyrus of hippocampus
DI Dysgranular insular cortex
DR Dorsal raphe nucleus
EC,l,m Entorhinal cortex, lateral, medial divisions
ECT Ectorhinal cortex
EN Endopiriform nucleus
FP,l,m Frontal polar cortex, lateral, medial divisions
FR Fasciculus retroflexus
FS Fundus of the striatum
GI Granular insular cortex
GP Globus pallidus
HF Hippocampal formation
IAM Interanteromedial nucleus of thalamus
IC Inferior colliculus
IF Interfascicular nucleus
IL Infralimbic cortex
IMD Intermediodorsal necleus of thalamus
INC Insular cortex
IP Interpeduncular nucleus
LA Lateral nucleus of amygdala
LC Locus coeruleus
LD Lateral dorsal nucleus of thalamus
LDT Laterodorsal tegmental nucleus
LG,d Lateral geniculate nucleus, dorsal division
LH Lateral habenula
LHy Lateral hypothalamus
LM Lateral mammillary nucleus
LO Lateral orbital cortex
LP Lateral posterior nucleus of thalamus
LPO Lateral preoptic area
LS Lateral septum
LV Lateral ventricle
MA Magnocellular preoptic nucleus
MB Mammillary bodies
MD Mediodorsal nucleus of thalamus
MEA Medial nucleus of the amygdala
MFB Medial forebrain bundle
MG,v Medial geniculate nucleus, ventral division
MH Medial habenula
MO Medial orbital cortex
mPFC Medial prefrontal cortex
MPO Medial preoptic area
MR Median raphe nucleus
MRF Mesencephalic reticular formation
MS Medial septum
MT Mammillothalamic tract
NI Nucleus incertus
NLL Nucleus of lateral lemniscus
NP Nucleus of pons
OC,1,2 Occipital cortex, primary and secondary
divisions
OT Olfactory tubercle
PA Posterior nucleus of amygdala
PAG Periaqueductal gray
PAp Posterior parietal cortex
PARA Parasubiculum of HF
PB, l, m Parabrachial nucleus, lateral, medial divisions
PC Paracentral nucleus of thalamus
PF Parafascicular nucleus
PH Posterior nucleus of hypothalamus
PIR Piriform cortex
PL Prelimbic cortex
PN5 Principal sensory nucleus of trigeminal nerve
PO Posterior nucleus of thalamus
POST Postsubiculum of HF
PPT Pedunculopontine tegmental nucleus
PRC Perirhinal cortex
PRE Presubiculum of HF
PT Paratenial nucleus of thalamus
PV Paraventricular nucleus of thalamus
RAM Radial arm maze
RE Nucleus reuniens of thalamus
RF Rhinal fissue
RH Rhomboid nucleus of thalamus
RLi Rostral linear nucleus
RM Raphe magnus
RN Red nucleus
RPC Nucleus pontis caudalis
RPO Nucleus pontis oralis
RSC Retrosplenial cortex
RR Retrorubral area
RT Reticular nucleus of thalamus
SC Superior colliculus
SF Septofimbrial nucleus
SI Substantia innominata
sm Stria medullaris
SM Submedial nucleus of thalamus
SN,c,r Substantia nigra, pars compacta, pars
reticulata
SSI Primary somatosensory cortex
SSII Secondary somatosensory cortex
SUB,d,v Subiculum, dorsal, ventral parts
150 Brain Struct Funct (2007) 212:149–179
123

SUM Supramammillary nucleus
TE Temporal cortex
TR Amygdalo-piriform transition zone
TT,d,v Taenia tecta, dorsal, ventral parts
V3 Third ventricle
V4 Forth ventricle
VAL Ventral anterior-lateral complex of thalamus
VB Ventrobasal complex of thalamus
VLO Ventrolateral orbital cortex
VM Ventral medial nucleus of thalamus
VO Ventral orbital cortex
VTA Ventral tegmental area
ZI Zona incerta
Introduction
The prefrontal cortex (PFC) of the rat has been divided into
medial, orbital and lateral parts (Ongur and Price 2000).
The medial PFC (mPFC) consists of the four main divi-
sions which from dorsal to ventral are the medial agranular
(AGm) (or medial precentral), the anterior cingulate (AC),
the prelimbic (PL), the infralimbic (IL) cortices (Berendse
and Groenewegen 1991; Ray and Price 1992; Ongur and
Price 2000; Heidbreder and Groenewegen 2003).
The mPFC has been associated with diverse functions
including oculomotor control (frontal eye fields), atten-
tional processes, visceromotor activity, decision making,
goal directed behavior, and working memory (Fuster 1989;
Kolb 1990; Neafsey 1990; Goldman-Rakic 1994; Petrides
1998; Repovs and Baddeley 2006). Dorsal regions of
mPFC (AGm and AC) have been implicated in various
motor behaviors, while ventral regions of mPFC (IL and
PL) have been associated with diverse emotional, cogni-
tive, and mnemonic processes (Heidbreder and Groe-
newegen 2003).
The IL has been shown to profoundly influence visceral/
autonomic activity. IL stimulation produces changes in
respiration, gastrointestinal motility, heart rate, and blood
pressure (Terreberry and Neafsey 1983; Burns and Wyss
1985; Hurley-Gius and Neafsey 1986; Verberne et al.
1987; Hardy and Holmes 1988). IL is viewed as a vis-
ceromotor center (Hurley-Gius and Neafsey 1986; Neafsey
1990). PL, on the other hand, has been implicated in
cognitive processes. PL lesions have been shown to pro-
duce pronounced deficits in delayed response tasks (Brito
and Brito 1990; Seamans et al. 1995; Delatour and Gisquet-
Verrier 1996, 1999, 2000; Floresco et al. 1997; Ragozzino
et al. 1998), similar to those seen with lesions of the dor-
solateral PFC of primates (Kolb 1984; Goldman-Rakic
1994; Barbas 1995, 2000; Groenewegen and Uylings
2000).
Although efferent projections from the mPFC have been
well described in several species (Room et al. 1985; Sesack
et al. 1989; Reep et al. 1990, 2003; Chiba et al. 2001;
Hurley et al. 1991; Takagishi and Chiba 1991; Buchanan
et al. 1994; Guandalini 1998; Vertes 2002, 2004; Cheat-
wood et al. 2003; Gabbott et al. 2003, 2005), few reports
have examined afferent projections to the mPFC, specifi-
cally to its various subdivisions. To our knowledge, only a
single study (Conde et al. 1995) has compared afferents to
the four divisions of the mPFC: IL, PL, AC, and AGm.
While Conde et al. (1995) described inputs to subregions of
the mPFC in the rat, their injections were relatively large
and as they indicated often included more than one mPFC
field. By making discrete injections of the retrograde tracer,
Fluorogold (FG), into select subfields of the mPFC, we
sought to define patterns of (differential) input to the IL,
PL, AC, and AGm cortices in rats. We show that each of
the mPFC subfields exhibited a quite unique pattern of
afferent projections. These distinctive sets of afferents
undoubtedly contribute to functional differences among
mPFC fields.
Materials and methods
Sixty-two male Sprague-Dawley rats (Charles River, Wil-
mington, MA) weighing 350–450 g were injected with the
retrograde fluorescent tracer FG (Fluorochrome, Denver,
CO). These experiments were approved by the Florida
Atlantic University Institutional Animal Care and Use
Committee and conform to all federal regulations and
National Institutes of Health guidelines for the care and use
of laboratory animals.
Fluorogold was dissolved in a 0.1 M sodium acetate
buffer (pH 4.0–5) to yield a 4–5% concentration. Rats were
anesthetized using a 75 mg/kg dose of sodium pentobarbi-
tal. Single injections of FG were made iontophoretically
using glass micropipettes with outside tip diameters of 25–
50 lm into one of four medial prefrontal cortical areas in
separate rats: AGm, the AC, PL, and IL cortices. Positive
direct current (5–10 lA) was applied through a Grass
stimulator (model 88) coupled with a high-voltage stimu-
lator (FHC, Bowdoinham, ME) at 2 s ‘on’/2 s ‘off’ intervals
for 2–10 min. Following a survival time of 7 days, rats were
deeply anesthetized with sodium pentobarbital and perfused
transcardially with 100 ml of a heparinized saline wash
followed by 450 ml of fixative [4% paraformaldehyde in
0.01 M sodium phosphate buffer (PB), pH 7.4]. The brains
were then removed and stored for 48 h in a sucrose solution
(30% sucrose in 0.1 M PB) at 4�C. Following this, 50 lm
coronal sections were taken on a freezing microtome and
Brain Struct Funct (2007) 212:149–179 151
123

collected in 0.1 M PB and stored at 4�C. Six series of
sections were taken yielding a representative collection of
sections that were 300 lm apart. For the reaction, sections
of a representative series were incubated in a sodium
borohydride solution (1% sodium borohydride in 0.1 M PB)
for 30 min, and washed with 0.1 M PB four times at 6 min
each (4 · 6 min). The sections were then blocked in a Tris-
saline solution [0.5% bovine serum albumen (BSA) (Sigma
Chemicals, St. Louis, MO) 0.25% Triton X-100 (Sigma
Chemicals) in 0.1 M Tris-saline, pH 7.6] for 1 h. Following
the blocking procedure, the sections were incubated for
48 h at room temperature in primary antiserum directed
against FG (rabbit anti-FG) (Fluorochrome, LLC) at a
concentration of 1:200 in diluent (0.1% BSA and 0.25%
Triton X-100 in 0.1 M Tris-saline solution). Following
incubation in the primary antiserum, sections were washed
(4 · 6 min) in 0.1 M PB, and then incubated in a secondary
antiserum (biotinylated goat anti-rabbit IgG) (Vector Labs,
Burlingame CA) at a concentration of 1:400 in diluent for
2 h. Sections were then washed again (4 · 6 min) and
incubated in avidin-biotin complex (Vector Labs) at a 1:100
concentration in diluent for 1 h. After a final set of
4 · 6 min rinses, the peroxidase reaction product was
visualized by incubation in a solution containing 0.022% of
3,3¢ diaminobenzidine (DAB, Aldrich, Milwaukee, WI),
0.015% nickel chloride (NiCl), and 0.003% H2O2 in TBS
for 6 min. Sections were then rinsed again in PBS
(3 · 1 min) and mounted onto chrome-alum gelatin-coated
slides. An adjacent series of representative sections from
each rat was stained with cresyl violet for anatomical ref-
erence. Sections were examined using light and darkfield
optics. Injection sites and labeled cells were plotted on
representative schematic coronal sections through the brain
using sections adapted from the rat atlas of Swanson (1998).
The brightfield photomicrographs of labeled cells were ta-
ken with a Nikon DXM1200 camera mounted on a Nikon
Eclipse E600 microscope. The photomontages were con-
structed using Image-Pro Plus 4.5.1.29 (Media Cybernetics
Inc., Silver Spring, MD) and adjusted for brightness and
contrast using Adobe PhotoShop 7.0 (Mountain View, CA).
Results
The pattern of retrogradely labeled cells throughout the
brain following injections of the retrograde tracer, FG, into
the four divisions of the mPFC are described. Four repre-
sentative cases with injections in the AGm, AC, PL, and IL
cortices are illustrated and discussed in detail. The patterns
of labeling obtained with the four schematically illustrated
cases are representative of patterns found with non-illus-
trated cases.
Afferents to the medial (frontal) agranular cortex
(AGm)
Figure 1 schematically depicts the pattern of retrogradely
labeled neurons in the brain following a FG injection in
AGm. At anterior levels of the forebrain (Fig. 1a–d),
labeling was pronounced within the medial frontal polar
(FPm), orbital, medial prefrontal, and dorsal agranular
insular (AId) cortices and the CLA, and generally heavier
ipsilateral than contralateral to the injection. Specifically,
significant numbers of labeled neurons were present within
FPm, the medial (MO), ventral (VO), ventrolateral (VLO)
and lateral (LO) orbital cortices, the AGm, AC, PL and IL
of the mPFC and the AId. Labeled cells of FPm, AGm and
AC spread throughout all cortical layers, while those in PL
and IL were largely concentrated in inner layers 5/6. A
small to moderate number of labeled neurons were also
present in the anterior olfactory nucleus (AON) (Fig. 1c).
Further caudally in the anterior forebrain (Fig. 1e–h),
labeled cells continued to be present in some of the same
sites, densely within caudal regions of AGm and AC (all
layers), the CLA and AId. Labeling was present but thin-
ned in PL and IL (Fig. 1e, f). Aside from moderate labeling
in the horizontal limb of the diagonal band nucleus (DBh)
(Fig. 1h), there was a virtual absence of labeled cells in
other regions of the rostral forebrain; that is, within the
lateral (frontal) agranular (AGl), piriform, and anterior
parts of the primary (SSI) and secondary (SSII) somato-
sensory cortices, the dorsal and ventral striatum (nucleus
accumbens ACC), the olfactory tubercle (OT), and medial
and lateral septum (LS).
At mid-levels of the forebrain (Fig. 1i–l), labeled cells
were localized to dorsomedial and ventrolateral regions of
the cortex, to CLA, to parts of the basal forebrain (BF), to
midline and lateral parts of the thalamus and to the baso-
lateral nucleus of the amygdala (BLA). Cortically, the
AGm, medial parts of AGl, and the posterior agranular
insular cortex (AIp) were densely labeled, whereas AC, the
SSI and the granular insular (GI) cortices were lightly to
moderately labeled. Labeled cells spread throughout the
BF, largely confined to the DBh, ventral pallidum (VP),
substantia innominata (SI), and the magnocellular preoptic
nucleus (MA). Of these, DBh and SI were the most heavily
labeled. Figure 2a, b shows a discrete group of labeled
neurons spanning SI and VP. The location, size and general
morphological characteristics of these neurons suggest that
they may belong to the cholinergic (ACh) population of
neurons of the BF (see Discussion). Within the thalamus,
the nucleus reuniens (RE), paratenial nucleus (PT), and
ventral anterior-lateral complex (VAL) were densely la-
beled (Fig. 1k, l). A few labeled cells were present within
the medial nucleus of amygdala (MEA) and the lateral
152 Brain Struct Funct (2007) 212:149–179
123

hypothalamus (LHy) (Fig. 1k, l). With few exceptions,
labeling was predominantly ipsilateral.
More caudally in the forebrain (Fig. 1m–p), prominent
numbers of labeled neurons were observed over the lateral
convexity of cortex, in CLA, in the midline, intralaminar
and lateral parts of thalamus and in BLA (see Fig. 2c), but
with the exception of a few cells in the posterior nucleus of
the hypothalamus (PH), were noticeably absent in the
hypothalamus. As depicted (Fig. 1m–p), a continuous
stream of labeled neurons extended dorsoventrally from the
retrosplenial (RSC) and motor cortices (AGm and AGl)
through primary/secondary somatosensory and auditory
(AUD) cortices, to AIp, the ectorhinal (ECT) and perirhinal
(PRC) cortices, adjacent to the rhinal fissure. Labeling was
dense in ECT and PRC, particularly in inner layers
(Fig. 1o, p). Several nuclei of the thalamus were strongly
labeled including the paraventricular (PV), mediodorsal
(MD), interanteromedial (IAM), anteromedial (AM),
paracentral (PC), central lateral (CL), central medial (CM)
intermediodorsal (IMD), rhomboid (RH), ventromedial
PL
MO
LO
AC
VLOAId
AON
SSI
AIv
SSI
PIR
ACC
AGl
NE
CP
PIR
OT
AId
CLA
SSI
DBh
D
FPm
FPl
FPm
AId
FS
AGl
AId
AC
PL
MO
AGm
AC
PL
ILCLA
VO
AC
AGl
CLA
EN
MS
LS
AC
AGm
CP
ACC
AC
PL
IL CLA
ACC
AGm
AC
PL
IL AId
CP
AGm
OT
SSII
AIp
A
B
C
E
F
G
H
Fig. 1 Series of representative rostro-caudally aligned schematic
transverse sections (a–x) depicting the location of retrogradely
labeled cells in the brain produced by a FG injection (c) in the
medial (frontal) agranular cortex (AGm). One dot = one cell. Sections
modified from the rat atlas of Swanson (1998). See list for
abbreviations
Brain Struct Funct (2007) 212:149–179 153
123

(VM), and RE (Fig. 1m–p). Labeling was particularly
pronounced within MD, VM, RH, and RE. Figure 2d
shows a tight cluster of labeled cells ventrally on the
midline in RE, others more diffusely distributed dorsolat-
erally in RE, and third population dorsally in RH, essen-
tially outlining RH. While the entire rostrocaudal extent of
BLA was densely labeled, considerably fewer labeled
neurons were present in other nuclei of the amygdala,
namely, in the basomedial (BMA) and posterior (PA) nu-
clei (Fig. 1m–p). Additional lightly labeled sites at these
levels were the lateral posterior nucleus (LP) of thalamus
and the zona incerta (ZI).
As observed rostrally, labeling within the cortex at the
caudal diencephalon-rostral midbrain (Fig. 1q–t) was fairly
widespread, but unlike rostrally was now predominantly
confined dorsomedially to RSC, the posterior parietal area
(PAp) and the secondary visual cortex (OC2), and ven-
trolaterally to the area bordering the rhinal fissure: ECT,
PRC, and the lateral entorhinal cortex (EC). Labeling was
light within temporal (TE) regions of the cortex. Within the
hippocampus, small numbers of reacted neurons were seen
in the postero-dorsal (Fig. 1q–s) and ventral CA1 (Fig. 1r–
t). Subcortically, labeling was mainly restricted to ventral
regions of the tegmentum; prominent in the substantia ni-
SSI
AC
AGm
SSII
AIp
CP
GPCLA
EN
BST
LHySI
MA
AGm
SSI
AIp
CP
SICEA
RE
PTAM
RT
AH
GP
CS
R
SSII
AIp
CP
SIBLA
RE
PT
AM
LHy
AC
AGm
MS
LSCP
CLA
EN
DBh
GI
GI
PAp
SI
PIR
VALAV
CEA
SSI
AIp
CP
VAL
VMRE
RH
IAM
CEA
BLA
BMA
LHy
RSC
RSC
SSII
PERIRE
RH VM
MD
CP
LHyMEA
BLALA
LD
CL
AGl
VAL
RSCSSI
PERI
MDIMD
CEM
ZILA
BMA
BLA
LHy
RECP
PV
VB
PIR
CEMPO
PH
RSC
SSI
ZI
IMD
PA BLA
PERI
EC
ECT
PFLP
I
J
K
L
M
N
O
P
Fig. 1 continued
154 Brain Struct Funct (2007) 212:149–179
123
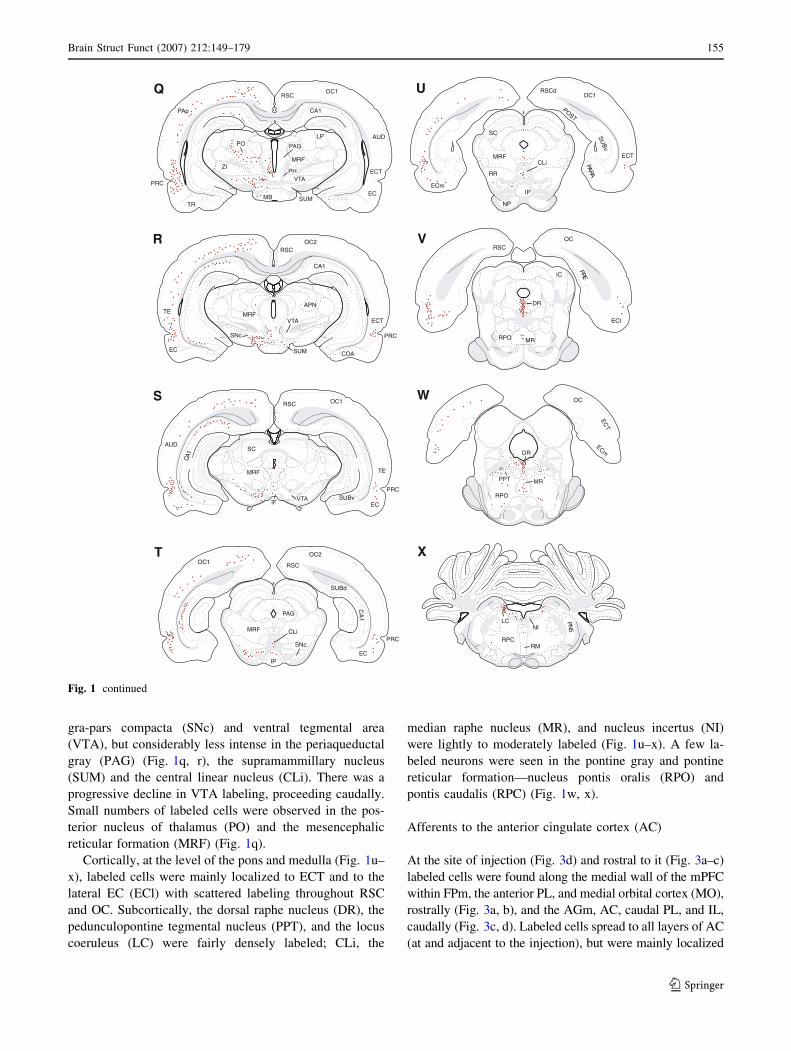
gra-pars compacta (SNc) and ventral tegmental area
(VTA), but considerably less intense in the periaqueductal
gray (PAG) (Fig. 1q, r), the supramammillary nucleus
(SUM) and the central linear nucleus (CLi). There was a
progressive decline in VTA labeling, proceeding caudally.
Small numbers of labeled cells were observed in the pos-
terior nucleus of thalamus (PO) and the mesencephalic
reticular formation (MRF) (Fig. 1q).
Cortically, at the level of the pons and medulla (Fig. 1u–
x), labeled cells were mainly localized to ECT and to the
lateral EC (ECl) with scattered labeling throughout RSC
and OC. Subcortically, the dorsal raphe nucleus (DR), the
pedunculopontine tegmental nucleus (PPT), and the locus
coeruleus (LC) were fairly densely labeled; CLi, the
median raphe nucleus (MR), and nucleus incertus (NI)
were lightly to moderately labeled (Fig. 1u–x). A few la-
beled neurons were seen in the pontine gray and pontine
reticular formation—nucleus pontis oralis (RPO) and
pontis caudalis (RPC) (Fig. 1w, x).
Afferents to the anterior cingulate cortex (AC)
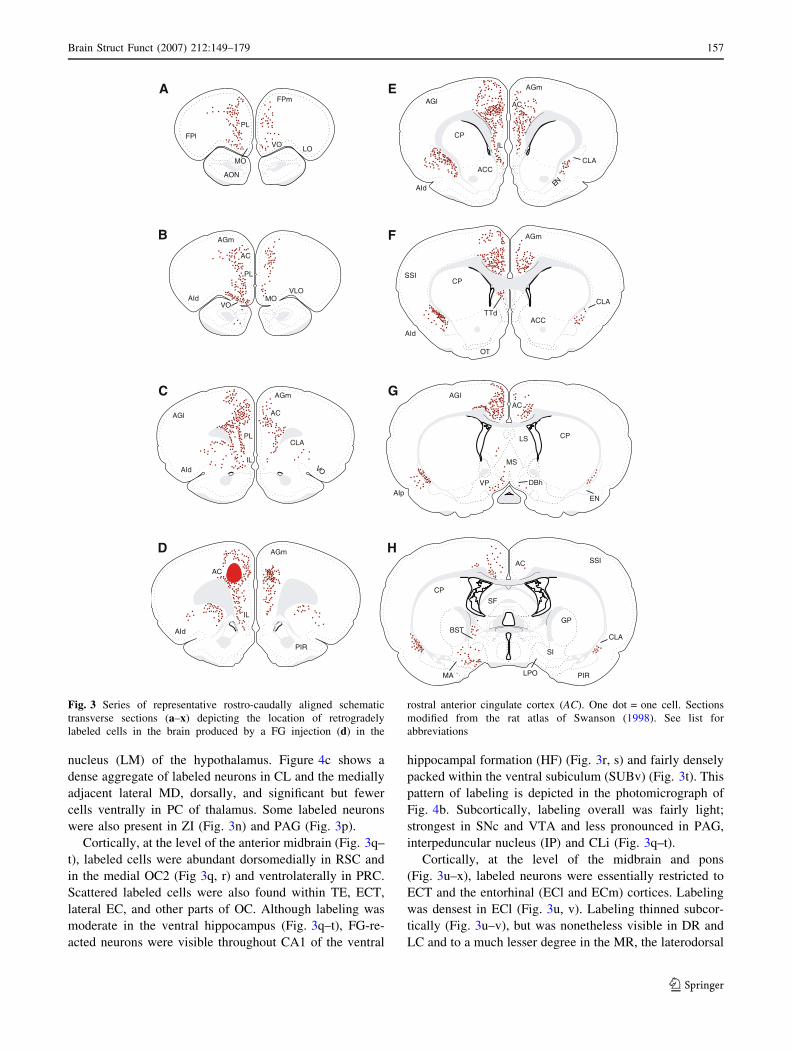
At the site of injection (Fig. 3d) and rostral to it (Fig. 3a–c)
labeled cells were found along the medial wall of the mPFC
within FPm, the anterior PL, and medial orbital cortex (MO),
rostrally (Fig. 3a, b), and the AGm, AC, caudal PL, and IL,
caudally (Fig. 3c, d). Labeled cells spread to all layers of AC
(at and adjacent to the injection), but were mainly localized
EC
PRC
ECT
OC1RSC
PAG
PH
SUMMB
VTA
MRF
POLP
ZI
AUD
PAp
TR
ECT
PRC
EC SUM
CA1
VTA
CA1
SNc
MRF
COA
OC2
TE
MRF
VTAIF EC
OC1
PRC
SC
1AC
SUBv
RSC
AUD
TE
RSC
EC
CLi
IP
PRC
OC2
1A
C
SUBd
PAG
MRF
SNc
OC1
OC1RSCd
TSOP
vB
US
ECm
ECTMRF
RR
CLi
IP
NP
SC
ARAP
DR
ECl
MR
TCE
IC ERP
OC
RPO
RSC
mCE
MRPPT
DR
OC
RPO
LCNI
RPC
5NP
RM
U
V
W
X
RSC
APN
Q
R
S
T
Fig. 1 continued
Brain Struct Funct (2007) 212:149–179 155
123
to inner layers of PL and IL. Although labeling was stronger
ipsi- than contralateral to the injection, relatively significant
numbers of labeled neurons were visible contralaterally in
the mPFC. The VO and CLA were moderately to densely
labeled; AId was lightly labeled (Fig. 3a–d).
More caudally in the anterior forebrain (Fig. 3e–h), la-
beled cells were pronounced within the caudal AC,
dorsomedially, and CLA, ventrolaterally. Light to moder-
ate numbers of labeled neurons were also present in AGm,
dorsal to AC, inner layers of AId, the dorsal taenia tecta
(TTd), and structures of the lateral BF: bed nucleus of stria
terminalis (BST), lateral preoptic area (LPO), MA, and SI.
At mid-levels of the forebrain (Fig. 3i–l), labeled cells
continued to be seen dorsomedially in the cortex, within
AC (Fig. 3i, j), extending caudally to the RSC. A few la-
beled cells were also present in motor cortices (AGm and
AGl) lateral to AC (Fig. 3k, l). Ventrolaterally, labeling
was virtually confined to CLA. As shown (Fig. 3j–l),
pronounced numbers of labeled neurons were observed
within the medial thalamus, mainly localized to PV, PT,
anteromedial (AM), IAM, lateral parts of MD, RH, CM,
and VM. The entire extent of AM was densely labeled,
particularly the ventral tier of AM (Fig. 3j, k). This is
depicted in the photomicrograph of Fig. 4a. Interestingly,
in contrast to heavy AM and IAM labeling, there was a
virtual absence of labeled neurons in the anterodorsal (AD)
and anteroventral (AV) nuclei of the anterior thalamus
(Fig. 3j–l).
Further caudally in the forebrain (Fig. 3m–p) labeled
cells were present in significant numbers in a region dorsal/
dorsolateral to the cingulum bundle (CB); that is, to AGm/
AGl, rostrally (Fig. 3m, n), and to RSC and the PAp
(Fig. 3o, p), caudally. The SSII, TE, ECT, and PRC were
moderately labeled. Subcortically, labeling was pro-
nounced in the medial and intralaminar nuclei of thalamus
and in the basal nuclei of the amygdala, but was light
within the hypothalamus (Fig. 3m–p). Specifically, labeled
neurons were fairly densely concentrated within lateral
MD, CL, PC, IMD, RH, and RE of thalamus (Fig. 3m–o)
and in the basomedial (BMA) and BLA of amygdala
(Fig. 3m, n), but loosely (and lightly) dispersed in CM of
thalamus and PH, LHy, SUM, and the lateral mammillary
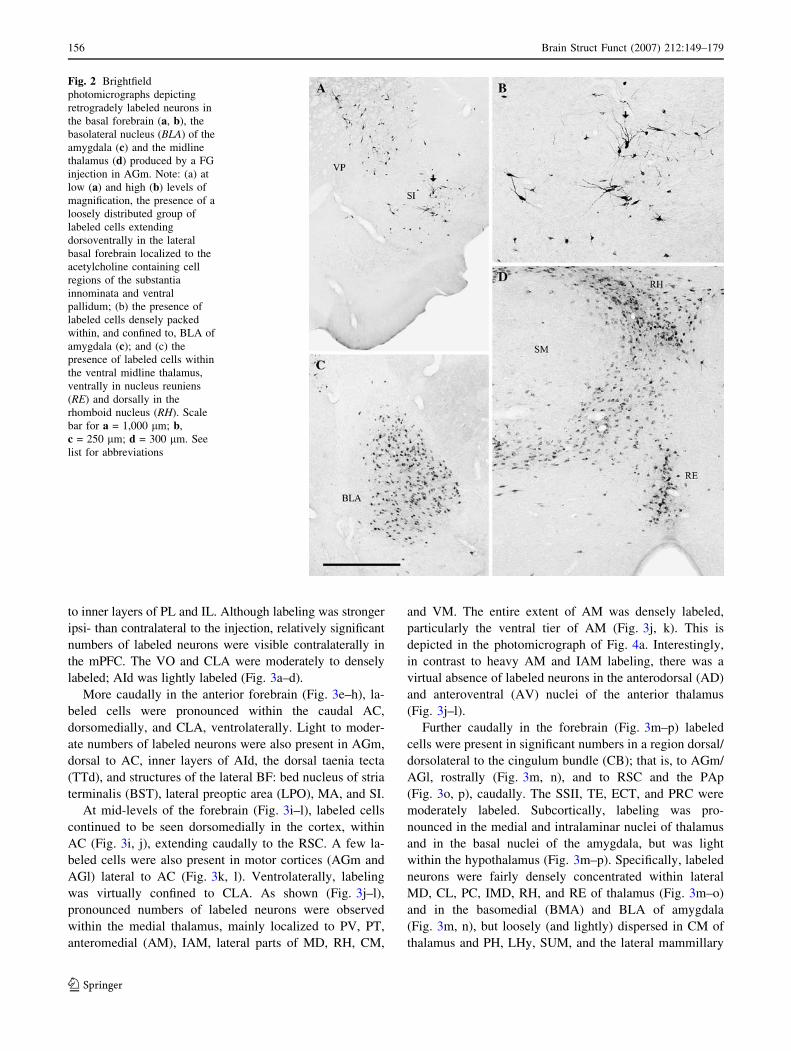
Fig. 2 Brightfield
photomicrographs depicting
retrogradely labeled neurons in
the basal forebrain (a, b), the
basolateral nucleus (BLA) of the
amygdala (c) and the midline
thalamus (d) produced by a FG
injection in AGm. Note: (a) at
low (a) and high (b) levels of
magnification, the presence of a
loosely distributed group of
labeled cells extending
dorsoventrally in the lateral
basal forebrain localized to the
acetylcholine containing cell
regions of the substantia
innominata and ventral
pallidum; (b) the presence of
labeled cells densely packed
within, and confined to, BLA of
amygdala (c); and (c) the
presence of labeled cells within
the ventral midline thalamus,
ventrally in nucleus reuniens
(RE) and dorsally in the
rhomboid nucleus (RH). Scale
bar for a = 1,000 lm; b,
c = 250 lm; d = 300 lm. See
list for abbreviations
156 Brain Struct Funct (2007) 212:149–179
123
nucleus (LM) of the hypothalamus. Figure 4c shows a
dense aggregate of labeled neurons in CL and the medially
adjacent lateral MD, dorsally, and significant but fewer
cells ventrally in PC of thalamus. Some labeled neurons
were also present in ZI (Fig. 3n) and PAG (Fig. 3p).
Cortically, at the level of the anterior midbrain (Fig. 3q–
t), labeled cells were abundant dorsomedially in RSC and
in the medial OC2 (Fig 3q, r) and ventrolaterally in PRC.
Scattered labeled cells were also found within TE, ECT,
lateral EC, and other parts of OC. Although labeling was
moderate in the ventral hippocampus (Fig. 3q–t), FG-re-
acted neurons were visible throughout CA1 of the ventral
hippocampal formation (HF) (Fig. 3r, s) and fairly densely
packed within the ventral subiculum (SUBv) (Fig. 3t). This
pattern of labeling is depicted in the photomicrograph of
Fig. 4b. Subcortically, labeling overall was fairly light;
strongest in SNc and VTA and less pronounced in PAG,
interpeduncular nucleus (IP) and CLi (Fig. 3q–t).
Cortically, at the level of the midbrain and pons
(Fig. 3u–x), labeled neurons were essentially restricted to
ECT and the entorhinal (ECl and ECm) cortices. Labeling
was densest in ECl (Fig. 3u, v). Labeling thinned subcor-
tically (Fig. 3u–v), but was nonetheless visible in DR and
LC and to a much lesser degree in the MR, the laterodorsal
PL
MO
VOLO
FPm
FPl
AON
AGm
AIdVLO
MO
PL
AC
VO
AC
PL
IL
AGm
AGl
AId
CLA
OV
IL
AGm
AGm
AC
ILCP
ACCCLA
NEAId
AGl
AGm
AId
ACC
CP
CLA
TTd
SSI
OT
AGlAC
MS
LS
DBh
CP
ENAIp
VP
AC
MA LPO
SI
CP
GP
PIR
BST
AC
AId
SSI
CLAPIR
SF
A
B
C
D
E
F
G
H
Fig. 3 Series of representative rostro-caudally aligned schematic
transverse sections (a–x) depicting the location of retrogradely
labeled cells in the brain produced by a FG injection (d) in the
rostral anterior cingulate cortex (AC). One dot = one cell. Sections
modified from the rat atlas of Swanson (1998). See list for
abbreviations
Brain Struct Funct (2007) 212:149–179 157
123

tegmental nucleus (LDT), the lateral parabrachial nucleus
(PBl), and NI.
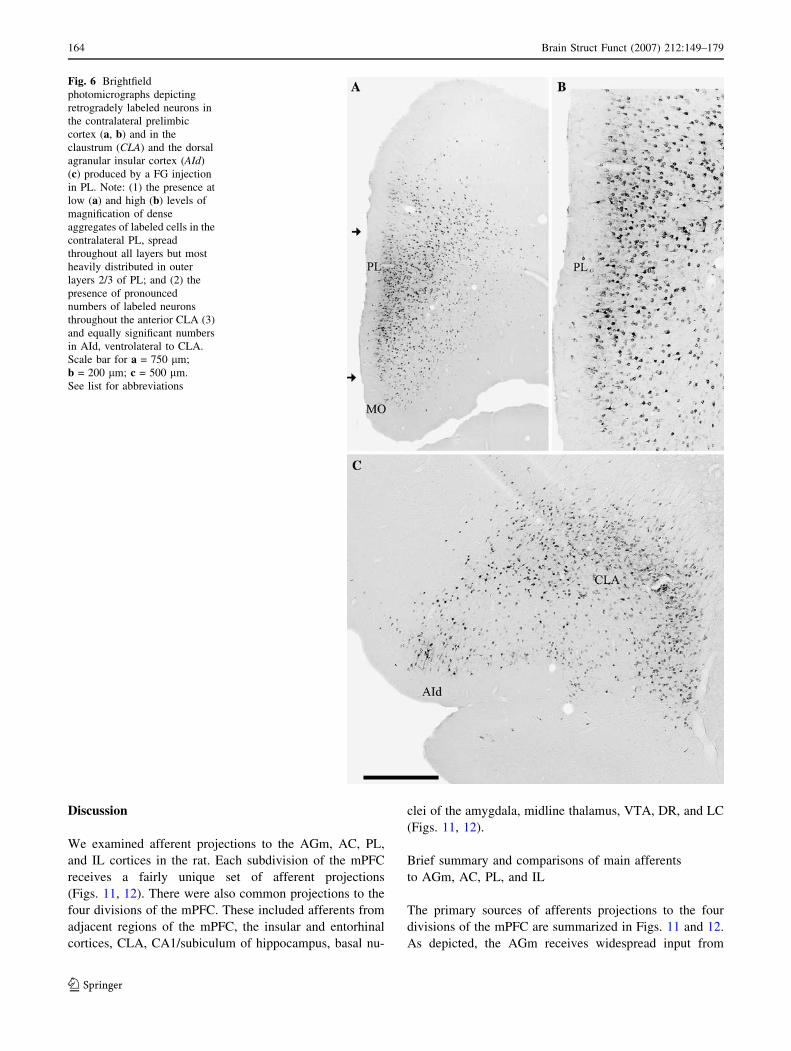
Afferents to the prelimbic cortex (PL)
As depicted (Fig. 5a–d), pronounced numbers of labeled
neurons were present in the PFC, mainly the medial PFC,
following a FG injection in PL. Labeling was predomi-
nantly localized to FPm, MO, VO, AGm, AC, PL (adjacent
to the injection and contralaterally), and IL, spreading
fairly evenly throughout all cortical layers of these regions.
The photomicrographs of Fig. 6a, b depict heavy labeling
contralaterally in anterior PL and MO, rostral to the
injection. Light to moderate numbers of labeled neurons
were present in VLO, CLA, AId, and AON (Fig. 5c, d).
More caudally in the anterior forebrain (Fig. 5e–h),
labeling remained pronounced within the mPFC, strongest in
AC, PL and IL, but was also dense in ventrolateral aspects of
the cortex particularly in the agranular insular cortex (AId
and AIv), CLA, and the endopiriform nucleus (EN). Fig-
ure 6c shows heavy concentrations of labeled cells in CLA
and AId, ipsilaterally. Additional labeled sites included the
TTd, and the LS (Fig. 5f, g). Labeling was considerably
stronger ipsi- than contralaterally in each of these structures.
At mid-levels of the anterior forebrain (Fig. 5i–l), label-
ing was restricted to dorsomedial and ventrolateral aspects of
AC
RE
RSC
CEA
PTAM
AIp
AV
AH
GI
RSC
SSp
AIp
CP
CEA
RE
PT
AM IAM
LHy
RT
VAL
CLA
AGl
PIR
RSC
SSII CP
VMRE
MD
ALB
LHy
RH
AD
MD
RERHCM
BLA
AL
BMA
CL
PCIMDVB
PIR
PRC
SSIRSC
SSII
AUD
AGl
RSC
ECT
PRC
MD
ZI
LA
BMA
RE
CL
LHy
LD
CM
M
N
O
P
AUD
PV
PHLHy
RSC
ZI
IMD PF
CL
COA
LP
VB
EC
PRC
RSC
PAp
LP
AGm
SSI
PT
SSII
RE
PAG
SUM
VTA
LMEC
ECT
PV
SI
CP
LHyAIp
CLA
RT
CA1
ZI
1AC
PAp
MRF
I
J
K
L
AUD
Fig. 3 continued
158 Brain Struct Funct (2007) 212:149–179
123
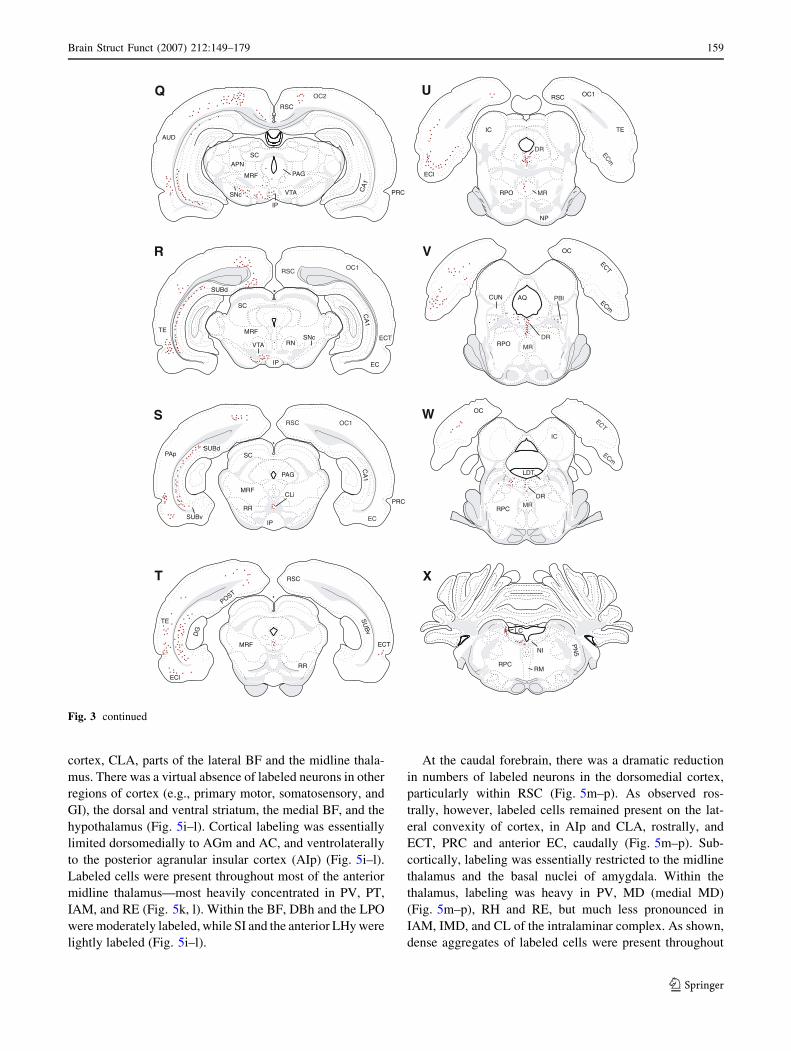
cortex, CLA, parts of the lateral BF and the midline thala-
mus. There was a virtual absence of labeled neurons in other
regions of cortex (e.g., primary motor, somatosensory, and
GI), the dorsal and ventral striatum, the medial BF, and the
hypothalamus (Fig. 5i–l). Cortical labeling was essentially
limited dorsomedially to AGm and AC, and ventrolaterally
to the posterior agranular insular cortex (AIp) (Fig. 5i–l).
Labeled cells were present throughout most of the anterior
midline thalamus—most heavily concentrated in PV, PT,
IAM, and RE (Fig. 5k, l). Within the BF, DBh and the LPO
were moderately labeled, while SI and the anterior LHy were
lightly labeled (Fig. 5i–l).
At the caudal forebrain, there was a dramatic reduction
in numbers of labeled neurons in the dorsomedial cortex,
particularly within RSC (Fig. 5m–p). As observed ros-
trally, however, labeled cells remained present on the lat-
eral convexity of cortex, in AIp and CLA, rostrally, and
ECT, PRC and anterior EC, caudally (Fig. 5m–p). Sub-
cortically, labeling was essentially restricted to the midline
thalamus and the basal nuclei of amygdala. Within the
thalamus, labeling was heavy in PV, MD (medial MD)
(Fig. 5m–p), RH and RE, but much less pronounced in
IAM, IMD, and CL of the intralaminar complex. As shown,
dense aggregates of labeled cells were present throughout
OC2
APN
IP
RSCOC1
SC
OC1RSC
EC
1A
C
SUBd
SUBv
SUBd
RR
MRF
SC
CLi
IP
PAG
AUD
TE
PAp
vB
US
ECT
GD
TE
ECl
TE
TSOP
RSC
PRC
EC
ECTVTA
SNc
1A
C
PRC
1A
CVTASNc
PAG
RSC
SC
MRF
RN
MRF
IP
MRF
RR
mCE
DR
MR
OC1
NP
RPO
IC
ECl
RSC
OC
MR
PBl
DR
TCE
AQCUN
RPO
OC
mCE
MR
IC
TCE
mCE
LDT
DR
RPC
NI
LC
RPC
5N
P
RM
Q
R
S
T
U
V
W
X
Fig. 3 continued
Brain Struct Funct (2007) 212:149–179 159
123

the extent of BLA (Figs. 5m–p, 7b). Light to moderate
numbers were also seen in BMA, the posterior (PA) and
anterior cortical nuclei (COA) of amygdala (Fig. 5o, p).
Cortically, at the level of the caudal diencephalon
(Fig. 5q–t), reacted cells were mainly restricted to the
parahippocampal cortices and HF; that is, moderate label-
ing in ECT, PRC and lateral EC, and dense labeling in CA1
of the ventral hippocampus extending dorso-ventrally
throughout CA1 of the ventral HF (Fig. 5s, t). The prom-
inent CA1 labeling is depicted in the photomicrograph of
Fig. 7a. There was a noticeable absence of labeling in
remaining regions of the cortex, including RSC, PAp, and
OC (Fig. 5q–t). With the exception of fairly dense labeling
of BLA/BMA (Fig. 7c) as well as the amygdalo-piriform
transition zone (TR), subcortical labeling was confined to
relatively few structures. Lightly to moderately labeled
sites were PAG, VTA, PH, IP, SUM, and LM (Fig. 5q–t).
At the pons-medulla (Fig. 5u–x), labeling was essen-
tially confined cortically to the ventral subiculum (Fig. 5u)
which was densely labeled, and ECl which was moderately
labeled. Subcortically, the DR was densely labeled; MR
and LC were moderately labeled (Fig. 5u–x).
Injections in other parts of PL resulted in the same
general pattern of labeling, but some differences in relative
densities of labeling. For instance, rostral (present case)
compared to caudal PL injections produced stronger cell
labeling in the CA1, BLA of amygdala, medial septum
(MS) and diagonal band nuclei and DR and MR of the
brainstem, while caudal injections gave rise to heavier
labeling in several regions of the BF including CLA, MA,
SI, and the VP.
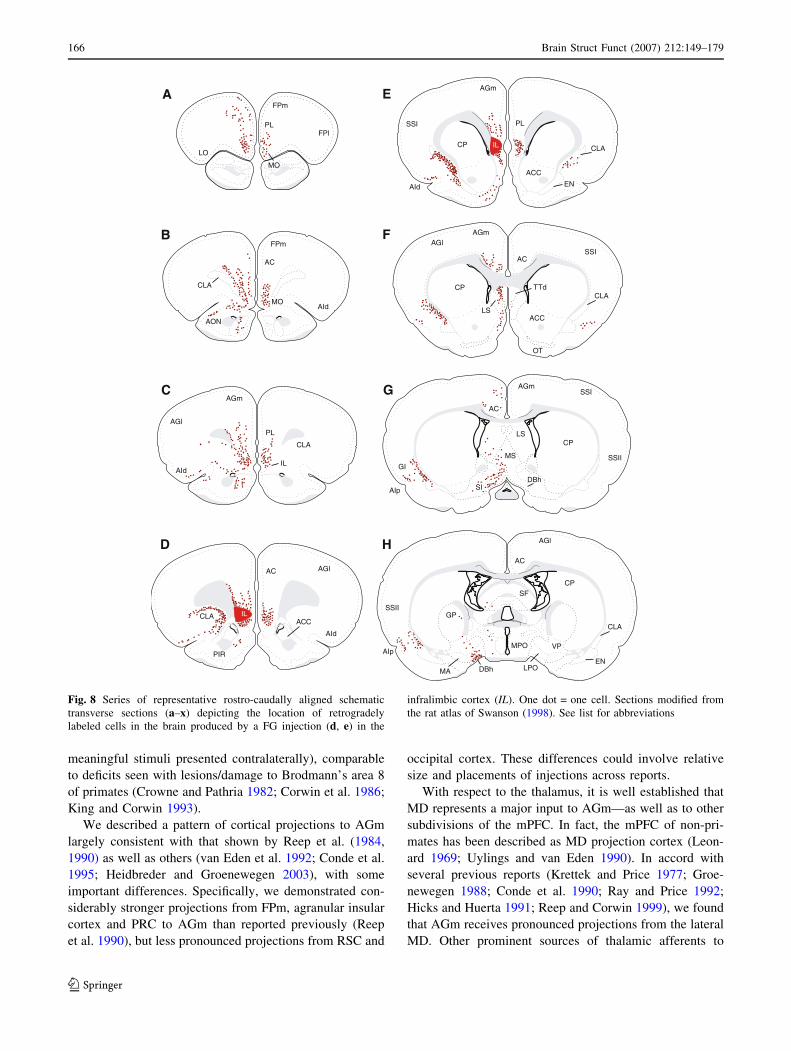
Afferents to the infralimbic cortex (IL)
Similar to injections in dorsal regions of the mPFC, sig-
nificant numbers of labeled cells were observed in anterior
regions of the forebrain (Fig. 8a–d) with IL injections
(Fig. 8d, e). Rostrally within mPFC, labeling extended
dorsoventrally throughout the mPFC to FPm, PL, and MO
(Fig. 8a, b), but caudally was mainly confined to the ven-
Fig. 4 Brightfield
photomicrographs depicting
retrogradely labeled neurons at
anterior (a) and posterior (c)
levels of the thalamus and
within the ventral hippocampus
(b) produced by a FG injection
in AC. Note: (1) the presence of
pronounced numbers of labeled
cells in the anteromedial
nucleus of thalamus (a) and
fewer numbers ventrally in
nucleus reuniens (a) and
dorsally in paratenial nucleus
(a) of thalamus; (2) the presence
of significant numbers of
labeled neurons in the lateral
mediodorsal nucleus (MD) and
the laterally adjacent central
lateral nucleus of thalamus (c)
as well as ventromedially in the
paracentral nucleus (c) but an
absence of labeling in the
medial and central MD; and (3)
the presence of moderate
numbers of labeled cells spread
dorsoventrally throughout CA1
of the ventral hippocampus (b).
Scale bar for a, b = 500 lm;
c = 300 lm. See list for
abbreviations
160 Brain Struct Funct (2007) 212:149–179
123
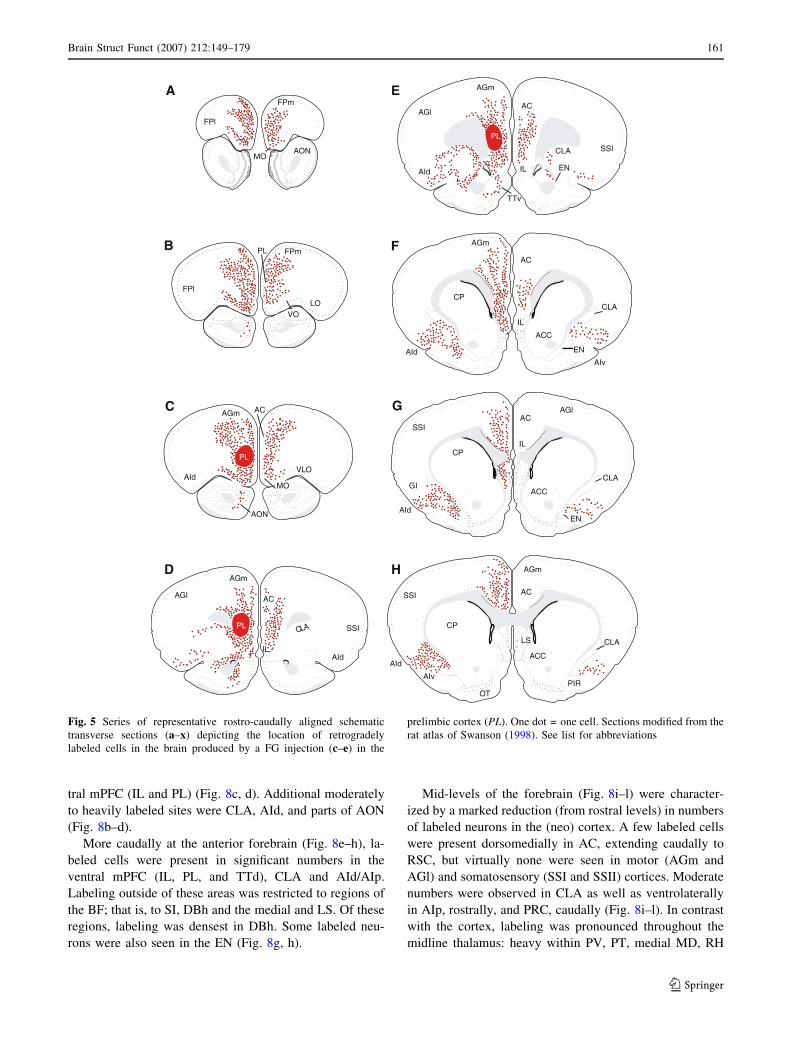
tral mPFC (IL and PL) (Fig. 8c, d). Additional moderately
to heavily labeled sites were CLA, AId, and parts of AON
(Fig. 8b–d).
More caudally at the anterior forebrain (Fig. 8e–h), la-
beled cells were present in significant numbers in the
ventral mPFC (IL, PL, and TTd), CLA and AId/AIp.
Labeling outside of these areas was restricted to regions of
the BF; that is, to SI, DBh and the medial and LS. Of these
regions, labeling was densest in DBh. Some labeled neu-
rons were also seen in the EN (Fig. 8g, h).
Mid-levels of the forebrain (Fig. 8i–l) were character-
ized by a marked reduction (from rostral levels) in numbers
of labeled neurons in the (neo) cortex. A few labeled cells
were present dorsomedially in AC, extending caudally to
RSC, but virtually none were seen in motor (AGm and
AGl) and somatosensory (SSI and SSII) cortices. Moderate
numbers were observed in CLA as well as ventrolaterally
in AIp, rostrally, and PRC, caudally (Fig. 8i–l). In contrast
with the cortex, labeling was pronounced throughout the
midline thalamus: heavy within PV, PT, medial MD, RH
VOLO
FPl
FPmPL
AGm
AIdMO
PL
AC
VLO
AON
AGl
ALC SSI
AGm
EN
CLA
FPm
FPl
AGm
AId
PL
IL
AC
AGm
AC
LS
CP
AC
IL
AGl
GI
AId
ACC
CP
CLA
AGm
AC
IL
CPCLA
AC
PL
ILAId
CLAAONMO
EN
TTv
AGl
ACC
ENAIdAIv
SSI
OTPIR
AId
AIv
SSI
ACC
A
B
C
D
E
F
G
H
SSI
Fig. 5 Series of representative rostro-caudally aligned schematic
transverse sections (a–x) depicting the location of retrogradely
labeled cells in the brain produced by a FG injection (c–e) in the
prelimbic cortex (PL). One dot = one cell. Sections modified from the
rat atlas of Swanson (1998). See list for abbreviations
Brain Struct Funct (2007) 212:149–179 161
123
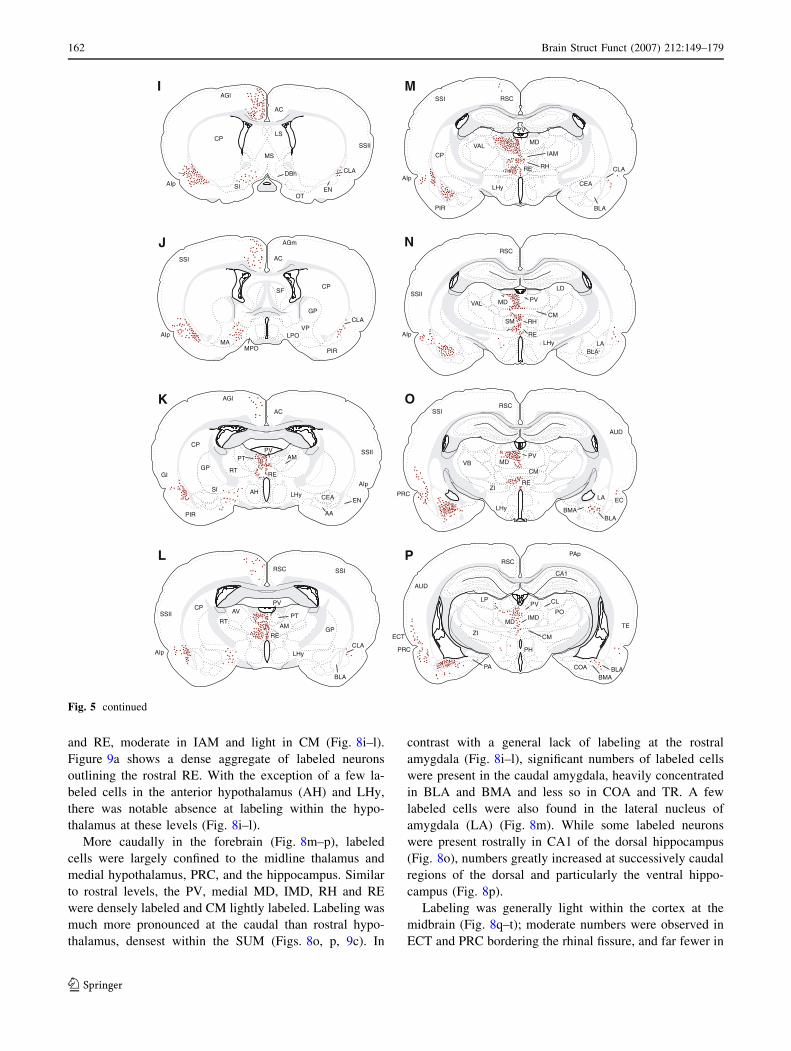
and RE, moderate in IAM and light in CM (Fig. 8i–l).
Figure 9a shows a dense aggregate of labeled neurons
outlining the rostral RE. With the exception of a few la-
beled cells in the anterior hypothalamus (AH) and LHy,
there was notable absence at labeling within the hypo-
thalamus at these levels (Fig. 8i–l).
More caudally in the forebrain (Fig. 8m–p), labeled
cells were largely confined to the midline thalamus and
medial hypothalamus, PRC, and the hippocampus. Similar
to rostral levels, the PV, medial MD, IMD, RH and RE
were densely labeled and CM lightly labeled. Labeling was
much more pronounced at the caudal than rostral hypo-
thalamus, densest within the SUM (Figs. 8o, p, 9c). In
contrast with a general lack of labeling at the rostral
amygdala (Fig. 8i–l), significant numbers of labeled cells
were present in the caudal amygdala, heavily concentrated
in BLA and BMA and less so in COA and TR. A few
labeled cells were also found in the lateral nucleus of
amygdala (LA) (Fig. 8m). While some labeled neurons
were present rostrally in CA1 of the dorsal hippocampus
(Fig. 8o), numbers greatly increased at successively caudal
regions of the dorsal and particularly the ventral hippo-
campus (Fig. 8p).
Labeling was generally light within the cortex at the
midbrain (Fig. 8q–t); moderate numbers were observed in
ECT and PRC bordering the rhinal fissure, and far fewer in
AGl
AC
CLA
MS
LS
DBh
CP
ENAIp
OT
SI
SSII
CP
GPCLA
AGm
AC
AIp LPO
SF
MPOMA
SSI
J
K
L
M
N
O
P
VP
PIR
SSII
SSI
AC
AGl
AIp
CP
SI
RE
PVPT AM
RT
LHy
GI
ENCEA
AAPIR
GP
AH
CLA
BLA
PT
AM
AIp
RSC
RE
AV
RT
PV
LHy
CP
GP
SSII
RSCSSI
AIp
CP
RE RH
MD
CEA
BLA
CLA
LHy
VALIAM
PV
PIR
RSC
SSII
AIp RE
RH
MD
CM
LHyBLA
LA
LD
PVVAL
SM
RSCSSI
PRCEC
MD
CM
ZI
LA
BMABLA
LHy
PV
RE
VB
AUD
RSC
MD
CMECT
PRC
PA
PH
BMA
PV
IMD
LP
ZI
CA1
PO
CL
COA BLA
I
TE
PAp
AUD
Fig. 5 continued
162 Brain Struct Funct (2007) 212:149–179
123

RSC and ECl. As seen rostrally (Fig. 8p), dorsal and
ventral aspects of CA1 were densely labeled and merged to
form a continuous band of labeled cells, dorsoventrally in
CA1 (Fig. 8q–s). This is depicted in Fig. 10a. As with
CA1, labeled cells spread heavily throughout the extent of
the ventral subiculum (Figs. 8t, 10b). Subcortically, label-
ing was predominantly confined medially to the rostral and
central linear nuclei and to VTA (Fig. 8q–t). Some labeled
cells were present in the mesencephalic PAG, rostrally
(Fig. 8o, p) as well as caudally (Fig. 8t).
Labeling within the cortex at the level of the pons/me-
dulla (Fig. 8u–x) was essentially confined to ECT and ECl.
A few labeled cells were also found scattered throughout
OC (Fig. 8u, v). Subcortically, labeling was prominent in
DR (Figs. 8u–w, 9b), in the isthmus region between DR
and MR (Fig. 8w), in LDT and in LC (Fig. 9d), moderate
in MR (Fig. 9c) and fairly light in the pontine central gray
and NI (Fig. 8x).
Similar to PL (see above), there were differences in
relative densities of labeling, but not overall patterns of
labeling with injections in different parts of IL. Specifi-
cally, denser labeling was observed in rostral parts of BLA
with rostral compared to caudal (above case) IL injections,
and considerably fewer labeled cells were seen in the
caudal midline thalamus (caudal PV and IMD) with
superficial (layers 1–3) relative to deep IL injections.
RSC
CA1ECTVTA
PAG
ZI
SUMLM MB
COABLA
TR
EC
MRF
LP
PAp
AUD
SUM
CA1
1AC
ECl
PAG
PH
VTA
PRC
MRF
LP
TR
AUD
OC2
TE
RSCOC1
PRC
1A
C
SUBv
VTA
SNcRN
MRF
SC
ECl
S
T
U
V
W
XOC2RSC
PRC
EClVTA
1A
C
SUBd
SUBvIP
MRF
RR
SC
GD
Q
R
TE
PAG vB
US
ECT
EClCLi
SC
RR
MRF
IP
NP
RSCOC1
mCE
TE
TSOP
DR
SC
MRRPO
PPT
OC1
ERP
RSC
ECT
DR
MR
IC
PAG
RPO
NLL
OC
mCE
LC
rNS
RPC
V4
RM
Fig. 5 continued
Brain Struct Funct (2007) 212:149–179 163
123
Discussion
We examined afferent projections to the AGm, AC, PL,
and IL cortices in the rat. Each subdivision of the mPFC
receives a fairly unique set of afferent projections
(Figs. 11, 12). There were also common projections to the
four divisions of the mPFC. These included afferents from
adjacent regions of the mPFC, the insular and entorhinal
cortices, CLA, CA1/subiculum of hippocampus, basal nu-
clei of the amygdala, midline thalamus, VTA, DR, and LC
(Figs. 11, 12).
Brief summary and comparisons of main afferents
to AGm, AC, PL, and IL
The primary sources of afferents projections to the four
divisions of the mPFC are summarized in Figs. 11 and 12.
As depicted, the AGm receives widespread input from
Fig. 6 Brightfield
photomicrographs depicting
retrogradely labeled neurons in
the contralateral prelimbic
cortex (a, b) and in the
claustrum (CLA) and the dorsal
agranular insular cortex (AId)
(c) produced by a FG injection
in PL. Note: (1) the presence at
low (a) and high (b) levels of
magnification of dense
aggregates of labeled cells in the
contralateral PL, spread
throughout all layers but most
heavily distributed in outer
layers 2/3 of PL; and (2) the
presence of pronounced
numbers of labeled neurons
throughout the anterior CLA (3)
and equally significant numbers
in AId, ventrolateral to CLA.
Scale bar for a = 750 lm;
b = 200 lm; c = 500 lm.
See list for abbreviations
164 Brain Struct Funct (2007) 212:149–179
123

non-limbic and limbic regions of the cortex as well as
from specific (relay) and ‘non-specific’ nuclei of the
thalamus. In like manner, AC receives afferents from
diverse regions of cortex, but less dense from non-limbic
cortex and more restricted from limbic cortex than those
to AGm (Fig. 11b). There is a considerable overlap in
thalamic projections to AGm and AC, with the notable
exception that the anterior medial nucleus of thalamus
distributes densely to AC. There is a dramatic shift in
cortical and thalamic projections to the ventral (PL and
IL) compared to the dorsal (AGm and AC) mPFC, such
that major inputs to ventral mPFC predominantly origi-
nate from limbic cortices and from the midline thalamus.
All regions of the mPFC receive complementary (over-
lapping) projections from the BF, amygdala, hypothala-
mus and brainstem (Figs. 11, 12), with some important
differences among divisions. For example, regions of the
lateral BF primarily target AGm/AC, while the medial BF
targets PL/IL, and the amygdala distributes more heavily
to the ventral than to the dorsal mPFC. Overall hypo-
thalamic projections to the mPFC are light, with the
exception of relatively pronounced projections from the
SUM to IL. Finally, the VTA and the DR distribute
significantly to all divisions of the mPFC.
Afferents to the four divisions of the mPFC:
comparisons with previous studies and functional
implications
Medial agranular cortex
The main sources of input to AGm were from the cortex
and thalamus. Extra-thalamic subcortical afferents to AGm
were moderate and less pronounced than to other divisions
of the mPFC.
The AGm of rats is also termed the medial precentral
area (or Fr2 region of Zilles 1985) and is partially coex-
tensive with the secondary motor cortex (Zilles and Wree
1995; Gabbott et al. 2005). In an early examination of
cortical connections of AGm, Reep et al. (1990) described
diverse cortical inputs to AGm from the cortex FPm, the
medial and ventrolateral orbital cortices, AGl (or primary
motor cortex), insular cortex (INC), SSI and SSII, RSC,
auditory and occipital cortices. Based on ‘extensive cor-
tico-cortical connections,’ Reep et al. (1990) proposed that
AGm is a multimodal association area with direct actions
on motor systems, and accordingly serves a central role in
directed spatial attention. Supporting this, lesions of AGm
in rats produce contralateral neglect (failure to attend to
Fig. 7 Brightfield
photomicrographs depicting
retrogradely labeled neurons in
CA1and the dorsal and ventral
subiculum of the ventral
hippocampus (a) and rostral (b)
and caudal (c) levels of the
amygdala produced by a FG
injection in PL. Note: (1) the
presence of pronounced cell
labeling throughout the extent
of the ventral hippocampus,
from the dorsal subiculum
through CA1 to the ventral
subiculum (a); (2) the presence
of labeled neurons, confined to,
and distributed throughout the
anterior basolateral nucleus of
amygdala (BLA); and (3) the
presence of labeled cells
caudally in the amygdala,
mediolaterally spanning the
posterior part of the basomedial
nucleus (BMAp), posterior BLA
and the amygdalo-piriform
transition zone (TR). Scale bar
for a = 1,100 lm; b = 500 lm;
c = 750 lm. See list for
abbreviations
Brain Struct Funct (2007) 212:149–179 165
123
meaningful stimuli presented contralaterally), comparable
to deficits seen with lesions/damage to Brodmann’s area 8
of primates (Crowne and Pathria 1982; Corwin et al. 1986;
King and Corwin 1993).
We described a pattern of cortical projections to AGm
largely consistent with that shown by Reep et al. (1984,
1990) as well as others (van Eden et al. 1992; Conde et al.
1995; Heidbreder and Groenewegen 2003), with some
important differences. Specifically, we demonstrated con-
siderably stronger projections from FPm, agranular insular
cortex and PRC to AGm than reported previously (Reep
et al. 1990), but less pronounced projections from RSC and
occipital cortex. These differences could involve relative
size and placements of injections across reports.
With respect to the thalamus, it is well established that
MD represents a major input to AGm—as well as to other
subdivisions of the mPFC. In fact, the mPFC of non-pri-
mates has been described as MD projection cortex (Leon-
ard 1969; Uylings and van Eden 1990). In accord with
several previous reports (Krettek and Price 1977; Groe-
newegen 1988; Conde et al. 1990; Ray and Price 1992;
Hicks and Huerta 1991; Reep and Corwin 1999), we found
that AGm receives pronounced projections from the lateral
MD. Other prominent sources of thalamic afferents to
PL
MO
LO
FPm
FPl
AGl
CLA
FPm
CLATTd
OT
AC
SI
SSI
AC
LPOMA DBh
AIdMO
AC
AGl
AGm
PL
ILAId
CLA
AId
AC
IL
MPO
EN
CLA
AGm
AIp
MS
DBh
CPLS
AGm
AC
CP
ACCLS
IL
PL
CP CLA
ACC
EN
AGm
AON
CLAACC
PIR
AId
AGl
GP
SFCP
AIp
SSII
A
B
C
D
E
F
G
H
SSI
SSI
SSIIGI
AGl
VP
Fig. 8 Series of representative rostro-caudally aligned schematic
transverse sections (a–x) depicting the location of retrogradely
labeled cells in the brain produced by a FG injection (d, e) in the
infralimbic cortex (IL). One dot = one cell. Sections modified from
the rat atlas of Swanson (1998). See list for abbreviations
166 Brain Struct Funct (2007) 212:149–179
123

AGm include the CL, PC, CM, posterior (PO), VAL, and
VM nuclei of thalamus (Herkenham 1979; Conde et al.
1990; Hicks and Huerta 1991; Reep and Corwin 1999,
present results). Reep and Corwin (1999) reported that
afferents to successively more caudal regions of AGm
originate from more caudal and lateral parts of the thala-
mus. Consistent with this, Hicks and Huerta (1991) de-
scribed projections from the lateral dorsal (LD) and LP
nuclei of thalamus to the caudal but not to the rostral AGm,
and we found that injections in the rostral AGm gave rise to
few labeled cells in LD or LP. Hicks and Huerta (1991)
proposed that visuomotor thalamic input (LD/LP) to the
caudal AGm supports a role for this area in visuomotor
functions.
In addition to prominent afferents from somatomotor/
visuomotor regions of thalamus to AGm, some reports
(Conde et al. 1990; Hicks and Huerta 1991; Vertes et al.
2006), but not others (Reep and Corwin 1999), have
demonstrated significant input to AGm from the midline
thalamus. We described projections from the PV, PT, IAM,
IMD, CM, rhomboid (RH) and reuniens (RE) nuclei of the
midline thalamus to AGm, most heavily from RH and RE.
The latter is consistent with our recent demonstration
(Vertes et al. 2006), using anterograde tracers, of pro-
nounced RH and RE projections to AGm, mainly to layers
1 and 5/6. Other studies have similarly described IAM,
CM, RH, and RE projections to AGm (Conde et al. 1990;
Hicks and Huerta 1991).
GI
AC
FI
MD
RSCSSI
AIp
ALB
RERH CLA
PV
IAM
VM
LHy
VAL
PIR
CEA
CP
TE
SSII
PV
CM
MD
RERH
CM
IMD
BLALA
PIR
VB
RSC
L
RSC
MD
CMECT
PHLHy
BMABLA
PA
PV
IMD
ZI
LP
AUD
PAp
PAp
TE
AUD
SUMm
SUMm
RSC
CA1
1
CA1
SUBvSUM
VTA
SNc
BLAAEM
SC3A
C
TRCOA
EC
RSCOC2
PAp
AUD
GI
AIp
CP
RE
AMPT
GP
SI
CLA
BLA
LHy
AGm
SSI
AIpLHy
PV
IAM
RE
AM
RSC
GI
PRC
MD
RH
RE
PRC
PRC
ECT
SUM
PH
PAG
PV
PIR
CEAAH
AGl
AV
VAL
RH
CEA
PIR
VBSSII
LHy
LD
MB
BLACOA
ZI
TR
EC
LP
MRF
I
J
K
M
N
O
P
SSII
AGl
Fig. 8 continued
Brain Struct Funct (2007) 212:149–179 167
123
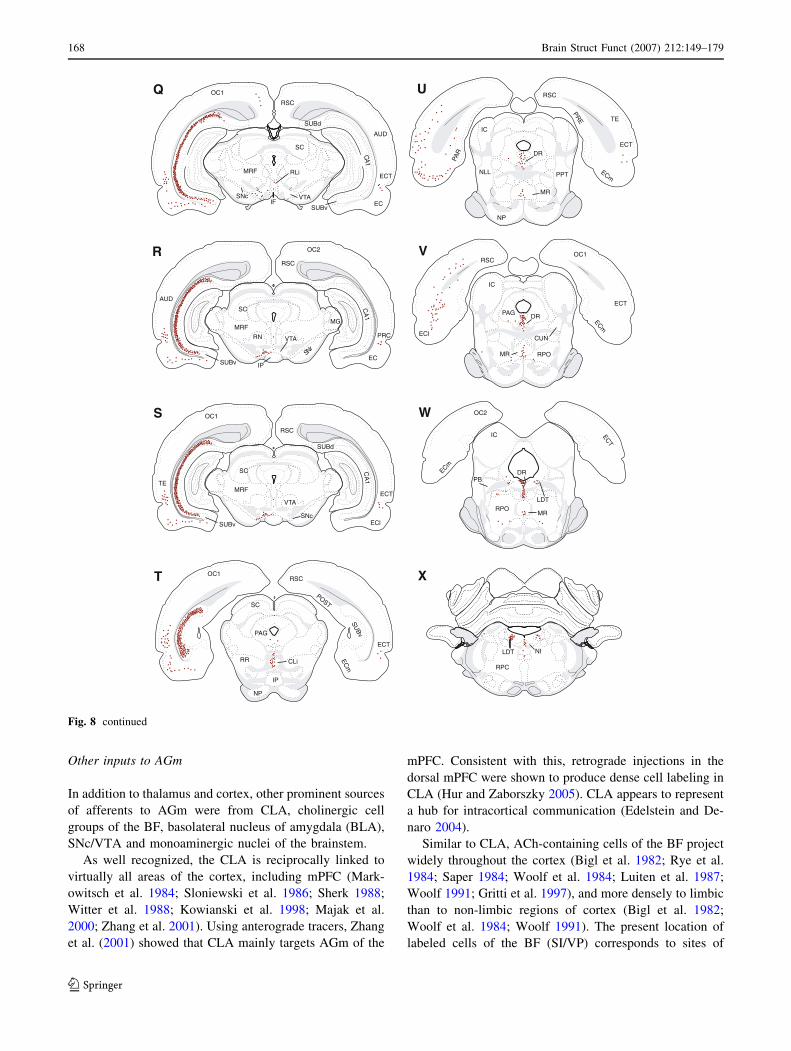
Other inputs to AGm
In addition to thalamus and cortex, other prominent sources
of afferents to AGm were from CLA, cholinergic cell
groups of the BF, basolateral nucleus of amygdala (BLA),
SNc/VTA and monoaminergic nuclei of the brainstem.
As well recognized, the CLA is reciprocally linked to
virtually all areas of the cortex, including mPFC (Mark-
owitsch et al. 1984; Sloniewski et al. 1986; Sherk 1988;
Witter et al. 1988; Kowianski et al. 1998; Majak et al.
2000; Zhang et al. 2001). Using anterograde tracers, Zhang
et al. (2001) showed that CLA mainly targets AGm of the
mPFC. Consistent with this, retrograde injections in the
dorsal mPFC were shown to produce dense cell labeling in
CLA (Hur and Zaborszky 2005). CLA appears to represent
a hub for intracortical communication (Edelstein and De-
naro 2004).
Similar to CLA, ACh-containing cells of the BF project
widely throughout the cortex (Bigl et al. 1982; Rye et al.
1984; Saper 1984; Woolf et al. 1984; Luiten et al. 1987;
Woolf 1991; Gritti et al. 1997), and more densely to limbic
than to non-limbic regions of cortex (Bigl et al. 1982;
Woolf et al. 1984; Woolf 1991). The present location of
labeled cells of the BF (SI/VP) corresponds to sites of
SNc
MRF
VTAIF
RSC
EC
ECT
SUBv
SC
RLi
SUBd
Q
R
OC1
AUD
AUD
RSC
1AC
RN
MRFMG
IP
rNS
SC
OC2
RSC
OC1
TE
CLi
1A
C
SUBd
SUBv
VTA
ECl
MRF
SC
SNc
ECT
RR
PAG
SC
DR
RSC
ERP
RAP
mCE
ECT
MR
NLL
NP
PPT
IC
mCE
ECl
ECT
DR
MR
PAG
IC
RPO
CUN
OC1RSC
MR
PB
IC
RPO
DR
LDT
OC2
mCE
TCE
S
T
U
V
W
X
NILDT
RPC
PRC
EC
1A
C
VTA
RSC
TSOP
mC
E
ECT
vB
US
SUBv
NP
IP
OC1
TE
Fig. 8 continued
168 Brain Struct Funct (2007) 212:149–179
123
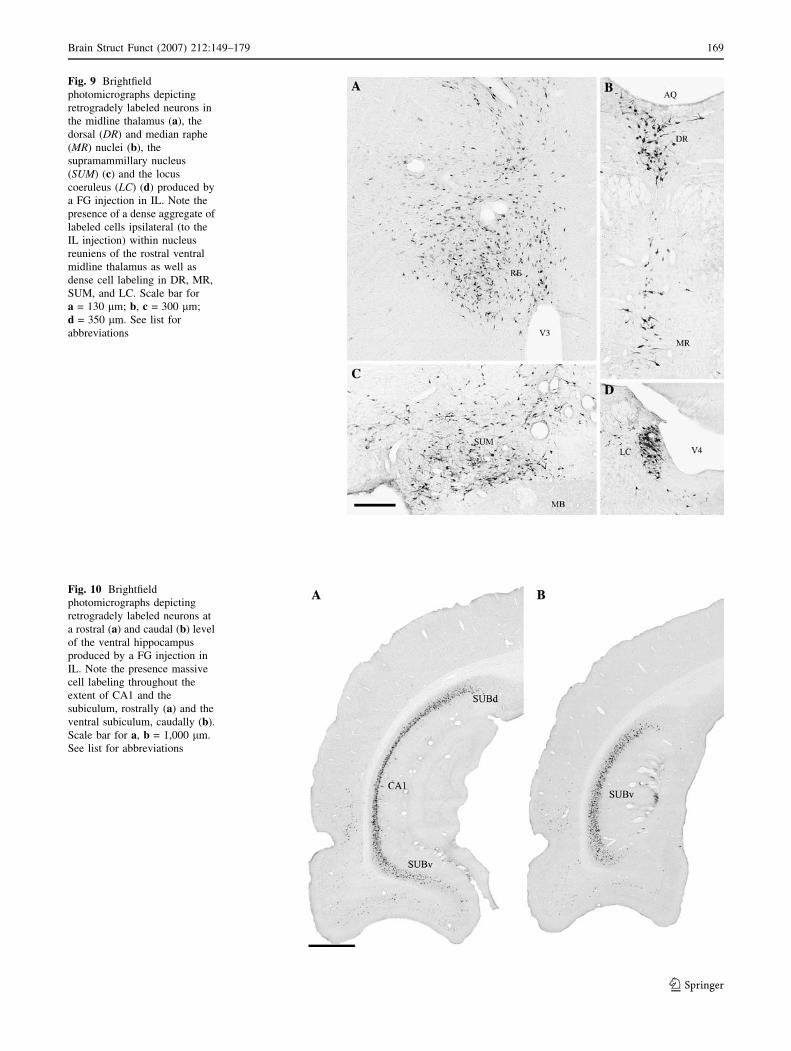
Fig. 10 Brightfield
photomicrographs depicting
retrogradely labeled neurons at
a rostral (a) and caudal (b) level
of the ventral hippocampus
produced by a FG injection in
IL. Note the presence massive
cell labeling throughout the
extent of CA1 and the
subiculum, rostrally (a) and the
ventral subiculum, caudally (b).
Scale bar for a, b = 1,000 lm.
See list for abbreviations
Fig. 9 Brightfield
photomicrographs depicting
retrogradely labeled neurons in
the midline thalamus (a), the
dorsal (DR) and median raphe
(MR) nuclei (b), the
supramammillary nucleus
(SUM) (c) and the locus
coeruleus (LC) (d) produced by
a FG injection in IL. Note the
presence of a dense aggregate of
labeled cells ipsilateral (to the
IL injection) within nucleus
reuniens of the rostral ventral
midline thalamus as well as
dense cell labeling in DR, MR,
SUM, and LC. Scale bar for
a = 130 lm; b, c = 300 lm;
d = 350 lm. See list for
abbreviations
Brain Struct Funct (2007) 212:149–179 169
123

anterograde BF injections (Luiten et al. 1987) that gave rise
to significant labeling of mPFC, particularly within AGm
(see their Fig. 6, p. 240). Cholinergic projections to cortex
reportedly serve important roles in behavioral/EEG arousal
and attentional mechanisms (Woolf 1991; Nunez 1996;
Jimenez-Capdeville et al. 1997; Zaborszky et al. 1999;
Cape et al. 2000; Zaborszky 2002; Jones 2004; Sarter et al.
2005). It worth noting, however, that the ACh region of
BF, contains other types of neurons, prominently including
GABAergic and glutamatergic cells, that project to most of
the cortical sites as ACh neurons (Brashear et al. 1986;
Zaborszky et al. 1986; Gritti et al. 1997, 2003; Zaborszky
2002).
Although earlier reports have described projections from
the (BLA) to mPFC (Kita and Kitai 1990; McDonald 1987,
1991; Bacon et al. 1996; Gabbott et al. 2006), those to
AGm appear to be considerably less pronounced than
shown here. Evidence suggests that BLA to mPFC pro-
jections (particularly to IL/PL) convey information on the
emotional properties of sensory stimuli (Garcia et al. 1999;
LeDoux 2000; Pare et al. 2004; Gabbott et al. 2006; Vertes
2006), involved in executive functions of the mPFC
Non-LimbicCortex
BasalForbrain
Thalamus
Hypothalamus
LimbicCortex
Midbrain Pons/medulla
AGm
FPmAGmSSISSIIOC2RSC
AGlGIAUDPApOC1TE
CLA
DBhSIMA
GPVP
CLMDCMPVRHREVMPT
AMIAMIMDPCVAL
LP
LHyPH
ZISUM
AmygdalaBLA
BMAMEAPATR
VTA
SNcPAGSUMCLi
MRFIP
DRPPTLC
MRNI
RPORPC
MOVOVLOACPLAIdAIpPRC
ILECTEC
CA1LO
Non-LimbicCortex
LimbicCortex
BasalForbrain
Thalamus
Hypothalamus Midbrain Pons/medulla
AC
ACPL
ILMOVOPRCECCA1
AIpECT
CLA
SITTd
BSTDBhLPOMA
AMCLRERHIAMMDPC
CLCMIMDPVPCVM
PT
AmygdalaLHyPAGPH
SUMLM
BLABMA
AGmPApRSCOC2FPmAUDAGlGITESSII
VTA
SNcIPCLi
DR
LCNILDT
MRPB
A
B
heavy labeling
light labeling
moderate labeling
Fig. 11 Summary of main
sources of afferent projections
to AGm (a) and AC (b) from the
cortex (non-limbic and
‘limbic’), basal forebrain,
amygdala, thalamus,
hypothalamus, midbrain, and
pons/medulla. Red, green, and
blue represents heavy,
moderate, and light projections,
respectively, to AGm and AC.
Non-limbic cortex is defined as
motor, somatosensory, special
sensory, and associational
regions of cortex, while ‘limbic’
cortex is defined as remaining
regions of cortex including
orbital cortices and the
hippocampal formation. See list
for abbreviations
170 Brain Struct Funct (2007) 212:149–179
123

(Salinas et al. 1993; Balleine et al. 2003; Pare 2003;
Floresco and Ghods-Sharifi 2007).
Anterior cingulate cortex
Similar to AGm, the main sources of afferents to the
anterior AC were from regions of cortex and the thalamus.
As discussed below, however, cortical input to AC differs
from that to AGm. The primary sources of cortical affer-
ents to AC were from FPm, other regions of mPFC (mainly
from AC and dorsal PL), PAp, RSC, PRC, entorhinal and
secondary visual cortices, as well as CA1/subiculum of HF.
Projections were strongest from AC, PAp, and RSC.
In an examination of cortical afferents to AGm, Reep
et al. (1990) made control injections in AC. In general
accord with present findings, they described afferents to
AC from AGm, various regions of RSC, and from
primary and secondary visual cortices, but failed to
identify them from PAp, parahippocampal cortices (PRC
and EC) and HF. These differences likely involve the
fact that our injections were mainly rostral, theirs caudal,
in AC.
In a recent comprehensive examination of intrinsic AC
connections, Jones et al. (2005) reported that: (1) the rostral
one third of AC is primarily connected with IL, PL and
itself (rostral AC); (2) dorsal and ventral zones of the
Non-LimbicCortex
BasalForbrain
Thalamus
Hypothalamus
LimbicCortex
Midbrain Pons/medullaAmygdala
A
IL
FPmRSC
DBhCLA
TTdEN
MSLSMPOSI
RERHPTPVMD
IAM
IMDCM
SUM
LHyAHPAG
BLABMATR
COALAPA
ILCA1SUB
PLAIpPRCEC
ACECT
VTACLi
PAGSNc
DRLDT
MRLC
NIRPO
Non-LimbicCortex
LimbicCortex
BasalForbrain
Thalamus
Hypothalamus Midbrain Pons/medullaAmygdala
B
PL
FPm
AGm
RSC
CLA
TTdDBhLPO
ENVP
PTPVMDRERH
IMDIAM
CL
SUM
LMLHyPH
BLABMA
TR
PACOA
MOVOACPLILAIdAIvAIpCA1SUBPRC
EC
ECT
VTA
PAGIPSNc
DR
LCMR
heavy labeling
light labeling
moderate labeling
Fig. 12 Summary of main
sources of afferent projections
to PL (a) and IL (b) from the
cortex (non-limbic and
‘limbic’), basal forebrain,
amygdala, thalamus,
hypothalamus, midbrain, and
pons/medulla. Red, green, and
blue represents heavy,
moderate, and light projections,
respectively, to PL and IL. Non-
limbic cortex is defined as
motor, somatosensory, special
sensory, and associational
regions of cortex, while ‘limbic’
cortex is defined as remaining
regions of cortex including
orbital cortices and the
hippocampal formation. See list
for abbreviations
Brain Struct Funct (2007) 212:149–179 171
123

caudal two-thirds of AC are extensively interconnected;
and (3) the caudal RSC projects to the rostral ACm, while
the rostral RSC projects to the caudal AC. The latter
findings are consistent with present and previous results
(van Groen and Wyss 1990a, 1992, 2003; Risold et al.
1997; Shibata et al. 2004) showing that RSC strongly tar-
gets AC. Based on extensive RSC connections with AC,
and additionally with parts of the limbic thalamus, subic-
ulum/postsubiculum, and occipital cortex, van Groen and
Wyss (2003) proposed that RSC is a focal point for the
integration of limbic information. As they noted, RSC is an
essential component of Papez’s circuit (Papez 1937) and
RSC lesions produce marked deficits in spatial navigation
and memory (Sutherland et al. 1988; Cooper and Mizumori
2001; Vann and Aggleton 2002).
The PAp is a large area, bordered rostrally by the
hindlimb sensorimotor cortex, caudally by primary/sec-
ondary visual cortices, medially by RSC and laterally by
SSI (Corwin and Reep 1998; Swanson 1998). Reep and
colleagues (Reep et al. 1990, 1994, 2003; Corwin and Reep
1998; Cheatwood et al. 2005) have described an extended
circuitry involving the medial PAp, AGm, Oc2M, VLO
and the dorsocentral striatum that participates in directed
spatial attention, and when disrupted, produces spatial ne-
glect (Corwin et al. 1986; Crowne et al. 1986; King et al.
1989; King and Corwin 1993; Van Vleet et al. 2003).
In addition to afferents from orbital, mPFC and visual
cortices (and associated thalamic nuclei), PAp receives
input from somatosensory and auditory regions of cortex
(Chandler et al. 1992; Reep et al. 1994). In this respect,
PAp, like RSC, is a mulitmodal integration zone. In accord
with earlier reports (Reep et al. 1990; Corwin and Reep
1998), we described strong PAp to AGm projections, but
also observed equally dense medial PAp projections to AC.
This projection does not seem to have been reported pre-
viously.
As indicated, there is a progressive increase in the
strength of hippocampal and parahippocampal projections
from the dorsal to the ventral mPFC. Similar to AGm,
parahippocampal afferents to AC were shown to primarily
originate from PRC and lateral EC and to a much lesser
degree from the ectorhinal cortex (ECT). This is consistent
with the findings of several earlier reports (Swanson and
Kohler 1986; Insausti et al. 1997; Delatour and Witter
2002). While previous studies have demonstrated projec-
tions from the hippocampus (CA1/subiculum) to the ven-
tral mPFC (IL/PL) (Swanson 1981; Irle and Markowitsch
1982; Ferino et al. 1987; Jay et al. 1989; van Groen and
Wyss 1990b; Jay and Witter 1991; Carr and Sesack 1996),
this is the first report to describe them to the dorsal mPFC:
moderate to AC and light to AGm.
In general accord with previous reports, we showed that
afferents to AC from ‘relay’ nuclei of thalamus arise from
mid-lateral regions of the thalamus. This involves projec-
tions from lateral MD, VAL (lightly), VM, CL, PC, and
anteromedial (AM) nuclei of thalamus. Conde et al. (1990)
described similar findings, but also projections (not seen
here) from PO and LD of thalamus. The conflicting results
could involve the (partial) inclusion of AGm in their AC
injections (Conde et al. 1990, 1995).
In accord with present findings, van Groen et al. (1999)
described massive AM projections to AC, mainly origi-
nating from the ventromedial AM. As has been demon-
strated (Sripanidkulchai and Wyss 1986; Shibata and Kato
1993; van Groen et al. 1999), there are marked differ-
ences in projections from the anterior thalamus to the
anterior and posterior AC (or RSC), such that AM dis-
tributes selectively to AC and AV/AD to RSC. While
earlier reports described (at best) modest projections from
the midline thalamus to AC (Conde et al. 1990, 1995), we
showed that several nuclei of the midline thalamus,
including PV, PT, IAM, CM, RH, and RE, distribute
significantly to AC. RE/RH projections to AC have re-
cently been demonstrated using anterograde tracers
(Vertes et al. 2006).
Other inputs to AC
Similar to AGm, AC receives input from several (non-
thalamic) subcortical sites including the CLA, TTd, MA,
SI, BST of the BF, the BLA/BMA of amygdala, the pos-
terior nucleus (PH) of hypothalamus, the mesencephalic
PAG, SNc and VTA of the midbrain, and monoaminergic
groups of the brainstem. This is consistent with the findings
of several previous reports of diverse subcortical forebrain
(Bigl et al. 1982; Markowitsch et al. 1984; Rye et al. 1984;
Saper 1984; Woolf et al. 1984; Sloniewski et al. 1986;
Luiten et al. 1987; McDonald 1987, 1991; Sherk 1988;
Witter et al. 1988; Woolf 1991; Vertes et al. 1995; Bacon
et al. 1996; Gritti et al. 1997; Kowianski et al. 1998; Majak
et al. 2000; Zhang et al. 2001; Dong and Swanson 2006a, b;
Gabbott et al. 2006) and brainstem inputs to AC (Swanson
1982; Foote et al. 1983; Waterhouse et al. 1983; Vertes and
Martin 1988; Herrero et al. 1991a; Vertes 1991; Cameron
et al. 1995; Morin and Meyer-Bernstein 1999; Vertes et al.
1999; Carr and Sesack 2000a, b; Berridge and Waterhouse
2003).
Regarding VTA and SNc, it is well documented that
VTA is a major source of projections to the mPFC (for
review, Seamans and Yang 2004), but projections from
SNc to mPFC are less well established. In support of
present findings, however, others have demonstrated
moderate SNc projections to the mPFC (Loughlin and
Fallon 1984; Conde et al. 1995) and, like here, have shown
that they predominately originate from medial parts of SNc
and distribute to the dorsal and ventral mPFC.
172 Brain Struct Funct (2007) 212:149–179
123

Prelimbic cortex
Probably the most significant change in the distribution of
afferents to PL (and IL) from those to AGm/AC was a
progressive decline in cortical afferents to the ventral
mPFC. Specifically, there was a marked reduction in
cortical projections, mainly involving sensory (special
sensory and somatosensory), motor or associational re-
gions (RSC and PAp) of cortex, to the ventral as com-
pared to the dorsal mPFC.
With respect to cortical input, however, PL nonetheless
receives projections from several regions of cortex
including FPm, anterior PL, medial (MO) and VO, IL,
dorsal and (rostral) posterior agranular insular, perirhinal
and entorhinal cortices. In addition, the CA1/subiculum
distributes densely to PL—much heavier than to AC or
AGm. These findings are consistent with previous dem-
onstrations of significant orbital and limbic cortical affer-
ents to PL (Reep and Winans 1982; Swanson and Kohler
1986; Jay et al. 1989; Hurley et al. 1991; Jay and Witter
1991; Yasui et al. 1991; Insausti et al. 1997; Shi and
Cassell 1998; Delatour and Witter 2002; Gabbott et al.
2003; Jasmin et al. 2004; Vertes 2004).
The insular cortex (INC) interconnects with the ventral
mPFC (PL and IL) in a topographically organized manner
(Allen et al. 1991; Yasui et al. 1991; Shi and Cassell 1998;
Gabbott et al. 2003; Jasmin et al. 2004, present results). For
instance, we showed that the rostral INC (mainly AId) pri-
marily targets PL and the caudal INC (mainly AIp) targets
IL. Consistent with this, Gabbott et al. (2003) demonstrated
that: (1) AId primarily projects to PL and dysgranular
insular (DI)/AIp mainly to IL; (2) AIv and the GI distribute
lightly to IL/PL; and (3) AId fibers to PL mainly terminate
in layers 2/3 and form asymmetric connections with
dendritic spines of PL cells. In a complementary manner,
there are strong (and selective) return projections from PL to
AId, and from IL to AIp (Sesack et al. 1989; Hurley et al.
1991; Gabbott et al. 2003; Vertes 2004).
Supporting present findings, several reports have dem-
onstrated massive hippocampal projections (CA1/subicu-
lum) to PL (Swanson 1981; Irle and Markowitsch 1982;
Ferino et al. 1987; Jay et al. 1989; van Groen and Wyss
1990b; Jay and Witter 1991; Carr and Sesack 1996). In
contrast, however, with earlier reports (Ferino et al. 1987;
Jay et al. 1989), we showed stronger CA1/subicular pro-
jections to IL than to PL. Interestingly, despite strong HF to
mPFC projections, there are no direct return projections
from the mPFC to the hippocampus (Beckstead 1979;
Room et al. 1985; Sesack et al. 1989; Hurley et al. 1991;
Takagishi and Chiba 1991; Vertes 2004). In the absence of
such projections, we have suggested (Vertes 2002, 2004,
2006; Vertes et al. 2007) that the RE of the midline thal-
amus is an important relay in the transfer of information
from the mPFC to the hippocampus (Wouterlood et al.
1990; Bokor et al. 2002). For instance, we demonstrated
that all divisions of the mPFC distribute heavily to RE
(Vertes 2002; McKenna and Vertes 2004), and RE in turn
is the source of pronounced projections to HF (CA1/su-
biculum) (Vertes et al. 2006, 2007).
Thalamic afferents to PL originate almost entirely from
the midline thalamus and MD. Of the midline groups, the
PT, PV, IAM, CM, rhomboid, and reuniens nuclei dis-
tribute densely to PL. Previous reports using anterograde or
retrograde tracers have described similar findings (Her-
kenham 1978; Ohtake and Yamada 1989; Berendse and
Groenewegen 1991; Conde et al. 1995; Moga et al. 1995;
Risold et al. 1997; Van der Werf et al. 2002; Vertes et al.
2006). We recently demonstrated that RE and RH distrib-
ute throughout the mPFC, terminating heavily in IL/PL,
mainly within layers 1 and 5/6 of these regions (Vertes
et al. 2006). It has been suggested that midline thalamic
input to the mPFC (and other parts of limbic cortex) par-
ticipate in processes of arousal and attention (Van der Werf
et al. 2002) and/or serve to gate the flow of information to
and among limbic forebrain structures (Vertes 2006; Vertes
et al. 2006).
Other inputs to PL
We recently reviewed evidence indicating that IL and PL
of rats serve separate and distinct functions (Vertes 2006).
IL is primarily involved in affective/visceromotor func-
tions, homologous to the orbitomedial PFC of primates,
while PL (and ventral AC) participates in cognitive/limbic
functions homologous to the lateral/dorsolateral cortex of
primates. Associated with its role in cognition, PL dis-
tributes to a relatively small groups of structures that
subserve cognition and have been designated the ‘PL cir-
cuit’ (Alexander et al. 1990; Groenewegen et al. 1990).
They mainly include the insular cortex (AId), the hippo-
campus (via RE), CLA, ACC, BLA, MD, RE, and VTA.
Lesions of each of these structures, like those of PL, pro-
duce deficits in delayed response tasks and memory (Har-
rison and Mair 1996; Seamans et al. 1998; Floresco et al.
1999; Romanides et al. 1999; Kalivas et al. 2001; Barros
et al. 2002; Pare 2003; Roozendaal et al. 2004; Seamans
and Yang 2004; Cain et al. 2006).
As demonstrated here, PL receives input from each of its
major targets including AId, HF, CLA, ACC (via VP and
MD), RE, and VTA. Dopaminergic VTA afferents to PL
(and mPFC) have been extensively examined (Seamans
and Yang 2004) and appear to play a critical role in PL-
associated behaviors. Using a disconnection procedure
wherein the VTA and HF were temporarily disrupted on
opposite sides of the brain, Seamans et al. (1998) showed
that the simultaneous blockade of hippocampal inputs to
Brain Struct Funct (2007) 212:149–179 173
123

PL and dopamine (D1) receptors at PL disrupted perfor-
mance on delayed, but not on non-delayed, versions of the
the radial arm maze (RAM) task. Regarding the possible
role of dopamine in hippocampal-prefrontal interactions,
Seamans et al. (1998) suggested that: ‘D1 receptors in the
PFC may modulate the transfer of spatial information from
the hippocampus to the PFC at a time when a prospective
series of response must be organized and executed.’
Infralimbic cortex
By comparison with other subdivisions of the mPFC, IL
receives considerably fewer inputs from adjacent regions
of the mPFC. Within the mPFC, PL is the main source of
afferents to IL. In a similar manner, cortical inputs to IL are
essentially limited to PL, AId, HF, and parahippocampal
structures. There is a virtual absence of projections from
sensorimotor, special sensory or associational regions of
cortex to IL.
Insular cortical projections to IL primarily originate
from AIp and to a much lesser extent from the dysgranular
INC, dorsal to AIp. Consistent with this, using anterograde
tracers, Shi and Cassell (1998) demonstrated that AIp
selectively targets IL of mPFC. AIp receives convergent
visceral and limbic input (Saper 1982; Ruggiero et al.
1987; Allen et al. 1991), and reportedly represents a major
source of viscerosensory information to the visceromotor
cortex—or IL.
Despite limited (neo/allo) cortical input to IL, the hip-
pocampus (CA1/subiculum) distributes massively to IL. If,
as indicated, IL represents a visceromotor center, hippo-
campal projections to IL may serve to associate past events
(including their affective quality) to present ones for
impending actions. In this regard, a characteristic feature of
bilateral damage to the ventromedial prefrontal cortex in
humans is a pervasive blunted affect (hypoemotionality)
coupled with generally inappropriate and often strongly
negative emotional reactions to relatively minor frustra-
tions (Damasio et al. 1990; Barrash et al. 2000; Berlin et al.
2004; Anderson et al. 2006; Koenigs and Tranel 2007).
Related to the foregoing, Mayberg et al. (2005) recently
demonstrated that deep brain stimulation localized to IL
(presumably suppressing IL activity) produced a marked
remission of depression in human subjects.
Thalamic afferents to IL largely originate from the same
midline thalamic groups that project to PL. They primarily
include medial MD, PV, PT, IAM, RH, and RE. Only
minor differences were observed in projections to IL and
PL. Compared with PL, cells projecting to IL were directly
aligned along the midline (particularly in MD), and CM
distributes much less densely to IL than to PL.
As a putative visceromotor center, IL receives visceral
afferent information from the INC (AIp). There are rela-
tively few additional sources of visceral/limbic input to IL
(or to the mPFC). They include the LS and horizontal
nucleus of the diagonal band (DBh) of the BF, the baso-
lateral (BLA) and TR of amygdala, and the SUM of the
hypothalamus. In contrast, the midline thalamus, particu-
larly the ventral midline thalamus, receives widespread
‘limbic’ afferents from diverse structures of the BF,
amygdala, hypothalamus and brainstem (Cornwall and
Phillipson 1988; Chen and Su 1990; Risold et al. 1997;
Krout et al. 2002; Van der Werf et al. 2002; McKenna and
Vertes 2004). These findings, coupled with the demon-
stration that the midline thalamus projects heavily to IL
(and to PL), suggest that midline thalamus is a primary
route by which limbic information reaches the mPFC
(Vertes 2006).
Other inputs to IL
Although IL receives fewer overall inputs than do other
divisions of mPFC, it is nonetheless the target of some
subcortical limbic structures that do not project elsewhere
in the mPFC—or certainly to the same degree as to IL.
These include the LS, DBh, BLA, TR, LHy, SUM, VTA,
PAG, DR, MR, NI, and the LDT. This is consistent with
previous demonstrations of relatively substantial projec-
tions from these sites to IL (Swanson 1982; Rye et al. 1984;
McDonald 1987, 1991; Bacon et al. 1996; Herrero et al.
1991a, b; Vertes 1991, 1992; Cameron et al. 1995; Morin
and Meyer-Bernstein 1999; Vertes et al. 1999; Carr and
Sesack 2000a, b; Goto et al. 2001; Olucha-Bordonau et al.
2003; Gabbott et al. 2006). Regarding LDT, it is well
recognized that LDT is a major source of cholinergic
afferents to the thalamus and parts of the BF (Hallanger
and Wainer 1988), but direct LDT projections to the cortex
(or to mPFC) are not well documented. In line, however,
with the present demonstration of moderate LDT projec-
tions (and some PPT projections) to the mPFC, previous
reports using various tracers, have similarly identified a
relatively prominent LDT input to the mPFC (Satoh and
Fibiger 1986; Cornwall et al. 1990; Herrero et al. 1991b).
As shown here, they mainly target the ventral mPFC or IL.
These additional afferents undoubtedly supplement those
from AIp and the midline thalamus to IL in relaying limbic
afferent information to IL in visceromotor control.
General summary: an integrative role for the mPFC
in goal directed behavior
Each of the subdivisions of the mPFC receives a fairly
unique set of afferent projections. There is a shift dorso-
ventrally along the mPFC from predominantly sensorimo-
tor (non-limbic) cortical and thalamic input to dorsal
174 Brain Struct Funct (2007) 212:149–179
123

mPFC, to limbic cortical and thalamic (midline thalamus)
input to the ventral mPFC. Each division of mPFC strongly
communicates with immediately adjacent regions, and with
the possible exception of IL, each division interconnects
with all others. The hippocampus (CA1/subiculum) pro-
jects heavily to IL and PL, and considerably less so to
dorsal regions of the mPFC. Sites projecting commonly to
the four divisions of mPFC include INC, CLA, BLA, and
TR of the amygdala, parts of the midline thalamus, SUM,
VTA, PAG, DR, MR, and LC of the brainstem.
The AGm (and dorsal AC) receives a vast array of
information both directly and indirectly from all sensory
modalities and presumably utilizes this information in sit-
uations demanding immediate attention for appropriate
actions. As discussed, Reep et al. (1990) view AGm as a
multisensory integration region. Stimulation of AGm (and
dorsal AC) produces movements (and generally coordi-
nated movements) of the head, eyes and vibrissa, having
the characteristics of orienting responses. Accordingly,
unilateral AGm lesions disrupt orienting movements and
produce contralateral neglect to visual, auditory and
somatosensory stimuli (Corwin et al. 1986). AGm it is
thought to be homologous to the premotor, supplementary
motor and frontal eye fields of primates (Vertes 2006).
As described, there is a dramatic shift in sources of
afferent information from the AGm/dorsal AC to PL (and
ventral AC), from multisensory afferents dorsally, to a
combination of sensory and limbic input (subcortical/cor-
tical) ventrally. PL is strategically positioned to integrate
information across modalities and compare present and
past events for appropriate actions. In this regard, cells of
PL (and ventral AC) respond selectively during the delay
period of delay response tasks, and PL lesions produce
marked deficits in delayed responses tasks involving short
and long delay—as do lesions of major PL targets (or the
PL circuit) (Vertes 2006). PL (and ventral AC) are thought
to be homologous to the lateral/dorsolateral PFC of pri-
mates (Vertes 2006).
Unlike AGm/AC (and PL), afferents to IL almost en-
tirely originate from limbic subcortical and cortical sites.
Visceral afferent information primarily reaches IL via AIp,
BLA and the midline thalamus, and together undoubtedly
represent important sources of limbic input to IL in vis-
ceromotor control. As discussed, IL profoundly influences
visceral/autonomic activity (see Introduction). IL is
thought to be homologous to the orbitomedial PFC of
primates (Vertes 2006).
The mPFC of rats, like the prefrontal cortex of primates,
would appear to be directly involved in higher order cog-
nitive functioning, and through interconnections among the
four divisions, would be capable of exerting control over
all aspects, including affective components, of goal direc-
ted behavior.
References
Alexander GE, Crutcher MD, DeLong MR (1990) Basal ganglia-
thalamocortical circuits: parallel substrates for motor, oculomo-
tor, ‘‘prefrontal’’ and ‘‘limbic’’ functions. Prog Brain Res
85:119–146
Allen GV, Saper CB, Hurley KM, Cechetto DF (1991) Organization
of visceral and limbic connections in the insular cortex of the rat.
J Comp Neurol 311:1–16
Anderson SW, Barrash J, Bechara A, Tranel D (2006) Impairments of
emotion and real-world complex behavior following childhood-
or adult-onset damage to ventromedial prefrontal cortex. J Int
Neuropsychol Soc 12:224–235
Bacon SJ, Headlam AJN, Gabbott PLA, Smith AD (1996) Amygdala
input to medial prefrontal cortex (mPFC) in the rat: a light and
electron microscope study. Brain Res 720:211–219
Balleine BW, Killcross AS, Dickinson A (2003) The effect of lesions
of the basolateral amygdala on instrumental conditioning. J
Neurosci 23:666–675
Barbas H (1995) Anatomical basis of cognitive-emotional interactions
in the primate prefrontal cortex. Neurosci Biobehav Rev 19:499–
510
Barbas H (2000) Connections underlying the synthesis of cognition,
memory, and emotion in primate prefrontal cortices. Brain Res
Bull 52:319–330
Barrash J, Tranel D, Anderson SW (2000) Acquired personality
disturbances associated with bilateral damage to the ventrome-
dial prefrontal region. Dev Neuropsychol 18:355–381
Barros DM, Pereira P, Medina JH, Izquierdo I (2002) Modulation of
working memory and of long- but not short-term memory by
cholinergic mechanisms in the basolateral amygdala. Behav
Pharmacol 13:163–167
Beckstead RM (1979) Autoradiographic examination of corticocor-
tical and subcortical projections of the mediodorsal-projection
(prefrontal) cortex in the rat. J Comp Neurol 184:43–62
Berendse HW, Groenewegen HJ (1991) Restricted cortical termina-
tion fields of the midline and intralaminar thalamic nuclei in the
rat. Neuroscience 42:73–102
Berlin HA, Rolls ET, Kischka U (2004) Impulsivity, time perception,
emotion and reinforcement sensitivity in patients with orbito-
frontal cortex lesions. Brain 127:1108–1126
Berridge CW, Waterhouse BD (2003) The locus coeruleus-noradren-
ergic system: modulation of behavioral state and state-dependent
cognitive processes. Brain Res Rev 42:33–84
Bigl V, Woolf NJ, Butcher LL (1982) Cholinergic projections from
the basal forebrain to frontal, parietal, temporal, occipital, and
cingulate cortices: a combined fluorescent tracer and acetylcho-
linesterase analysis. Brain Res Bull 8:727–749
Bokor H, Csaki A, Kocsis K, Kiss J (2002) Cellular architecture of the
nucleus reuniens thalami and its putative aspartatergic/glutama-
tergic projection to the hippocampus and medial septum in the
rat. Eur J Neurosci 16:1227–1239
Brashear HR, Zaborszky L, Heimer L (1986) Distribution of
GABAergic and cholinergic neurons in the rat diagonal band.
Neuroscience 17:439–451
Brito GNO, Brito LSO (1990) Septohippocampal system and the
prelimbic sector of frontal cortex: a neuropsychological battery
analysis in the rat. Behav Brain Res 36:127–146
Buchanan SL, Thompson RH, Maxwell BL, Powell DL (1994)
Efferent connections of the prefrontal cortex in the rabbit. Exp
Brain Res 100:469–483
Burns SM, Wyss JM (1985) The involvement of the anterior cingulate
cortex in blood pressure control. Brain Res 370:71–77
Cain DP, Boon F, Corcoran ME (2006) Thalamic and hippocampal
mechanisms in spatial navigation: a dissociation between brain
Brain Struct Funct (2007) 212:149–179 175
123

mechanisms for learning how versus learning where to navigate.
Behav Brain Res 170:241–256
Cameron AA, Khan IA, Westlund KN, Cliffer KD, Willis WD (1995)
The efferent projections of the periaqueductal gray in the rat: a
Phaseolus vulgaris-leucoagglutinin study. I. Ascending projec-
tions. J Comp Neurol 351:568–584
Cape EG, Manns ID, Alonso A, Beaudet A, Jones BE (2000)
Neurotensin-induced bursting of cholinergic basal forebrain
neurons promotes gamma and theta cortical activity together
with waking and paradoxical sleep. J Neurosci 20:8452–8461
Carr DB, Sesack SR (1996) Hippocampal afferents to the rat
prefrontal cortex: synaptic targets and relation to dopamine
terminals. J Comp Neurol 369:1–15
Carr DB, Sesack SR (2000a) GABA-containing neurons in the rat
ventral tegmental area project to the prefrontal cortex. Synapse
38:114–123
Carr DB, Sesack SR (2000b) Dopamine terminals synapse on callosal
projection neurons in the rat prefrontal cortex. J Comp Neurol
425:275–283
Chandler HC, King V, Corwin JV, Reep RL (1992) Thalamocortical
connections of rat posterior parietal cortex. Neurosci Lett
143:237–242
Cheatwood JL, Reep RL, Corwin JV (2003) The associative striatum:
cortical and thalamic projections to the dorsocentral striatum in
rats. Brain Res 968:1–14
Cheatwood JL, Corwin JV, Reep RL (2005) Overlap and interdig-
itation of cortical and thalamic afferents to dorsocentral striatum
in the rat. Brain Res 1036:90–100
Chen S, Su HS (1990) Afferent connections of the thalamic
paraventricular and parataenial nuclei in the rat—a retrograde
tracing study with iontophoretic application of Fluoro-Gold.
Brain Res 522:1–6
Chiba T, Kayahara T, Nakanoh K (2001) Efferent projections of
infralimbic and prelimbic areas of the medial prefrontal cortex
in the Japanese monkey, Macaca fuscata. Brain Res 888:83–
101
Conde F, Audinat E, Maire-Lepoivre E, Crepel F (1990) Afferent
connections of the medial frontal cortex of the rat. A study using
retrograde transport of fluorescent dyes. I. Thalamic afferents.
Brain Res Bull 24:341–354
Conde F, Maire-Lepoivre E, Audinat E, Crepel F (1995) Afferent
connections of the medial frontal cortex of the rat. II. Cortical
and subcortical afferents. J Comp Neurol 352:567–593
Cooper BG, Mizumori SJ (2001) Temporary inactivation of the
retrosplenial cortex causes a transient reorganization of spatial
coding in the hippocampus. J Neurosci 21:3986–4001
Cornwall J, Phillipson OT (1988) Afferent projections to the dorsal
thalamus of the rat as shown by retrograde lectin transport. II.
The midline nuclei. Brain Res Bull 21:147–161
Cornwall J, Cooper JD, Phillipson OT (1990) Afferent and efferent
connections of the laterodorsal tegmental nucleus of the rat.
Brain Res Bull 25:271–284
Corwin JV, Reep RL (1998) Rodent posterior parietal cortex as a
component of a cortical network mediating directed spatial
attention. Psychobiology 26:87–102
Corwin JV, Kanter S, Watson RT, Heilman KM, Valenstein E,
Hashimoto A (1986) Apomorphine has a therapeutic effect on
neglect produced by unilateral dorsomedial prefrontal cortex
lesions in rats. Exp Neurol 94:683–698
Crowne DP, Pathria MN (1982) Some attentional effects of unilateral
frontal lesions in the rat. Behav Brain Res 6:25–39
Crowne DP, Richardson CM, Dawson KA (1986) Parietal and frontal
eye field neglect in the rat. Behav Brain Res 22:227–231
Damasio AR, Tranel D, Damasio H (1990) Individuals with
sociopathic behavior caused by frontal damage fail to respond
autonomically to social stimuli. Behav Brain Res 41:81–94
Delatour B, Gisquet-Verrier P (1996) Prelimbic cortex specific
lesions disrupt delayed-variable response tasks in the rat. Behav
Neurosci 110:1282–1298
Delatour B, Gasket-Verrier P (1999) Lesions of the prelimbic-
infralimbic cortices in rats do not disrupt response selection
processes but induce delay-dependent deficits: evidence for a
role in working memory? Behav Neurosci 113:941–955
Delatour B, Gisquet-Verrier P (2000) Functional role of rat prelimbic-
infralimbic cortices in spatial memory: evidence for their
involvement in attention and behavioral flexibility. Behav Brain
Res 109:113–128
Delatour B, Witter MP (2002) Projections from the parahippocampal
region to the prefrontal cortex in the rat: evidence of multiple
pathways. Eur J Neurosci 15:1400–1407
Dong HW, Swanson LW (2006a) Projections from bed nuclei of the
stria terminalis, magnocellular nucleus: implications for cerebral
hemisphere regulation of micturition, defecation, and penile
erection. J Comp Neurol 494:108–141
Dong HW, Swanson LW (2006b) Projections from bed nuclei of the
stria terminalis, anteromedial area: cerebral hemisphere integra-
tion of neuroendocrine, autonomic, and behavioral aspects of
energy balance. J Comp Neurol 494:142–178
Edelstein LR, Denaro FJ (2004) The claustrum: a historical review of
its anatomy, physiology, cytochemistry and functional signifi-
cance. Cell Mol Biol 50:675–702
Ferino F, Thierry AM, Glowinski J (1987) Anatomical and electro-
physiological evidence for a direct projection from Ammon’s
horn to the medial prefrontal cortex in the rat. Exp Brain Res
65:421–426
Floresco SB, Seamans JK, Phillips AG (1997) Selective roles for
hippocampal, prefrontal cortical, and ventral striatal circuits in
radial-arm maze tasks with or without a delay. J Neurosci
17:1880–1890
Floresco SB, Braaksma DN, Phillips AG (1999) Thalamic-cortical-
striatal circuitry subserves working memory during delayed
responding on a radial arm maze. J Neurosci 19:11061–11071
Floresco SB, Ghods-Sharifi S (2007) Amygdala-prefrontal cortical
circuitry regulates effort-based decision making. Cereb Cortex
17:251–260
Foote SL, Bloom FE, Aston-Jones G (1983) Nucleus locus coeruleus:
new evidence of anatomical and physiological specificity.
Physiol Rev 63:844–914
Fuster JM (1989) The prefrontal cortex. Anatomy, physiology and
neuropsychology of the frontal lobe, 2nd edn. Raven Press, New
York
Gabbott PLA, Warner TA, Jays PRL, Bacon SJ (2003) Areal and
synaptic interconnectivity of paralimbic (area 32), infralimbic
(area 25) and insular cortices in the rat. Brain Res 993:59–71
Gabbott PLA, Warner TA, Jays PRL, Salway P, Busby SJ (2005)
Prefrontal cortex in the rat: projections to subcortical autonomic,
motor, and limbic centers. J Comp Neurol 492:145–177
Gabbott PLA, Warner TA, Busby SJ (2006) Amygdala input
monosynaptically innervates parvalbumin immunoreactive local
circuit neurons in rat medial prefrontal cortex. Neuroscience
139:1039–1048
Garcia R, Vouimba RM, Baudry M, Thompson RF (1999) The
amygdala modulates prefrontal cortex activity relative to con-
ditioned fear. Nature 402:294–296
Goldman-Rakic PS (1994) The issue of memory in the study of
prefrontal function. In: Thierry AM, Glowinsky J, Goldman-
Rakic PS, Christen Y (eds) Motor and cognitive functions of the
prefrontal cortex. Springer, Berlin, pp 112–123
Goto M, Swanson LW, Canteras NS (2001) Connections of the
nucleus incertus. J Comp Neurol 438:86–122
Gritti I, Mainville L, Mancia M, Jones BE (1997) GABAergic and
other noncholinergic basal forebrain neurons, together with
176 Brain Struct Funct (2007) 212:149–179
123

cholinergic neurons, project to the mesocortex and isocortex in
the rat. J Comp Neurol 383:163–177
Gritti I, Manns ID, Mainville L, Jones BE (2003) Parvalbumin,
calbindin, or calretinin in cortically projecting and GABAergic,
cholinergic, or glutamatergic basal forebrain neurons of the rat. J
Comp Neurol 458:11–31
Groenewegen HJ (1988) Organization of the afferent connections of
the mediodorsal thalamic nucleus in the rat, related to the
mediodorsal prefrontal topography. Neuroscience 24:379–431
Groenewegen HJ, Uylings HBM (2000) The prefrontal cortex and the
integration of sensory, limbic and autonomic information. Prog
Brain Res 126:3–28
Groenewegen HJ, Berendse HW, Wolters JG, Lohman AHM (1990)
The anatomical relationship of the prefrontal cortex with the
striatopallidal system, the thalamus and the amygdala: evidence
for a parallel organization. Prog Brain Res 85:95–118
Guandalini P (1998) The corticocortical projections of the physio-
logically defined eye field in the rat medial frontal cortex. Brain
Res Bull 47:377–385
Hallanger AE, Wainer BH (1988) Ascending projections from the
pedunculopontine tegmental nucleus and the adjacent mesopon-
tine tegmentum in the rat. J Comp Neurol 274:483–515
Hardy SGP, Holmes DE (1988) Prefrontal stimulus-produced hypo-
tension in rat. Exp Brain Res 73:249–255
Harrison LM, Mair RG (1996) A comparison of the effects of frontal
cortical and thalamic lesions on measures of spatial learning and
memory in the rat. Behav Brain Res 75:195–206
Heidbreder CA, Groenewegen HJ (2003) The medial prefrontal
cortex in the rat: evidence for a dorso-ventral distinction based
upon functional and anatomical characteristics. Neurosci Biobe-
hav Rev 27:555–579
Herkenham M (1978) The connections of the nucleus reuniens
thalami: evidence for a direct thalamo-hippocampal pathway in
the rat. J Comp Neurol 177:589–610
Herkenham M (1979) The afferent and efferent connections of the
ventromedial thalamic nucleus in the rat. J Comp Neurol
183:487–517
Herrero MT, Insausti R, Gonzalo LM (1991a) Cortically projecting
cells in the periaqueductal gray matter of the rat. A retrograde
fluorescent tracer study. Brain Res 543:201–212
Herrero MT, Insausti R, Gonzalo LM (1991b) Cortical projections
from the laterodorsal and dorsal tegmental nuclei. A fluorescent
retrograde tracing study in the rat. Neurosci Lett 123:144–147
Hicks RR, Huerta MF (1991) Differential thalamic connectivity of
rostral and caudal parts of cortical area Fr2 in rats. Brain Res
568:325–329
Hur EE, Zaborszky L (2005) Vglut2 afferents to the medial prefrontal
and primary somatosensory cortices: a combined retrograde
tracing in situ hybridization study. J Comp Neurol 483:351–373
Hurley KM, Herbert H, Moga MM, Saper CB (1991) Efferent
projections of the infralimbic cortex of the rat. J Comp Neurol
308:249–276
Hurley-Gius KM, Neafsey EJ (1986) The medial frontal cortex and
gastric motility: microstimulation results and their possible
significance for the overall pattern of organization of rat frontal
and parietal cortex. Brain Res 365:241–248
Insausti R, Herrero MT, Witter MP (1997) Entorhinal cortex of the
rat: cytoarchitectonic subdivisions and the origin and distribution
of cortical efferents. Hippocampus 7:146–183
Irle E, Markowitsch HJ (1982) Connections of the hippocampal
formation, mamillary bodies, anterior thalamus and cingulate
cortex. A retrograde study using horseradish peroxidase in the
cat. Exp Brain Res 47:79–94
Jasmin L, Granato A, Ohara PT (2004) Rostral agranular insular
cortex and pain areas of the central nervous system: a tract-
tracing study in the rat. J Comp Neurol 468:425–440
Jay TM, Witter MP (1991) Distribution of hippocampal CA1 and
subicular efferents in the prefrontal cortex of the rat studied by
means of anterograde transport of Phaseolus vulgaris-leucoag-
glutinin. J Comp Neurol 313:574–586
Jay TM, Glowinski J, Thierry AM (1989) Selectivity of the
hippocampal projection to the paralimbic area of the prefrontal
cortex in the rat. Brain Res 505:337–340
Jimenez-Capdeville ME, Dykes RW, Myasnikov AA (1997) Differ-
ential control of cortical activity by the basal forebrain in rats: a
role for both cholinergic and inhibitory influences. J Comp
Neurol 381:53–67
Jones BE (2004) Activity, modulation and role of basal forebrain
cholinergic neurons innervating the cerebral cortex. Prog Brain
Res 145:157–169
Jones BF, Groenewegen HJ, Witter MP (2005) Intrinsic connections
of the cingulate cortex in the rat suggest the existence of multiple
functionally segregated networks. Neuroscience 133:193–207
Kalivas PW, Jackson D, Romanidies A, Wyndham L, Duffy P (2001)
Involvement of pallidothalamic circuitry in working memory.
Neuroscience 104:129–136
King VR, Corwin JV (1993) Comparisons of hemi-inattention
produced by unilateral lesions of the posterior parietal cortex
or medial agranular prefrontal cortex in rats: neglect, extinction,
and the role of stimulus distance. Behav Brain Res 54:117–131
King V, Corwin JV, Reep RL (1989) Production and characterization
of neglect in rats with unilateral lesions of ventrolateral orbital
cortex. Exp Neurol 105:287–299
Kita H, Kitai ST (1990) Amygdaloid projections to the frontal cortex
and the striatum in the rat. J Comp Neurol 298:40–49
Koenigs M, Tranel D (2007) Irrational economic decision-making
after ventromedial prefrontal damage: evidence from the Ulti-
matum Game. J Neurosci 27:951–956
Kolb B (1984) Functions of the frontal cortex of the rat: a
comparative review. Brain Res Rev 8:65–98
Kolb B (1990) Animal models for human PFC-related disorders. Prog
Brain Res 85:501–519
Kowianski P, Morys J, Karwacki Z, Dziewiatkowski J, Narkiewicz O
(1998) The cortico-related zones of the rabbit claustrum: study
of the claustrocortical connections based on the retrograde
axonal transport of fluorescent tracers. Brain Res 784:199–209
Krettek JE, Price JL (1977) The cortical projections of the medio-
dorsal nucleus and adjacent thalamic nuclei in the rat. J Comp
Neurol 171:157–191
Krout KE, Belzer RE, Loewy AD (2002) Brainstem projections to
midline and intralaminar thalamic nuclei of the rat. J Comp
Neurol 448:53–101
LeDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci
23:55–184
Leonard CM (1969) The prefrontal cortex of the rat. I. Cortical
projection of the mediodorsal nucleus. II. Efferent connections.
Brain Res 12:321–343
Loughlin SE, Fallon JH (1984) Substantia nigra and ventral tegmental
area projections to cortex: topography and collateralization.Neuroscience 11:425–435
Luiten PGM, Gaykema RPA, Traber J, Spencer DG Jr (1987) Cortical
projection patterns of magnocellular basal nucleus subdivisions
as revealed by anterogradely transported Phaseolus vulgaris
leucoagglutinin. Brain Res 413:229–250
Majak K, Kowianski P, Morys J, Spodnik J, Karwacki Z, Wisniewski
HM (2000) The limbic zone of the rabbit and rat claustrum: a study
of the claustrocingulate connections based on the retrograde axonal
transport of fluorescent tracers. Anat Embryol 201:15–25
Markowitsch HJ, Irle E, Bangolsen R, Flindtegebak P (1984)
Claustral efferents to the cats limbic cortex studied with
retrograde and anterograde tracing techniques. Neuroscience
12:409–425
Brain Struct Funct (2007) 212:149–179 177
123

Mayberg HS, Lozano AM, Voon V, McNeely HE, Seminowicz D,
Hamani C, Schwalb JM, Kennedy SH (2005) Deep brain
stimulation for treatment-resistant depression. Neuron 45:651–660
McDonald AJ (1987) Organization of amygdaloid projections to the
mediodorsal thalamus and prefrontal cortex: a fluorescence
retrograde transport study in the rat. J Comp Neurol 262:46–58
McDonald AJ (1991) Organization of amygdaloid projections to the
prefrontal cortex and associated striatum in the rat. Neuroscience
44:1–14
McKenna JT, Vertes RP (2004) Afferent projections to nucleus
reuniens of the thalamus. J Comp Neurol 480:115–142
Moga MM, Weis RP, Moore RY (1995) Efferent projections of the
paraventricular thalamic nucleus in the rat. J Comp Neurol
359:221–238
Morin LP, Meyer-Bernstein EL (1999) The ascending serotonergic
system in the hamster: comparison with projections of the dorsal
and median raphe nuclei. Neuroscience 91:81–105
Neafsey EJ (1990) Prefrontal cortical control of the autonomic
nervous system: anatomical and physiological observations. Prog
Brain Res 85:147–166
Nunez A (1996) Unit activity of rat basal forebrain neurons:
relationship to cortical activity. Neuroscience 72:757–766
Ohtake T, Yamada H (1989) Efferent connections of the nucleus
reuniens and the rhomboid nucleus in the rat: an anterograde
PHA-L tracing study. Neurosci Res 6:556–568
Olucha-Bordonau FE, Teruel V, Barcia-Gonzalez J, Ruiz-Torner A,
Valverde-Navarro AA, Martinez-Soriano F (2003) Cytoarchi-
tecture and efferent projections of the nucleus incertus of the rat.
J Comp Neurol 464:62–97
Ongur D, Price JL (2000) The organization of networks within the
orbital and medial prefrontal cortex of rats, monkeys and
humans. Cereb Cortex 10:206–219
Papez JW (1937) A proposed mechanism of emotion. Arch Neurol
Psychiatr 38:725–743
Pare D (2003) Role of the basolateral amygdala in memory
consolidation. Prog Neurobiol 70:409–420
Pare D, Quirk GJ, LeDoux JE (2004) New vistas on amygdala
networks in conditioned fear. J Neurophysiol 92:1–9
Petrides M (1998) Specialized systems for the processing of
mnemonic information within the primate frontal cortex. In:
Roberts AC, Robbins TW, Weiskrantz L (eds) The prefrontal
cortex: executive and cognitive functions. Oxford University
Press, New York, pp 103–116
Ragozzino ME, Adams S, Kesner RP (1998) Differential involvement
of the dorsal anterior cingulate and prelimbic-infralimbic areas
of the rodent prefrontal cortex in spatial working memory. Behav
Neurosci 112:293–303
Ray JP, Price JL (1992) The organization of the thalamocortical
connections of the mediodorsal thalamic nucleus in the rat,
related to the ventral forebrain prefrontal cortex topography. J
Comp Neurol 323:167–197
Reep RL, Winans SS (1982) Efferent connections of dorsal and
ventral agranular insular cortex in the hamster, Mesocricetus
auratus. Neuroscience 7:2609–2635
Reep RL, Corwin JV (1999) Topographic organization of the striatal
and thalamic connections of rat medial agranular cortex. Brain
Res 841:43–52
Reep RL, Corwin JV, Hashimoto A, Watson RT (1984) Afferent
connections of medial precentral cortex in the rat. Neurosci Lett
44:247–252
Reep RL, Goodwin GS, Corwin JV (1990) Topographic organization
in the corticocortical connections of medial agranular cortex in
rats. J Comp Neurol 294:262–280
Reep RL, Chandler HC, King V, Corwin JV (1994) Rat posterior
parietal cortex: topography of corticocortical and thalamic
connections. Exp Brain Res 100:67–84
Reep RL, Cheatwood JL, Corwin JV (2003) The associative striatum:
organization of cortical projections to the dorsocentral striatum
in rats. J Comp Neurol 467:271–292
Repovs G, Baddeley A (2006) The multi-component model of
working memory: explorations in experimental cognitive psy-
chology. Neuroscience 139:5–21
Risold PY, Thompson RH, Swanson LW (1997) The structural
organization of connections between hypothalamus and cerebral
cortex. Brain Res Rev 24:197–254
Romanides AJ, Duffy P, Kalivas PW (1999) Glutamatergic and
dopaminergic afferents to the prefrontal cortex regulate spatial
working memory in rats. Neuroscience 92:97–106
Room P, Russchen FT, Groenewegen HJ, Lohman AHM (1985)
Efferent connections of the paralimbic (area 32) and the
infralimbic (area 25) cortices: an anterograde tracing study in
the cat. J Comp Neurol 242:40–55
Roozendaal B, McReynolds JR, McGaugh JL (2004) The basolateral
amygdala interacts with the medial prefrontal cortex in regulat-
ing glucocorticoid effects on working memory impairment. J
Neurosci 24:1385–1392
Ruggiero DA, Mraovitch S, Granata AR, Anwar M, Reis DJ (1987) A
role of insular cortex in cardiovascular function. J Comp Neurol
257:189–207
Rye DB, Wainer BH, Mesulam MM, Mufson EJ, Saper CB (1984)
Cortical projections arising from the basal forebrain: a study of
cholinergic and noncholinergic components employing com-
bined retrograde tracing and immunohistochemical localization
of choline acetyltransferase. Neuroscience 13:627–643
Salinas JA, Packard MG, McGaugh JL (1993) Amygdala modulates
memory for changes in reward magnitude: reversal post-training
inactivation with lidocaine attenuates the response to a reduction
in reward. Behav Brain Res 59:153–159
Saper CB (1982) Convergence of autonomic and limbic connections
in the insular cortex of the rat. J Comp Neurol 210:163–173
Saper CB (1984) Organization of cerebral cortical afferent systems in
the rat. I. Magnocellular basal nucleus. J Comp Neurol 222:313–
342
Sarter M, Hasselmo ME, Bruno JP, Givens B (2005) Unraveling the
attentional functions of cortical cholinergic inputs: interactions
between signal-driven and cognitive modulation of signal
detection. Brain Res Rev 48:98–111
Satoh K, Fibiger HC (1986) Cholinergic neurons of the laterodorsal
tegmental nucleus: efferent and afferent connections. J Comp
Neurol 253:277–302
Seamans JK, Yang CR (2004) The principal features and mechanisms
of dopamine modulation in the prefrontal cortex. Prog Neurobiol
74:1–58
Seamans JK, Floresco SB, Phillips AG (1995) Functional differences
between the paralimbic and anterior cingulate regions of the rat
prefrontal cortex. Behav Neurosci 109:1063–1073
Seamans JK, Floresco SB, Phillips AG (1998) D1 receptor modula-
tion of hippocampal-prefrontal cortical circuits integrating
spatial memory with executive functions in the rat. J Neurosci
18:1613–1621
Sesack SR, Deutch AY, Roth RH, Bunney BS (1989) Topographical
organization of the efferent projections of the medial prefrontal
cortex in the rat: an anterograde tract-tracing study with
Phaseolus vulgaris leucoagglutinin. J Comp Neurol 290:213–
242
Sherk H (1988) The claustrum and the cerebral cortex. In: Jones EG,
Peters A (eds) Cerebral cortex, vol 5. Sensory-motor areas and
aspects of cortical connectivity. Plenum Press, New York, pp
467–499
Shi CJ, Cassell MD (1998) Cortical, thalamic, and amygdaloid
connections of the anterior and posterior insular cortices. J Comp
Neurol 399:440–468
178 Brain Struct Funct (2007) 212:149–179
123

Shibata H, Kato A (1993) Topographic relationship between antero-
medial thalamic nucleus neurons and their cortical terminal
fields in the rat. Neurosci Res 17:63–69
Shibata H, Kondo S, Naito J (2004) Organization of retrosplenial
cortical projections to the anterior cingulate, motor, and
prefrontal cortices in the rat. Neurosci Res 49:1–11
Sloniewski P, Usunoff KG, Pilgrim C (1986) Retrograde transport of
fluorescent tracers reveals extensive ipsilateral and contralateral
claustrocortical connections in the rat. J Comp Neurol 246:467–477
Sripanidkulchai K, Wyss JM (1986) Thalamic projections to retro-
splenial cortex in the rat. J Comp Neurol 254:143–165
Sutherland RJ, Whishaw IQ, Kolb B (1988) Contributions of
cingulate cortex to two forms of spatial learning and memory.
J Neurosci 8:1863–1872
Swanson LW (1982) The projections of the ventral tegmental area and
adjacent regions: a combined fluorescent retrograde tracer and
immunofluorescence study in the rat. Brain Res Bull 9:321–353
Swanson LW (1981) A direct projection from Ammon’s horn to
prefrontal cortex in the rat. Brain Res 217:150–154
Swanson LW (1998) Brain maps: structure of the rat brain. Elsevier,
New York
Swanson LW, Kohler C (1986) Anatomical evidence for direct
projections from the entorhinal area to the entire cortical mantle
in the rat. J Neurosci 6:3010–3023
Takagishi M, Chiba T (1991) Efferent projections of the infralimbic
(area 25) region of the medial prefrontal cortex in the rat: an
anterograde tracer PHA-L study. Brain Res 566:26–39
Terreberry RR, Neafsey EJ (1983) Rat medial frontal cortex: a
visceral motor region with a direct projection to the solitary
nucleus. Brain Res 278:245–249
Uylings HBM, van Eden CG (1990) Qualitative and quantitative
comparison of the prefrontal cortex in rat and in primates,
including humans. Prog Brain Res 85:31–62
Van der Werf YD, Witter MP, Groenewegen HJ (2002) The
intralaminar and midline nuclei of the thalamus. Anatomical
and functional evidence for participation in processes of arousal
and awareness. Brain Res Rev 39:107–140
van Eden CG, Lamme VA, Uylings HB (1992) Heterotopic cortical
afferents to the medial prefrontal cortex in the rat. A combined
retrograde and anterograde tracer study. Eur J Neurosci 4:77–97
van Groen T, Wyss JM (1990a) Connections of the retrosplenial
granular a cortex in the rat. J Comp Neurol 300:593–606
van Groen T, Wyss JM (1990b) The connections of presubiculum and
parasubiculum in the rat. Brain Res 518:227–243
van Groen T, Wyss JM (1992) Connections of the retrosplenial
dysgranular cortex in the rat. J Comp Neurol 315:200–216
van Groen T, Wyss JM (2003) Connections of the retrosplenial
granular b cortex in the rat. J Comp Neurol 463:249–263
van Groen T, Kadish I, Wyss JM (1999) Efferent connections of the
anteromedial nucleus of the thalamus of the rat. Brain Res Rev
30:1–26
Van Vleet TM, Heldt SA, Corwin JV, Reep RL (2003) Infusion of
apomorphine into the dorsocentral striatum produces acute drug-
induced recovery from neglect produced by unilateral medial
agranular cortex lesions in rats. Behav Brain Res 143:147–157
Vann SD, Aggleton JP (2002) Extensive cytotoxic lesions of the rat
retrosplenial cortex reveal consistent deficits on tasks that tax
allocentric spatial memory. Behav Neurosci 116:85–94
Verberne AJ, Lewis SJ, Worland PJ, Beart PM, Jarrott B, Christie MJ,
Louis WJ (1987) Medial prefrontal cortical lesions modulate
baroreflex sensitivity in the rat. Brain Res 426:243–249
Vertes RP (1991) A PHA-L analysis of ascending projections of the
dorsal raphe nucleus in the rat. J Comp Neurol 313:643–668
Vertes RP (1992) PHA-L analysis of projections from the supra-
mammillary nucleus in the rat. J Comp Neurol 326:595–622
Vertes RP (2002) Analysis of projections from the medial prefrontal
cortex to the thalamus in the rat, with emphasis on nucleus
reuniens. J Comp Neurol 442:163–187
Vertes RP (2004) Differential projections of the infralimbic and
paralimbic cortex in the rat. Synapse 51:32–58
Vertes RP (2006) Interactions among the medial prefrontal cortex,
hippocampus and midline thalamus in emotional and cognitive
processing in the rat. Neuroscience 142:1–20
Vertes RP, Martin GF (1988) Autoradiographic analysis of ascending
projections from the pontine and mesencephalic reticular
formation and the median raphe nucleus in the rat. J Comp
Neurol 275:511–541
Vertes RP, Crane AM, Colom LV, Bland BH (1995) Ascending
projections of the posterior nucleus of the hypothalamus: PHA-L
analysis in the rat. J Comp Neurol 359:90–116
Vertes RP, Fortin WJ, Crane AM (1999) Projections of the median
raphe nucleus in the rat. J Comp Neurol 407:555–582
Vertes RP, Hoover WB, Do Valle AC, Sherman A, Rodriguez JJ
(2006) Efferent projections of reuniens and rhomboid nuclei of
the thalamus in the rat. J Comp Neurol 499:768–796
Vertes RP, Hoover WB, Szigeti-Buck K, Leranth C (2007) Nucleus
reuniens of the midline thalamus: Link between the medial
prefrontal cortex and the hippocampus. Brain Res Bull 71:601–
609
Waterhouse BD, Lin CS, Burne RA, Woodward DJ (1983) The
distribution of neocortical projection neurons in the locus
coeruleus. J Comp Neurol 217:418–431
Witter MP, Room P, Groenewegen HJ, Lohman AHM (1988)
Reciprocal connections of the insular and piriform claustrum
with limbic cortex: an anatomical study in the cat. Neuroscience
24:519–539
Woolf NJ (1991) Cholinergic systems in mammalian brain and spinal
cord. Prog Neurobiol 37:475–524
Woolf NJ, Eckenstein F, Butcher LL (1984) Cholinergic systems in
the rat brain: I. projections to the limbic telencephalon. Brain
Res Bull 13:751–784
Wouterlood FG, Saldana E, Witter MP (1990) Projection from the
nucleus reuniens thalami to the hippocampal region: Light and
electron microscopic tracing study in the rat with the anterograde
tracer Phaseolus vulgaris-leucoagglutinin. J Comp Neurol
296:179–203
Yasui Y, Breder CD, Saper CB, Cechetto DF (1991) Autonomic
responses and efferent pathways from the insular cortex in the
rat. J Comp Neurol 303:355–374
Zaborszky L (2002) The modular organization of brain systems. Basal
forebrain: the last frontier. Prog Brain Res 136:359–372
Zaborszky L, Carlsen J, Brashear HR, Heimer L (1986) Cholinergic
and GABAergic afferents to the olfactory bulb in the rat with
special emphasis on the projection neurons in the nucleus of the
horizontal limb of the diagonal band. J Comp Neurol 243:488–
509
Zaborszky L, Pang K, Somogyi J, Nadasdy Z, Kallo I (1999) The
basal forebrain corticopetal system revisited. Ann NY Acad Sci
877:339–367
Zhang X, Hannesson DK, Saucier DM, Wallace AE, Howland J,
Corcoran ME (2001) Susceptibility to kindling and neuronal
connections of the anterior claustrum. J Neurosci 21:3674–3687
Zilles K (1985) The cortex of the rat: a stereotaxic atlas. Springer,
Berlin, Heidelberg, New York, Tokyo
Zilles K, Wree A (1995) Cortex. In: Paxinos G (ed) The rat nervous
system, 2nd edn. Academic, New York, pp 649–685
Brain Struct Funct (2007) 212:149–179 179
123


















