
47–60 Blackwell Munksgaard Rab10 is Involved … Munksgaard Rab10 is Involved in Basolateral...
14
# 2006 The Authors Journal compilation # 2006 Blackwell Publishing Ltd doi: 10.1111/j.1600-0854.2006.00506.x Traffic 2007; 8: 47–60 Blackwell Munksgaard Rab10 is Involved in Basolateral Transport in Polarized Madin–Darby Canine Kidney Cells Sebastian Schuck 1,† , Mathias J. Gerl 1,† , Agnes Ang 2 , Aki Manninen 3 , Patrick Keller 4 , Ira Mellman 2 and Kai Simons 1, * 1 Max Planck Institute of Molecular Cell Biology and Genetics, 01307 Dresden, Germany 2 Department of Cell Biology, Yale University School of Medicine, New Haven, CT 06520, USA 3 Biocenter Oulu, University of Oulu, 90220 Oulu, Finland 4 Meso Scale Discovery, Gaithersburg, MD 20877, USA *Corresponding author: Kai Simons, [email protected] † These authors contributed equally to this work. The sorting of newly synthesized membrane proteins to the cell surface is an important mechanism of cell polar- ity. To identify more of the molecular machinery involved, we investigated the function of the small GTPase Rab10 in polarized epithelial Madin–Darby canine kidney cells. We find that GFP-tagged Rab10 localizes primarily to the Golgi during early cell polarization. Expression of an activated Rab10 mutant inhibits biosynthetic transport from the Golgi and missorts basolateral cargo to the apical membrane. Depletion of Rab10 by RNA interfer- ence has only mild effects on biosynthetic transport and epithelial polarization, but simultaneous inhibition of Rab10 and Rab8a more strongly impairs basolateral sorting. These results indicate that Rab10 functions in trafficking from the Golgi at early stages of epithelial polarization, is involved in biosynthetic transport to the basolateral membrane and may co-operate with Rab8. Key words: basolateral membrane, biosynthetic trans- port, epithelial polarity, Golgi, Rab GTPases Received 2 August 2006; revised and accepted for publi- cation 10 October 2006; published online 21 November 2006 Eukaryotic cells use an elaborate transport system to control the distribution of their proteins and lipids. Once synthesized, these molecules are sent to many different destinations, including the endoplasmic reticulum (ER), the Golgi, endosomes and the plasma membrane. Additional complexity is found in polarized cells, which typically possess distinct surface domains. Epithelial cells, for example, have apical and basolateral membranes, whereas neurons have axonal and somatodendritic do- mains. To generate and maintain such asymmetry, polar- ized cells sort newly synthesized cargo to different parts of their plasma membrane. Biosynthetic sorting therefore is an important determinant of cell polarity (1–3). The trafficking routes and sorting sites for biosynthetic cargo in polarized cells are still being defined. Even in the well-studied epithelial Madin–Darby canine kidney (MDCK) cell line, it is not entirely clear which apical and basolateral proteins travel from the Golgi to the plasma membrane by direct pathways, which proteins reach the surface via endosomes and which proteins use a transcytotic mode of delivery (4–6). In addition, it has long been debated where apical and basolateral cargo is separated, and both the trans Golgi network (TGN) and recycling endosomes have been proposed as major biosynthetic sorting sites (7,8). Although parts of the molecular machinery for polar- ized surface transport are already known, more of the missing players need to be identified to resolve these issues. Rab GTPases are key regulators of membrane trafficking (9,10). Rabs cycle between an inactive GDP-bound and an active GTP-bound state. They are recruited from the cytosol onto cell membranes in the inactive form, are activated by guanine nucleotide exchange factors and then bind a large number of Rab effector proteins. In this way, Rabs co-ordinate the assembly of effector complexes and generate functional membrane domains. Following inacti- vation by GTP hydrolysis, Rabs release their effectors and become available for the next round of activation and effector binding. Rab effector proteins are functionally diverse, which allows Rabs to co-ordinate multiple aspects of membrane trafficking, including transport vesicle for- mation, motility, docking and fusion. Many Rabs have been specifically linked to particular transport pathways and are therefore useful landmarks to map out the intracellular trafficking network. The Rab GTPase Sec4p is essential for post-Golgi traffick- ing in yeast (11). The closest Sec4p homologues in higher organisms are Rab8, Rab10 and Rab13 (12). Rab8 is involved in transport to the basolateral membrane in MDCK cells (13), and Rab13 functions in tight junction assembly (14). The role of Rab10 is less clear. Rab10 has a single isoform, is conserved throughout metazoan evo- lution and is ubiquitously expressed in mouse and human tissues (12,15). The mammalian Rab10 has been found at the late Golgi in fibroblasts (16). More recently, two studies have provided evidence that interfering with Rab10 func- tion leads to defects in endocytosis in polarized cells. In Caenorhabditis elegans, lack of Rab10 impairs endocytic recycling in the intestine (17). In MDCK cells, mutant vari- ants of Rab10 affect early endocytic events (18). Neverthe- less, the homology to Sec4p suggests that Rab10 may also play a role in exocytic trafficking in MDCK cells. Here, we test this hypothesis to gain more insight into the molecular www.traffic.dk 47
Transcript of 47–60 Blackwell Munksgaard Rab10 is Involved … Munksgaard Rab10 is Involved in Basolateral...

# 2006 The Authors
Journal compilation# 2006 Blackwell Publishing Ltd
doi: 10.1111/j.1600-0854.2006.00506.xTraffic 2007; 8: 47–60Blackwell Munksgaard
Rab10 is Involved in Basolateral Transport in PolarizedMadin–Darby Canine Kidney Cells
Sebastian Schuck1,†, Mathias J. Gerl1,†,
Agnes Ang2, Aki Manninen3, Patrick Keller4,
Ira Mellman2 and Kai Simons1,*
1Max Planck Institute of Molecular Cell Biology andGenetics, 01307 Dresden, Germany2Department of Cell Biology, Yale University School ofMedicine, New Haven, CT 06520, USA3Biocenter Oulu, University of Oulu, 90220 Oulu, Finland4Meso Scale Discovery, Gaithersburg, MD 20877, USA*Corresponding author: Kai Simons, [email protected]†These authors contributed equally to this work.
The sorting of newly synthesized membrane proteins to
the cell surface is an important mechanism of cell polar-
ity. To identifymore of themolecular machinery involved,
we investigated the function of the small GTPase Rab10
in polarized epithelial Madin–Darby canine kidney cells.
We find that GFP-tagged Rab10 localizes primarily to
the Golgi during early cell polarization. Expression of an
activated Rab10 mutant inhibits biosynthetic transport
from the Golgi and missorts basolateral cargo to the
apical membrane. Depletion of Rab10 by RNA interfer-
ence has only mild effects on biosynthetic transport
and epithelial polarization, but simultaneous inhibition
of Rab10 and Rab8a more strongly impairs basolateral
sorting. These results indicate that Rab10 functions
in trafficking from the Golgi at early stages of epithelial
polarization, is involved in biosynthetic transport to
the basolateral membrane and may co-operate with
Rab8.
Key words: basolateral membrane, biosynthetic trans-
port, epithelial polarity, Golgi, Rab GTPases
Received 2 August 2006; revised and accepted for publi-
cation 10 October 2006; published online 21 November
2006
Eukaryotic cells use an elaborate transport system to
control the distribution of their proteins and lipids. Once
synthesized, these molecules are sent to many different
destinations, including the endoplasmic reticulum (ER), the
Golgi, endosomes and the plasma membrane. Additional
complexity is found in polarized cells, which typically
possess distinct surface domains. Epithelial cells, for
example, have apical and basolateral membranes,
whereas neurons have axonal and somatodendritic do-
mains. To generate and maintain such asymmetry, polar-
ized cells sort newly synthesized cargo to different parts of
their plasma membrane. Biosynthetic sorting therefore is
an important determinant of cell polarity (1–3).
The trafficking routes and sorting sites for biosynthetic
cargo in polarized cells are still being defined. Even in the
well-studied epithelial Madin–Darby canine kidney (MDCK)
cell line, it is not entirely clear which apical and basolateral
proteins travel from the Golgi to the plasma membrane by
direct pathways, which proteins reach the surface via
endosomes and which proteins use a transcytotic mode
of delivery (4–6). In addition, it has long been debated
where apical and basolateral cargo is separated, and both
the trans Golgi network (TGN) and recycling endosomes
have been proposed as major biosynthetic sorting sites
(7,8). Although parts of the molecular machinery for polar-
ized surface transport are already known, more of the
missing players need to be identified to resolve these issues.
Rab GTPases are key regulators of membrane trafficking
(9,10). Rabs cycle between an inactive GDP-bound and
an active GTP-bound state. They are recruited from the
cytosol onto cell membranes in the inactive form, are
activated by guanine nucleotide exchange factors and then
bind a large number of Rab effector proteins. In this way,
Rabs co-ordinate the assembly of effector complexes and
generate functional membrane domains. Following inacti-
vation by GTP hydrolysis, Rabs release their effectors and
become available for the next round of activation and
effector binding. Rab effector proteins are functionally
diverse, which allows Rabs to co-ordinate multiple aspects
of membrane trafficking, including transport vesicle for-
mation, motility, docking and fusion. Many Rabs have been
specifically linked to particular transport pathways and are
therefore useful landmarks to map out the intracellular
trafficking network.
The Rab GTPase Sec4p is essential for post-Golgi traffick-
ing in yeast (11). The closest Sec4p homologues in higher
organisms are Rab8, Rab10 and Rab13 (12). Rab8 is
involved in transport to the basolateral membrane in
MDCK cells (13), and Rab13 functions in tight junction
assembly (14). The role of Rab10 is less clear. Rab10 has
a single isoform, is conserved throughout metazoan evo-
lution and is ubiquitously expressed in mouse and human
tissues (12,15). The mammalian Rab10 has been found at
the late Golgi in fibroblasts (16). More recently, two studies
have provided evidence that interfering with Rab10 func-
tion leads to defects in endocytosis in polarized cells. In
Caenorhabditis elegans, lack of Rab10 impairs endocytic
recycling in the intestine (17). In MDCK cells, mutant vari-
ants of Rab10 affect early endocytic events (18). Neverthe-
less, the homology to Sec4p suggests that Rab10may also
play a role in exocytic trafficking in MDCK cells. Here, we
test this hypothesis to gain more insight into the molecular
www.traffic.dk 47

machinery for biosynthetic transport in polarized epithelial
cells.
Results
Rab10 localizes primarily to the Golgi in MDCK cells
Rab proteins usually localize to the sites at which they
regulate membrane trafficking events. Mutants of a given
Rab that prefer the inactive GDP-bound state additionally
indicate where membrane recruitment takes place,
whereas mutants that prefer the active GTP-bound state
show where effector binding can occur [e.g. (19,20)]. We
therefore determined the distribution of green fluorescent
protein (GFP)-tagged versions of wild-type canine Rab10,
inactivated Rab10T23N and activated Rab10Q68L in
MDCK–TfR cells. These cells produce the human trans-
ferrin receptor, which facilitates labeling of the recycling
endosomes with fluorescent transferrin. The different
Rab10 constructs were introduced into MDCK–TfR cells
on coverslips by transient transfection and analyzed by
immunofluorescence microscopy. Weakly expressing
cells were chosen for imaging to avoid mislocalization
due to high levels of expression.
GFP-Rab10 was present in perinuclear tubular structures
that overlapped with those positive for giantin, a marker for
the Golgi (Figure 1A). GFP-Rab10 also showed substantial
co-localization with furin, a marker for the TGN, but only
minor overlap with endocytosed transferrin that had been
accumulated in recycling endosomes (Figure 1B). In addi-
tion, no co-localization was observed with the early endo-
some marker EEA1 (data not shown). Activated GFP-
Rab10Q68L co-localized well with furin, but not with
endocytosed transferrin (Figure 1C). Inactivated GFP-
Rab10T23N was in two pools, one that was dispersed
throughout the cytosol and one that coincided with giantin
as well as furin (Figure 1D and data not shown).
These observations indicate that Rab10 localizes primarily
to the Golgi, including the TGN, at least in incompletely
polarizedMDCK cells grown on coverslips. The distribution
of GFP-Rab10T23N and GFP-Rab10Q68L further indicates
that Rab10 is recruited onto Golgi membranes and is also
active at the Golgi. Thus, the localization of the different
Rab10 variants suggests that Rab10 is involved in mem-
brane trafficking events that take place at, or emanate
from, the Golgi.
Activated Rab10 inhibits biosynthetic transport from
the Golgi to plasma membrane
The normal nucleotide cycle of Rabs can often be inhibited
by the expression of inactivated or activated mutant
variants. At sufficiently high levels, inactivated GDP-bound
mutants are thought to sequester guanine nucleotide
exchange factors, so that the endogenous Rab can no
longer be activated (21,22). The effects of activated GTP-
bound mutants may depend on their expression levels.
At low levels, they can support the activity of the endog-
enous Rab (21). At high levels, they can act as inhibitors
(13,23), presumably by sequestering effector proteins and
thus interrupting the cycle of effector binding and release.
To explore the possible role of Rab10 in biosynthetic
transport, we introduced Rab10T23N and Rab10Q68L into
MDCK cells using recombinant adenoviruses to ensure
strong expression. We then analyzed the trafficking of
yellow fluorescent protein (YFP)-tagged temperature-
sensitive vesicular stomatitis virus glycoprotein (VSV-G)
in these cells. VSV-G is a transmembrane protein that can
be reversibly blocked in the ER at 39.58C and is sorted
mostly to the basolateral domain in polarized MDCK cells
at 328C, the permissive temperature for VSV-G transport
(24). Cells grown to confluence on coverslips were in-
fected with adenoviruses encoding red fluorescent protein
(RFP)-tagged Rab10T23N or Rab10Q68L, using conditions
that achieved infection rates close to 100%. Twelve hours
later, they were infected with a second adenovirus,
encoding VSV-G, and incubated at 39.58C for 6 h to
accumulate newly synthesized VSV-G in the ER. Protein
synthesis was blocked with cycloheximide and the tem-
perature was shifted to 328C, thus chasing VSV-G through
the biosynthetic pathway as a synchronous wave (25).
Cells were fixed after various times and analyzed by
immunofluorescence microscopy.
In control cells, VSV-G was found throughout the cytosol
immediately after the shift to 328C, exhibiting a pattern
characteristic of the ER and distinct from the compact
perinuclear distribution of giantin (Figure 2A, top panel).
After 25 min, VSV-G had moved to the Golgi, as indicated
by co-localization with giantin (Figure 2A, middle panel).
After 60 min, the bulk of VSV-G had reached the plasma
membrane (Figure 2A, bottom panel). In contrast, VSV-G
transport was strongly delayed in cells expressing acti-
vated Rab10Q68L. VSV-G accumulated in the ER and was
transported to the Golgi essentially as in control cells
(Figure 2B, top and middle panel). However, it then
persisted in perinuclear giantin-positive structures (Fig-
ure 2B, bottom panel). In addition, the giantin staining often
appeared more fragmented, possibly because the structure
of the Golgi is disturbed by the accumulation of VSV-G as
well as endogenous cargo.
Inactivated Rab10T23N caused no obvious delay of VSV-G
transport. Given the qualitative nature of the microscopic
assay, however, we cannot exclude that a slight delay of
VSV-G transport was missed. In addition, we noticed that
the expression of Rab10T23N, while it was quite strong
and easily detectable in almost all cells by the fluorescent
signal of the RFP tag, was much weaker than that of
Rab10Q68L. A comparison of the mRNA and protein levels
of the two Rab10 variants determined by quantitative
reverse transcriptase polymerase chain reaction (RT–PCR)
and immunoblotting with an anti-RFP antibody indicated
that RabT23N is less stable than Rab10Q68L (data not
48 Traffic 2007; 8: 47–60
Schuck et al.
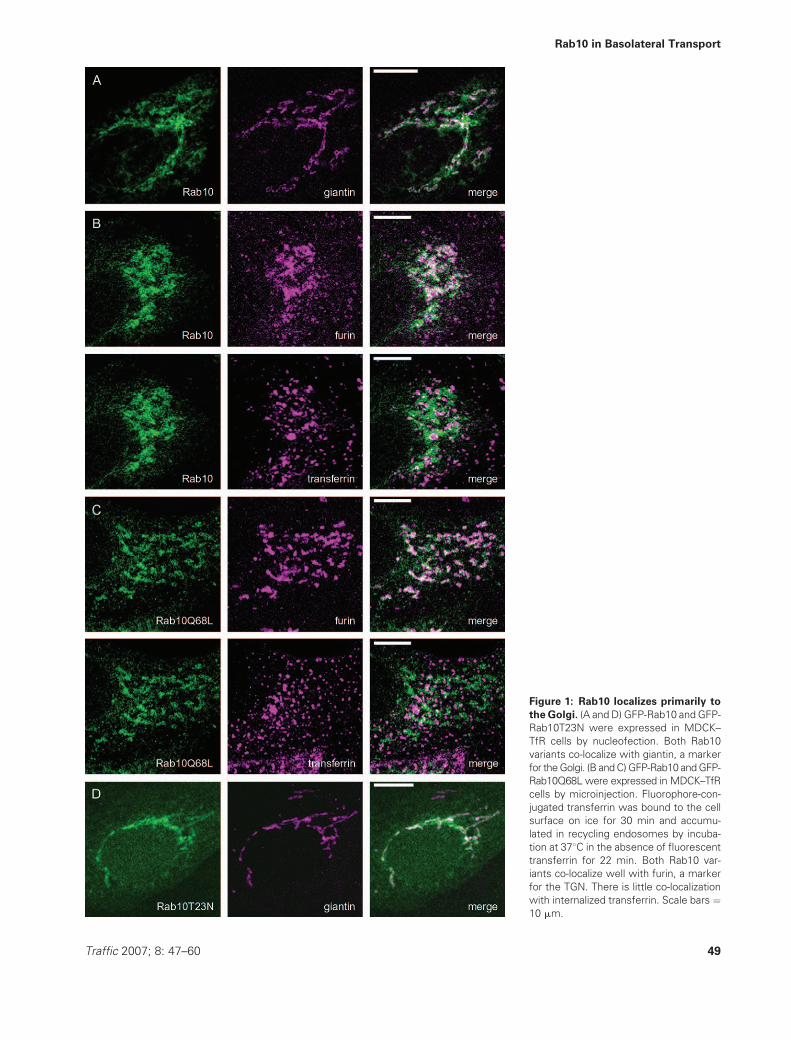
Figure 1: Rab10 localizes primarily to
the Golgi. (A and D) GFP-Rab10 and GFP-
Rab10T23N were expressed in MDCK–
TfR cells by nucleofection. Both Rab10
variants co-localize with giantin, a marker
for theGolgi. (B andC)GFP-Rab10 andGFP-
Rab10Q68L were expressed in MDCK–TfR
cells by microinjection. Fluorophore-con-
jugated transferrin was bound to the cell
surface on ice for 30 min and accumu-
lated in recycling endosomes by incuba-
tion at 378C in the absence of fluorescent
transferrin for 22 min. Both Rab10 var-
iants co-localize well with furin, a marker
for the TGN. There is little co-localization
with internalized transferrin. Scale bars ¼10 mm.
Traffic 2007; 8: 47–60 49
Rab10 in Basolateral Transport

shown). It is possible, therefore, that we did not reach
levels of Rab10T23N sufficient to delay VSV-G transport.
Finally, Rab10 may be involved in a trafficking step that is
not rate limiting for VSV-G transport and that can still
support overall cargo flow through the biosynthetic path-
way without Rab10. Only blocking this step by sequester-
ing multiple effectors with activated Rab10Q68L may lead
to transport inhibition. Similar cases, in which an activated
Rabmutant caused clear alterationswhereas a correspond-
ing inactivated mutant had subtle or no effects, have been
reported before (13,14,26,27).
To confirm the inhibition of VSV-G transport by Rab10Q68L
biochemically, VSV-G delivery to the plasma membrane
was measured using surface biotinylation. As before,
confluent coverslip-grown control and Rab10Q68L cells
were infected with VSV-G adenovirus, the cargo was
accumulated in the ER at 39.58C and chased to the plasma
membrane by shifting the temperature to 328C. After thechase, tight junctions were opened with ethylenediamine-
tetraacetic acid (EDTA) and the whole cell surface was
biotinylated. Total and biotinylated VSV-G were quantified
by electrochemiluminescence. Following release from the
Figure 2: Activated Rab10 inhibits transport from theGolgi to the plasmamembrane. Coverslip-grown control and Rab10Q68L cells
were infected with VSV-G adenovirus. VSV-G was accumulated in the ER at 39.58C for 6 h. A) Control cells. VSV-G was chased out of the
ER at 328C for 0, 25 or 60min. VSV-G transiently co-localizes with giantin as it moves to the Golgi and reaches the plasmamembrane. Scale
bars¼ 10mm. B) Rab10Q68L cells. VSV-Gwas chased out of the ER as above. VSV-Gmoves to the Golgi as in control cells, but then fails to
reach the plasmamembrane and remains in the Golgi. Scale bars¼ 10 mm. C) VSV-Gwas chased out of the ER at 328C for 0, 30, 40, 50, 60
or 80 min. To measure arrival at the plasma membrane, the cell surface was biotinylated, and surface and total VSV-G were quantified by
electrochemiluminescence. ER to plasma membrane transport is strongly inhibited in Rab10Q68L cells. Data are mean � SEM from an
experiment performed in quadruplicate. D) VSV-Gwas accumulated in the Golgi at 19.58C for 90min and chased out of the Golgi at 328C for
0, 15, 30, 45, 60 or 75 min. Arrival at the plasma membrane was measured as above. Golgi to plasma membrane transport is strongly
inhibited in Rab10Q68L cells. Data are mean � SEM from an experiment performed in quadruplicate.
50 Traffic 2007; 8: 47–60
Schuck et al.

ER and a certain lag period, VSV-G steadily accumulated at
the plasma membrane in control cells, reaching maximum
levels after about 60 min. Surface arrival was strongly in-
hibited in cells expressing Rab10Q68L (Figure 2C). To test
if this was due to a delay in ER to Golgi transport, we made
use of the fact that Golgi exit but not ER exit of VSV-G is
blocked at 19.58C. VSV-G was accumulated in the ER at
39.58C, protein biosynthesis was shut off and the temper-
ature was shifted to 19.58C for 90 min. This led to an
accumulation of VSV-G in the Golgi in both control and
Rab10Q68L cells (Figure S1). When cells were then
incubated at 328C, VSV-G rapidly appeared on the surface
in control cells, but still did so only slowly in the presence
of Rab10Q68L (Figure 2D). Hence, the inhibition of VSV-G
surface delivery is not due to impaired ER to Golgi trans-
port, but more likely reflects an inhibition of a later trans-
port step.
Activated Rab10 retains biosynthetic cargo
in the TGN
Rab10Q68L could accumulate VSV-G in the Golgi by
inhibiting intra-Golgi transport, by delaying Golgi exit or
by interfering with post-Golgi trafficking. To narrow down
which transport step is most severely affected by
Rab10Q68L, we monitored the glycosylation of VSV-G
during surface delivery. VSV-G is modified in the ER by the
addition of two N-linked core glycans, which are converted
to complex type glycans in the Golgi (28). The glycans are
sensitive to cleavage by endoglycosidase H (endoH) until
they are rendered endoH resistant bymannosidase II in the
medial Golgi. EndoH resistance therefore indicates that
VSV-G has passed through the early Golgi. Control and
Rab10Q68L cells were infected with VSV-G adenovirus,
newly synthesized VSV-G was accumulated in the ER at
39.58C, chased to the Golgi at 19.58C and finally chased to
the plasma membrane at 328C. At each stage, cells were
lysed, treated with endoH and subjected to immunoblot-
ting with an anti-VSV-G antibody, or fixed and processed for
immunofluorescence microscopy.
In control cells, VSV-G was entirely endoH sensitive when
accumulated in the ER, as indicated by a shift in its
electrophoretic mobility upon endoH treatment (Figure 3A,
lane 1 and 2). Incubation at 19.58C for 120 min led to trans-
port from the ER to the Golgi as judged by co-localization
with giantin (see Figure S1). At this point, about half of the
VSV-G had become endoH resistant (Figure 3A, lane 3).
This incomplete conversion into the endoH-resistant form
has been observed before (28) and likely reflects the high
demand on the machinery for protein transport and glyco-
sylation during release of the accumulated VSV-G from the
ER. When cells were chased for another 60 min at 328C,VSV-G reached the plasma membrane (see Figure 2A and
D) and became fully endoH resistant (Figure 3A, lane 4). In
the presence of Rab10Q68L, VSV-G stayed endoH sensi-
tive during the incubation at 19.58C (Figure 3A, lane 5–7),
although it had reached the Golgi (see Figure S1). This
observation is consistent with a defect in transport through
the early Golgi. However, it is equally consistent with an
inhibition of Golgi exit, which could cause cargo to build up
in the TGN and subsequently lead to an accumulation of
incoming cargo also in the early Golgi. It is possible that
endogenous cargo had already obstructed the late as well
as medial Golgi at the timewhen VSV-G was released from
the ER, thus preventing VSV-G from becoming endoH re-
sistant.When the temperaturewas shifted to 328C for 60min,
most of the VSV-G acquired endoH resistance (Figure 3A,
lane 8). In contrast to control cells, however, VSV-G re-
mained intracellular and exhibited a distribution similar to
that of giantin (Figure 3B, top panel). These observations
show that VSV-G had encountered mannosidase II at this
stage, but was unable to exit the Golgi. Nevertheless, VSV-G
and giantin showed little overlap. Instead, they were
typically found in close apposition, with the more tubular
giantin-positive structures surrounding those positive for
VSV-G. When the experiment was repeated with MDCK–
TGN38 cells, which express the human TGN38 as amarker
for the TGN, extensive co-localization between VSV-G
and TGN38 was observed (Figure 3B, bottom panel).
These results show that VSV-G is able to pass through
the cis Golgi in Rab10Q68L cells, but is then retained in
the TGN. Hence, the step most severely affected by
Rab10Q68L is downstream of transport through the early
Golgi. These effects of Rab10Q68L suggest that Rab10
functions in transport from the Golgi, although we cannot
rule out that the inhibition of Golgi exit is caused indirectly
by a block at a later step.
Activated Rab10 selectively disrupts
basolateral sorting
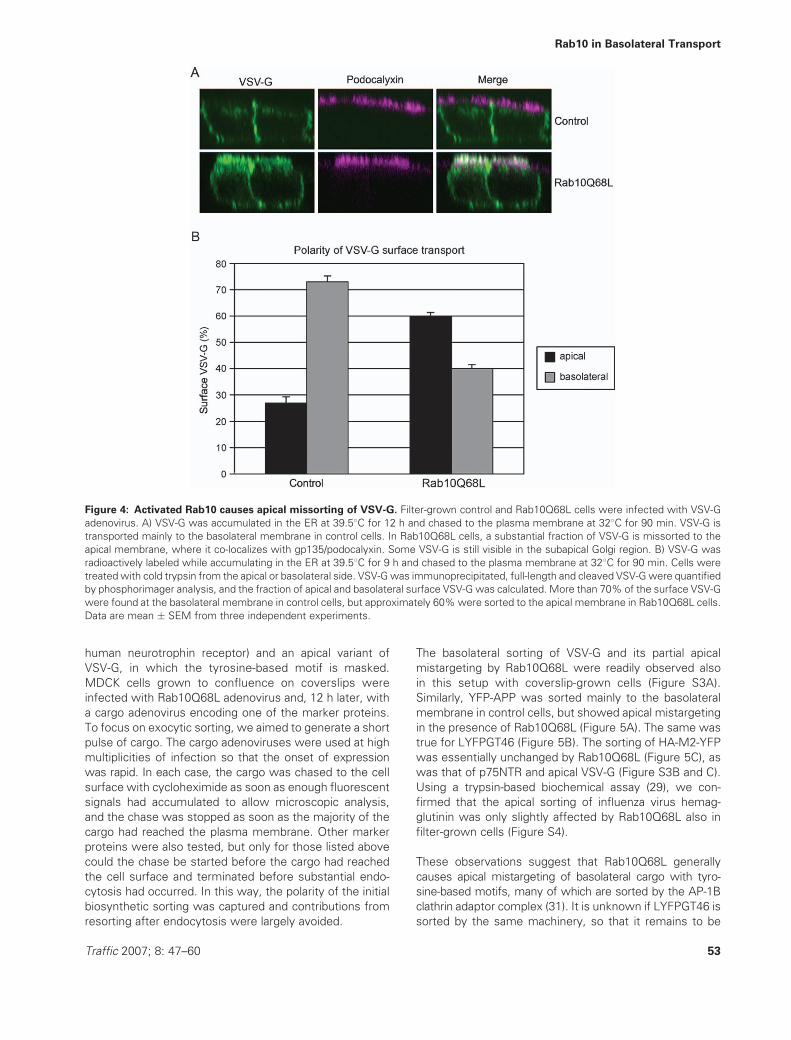
At long chase times, VSV-G eventually reached the plasma
membrane in Rab10Q68L cells. However, instead of being
sorted mostly to the basolateral side as in control cells, it
was partially mistargeted to the apical surface. This was
already obvious in confluent cells on coverslips, but is best
appreciated in MDCK cells grown on filters to generate
a well-polarized epithelial monolayer. When VSV-G was
expressed in control cells, accumulated in the ER and
chased to the plasma membrane at 328C for 90 min, it
was mainly at the basolateral domain, and there was
little overlap with gp135/podocalyxin, a marker for the
apical membrane (Figure 4A, top panel). The VSV-G visible
underneath the apical membrane was still in the subapical
Golgi region, as judged by co-localization with giantin (data
not shown). In contrast, a large fraction of VSV-G was
transported to the apical membrane in cells expressing
Rab10Q68L (Figure 4A, bottom panel). Rab10Q68L cells
were somewhat less well polarized than control cells, pre-
sumably as a result of the disturbed biosynthetic sorting,
but overall polarity of the epithelial monolayer was main-
tained. The intracellular retention of VSV-G by Rab10Q68L
was less pronounced than in coverslip-grown cells (see
Figure 2), likely because infection with the Rab10Q68L
Traffic 2007; 8: 47–60 51
Rab10 in Basolateral Transport

adenovirus was less efficient in filter-grown cells and
yielded lower Rab10Q68L expression levels.
To quantify the mistargeting caused by Rab10Q68L, VSV-G
was expressed in filter-grown MDCK cells, radioactively
labeled during accumulation in the ER at 39.58C and chased
to the plasma membrane at 328C for 90 min. After the
chase, cells were placed on ice and treated with trypsin
from the apical or basolateral side. Low trypsin concentra-
tions were used that preserve the separation of apical and
basolateral domains (29). This treatment resulted in partial
cleavage of the VSV-G ectodomain. Full-length and cleaved
VSV-G were immunoprecipitated with an antibody against
the cytoplasmic YFP tag and the fraction of VSV-G cleavable
from the apical or basolateral sidewas quantified (Figure S2).
In control cells, approximately 70% of the surface VSV-G
were basolateral (Figure 4B), in agreement with previous
data obtained by domain-selective surface biotinylation
(24). In Rab10Q68L cells, however, the majority of the
surface VSV-G was found at the apical membrane, and only
40% were basolateral. As the basolateral domain is larger
than the apical, this distribution does not reflect random
delivery, but preferential apical transport. In addition, only
about 80% of the cells were infected by the Rab10Q68L
adenovirus, so that the effect of the activated Rab10mutant
is probably underestimated due to the presence of cells
that express VSV-G but not Rab10Q68L. These results
confirm that activated Rab10 causes apical mistargeting of
VSV-G and suggest that Rab10 is involved in the polarized
sorting of newly synthesized membrane proteins.
To test if Rab10Q68L also affects the polarity of other
basolateral or apical cargo, we analyzed the targeting of
five additional transmembrane proteins. Besides VSV-G,
we used YFP-tagged amyloid precursor protein (YFP-APP)
and LYFPGT46 as basolateral markers. Both VSV-G and
YFP-APP are sorted basolaterally by means of tyrosine-
based sorting motifs in their cytoplasmic tails. LYFPGT46
is a chimeric fusion protein that contains the atypical FTSL
basolateral sorting motif at the extreme C-terminus of its
cytoplasmic part (30). Apical markers were HA-M2-YFP
(based on influenza virus hemagglutinin), p75NTR (the
Figure 3: Activated Rab10 retains VSV-G in the TGN. A) Coverslip-grown control and Rab10Q68L cells were infected with VSV-G
adenovirus. VSV-Gwas accumulated in the ER at 39.58C for 6 h, accumulated in the Golgi at 19.58C for 120min and chased out of the Golgi at
328C for 60min.After eachstage,cell lysateswere treatedwithendoHandVSV-Gwasanalyzedby immunoblotting.Transport beyond theearly
Golgi renders VSV-G resistant to degradation to its faster migrating de-glycosylated form (VSV-Ge). In control cells, VSV-G starts to acquire
endoH resistance during Golgi accumulation and becomes entirely endoH resistant during the chase at 328C. In Rab10Q68L cells, VSV-G
remains endoH sensitive during Golgi accumulation, but is also mostly endoH resistant after the chase at 328C, showing that it has passed
the early Golgi. B) Rab10Q68L was expressed in MDCK or MDCK–TGN38 cells (upper and lower panel, respectively). VSV-G was accu-
mulated in the Golgi and chased at 328C for 60 min as above. After the chase at 328C, VSV-G is found in apposition to giantin and co-
localizes with TGN38, indicating that it has reached the TGN but is unable to leave the Golgi. Scale bars are 10 mm in the merge and 2 mm in
the zoom.
52 Traffic 2007; 8: 47–60
Schuck et al.
human neurotrophin receptor) and an apical variant of
VSV-G, in which the tyrosine-based motif is masked.
MDCK cells grown to confluence on coverslips were
infected with Rab10Q68L adenovirus and, 12 h later, with
a cargo adenovirus encoding one of the marker proteins.
To focus on exocytic sorting, we aimed to generate a short
pulse of cargo. The cargo adenoviruses were used at high
multiplicities of infection so that the onset of expression
was rapid. In each case, the cargo was chased to the cell
surface with cycloheximide as soon as enough fluorescent
signals had accumulated to allow microscopic analysis,
and the chase was stopped as soon as the majority of the
cargo had reached the plasma membrane. Other marker
proteins were also tested, but only for those listed above
could the chase be started before the cargo had reached
the cell surface and terminated before substantial endo-
cytosis had occurred. In this way, the polarity of the initial
biosynthetic sorting was captured and contributions from
resorting after endocytosis were largely avoided.
The basolateral sorting of VSV-G and its partial apical
mistargeting by Rab10Q68L were readily observed also
in this setup with coverslip-grown cells (Figure S3A).
Similarly, YFP-APP was sorted mainly to the basolateral
membrane in control cells, but showed apical mistargeting
in the presence of Rab10Q68L (Figure 5A). The same was
true for LYFPGT46 (Figure 5B). The sorting of HA-M2-YFP
was essentially unchanged by Rab10Q68L (Figure 5C), as
was that of p75NTR and apical VSV-G (Figure S3B and C).
Using a trypsin-based biochemical assay (29), we con-
firmed that the apical sorting of influenza virus hemag-
glutinin was only slightly affected by Rab10Q68L also in
filter-grown cells (Figure S4).
These observations suggest that Rab10Q68L generally
causes apical mistargeting of basolateral cargo with tyro-
sine-based motifs, many of which are sorted by the AP-1B
clathrin adaptor complex (31). It is unknown if LYFPGT46 is
sorted by the same machinery, so that it remains to be
Figure 4: Activated Rab10 causes apical missorting of VSV-G. Filter-grown control and Rab10Q68L cells were infected with VSV-G
adenovirus. A) VSV-G was accumulated in the ER at 39.58C for 12 h and chased to the plasma membrane at 328C for 90 min. VSV-G is
transported mainly to the basolateral membrane in control cells. In Rab10Q68L cells, a substantial fraction of VSV-G is missorted to the
apical membrane, where it co-localizes with gp135/podocalyxin. Some VSV-G is still visible in the subapical Golgi region. B) VSV-G was
radioactively labeled while accumulating in the ER at 39.58C for 9 h and chased to the plasma membrane at 328C for 90 min. Cells were
treatedwith cold trypsin from the apical or basolateral side. VSV-Gwas immunoprecipitated, full-length and cleaved VSV-Gwere quantified
by phosphorimager analysis, and the fraction of apical and basolateral surface VSV-G was calculated. More than 70% of the surface VSV-G
were found at the basolateral membrane in control cells, but approximately 60%were sorted to the apical membrane in Rab10Q68L cells.
Data are mean � SEM from three independent experiments.
Traffic 2007; 8: 47–60 53
Rab10 in Basolateral Transport

determined if the effect of Rab10Q68L is restricted to AP-
1B-dependent cargo. The finding that the polarity of three
apical markers is essentially unaffected by Rab10Q68L
argues that interfering with Rab10 function specifically
disrupts basolateral sorting. Missorting of basolateral but
not apical cargo has also been found with activated Rab8.
Activated Rab8 does not, however, inhibit exit from the
Golgi (13).
Inhibition of Rab10 and Rab8a disturb basolateral
sorting in an additive manner
An activated Rab mutant can sequester multiple effector
proteins and thus have pleiotropic effects. To more faith-
fully assess the role of Rab10, we used RNA interference
(RNAi) to deplete the endogenous protein in MDCK cells.
Retrovirus-mediated expression of small hairpin RNAs
followed by elimination of non-transduced cells by anti-
biotic selection reduced Rab10 mRNA levels by 95 � 1%
[mean � standard error of the mean (SEM), n ¼ 5]. We
could not measure Rab10 protein levels directly as no
suitable antibody was available. Yet, expression of trans-
fected GFP-Rab10 was almost completely suppressed in
Rab10 knockdown cells (Figure 6A), suggesting that the
endogenous Rab10 had largely been removed.
Rab10 knockdown cells displayed no obvious delay in
biosynthetic transport of VSV-G and established a normal
epithelial monolayer when seeded onto filters (data not
shown). In addition, they sorted VSV-G with only slightly
lower accuracy than control cells (Figure 6B). This finding
suggests that Rab10 is not essential for basolateral sort-
ing, although we cannot exclude that the residual Rab10
was sufficient to maintain its function.
One reason for the mild sorting phenotype in Rab10
knockdown cells could be that the function of Rab10 is
partially redundant with that of another Rab protein,
particularly Rab8. MDCK cells express two Rab8 isoforms,
Rab8a and Rab8b, of which Rab8a is the major form as
judged by quantitative RT–PCR (data not shown). Retrovirus-
mediated depletion of Rab8a mRNA by 91 � 1% (mean �SEM, n ¼ 3) caused minor missorting of VSV-G, indicating
that also depletion of Rab8a alone does not disrupt baso-
lateral sorting (Figure 6C). When we targeted both Rab10
Figure 5: Activated Rab10 causes missort-
ing of basolateral but not apical cargo.
Coverslip-grown control and Rab10Q68L cells
were infected with A) YFP-APP, B) LYFPGT46
or C) HA-M2-YFP adenovirus. Cargo was accu-
mulated for the minimum time necessary to
allow microscopic analysis and chased to the
plasma membrane for 90 min. An apical and
a basal optical section are shown for each
condition. YFP-APP and LYFPGT46 are trans-
ported mostly to the basolateral membrane in
control cells, but are partially missorted to the
apical membrane in Rab10Q68L cells. The
polarity of HA-M2-YFP is essentially unchanged
in Rab10Q68L cells. YFP-APP consists of the
YFP tag (yellow) and the full-length APP (gray)
with the YTSI sorting motif (red). LYFPGT46 is
YFP (yellow) fused to the LDL receptor trans-
membrane domain (gray) and the CD46 cyto-
plasmic tail (black), including the FTSL sorting
motif (green). HA-M2-YFP is hemagglutinin
(gray) fused to the cytoplasmic tail of influenza
virus M2 (black) and YFP (yellow). Scale bars ¼10 mm.
54 Traffic 2007; 8: 47–60
Schuck et al.

and Rab8a using two different retroviruses, the fraction of
apical surface VSV-G increased from 30% to slightly over
40% (data not shown). Thus, the combined depletion of
Rab10 and Rab8a affects VSV-G sorting somewhat more
strongly than the depletion of either of the two Rabs alone.
However, we were unable to simultaneously reduce the
mRNA levels of both Rab10 and Rab8a by more than 90%,
presumably because cells with a strong depletion of both
Rabs are rapidly eliminated from the cell population. The
observed VSV-G missorting in Rab10/Rab8a double knock-
down cells is therefore likely an underestimate of the true
effect of removing both Rabs.
We reasoned that the expression of inactivated Rab10T23N
might be equivalent to Rab10 RNAi as it sequesters the
guanine nucleotide exchange factor necessary for the con-
version of Rab10 to the active GTP-bound form. Consistent
with this idea, adenovirally expressed Rab10T23N mis-
sorted VSV-G to a similar extent as Rab10 RNAi (Figure 6B
and C). We then combined Rab8a RNAi with adenoviral
expression of Rab10T23N. In this way, we aimed at
interfering with the function of Rab10 more acutely than
is possible with RNAi. This treatment increased the
fraction of surface VSV-G at the apical membrane from
30 to 45% (Figure 6C). Interference with both Rab8a and
Rab10 thus augments the effects of inhibiting only one of
the two Rabs and causes a more pronounced apical mis-
sorting of VSV-G. This effect suggests that Rab8a and
Rab10 are functionally related, either co-operating along
the same pathway or acting in parallel pathways.
Depletion of Rab10 impairs polarization of
MDCK cysts
One reason for the absence of an obvious morphological
phenotype in filter-grown Rab10 knockdown cells could be
that cell polarization is very robust in the presence of the
strong spatial cue provided by the filter support. It has been
noted before that growth in a three-dimensional matrix,
in which MDCK cells develop into hollow spheres of cells
called cysts, places greater demands on the machinery for
epithelial polarization than the two-dimensional filter sys-
tem and provides a more sensitive assay for cell polarity
(32). When single MDCK cells were cultured in collagen
gels, Rab10 knockdown cells indeed exhibited polarization
defects not seen on filters. After 10 days of culture, control
cells had formed well-organized cysts with a smooth apical
membrane facing the cyst lumen. Rab10 knockdown cysts
often had irregular shapes and their apical surface was
frequently bulged out (Figure 7). This difference between
control and Rab10 knockdown cells persisted up to at least
13 days of culture, the longest time tested.
More work is needed to understand the immediate
cause for the observed morphological aberrations and
potentially link them to a role of Rab10 in polarized sorting.
Nevertheless, these results show that Rab10 has a non-
redundant function in MDCK cells that is required for
proper epithelial polarization in cysts.
Discussion
The aim of this study was to better understand the
molecular machinery for biosynthetic transport in polarized
Figure 6: Depletion of Rab10 or Rab8a by RNAi causes minor
missorting of VSV-G, but simultaneous inhibition of both
Rabs has a clear effect. Knockdown cells were generated by
retrovirus-mediated RNAi. MDCK cells transduced with empty
retrovirus served as control. A) Rab10 knockdown cells suppress
expression of GFP-Rab10. Control and Rab10 knockdown cells
were transfected with GFP-Rab10 by nucleofection. Equivalent
amounts of cell lysate were separated by SDS–PAGE and ana-
lyzed by immunoblotting. Lysate from untransfected cells (MDCK)
served as background control. B) Rab10 knockdown only slightly
affects VSV-G sorting. VSV-G transport was assayed in filter-
grown control and Rab10 knockdown cells as in Figure 4B. Data
are mean � SEM from four independent experiments. C) Rab8a
knockdown and expression of Rab10T23N only slightly affect VSV-
G sorting, but combined treatment more strongly impairs baso-
lateral targeting. VSV-G transport was assayed in filter-grown
control and Rab8a knockdown cells that expressed Rab10T23N
or not as in Figure 4B. Data are mean � SEM from three
independent experiments. KD, knockdown.
Traffic 2007; 8: 47–60 55
Rab10 in Basolateral Transport

epithelial cells. We focused on Rab proteins, which are key
regulators of the intracellular membrane trafficking net-
work. We analyzed Rab10 because it is homologous to
Sec4p, a central component of post-Golgi trafficking in
yeast. Our results indicate that Rab10 regulates transport
from the Golgi, at least at early stages of MDCK cell
polarization, and is involved in basolateral sorting. How-
ever, Rab10 is not strictly required for biosynthetic trans-
port, as depletion of Rab10 yields only mild sorting and
polarity phenotypes. Depletion of Rab8a also causes only
minor missorting of basolateral cargo, but simultaneous
inhibition of the two Rabs results in a more pronounced
sorting defect. These findings suggest that Rab10 and
Rab8 are functionally related and point to considerable
robustness built into post-Golgi trafficking in polarized cells.
Where might Rab10 act? We observed GFP-tagged wild-
type Rab10, activated Rab10Q68L and inactivated
Rab10T23N mostly at the Golgi in coverslip-grown MDCK
cells. A minor population of GFP-Rab10 and GFP-
Rab10Q68L was found on recycling endosomes. Babbey
et al. recently reported that GFP-Rab10T23N localizes to
the Golgi also in filter-grown, fully polarized MDCK cells.
However, they observed GFP-Rab10 and GFP-Rab10Q68L
mostly on recycling endosomes under these conditions
(18). We have confirmed that transiently transfected GFP-
Rab10 is on recycling endosomes in more fully polarized
MDCK cells grown on filters (our unpublished data). It
therefore appears that Rab10 is at the Golgi at early stages
of MDCK cell polarization, but then shifts towards recy-
cling endosomes. This view is consistent with the other
previous studies of Rab10 localization, which showed that
HA-tagged Rab10 is at the late Golgi in non-polarized cells
(16) and that GFP-Rab10 is distributed between the Golgi
and endosomes in polarized intestinal cells in C. elegans
(17). Similar changes in the localization of Rab proteins
during cell polarization have been observed before. Rab11,
for instance, is found at recycling endosomes and the Golgi
in non-polarized cells, but is restricted to recycling endo-
somes in fully polarized MDCK cells [(33) and references
therein].
In the presence of Rab10Q68L, newly synthesized VSV-G
accumulates in the Golgi of incompletely polarized MDCK
cells, indicating a role of Rab10 in anterograde trafficking
through the secretory pathway. The step most severely
affected by Rab10Q68L is downstream of transport through
the early Golgi. It is difficult to determine if Rab10Q68L
retains VSV-G in the Golgi directly, by slowing down export
from the TGN, or indirectly, by inhibiting a later trafficking
step. However, as we found Rab10 primarily at the Golgi,
the most straightforward explanation for the effect of
Rab10Q68L is that Rab10 functions in transport from the
Golgi at early stages of MDCK cell polarization.
There is evidence that biosynthetic cargo such as VSV-G
reaches the basolateral membrane of MDCK cells via
recycling endosomes (25,34). As Rab10Q68L causes
apical missorting of VSV-G, one possibility is that Rab10
functions in trafficking from the Golgi to recycling endo-
somes during early epithelial polarization. Disrupting this
pathway by the expression of Rab10Q68L could force
basolateral cargo into a direct route from the Golgi to the
apical membrane. Alternatively, Rab10 could function in
trafficking from the Golgi to the apical membrane and
activated Rab10Q68L could draw basolateral cargo into
this pathway. While we cannot rule out this explanation,
we think it less likely as it does not account for the
observed Golgi retention of basolateral cargo. In addition,
the delivery of apical cargo is slowed down rather than
accelerated by Rab10Q68L (our unpublished data), pos-
sibly as a secondary consequence of an accumulation
of endogenous basolateral cargo in the Golgi. In either
scenario, the decision of apical versus basolateral delivery
would be associated with Golgi exit, arguing that the Golgi
is an important biosynthetic sorting station.
We observed the missorting of basolateral but not
apical cargo by Rab10Q68L in both coverslip-grown and
Figure 7: Depletion of Rab10 by RNAi disturbs epithelial
polarization. Control cells transduced with empty retrovirus and
Rab10 knockdown cells were grown in collagen gels to form
cysts. A) Apical and basolateral membranes were stained with
antibodies against podocalyxin (red) and E-cadherin (green),
respectively, and cell nuclei with DAPI (blue). Rab10 knockdown
cysts are typically less well organized and have curved apical
membranes. B) Cysts with flat or curved apical membranes were
counted. Data are mean � SEM from three independent experi-
ments, at least 50 cysts per condition were counted in each
experiment. Scale bars ¼ 30 mm. KD, knockdown.
56 Traffic 2007; 8: 47–60
Schuck et al.

filter-grown cells, suggesting that Rab10 plays a similar
role in basolateral sorting at early as well as late stages of
polarization. However, given the change in its localization
during polarization, Rab10 may be able to function at
different stations of the biosynthetic pathway. Epithelial
cells clearly make differential use of their repertoire of
trafficking pathways depending on their polarization state
(6). Biosynthetic sorting may likewise be accomplished at
different trafficking stations during polarization and the
involved sorting machinery may be relocated accordingly.
We therefore speculate that the sorting events involving
Rab10 occur at the Golgi during early polarization, but shift
towards recycling endosomes when MDCK cells achieve
full epithelial polarity.
We did not analyze possible effects of Rab10Q68L on
endocytosis. However, as Rab10Q68L missorts certain
biosynthetic cargo, interfering with Rab10 function could
disrupt the supply of components of the endocytic machin-
ery and in this way indirectly inhibit endocytosis and
recycling. This view is compatible with the work in C.
elegans, which showed that lack of Rab10 causes shrink-
age of recycling endosomes and swelling of early endo-
somes in intestinal cells (17). Rab10 is normally distributed
between the Golgi and endosomes in these cells, so that
the decrease in recycling endosome size may result from
an inhibition of Golgi to endosome transport. Internalized
cargo could subsequently accumulate in early endosomes
because transport through recycling endosomes is impaired.
Such indirect effects may also explain why the only other
study on Rab10 in MDCK cells reported that Rab10T23N is
at the Golgi and affects endocytic recycling from early
endosomes (18). We conclude that Rab10 functions in
exocytic transport, but we do not rule out that it may also
be needed, directly or indirectly, for endocytic trafficking.
The effects of Rab10Q68L indicate that Rab10 is involved
in basolateral sorting in MDCK cells, but do not answer the
question if Rab10 is also required for this process. At high
expression levels, activated Rab mutants are likely to
sequester and thus inactivate a number of effector pro-
teins. This may generally be more disruptive than interfer-
ing with the co-ordination of effector proteins by inhibiting
the corresponding endogenous Rab, be it with an inacti-
vated mutant or by RNAi. Inactivated Rab10T23N and
Rab10 RNAi had only minor effects on basolateral sorting,
suggesting that Rab10 is not essential for polarized trans-
port in MDCK cells. Inhibition of Rab10 may be tolerated
due to redundancy with other Rabs. One Rab that likely has
a related function is Rab8, as suggested by the more
pronounced missorting of basolateral cargo after simulta-
neous inhibition of both Rab10 and Rab8a. Accordingly,
expression of either inactivated Rab8 or inactivated Rab10
in the Drosophila wing epithelium cause relatively mild
problems in wing morphogenesis, whereas combined
expression of the two mutant Rabs has much more severe
effects (E.Marois andS. Eaton,MPI-CBG,Dresden,Germany,
personal communication). It therefore appears that Rab10,
although it is ubiquitously expressed (15), is indispensable
only in particular cell types, such as C. elegans intestinal
cells (17), during certain developmental processes, such as
cyst formation in MDCK cells, or after inhibition of func-
tionally related Rabs, such as Rab8.
The interplay of Rab10 and Rab8 deserves further analysis.
It is possible that the two Rabs can replace each other or
that they act in parallel basolateral pathways. A third
possibility is that they co-operate along the same pathway.
As discussed above, Rab10 likely functions transport
between the Golgi and recycling endosomes, whereas
Rab8 is associated with recycling endosomes (13). An
attractive hypothesis is that Rab10 and Rab8 co-operate
during basolateral transport as part of a Rab cascade, as
proposed for Rab5 and Rab4 as well as Rab5 and Rab7 in
the endocytic pathway in mammalian cells (35,36) and
for Ypt31/32p and Sec4p in the exocytic pathway in
yeast (37). A further speculation is that the multiple func-
tions of Sec4p in yeast are carried out by separate Rabs
in higher organisms, so that Rab8, Rab10 and possibly
Rab13 could represent a split version of Sec4p. Several
other Rab proteins will ultimately have to be integrated
into this model to obtain a more complete map of the
biosynthetic transport routes in polarized cells. These
include Rab11a and Rab11b, at least one of which seems
to function in basolateral transport (19,23), and Rab14,
which also could have a role in biosynthetic post-Golgi
trafficking (20).
Functional similarity and redundancy between Rab pro-
teins is a recurrent theme. Different isoforms of the same
Rab often have the same function andmay be interchange-
able (38–40). Distinct Rabs sometimes have related func-
tions and may be able to fulfill the same task by similar
means (26,41). Thus, it is likely that the intracellular
trafficking network has a number of redundant elements.
The resulting robustness may allow cells to tolerate the
inhibition of certain parts of their trafficking machinery
without overall disruption of sorting and transport. The Rab
family has expanded substantially during evolution (12).
This expansion presumably reflects the diversification of
membrane trafficking pathways in higher organisms, but
the benefits of increased robustness afforded by redun-
dancy may also have been a driving force.
In summary, this work defines Rab10 as a new component
of the machinery for polarized biosynthetic transport. Under-
standing the precise function of Rab10 and how it is
connected with other parts of the transport machinery, such
as cargo adaptors, molecular motors, vesicle fusion proteins
and other Rab GTPases, will require the identification of
Rab10 effector proteins. Given the complex morphology
of the post-Golgi membrane system, it will be a continuing
challenge to map transport and sorting events onto in-
tracellular locations. In addition, we consider it likely that
sorting decisions are made in different places depending on
cell type and developmental stage. To come to grips
Traffic 2007; 8: 47–60 57
Rab10 in Basolateral Transport

with this complexity, we need a more detailed understand-
ing of the molecular sorting machinery and its modular
construction.
Materials and Methods
AntibodiesMouse monoclonals were anti-gp135/podocalyxin (G. Ojakian, SUNY
Downstate Medical Center, Brooklyn, NY, USA), anti-giantin (H. P. Hauri,
University of Basel, Basel, Switzerland), anti-E-cadherin (Simons lab,
MPI-CBG, Dresden, Germany), anti-acetylated b-tubulin (Sigma, Munich,
Germany), anti-p75NTR and anti-myc (both from Santa Cruz Biotechnology,
Santa Cruz, CA, USA). Polyclonals were rabbit anti-mRFP (M. Zerial, MPI-
CBG), goat anti-GFP (D. Drechsel, MPI-CBG), rabbit anti-VSVG (Simons lab)
and rabbit anti-furin (Dianova, Hamburg, Germany). The rabbit anti-podoca-
lyxin antibody was raised against the CDNLAKDDLDEEEDTHL epitope.
Plasmids and adenovirusesCanine Rab10 cDNA M. Zerial was cloned into pEGFP-C1 (Clontech,
Moutain View, CA, USA) to generate pGFP-Rab10. Mutations yielding
pGFP-Rab10T23N and pGFP-Rab10Q68L were introduced using the Quik-
Change kit (Stratagene, La Jolla, CA, USA). RFP-tagged variants were
constructed by replacing the GFP with monomeric RFP1. They were
subcloned into pAdEasy-1 (Qbiogene, Heidelberg, Germany) and adenovi-
ruses were generated according to the Qbiogene manual. The p75NTR
adenovirus was from E. Rodriguez-Boulan (Cornell University, New York,
NY, USA). All other adenoviruses have been described before (24,42–44).
Cell cultureMDCK cells, MDCK–TfR cells expressing the human transferrin receptor
and MDCK–TGN38 cells expressing myc-tagged human TGN38 were
cultured as described (13). For transport assays, cells were seeded at
1.5 � 105 per 24 well containing a glass coverslip or at 5 � 105 per 12-mm
Transwell filter (Corning Life Sciences, Corning, NY, USA). After 24 h, cells
were infected or not with Rab10Q68L adenovirus in OptiMEM at 378C for
1 h and grown for another 12 h.
Transfection and immunofluorescence microscopyGFP-Rab10 constructs were introduced into MDCK–TfR cells by nucleo-
fection or microinjection. Nucleofection, an electroporation-based method,
was done as recommended by the manufacturer (Amaxa, Cologne,
Germany), cells were grown on coverslips for 20 h and immunostained.
Low expression levels of GFP-Rab10 and especially GFP-Rab10Q68L were
difficult to achieve by this method and the proteins were often mainly
cytosolic. Microinjection was therefore preferred to more tightly control
their levels of expression. Coverslip-grown cells were microinjected, grown
for 2 h, labeled with fluorescent transferrin and immunostained as
described (13). Confocal microscopy was performed with a laser scanning
microscope (LSM510; Carl Zeiss, Jena, Germany) using 40� water immer-
sion (n¼ 1.5) or 63� oil immersion (n¼ 1.4) objectives at 258C. Imageswere
processed with Adobe Photoshop software (Adobe, San Jose, CA, USA).
Microscopic transport assays on coverslipsTo assay VSV-G transport, coverslip-grown control and Rab10Q68L cells
were infected with VSV-G adenovirus in OptiMEM at 378C for 2 h. For
microscopic analysis, cells were incubated in normal medium at 39.58C for
6 h, chased in medium containing 40 mg/mL cycloheximide at 328C for
90 min and immunostained. To monitor its glycosylation, VSV-G was accu-
mulated in the ER, chased with cycloheximide at 19.58C for 120 min
and allowed to exit the Golgi at 328C for 60 min. Cells were collected at the
end of each stage and treated with endoH as described (28). The same
conditions as above were applied for apical VSV-G. For YFP-APP,
LYFPGT46, HA-M2-YFP and p75NTR, cells were infected with the respec-
tive adenovirus for 1–3 h, incubated at 378C for 2–9 h, chased with
cycloheximide for 90 min and immunostained. In each case, expression of
the marker was allowed for the minimum time necessary to produce a
sufficient amount for microscopic analysis, and the chase was started
when no marker protein was yet visible at the cell surface. Endocytosis
of these markers was relatively slow. This allowed most of the cargo
to reach theplasmamembranebeforesubstantial endocytosis hadoccurred.
Biochemical VSV-G transport assay on coverslipsVSV-G was accumulated in the ER of coverslip-grown control and
Rab10Q68L cells as described above. To assay ER to plasma membrane
transport, cells were chased in medium containing 40 mg/mL cycloheximide
at 328C for 0, 30, 40, 50, 60 and 80 min. To assay Golgi to plasma
membrane transport, cells were incubated in medium with cycloheximide
at 19.58C for 90 min to accumulate VSV-G in the Golgi and chased at 328Cfor 0, 15, 30, 45, 60 and 75min. Coverslips were washed with cold PBS and
incubated with 1 mM EDTA/PBS on ice for 10 min to open the tight
junctions and make the entire cell surface biochemically accessible. Cells
were surface biotinylated with 1 mg/mL Sulfo-NHS-LC-biotin (Pierce,
Rockford, IL, USA) on ice for 2 � 20 min. Unreacted biotin was quenched
with cold 0.3% BSA/0.1 M glycine for 2 � 10 min and cells were lysed with
PBS containing 2% Nonidet P-40 (NP-40), 0.2% sodium dodecyl sulphate
(SDS) and protease inhibitors. Total and biotinylated VSV-G were quantified
using electrochemiluminescence technology from Meso Scale Discovery
(Gaithersburg, MD, USA). To determine total VSV-G, MA6000 96-well
plates were coated with 10 ng affinity-purified rabbit anti-GFP antibody per
well, quantification of biotinylated VSV-G was done on avidin-coated
MA6000 high bind 96-well plates. Wells were incubated with 3% BSA in
PBS/0.2% NP-40 for 1 h to prevent non-specific binding. Lysates were
diluted in PBS containing 0.06% NP-40, 0.006% SDS and 100 mM HEPES
and added to the wells for 1 h to allow capture of the VSV-G by means of
the YFP tag or the biotin moiety. Wells were washed with PBS/0.2%
NP-40, and 20 ng rabbit anti-GFP antibody labeled with MSD Sulfo-Tag
were added for detection of the immobilized VSV-G. After washing, MSD
Read Buffer T with surfactant was added, and the electrochemilumines-
cence signal was detected using a Sector Imager 6000. Values for surface
VSV-G were normalized for total VSV-G to eliminate variations in expression
levels.
Transport assays on filtersTo measure the polarity of VSV-G surface transport, filter-grown control and
Rab10Q68L cells were infected with VSV-G adenovirus in OptiMEM at
378C for 2 h. For microscopic analysis, cells were incubated at 39.58C for
12 h, chased with cycloheximide at 328C for 90 min and immunostained.
For biochemical analysis, VSV-G was radioactively labeled and accumulated
in the ER by incubation at 39.58C for 9 h in labeling medium (methionine/
cysteine-free MEM, 1.5 mg/mL methionine, 2.5% fetal calf serum, 20 mM
HEPES pH 7.2) with 50 mCi 35S-methionine added to the basolateral side.
Cells were washed with PBS containing Mg2þ and Ca2þ (PBSþ) and
incubated in chase medium containing excess non-radioactive methionine
(methionine/cysteine-free MEM, 150 mg/mL methionine, 20 mM HEPES
pH 7.2) at 328C for 90 min. Cells were washed 3 � 5 min with cold PBSþand treated with 100 mg/mL trypsin (Worthington, Lakewood, NJ, USA) in
PBSþ from the apical or basolateral side on ice for 30 min. Tight junctions
remain intact under these conditions and restrict trypsin to the side to
which it has been added (29). Cells were washed 3� 5 min with 0.1 mg/mL
soy bean trypsin inhibitor (Worthington) and full-length and cleaved VSV-G
were immunoprecipitated with an anti-GFP antibody against the cytosolic
YFP tag that is protected from cleavage. Immunoprecipitates were
separated by SDS–PAGE and full-length and cleaved VSV-G were quantified
by phosphorimager analysis. The ratio of apical to basolateral surface VSV-G
was calculated from the fraction of VSV-G cleaved by trypsin from the apical
or basolateral side. The polarity of influenza virus hemagglutinin surface
transport was measured as described (29). Briefly, filter-grown MDCK cells
were infected with influenza virus and incubated for 3 h to allow host
protein shut-off to occur. Newly synthesized virus proteins were labeled
with 35S-methionine for 10 min and chased to the plasma membrane for
30 min. Cells were trypsinized as above, cell lysates were separated by
58 Traffic 2007; 8: 47–60
Schuck et al.

SDS–PAGE and the fraction of HA cleaved from the apical or basolateral side
was quantified by phosphorimager analysis.
RNAiThe Rab10 and Rab8a target sequences were GCTGAAGATATCCTTC-
GAAAG and AAGACAAGTTTCCAAGGAACG, respectively. Retrovirus-
mediated RNAi and quantitative RT–PCR were done as before (45).
Culture of MDCK cystsHydrated collagen I solution (2 mg/mL) was prepared by mixing 16 volumes
ice-cold VitrogenTM (3mg/mL collagen I; Cohesion, Palo Alto, CA, USA) with
2 volumes chilled 10� DMEM (Invitrogen, Karlsruhe, Germany), 2 volumes
0.1 M NaOH, 1 volume 7.5% NaHCO3 and 1 volume 1 M HEPES pH 7.2.
Fetal calf serum was added to a final concentration of 1% (v/v) and the
mixture was kept on ice until use. Subconfluent MDCK cells were trypsi-
nized and a single cell suspension containing 5� 106 cells/mLwas prepared
in PBS. Cells were pipeted into the collagen solution to yield a mixture
containing 2 � 105 cells/mL. The mixture was pipeted onto 24-well plates
and incubated at 378C for 30–45 minutes to allow the collagen to solidify.
Medium was added and the incubation was continued for 10 days. Medium
was exchanged every other day.
Immunostaining of MDCK cystsCollagen gels were rinsed twice with PBSþ and incubated in 0.05%
collagenase A in PBSþ for 5 min at 378C. After one wash with PBSþ, cells
were fixed in 4% paraformaldehyde for 30 minutes and excess aldehyde
was quenched with 200 mM glycine in PBS. Cells were permeabilized for
30 min in PBS containing 0.1% Triton-X-100 and non-specific binding was
blocked by incubation in PBS containing 0.5% BSA, 0.2% fish skin gelatin
and 0.01% Triton-X-100 (blocking solution) for 2 h. Cells were incubated
overnight with primary antibodies diluted in blocking solution. Dilutions
were 1:100 for rabbit anti-podocalyxin, and 1:50 for mouse anti-E-Cadherin
and mouse anti-acetylated b-tubulin, 4’,6-diamidino-2-phenylindole (DAPI,
Sigma) was diluted 1:10 000 and TRITC-Phalloidin (Sigma) 1:1000. Gels
were washed extensively with blocking solution, followed by overnight
incubation with fluorophore-conjugated secondary antibodies diluted 1:300
in blocking solution. Gels were washed with PBS and mounted onto slides
with Mowiol. Images were acquired with an OLYMPUS FluoView-1000
laser scanning confocal microscope (Olympus, Hamburg, Germany) using
a 60� PlanApo oil objective (n ¼ 1.1) at 258C.
Acknowledgments
We thank George Ojakian, Hans-Peter Hauri, Enrique Rodriguez-Boulan and
Marino Zerial for antibodies, adenoviruses and plasmids. We are grateful to
Marino Zerial for discussion and comments on the manuscript, and to Eric
Marois and Suzanne Eaton for communicating unpublished results. This
work was supported by EU FP5 contract no. HPRN-CT-2002-00259 and
Transregio SFB-TR13-TPA1.
Supplementary Materials
Figure S1: Accumulation of VSV-G in the Golgi is unaffected by
Rab10Q68L. Coverslip-grown control and Rab10Q68L cells were infected
with VSV-G adenovirus. VSV-G was accumulated in the ER at 39.58C for
6 h and chased out of the ER at 19.58C for 90 min. VSV-G co-localizes with
giantin in control and Rab10Q68L cells (upper and lower panel, respec-
tively), showing that it has left the ER and reached the Golgi. No change
occurred when the incubation at 19.58C was prolonged to 120 min. Scale
bars ¼ 20 mm.
Figure S2: Trypsin-based assay for biosynthetic sorting of VSV-G.
Filter-grown control and Rab10Q68L cells were infected with VSV-G
adenovirus. VSV-G was radioactively labeled for 9 h while accumulating in
the ER at 39.58C and chased to the plasma membrane at 328C for 90 min.
Cells were treated with trypsin from the apical or basolateral side at 48C for
30 min. Full-length VSV-G (VSV-G0) and cleavage products (VSV-GC) were
precipitated with an antibody against the cytoplasmic YFP tag, separated by
SDS–PAGE and quantified by phosphorimager analysis. The ratio of cleaved
to full-length VSV-G was calculated for each sample, so that a measure for
the fraction of VSV-G accessible by trypsin from the apical and basolateral
side was obtained. Samples not treated with trypsin served as background
controls.
Figure S3: Activated Rab10 causes missorting of basolateral but not
apical cargo. Coverslip-grown control and Rab10Q68L cells were infected
with A) VSV-G, B) p75NTR or C) apical VSV-G adenovirus. Cargo was
accumulated for theminimum time necessary to allowmicroscopic analysis
and chased to the plasma membrane at 328C (A, C) or 378C (B) for 90 min.
Immunostaining for podocalyxin and, in (B), neurotrophin receptor. An
apical and a basal optical section are shown for each condition. VSV-G is
missorted to the apical membrane in the presence of Rab10Q68L, but the
polarity of p75NTR and apical VSV-G is largely unchanged. VSV-G is VSV-G
ts045 (gray) with the YTDI sorting motif (red), fused to a spacer (black) and
YFP (yellow). p75NTR is the human neurotrophin receptor (gray). Apical
VSV-G is VSV-G ts045 (gray) fused to YFP (yellow) without a spacer, so that
the basolateral sorting motif (red) is masked. Scale bars ¼ 10 mm.
Figure S4: Apical sorting of influenza virus HA is essentially normal in
the presence of activated Rab10. Filter-grown control and Rab10Q68L
cells were infected with influenza virus and the polarity of HA surface
transport was determined. The accuracy of HA targeting to the apical
membrane is slightly reduced in cells expressing Rab10Q68L. This effect
is minor compared with the severe missorting observed for VSV-G (see
Figure 4B). Data are from an experiment performed in duplicate.
Supplemental materials are available as part of the online article at http://
www.blackwell-synergy.com
References
1. Mostov K, Su T, ter Beest M. Polarized epithelial membrane traffic:
conservation and plasticity. Nat Cell Biol 2003;5:287–293.
2. Horton AC, Ehlers MD. Neuronal polarity and trafficking. Neuron
2003;40:277–295.
3. Schuck S, Simons K. Polarized sorting in epithelial cells: raft clustering
and the biogenesis of the apical membrane. J Cell Sci 2004;117:
5955–5964.
4. Traub LM, Apodaca G. AP-1B: polarized sorting at the endosome.
Nat Cell Biol 2003;5:1045–1047.
5. Rodriguez-Boulan E, Kreitzer G, Musch A. Organization of vesicular
trafficking in epithelia. Nat Rev Mol Cell Biol 2005;6:233–247.
6. Schuck S, Simons K. Controversy fuels trafficking of GPI-anchored
proteins. J Cell Biol 2006;172:963–965.
7. Griffiths G, Simons K. The trans Golgi network: sorting at the exit site of
the Golgi complex. Science 1986;234:438–443.
8. Matter K, Mellman I. Mechanisms of cell polarity: sorting and transport
in epithelial cells. Curr Opin Cell Biol 1994;6:545–554.
9. Zerial M, McBride H. Rab proteins as membrane organizers. Nat Rev
Mol Cell Biol 2001;2:107–117.
10. Grosshans BL, Ortiz D, Novick P. Rabs and their effectors: achieving
specificity in membrane traffic. Proc Natl Acad Sci U S A 2006;103:
11821–11827.
Traffic 2007; 8: 47–60 59
Rab10 in Basolateral Transport

11. Salminen A, Novick PJ. A ras-like protein is required for a post-Golgi
event in yeast secretion. Cell 1987;49:527–538.
12. Pereira-Leal JB, Seabra MC. Evolution of the Rab family of small
GTP-binding proteins. J Mol Biol 2001;313:889–901.
13. AngAL, Folsch H, Koivisto UM, PypaertM,Mellman I. The Rab8GTPase
selectively regulates AP-1B-dependent basolateral transport in polarized
Madin-Darby canine kidney cells. J Cell Biol 2003;163:339–350.
14. Kohler K, Louvard D, Zahraoui A. Rab13 regulates PKA signaling during
tight junction assembly. J Cell Biol 2004;165:175–180.
15. Gurkan C, Lapp H, Alory C, Su AI, Hogenesch JB, Balch WE. Large-
scale profiling of Rab GTPase trafficking networks: the membrome.
Mol Biol Cell 2005;16:3847–3864.
16. Chen YT, Holcomb C, Moore HP. Expression and localization of two
low molecular weight GTP-binding proteins, Rab8 and Rab10, by
epitope tag. Proc Natl Acad Sci U S A 1993;90:6508–6512.
17. Chen CC, Schweinsberg PJ, Vashist S, Mareiniss DP, Lambie EJ, Grant
BD. RAB-10 is required for endocytic recycling in the C. elegans
intestine. Mol Biol Cell 2006;17:1286–1297.
18. Babbey CM, Ahktar N, Wang E, Chen CC, Grant B, Dunn KW. Rab10
regulates membrane transport through early endosomes of polarized
Madin-Darby canine kidney cells. Mol Biol Cell 2006;17:3156–3175.
19. Chen W, Feng Y, Chen D, Wandinger-Ness A. Rab11 is required for
trans-Golgi network-to-plasma membrane transport and a preferential
target for GDP dissociation inhibitor. Mol Biol Cell 1998;9:3241–3257.
20. Junutula JR, de Maziere AM, Peden AA, Ervin KE, Advani RJ, van Dijk
SM, Klumperman J, Scheller RH. Rab14 is involved in membrane
trafficking between the Golgi complex and endosomes. Mol Biol Cell
2004;15:2218–2229.
21. Stenmark H, Parton RG, Steele-Mortimer O, Lutcke A, Gruenberg J,
Zerial M. Inhibition of rab5 GTPase activity stimulates membrane fusion
in endocytosis. EMBO J 1994;13:1287–1296.
22. Riederer MA, Soldati T, Shapiro AD, Lin J, Pfeffer SR. Lysosome
biogenesis requires Rab9 function and receptor recycling from endo-
somes to the trans-Golgi network. J Cell Biol 1994;125:573–582.
23. Lock JG, Stow JL. Rab11 in recycling endosomes regulates the sorting
and basolateral transport of E-cadherin.Mol Biol Cell 2005;16:1744–1755.
24. Keller P, Toomre D, Diaz E, White J, Simons K. Multicolour imaging of
post-Golgi sorting and trafficking in live cells. Nat Cell Biol 2001;3:140–149.
25. Ang AL, Taguchi T, Francis S, Folsch H, Murrells LJ, Pypaert M,Warren
G, Mellman I. Recycling endosomes can serve as intermediates during
transport from the Golgi to the plasmamembrane of MDCK cells. J Cell
Biol 2004;167:531–543.
26. Kauppi M, Simonsen A, Bremnes B, Vieira A, Callaghan J, Stenmark H,
Olkkonen VM. The small GTPase Rab22 interacts with EEA1 and
controls endosomalmembrane trafficking. J Cell Sci 2002;115:899–911.
27. Yi Z, Yokota H, Torii S, Aoki T, Hosaka M, Zhao S, Takata K, Takeuchi T,
Izumi T. The Rab27a/granuphilin complex regulates the exocytosis of
insulin-containing dense-core granules. Mol Cell Biol 2002;22:
1858–1867.
28. Griffiths G, Pfeiffer S, Simons K, Matlin K. Exit of newly synthesized
membrane proteins from the trans cisterna of the Golgi complex to the
plasma membrane. J Cell Biol 1985;101:949–964.
29. Matlin KS, Simons K. Sorting of an apical plasma membrane glycopro-
tein occurs before it reaches the cell surface in cultured epithelial cells.
J Cell Biol 1984;99:2131–2139.
30. Maisner A, Zimmer G, Liszewski MK, Lublin DM, Atkinson JP,
Herrler G. Membrane cofactor protein (CD46) is a basolateral protein
that is not endocytosed. Importance of the tetrapeptide FTSL at the
carboxyl terminus. J Biol Chem 1997;272:20793–20799.
31. Folsch H, Ohno H, Bonifacino JS, Mellman I. A novel clathrin adaptor
complex mediates basolateral targeting in polarized epithelial cells. Cell
1999;99:189–198.
32. Roh MH, Fan S, Liu CJ, Margolis B. The Crumbs3-Pals1 complex
participates in the establishment of polarity in mammalian epithelial
cells. J Cell Sci 2003;116:2895–2906.
33. Lapierre LA, Dorn MC, Zimmerman CF, Navarre J, Burnette JO,
Goldenring JR. Rab11b resides in a vesicular compartment distinct
from Rab11a in parietal cells and other epithelial cells. Exp Cell Res
2003;290:322–331.
34. Futter CE, Connolly CN, Cutler DF, Hopkins CR. Newly synthesized
transferrin receptors can be detected in the endosome before they
appear on the cell surface. J Biol Chem 1995;270:10999–11003.
35. Vitale G, Rybin V, Christoforidis S, Thornqvist P, McCaffrey M,
Stenmark H, Zerial M. Distinct Rab-binding domains mediate the inter-
action of Rabaptin-5 with GTP-bound Rab4 and Rab5. EMBO J 1998;
17:1941–1951.
36. Rink J, Ghigo E, Kalaidzidis Y, Zerial M. Rab conversion as a mecha-
nism of progression from early to late endosomes. Cell 2005;122:
735–749.
37. Ortiz D, Medkova M, Walch-Solimena C, Novick P. Ypt32 recruits the
Sec4p guanine nucleotide exchange factor, Sec2p, to secretory
vesicles; evidence for a Rab cascade in yeast. J Cell Biol 2002;157:
1005–1015.
38. Nuoffer C, Davidson HW, Matteson J, Meinkoth J, Balch WE. A
GDP-bound of rab1 inhibits protein export from the endoplasmic
reticulum and transport between Golgi compartments. J Cell Biol 1994;
125:225–237.
39. Barral DC, Ramalho JS, Anders R, Hume AN, Knapton HJ, Tolmachova
T, Collinson LM, Goulding D, Authi KS, Seabra MC. Functional redun-
dancy of Rab27 proteins and the pathogenesis of Griscelli syndrome.
J Clin Invest 2002;110:247–257.
40. Schluter OM, Schmitz F, Jahn R, Rosenmund C, Sudhof TC. A
complete genetic analysis of neuronal Rab3 function. J Neurosci 2004;
24:6629–6637.
41. Wang X, Kumar R, Navarre J, Casanova JE, Goldenring JR. Regulation
of vesicle trafficking in madin-darby canine kidney cells by Rab11a and
Rab25. J Biol Chem 2000;275:29138–29146.
42. Pralle A, Keller P, Florin EL, Simons K, Horber JK. Sphingolipid-cholesterol
rafts diffuse as small entities in the plasma membrane of mammalian
cells. J Cell Biol 2000;148:997–1008.
43. Ehehalt R, Keller P, Haass C, Thiele C, Simons K. Amyloidogenic
processing of the Alzheimer beta-amyloid precursor protein depends
on lipid rafts. J Cell Biol 2003;160:113–123.
44. Meder D, Moreno MJ, Verkade P, Vaz WL, Simons K. Phase coexis-
tence and connectivity in the apical membrane of polarized epithelial
cells. Proc Natl Acad Sci U S A 2006;103:329–334.
45. Schuck S, Manninen A, Honsho M, Fullekrug J, Simons K. Generation
of single and double knockdowns in polarized epithelial cells by
retrovirus-mediated RNA interference. Proc Natl Acad Sci U S A 2004;
101:4912–4917.
60 Traffic 2007; 8: 47–60
Schuck et al.


















