
A 23,000-yr pollen record from Lake Euramoo, Wet Tropics of NE Queensland,...
14
A 23,000-yr pollen record from Lake Euramoo, Wet Tropics of NE Queensland, Australia Simon G. Haberle * Resource Management in Asia-Pacific Program, Research School of Pacific and Asian Studies, Australian National University, Canberra, ACT 0200, Australia Received 17 January 2005 Abstract A new extended pollen and charcoal record is presented from Lake Euramoo, Wet Tropics World Heritage rainforest of northeast Queensland, Australia. The 8.4-m sediment core taken from the center of Lake Euramoo incorporates a complete record of vegetation change and fire history spanning the period from 23,000 cal yr B.P. to present. The pollen record is divided into five significant zones; 23,000–16,800 cal yr B.P., dry sclerophyll woodland; 16,800 –8600 cal yr B.P., wet sclerophyll woodland with marginal rainforest in protected pockets; 8600–5000 cal yr B.P., warm temperate rainforest; 5000–70 cal yr B.P., dry subtropical rainforest; 70 cal yr B.P.– A.D 1999, degraded dry subtropical rainforest with increasing influence of invasive species and fire. The process of rainforest development appears to be at least partly controlled by orbital forcing (precession), though more local environmental variables and human activity are also significant factors. This new record provides the opportunity to explore the relationship between fire, drought and rainforest dynamics in a significant World Heritage rainforest region. D 2005 University of Washington. All rights reserved. Keywords: Lake Euramoo; Pollen; Charcoal; Tropical rainforest; Holocene; Atherton Tablelands; Australia Introduction In 1970, the first pollen record from the Australian wet tropical rainforest (Lake Euramoo: Kershaw, 1970) was published and demonstrated that tropical palynology was indeed a viable approach to understanding rainforest dynamics over timescales only imagined in ecological studies. Since then, sediments from volcanic craters in the Atherton Table- lands of northeast Queensland have yielded multi-proxy data sets that have made an important contribution to our understanding of Australian palaeoclimates and rainforest vegetation history (Chen, 1988; Moss and Kershaw, 2000; Kershaw et al., 2002; Turney et al., 2004). One of the most outstanding attributes of these data is that they present a picture of a highly dynamic landscape, sensitive to both climate change and human activity on timescales ranging from millennia to decades (Haberle et al. in preparation; Turney et al., 2004). The late Quaternary vegetation history of the Atherton Tablelands is characterized by the expansion of rainforest taxa during warm and wet interglacial periods and contraction of these same taxa, to as yet unknown refugial locations, during cool and dry glacial periods (Kershaw, 1994). The rate at which these changes take place and the nature of succession from one vegetation community to another has been examined in greatest detail over the late glacial transition. This period is represented in five pollen records from the Atherton Tablelands and shows that the transition from sclerophyll woodland to rainforest began as early as 11,500 cal yr B.P., took from 400 to 1000 yr to complete and was regionally concluded by 7000 cal yr B.P. (Hiscock and Kershaw, 1992). An increase in effective precipitation during the late glacial transition is considered the primary driver of vegetation change (Kershaw et al., 2002), though local factors including development of organic soils and fire frequencies (either natural or anthropogenic) may have contributed to the apparent inhibition of rainforest advance during the early Holocene. Rainforest reach their maximum extent on the Atherton Tablelands by the around 7000 cal yr B.P. under a climate 0033-5894/$ - see front matter D 2005 University of Washington. All rights reserved. doi:10.1016/j.yqres.2005.08.013 * Fax: +61 2 6125 1635. E-mail address: [email protected]. Quaternary Research 64 (2005) 343 – 356 www.elsevier.com/locate/yqres
Transcript of A 23,000-yr pollen record from Lake Euramoo, Wet Tropics of NE Queensland,...

lsevier.com/locate/yqres
Quaternary Research 64
A 23,000-yr pollen record from Lake Euramoo,
Wet Tropics of NE Queensland, Australia
Simon G. Haberle *
Resource Management in Asia-Pacific Program, Research School of Pacific and Asian Studies,
Australian National University, Canberra, ACT 0200, Australia
Received 17 January 2005
Abstract
A new extended pollen and charcoal record is presented from Lake Euramoo, Wet Tropics World Heritage rainforest of northeast Queensland,
Australia. The 8.4-m sediment core taken from the center of Lake Euramoo incorporates a complete record of vegetation change and fire history
spanning the period from 23,000 cal yr B.P. to present. The pollen record is divided into five significant zones; 23,000–16,800 cal yr B.P., dry
sclerophyll woodland; 16,800–8600 cal yr B.P., wet sclerophyll woodland with marginal rainforest in protected pockets; 8600–5000 cal yr B.P.,
warm temperate rainforest; 5000–70 cal yr B.P., dry subtropical rainforest; 70 cal yr B.P.–A.D 1999, degraded dry subtropical rainforest with
increasing influence of invasive species and fire.
The process of rainforest development appears to be at least partly controlled by orbital forcing (precession), though more local environmental
variables and human activity are also significant factors. This new record provides the opportunity to explore the relationship between fire,
drought and rainforest dynamics in a significant World Heritage rainforest region.
D 2005 University of Washington. All rights reserved.
Keywords: Lake Euramoo; Pollen; Charcoal; Tropical rainforest; Holocene; Atherton Tablelands; Australia
Introduction
In 1970, the first pollen record from the Australian wet
tropical rainforest (Lake Euramoo: Kershaw, 1970) was
published and demonstrated that tropical palynology was
indeed a viable approach to understanding rainforest dynamics
over timescales only imagined in ecological studies. Since
then, sediments from volcanic craters in the Atherton Table-
lands of northeast Queensland have yielded multi-proxy data
sets that have made an important contribution to our
understanding of Australian palaeoclimates and rainforest
vegetation history (Chen, 1988; Moss and Kershaw, 2000;
Kershaw et al., 2002; Turney et al., 2004). One of the most
outstanding attributes of these data is that they present a picture
of a highly dynamic landscape, sensitive to both climate
change and human activity on timescales ranging from
millennia to decades (Haberle et al. in preparation; Turney et
al., 2004).
0033-5894/$ - see front matter D 2005 University of Washington. All rights reserv
doi:10.1016/j.yqres.2005.08.013
* Fax: +61 2 6125 1635.
E-mail address: [email protected].
The late Quaternary vegetation history of the Atherton
Tablelands is characterized by the expansion of rainforest taxa
during warm and wet interglacial periods and contraction of
these same taxa, to as yet unknown refugial locations, during
cool and dry glacial periods (Kershaw, 1994). The rate at which
these changes take place and the nature of succession from one
vegetation community to another has been examined in greatest
detail over the late glacial transition. This period is represented
in five pollen records from the Atherton Tablelands and shows
that the transition from sclerophyll woodland to rainforest
began as early as 11,500 cal yr B.P., took from 400 to 1000 yr
to complete and was regionally concluded by 7000 cal yr B.P.
(Hiscock and Kershaw, 1992). An increase in effective
precipitation during the late glacial transition is considered
the primary driver of vegetation change (Kershaw et al., 2002),
though local factors including development of organic soils and
fire frequencies (either natural or anthropogenic) may have
contributed to the apparent inhibition of rainforest advance
during the early Holocene.
Rainforest reach their maximum extent on the Atherton
Tablelands by the around 7000 cal yr B.P. under a climate
(2005) 343 – 356
www.e
ed.

S.G. Haberle / Quaternary Research 64 (2005) 343–356344
regime characterized by the establishment of modern day
temperatures, possibly higher precipitation than present (or at
least reduced seasonality in precipitation: Kershaw and Nix,
1988), and low frequency of fires (every 230 yr or so: Chen,
1988). Mid–Late Holocene rainforest composition was far
from stable with high variability evident in rainforest pollen
assemblages recorded during this period (Hiscock and Ker-
shaw, 1992). Around 5000–6000 cal yr B.P. sustained rises in
the emergent conifer Agathis and the early successional taxa
Urticaceae/Moraceae, point to an opening up of the rainforest
canopy perhaps due to reduced precipitation or increased
disturbance. Rainforest disturbance may have had its origin in
natural climate variability (El Nino-related storm and drought
events: Gagan et al., 2004) or through increased intensity of
human activity in rainforest environments (Haberle and David,
2004; Horsfall, 1987).
This paper describes a new core taken from the centre of
Lake Euramoo in 1999, which is part of a study directed at
understanding the origin and role of disturbance within tropical
rainforests through fine resolution palaeoecology. The new
core represents a complete record of sediment accumulation
from the Last Glacial Maximum to the present (post-European
period), which is unlike any other palaeoecological record from
the Atherton Tablelands (Hiscock and Kershaw, 1992).
The site
Lake Euramoo is located near the western edge of the World
Heritage Wet Tropics Bioregion of North East Queensland (718
m above sea level, 17-10V S, 146-38V E) on the Tertiary uplifted
highlands of the Atherton Tablelands (Fig. 1) and within area
that contains Australia’s most significant expanse of tropical
rainforest. Lake Euramoo is a maar described as an ovate
double explosion crater that formed during the Late Pleistocene
period (Kershaw, 1970), with a relatively small catchment area
of ¨4500 m2 and no inflow or outflow channels. The lake is
warm monomictic with a water depth averaging around 20 m in
Figure 1. Map of northeast Queensland showing locat
the northern basin and 16 m in the southern basin, though there
are seasonal fluctuations in water depth of between 2 and 3 m
(Timms, 1976).
An annual average rainfall of 1500 mm is estimated to occur
at Lake Euramoo with a distinctly dry winter rainfall regime as
more than 60% falls between the months of January to March
(Kershaw, 1970). The lake lies at the lower rainfall end of a
steep east to west rainfall gradient. The dominant source of
precipitation is the Southeast Trades interspersed with occa-
sional Northwesterly monsoonal flows and associated tropical
cyclones which also bring high but infrequent rainfall events
during the austral summer months when the intertropical
convergence zone (ITCZ) is at its most southerly extent
(Godfred-Spenning and Reason, 2002). During El Nino
episodes, a northward movement of the ITCZ and a northeast-
ward migration of the South Pacific convergence zone result in
a significant decrease in summer precipitation (typically 150–
300 mm below seasonal average) over the region (Dai and
Wigley, 2000). Mean daily maximum and minimum tempera-
tures are around 25.9- and 14.4-C and frosts occur infrequently
during the austral winter months at times of weak trade winds
and low cloud cover.
The terrestrial vegetation surrounding Lake Euramoo is a
remnant of moist sub-montane rainforest surrounded by
previously cleared land that within the last 50 years has been
either planted with endemic (Araucaria cunninghamii) and
exotic confers or has undergone secondary colonization
towards re-establishing rainforest. Over 100 species of rain-
forest tree and shrub have been recorded within 100 m of Lake
Euramoo (Kershaw personal communication). Typical moist
sub-montane rainforest species found near Lake Euramoo
include members of the Arailiaceae (e.g., Polyscias australi-
ana, Schefflera actinophylla), Araucariaceae (e.g., Agathis
robusta), Moraceae (e.g., Ficus sp.) Elaeocarpaceae (e.g.,
Elaeocarpus grandis), Euphorbiaceae (e.g., Aleutites mollu-
cana, Macaranga sp.), Myrtaceae (e.g., Austromyrtus sp.,
Eugenia cormiflora), and Rubiaceae (e.g., Flindersia bray-
ion of wet tropical rainforests and Lake Euramoo.

S.G. Haberle / Quaternary Research 64 (2005) 343–356 345
leyana, Euodia bonwickii). The nearby (<1 km) drier
vegetation communities include tall open forest (sclerophyll
woodland) dominated by Casuarina sp. and Eucalyptus sp. At
higher altitude, cool lower montane rainforest is increasingly
dominated by members of families such as the Cunoniaceae
(e.g., Caldcluvia australiensis, Schizomeria whitei) Podocar-
paceae (e.g., Podocarpus sp., Prumnopitys sp.) and Rhamna-
ceae (e.g., Alphitonia sp.).
The vegetation around the lake margin is made of
conspicuous zones of aquatic plant communities governed by
water depth and seasonal fluctuations in water level (Kershaw,
1978). Rainforest lianas (e.g., Parsonia sp.) intertwine with tall
swamp grass Phragmites sp. around the shallow lake edge.
Hibiscus sp. and Ludwigia sp. become more common outside
the influence of canopy shade and liana growth in the deeper
(less than 1 m water depth) swamp margin. A zone of rooted
emergent aquatics and floating vegetation mats is characterized
by Cyclosorus gongylodes and Oenanthe sp., with Blechnum
sp. and Elaeocharis equiseteum becoming common on less
stable floating mats. As water depth increases to greater than 1
m, there is a transition from rooted emergent aquatics to
floating aquatics, mainly Nymphoides sp. which can occur as
much as 30 m from the lake shore.
Methods
Fieldwork was conducted in the southern hemisphere
winter of 1999. Parallel cores were taken from the center of
the lake with a modified Livingstone corer with the mud–
water interface section taken with a 1-m plastic piston corer
and extruded at 1-cm intervals on site. Each core was
described in detail and scanned by a magnetic susceptibility
core-scanning loop at 1-cm intervals that identifies changes in
the abundance of ferrimagnetic minerals, which was used as
an indication of changing erosional input to the lake. The
cores were also sampled at 1-cm intervals for loss on ignition
analysis (at 550-C) to identify changes in the amount of
combustible organic matter. Sampling for pollen analysis
ranged from 2- to 8-cm intervals. Chronological control is
provided by using 210Pb dating and AMS 14C analysis of bulk
organic and pollen preparation samples extracted from the
central portion of the sediment core (Table 1, calibrated years
before present, where 0 cal yr B.P. = 1950 A.D, Stuiver et al.,
1998).
Pollen analysis follows the standard KOH, HF, HCl and
acetolysis method described by Bennett and Willis (2001).
Samples from these lake deposits are generally organic-rich
with high pollen concentrations calculated volumetrically with
the addition of an exotic spike (Lycopodium clavatum). Pollen
identification and nomenclature follows regional reference
collections currently held at the School of Geography and
Environmental Science, Monash University and the Depart-
ment of Archaeology and Natural History at the Australian
National University. Ecological information on the taxa
represented in the pollen diagrams can be found in Kershaw
(1970, 1971, 1975, 1983). Additional taxa identified in this
analysis include Melalueca comp. (Myrtaceae; a wet scler-
ophyll woodland tree in poorly drained soils), and Colocasia
(Araceae, herb of poorly drained soils). Pollen counts are
expressed as percentages of the total pollen sum (excluding
pollen of aquatic vascular plants and spores of ferns and fern
allies), which reaches a minimum of 300 in all samples.
Microscopic charcoal particle accumulation rates were
calculated as concentrations of particles 10–125 Am encoun-
tered during pollen counting compared to exotic Lycopodium
spike divided by the sediment accumulation rate. Macroscopic
charcoal particle accumulation rates were calculated as
concentrations of particles >125 Am determined at contiguous
1-cm intervals by treating 4 cm3 sediment with KOH, mild
bleach (6% Potassium Hypochlorite) overnight, sieved
through a 125-Am sieve and counting total particles encoun-
tered under a low-powered (¨10�) stereomicroscope. This
provides basic data on the abundance of charcoal of two
different size fractions, the finer of which is most likely to be
derived from long distance as well as local fires and the
coarser fraction is likely to be derived from more local fire
events.
Numerical zonation, rarefaction and Principle Components
Analysis (PCA) were performed with only major taxa whose
pollen or spore values exceeded 5% at least once. Numerical
zonation employed optimal splitting by sum-of-squares anal-
ysis to partition the stratigraphically constrained fossil pollen
assemblage data into significantly different pollen zones (after
Bennett, 1996). Rarefaction analysis provides an estimate of
the diversity of pollen types or palynological richness of each
sample (Birks and Line, 1992). PCA is used to reduce the
pollen and spore data to a two-dimensional plot and the
resulting data set is displayed as a plot for samples and taxa
(Birks and Gordon, 1985). All numerical analyses have been
implemented within psimpoll, a program for plotting pollen
data, developed by Bennett (1994).
Results and discussion
Lithology, sediment analysis and chronology
A total of 840 cm of lake muds and clays were recovered
from the centre of the northern basin of Lake Euramoo from
1600 to 2440 cm bws (below water surface). The basal core
encountered solid bedrock at a depth of 2440 cm bws.
Overlying the bedrock are sediments composed of grey-green
massive silty clays to 2365 cm bws that are characterized by
low LOI values and peak magnetic susceptibility values (Fig.
2). Between 2365 to 2265 cm bws, the sediments are black lake
muds with occasional bands (<1 cm thick) of light brown
clayey muds decreasing in frequency up-core. Magnetic
susceptibility values show a decreasing trend to minimum
values at around 2320 cm bws. The LOI values increase up
core with a sharp peak (60% organics) from 2320 to 2290 cm
bws, followed by a reversal to lower LOI (30% organics)
between 2290 to 2265 cm bws. Above 2265 cm bws to the mud
water interface at 1600 cm bws, the sediments are uniformly
dark brown lake muds with LOI values around 80–90%. A
zone of coarse detritus and wood is encountered at 2000 to

Table 114C Radiocarbon AMS sample results from the ANSTO radiocarbon lab
ANSTO code Sample Type and ID
(cm below water level)
y(13C) per mil Conventional 14C age Calibrated age, cal yr B.P.
(CALIB v4.4, 2j error)14C yr B.P. 1y error
OZE682 EU 1695–1696 �27.9 570 50 490–570 (80%)
590–630 (20%)
OZE683 EU 1790–1790 �30.1 1320 40 1080–1110 (5%)
1120–1160 (9%)
1170–1290 (86%)
OZE684 EU 1857–1858 �31.8 2410 40 2320–2490 (85%)
2630–2710 (15%)
OZE685 EU 1925–1926 �33.5 3510 40 3630–3840 (100%)
OZE686 EU 2040–2041 �31.7 4090 40 4420–4630 (92%)
4760–4800 (8%)
OZE687 EU 2157–2158 �33.9 6210 50 6900–7210 (99%)
7220–7230 (1%)
OZH310 EU 2201–2202 �31.1 5690 50 6640–6580 (9%)
6570–6400 (86%)
6370–6350 (4%)
6330–6320 (1%)
OZH311 *EU 2219–2220 �31.7 7860 60 8980–8910 (11%)
8900–8880 (4%)
8870–8830 (7%)
8810–8540 (78%)
OZH312 EU 2246–2247 �21.3 9400 80 11,070–10,940 (11%)
10,860–10,830 (2%)
10,810–10,800 (1%)
10,790–10,400 (86%)
OZH313 EU 2255–2256 �25.5 9530 60 11,110–10,660 (99%)
10,620–10,610 (1%)
OZH688 EU 2263–2264 �28.5 9640 60 11,190–11,050 (40%)
11,040–10,780 (60%)
OZH314 *EU 2270–2271 �23.2 9800 90 11,550–11,500 (4%)
11,450–11,390 (3%)
11,360–11,060 (81%)
10,950–10,840 (9%)
10,830–10,760 (3%)
OZH315 *EU 2275–2276 �29.1 10,130 60 12,280–12,220 (3%)
12,130–11,540 (83%)
11,520–11,400 (10%)
11,390–11,340 (4%)
OZH316 Eu 2282–2283 �27.2 10,640 60 12,950–12,600 (76%)
12,500–12,340 (24%)
OZE838 EU 2292–2293 �24.0 11,100 60 12,670–12,720 (3%)
12,870–13,190 (96%)
13,310–13,340 (1%)
OZH317 *EU 2303–2304 �26.6 9110 60 10,470–10,460 (2%)
10,430–10,180 (98%)
OZH318 EU 2315–2316 24.6 11,230 60 13,760–13,700 (3%)
13,460–13,000 (97%)
OZE689 EU 2341–2342 �26.4 12,600 70 14,240–15,540 (100%)
OZH319 EU 2361–2362 �25.0 15,820 120 19,550–18,270 (100%)
OZH320 EU 2389–2390 �24.8 11,310 70 13,310–13,070 (100%)
OZH321 EU 2413–2414 �18.6 18,960 160 22,940–22,170 (100%)
OZE690 EU 2430–2435 �24.1 19,130 90 21,930–23,430 (100%)
All samples are derived from bulk organic detritus with the exception of 4 pollen preparation samples*. Calibration results from CALIB v4.4 (Stuiver and Reimer,
1993). The midpoint of calendar yr range marked in bold are used in age-depth model calculations (3 samples excluded from the age-depth model are not in bold).
S.G. Haberle / Quaternary Research 64 (2005) 343–356346
1940 cm bws and is considered to be debris from a fallen tree
or floating root mat and is excluded from the analysis.
Core chronology is based on a total of 22 AMS 14C dates
on bulk organic detritus and pollen preparation samples
(Table 1, derived from 5-mm-thick sample extractions).
Fourteen sediment samples from the upper 46 cm of the
core were dried overnight at 75-C, finely ground and
analysed for 210Pb by ANSTO (Lucas Heights, Sydney).
Sedimentation rates for the top 46 cm were derived from the
analysis of 210Pb activity using the constant initial concen-
tration (CIC) model (Appleby and Oldfield, 1983) and
produced a basal 210Pb age of A.D 1880 at 1646 cm bws.
The final age-depth model adopted for this study (Fig. 2) is
derived by linear interpolation between a mud–water inter-

Figure 2. Lake Euramoo stratigraphy, sediment description, 14C age yr B.P., water content, loss on ignition and age-depth model.
S.G. Haberle / Quaternary Research 64 (2005) 343–356 347
face age of A.D 1999, the first appearance of exotic pollen
types taxa (Lantana, Mimosa, Pinus and Plantago, details in
Haberle et al. in prep), the 210Pb profile, and the 14C age
determinations. Three AMS 14C dates were excluded from the
age-depth profile on the basis of poorest fit with the linear
interpolation and may be the result of down-core contamina-
tion with younger carbon.
Pollen and charcoal record
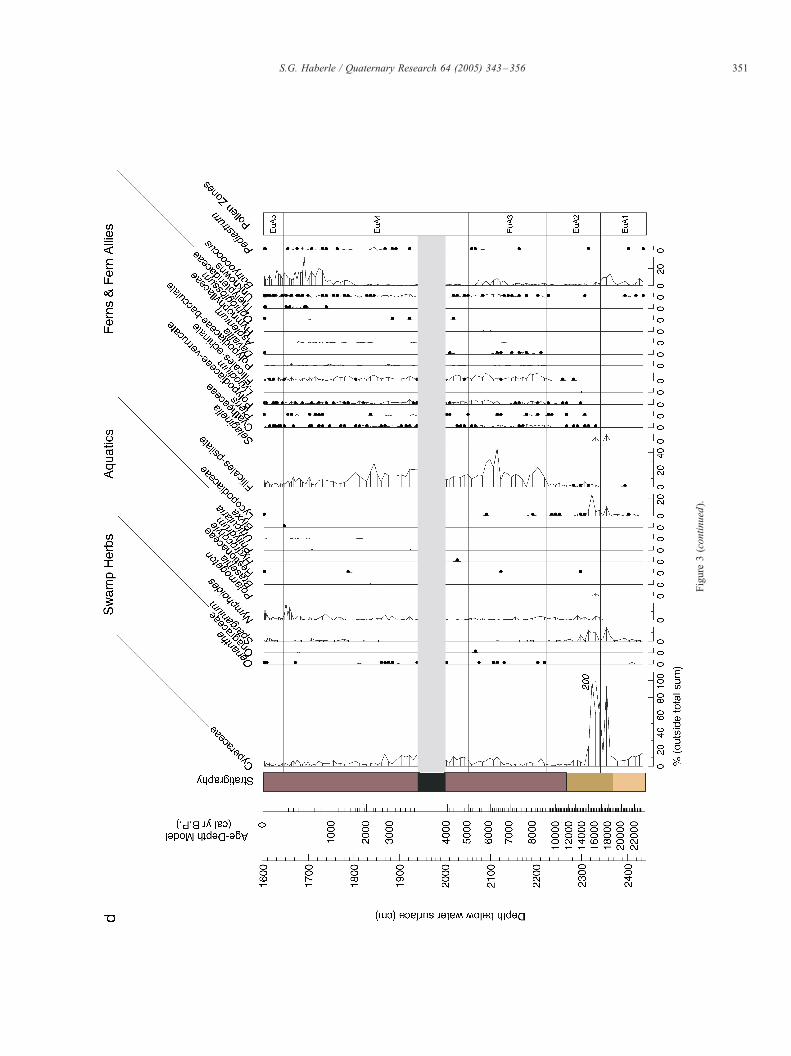
The pollen record for Lake Euramoo is presented in Figures
3a–d and is divided into five zones that are described below. A
summary interpretation of each zone is presented in Table 2.
Lake Euramoo 23,000–16,800 cal yr B.P.
(Zone Eu-A1, 2432–2340 cm bws)
Grasses and the sclerophyll woodland taxa, mainly
Casuarina, Myrtaceae (dominated by Melalueca comp.) and
to a lesser extent Callitris dominate the diagram forming
>95% of the pollen sum. Melalueca comp. and the grasses
alternate peak values with Casuarina. There is no significant
presence of rainforest taxa and the palynological richness
reaches a minimum in this zone. Locally, the presence of
Cyperaceae, increasing sharply towards the top of the zone,
and aquatic taxa such as Sparganium, Botryococcus and
Pediastrum suggest that a swampy margin developed around
a small open water body. The Cyperaceous swamp expanded
markedly after about 18,000 cal yr B.P. and included a greater
presence of Lycopodiaceae, Sparganium and the aquatic fern
Selaginella. Fires appear to be infrequent throughout this
zone.
Lake Euramoo 16,800–8700 cal yr B.P.
(Zone Eu-A2, 2340–2220 cm bws)
High values for sclerophyll taxa are maintained though
there is an increase in the importance of Mrytaceous taxa,
including the wet sclerophyll Eucalyptus types. Callitris
increases to its highest values in the record. Grasses and
Compositae decrease with minor increases recorded in Alter-
nathera and Liliaceae between 16,000 and 10,600 cal yr B.P.
The first appearance of 36 rainforest taxa, dominated by
Urticaceae/Moraceae, is recorded in this zone which is

S.G. Haberle / Quaternary Research 64 (2005) 343–356348
reflected in a doubling of the palynological richness by 15,500
cal yr B.P. The rainforest gymnosperms Agathis and Podo-
carpus appear for the first time by 15,500 cal yr B.P. but then
are absent between 12,600 and 9600 cal yr B.P. Rainforest
taxa as a whole make up between 10% and 20% of the pollen
sum between 16,800 and 11,500 cal yr B.P., but then show a
reversal to dry sclerophyll taxa dominance (mainly Casuarina)
between 11,500 and 9600 cal yr B.P. Regional pollen dispersal
from the lowland mangrove swamps is also incorporated into
the Lake Euramoo sediments and may reflect increased
convection or more extensive mangrove forests along the
coastal margins as sea level stabilizes during the early
Holocene (Clark and Guppy, 1988). The Cyperaceae swamp
dominates the lake margin until 15,500 cal yr B.P. but then
becomes a less significant feature of the lake edge vegetation
after this time. Ferns also begin to become important in this
zone. Fire is much more prominent in the landscape after
16,800 cal yr B.P. The fire record is dominated by fine
charcoal particles derived from regional as well as local
(probably grass dominated) fires. By around 11,500 cal yr B.P.
the coarser charcoal fraction begins to increase perhaps in
response to reduced grassy understorey and the development
of a more dominant woody understorey.
Figure 3. Lake Euramoo pollen diagram. (a) Summary pollen diagram with rarefactio
rates (2 size fractions: 10–125 Am and >125 Am). (b) % diagram of herbaceous, scle
<1%). (c) % diagram of minor rainforest taxa included within the total pollen s
palynomorphs excluded from the total pollen sum.
Lake Euramoo 8700–5000 cal yr B.P.
(Zone Eu-A3, 2220–2050 cm bws)
At 8700 cal yr B.P. there is a rapid shift from sclerophyll
to rainforest dominance in the pollen spectrum. Casuarina,
Myrtaceae, Callitris and the grass percentages all fall to low
values, reaching a minimum by 7300 cal yr B.P., as the
total rainforest taxa contribution to the pollen sum rise
sharply to around 80–90%. Urticaceae/Moraceae, Mallotus/
Macaranga, Trema, Cunoniaceae, Elaeocarpus and ferns
(mainly Filicales-psilate type) all show sustained increase in
percentages through this zone. Podocarpus shows an initial
increase and then declines by around 7300 cal yr B.P. A
further 19 rainforest taxa make their first appearance in this
zone, though the palynological richness reaches a high level
at the base of this zone that is maintained for the rest of the
record, reflecting a balance of taxa losses as well as gains
during this transition. The swamp herbs Colocasia, Halor-
agis and an undetermined Umbelliferae are incorporated into
the local cyperaceous swamp. The charcoal record is
dominated by the coarse charcoal fraction through to about
7300 cal yr B.P. possibly reflecting continued burning of the
rainforest-sclerophyll woodland margin during the early
Holocene. Total charcoal reaches a minimum between about
n analysis and principal components analysis results, and charcoal accumulation
rophyll woodland and rainforest taxa included within the total pollen sum (dot =
um (dot = <1%). (d) % diagram of swamp herbs, aquatics, ferns and other

Figure
3(continued).
S.G. Haberle / Quaternary Research 64 (2005) 343–356 349

Figure
3(continued).
S.G. Haberle / Quaternary Research 64 (2005) 343–356350
Figure
3(continued).
S.G. Haberle / Quaternary Research 64 (2005) 343–356 351

Table 2
Interpretation of Lake Euramoo pollen zones and equivalent zones represented in the Lake Euramoo edge core (Kershaw, 1970)
Pollen zone Age cal yr B.P. Interpretation of pollen spectra Equivalent pollen
zone Kershaw 1970
Eu-A1 23,000–16,800 Dry sclerophyll woodland dominated by fire-sensitive Casuarina with a grassy understorey. A sedge
swamp surrounds a small permanent open water body with local Melalueca swamp around the lake
edge margins.
Not evident
Eu-A2 16,800–8700 Wet sclerophyll woodland with marginal rainforest in protected pockets or forming rainforest mosaics
with drier woodland formations. Fire becoming increasingly prevalent with increase in fire-promoting
woody flora.
Zone 1
Eu-A3 8700–5000 Lower montane rainforest with relative dominance by Cunoniaceae and Urticaceae/Moraceae with fire
virtually absent.
Zone 2, 3
Eu-A4 5000–70 Sub-montane rainforest dominated by Elaeocarpaceae, Agathis and Trema with fire increasingly
prevalent.
Zone 4, 5
Eu-A5 AD 1880–1999 Degraded sub-montane rainforest with increasing influence of invasive species and fire. Surface samples only
S.G. Haberle / Quaternary Research 64 (2005) 343–356352
7300 and 6300 cal yr B.P. at the same time as rainforest
taxa attain maximum percentage representation of close to
100% of the pollen sum. The coarse charcoal fraction then
makes a sustained increase after 6300 cal yr B.P. and is
accompanied by rises in Trema a pioneer rainforest taxon.
Lake Euramoo 5000–70 cal yr B.P.
(Zone Eu-A4, 2050–1646 cm bws)
This zone is separated from the previous zone by a sustained
rise in Elaeocarpus, closely associated with slight rises in
Agathis and Mallotus/Macaranga. Cunoniaceae generally
decrease while Urticaceae/Moraceae, Cunoniaceae and Podo-
Figure 4. Principle components analysis of the Lake Euramoo pollen data from the K
paper. The analysis included only taxa with >5% representation common to both rec
1 = 63% and axis 2 = 8%. Sample symbols distinguish pollen zones from each site a
dashed (lake edge core) arrow.
carpus maintain constant percentage values in this zone. Fine
and coarse charcoal particles are present throughout with a
series of peaks in course charcoal deposition recorded from
4000 cal yr B.P. onwards and a rise in accumulation rate
between 2700 and 1200 cal yr B.P. At around 1200 cal yr B.P.,
a further rise in Elaeocarpus is accompanied by slight
increases in sclerophyll taxa and a decrease in Agathis,
Mallotus/Macaranga and Trema. A decrease in Cyperaceae
and an increase in the aquatics, Sparganium, Nymphoides and
Botryococcus, suggest a return to more open water conditions
at this time. The appearance of the swamp fern Thelypter-
idaceae (most likely Cyclosorus gongylodes), a weedy fern
ershaw (1970) lake edge core and the lake centre core taken in 1999 from this
ords (Rut. = Rutaceae/Arailiaceae, Cunon. = Cunoniaceae) with eigenvalue axis
nd the direction of change is indicated by grey solid (lake centre core) and grey

S.G. Haberle / Quaternary Research 64 (2005) 343–356 353
recorded in disturbed swampland across the Pacific (Powell,
1982; Mueller-Dombois and Fosberg, 1998) around 1000 cal yr
B.P. suggests the floating mat habitat may have expanded as
well in the last millennia.
Lake Euramoo A.D 1880–1999
(Zone Eu-A5, 1646–1600 cm bws)
This zone is separated from the preceding zone by the
appearance of exotic species Lantana, Mimosa, Plantago,
Pinus, Solonaceae and Rumex, introduced during the period
of European settlement, beginning in the early A.D 1880s.
Sclerophyll taxa maintain constantly low values whereas
several rainforest taxa show marked declines including
Urticaceae/Moraceae, Elaeocarpus and Agathis, most likely
as a result of removal of the taxa due to selective logging
during the 19th Century (Birtles, 1988). Increases are
recorded in Rutaceae/Araliaceae, Trema, Cunoniaceae, Sloa-
nea and Eugenia. Charcoal particle accumulation rates return
to high values comparable with those recorded in the early
Holocene, though there is little apparent impact on the total
rainforest taxa percentage of the pollen sum.
Comparison between lake edge and lake centre pollen record
The PCA ordination (Fig. 4) incorporates two fossil pollen
datasets from Lake Euramoo. The diagram compares the
extended Lake Euramoo pollen record from the lake centre
(23,000 cal yr B.P. to present) with the lake edge core
collected by Kershaw (1970) that spanned the Holocene from
11,000 to 1500 cal yr B.P. and was also divided into five
pollen zones. Axis 1 contrasts the sclerophyll woodland taxa,
Casuarina and Gramineae, with rainforest taxa and appears to
be related primarily to precipitation. The intermediate position
of Eucalyptus taxa along axis 1 reflects the widespread
association of Eucalyptus with both woodland and rainforest
habitats. Axis 2 explains much less variance though it appears
to distinguish mid–late Holocene dominance of sub-montane
rainforest gymnosperms and secondary rainforest angiosperms
such as Mallotus/Macaranga and Trema, from the early-mid
Holocene lower montane rainforest taxa dominated by
Cunoniaceae and Urticaceae/Moraceae. Axis 2 may relate to
disturbance factors such as increased burning in the mid–late
Holocene or a decrease in dry season precipitation proposed
by Kershaw and Nix (1988) bioclimatic analysis of the Lake
Euramoo edge core record.
The two records are readily comparable through PCA
analysis (Fig. 4, Table 2) though the lake edge core samples
are overall more negative on axis 2 than the lake centre core
samples. One explanation may be that the catchment area
for pollen in the edge core is more sensitive to sub-canopy
and surface wash dispersed pollen including taxa such as
Trema, Mallotus/Macaranga and Podocarpus and the local
swamp Gramineae. The lake centre core is likely to be more
sensitive to pollen wind dispersed from the local emergent
and canopy taxa such as the Cunoniaceae and Urticaceae/
Moraceae, as well as regionally dispersed taxa such as
Eucalyptus.
Discussion
An extended sclerophyll woodland phase at Lake Euramoo
The new pollen record from Lake Euramoo extends the old
record by around 12,000 cal yr B.P. to include the Last Glacial
Maximum. Kershaw (1970: 797) originally hypothesized that
the persistence of sclerophyll woodland in the early Holocene
part of the record was a consequence of recent (early
Holocene) volcanic activity at the site and that the earliest
vegetation then evident from the pollen record represented
pioneer vegetation establishing on immature soils. The new
core from the lake centre clearly refutes the possibility of early
Holocene volcanic activity from the Euramoo crater, placing a
minimum age for crater formation of >23,000 cal yr B.P. There
is no indication in the pollen record that vegetation went
through successional phases of establishment on maturing
basalt soils around the site after 23,000 cal yr B.P. As
sediments accumulated over basalt bedrock the regional
vegetation remains dominated by fire-sensitive Casuarina with
a grassy understorey, though the alternating peaks in Mela-
lueca comp. and the grasses with Casuarina, possibly reflect
millennial-scale shifts in regional soil moisture or salinity
during this period (Crowley, 1994). Swamp vegetation persists
locally throughout this early period.
Pollen and sediment evidence from Lynch’s Crater (Turney
et al., 2004; Kershaw, 1994), sea surface temperature
reconstructions from the Coral Sea (Barrows and Juggins, in
press; Anderson et al., 1989) and fluvial geomorphology from
NE Queensland (Thomas et al., 2001) all point to conditions
on the Atherton Tablelands at this time being much drier (up to
50% reduced precipitation) and cooler (2-–5-C) than today.
The low levels of charcoal preserved in the Lake Euramoo
sediments suggests that the maintenance of the sclerophyll
woodland community around Lake Euramoo was most
strongly associated with the climatic controls of low precip-
itation rather than a high burning frequency. Similar conditions
are reflected in the Lynch’s Crater (zone L2 lower section) and
Strenekoff’s Crater (zone S2 lower sample) record, where
samples closest to the LGM in these records show relatively
high Casuarina–low Eucalyptus pollen percentages accom-
panied by low charcoal densities (Kershaw, 1994). This is at
odds with reconstructions of biomass burning further to the
north in the equatorial highlands of New Guinea where
cumulative charcoal records from wet montane forest/grass-
land settings suggest a high rate of charcoal production
between 20,000 and 10,000 cal yr B.P. The New Guinea
records have been interpreted as resulting from greater
variability in rainfall patterns, during the late glacial period,
enhancing the potential for frequent fires (Haberle et al., 2001).
The combined effect of relatively lower annual precipitation
and the lower biomass of sclerophyll woodland in the Atherton
Tablelands may have been sufficient to reduce charcoal
deposition during the late glacial period in Lake Euramoo
and it is not until increased woodiness, in response to increased
precipitation of the late glacial transition phase, that charcoal
production also increased.

S.G. Haberle / Quaternary Research 64 (2005) 343–356354
Four phases of rainforest development
The transition from sclerophyll woodland to rainforest as
recorded at Lake Euramoo can be considered to have taken
9500 yr, with the initial indication that rainforest was re-
invading the Atherton Tablelands from glacial refugia begin-
ning at 16,800 cal yr B.P. and being completed by 7300 cal yr
B.P. The development of rainforest appears to occur in four
phases.
The first phase begins at 16,800 cal yr B.P., when a limited
suite of rainforest angiosperm and gymnosperm taxa and wet
sclerophyll taxa first appear near the site, possibly as insipient
rainforest patches forming mosaics with wet sclerophyll
woodland. This is almost 4000 yr earlier than similar changes
noted in other Atherton Tablelands records (Walker and Chen,
1987; Hiscock and Kershaw, 1992) and may reflect closer
proximity of Lake Euramoo to glacial rainforest refugia, more
favorable geology (basalts) for early establishment of rain-
forest, or the wet swampy lake margins providing restricted
refugia for rainforest taxa as precipitation and temperature
increased during the late glacial transition. Increased organic
accumulation and biomass burning are associated with this
phase, which supports the suggestion that a rise in woody plant
biomass may have been a factor in rising charcoal particle
accumulation rates in the sediments.
The second phase spans the period from 12,600 to 9600 cal
yr B.P. and is marked by the disappearance of a number of
rainforest taxa from the record, including Agathis and
Podocarpus, and a return to peak representation of Casuarina
in place of wet scleophyll woodland. This may represent a
reversal towards drier climatic conditions, however, rainforest
taxa continue to make initial appearances in the record
suggesting that taxa disappearances may represent shifts in
canopy dominance through competitive advantage rather than a
reversal in climatic conditions necessarily restricting rainforest
advancement into the area.
The third phase occurs between 9600 and 8700 cal yr B.P.
when the maximum rate of increase in rainforest, led by taxa
typical of lower montane rainforest, is recorded at Lake
Euramoo. This is most likely to represent the local establish-
ment of closed canopy rainforest under the influence of
increasing precipitation at the site. This occurs within a 900-
yr period and is comparable to the period of time recorded at
other sites on the Atherton Tablelands where the transition
from sclerophyll woodland to rainforest dominance has been
estimated to have taken between about 400 and 1000 yr on the
Atherton Tablelands (Hiscock and Kershaw, 1992).
The final phase of rainforest development involves the
gradual exclusion of sclerophyll woodland prior to the peak
development of lower montane rainforest at 7300 cal yr B.P.
Local dominance of rainforest was achieved despite the
persistent recurrence of fires that appear to have maintained
local patches of sclerophyll woodland and may have retarded
the encroachment of rainforest onto the site.
The non-synchronous and non-linear nature of the scler-
ophyll to rainforest transition recorded at five sites on the
Tablelands points to local as well as regional environmental
variables influencing the development of rainforest through
time. The apparent contradiction that the most intensive fire
period occurs during the phase of most rapid increase in fire
sensitive rainforest suggests that fire may have become
decoupled from the natural climate regime and was at least
partially controlled by aboriginal fire practices. The motivation
for maintaining open sclerophyll woodlands in an ever
encroaching rainforest landscape may have stemmed from the
need to maintain open habitats that were so prevalent during
the late glacial period for hunting, mobility and to exploit
woodland plant diversity for food procurement (Hill and Baird,
2003; Horsfall, 1987).
Rainforest dynamics during the mid–late Holocene
Around 7300 to 6300 cal yr B.P. rainforest achieved its
maximum extent across the Atherton Tablelands under condi-
tions of higher than present wet season temperatures and higher
dry season precipitation (Kershaw and Nix, 1988). This
effectively would have reduced annual climate seasonality
resulting in low potential for dry-season burning of sclerophyll
woodland and limited moisture loss from rainforest habitats
reduced its vulnerability to fire. Reduced seasonality may have
been partially driven by orbital forcing according to the
Milankovitch theory, which predicts a minimum in Southern
Hemisphere summer insolation from ¨12,000 to 9000 cal yr
B.P., with an increase toward the present (Berger and Loutre,
1991). In the Lake Euramoo record, the maximum rainforest
extent of rainforest occurs at least 2000 yr after the insolation
minimum. One explanation for this apparent lag in vegetation
response to Milankovich forcing lies in the strong influence of
southeast trade winds on precipitation in the Atherton Table-
lands today. In the past precipitation at Lake Euramoo would
have been influenced by the proximity of the coastline. Rising
sea levels during the late-glacial transition flooded the
continental shelf adjacent to the Atherton Tablelands as well
as flooding the Torres Strait, between northern Australia and
New Guinea, with current conditions only being attained by
around 8000 to 7000 cal yr B.P. (Woodroffe et al., 2000). The
combined impact of reduced continentality, increased moisture
convection over the Torres Strait leading to greater southern
penetration of the ITCZ during the southern summer, may have
been the primary driver increasing delivery of rainfall to Lake
Euramoo after 8700 cal yr B.P. and during the mid Holocene
period.
The evidence for a sustained onset of increased burning
after 6300 cal yr B.P. and the appearance for major infrequent
(every 250–1000 yr) peaks of charcoal after 4000 cal yr B.P. is
accompanied by evidence for greater disturbance of the
rainforest, suggesting that the charcoal source was not
restricted to patches of sclerophyll woodland (currently within
1 km from the site) but may indicate fires occurring within the
rainforest resulting in an opening up of the canopy. Similar fire
frequencies are suggested for the Lake Barrine catchment area
during the period of rainforest encroachment (Chen, 1988) and
this may represent a general fire return time for rainforests in
the region. Increased rainforest disturbance may have been

S.G. Haberle / Quaternary Research 64 (2005) 343–356 355
driven by overall drier conditions or increased rainfall
seasonality (Kershaw and Nix, 1988). In addition, the potential
influence of intensified El Nino activity after ¨5000 cal yr B.P.
that has been suggested from eastern (Sandweiss et al., 1996;
Rodbell et al., 1999) and western Pacific (Haberle et al., 2001;
Hayne and Chappell, 2001) proxy records and would likely
have a significant impact on rainforest dynamics in the
Atherton Tablelands region. The shift in the Lake Euramoo
pollen assemblage towards greater representation of short-lived
taxa such as Mallotus/Macaranga, after 5000 cal yr B.P. may
well be indicative of an overall increase in ecosystem turnover
rates under the influence of more frequent El Nino-related
disturbance events. Alternatively, or at least overlaying these
climatic factors there may have been a greater intensity of
occupation and resource management by aboriginal popula-
tions after about 4000 cal yr B.P. facilitated through a
broadening of the range of foodstuffs consumed and the
commencement of toxic plant exploitation (Horsfall, 1987;
Haberle and David, 2004). Destruction of the swamp forest on
Lynch’s Crater at about 5000 cal yr B.P. has also been linked to
aboriginal exploitation and intensified use of fire at this time
(Hiscock and Kershaw, 1992). The slight rise in total charcoal
accumulation rate and increased frequency of major charcoal
peaks between 2500 and 1500 cal yr B.P. may also point to a
period of intensified burning activity near Lake Euramoo. The
relative contribution of climate change and human activity to
these changes remains a complex question that ultimately may
not be resolved if we assume an interdependence of human
response to environmental conditions (Haberle and Chepstow-
Lusty, 2000; Haberle and David, 2004).
The pollen record of the last few hundred years at Lake
Euramoo, missing in the original lake edge core (Kershaw,
1970), details the first appearances of exotic pollen (Lantana,
Mimosa and Pinus/Plantago) and the dramatic rise in charcoal
accumulation rate in the lake sediments provide a clear
stratigraphic marker for European occupation and land use in
the region. The fine-resolution record of the upper 100 cm
shows that the present composition of rainforest in this region
has been significantly altered over the last 120 yr, most notably
evident through the reduction of key taxa in the pollen record,
including Agathis and Podocarpus (Figs. 3b and 4), both
exploited for the timber industry during the early 19th Century
(Birtles, 1988). The most significant changes are recorded at
the onset of European occupation between about A.D 1880 and
1920, through clearance and burning activities. These changes
were rapid and their impact appears to have persisted through
the last century with no sign of recovery to a pre-European
assemblage despite nearly 50 yr of fire management and
rainforest conservation.
Conclusions
The extended pollen record from Lake Euramoo represents
a complete record of vegetation change from the LGM to
present incorporating the transition from sclerophyll woodlands
to rainforest. Detailed analysis of the transition including a
high-resolution contiguous record of charcoal particle accu-
mulation rates suggests that the progression of rainforest
expansion at the beginning of the Holocene was neither
synchronous with climate change nor unidirectional implying
a much more complex model for rainforest development
incorporating external (regional climatic) as well as local
(topographic, anthropogenic) factors. The process of rainforest
expansion takes about 9500 yr, beginning around 16,800 cal yr
B.P. and proceeding through a non-linear four-phase develop-
ment until maximum rainforest extent is achieved by 7300 cal
yr B.P. Fire appears to play a major role in the development of
early rainforest cover, both through general retardation of
rainforest expansion and by providing competitive advantage
to fire-tolerant and pioneer species. Tropical biomass burning
appears to be at least partly controlled by orbital forcing
(precession), though biomass change and human activity are
also significant factors. Shifts towards competitive dominance
of taxa adapted to drought/fire (e.g., Agathis, Mallotus/
Macaranga, Podocarpus and Trema) are most prevalent in
the Lake Euramoo catchment after 5000 cal yr B.P., which may
be a response to the intensification of El Nino-related climate
variability and/or increased human activity within north
Queensland tropical rainforest.
This new record provides the opportunity to explore the
relationship between fire, drought and rainforest dynamics in a
significant World Heritage rainforest region. That vegetation
changes and fire episodes are not always synchronous with
regional climate change through time supports the notion that
climate thresholds and ecological or vegetation thresholds are
not necessarily the same, nor once they are crossed are they
necessarily reversible (Maslin, 2004).
Acknowledgments
The author thanks the editors of this volume Hermann
Behling and Robert Marchant. Thanks also to Peter Kershaw
for encouraging me to re-visit Lake Euramoo and providing
data and advice along the way. Nick Porch, Henk Heijnis,
Sophia Dimitriadis and Ainsley Noakes helped with fieldwork
and Timme Donders made helpful comments on the manu-
script. Permission to core at Lake Euramoo was granted
through the Queensland Parks and Wildlife Service, Queens-
land Government. Radiocarbon and Lead-210 analysis was
completed at the Australian Nuclear Science and Technology
Organisation funded through an AINSE Grant 00/60. The
research was completed while the author was supported by an
Australian Research Council QEII Fellowship at Monash
University and the Australian National University.
References
Anderson, D.M., Prell, W.L., Barratt, N.J., 1989. Estimates of sea surface
temperature in the Coral Sea at the last glacial maximum. Paleoceanography
4, 615–627.
Appleby, P.G., Oldfield, F., 1983. Assessment of 210Pb from sites with varying
sediment accumulation rates. Hydrobiologia 103, 29–35.
Barrows, T.T., Juggins, S., 2005. Sea-surface temperatures around the
Australian margin and Indian Ocean during the Last Glacial Maximum.
Quaternary Science Reviews 24, 1017–1047.

S.G. Haberle / Quaternary Research 64 (2005) 343–356356
Bennett, K.D., 1994. Fpsimpoll_ version 2.23: a C program for analysing
pollen data and plotting pollen diagrams. INQUA Commission for the
study of the Holocene: working group on data-handling methods.
Newsletter 11, 4–6.
Bennett, K.D., 1996. Determination of the number of zones in a biostrati-
graphical sequence. New Phytologist 132, 155–170.
Bennett, K.D.,Willis, K.J., 2001. Pollen. In: Smol, J.P., Birks, H.J.B., Last, W.M.
(Eds.), Tracking Environmental Change Using Lake Sediments. Terrestrial,
Algal, and Siliceous Indicators, vol. 3. Kluwer, Dordrecht, pp. 5–32.
Berger, A., Loutre, M.F., 1991. Insolation values for the climate of the last 10
million of years. Quaternary Science Reviews 10, 297–317.
Birks, H.J.B., Gordon, A.D., 1985. Numerical Methods in Quaternary Pollen
Analysis. Academic Press, London (317p).
Birks, H.J.B., Line, J.M., 1992. The use of rarefaction analysis for estimating
palynological richness from Quaternary pollen-analytical data. The Holo-
cene 2, 1–10.
Birtles, T., 1988. European interpretation of the Atherton-Evelyn vine scrub of
tropical north Queensland, 1880–1920. In: Dargavel, J., Dixon, K.,
Semple, N. (Eds.), Changing Tropical Forests: Historical Perspectives on
Today’s Challenges in Asia, Australasia and Oceania. Centre for Resource
and Environmental Studies Publication, Australian National University,
Canberra, pp. 197–216.
Chen, Y., 1988. Early Holocene population expansion of some rainforest
trees at Lake Barrine basin, Queensland. Australian Journal of Ecology
13, 225–233.
Clark, R.L., Guppy, J.C., 1988. A transition from mangrove forest to freshwater
wetland in monsoon tropics of Australia. Journal of Biogeography 15,
665–684.
Crowley, G.M., 1994. Quaternary soil salinity events and Australian vegetation
history. Quaternary Science Reviews 13, 15–22.
Dai, A., Wigley, T.M.L., 2000. Global patterns of ENSO-induced precipitation.
Geophysical Research Letters 27, 1283–1286.
Gagan, M.K., Hendy, E.J., Haberle, S.G., Hantoro, W.S., 2004. Post-glacial
evolution of the Indo-Pacific Warm Pool and El Nino-Southern Oscillation.
Quaternary International 118–119, 127–143.
Godfred-Spenning, C.R., Reason, C.J.C., 2002. Interannual variability of
lower-tropospheric moisture transport during the Australian Monsoon.
International Journal of Climatology 22, 509–532.
Haberle, S.G., Chepstow-Lusty, A., 2000. Can climate influence cultural
development?: a view through time. Environment and History 6, 349–369.
Haberle, S.G., David, B., 2004. Climates of change: human dimensions of
Holocene environmental change in low latitudes of the PEPII transect.
Quaternary International 118–119, 165–179.
Haberle, S.G., Hope, G.S., van der Kaars, W.A., 2001. Biomass burning in
Indonesia and Papua New Guinea: natural and human induced fire events in
the fossil record. Palaeogeography, Palaeoclimatology, Palaeoecology 171,
259–268.
Hayne, M., Chappell, J., 2001. Cyclone frequency during the last 5000 years at
Curacoa Island, north Queensland, Australia. Palaeogeography, Palaeocli-
matology, Palaeoecology 168, 207–219.
Hill, R., Baird, A., 2003. Kuku-Yalanji rainforest aboriginal people and
carbohydrate resource management in the Wet Tropics of Queensland,
Australia. Human Ecology 31, 27–52.
Hiscock, P., Kershaw, A.P., 1992. Palaeoenvironments and prehistory of
Australia’s tropical Top End. In: Dodson, J. (Ed.), The Niave Lands:
Prehistory and Environmental Change in Australia and the Southwest
Pacific. Longman Cheshire, Melbourne, pp. 43–75.
Horsfall, N., 1987. Aborigines and toxic north-eastern Queensland rainforest
plants. In: Covacevich, J., Davie, P., Pearn, J. (Eds.), Toxic Plants and
Animals: A Guide for Australia. QueenslandMuseum, Brisbane, pp. 57–63.
Kershaw, A.P., 1970. A pollen diagram from Lake Euramoo, North-East
Queensland, Australia. New Phytologist 69, 785–805.
Kershaw, A.P., 1971. A pollen record from Quincan Crater, north-east
Queensland, Australia. New Phytologist 70, 669–805.
Kershaw, A.P., 1975. Stratigraphy and pollen analysis of Bromfield Swamp,
north-eastern Queensland, Australia. New Phytologist 75, 173–191.
Kershaw, A.P., 1978. The analysis of aquatic vegetation on the Atherton
Tablelands, north-east Queensland, Australia. Australian Journal of Ecology
3, 23–42.
Kershaw, A.P., 1983. A Holocene pollen record from Lynch’s Crater, north-
eastern Queensland, Australia. New Phytologist 94, 669–682.
Kershaw, A.P., 1994. Pleistocene vegetation of the humid tropics of
northeastern Queensland, Australia. Palaeogeography, Palaeoclimatology,
Palaeoecology 109, 399–412.
Kershaw, A.P., Nix, H.A., 1988. Quantitative palaeoclimatic estimates from
pollen using bioclimatic profiles of extant taxa. Journal of Biogeography
15, 589–602.
Kershaw, A.P., van der Kaars, S., Moss, P.T., Wang, X., 2002. Quaternary
records of vegetation, biomass burning, climate and possible human impact
in the Indonesian-Northern Australian region. In: Kershaw, A.P., David, B.,
Tapper, N.J., Penny, D., Brown, J. (Eds.), Bridging Wallace’s Line: The
Environmental and Cultural History and Dynamics of the Southeast Asian-
Australian Region. Catena Verlag, Reiskirchen, Germany, pp. 97–118.
Maslin, M., 2004. Ecological versus climatic thresholds. Science 306,
2197–2198.
Moss, P.T., Kershaw, A.P., 2000. The last glacial cycle from the humid tropics
of Australia: comparison of a terrestrial and marine record. Palaeogeo-
graphy, Palaeoclimatology, Palaeoecology 155, 155–176.
Mueller-Dombois, D., Fosberg, F.R., 1998. Vegetation of the Tropical Pacific
Islands. Springer, New York.
Powell, J.M., 1982. The history of plant use and man’s impact on the
vegetation. In: Gressitt, J.L. (Ed.), Biogeography and Ecology of New
Guinea. Junk, The Hague, pp. 207–227.
Rodbell, D.T., Seltzer, G.O., Anderson, D.M., Abbott, M.B., Enfield, D.B.,
Newman, J.H., 1999. An¨15,000-year record of El Nino-driven alluviation
in southwestern Ecuador. Science 283, 516–520.
Sandweiss, D.H., Richardson III, J.B., Reitz, E.J., Rollins, H.B., Maasch, K.A.,
1996. Geoarchaeological evidence from Peru for a 5000 years B.P. onset of
El Nino. Science 273, 1531–1533.
Stuiver, M., Reimer, P.J., 1993. Extended 14C database and revised CALIB
radiocarbon calibration program. Radiocarbon 35, 215–230.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A.,
Kromer, B., McCormac, F.G., van der Plicht, J., Spurk, M., 1998.
INTCAL98 Radiocarbon age calibration 24,000-0 cal BP. Radiocarbon
40, 1041–1083.
Thomas, M.F., Nott, J., Price, D.M., 2001. Late Quaternary stream sedimen-
tation in the humid tropics: a review with new data from NE Queensland,
Australia. Geomorphology 39, 53–68.
Timms, B.V., 1976. Morphology of Lakes Barrine, Eacham and Euramoo,
Atherton Tableland, North Queensland. Proceedings of the Royal Society of
Queensland 87, 81–84.
Turney, C.S.M., Kershaw, A.P., Clemens, S.C., Branch, N., Moss, P.T., Fifield,
L.K., 2004. Millennial and orbital variations of El Nino/Southern
Oscillation and high-latitude climate in the last glacial period. Nature 428,
306–310.
Walker, D., Chen, Y., 1987. Palynological light on tropical rainforest dynamics.
Quaternary Science Reviews 6, 77–92.
Woodroffe, C.D., Kennedy, D.M., Hopley, D., Rasmussen, C., Smithers,
S.G., 2000. Holocene reef growth in Torres Strait. Marine Geology 170,
331–346.


















