
3 sem final version
51
Structural Basis for Cooperation of Hsp70 and Hsp110 in Protein Folding Volume 133, Issue 6, 13 June 2008, Pages 1068-1079 Sigrun Polier, Zdravko Dragovic, F. Ulrich Hartl and Andreas Bracher Department of Cellular Biochemistry, Max-Planck-Institute of Biochemistry, Germany
Transcript of 3 sem final version

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 1/51
Structural Basis for Cooperation of
Hsp70 and Hsp110 in Protein Folding
Volume 133, Issue 6, 13 June 2008, Pages 1068-1079
Sigrun Polier, Zdravko Dragovic, F. Ulrich Hartl and Andreas Bracher
Department of Cellular Biochemistry, Max-Planck-Institute of Biochemistry,Germany

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 2/51
Hsp70 chaperones
� protein folding,
� prevention of aggregation,
� protein transport across membranes,� and protein degradation
[Bakau et al, 2006] and [ Hartl and Haver-Hartl,2002]
8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 3/51
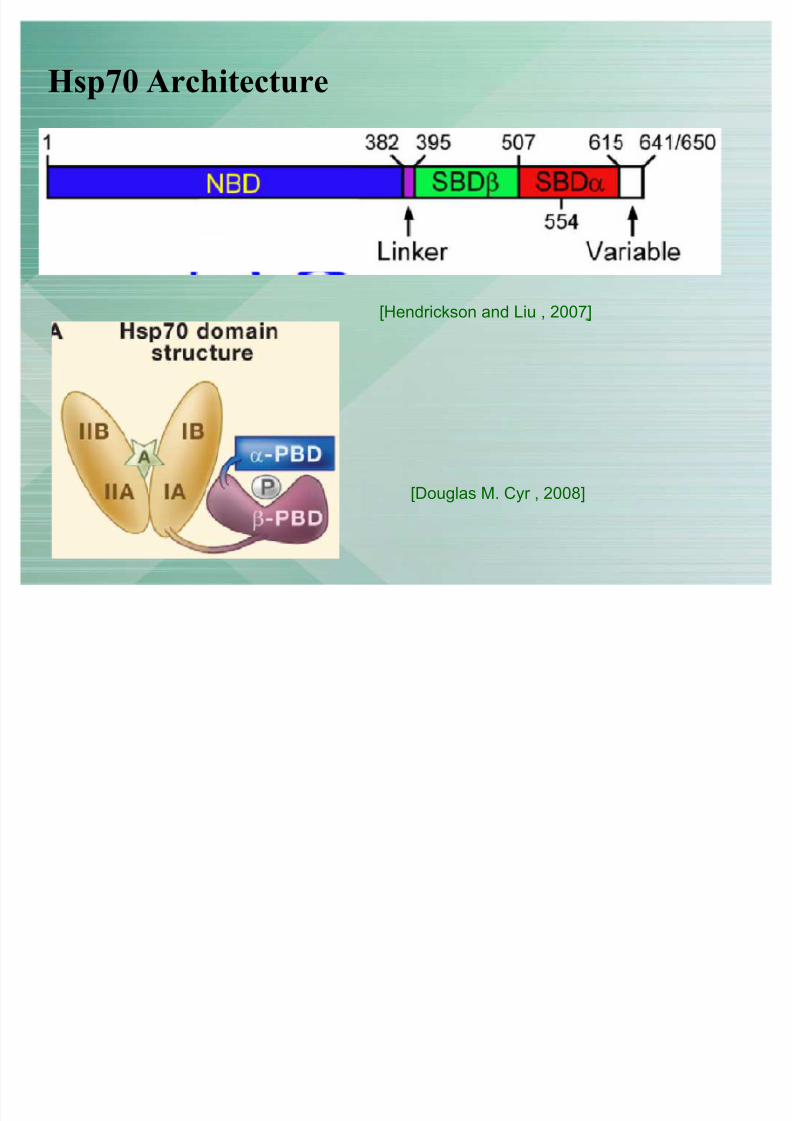
Hsp70 Architecture
[Douglas M. Cyr , 2008]
[Hendrickson and Liu , 2007]

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 4/51
� J-domain proteins (eg. Hsp40) trigger ATP hydrolysis by Hsp70.[Mayer and Bukau, 2005]
� NEFs (eg Hsp110) catalyze the release of ADP from Hsp70 [Liberek et al.,1991] and [Szabo et al., 1994]
Hsp70s require two additional proteins

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 5/51
Hsp70 and corresponding Hsp110 NEFs
[Wayne A. Hendrickson, Qinglian Liu 2008]

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 6/51
� These NEF proteins are only partially redundant and fulfill specialized roles. (Fes1p,Sse1p, Sse2p )
� Dysfunction of the ER NEF (Sil1/BAP) causes neurodegenerative diseaseMarinesco-Sjögren syndrome (Grp170 is working) [Anttonen et al., 2005] and[Senderek et al., 2005]
� Hsp110/Grp170 proteins are themselves Hsp70 homologs and were reported to bindand stabilize unfolded proteins [Goeckeler et al., 2002], [Oh et al., 1997] and [Ohet al., 1999]
� However, Hsp110 homologs were found unable to fold proteins independently of regular Hsp70s (Easton et al., 2000).
Previous sudies on NEF and Hsp110

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 7/51
� The recently determined crystal structure of Sse1p.ATP confirmed theHsp70-like domain composition of Hsp110 [Liu and Hendrickson, 2007].
� The NBD and PBD being tightly associated. In particular, the -helical partof the PBD does not form a ³lid´ over the peptide-binding pocket
� as observed in the structure of the isolated PBD of Hsp70 [Zhu et al.,1996], but instead interacts with the flank of the NBD and points awayfrom the sandwich domain.
Previous sudies cont..

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 8/51
Comparison of Sse1p structure with DnaK(Hsp70)
[Douglas M. Cyr , 2008]
[Hendrickson and Lir,2007]
Sse1p
Hsp70-PBD

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 9/51
Objective of study
� How Hsp70 and Hsp110 interact
� Conformational changes in the interaction partners
� Find role of Sse1p in protein release,
� Protein folding
� Understand Sse1p holdase activity
� Make mutations at key residues and see in vitro and in vivo effects.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 10/51
Approach in Short
� Solved structure of Sse1p.ATP(y) + Hsp70N(h)
� Use the structural information to make mutations in Sse1p partner Ssa1N(yHsp70)

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 11/51
What are the requirements for for Sse1p-
Hsp70 complex formation and NEF activity ?

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 12/51
1. Size exclusion chromatography/ Gel filtration.
� no nucleotide- Sse1p and Ssa1N eluted seperatly
� ATP - Sse1p and Ssa1N eluted together
� ADP - Sse1p and Ssa1N eluted together ± lesser extent compared to ATP
2. Free flow electrophoresis MALLS
binary complex of approximately 101.1 kDa
3. Kd-0.4 M for this complex
Determining requirements for Complex formation
beteween Sse1p and Ssa1p

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 13/51
MABA-ADP exhange from Ssa1- NEF activity
�Interaction of cytosolic Hsp110swith the NBD of Hsp70 is sufficient
for nucleotide exchange
�For NEF activity Sse1p must be inSse1p.ATP form
�Structure of Ssa1pN and Hsp70N
is similar
N8-(4-N -methylanthraniloylaminobutyl)-8-aminoadenosine 5 -diphosphate (MABA-ADP)

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 14/51
Crystallization
� Hsp70N and Sse1-loop, in which the poorly conserved and presumablyflexible insertion (residues 503±524) found in all Hsp110/Hsp170 homologsis replaced by the peptide linker sequence AGSD
� The structure was solved by Se-MAD, and the model was refined against
isomorphous native data at 2.3 Å resolution to a crystallographic R factor of 19.7% (Rfree 24.4%)
� The asymmetric unit contains two virtually identical copies of the complex
� clear electron density could not be assigned to the non-natural linker sequence and to the C-terminal segment of Sse1p (residues 656±693)

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 15/51

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 16/51
Domain organization of Sse1p is like Hsp70 proteins
�N-terminal Hsp70/actin-type NBD, a sandwich domain, and a C-terminalthree helix bundle domain (3HBD)
� sandwich and 3HBD are arrangedalong the NBD and point intoopposite directions.
�All interactions with the N of Hsp70are mediated by the N and the 3H of Sse1p.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 17/51
NBDs of Sse1p and Hsp70 are oriented
toward each other
IIb and Ib, IIa and Ia, and Ib and IIb of Sse1p and Hsp70, respectively
Sse1p Hsp70

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 18/51
3HBD contacts Subdomain IIb

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 19/51
3HBD-IIb interface
�1760 Å2 of surface on each bindingpartner Sc - 0.74
�N terminus of helix 9IIb is in closecontact to the side chain of Asn572S
�Mainly H-bonds and wdWaalscontacts
�Mutation Asn572S, Glu575, Ala300

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 20/51
NBD-NBD interface
�Gln33H direct contacts adenineNH2
�Gln33H vdWaals Ala280S and Ser279S
�Ala280S intercalating between Ala54Hand Gln33H
�Asn281S, hydrogen bond Asn57H.
�Arg346S side chain flanks the nucleobaseof ATP and forms a -stacking interaction
with the peptide bond between Gln33Hand Gly34H
�Mutation A280, N281, Q33

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 21/51
At NBD-NBD interface Sse1p and Hsp70 contact
with the nucleotide which explains the
requirement of ATP for complex formation

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 22/51
One more area of surface complementarity at
residues 364± 367 in Sse1p
�Sse1p contact Hsp70 backbone at133H and 134H as well as the sidechain of Tyr134H
�Hydrogen bonds of Thr365S andAsn367S shape the surface of Sse1pinto a conformation that cradlesTyr134H
�Mutatiion T365, N367

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 23/51
Conformational Features of Sse1p Critical for NEF
Activity
�Sse1p·ATP is characterized by atwisted arrangement of the twolobes of the NBD, in whichsubdomain IIb is significantlydisplaced when compared with the
structure of the ADP-NBD of Hsc70
�twisted lobe arrangement isstabilized by backbone contactsfrom both lobes to the phosphate
of ATP
�and by interdomain interactionswith the linker segment (residues385±399), the sandwich domain,
and the N-terminal segment of the3HBD

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 24/51
Sse1p interface N-ter 3HBD and flank of lobe I
NBD ensures the correct positioning of the 3HBD
�Polar interactions of residue Arg47 withGlu554 and Asp561
�Glu554 is replaced by Ala in canonicalHsp70s and by larger hydrophobic residues,
such as Leu and Val, in ER-residentHsp110s
�in the PBD of the bacterial Hsp70, DnaK,the equivalent residues of the 3HBD bind tothe sandwich domain, thereby enclosingthe bound substrate
Mutation E554, M557, L558

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 25/51
Mutations studied for Sse1p and Ssa1p

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 26/51
Mutations affecting the complex formation
�Mutations targeting the Sse1p-Hsp70 NBD interface (Sse1-2, Sse1-3,Sse1-4, Ssa1N[Q31A], and Ssa1N[A300E]) reduced binding to a low basallevel.
�structural integrity of Sse1p appears to be essential for the high-affinityinteraction with Hsp70(Sse1-5, Sse1-7, and Sse1-8)
�ATP hydrolysis (Sse1-1) is not required for complex formaion.
�Mutations in the sandwich domain(Sse1-9 and Sse1-10) did not impair
binding to Hsp70NBD
8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 27/51
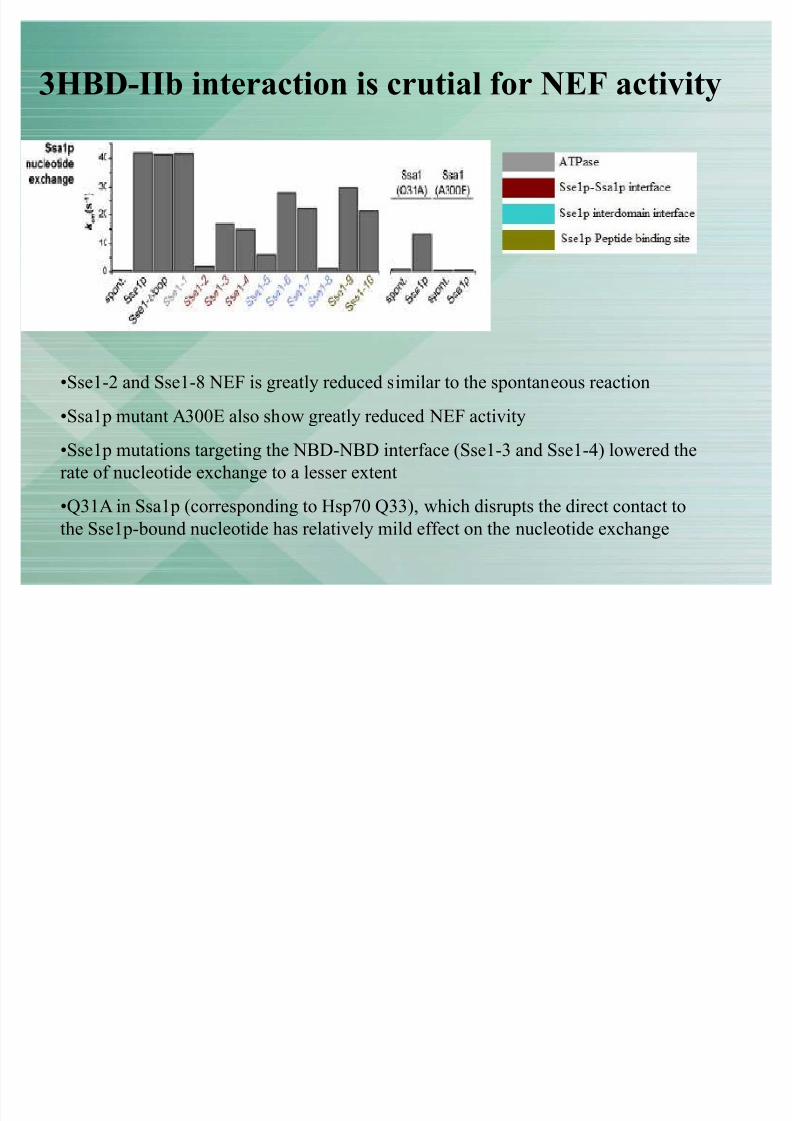
3HBD-IIb interaction is crutial for NEF activity
�Sse1-2 and Sse1-8 NEF is greatly reduced similar to the spontaneous reaction
�Ssa1p mutant A300E also show greatly reduced NEF activity�Sse1p mutations targeting the NBD-NBD interface (Sse1-3 and Sse1-4) lowered therate of nucleotide exchange to a lesser extent
�Q31A in Ssa1p (corresponding to Hsp70 Q33), which disrupts the direct contact tothe Sse1p-bound nucleotide has relatively mild effect on the nucleotide exchange

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 28/51
Linker-NBD interaction is important for NEF
activity
�Mutation of the linker-NBD interface of Sse1p (Sse1-5) also strongly attenuated
nucleotide exchange
�other domain-domain interface mutants (Sse1-6 and Sse1-7) as well as
mutations directed at the putative peptide-binding site (Sse1-9 and Sse1-10)
exhibited nucleotide exchange activities more similar to unmodified Sse1p.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 29/51
Multiple mutations are needed to disrupt Sse1p
and Ssa1p interaction
�Sse1-(2+3) and Sse1-(2+4) have similarly low nucleotide exchange activities asmutants Sse1-2 and Sse1-8
�10-fold molar excess of Sse1-2 or Sse1-8 accelerated nucleotide exchangeroughly by a factor of 10, this effect cannot be seen for Sse1-(2+3) and Sse1-(2+4)

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 30/51
R ole of Sse1p NEF activity in
substrate turnover by Hsp70(Ssa1p) ?

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 31/51
Dissociation of D-NR from Hsp70 in the presence
of excess NR
�Sse1p mutants with a mild NEF defect were still able to stimulate peptide release
�However, in mutants with severely impaired NEF activity, nucleotide exchangebecame rate limiting for peptide release.
D-NR (dansyl-NRLLLTGC)

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 32/51
Sse1p contribution to Hsp70-assisted protein
folding?

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 33/51
Sse1p:Ssa1P ratio 1:6 shows maximum folding
activity
whereas equimolar concentrations of Sse1p with Ssa1P inhibited refolding
8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 34/51
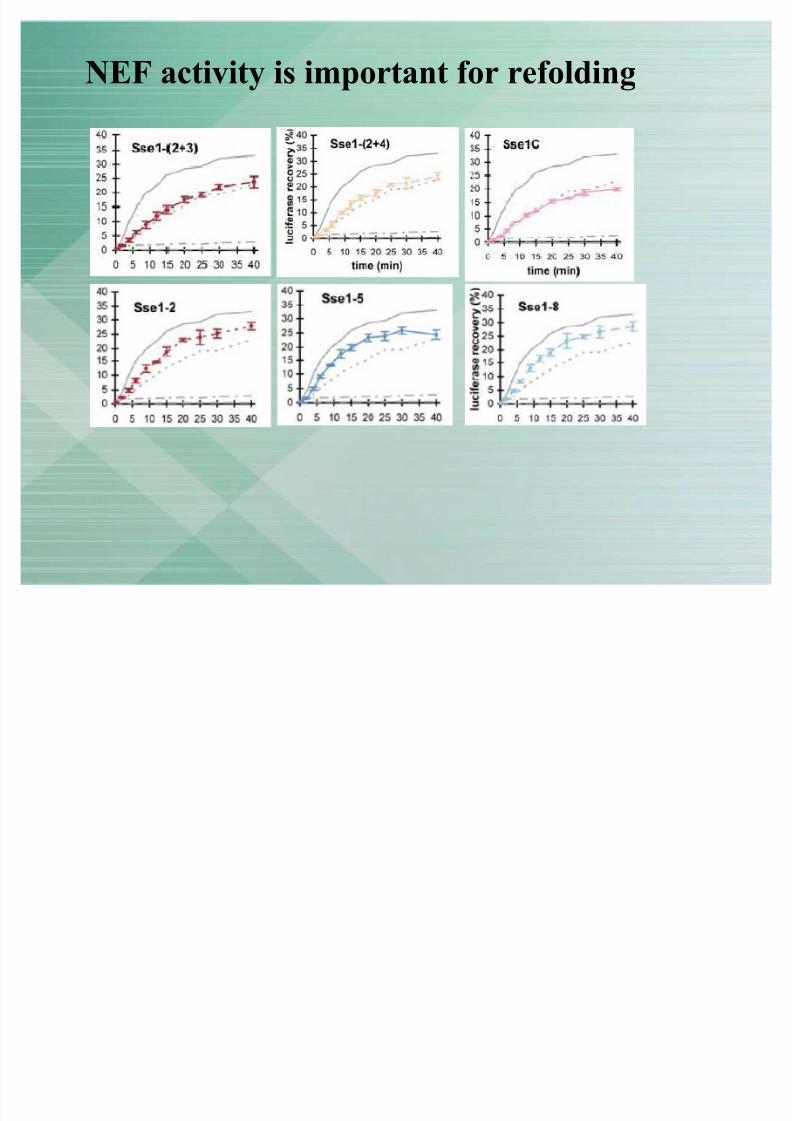
NEF activity is important for refolding

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 35/51
Peptide binding site mutation affects refolding
ATPase activity of Sse1p is not required for refolding activity

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 36/51
Thermal transition in Sse1p is requried for
Holdase activity

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 37/51
Invivo Effects ± NEF activity is critical for cell
vialibilty
�NEF is crutial for cell survival
�3HBD-IIb interaction is crutial for cell survival at higher temperature
�PBD mutations become apparent only
when combined with severe mutationin NEF activity

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 38/51
Model for the Cooperation of Hsp110 and Hsp70 in Protein Folding
Step1. Recruitment of Hsp70 (red)
to unfolded substrate protein
(green) is assisted by J-domain
proteins (Hsp40)
Step2. Complex formation
between Hsp70 and Hsp110 (blue)
displaces ADP from the Hsp70
Step3. upon binding of ATP to
Hsp70, the Hsp70-Hsp110
complex dissociates and the
substrate protein is released for folding

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 39/51
Summary
NBD of Hsp70 is embraced by the NBD and the 3HBD of Sse1pATP in NBD of Sse1p is required for complex formation
ATP in (Sse1p) NBD causes the two NBD lobes to adopt twistedconformation which inturn induces nucleotide free state in Hsp70NBD
Hsp110 assist protein folding by-
1. by transient interaction with Hsp70 ±nucleotide exhange(major function)
2. together with Hsp70 it can provide platform for conformationalremodeling of the substrate
Refolding occurs at stoichiometric concentrations of Sse1p to Ssa1p and
high concentrations of Sse1p inhibits refoldingSse1p efficiently stabilized thermally denatured luciferase in a folding-competent state, demonstrating the ability of Hsp110 to interact withunfolded proteins
ATP hydrolysis and allosteric rearrangements is dispensable for the
functional cycle of Sse1p

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 40/51
Future directions
Sse1p structure shows groove-like indentions running
between domain protrusions that could possibly
accommodate exposed helices or hairpins in
unfolded substrates ?
The role of Sse1p ATPase and its regulation ?
Determine the specialized role of NEFs in cellular compartment ?

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 41/51
Sse1p structure provides a groove-like indention
running between domain protrusions

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 42/51
Thank you !!

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 43/51
Additionally, all Hsp110/Grp170 proteins share a
large insertion in the sandwich domain and a
C-terminal extension, features that are not
present in canonical Hsp70s (Easton et al.,2000).

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 44/51

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 45/51
Hsp70 subdomains

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 46/51
Method of seperation based on EPM/pIs
�no stationary phase
�Sample to buffer film
�Electric field applied IIr toelectrolyte flow

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 47/51
MALLS
� scattering measurements are made at a number of differentangles, none of which are close to the incident light.
� This significantly reduces problems associated with light
scattering from particulate contaminants.
� The intensity of the scattered light is proportional to themolecular weight.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 48/51
MAD
� Multi-wavelength anomalous dispersion (sometimesMulti-
wavelength anomalous diffraction; abbreviated MAD) is a
technique used in X-ray crystallography that facilitates the
determination of the structure of proteins by allowing the solution of
the phase problem. This is possible if the structure contains one or
more atoms that cause significant anomalous scattering from
incoming X-rays at the wavelength used for the diffraction experiment.
Atoms in proteins which are suitable for this purpose are sulfur or
heavier atoms, for example metal ions in metalloproteins. The most
commonly used atom for phase determination via MAD, however, is
selenium, since it is usually possible to replace the natural sulfur containing amino acid methionine by selenomethionine.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 49/51
R-Factor
� In crystallography, the R-factor (sometimes called residualfactor or reliability factor) is a measure of the agreementbetween the crystallographic model and the experimental X-raydiffraction data.
� The R factor usually ranges between 0.6 (when comparing arandom set of reflections with a given model) and 0.2 (for example for a well refined macro-molecular model at aresolution of 2.5 Angstrom). When working on small molecules(up to 300 atoms), which form extremely well ordered crystals,
it is possible to attain R-factors as low as 0.015.Crystallographers also use the Free R-Factor and thesymmetric R-Factor to describe the quality of a model.

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 50/51
II stacking interaction
� an aromatic interaction (or - interaction) is a
noncovalent interaction between organic compounds
containing aromatic moieties.
� - interactions are caused by intermolecular overlapping
of p-orbitals in -conjugated systems, so they become
stronger as the number of -electrons increases

8/7/2019 3 sem final version
http://slidepdf.com/reader/full/3-sem-final-version 51/51
X-ray
� the study of crystal structures through X-ray diffraction
techniques. When an X-ray beam bombards a crystalline
lattice in a given orientation, the beam is scattered in a
definite manner characterized by the atomic structure of the
lattice. This phenomenon, known as X-ray diffraction,occurs when the wavelength of X-rays and the interatomic
distances in the lattice have the same order of magnitude.


















