
A Heterodimer of HEB and an E12-Related Protein Interacts with the ...
9
MOLECULAR AND CELLULAR BIOLOGY, Sept. 1993, p. 5620-5628 0270-7306/93/095620-09$02.00/0 Copyright C 1993, American Society for Microbiology A Heterodimer of HEB and an E12-Related Protein Interacts with the CD4 Enhancer and Regulates Its Activity in T-Cell Lines SHINICHIRO SAWADA1' 2 AND DAN R. LITTMAN' .23* Departments of Microbiology and Immunology1 and of Biochemistry and Biophysics3 and Howard Hughes Medical Institute,2 University of California, San Francisco, California 94143-0414 Received 31 December 1992/Returned for modification 23 February 1993/Accepted 21 June 1993 A T-lymphocyte-specific enhancer located 13 kb upstream of the murine CD4 gene was recently shown to be required for the developmentally regulated expression of CD4. We have previously identified three nuclear protein binding sites in this enhancer, one of these sites, CD4-3, is essential for expression and contains two E-box core motifs (CANNTG) adjacent to each other in the sequence TAACAGGTGT-TAGCIGT. In electrophoretic mobility shift assays using the CD4-3 oligonucleotide as a probe, three nuclear protein complexes, termed CD4-3A, -B, and -C, were detected with nuclear extracts from T-cell lines. CD4-3A, which involves nuclear protein binding to the 5' E-box, was detected only with nuclear extracts from lymphoid cells. Specific antisera were used to show that the CD4-3A complex contains a heterodimer or heterooligomer of basic helix-loop-helix transcriptional factors, E12 or a related factor and HEB, which is expressed predominantly in thymus. Consistent with this finding, in vitro-translated E12 and HEB proteins, as homodimers or het- erodimers, bound preferentially to the 5' E-box. Point mutations in the 5' E-box, but not in the 3' E-box, abolished CD4 enhancer activity. Furthermore, overexpression of Id, a protein that forms inactive het- erodimers with E12/E47, blocked CD4 enhancer activity in T cells. These results suggest that a heterodimer composed of HEB and E12 or a closely related protein plays a critical role in CD4 enhancer function by interacting with the 5' E-box motif of the CD4-3 site in vivo. Development of T lymphocytes within the thymus re- quires appropriately timed expression of several T-cell- specific surface glycoproteins involved in cell-cell interac- tions. Two of these, CD4 and CD8, are coexpressed on the majority of immature thymocytes and have critical roles in the ensuing progression to functional T cells that express one glycoprotein to the exclusion of the other. The CD4 glyco- protein is specifically expressed on the surface of helper T cells, while CD8 is expressed on cytotoxic T cells. CD4 and CD8 bind to nonpolymorphic regions of major histocompat- ibility complex class II and class I molecules, respectively (7, 11, 15, 25, 31, 34, 43, 46). In addition, a cytoplasmic protein tyrosine kinase, p56k , is noncovalently associated with the cytoplasmic domains of both CD4 and CD8 (45, 49, 54, 55). Upon interaction of the T-cell antigen receptor (TCR) with major histocompatibility complex-bound pep- tides, CD4 or CD8 and the associated kinase migrate into the vicinity of the TCR complex, increasing its avidity for the major histocompatibility complex molecule (40, 41, 61). The signals transduced through the collaboration of CD4 and CD8 with the TCR are critical for selection of the T-cell repertoire during thymic ontogeny and for the activation of mature T cells (1, 17, 20, 24, 27, 44). Recent evidence suggests that commitment of a thymo- cyte to express either CD4 or CD8 occurs stochastically and is coordinated with commitment to become a helper or cytotoxic cell, respectively (13). Elucidation of the mecha- nism involved in the regulation of CD4 and CD8 gene expression is likely to yield important insight into processes involved in the functional differentiation of T-cell subsets. In earlier work, we have identified and characterized a T-cell- * Corresponding author. specific enhancer located 13 kb 5' of the transcriptional initiation site of the mouse CD4 gene (47). A minimal element of 339 bp enhanced transcription from the CD4 promoter in both CD4+ CD8- and CD4- CD8+ T-cell lines. The same element, in the context of a CD4 minigene, directs correct subset-specific expression in transgenic mice, sug- gesting that sequences outside of the minimal enhancer are involved in preventing CD4 gene expression in CD4- CD8+ T cells (28). Three nuclear protein binding sites (CD4-1, CD4-2, and CD4-3) were identified in the minimal enhancer element by DNase I footprinting (Fig. 1). The T- and pre-B-cell-specific nuclear factor LEF (53, 57) was shown to bind to the CD4-2 site through its binding motif, CANAG (47). Both the CD4-1 and CD4-3 sites were found to contain E-box motifs, CANNTG. The CD4-3 site contains two E-boxes separated by a single base pair. Mutational analysis of the protein binding sites indicated that CD4-1 and CD4-2 were not essential for transcriptional activity in T cells; in contrast, mutation of both E-boxes within the CD4-3 sequence abol- ished enhancer activity. This result suggested that the bind- ing of one or more E-box-binding proteins, which are mem- bers of the bHLH (basic helix-loop-helix) family of transcription factors, to one or both motifs in CD4-3 is essential for the T-cell-specific activity of this enhancer (47). The E-box-binding proteins of the bHLH family are in- volved in regulating tissue-specific transcription of numer- ous genes, and some of these factors are thought to function as central regulators of cell type determination (36). For example, members of the MyoD family and of the achaete- scute complex mediate myogenesis and neurogenesis, re- spectively (8, 58), and other bHLH factors are involved in the regulation of expression of immunoglobulin and insulin genes. The bHLH transcription factors bind to DNA as 5620 Vol. 13, No. 9
-
Upload
nguyendung -
Category
Documents
-
view
215 -
download
0
Transcript of A Heterodimer of HEB and an E12-Related Protein Interacts with the ...

MOLECULAR AND CELLULAR BIOLOGY, Sept. 1993, p. 5620-56280270-7306/93/095620-09$02.00/0Copyright C 1993, American Society for Microbiology
A Heterodimer of HEB and an E12-Related Protein Interactswith the CD4 Enhancer and Regulates Its Activity in
T-Cell LinesSHINICHIRO SAWADA1' 2 AND DAN R. LITTMAN'.23*
Departments ofMicrobiology and Immunology1 and ofBiochemistry and Biophysics3 and Howard HughesMedical Institute,2 University of California, San Francisco, California 94143-0414
Received 31 December 1992/Returned for modification 23 February 1993/Accepted 21 June 1993
A T-lymphocyte-specific enhancer located 13 kb upstream of the murine CD4 gene was recently shown to berequired for the developmentally regulated expression of CD4. We have previously identified three nuclearprotein binding sites in this enhancer, one of these sites, CD4-3, is essential for expression and contains twoE-box core motifs (CANNTG) adjacent to each other in the sequence TAACAGGTGT-TAGCIGT. Inelectrophoretic mobility shift assays using the CD4-3 oligonucleotide as a probe, three nuclear proteincomplexes, termed CD4-3A, -B, and -C, were detected with nuclear extracts from T-cell lines. CD4-3A, whichinvolves nuclear protein binding to the 5' E-box, was detected only with nuclear extracts from lymphoid cells.Specific antisera were used to show that the CD4-3A complex contains a heterodimer or heterooligomer ofbasichelix-loop-helix transcriptional factors, E12 or a related factor and HEB, which is expressed predominantly inthymus. Consistent with this finding, in vitro-translated E12 and HEB proteins, as homodimers or het-erodimers, bound preferentially to the 5' E-box. Point mutations in the 5' E-box, but not in the 3' E-box,abolished CD4 enhancer activity. Furthermore, overexpression of Id, a protein that forms inactive het-erodimers with E12/E47, blocked CD4 enhancer activity in T cells. These results suggest that a heterodimercomposed of HEB and E12 or a closely related protein plays a critical role in CD4 enhancer function byinteracting with the 5' E-box motif of the CD4-3 site in vivo.
Development of T lymphocytes within the thymus re-quires appropriately timed expression of several T-cell-specific surface glycoproteins involved in cell-cell interac-tions. Two of these, CD4 and CD8, are coexpressed on themajority of immature thymocytes and have critical roles inthe ensuing progression to functional T cells that express oneglycoprotein to the exclusion of the other. The CD4 glyco-protein is specifically expressed on the surface of helper Tcells, while CD8 is expressed on cytotoxic T cells. CD4 andCD8 bind to nonpolymorphic regions of major histocompat-ibility complex class II and class I molecules, respectively(7, 11, 15, 25, 31, 34, 43, 46). In addition, a cytoplasmicprotein tyrosine kinase, p56k , is noncovalently associatedwith the cytoplasmic domains of both CD4 and CD8 (45, 49,54, 55). Upon interaction of the T-cell antigen receptor(TCR) with major histocompatibility complex-bound pep-tides, CD4 or CD8 and the associated kinase migrate into thevicinity of the TCR complex, increasing its avidity for themajor histocompatibility complex molecule (40, 41, 61). Thesignals transduced through the collaboration of CD4 andCD8 with the TCR are critical for selection of the T-cellrepertoire during thymic ontogeny and for the activation ofmature T cells (1, 17, 20, 24, 27, 44).
Recent evidence suggests that commitment of a thymo-cyte to express either CD4 or CD8 occurs stochastically andis coordinated with commitment to become a helper orcytotoxic cell, respectively (13). Elucidation of the mecha-nism involved in the regulation of CD4 and CD8 geneexpression is likely to yield important insight into processesinvolved in the functional differentiation of T-cell subsets. Inearlier work, we have identified and characterized a T-cell-
* Corresponding author.
specific enhancer located 13 kb 5' of the transcriptionalinitiation site of the mouse CD4 gene (47). A minimalelement of 339 bp enhanced transcription from the CD4promoter in both CD4+ CD8- and CD4- CD8+ T-cell lines.The same element, in the context of a CD4 minigene, directscorrect subset-specific expression in transgenic mice, sug-gesting that sequences outside of the minimal enhancer areinvolved in preventing CD4 gene expression in CD4- CD8+T cells (28).Three nuclear protein binding sites (CD4-1, CD4-2, and
CD4-3) were identified in the minimal enhancer element byDNase I footprinting (Fig. 1). The T- and pre-B-cell-specificnuclear factor LEF (53, 57) was shown to bind to the CD4-2site through its binding motif, CANAG (47). Both the CD4-1and CD4-3 sites were found to contain E-box motifs,CANNTG. The CD4-3 site contains two E-boxes separatedby a single base pair. Mutational analysis of the proteinbinding sites indicated that CD4-1 and CD4-2 were notessential for transcriptional activity in T cells; in contrast,mutation of both E-boxes within the CD4-3 sequence abol-ished enhancer activity. This result suggested that the bind-ing of one or more E-box-binding proteins, which are mem-bers of the bHLH (basic helix-loop-helix) family oftranscription factors, to one or both motifs in CD4-3 isessential for the T-cell-specific activity of this enhancer (47).The E-box-binding proteins of the bHLH family are in-
volved in regulating tissue-specific transcription of numer-ous genes, and some of these factors are thought to functionas central regulators of cell type determination (36). Forexample, members of the MyoD family and of the achaete-scute complex mediate myogenesis and neurogenesis, re-spectively (8, 58), and other bHLH factors are involved inthe regulation of expression of immunoglobulin and insulingenes. The bHLH transcription factors bind to DNA as
5620
Vol. 13, No. 9

CD4 ENHANCER AND bHLH PROTEINS 5621
homodimers or heterodimers. Dimerization occurs via theHLH motif, which, along with the basic region, is requiredfor DNA binding (6, 9, 36, 56).A series of closely related bHLH proteins, E12/E47/ITF1
(22, 36), ITF2 (22), and HTF4/HEB (23, 62), have beencloned from human cDNA expression libraries by screeningfor binding of labeled E-box sequences. Although thesefactors are expressed ubiquitously, they are believed toparticipate in the formation of heterodimeric transcriptionfactors that direct tissue-specific regulation of expression ofthe immunoglobulin p, and K chain genes, muscle-specificgenes, and the insulin gene (2, 12, 33, 37, 50). In musclecells, E12-like proteins dimerize with the muscle-cell-spe-cific MyoD family proteins (Myf-5, myogenin, MyoD, andMRF/herculin/Myf-6) and thus mediate expression of mus-cle- specific genes (14, 33, 59). The function of such het-erodimers is inhibited by expression of the Id proteins, HLHproteins that lack the basic region and dimerize with E12/E47 to form inactive complexes devoid of DNA bindingactivity (3, 4, 10, 16, 51). It has been proposed that throughthis process Id proteins regulate tissue-specific gene expres-sion during development (26, 32, 51, 60).
In this study, we have begun to analyze the nuclear factorsthat bind to the critical CD4-3 site. We have found that anoligomer containing the bHLH proteins HEB and E12, or aprotein immunologically related to E12, binds to the 5'E-box of the CD4-3 site and is present in a lymphoidcell-specific nuclear protein complex. Enhancer functionwas disrupted by mutations that blocked formation of thiscomplex and by expression of Id. These studies suggest thatthe interaction of HEB and an E12-like protein with theCD4-3 site plays a crucial role in CD4 enhancer function.
MATERIALS AND METHODS
Cell culture. All cell lines were grown in RPMI 1640medium containing 10% fetal calf serum, 50 puM 2-mercap-toethanol, and antibiotics.
Construction of CAT expression vectors. The pCD4CATvector, in which the chloramphenicol acetyltransferase(CAT) gene is driven by the CD4 promoter, was prepared byreplacing the thymidine kinase promoter fragment ofpBLCAT2 (35) with the XbaI-to-XhoI fragment (487 bp) ofthe CD4 promoter region, which contains the transcriptioninitiation site and 70 bp of the untranslated first exon (33a).The 339-bp minimal CD4 enhancer fragments (BstXI toAvaII) with or without mutations were inserted in thepolylinker site at the 5' end of pCD4CAT in a senseorientation.
Transfections and CAT assays. Transfections were per-formed with the DEAE-dextran procedure (21) with minormodifications (47). Rous sarcoma virus-luciferase plasmid (2p,g) was cotransfected to control for transfection efficiency.After 48 h, cells were harvested and lysed by four cycles offreezing and thawing. The lysates were centrifuged at 4°C,and the supernatants were recovered. For the CAT assays,the extracts were further incubated at 56°C for 10 min andcentrifuged to remove debris. Protein concentration wasdetermined, and extract containing 100 p,g of protein wasassayed for its ability to convert [14C]chloramphenicol to theacetylated form overnight. Quantitation of acetylation ratioswas performed with a PhosphorImager system (MolecularDynamics). Luciferase assays were performed according tothe method of Ow et al. (42). For Id cotransfection studies,10 p.g of reporter plasmids and 5 p.g of Rous sarcomavirus-luciferase plasmid were transfected with 10 and 15 p.g
of either a plasmid expressing the Id cDNA (pE:Id[S]) or aplasmid with the Id cDNA in the antisense orientation(pE:Id[A]). The total amount of DNA in each of the sampleswas adjusted to 30 ,ug with pBluescript DNA. The pE:Id[S]and pE:Id[A] were in the plasmid pEMSV-scribe a2, underthe transcriptional regulation of the murine sarcoma viruslong terminal repeat promoter (3). These plasmids werekindly provided by R. Benezra (Memorial Sloan-KetteringCancer Center [3]).
Electrophoretic mobility shift assay (EMSA). End-labeleddouble-stranded oligonucleotide probes (10,000 or 50,000cpm) were incubated in the binding reaction buffer (10%glycerol, 10 mM HEPES [N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid]-NaOH [pH 7.9], 4 mM Tris-HCl [pH7.9], 80 mM NaCl, 1 mM EDTA, 1 mM dithiothreitol, 300 p.gof bovine serum albumin per ml) with 2 p,g of poly(dI-dC)poly(dI-dC) and 5 pg of a crude nuclear extract for 15 min ina 30°C water bath. The nuclear extracts were prepared asdescribed by Schreiber et al. (48). Following binding, themixture was electrophoresed through a native high-ionic-strength gel (4% acrylamide, 0.05% bisacrylamide, 2.5%glycerol, 50 mM Tris base, 380 mM glycine, 2 mM EDTA).Electrophoresis was carried out at 30 mA for 2 to 3 h at 4°C.The gel was dried and autoradiographed with an intensifyingscreen at -70°C. For competition experiments, specific andnonspecific competitor DNAs (5 to 50 ng) were incubated inthe mixture prior to addition of the nuclear extracts. Thesequences of wild-type and mutant oligonucleotides forCD4-1 and CD4-3 sites are shown in Fig. 2A. The wild-typeoligonucleotides for the KE2 and CD4-2 binding sites are5'-CAAGGCAGGTGGCCCAC-3' and 5'-AAAAAAGAACAAAGGGCCTAGAT1T-3', respectively. For the supershiftassay, 1 ,ul of antiserum against E12 (16 mg/ml) or ofpreimmune serum (19 mg/ml), undiluted or diluted 1:5 or1:25, was added to the mixture. These sera were kindlyprovided by C. Murre (38).
Expression screening. Purified fusion protein containingthe HLH motif of Syrian hamster shPAN-2 (amino acids 509to 649; R-554 -- A, R-556 -* A [18]), a heart muscle kinaserecognition site, and the FLAG epitope was kindly providedby M. A. Blanar and was labeled with [y-32P]ATP and usedto screen 1.8 x 106 plaques of a Xgtll human thymus cDNAlibrary (Clontech, Palo Alto, Calif.) as described previously(5). The cDNA inserts of clones identified in the screen wereinserted into the EcoRI cloning site of pBluescript, anddouble-stranded sequencing was performed with Sequenase(United States Biochemical, Cleveland, Ohio).
In vitro transcription and translation. pHEB was con-structed by inserting a cDNA fragment (nucleotides 140 to2306) encoding the full-length HEB protein into the cloningsite of pBluescript. pE12R, containing a partial E12 cDNA,was kindly provided by C. Murre (36). The ITF2 cDNA wasa gift of T. Kadesch (22). In vitro transcription-translationwas performed in a final volume of 25 p.l with T3 polymeraseand rabbit reticulocyte lysate, as described in the Promegatechnical manual (Promega Corp., Madison, Wis.). Proteinwas synthesized at 30°C for 2 h. One microliter of thereaction mixture was used for electrophoretic mobility shiftassays (EMSA). To compare the mobilities of closely mi-grating complexes (Fig. 5), separation was performed on a3.5% acrylamide gel for 7 h at 30 mA.
Preparation of antisera. A plasmid expressing a glutathi-one S-transferase (GST)-HEB peptide-1 (amino acids 530 to554, HEPPSSDDMKSDDESSQKDIKVSSR [23]) fusionprotein was prepared by inserting the corresponding double-stranded oligonucleotide between the BamHI and EcoRI
VOL. 13, 1993

5622 SAWADA AND LITTMAN
CD4-1 CD4-2BXI -C)----C -
100 200 300
CD4-1 CCCCAGCTGTTAGCCCTCT
CD4-2 AAGAACAAAGGGCCTAGATTT
CD4-3 TAACAGGTGTCAGCTGGCT
FIG. 1. Nuclear protein binding sites in the minimal CD4 en-hancer. The binding sites are represented as open boxes in the339-bp BstXI-to-AvaII fragment of the CD4 enhancer. The se-
quences protected in the DNase I footprinting assay are shownbelow the map. The E-box core motifs (CD4-1 and CD4-3) and LEFbinding motif (CD4-2) are underlined.
cloning sites of pGEX-2T (Pharmacia, Piscataway, N.J.).Fusion protein was induced and purified with glutathioneagarose beads (Sigma, St. Louis, Mo.) as described previ-ously (30). Antiserum against the fusion protein was pro-duced by immunizing rabbits (CALTAG Laboratories,Healdsburg, Calif.). GST-HEB (amino acids 254 to 682 [23])was prepared by inserting the HEB cDNA (nucleotides 966to 2306) at the EcoRI cloning site of pGEX-2T. Purifiedfusion protein was coupled with beaded agarose with theReacti-Gel (6X) carbodiimidazole system (Pierce, Rockford,Ill.). GST-HEB beads (50 ,ll) were added into 100 ,ul ofeightfold-diluted anti-E12 antiserum (kindly provided by C.Murre) at 4°C for 1 h to absorb antibodies against HEB.Absorption was repeated three times.
Site-directed mutagenesis. The minimal enhancer fragmentwas inserted into the polylinker site of pBluescript KS(+)vector, and single-strand DNA was prepared by infection ofthe bacteria with helper phage. Mutants were produced byusing an oligonucleotide-mediated, gapped heteroduplexmutagenesis protocol according to the manual of the Muta-Gene in vitro mutagenesis kit (Bio-Rad). All mutants wereconfirmed by dideoxy-DNA sequence analysis with com-mercially available Sequenase reagents (United States Bio-chemical).
RESULTS
Three nuclear protein-DNA complexes are detected byEMSA with a CD4-3 oligonucleotide. The CD4-3 site, one ofthree nuclear protein binding sites in the minimal enhancerelement, contains two adjacent E-box motifs (Fig. 1). Tocharacterize proteins that bind to the CD4-3 site, a labeledsynthetic oligonucleotide corresponding to the CD4-3 se-
quence was incubated with nuclear extract from the T-cellline SL3B and subjected to EMSA (Fig. 2B). Using bindingconditions with NaCl at 80 mM, we could detect a slowlymigrating complex, CD4-3A, which was barely detectable inour previous analysis with NaCl at 200 mM (Fig. 2B, lane 1)(47). The other complexes, CD4-3B and CD4-3C, have beendetected with either condition (47). All three complexeswere specific, since they were inhibited by the wild-typeCD4-3 oligonucleotide but not by a CD4-3 oligonucleotidemutated in both E-boxes or by control salmon sperm DNA(Fig. 2B, lanes 6 to 11).The CD4-1 binding site also contains an E-box motif and
was hence tested for its ability to compete for proteinbinding to the CD4-3 site. The wild-type, but not the mutant,CD4-1 oligonucleotide competed for both CD4-3A andCD4-3C binding (Fig. 2B, lanes 2 to 5), suggesting that both
the CD4-3A and CD4-3C complexes contain factors whichbind to the CD4-1 sequence. However, the CD4-1 oligonu-cleotide competed for CD4-3A binding less effectively thandid the CD4-3 oligonucleotide, suggesting that CD4-3A nu-clear protein interacts preferentially with the CD4-3 se-quence (Fig. 2B, lanes 2 and 6). No competition wasobserved between CD4-1 and the CD4-3B complex.Mapping of the binding sites for the CD4-3 nuclear factors.
A panel of CD4-3 double-stranded oligonucleotides withE-box mutations was synthesized and used in competitionassays to localize sequences involved in the formation of thecomplexes. Ml has mutations in both E-boxes; M2, M4, andM6 have point mutations in the 5' E-box; and M3, M5, andM7 have point mutations in the 3' E-box (Fig. 2A). The 5'E-box mutants (M2, M4, and M6) competed efficiently forCD4-3C binding but not for CD4-3A binding (Fig. 2C). Incontrast, the 3' E-box mutants (M3, M5, and M7) competedefficiently for CD4-3A binding but not for CD4-3C binding.None of the mutant oligonucleotides competed efficiently forCD4-3B binding. Thus, single nucleotide substitutions in theupstream E-box, including a substitution inside the motif(M6), significantly reduced the level of CD4-3A binding,whereas those in the downstream E-box likewise affectedCD4-3C binding. These results therefore indicate that theCD4-3A complex results from protein binding to the 5'E-box, whereas the CD4-3C complex involves binding to the3' E-box.Some cross-competition was observed in this assay when
50 ng of competing oligonucleotide was used. Therefore,nuclear proteins that bind strongly to one E-box motifappear to also bind weakly to the other E-box. The Mloligonucleotide, which is mutated in both E-boxes, did notcompete at all (Fig. 2C, lane 3). Point mutations in either ofthe two E-boxes abolished competition for CD4-3B binding,suggesting that this complex involves protein interactionswith sequences contained in both motifs.
In addition, oligonucleotides from both the CD4-1 site andthe immunoglobulin enhancer cE2 site competed forCD4-3A binding, but only the CD4-1 oligonucleotide com-peted for CD4-3C binding (Fig. 2C, lanes 10 and 11).Therefore, CD4-3A, but not CD4-3C, appears to contain KE2binding factors, whereas both complexes appear to containCD4-1 binding factors. The CD4-2 oligonucleotide contain-ing the LEF binding site and control salmon sperm DNA didnot compete for the formation of any of the complexes (Fig.2C, lanes 12 and 13).Lymphoid specificity of the CD4-3A complex. We next
examined the cell type specificity of the factors binding tothe CD4-3 site. Nuclear extracts from various cell lines wereincubated with the labeled CD4-3 oligonucleotide and sub-jected to EMSA (Fig. 3). CD4-3B and CD4-3C were detect-able in extracts from lymphoid and nonlymphoid cell lines,with the exception of the absence of CD4-3C from NIH 3T3cells (Fig. 3) (47). In contrast, the CD4-3A complex wasdetected, albeit at various levels, in extracts from all T- andB-cell lines, but not in extracts from the nonlymphoid celllines P815, NIH 3T3, HeLa, K562, and U937 (Fig. 3). Thus,CD4-3A is expressed in a lymphoid cell-specific manner,whereas CD4-3B and CD4-3C appear to be widely ex-pressed.The CD4-3A complex contains an oligomer of HEB and an
E12-like protein. To determine whether any of the complexescontain E12-like proteins, the labeled CD4-3 oligonucleotidewas incubated with nuclear extract from the T-cell line SL3Bin the presence of antiserum against E12 and subjected toEMSA (Fig. 4). The antiserum supershifted the CD4-3A
MOL. CELL. BIOL.
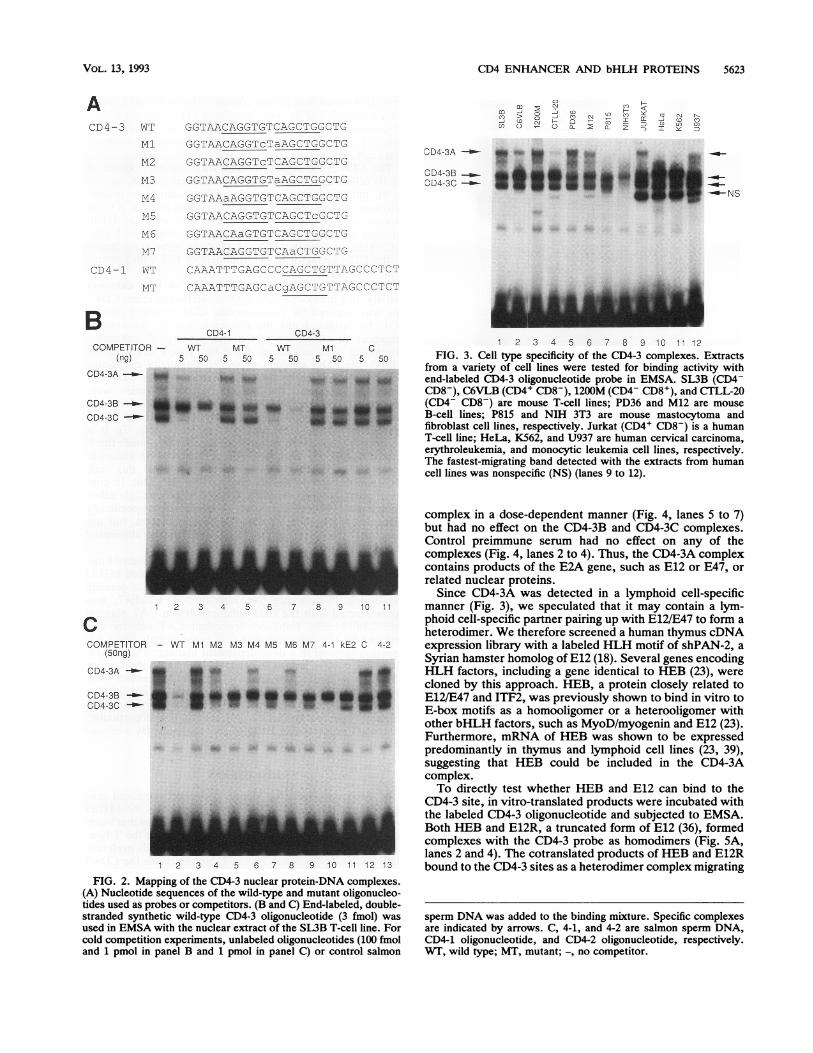
CD4 ENHANCER AND bHLH PROTEINS 5623
ACD4-3 WT GGTAACAGGTGTCAGCTGG-CTG
Ml GGTAACAGGTcTaAGCTGGCTG
M2 GGTAACAGGTcTCAGCTGGCTG
M3 GGTAACAGGTGTaAGCTGGCTG
M4 C-GTAAaAGGTGTCAGCTGGCTG
Ms GGTAACAGGTGTCAGCTcGCTG-
M6 GGTAACAaGTGTCAGCTGGCTG
M7 GGTAACAGGTGTCAaCTGGCTG
CD4-1 WT CAAATTTGAGCCCCAGCTGTTACGCCCTCT
MT CAAATTTGAGCaCgAGC-TST-AGCCTC
C)
co i1J <
>i co L.0 Cl
C) CE) m- _D D C0 2 -- o
C~) 0 EL )--- :0 ~ c
vD4-3A -
CD4-3B _3D4-3C-&
AI .0
BCOMPETITOR -
(ng)
CD4-1 CWT MT WT
5 50 5 50 5 50
'D4-3Ml C
5 50 5 50
CD4-3A -_.- gpl wo tw_ I0~~~~~~~~~~~~~..
CD4-3B---
CD4-3C -= R..f.
1 2 3 4 5 6 7 8 9 10 11 12FIG. 3. Cell type specificity of the CD4-3 complexes. Extracts
from a variety of cell lines were tested for binding activity withend-labeled CD4-3 oligonucleotide probe in EMSA. SL3B (CD4-CD8-), C6VLB (CD4+ CD8-), 1200M (CD4- CD8+), and CTLL-20(CD4- CD8-) are mouse T-cell lines; PD36 and M12 are mouseB-cell lines; P815 and NIH 3T3 are mouse mastocytoma andfibroblast cell lines, respectively. Jurkat (CD4+ CD8-) is a humanT-cell line; HeLa, K562, and U937 are human cervical carcinoma,erythroleukemia, and monocytic leukemia cell lines, respectively.The fastest-migrating band detected with the extracts from humancell lines was nonspecific (NS) (lanes 9 to 12).
2 3 4 5
CCOMPETITOR -
(50ng)
6 7 8 9 10 11
WT Ml M2 M3 M4 MS M6 M7 4-1 kE2 C 4-2
ICD4-3A- - *P -P.
CD4-3B_ x34-3C-
i-D4-3C-)5
1 2 3 4 5 6 7 8 9 10 11 12 13
FIG. 2. Mapping of the CD4-3 nuclear protein-DNA complexes.(A) Nucleotide sequences of the wild-type and mutant oligonucleo-tides used as probes or competitors. (B and C) End-labeled, double-stranded synthetic wild-type CD4-3 oligonucleotide (3 fmol) wasused in EMSA with the nuclear extract of the SL3B T-cell line. Forcold competition experiments, unlabeled oligonucleotides (100 fmoland 1 pmol in panel B and 1 pmol in panel C) or control salmon
complex in a dose-dependent manner (Fig. 4, lanes 5 to 7)but had no effect on the CD4-3B and CD4-3C complexes.Control preimmune serum had no effect on any of thecomplexes (Fig. 4, lanes 2 to 4). Thus, the CD4-3A complexcontains products of the E2A gene, such as E12 or E47, orrelated nuclear proteins.
Since CD4-3A was detected in a lymphoid cell-specificmanner (Fig. 3), we speculated that it may contain a lym-phoid cell-specific partner pairing up with E12/E47 to form aheterodimer. We therefore screened a human thymus cDNAexpression library with a labeled HLH motif of shPAN-2, aSyrian hamster homolog of E12 (18). Several genes encodingHLH factors, including a gene identical to HEB (23), werecloned by this approach. HEB, a protein closely related toE12/E47 and ITF2, was previously shown to bind in vitro toE-box motifs as a homooligomer or a heterooligomer withother bHLH factors, such as MyoD/myogenin and E12 (23).Furthermore, mRNA of HEB was shown to be expressedpredominantly in thymus and lymphoid cell lines (23, 39),suggesting that HEB could be included in the CD4-3Acomplex.To directly test whether HEB and E12 can bind to the
CD4-3 site, in vitro-translated products were incubated withthe labeled CD4-3 oligonucleotide and subjected to EMSA.Both HEB and E12R, a truncated form of E12 (36), formedcomplexes with the CD4-3 probe as homodimers (Fig. SA,lanes 2 and 4). The cotranslated products of HEB and E12Rbound to the CD4-3 sites as a heterodimer complex migrating
sperm DNA was added to the binding mixture. Specific complexesare indicated by arrows. C, 4-1, and 4-2 are salmon sperm DNA,CD4-1 oligonucleotide, and CD4-2 oligonucleotide, respectively.WT, wild type; MT, mutant; -, no competitor.
VOL. 13, 1993

5624 SAWADA AND LI1TMAN
CONTROL ANTI-El 2/E47X2= X5 Xl X25 X5 Xl
ii.s
1 2 3 4 5 6 7FIG. 4. Supershifting of the CD4-3A complex with an anti-E12/
E47 antibody. End-labeled CD4-3 oligonucleotide was incubatedwith the nuclear extract from SL3B cells in the presence of seriallydiluted preimmune (lanes 2 to 4) or immune (lanes 5 to 7) serum priorto EMSA. Lane 1, no serum.
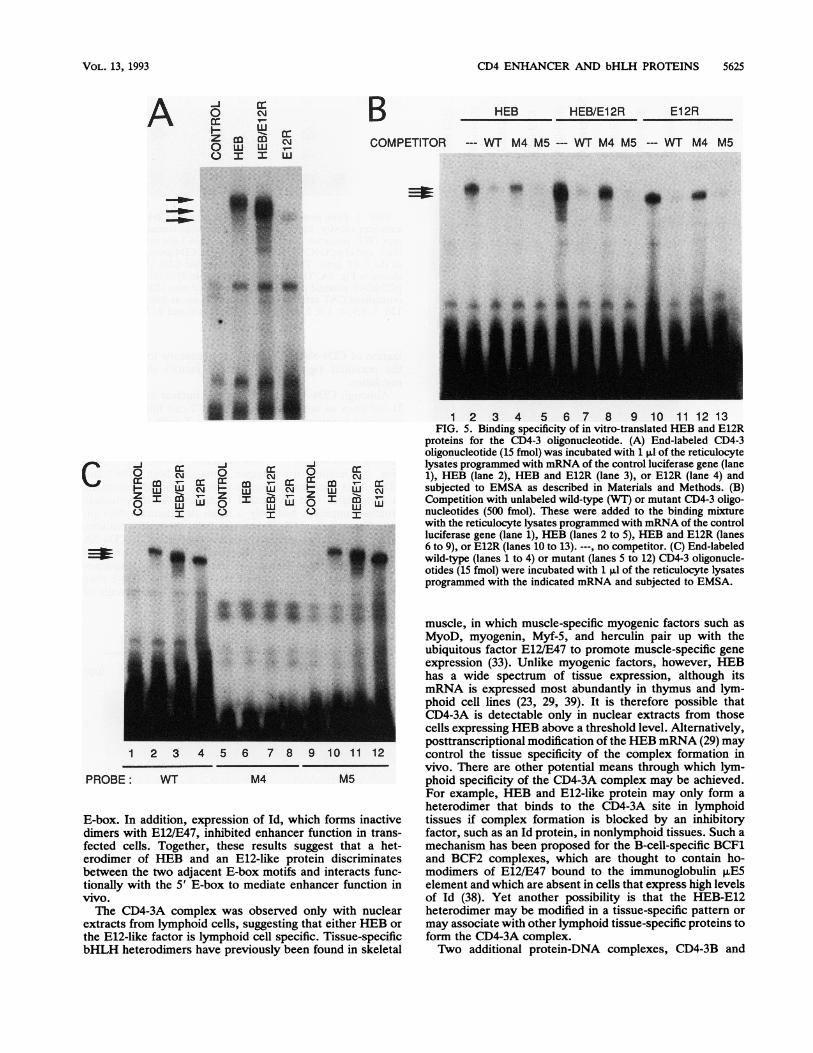
with a mobility intermediate between those of the ho-modimer complexes (Fig. 5A, lane 3). Additional bandsmigrating faster may represent products of alternative trans-lation initiation codons, as previously suggested (23). Tolocalize the sequences interacting with these factors, CD4-3oligonucleotides mutated in the E-boxes were used in com-petition assays (Fig. 5B). The M4 oligonucleotide, which hasa point mutation in the 5' E-box, competed poorly forbinding of all factors. In contrast, the M5 oligonucleotide,which has a point mutation in the 3' E-box, competed asefficiently as the wild-type oligonucleotide. The ability of thein vitro-translated factors to directly bind to labeled mutantCD4-3 oligonucleotides was also tested (Fig. 5C). Ho-modimers or heterodimers of HEB and E12R were clearlydetected with the wild-type probe and the M5 probe but notwith the M4 probe. These results indicate that HEB and E12,as homodimers or heterodimers, interact preferentially withthe 5' E-box of the CD4-3 site, similar to the CD4-3A factorin nuclear extracts of lymphoid cell lines.To determine whether HEB is present in the CD4-3A
complex, a specific antiserum against HEB was raised byimmunizing rabbits with a GST-HEB peptide-1 (amino acids530 to 554) fusion protein. This peptide was selected fromthe HEB amino acid sequence by virtue of its relatively lowhomology with the closely related bHLH factors E12/E47and ITF2. The antiserum against E12, which cross-reactswith in vitro-translated HEB (data not shown), was depletedof its cross-reactivity by preabsorption with a GST-HEBfusion protein coupled with agarose beads. The specificity ofthese antisera was tested on in vitro-translated proteins inEMSA with the CD4-3 oligonucleotide. The anti-HEB serumbut not the preabsorbed anti-E12 serum supershifted invitro-translated HEB, whereas the preabsorbed anti-E12serum but not the anti-HEB serum recognized in vitro-translated E12R (Fig. 6, lanes 1 to 6). The specificity of the
MOL. CELL. BIOL.
anti-HEB serum was further confirmed by its failure to reactwith a complex of in vitro-translated ITF2 and the CD4-3oligonucleotide (data not shown). In contrast, the absorbedanti-E12 antiserum disrupted the ITF2-containing complex(data not shown).The CD4-3A complex formed with nuclear extract of
SL3B cells was recognized by both antisera (Fig. 6, lanes 7to 9), indicating that it contains a heterodimer or hetero-oligomer of HEB and E12 or an E12-related protein. A smallamount of CD4-3A was not supershifted with anti-HEB,whereas all detectable CD4-3A was supershifted with anti-E12. This suggests that CD4-3A contains some homodimersof the E12-like factor or heterodimers of E12 with proteinsother than HEB.
Effects of mutations in the CD4-3 site on CD4 enhanceractivity. In a previous study, we showed that an intact CD4-3site was required for T-cell-specific CD4 enhancer activity(47). To further define the elements that are involved inenhancer function, we tested the activity of enhancerscontaining point mutations in the two E-box motifs of theCD4-3 site. The minimal enhancer fragments containingthese mutations were subcloned upstream of the CD4 pro-moter-CAT plasmid (pCD4CAT). These plasmids weretransfected into the T-cell line 1200M, and CAT activity wasmeasured 2 days later. The point mutations in the 5' E-boxabolished the enhancer activity completely (Fig. 7). It isnoteworthy that the substitution of a nucleotide inside the 5'E-box (CAGGIX to CAAG2i) also abolished enhanceractivity (Fig. 7, M6). In the competition analysis, this samemutation disrupted CD4-3A binding (Fig. 2C, M6). In con-trast, point mutations in the 3' E-box had only a slight effecton enhancer activity. Therefore, mutations that specificallydisrupted the binding of CD4-3A nuclear factors, but notthose that disrupted binding of CD4-3C factors, eliminatedenhancer activity.
Overexpression of Id protein inhibits CD4 enhancer func-tion. To test the potential role of E12/E47 or related bHLHproteins in the function of the CD4 enhancer, a plasmidexpressing Id-1, a negative regulator of E12/E47, wascotransfected with the CD4 enhancer-driven CAT plasmidinto 1200M cells. Enhancer-dependent CAT activity wasreduced by the cotransfection of a sense-oriented Id-1 ex-pression plasmid in a dose-dependent manner. The expres-sion of a control antisense-oriented Id-1 had only a minoreffect on enhancer activity (Fig. 8). Similar results wereobtained with the human T-cell line Jurkat (data not shown).As Id-1 is known to form inactive heterodimers with bHLHproteins in general and with E2A gene products in particular,this result suggests that E12/E47-like proteins play a crucialrole in CD4 enhancer activity in T cells.
DISCUSSION
In this study, we have identified a novel lymphoid cell-specific nuclear factor, CD4-3A, that contains the bHLHproteins HEB and E12/E47 or related proteins and that bindsto a DNA sequence essential for the activity of the T-lym-phocyte-specific CD4 enhancer. This factor binds preferen-tially to one of the two adjacent E-box motifs in the CD4-3site of the enhancer; point mutations in this 5' E-box, but notin the 3' E-box, resulted in both the loss of formation of theCD4-3A complex on EMSA and the loss of CD4 enhanceractivity in T cells. The CD4-3A complex was reactive withspecific antibodies against HEB and E12/E47. Consistentwith these results, in vitro-translated E12 and HEB, as eitherhomodimers or heterodimers, bound specifically to the 5'
CD4-3A -_
CD4 ENHANCER AND bHLH PROTEINS 5625
A°0 C%
(_) I I
B- COMPETITOR
HEB HEB/EI2R EI2R
--- WT M4 M5--- WT M4 M5 --- WT M4 M5
a:ac a cc cc m_ rN- uH LL CK wL Wi
0
_ _ s_ i_.F _ __ F . w . -.w m. ........-F:
,; =
§ t lllX |Z ', 5'
1 2 3 4 5 6 7 8 9 10 11 12
PROBE: WT M4 M5
E-box. In addition, expression of Id, which forms inactivedimers with E12/E47, inhibited enhancer function in trans-fected cells. Together, these results suggest that a het-erodimer of HEB and an E12-like protein discriminatesbetween the two adjacent E-box motifs and interacts func-tionally with the 5' E-box to mediate enhancer function invivo.The CD4-3A complex was observed only with nuclear
extracts from lymphoid cells, suggesting that either HEB orthe E12-like factor is lymphoid cell specific. Tissue-specificbHLH heterodimers have previously been found in skeletal
1 2 3 4 5 6 7 8 9 10 1112 13FIG. 5. Binding specificity of in vitro-translated HEB and E12R
proteins for the CD4-3 oligonucleotide. (A) End-labeled CD4-3oligonucleotide (15 fmnol) was incubated with 1 p.1 of the reticulocytelysates programmed with mRNA of the control luciferase gene (lane1), HEB (lane 2), HEB and E12R (lane 3), or E12R (lane 4) andsubjected to EMSA as described in Materials and Methods. (B)Competition with unlabeled wild-type (WT) or mutant CD4-3 oligo-nucleotides (500 fmnol). These were added to the binding mixturewith the reticulocyte lysates programmed with mRNA of the controlluciferase gene (lane 1), HEB (lanes 2 to 5), HEB and E12R (lanes6 to 9), or E12R (lanes 10 to 13). ---, no competitor. (C) End-labeledwild-type (lanes 1 to 4) or mutant (lanes 5 to 12) CD4-3 oligonucle-otides (15 fmnol) were incubated with 1 p.l of the reticulocyte lysatesprogrammed with the indicated mRNA and subjected to EMSA.
muscle, in which muscle-specific myogenic factors such asMyoD, myogenin, Myf-5, and herculin pair up with theubiquitous factor E12/E47 to promote muscle-specific geneexpression (33). Unlike myogenic factors, however, HEBhas a wide spectrum of tissue expression, although itsmRNA is expressed most abundantly in thymus and lym-phoid cell lines (23, 29, 39). It is therefore possible thatCD4-3A is detectable only in nuclear extracts from thosecells expressing HER above a threshold level. Alternatively,posttranscriptional modification of the HEB mRNA (29) maycontrol the tissue specificity of the complex formation invivo. There are other potential means through which lym-phoid specificity of the CD4-3A complex may be achieved.For example, HEB and E12-like protein may only form aheterodimer that binds to the CD4-3A site in lymphoidtissues if complex formation is blocked by an inhibitoryfactor, such as an Id protein, in nonlymphoid tissues. Such amechanism has been proposed for the B-cell-specific BCF1and BCF2 complexes, which are thought to contain ho-modimers of E12/E47 bound to the immunoglobulin p.E5element and which are absent in cells that express high levelsof Id (38). Yet another possibility is that the HEB-E12heterodimer may be modified in a tissue-specific patter ormay associate with other lymphoid tissue-specific proteins toform the CD4-3A complex.T`wo additional protein-DNA complexes, CD4-3B and
a*....
C c°O w
o
VOL. 13, 1993
5626 SAWADA AND LI1TMAN
-4
W
SERUM: TIi
o Cu C
PROTEIN: HEB
-4
o cu CuJE CO
El 2R
-4
o Cu Cu
SL3B N.E.
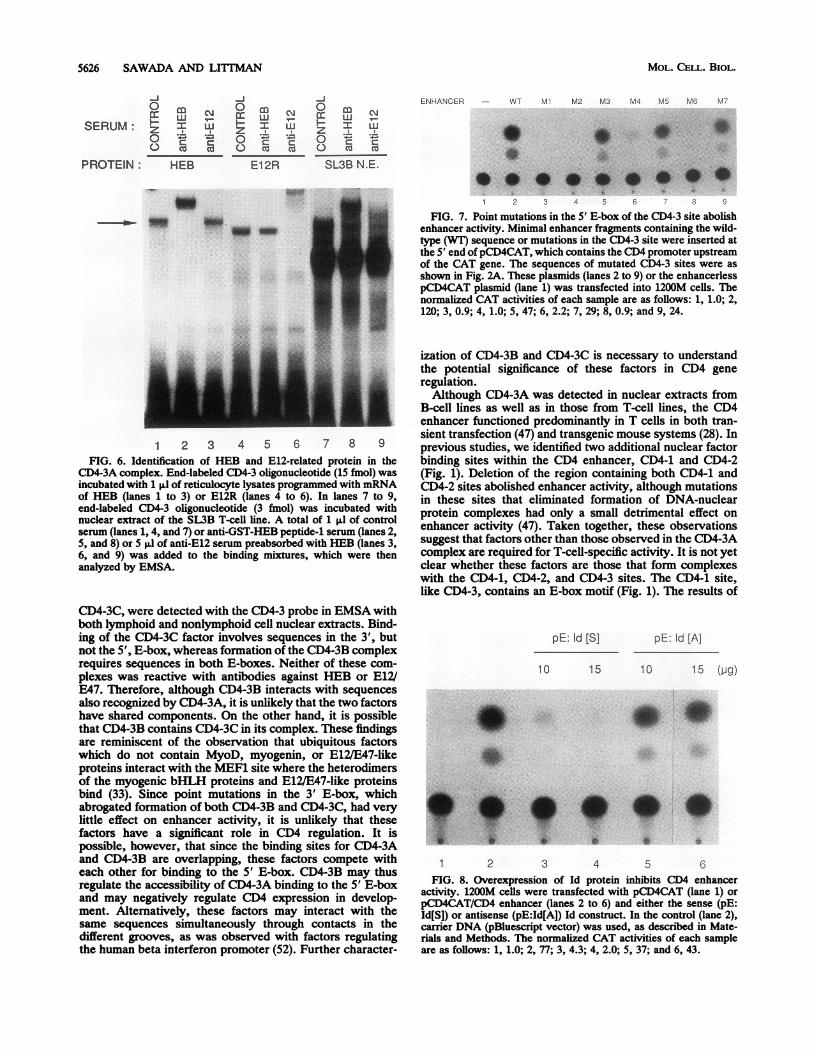
1 2 3 4 5 6 7 8 9FIG. 6. Identification of HEB and E12-related protein in the
CD4-3A complex. End-labeled CD4-3 oligonucleotide (15 fmol) wasincubated with 1 ,l of reticulocyte lysates programmed with mRNAof HEB (lanes 1 to 3) or E12R (lanes 4 to 6). In lanes 7 to 9,end-labeled CD4-3 oligonucleotide (3 fmol) was incubated withnuclear extract of the SL3B T-cell line. A total of 1 Id of controlserum (lanes 1, 4, and 7) or anti-GST-HEB peptide-1 serum (lanes 2,5, and 8) or 5 pl of anti-E12 serum preabsorbed with HEB (lanes 3,6, and 9) was added to the binding mixtures, which were thenanalyzed by EMSA.
CD4-3C, were detected with the CD4-3 probe in EMSA withboth lymphoid and nonlymphoid cell nuclear extracts. Bind-ing of the CD4-3C factor involves sequences in the 3', butnot the 5', E-box, whereas formation of the CD4-3B complexrequires sequences in both E-boxes. Neither of these com-plexes was reactive with antibodies against HEB or E12/E47. Therefore, although CD4-3B interacts with sequencesalso recognized by CD4-3A, it is unlikely that the two factorshave shared components. On the other hand, it is possiblethat CD4-3B contains CD4-3C in its complex. These findingsare reminiscent of the observation that ubiquitous factorswhich do not contain MyoD, myogenin, or E12/E47-likeproteins interact with the MEF1 site where the heterodimersof the myogenic bHLH proteins and E12/E47-like proteinsbind (33). Since point mutations in the 3' E-box, whichabrogated formation of both CD4-3B and CD4-3C, had verylittle effect on enhancer activity, it is unlikely that thesefactors have a significant role in CD4 regulation. It ispossible, however, that since the binding sites for CD4-3Aand CD4-3B are overlapping, these factors compete witheach other for binding to the 5' E-box. CD4-3B may thusregulate the accessibility of CD4-3A binding to the 5' E-boxand may negatively regulate CD4 expression in develop-ment. Alternatively, these factors may interact with thesame sequences simultaneously through contacts in thedifferent grooves, as was observed with factors regulatingthe human beta interferon promoter (52). Further character-
ENHANCER WT Ml M2 M3 M4 M5 M6 M7
*~~S..
1 2 3 4 5 6 7 8 9
FIG. 7. Point mutations in the 5' E-box of the CD4-3 site abolishenhancer activity. Minimal enhancer fragments containing the wild-type (WT) sequence or mutations in the CD4-3 site were inserted atthe 5' end ofpCD4CAT, which contains the CD4 promoter upstreamof the CAT gene. The sequences of mutated CD4-3 sites were asshown in Fig. 2A. These plasmids (lanes 2 to 9) or the enhancerlesspCD4CAT plasmid (lane 1) was transfected into 1200M cells. Thenormalized CAT activities of each sample are as follows: 1, 1.0; 2,120; 3, 0.9; 4, 1.0; 5, 47; 6, 2.2; 7, 29; 8, 0.9; and 9, 24.
ization of CD4-3B and CD4-3C is necessary to understandthe potential significance of these factors in CD4 generegulation.Although CD4-3A was detected in nuclear extracts from
B-cell lines as well as in those from T-cell lines, the CD4enhancer functioned predominantly in T cells in both tran-sient transfection (47) and transgenic mouse systems (28). Inprevious studies, we identified two additional nuclear factorbinding sites within the CD4 enhancer, CD4-1 and CD4-2(Fig. 1). Deletion of the region containing both CD4-1 andCD4-2 sites abolished enhancer activity, although mutationsin these sites that eliminated formation of DNA-nuclearprotein complexes had only a small detrimental effect onenhancer activity (47). Taken together, these observationssuggest that factors other than those observed in the CD4-3Acomplex are required for T-cell-specific activity. It is not yetclear whether these factors are those that form complexeswith the CD4-1, CD4-2, and CD4-3 sites. The CD4-1 site,like CD4-3, contains an E-box motif (Fig. 1). The results of
pE: Id [S] pE: Id [A]
10 15 10 15 (GJg)
S
1 2 3 4 5 6FIG. 8. Overexpression of Id protein inhibits CD4 enhancer
activity. 1200M cells were transfected with pCD4CAT (lane 1) orpCD4CAT/CD4 enhancer (lanes 2 to 6) and either the sense (pE:Id[S]) or antisense (pE:Id[A]) Id construct. In the control (lane 2),carrier DNA (pBluescript vector) was used, as described in Mate-rials and Methods. The normalized CAT activities of each sampleare as follows: 1, 1.0; 2, 77; 3, 4.3; 4, 2.0; 5, 37; and 6, 43.
MOL. CELL. BIOL.
4 0.

CD4 ENHANCER AND bHLH PROTEINS 5627
the competition assays indicated that both CD4-3A andCD4-3C factors could bind to the CD4-1 site, although theCD4-3A factor bound to this site less efficiently than to theCD4-3 sequence (Fig. 2B, lanes 2 and 6). Recently, thepre-B- and T-cell-specific high-mobility-group (HMG) familyfactor that binds to the CD4-2 site, LEF, was shown to haveDNA bending activity (19). It was proposed that, in theTCR-a enhancer, this factor facilitates the interaction of twoflanking protein-DNA complexes. In the CD4 enhancer, theLEF binding site is similarly situated between two transcrip-tion factor binding sites (Fig. 1). Binding of LEF to theCD4-2 site may thus bend the DNA to facilitate an interac-tion between the nuclear protein complexes binding to theCD4-1 and CD4-3 sites.
Expression of the CD4 gene is unusually regulated duringT-cell development. It is positively regulated in the transi-tion of immature CD4- CD8- to CD4+ CD8+ thymocytesand then apparently negatively regulated in a subset ofdouble-positive cells that yield CD4- CD8+ cytotoxic Tcells. Evidence of apparent transcriptional silencing of CD4in the mature CD8+ cells comes from transgenic studies inwhich the CD4 enhancer, linked to the CD4 promoter,directed expression of a CD4 cDNA-genomic minigene in allT cells (12a). In contrast, a similar combination of the CD4enhancer and promoter directed subset-specific expressionof a CD4 genomic construct, suggesting that a transcriptionalsilencer, activated in CD8+ cells, acts on a discrete genomicsequence adjacent to the CD4 promoter (46a). During thetransition from CD4- CD8- to CD4+ CD8+ cells, the CD4enhancer appears to up-regulate CD4 gene transcription,most likely by interacting with multiple transcription factorsincluding HEB, E12-like protein, and LEF. Further differ-entiation may involve activation of additional factors thatmodulate the interactions of the CD4 enhancer/promoterwith its transcription factors, resulting in the down-regula-tion of CD4 gene expression in the CD4- CD8+ lineage.Regulation of the binding of bHLH factors to the CD4enhancer may play a central role during this process ofdifferentiation.
ACKNOWLEDGMENTS
We thank J. Allison and J. Lenz for providing the 1200M, C6VLB,and SL3B cell lines; R. Grosschedl for the PD36 and M12 cell lines;R. Benezra and T. Kadesch for the plasmids expressing Id and ITF2cDNAs, respectively; and C. Murre for the plasmid expressingE12R and the antiserum against E12. We are especially grateful toM. A. Blanar for providing the fusion protein of the HLH motif fromshPAN-2 and for much patient advice. We also thank M. Hill, N.Killeen, and S. Robbins for their advice; R. Grosschedl, R. Benezra,and N. Killeen for critical reading of the manuscript; and G. Mosleyfor excellent secretarial help.
This work was supported by a grant from the National Institutesof Health (Al 23513). D.R.L. is an investigator of the HowardHughes Medical Institute.
REFERENCES1. Aldrich, C. J., R. E. Hammer, S. Jones-Youngblood, U. Koszi-
nowski, L. Hood, I. Stroynowski, and J. Forman. 1991. Negativeand positive selection of antigen-specific cytotoxic T lympho-cytes affected by the a3 domain of MHC I molecules. Nature(London) 352:718-721.
2. Aronheim, A., H. Ohisson, C. W. Park, T. Edlund, and M. D.Walker. 1991. Distribution and characterization of helix-loop-helix enhancer-binding proteins from pancreatic d cells andlymphocytes. Nucleic Acids Res. 19:3893-3899.
3. Benezra, R., R. L. Davis, D. Lockshon, D. L. Turner, and H.Weintraub. 1990. The protein Id: a negative regulator of helix-loop-helix DNA binding proteins. Cell 61:49-59.
4. Biggs, J., E. V. Murphy, and M. A. Israel. 1992. A human Id-likehelix-loop-helix protein expressed during early development.Proc. Natl. Acad. Sci. USA 89:1512-1516.
5. Blanar, M. A., and W. J. Rutter. 1992. Interaction cloning:identification of a helix-loop-helix zipper protein that interactswith c-Fos. Science 256:1014-1018.
6. Cabrera, C. V., and M. C. Alonso. 1991. Transcriptional acti-vation by heterodimers of the achaete-scute and daughterlessgene products of Drosophila. EMBO J. 10:2965-2973.
7. Cammarota, G., A. Scheirle, B. Takacs, D. M. Doran, R. Knorr,W. Bannwarth, J. Guardiola, and F. Sinigaglia. 1992. Identifi-cation of a CD4 binding site on the P2 domain of HLA-DRmolecules. Nature (London) 356:799-801.
8. Campuzano, S., and J. Modolell. 1992. Patterning of the Dro-sophila nervous system: the achaete-scute gene complex.Trends Genet. 8:202-208.
9. Chakraborty, T., T. J. Brennan, L. Li, D. Edmondson, and E. N.Olson. 1991. Inefficient homooligomerization contributes to thedependence of myogenin on E2A products for efficient DNAbinding. Mol. Cell. Biol. 11:3633-3641.
10. Christy, B. A., L. K Sanders, L. F. Lau, N. G. Copeland, N. A.Jenkins, and D. Nathans. 1991. An Id-related helix-loop-helixprotein encoded by a growth factor-inducible gene. Proc. Natl.Acad. Sci. USA 88:1815-1819.
11. Connolly, J. M., T. H. Hansen, A. L. Ingold, and T. A. Potter.1990. Recognition by CD8 on cytotoxic T lymphocytes isablated by several substitutions in the class I a3 domain: CD8and the T-cell receptor recognize the same class I molecule.Proc. Natl. Acad. Sci. USA 87:2137-2141.
12. Cordle, S. R., E. Henderson, H. Masuoka, P. A. Weil, and R.Stein. 1991. Pancreatic ,3-cell-type-specific transcription of theinsulin gene is mediated by basic helix-loop-helix DNA-bindingproteins. Mol. Cell. Biol. 11:1734-1738.
12a.Crooks, M. E. C., R W. Karr, and D. R. Lttman. Unpublishedresults.
13. Davis, C. B., N. Kifleen, M. E. C. Crooks, D. Raulet, and D. RLittman. 1993. Evidence for a stochastic mechanism in thedifferentiation of mature subsets of T lymphocytes. Cell 73:237-247.
14. Davis, R L., and H. Weintraub. 1992. Acquisition of myogenicspecificity by replacement of three amino acid residues fromMyoD into E12. Science 256:1027-1030.
15. Doyle, C., and J. L. Strominger. 1987. Interaction between CD4and class II MHC molecules mediates cell adhesion. Nature(London) 330:256-259.
16. Ellmeler, W., A. Aguzzi, E. Kleiner, R. Kurzbauer, and A.Welth. 1992. Mutually exclusive expression of a helix-loop-helixgene and N-myc in human neuroblastomas and in normaldevelopment. EMBO J. 11:2563-2571.
17. Fung-Leung, W. P., M. W. Schllham, A. Rahemtulla, T. M.Kundig, M. Vollenweider, J. Potter, W. van EwUk, and T. W.Mak. 1991. CD8 is needed for development of cytotoxic T cellsbut not helper T cells. Cell 65:443-449.
18. German, M. S., M. A. Blanar, C. Nelson, L. G. Moss, and W. J.Rutter. 1991. Two related helix-loop-helix proteins participatein separate cell-specific complexes that bind the insulin en-hancer. Mol. Endocrinol. 5:292-299.
19. Giese, K, J. Cox, and R Grosschedl. 1992. The HMG domain oflymphoid enhancer factor 1 bends DNA and facilitates assemblyof functional nucleoprotein structures. Cell 69:185-195.
20. Glaichenhaus, N., N. Shastri, D. R. Lttman, and J. M. Turner.1991. Requirement for association of p561k with CD4 in antigen-specific signal transduction in T cells. Cell 64:511-520.
21. Grosschedl, R., and D. Baltimore. 1985. Cell-type specificity ofimmunoglobulin gene expression is regulated by at least threeDNA sequence elements. Cell 41:885-897.
22. Henthorn, P., M. Kiledjian, and T. Kadesch. 1990. Two distincttranscription factors that bind the immunoglobulin enhancerFE5/KE2 motif. Science 247:467-470.
23. Hu, J.-S., E. N. Olson, and R. E. Kingston. 1992. HEB, ahelix-loop-helix protein related to E2A and ITF2 that canmodulate the DNA-binding ability of myogenic regulatory fac-tors. Mol. Cell. Biol. 12:1031-1042.
VOL. 13, 1993

5628 SAWADA AND LIITMAN
24. Ingold, A. L., C. Landel, C. Knall, G. A. Evans, and T. A.Potter. 1991. Co-engagement of CD8 with the T cell receptor isrequired for negative selection. Nature (London) 352:721-723.
25. Janeway, C. A. 1992. The T cell receptor as a multicomponentsignalling machine: CD4/CD8 coreceptors and CD45 in T cellactivation. Annu. Rev. Immunol. 10:645-674.
26. Jen, Y., H. Weintraub, and R. Benezra. 1992. Overexpression ofId protein inhibits the muscle differentiation program: in vivoassociation of Id with E2A proteins. Genes Dev. 6:1466-1479.
27. Killeen, N., A. Moriarty, H. S. Teh, and D. R. Littman. 1992.Requirement for CD8-major histocompatibility complex class Iinteraction in positive and negative selection of developing Tcells. J. Exp. Med. 176:89-97.
28. Killeen, N., S. Sawada, and D. R. Lttman. 1993. Regulatedexpression of human CD4 rescues T-helper cell development inmice lacking expression of endogenous CD4. EMBO J. 12:1547-1553.
29. Klein, E. S., D. M. Simmons, L. W. Swanson, and M. G.Rosenfeld. 1993. Tissue-specific RNA splicing generates anankyrin-like domain that affects the dimerization and DNA-binding properties of a bHLH protein. Genes Dev. 7:55-71.
30. Koland, J. G., K. M. O'Brien, and R. A. Cerione. 1990.Expression of epidermal growth factor receptor sequences as E.coli fusion proteins: applications in the study of tyrosine kinasefunction. Biochem. Biophys. Res. Commun. 166:90-100.
31. Konig, R., L.-Y. Huang, and R. N. Germain. 1992. MHC class IIinteraction with CD4 mediated by a region analogous to theMHC class I binding site for CD8. Nature (London) 356:796-798.
32. Kreider, B. L., R. Benezra, G. Rovera, and T. Kadesch. 1992.Inhibition of myeloid differentiation by the helix-loop-helixprotein Id. Science 255:1700-1702.
33. Lassar, A. B., R. L. Davis, W. E. Wright, T. Kadesch, C. Murre,A. Voronova, D. Baltimore, and H. Weintraub. 1991. Functionalactivity of myogenic HLH proteins requires hetero-oligomeriza-tion with E12/E47-like proteins in vivo. Cell 66:305-315.
33a.Littman, D. R. Unpublished results.34. Littman, D. R. 1987. The structure of the CD4 and CD8 genes.
Annu. Rev. Immunol. 5:561-584.35. Luckow, B., and G. Schutz. 1987. CAT constructions with
multiple unique restriction sites for the functional analysis ofeukaryotic promoters and regulatory elements. Nucleic AcidsRes. 15:5490.
36. Murre, C., P. S. McCaw, and D. Baltimore. 1989. A new DNAbinding and dimerization motif in immunoglobulin enhancerbinding, daughterless, MyoD, and myc proteins. Cell 56:777-783.
37. Murre, C., P. S. McCaw, H. Vaessin, M. Caudy, L. Y. Jan, Y. N.Jan, C. V. Cabrera, J. N. Buskin, S. D. Hauschka, A. B. Lassar,H. Weintraub, and D. Baltimore. 1989. Interaction betweenheterologous helix-loop-helix proteins generate complexes thatbind specifically to a common DNA sequence. Cell 58:537-544.
38. Murre, C., A. Voronova, and D. Baltimore. 1991. B-cell- andmyocyte-specific E2-box-binding factors contain E12/E47-likesubunits. Mol. Cell. Biol. 11:1156-1160.
39. Nielsen, A. L., N. Pallisgaard, F. S. Pedersen, and P. J0rgensen.1992. Murine helix-loop-helix transcriptional activator proteinsbinding to the E-box motif of the Akv murine leukemia virusenhancer identified by cDNA cloning. Mol. Cell. Biol. 12:3449-3459.
40. O'Rourke, A. M., and M. F. Mescher. 1992. Cytotoxic T-lym-phocyte activation involves a cascade of signalling and adhesionevents. Nature (London) 358:253-255.
41. O'Rourke, A. M., J. Rogers, and M. F. Mescher. 1990. Acti-vated CD8 binding to class I protein mediated by the T-cellreceptor results in signalling. Nature (London) 346:187-189.
42. Ow, D. W., K. V. Wood, M. DeLuca, J. R. de Wet, D. R.Helinski, and S. H. Howell. 1986. Transient and stable expres-sion of the firefly luciferase gene in plant cells and transgenicplants. Science 234:856-859.
43. Parnes, J. R. 1989. Molecular biology and function of CD4 andCD8. Adv. Immunol. 44:265-311.
44. Rahemtulla, A., W. P. Fung-Leung, M. W. Schilham, T. M.Kundig, S. R. Sambhara, A. Narendran, A. Arabian, A. Wake.
ham, C. J. Paige, R. M. Zinkernagel, R. G. Miller, and T. W.Mak. 1991. Normal development and function of CD8' cells butmarkedly decreased helper cell activity in mice lacking CD4.Nature (London) 353:180-184.
45. Rudd, C. E., J. M. Trevillyan, J. D. Dasgupta, L. L. Wong, andS. F. Schlossman. 1988. The CD4 receptor is complexed indetergent lysates to a protein-tyrosine kinase (pp58) from hu-man T lymphocytes. Proc. Natl. Acad. Sci. USA 85:5190-5194.
46. Salter, R. D., R. J. Benjamin, P. K. Wesley, S. E. Buxton, T. P.Garrett, C. Clayberger, A. M. Krensky, A. M. Norment, D. R.Littman, and P. Parham. 1990. A binding site for the T-cellco-receptor CD8 on the alpha 3 domain of HLA-A2. Nature(London) 345:41-46.
46a.Sawada, S., N. Killeen, and D. R. Littman. Unpublished results.47. Sawada, S., and D. R. Littman. 1991. Identification and charac-
terization of a T-cell-specific enhancer adjacent to the murineCD4 gene. Mol. Cell. Biol. 11:5506-5515.
48. Schreiber, E., P. Matthias, M. M. Muller, and W. Schaflher.1989. Rapid detection of octamer binding proteins with 'mini-extracts', prepared from a small number of cells. Nucleic AcidsRes. 17:6419.
49. Shaw, A. S., K. E. Amrein, C. Hammond, D. F. Stern, B. M.Sefton, and J. K. Rose. 1989. The lck tyrosine protein kinaseinteracts with the cytoplasmic tail of the CD4 glycoproteinthrough its unique amino-terminal domain. Cell 59:627-636.
50. Shieh, S.-Y., and M.-J. Tsai. 1991. Cell-specific and ubiquitousfactors are responsible for the enhancer activity of the ratinsulin II gene. J. Biol. Chem. 266:16708-16714.
51. Sun, X.-H., N. G. Copeland, N. A. Jenkdns, and D. Baltimore.1991. Id proteins Idl and Id2 selectively inhibit DNA binding byone class of helix-loop-helix proteins. Mol. Cell. Biol. 11:5603-5611.
52. Thanos, D., and T. Maniatis. 1992. The high mobility groupprotein HMG I(Y) is required for NF-KB-dependent virus in-duction of the human IFN-j gene. Cell 71:777-789.
53. Travis, A., A. Amsterdam, C. Belanger, and R. Grosschedl.1991. LEF-1, a gene encoding a lymphoid-specific protein withan HMG domain, regulates T-cell receptor a enhancer function.Genes Dev. 5:880-894.
54. Turner, J. M., M. H. Brodsky, B. A. Irving, S. D. Levin, R. M.Perlmutter, and D. R. Littman. 1990. Interaction of the uniqueN-terminal region of tyrosine kinase pS6lck with cytoplasmicdomains of CD4 and CD8 is mediated by cysteine motifs. Cell60:755-765.
55. Veillette, A., M. A. Bookman, E. M. Horak, and J. B. Bolen.1988. The CD4 and CD8 T cell surface antigens are associatedwith the internal membrane tyrosine-protein kinase p56Ick. Cell55:301-308.
56. Voronova, A., and D. Baltimore. 1990. Mutations that disruptDNA binding and dimer formation in the E47 helix-loop-helixprotein map to distinct domains. Proc. Natl. Acad. Sci. USA87:47224726.
57. Waterman, M. L., W. H. Fischer, and K. A. Jones. 1991. Athymus-specific member of the HMG protein family regulatesthe human T cell receptor Ca enhancer. Genes Dev. 5:656-669.
58. Weintraub, H., R. Davis, S. Tapscott, M. Thayer, M. Krause, R.Benezra, T. K Blackwell, D. Turner, R. Rupp, S. Hollenberg, Y.Zhuang, and A. Lassar. 1991. The myoD gene family: nodalpoint during specification of the muscle cell lineage. Science251:761-766.
59. Weintraub, H., V. J. Dwarki, I. Verma, R. Davis, S. Hollenberg,L. Snider, A. Lasser, and S. J. Tapscott. 1991. Muscle-specifictranscriptional activation by MyoD. Genes Dev. 5:1377-1386.
60. Wilson, R. B., M. Kiledjian, C.-P. Shen, R Benezra, P. Zwollo,S. M. Dymecki, S. V. Desiderio, and T. Kadesch. 1991. Repres-sion of immunoglobulin enhancers by the helix-loop-helix pro-tein Id: implications for B-lymphoid-cell development. Mol.Cell. Biol. 11:6185-6191.
61. Xu, H., and D. R. Littman.A kinase-independent function of lckin potentiating antigen-specific T cell activation. Cell, in press.
62. Zhang, Y., J. Babin, A. L. Feldhaus, H. Singh, P. A. Sharp, andM. Bina. 1991. HTF4: a new human helix-loop-helix protein.Nucleic Acids Res. 19:4555.
MOL. CELL. BIOL.


















