2008 Rev Mitoc Lactate ( Cajigal)

of 9
-
Upload
burgoschile -
Category
Documents
-
view
217 -
download
0
Transcript of 2008 Rev Mitoc Lactate ( Cajigal)
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
1/9
Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
Mitochondrial Lactate Oxidation Complex andan Adaptive Role for Lactate Production
TAKESHI HASHIMOTO and GEORGE A. BROOKS
Exercise Physiology Laboratory, Department of Integrative Biology, University of California, Berkeley, CA
ABSTRACT
HASHIMOTO, T. and G. A. BROOKS. Mitochondrial Lactate Oxidation Complex and an Adaptive Role for Lactate Production. Med.Sci. Sports Exerc. , Vol. 40, No. 3, pp. 486494, 2008. The intracellular lactate shuttle (ILS) hypothesis holds that lactate produced asthe result of glycolysis and glycogenolysis in the cytosol is balanced by oxidative removal in mitochondria of the same cell. Also, theILS is a necessary component of the previously described cellcell lactate shuttle (CCLS), because lactate supplied from the interstitiumand vasculature can be taken up and used in highly oxidative cells (red skeletal and cardiac myocytes, hepatocytes, and neurons). ThisILS emphasizes the role of mitochondrial redox in creating the proton and lactate anion concentration gradients necessary for theoxidative disposal of lactate in the mitochondrial reticulum during exercise and other conditions. The hypothesis was initially supported by direct measurement of lactate oxidation in isolated mitochondria as well as findings of the existence of mitochondrialmonocarboxylate transporters (mMCT) and lactate dehydrogenase (mLDH). Subsequently, the presence of a mitochondrial lactateoxidation complex (composed of mMCT1, CD147 (basigin), mLDH, and cytochrome oxidase (COX)) was discovered, which lends
support to the presence of the ILS. Most recently, efforts have been made to evaluate the role of lactate as a cell-signaling molecule (i.e.,a lactormone) that is involved in the adaptive response to exercise. Lactate is capable of upregulating MCT1 and COX gene and protein expression. Current findings allow us to understand how lactate production during exercise represents a physiological signal for the activation of a vast transcription network affecting MCT1 protein expression and mitochondrial biogenesis, thereby explaining howtraining increases the capacity for lactate clearance via oxidation. Key Words: SKELETAL MUSCLE, REACTIVE OXYGENSPECIES, MCT1, CD147, CYTOCHROME OXIDASE, LDH
W e thank Dr. Bruce Gladden for organizing thissymposium on the lactate shuttle and meetingorganizers for the opportunity to inform readersof recent work on the subject. As such, this paper is not a
comprehensive review, because such are available (10,33),and other papers in the symposium extend the scope of our present effort.
Once thought to be formed as the result of oxygen lack,we now know that lactate is produced continuously under fully aerobic conditions, especially during exercise whenrates of glycogenolysis and glycolysis are elevated (10,62).Also, from results of isotope tracer, arterialvenous differ-
ence mass balance, and biopsy studies (5,13), we now knowthat working skeletal muscle is not only the major site of lactate production, but also the major site of its removal,mainly via oxidation.
The carboxylic acids lactate and pyruvate are exchangedacross lipid bilayer membranes by facilitated, proton-linkedtransport (63,64,70), involving a family of monocarboxylatetransport (MCT) proteins (31). MCT1 is widely expressedin different tissues (34) and has been localized at plasma and mitochondrial membranes of muscle and other cells,including astrocytes and neurons (12,18,35,38,48,49,56).As part of the cellcell lactate shuttle (CCLS) mechanism,MCT1 facilitates uptake of lactate into working humanskeletal muscle from interstitium and plasma (5,27). A goodexample is the shuttle of lactate from working muscle to theheart, in which lactate is oxidized (32).
The seminal CCLS hypothesis (9) posits that lactate is
either used within cells of formation or is exported toadjacent and anatomically distributed cells, tissues, andorgans for use. Hence, lactate was thought to represent a carbon source for oxidation and gluconeogenesis as well asa vehicle for cellcell signaling via redox changes (10,11).At present, there is good agreement on key elements of theCCLS, but there are fewer data and less general agreement on the cellular sites of lactate oxidation (33). Therefore, themain purpose of this brief review is to highlight the recent evidence that cellular mitochondrial networks are sites of
Editors Note: This paper is an Editor-in-Chiefinvited contribution fromthe ACSMs conference on Integrative Physiology on Exercise held inIndianapolis, Indiana, September 2730, 2006.
Address for correspondence: George A. Brooks, Ph.D., Department of Integrative Biology, 5101 VLSB, University of California, Berkeley CA94720-3140 USA; E-mail: [email protected] for publication May 2007.Accepted for publication October 2007.
0195-9131/08/4003-0486/0MEDICINE & SCIENCE IN SPORTS & EXERCISE Copyright 2008 by the American College of Sports Medicine
486
B A S I C S C I E N C E S
DOI: 10.1249/MSS.0b013e31815fcb04
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
2/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
lactate disposal via a lactate oxidation complex (35). Asecondary purpose is further understanding of the coordi-nation between muscle lactate production and mitochondrial biogenesis (36).
Intracellular Lactate Shuttle Hypothesis
The intracellular lactate shuttle (ILS) hypothesis (11)
posits that lactate produced as the result of glycolysis andglycogenolysis in the cytosol is balanced by oxidation inmitochondria of the same cell. This mitochondrial lactateoxidation is plausible when energetics of lactate dehydro-genase (LDH) are considered. Lactate production occurs inmuscle cytosol because the K eq of LDH is very high (3.6 10 4), and muscle isoforms with low K M values for pyruvate predominate (27), making lactate, and not pyruvate, the predominant end-product of glycolysis. Especially duringexercise, muscle glucose uptake and glycogenolysis risemany times, causing pyruvate and lactate concentrationsand the [lactate]/[pyruvate] ratio to rise (39,68). Lactateoxidation to pyruvate is unlikely in the cytosol of contract-ing muscle. In contrast to NAD +/NADH redox couplechanges in the cytosol during muscle contractions, themitochondrial NAD + /NADH pool becomes relatively moreoxidized during exercise than at rest (41). The high[lactate], [lactate]/[pyruvate], and [H +] in cytosol are juxtaposed to low monocarboxylate and H + concentrationsin the mitochondrial matrix. These chemical and proton
concentration gradients favor mitochondrial lactate influxand oxidation because the environment facilitates removalof both pyruvate (via TCA cycle) and H + (via themitochondrial ATPase). The presence of MCT1 and LDHin mitochondria allows mitochondrial lactate influx andoxidation. Evidence supporting, and not supporting, the ILShypothesis is summarized in Tables 1 and 2, respectively.With regard to Table 2, Yoshida et al. (74) have recently
reported on mitochondrial preparations low in LDH content and with poor lactate oxidation capability. Our view is that the harsh homogenization, protease, and Percoll gradient treatments caused loss of mLDH (18), which is nowrecognized to be a constituent of the mitochondrial proteome (69). According to them, for their preparationsto respire lactate at a rate equivalent to pyruvate, the lactateconcentration would have to be one to two orders of magnitude greater; but such is the case in vivo, according toa previous paper from the same group (68). Therefore,while in their Table 2 the authors listed their results as not supporting the ILS hypothesis (74), in our view, even withLDH-depleted mitochondrial preparations, they essentially prove the ILS hypothesis for in vivo conditions. Conceptualand technical filings of the paper by Yoshida et al. aredescribed in our recent letter to editor of the Journal of Physiology (15), as well as on the Internet (http://ib. berkeley.edu/labs/brooks/Comnts_on_Yoshida.pdf). Also,in a soon-to-be-published report, Anna Atlante, Lidia deBari, Antonella Bobba, Ersilia Marra, and Salvatore
TABLE 1. Key evidence supporting the intracellular lactate shuttle hypothesis.
1. Localization of LDH in mitochondriaa. Electron microscopy showed LDH to be associated with the inner membrane and matrix of rat skeletal and cardiac muscle mitochondria (1).b. Electron microscopy showed intramitochondrial localization of LDH in rat heart and liver mitochondria (14).c. LDH isoforms found in isolated human skeletal muscle mitochondria (27).d. Confocal laser scanning microscopy (CLSM) detected mitochondrial LDH in rat musclederived L6 cells (35).
2. Localization of MCT1 in mitochondriaa. In discovery of MCT1, Garcia and colleagues (31) produced micrographs showing very similar localizations of MCT1 and the mitochondrial marker, SDH. Although not
colocalization studies, muscle SDH and MCT1 image patterns are remarkably similar.b. Electron microscopy showed that MCT1 is abundant in rat heart and skeletal muscle mitochondria (12).c. Western blotting shows MCT1 in isolated human skeletal muscle mitochondria (27).d. Confocal laser scanning microscopy (CLSM) showed MCT1 was colocalized with subsarcolemmal and intermyofibrillar mitochondria in rat plantaris muscle (38).e. CLSM showed colocalization of MCT1 and mitochondria in rat L6 cells (35).
3. Isolated mitochondria contain LDH, MCT1, and its chaperone protein CD147.a. LDH was present in rat liver, kidney, and heart mitochondria by agarose gel electrophoresis as well as spectrophotometric assays after cell fractionation (8,45).b. Electrophoretic analysis demonstrated mitochondrial LDH in rat liver, muscle, and heart (14,27).c. Western blots showed mitochondrial MCT1 and CD147 in rat, but proteolytic digestion resulted in loss of those proteins (18).d. Subsarcolemmal mitochondria obtained from rat skeletal muscle contained MCT1, but intermyofibrillar mitochondria did not contain MCT1 by Western blotting after cell
fractionation (3).e. Western blots and immunoprecipitation demonstrated that MCT1, CD147, and LDH were localized at mitochondria inner mitochondrial membrane associated with cytochrome
oxidase (COX) in L6 cells (35).4. Physiological data of mitochondrial lactate oxidation
a. Isolated rat liver mitochondria were capable of oxidizing lactate faster than pyruvate (45).b. Isolated rat muscle mitochondria were able to oxidize lactate faster than pyruvate (14).c. Isolated rat liver mitochondria were able to take up and metabolize L-lactate via mitochondrial LDH and an L-lactate/H+ symporter as well as an L-lactate/pyruvate antiporter (26).
5. Magnetic resonance spectroscopy (MRS) is supportive of mitochondrial, and not cytosolic, lactate oxidation.a. Infusion of 13C-pyruvate into working canine hearts labeled cytosolic alanine and lactate pools. In contrast, 13C-lactate did not label either pyruvate or alanine; instead,
glutamate was labeled with 13C from tracer lactate (46).b. Comparison of glycolytic and exogenous lactate oxidation rates in rat heart demonstrated preferential oxidation of exogenous lactate as well as mitochondrial, but not
cytosolic, lactate oxidation (21). Similar data were provided previously on working rat skeletal muscle (6).6. Studies of brain metabolism clearly show cell-to-cell and intracellular lactate shuttles (65).7. Results of studies on mitochondrial proteome list LDH, basigin, and MCT1 (51,68).8. The presence of mitochondrial LDH and the ability of mammalian (brain) mitochondria to oxidize lactate rereconfirmed (Anna Atlante A, de Bari L, Bobba A, Marra E, Passarella
S. Transport and metabolism of L-lactate occur in mitochondria from cerebellar granule cells and are modified in cells undergoing low potassium dependent apoptosis.Biochim.Biophys . (in press).
THE INTRACELLULAR LACTATE SHUTTLE Medicine & Science in Sports & Exercise d 487
BA SI C S CI EN CE S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
3/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
Passarella yet again show the presence of mLDH, this timein rat cerebellar granule cells.
CD147 (Basigin) Serves to Scaffold MCT1 to the
Mitochondria Inner MembraneAmong the unanswered questions regarding the mecha-
nism of lactate oxidation in vivo has been the identity of themitochondrial chaperone for MCT1 and other associatedlactate oxidation complex constituents. The single-spantransmembrane glycoprotein CD147 (BSG, basigin) isconsidered to be the chaperone protein for MCT1, localiz-ing it to the cell surface (29,44,73). However, few studieshave focused on the association between CD147 and MCT1in the mitochondrial reticulum of mammalian skeletalmuscle cells. As CD147 serves to anchor MCT1 to thesarcolemma, it was reasonable to evaluate whether CD147
also served to scaffold MCT1 to the mitochondrial inner membrane, particularly in subsarcolemmal domains of themitochondrial reticulum. Such a protein complex was posited to provide a means for lactate oxidation in themitochondrial reticulum. Accordingly, we evaluated andvisualized by confocal laser scanning microscopy (CLSM)that CD147 and MCT1 are colocalized throughout rat skeletal muscle-derived L6 cells, including the mitochon-drial reticulum and sarcolemma (35). Supporting that observation of CLSM, our Western blots of cell subfrac-tions demonstrated that CD147 was expressed in themitochondrial fractions of L6 cells and liver (35). Addi-tionally, immunoprecipitation of CD147 from the mito-
chondrial fraction of L6 cells coprecipitated MCT1 (35).Those results confirm and extend the observation of Kirk et al. (44) regarding CD147 as a sarcolemmal chaperonefor MCT1. Previously, they have shown colocalizationof MCT1 and CD147 at the cell surfaces of isolatedrat cardiac cells by using CLSM, and, although their em- phasis was on cell-surface protein expression, their micro-graphs also show colocalization of MCT1 and CD147throughout transfected Hela cells, a finding not mentionedin their text.
To some, the presence of the glycoprotein CD147 inmitochondria might have been a surprise, given its previously documented function as a highly glycosylated protein in the sarcolemma. Although attempts to determinewhether glycosylated proteins are present in mitochondria have been less convincing, the existence of a glycoproteinin rat liver mitochondria was previously demonstrated (20).The molecular weight of the glycoprotein approximated 45kDa, which approximates that of CD147 (42 kDa), and that glycoprotein is a component of the inner mitochondrialmembrane (20). Hence, it was not surprising to us that CLSM, Western blotting of cell subfractions, and immuno- precipitation techniques showed CD147 to colocalize withMCT1 as well as LDH and cytochrome oxidase (COX) at the mitochondrial inner membrane. However, how CD147interacts and functions with MCT1 in plasma and mito-chondrial membranes requires elucidation (44,47,72,73).
The Evidence for a Lactate Oxidation Complex atthe Inner Mitochondrial Membrane
In addition to CD147, LDH was found to be associatedwith the mitochondrial reticulum of L6 cells by immuno-cytochemistry, Western blotting after cell fractionation, andCLSM (35). The interaction of these two proteins (MCT1and CD147) and presence of LDH in mitochondria wasconfirmed by immunoprecipitation of mitochondrial frac-tions from L6 cells (35). Our findings obtained using bothCLSM and immunoprecipitation indicate that the terminalmitochondrial electron-transport chain constituent COX is
oriented to complex with MCT1, CD147, and LDH (35).However, we did not find an association between NADH-dh and MCT1 and LDH. The findings suggest the presenceof a previously unrecognized mitochondrial lactate oxida-tion complex associated with complex IV at the mitochon-drial inner membrane (Fig. 1). In this model, the chemicaland proton gradients across the inner membrane of respiringmitochondrial networks establish the conditions necessaryfor mitochondrial lactate and pyruvate uptake. Further, it issuggested that the oxidizing environment of COX permits
TABLE 2. Negative findings about a mitochondrial site of lactate oxidation.
1. Histochemical localization of MCT1 at cell surface but not in mitochondria.a. Immunofluorescence staining of MCT1 showed no labeling of intracellular compartments in cross-section of human (30,58) and rat (4,70) skeletal muscle. But, we (38) used
unfixed sections followed by ABC staining for histochemical protein detection, whereas others used formalin fixation before staining. This methodological distinction issignificant because it is known that antibody binding can be blocked in formalin-fixed sections. Additionally, it is likely that because of the presence of MCT1 in both thesarcolemma and subsarcolemmal mitochondria, investigators using less-than-ideal resolution and nonquantitative histochemical staining could not adequately distinguishbetween sarcolemmal, subsarcolemmal mitochondrial, and intermyofibrillar mitochondrial MCT1.
b. Immunohistochemistry showed that MCT1 and CD147 (basigin) were localized at cell surface of skeletal muscle or cardiac muscle of mice (52).c. Electron microscopy did not show MCT1 to be associated with mitochondria of rat heart (42). But, then again, the same investigators could not visualize sarcolemmal MCT1 in
sections of the same hearts that showed abundant MCT1 in Western blot analysis.2. Isolated mitochondria from acinar or any other type of cell did not show immunoreactivity to MCT1 (74). This result was subsequently shown to be incorrect by workers in a
collaborating group using the same antibodies (3).3. Physiological data of no or minimal mitochondrial lactate oxidation.a. Isolated mitochondria neither oxidized lactate nor contained LDH in human, mouse, or rat (60,64,73). But, the procedures used were those ideally suited to result in loss of
mLDH. And, the preparations were so poor, and the respiratory capacity so low, that one group reached the absurd conclusion that measurements of pulmonary oxygenconsumption during exercise were artifacts because muscle mitochondrial oxygen consumption could not possibly support the measured rates of pulmonary oxygenconsumption (60).
b. Lactate oxidation was low, approximately half that of pyruvate in saponin skinned fibers from rat skeletal and cardiac muscle (59). But, the detergent used is well known toresult in the loss of LDH. The loss of LDH cannot be assumed to come from cytosol exclusively, because the detergent would encounter subsarcolemmal mitochondria asreadily as the plasma membrane.
http://www.acsm-msse.org488 Official Journal of the American College of Sports Medicine
B A S I C S C I E N C E S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
4/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
oxidation of lactate to pyruvate for subsequent oxidativecatabolism of pyruvate in the TCA cycle.
By providing micrographs showing colocalization of MCT1, CD147, and COX in rat plantaris muscle (Fig. 2I),we extended the evidence for presence of a lactate oxidationcomplex to include mammalian skeletal muscle (16). Addi-
tionally, colocalization of COX and LDH was apparent inrat plantaris muscle (Fig. 2II). These micrographs support the existence of a lactate oxidation complex in adult mammalian skeletal muscle. Previously, Benton et al. (3)have reported that isolated rat muscle mitochondria con-tained MCT2; however, we (38) detected only faint signalsof MCT2 located at sarcolemma of rat plantaris muscles.Hence, we favor the interpretation that MCT1 is the primaryisoform for the lactate oxidation complex in muscle, but it may be that this is not universally the case for all tissues,such as in the brain (T. Hashimoto, H.-S. Cho, R. Hussien,and G.A. Brooks, unpublished data). So far, the ILS has been seen in liver, skeletal, and cardiac muscles (14,21),
and brain (66). Evidently, the ILS is most active whentissue energy demands increase, such as during exercise, bywhich glycolysis and glycogenolysis are accelerated andintracellular and blood lactate concentrations increase.Further, in terms of muscle physiology and metabolism,the differential cellular localizations and relative abundan-ces of the lactate oxidation complex (16,38) wouldcontribute to the CCLS (9). Lactate formed in some musclecells with high rates of glycolysis (e.g., type II fibers) could be readily released and transported into type I fibers, and
then imported lactate could be readily taken up andoxidized by subsarcolemmal mitochondria (14,38). Also,the CCLS between neighboring cells, even in the sametissue, would be relevant in the brain in terms of theastrocyteneuron lactate shuttle hypothesis (54). Hence, weare able to realize the centrality of the ILS by means of the
mitochondrial lactate oxidation complex in the regulation of energy substrate flux and its logical imperatives, such ashow to make a CCLS work in the absence of an ILS.Additionally, in combination with the well-known effect of endurance training on increasing the mass of the musclemitochondrial reticulum as well as muscle oxidativeenzymatic activities (24,25,40), the presence of the lactateoxidation complex helps explain why training increaseslactate clearance via oxidation in working muscle (5). In thelater part of this article, we provide the novel finding that mitochondrial constituents are physiologically coordinated by lactate.
Regulation of MCT1 and CD147 Expression
To understand lactate metabolism of muscle cells interms of the ILS, the precise mechanisms to upregulate theexpression of MCT (especially MCT1) and their chaperones(specifically CD147) need to be elucidated. Until recently,the focus has been on the ability of exercise training andmuscle contractions to increase the expression of the lactatetransporter MCT1 in mammalian skeletal and cardiacmuscles (2,7,17,22,27,37,57). In addition to exercise, cross
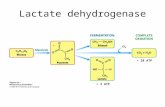
FIGURE 1Schematic showing the putative lactate oxidation complex: MCT1 is inserted into the mitochondrial inner membrane strongly
interacting with its chaperone protein CD147, and it is also associated with COX as well as mitochondrial LDH (mLDH), which could be located atthe outer side of the inner membrane. Lactate, which is always produced in cytosol of muscle and other tissues because of the abundance, activity,and characteristics of cytosolic LDH, is oxidized to pyruvate via the lactate oxidation complex in mitochondria of the same cell. This endergoniclactate oxidation reaction is coupled to the exergonic redox change in COX during mitochondrial electron transport. Transport of pyruvate acrossthe inner mitochondrial membrane is facilitated by MCT1. GP, glycerol phosphate; Mal-Asp, malateaspartate; ETC, electron-transport chain;TCA, tricarboxylic acid. Redrawn from Hashimoto et al. (35), with permission.
THE INTRACELLULAR LACTATE SHUTTLE Medicine & Science in Sports & Exercise d 489
BA SI C S CI EN CE S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
5/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
innervations (4), testosterone (28), and myocardial infarc-tion (42) have been observed to increase MCT1 expressionas well. In human colon cells, butyrate, which is another substrate for MCT1, increased MCT1 mRNA and proteinexpression (23), and, in thyroid cells, thyroid-stimulatinghormone (TSH) stimulated MCT1 expression at the tran-scription level (29). Among the various reports describedabove, it has been observed that MCT1 protein expressionwas rapidly upregulated in rat skeletal muscle by a single bout of exercise, but increases in MCT1 protein were not always accompanied by concomitant changes in transcript level (22). Similarly, it has been demonstrated in humancolonic epithelial cells that butyrate increased MCT1
mRNA expression by the dual control of MCT1 genetranscription and stability of the MCT1 transcript (23).Results of these investigations suggest that the MCT1 protein expression might be regulated by both transcrip-tional and posttranscriptional mechanisms.
The precise mechanisms regulating CD147 expression, aswell as its association with MCT1 expression in skeletal or cardiac muscles, have also not been fully elucidated.Jouaville et al. (43) have reported the effect of malnutritionon skeletal muscle MCT1 or CD147 mRNA expression in
rats showing muscle typespecific regulation. In extensor digitorum longus muscle, malnutrition decreased MCT1mRNA expression but increased CD147 mRNA expression.On the other hand, neither MCT1 nor CD147 mRNA waschanged by malnutrition in plantaris or soleus muscle.Initial findings with CD147-null mice were that MCT1 protein levels did not track levels of mRNA expressed (55).More recent reports on CD147 KO mice show large, tissue-specific alterations in MCT1 protein expression as deter-mined from Western blotting and immunohistochemistry(52). Also, with incubated thyroid cells, TSH increasedCD147 protein levels, but CD147 transcript levels did not respond to TSH (29). These findings suggest that MCT1
and CD147 genes and proteins are differentially regulated.Our most recent findings (36) of the regulation of MCT1and CD147 by lactate are described below.
Could It Be That Lactate Is Adaptive and RegulatesMCT1 Expression and Mitochondrial Biogenesis?
In addition to serving as an oxidizable substrate andgluconeogenic precursor, is it possible that lactate also has a signaling role as a pseudohormone lactormone (10) in the
FIGURE 2Cellular locations of MCT1, CD147, COX, and LDH were determined using confocal laser scanning microscopy (CLSM) andfluorescent probes for the respective proteins in rat plantaris muscle, as previously described (38). Panel I , MCT1 was detected throughout the cells,including subsarcolemmal and interfibrillar domains of oxidative fibers ( arrows: plates a and c ). CD147, chaperone protein of MCT1 ( plate b ) islocalized in association with MCT1 ( plate a ). When these fluorescences (MCT1 ( green ), CD147 ( red ), and COX ( blue )) were merged, superposition of the three probes was clear ( white , plate d ). Panel II , When the COX ( green ) and LDH ( red ) signals were merged, superposition of the two probes wasclear ( yellow, plate c ). These micrographs indicate the existence of a mitochondrial lactate oxidation complex in rat plantaris muscle. Scale bar = 20mm. From Brooks and Hashimoto (16), with permission.
http://www.acsm-msse.org490 Official Journal of the American College of Sports Medicine
B A S I C S C I E N C E S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
6/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
activation of genes known to respond to acute and chronic physical activity? In preliminary studies on rat muscle derived L6 cells to identify the physiological signalsaffecting lactate-transport protein expression, we noticedseveral things. First, the addition of Ca 2+ or a reactiveoxygen species (ROS) generator, such as H 2O2 , to theincubation medium caused rapid increases in MCT1 proteinexpression. Further, incubation of L6 cells in the high-
glucose-containing medium typically used in tissue cultureresulted in progressively rising lactate levels that wereaccompanied by increases in MCT1 and COX proteinlevels. Examination of the promoter areas for COX subunit IV and MCT1 revealed the presence of putative bindingsites for transcription factors that are known readouts of ROS- and Ca 2+ -signaling pathways. Therefore, to explorethe hypothesis that ROS generation, which may be induced by lactate, is involved in regulation of lactate oxidationcomplex proteins, we set about finding a means to controlthe culture lactate concentration levels and to determine theeffects of lactate anion on gene and protein expression. Wefound that elevated concentrations of lactate in culture asoccurs in contracting muscle in vivo is a key factor in thecoordination of lactate oxidation: lactate upregulated the
total mitochondrial mass and abundance of the lactateoxidation complex (MCT1, CD147, COX, and LDH) in L6cells through ROS-signaling mechanisms (Fig. 3) (36). Wealso found, using GeneChip analysis, that lactate incubationupregulated hundreds of ROS-sensitive genes, suggestingthe presence of a vast, lactate-activated transcription net-work, a lactate transcriptome. Specifically, these findingsindicate that lactate stimulates ROS generation, which
activates the transcription factors nuclear factor-kappaB(NF- J B), nuclear factor erythroid 2 (NF-E2, or Nrf2),nuclear respiratory factor (NRF)-2, and cAMP-responseelement-binding protein (CREB), leading, in turn, toincreases in MCT1 gene expression. With regard to NRF-2 and CREB, their involvement in mitochondrial biogenesis(19,50,53,67) is notable. MCT1 is predominant in slow-twitch oxidative fibers (38) and is a constituent of themitochondrial lactate oxidation complex (35). Coordinationof MCT1 and mitochondrial biogenesis by NRF-2 andCREB is likely physiologically relevant for increasing oxi-dative lactate-clearance capacity in skeletal muscle (5,27).
In the same study (36), increased CD147 and MCT1 protein contents were found in whole-muscle homogenatesof L6 cells after 1 h of incubation with lactate. Similarly,
FIGURE 3Schematic diagram summarizing the effects of lactate on intracellular signaling in muscle. Contractions stimulate glycolysis andsubsequent lactate production and accumulation. In combination, lactate accumulation and mitochondrial respiration induce ROS, which elicitsmany cell responses seen in the responses to exercise, including increased MCT1 expression, mitochondrial (mito) biogenesis, and the production of the antioxidant glutathione peroxidase (GPx). Also, lactate induces a large amount of gene expression, which is considered to be an adaptiveresponse. With regard to mitochondrial biogenesis, the lactate-signaling pathway merges with Ca
2+signaling as contractions increase cytosolic Ca
2+
flux. By itself, lactate increases expressions of slow-type troponin I (TnI) and myogenin, which are also know to be responsive to Ca2+
flux viacalcineurin (CaN). ROS can increase intracellular Ca
2+, which raises CaMK activity. Free Ca
2+can also activate CaMK. In conclusion, lactate elicits
a large number of adaptive responses and, thereby, would coordinate metabolism as a functional adaptation to exercise in skeletal muscle cells, suchas activation of the lactate oxidation complex. From Hashimoto et al. (36), with permission.
THE INTRACELLULAR LACTATE SHUTTLE Medicine & Science in Sports & Exercise d 491
BA SI C S CI EN CE S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
7/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
lactate incubation increased both MCT1 and CD147 tran-script levels within an hour. Therefore, at the tissue level,CD147 transcript and protein levels coincided.
Interactions among the insertions of scaffold (CD147)and transporter proteins (MCT1) into specific cell domains(mitochondria) are somewhat complex. In the mitochondrialfraction of L6 cells, increased MCT1 insertion was foundafter 1 h of incubation with lactate, but CD147 did not
increase, although it was abundant in mitochondria byWestern blotting. At 6 h of lactate incubation, mitochon-drial MCT1 and CD147 levels did not change; however,considering the increased mitochondrial mass represented by increased COX expression in the whole-muscle homo-genate of L6 cells, as well as increased gene expressions of mitochondrial import machinery TIM 13 and TIM 17 at 6 h(Fig. 3), lactate treatment upregulated the abundance of themitochondrial MCT1 and CD147, as indicated by Gene-Chip analysis. Mechanisms of the coordination of TOM/ TIM complexes with MCT1 and its accessory proteinCD147, as well as their targeting signals to mitochondria,remain to be elucidated.
Previously, Mootha et al. (51) and Taylor et al. (69) performed proteomic surveys of mitochondria from mouse brain, heart, kidney, liver, and human heart. These inves-tigators have provided evidence for more than 600 mito-chondrial or mitochondria-associated proteins, includingcomponents of the mitochondrial lactate oxidation complex(i.e., MCT1, CD147, COX, and LDH). We (36) expandedtheir demonstration, and, by explaining how these mito-chondrial constituents are assembled to form a lactateoxidation complex, we provided an insight on how thecomplex functions to facilitate lactate disposal in the L6 cellsand adult mammalian skeletal muscle (38). Again, exercise
training results in large increases in lactate clearance via oxidation (5) as well as in lactate oxidation complexconstituents, with increases in mitochondrial biogenesis(36). Such an adaptation would facilitate lactate oxidation inskeletal muscle cells, permitting high-power outputs andglycolytic fluxes to occur while minimizing acidosis, whichreveals the physiological significance of the ILS.
SUMMARY AND CONCLUSION
In summary, by using confocal laser scanning micro-scopy, Western blotting of cell fractions, and immunoblot-ting after immunoprecipitation from cell lysates, evidencehas been provided (35) of strong associations amongmitochondrial MCT1, CD147, LDH, and COX, but not NADH-dh. These findings advance our understanding of the mitochondrial role in cellular lactate oxidation, and theymay be interpreted to indicate the presence of a terminalmitochondrial electron-transport chain component, thelactate oxidation complex, which establishes the protonand lactate concentration gradients necessary for thefunctioning of the CCLS and ILS. The physiologicalsignificance of the ILS is that it provides the necessary link between glycolysis and oxidative metabolism, with the product of the former being a chief substrate for the latter.Further significance of the most recent work is that in itssignaling role (i.e., in its function as a lactormone), lactate production may be adaptive. During exercise, the transient elevations in tissue lactate concentration and mitochon-
drial O 2 consumption induce ROS generation, whichactivates a transcriptional network signaling adaptive cellresponses. Among these are increases in the lactateoxidation complex, mitochondrial biogenesis, and upregu-lation of antioxidant enzymes and moieties of Ca 2+ signal-ing (Fig. 3). The results we provide here can be interpretedto mean that elevated lactate flux and concentration signalslead to adaptation of pathways of lactate removal (lactateoxidation complex) and also signal many of the adaptationsin muscle found in response to endurance training.Although still controversial, the evidence provided in thisand other recent publications is supportive of the ILShypothesis, as well as for an adaptive role of muscle lactate production during exercise.
This work was supported by NIH R01 AR050459 to G.A.B. T.H.was supported by a grant from the Japan Society for the Promotionof Science. Thanks are due to Tamara Mau for reading andcommenting on the manuscript.
REFERENCES1. Baba N, Sharma HM. Histochemistry of lactic dehydrogenase in
heart and pectoralis muscles of rat. J Cell Biol . 1971;51:62165.2. Baker SK, McCullagh KJ, Bonen A. Training intensity-dependent
and tissue-specific increases in lactate uptake and MCT-1 in heart
and muscle. J Appl Physiol . 1998;84:98794.3. Benton CR, Campbell SE, Tonouchi M, Hatta H, Bonen A.Monocarboxylate transporters in subsarcolemmal and intermyofi- brillar mitochondria. Biochem Biophys Res Commun . 2004;323:24953.
4. Bergersen LH, Thomas M, Johannsson E, et al. Cross-reinnerva-tion changes the expression patterns of the monocarboxylatetransporters 1 and 4: an experimental study in slow and fast rat skeletal muscle. Neuroscience. 2006;138:110513.
5. Bergman BC, Wolfel EE, Butterfield GE, et al. Active muscle andwhole body lactate kinetics after endurance training in men. J Appl Physiol . 1999;87:168496.
6. Bertocci LA, Lujan BF. Incorporation and utilization of [3-13C]lactate and [1,2-13C]acetate by rat skeletal muscle. J Appl Physiol . 1999;86:207789.
7. Bonen A, Tonouchi M, Miskovic D, Heddle C, Heikkila JJ,
Halestrap AP. Isoform-specific regulation of the lactate trans- porters MCT1 and MCT4 by contractile activity. Am J Physiol Endocrinol Metab . 2000;279:E11318.
8. Brandt RB, Laux JE, Spainhour SE, Kline ES. Lactate dehydro-genase in rat mitochondria. Arch Biochem Biophys . 1987;259:41222.
9. Brooks GA. Glycolytic end product and oxidative substrate duringsustained exercise in mammalsthe lactate shuttle. In: GillesR, editor. Current Topics and Trends, Volume A, Respiration MetabolismCirculation . Berlin (Germany); 1985. p. 20218.
10. Brooks GA. Lactate shuttles in nature. Biochem Soc Trans .2002;30:25864.
http://www.acsm-msse.org492 Official Journal of the American College of Sports Medicine
B A S I C S C I E N C E S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
8/9Copyright @ 200 by the American College of Sports Medicine. Unauthorized reproduction of this article is prohibited.8
11. Brooks GA. Mammalian fuel utilization during sustained exercise.Comp Biochem Physiol B Biochem Mol Biol . 1998;120:89107.
12. Brooks GA, Brown MA, Butz CE, Sicurello JP, Dubouchaud H.Cardiac and skeletal muscle mitochondria have a monocarbox-ylate transporter MCT1. J Appl Physiol . 1999;87:17138.
13. Brooks GA, Butterfield GE, Wolfe RR, et al. Decreased relianceon lactate during exercise after acclimatization to 4,300 m. J Appl Physiol . 1991;71:33341.
14. Brooks GA, Dubouchaud H, Brown M, Sicurello JP, Butz CE.Role of mitochondrial lactate dehydrogenase and lactate oxidation
in the intracellular lactate shuttle. Proc Natl Acad Sci U. S. A .1999;96:112934.
15. Brooks GA, Hashimoto T. Investigation of the lactate shuttle inskeletal muscle mitochondria. J Physiol. 2007;584:7056.
16. Brooks GA, Hashimoto T. Organization of intermediary metab-olismlactate and its transporters. Physiol News 2006;65:1920.
17. Burgomaster KA, Cermak NM, Phillips SM, Benton CR, BonenA, Gibala MJ. Divergent response of metabolite transport proteinsin human skeletal muscle after sprint interval training anddetraining. Am J Physiol Regul Integr Comp Physiol. 2007;292(5):R19706.
18. Butz CE, McClelland GB, Brooks GA. MCT1 confirmed in rat striated muscle mitochondria. J Appl Physiol . 2004;97:105966.
19. Chabi B, Adhihetty PJ, Ljubicic V, Hood DA. How ismitochondrial biogenesis affected in mitochondrial disease? Med
Sci Sports Exerc . 2005;37(12):210210.20. Chandra NC, Spiro MJ, Spiro RG. Identification of a glycoprotein
from rat liver mitochondrial inner membrane and demonstration of its origin in the endoplasmic reticulum. J Biol Chem . 1998;273:1971521.
21. Chatham JC, Des Rosiers C, Forder JR. Evidence of separate pathways for lactate uptake and release by the perfused rat heart. Am J Physiol Endocrinol Metab . 2001;281:E794802.
22. Coles L, Litt J, Hatta H, Bonen A. Exercise rapidly increasesexpression of the monocarboxylate transporters MCT1 and MCT4in rat muscle. J Physiol . 2004;561:25361.
23. Cuff MA, Lambert DW, Shirazi-Beechey SP. Substrate-inducedregulation of the human colonic monocarboxylate transporter,MCT1. J Physiol . 2002;539:36171.
24. Davies KJ, Packer L, Brooks GA. Biochemical adaptation of
mitochondria, muscle, and whole-animal respiration to endurancetraining. Arch Biochem Biophys . 1981;209:53954.25. Davies KJ, Packer L, Brooks GA. Exercise bioenergetics follow-
ing sprint training. Arch Biochem Biophys . 1982;215:2605.26. De Bari L, Atlante A, Valenti D, Passarella S. Partial reconstruc-
tion of in vitro gluconeogenesis arising from mitochondrialL-lactate uptake/metabolism and oxaloacetate export via novel L-lactate translocators. Biochem J . 2004;380:23142.
27. Dubouchaud H, Butterfield GE, Wolfel EE, Bergman BC, BrooksGA. Endurance training, expression, and physiology of LDH,MCT1, and MCT4 in human skeletal muscle. Am J Physiol Endocrinol Metab . 2000;278:E5719.
28. Enoki T, Yoshida Y, Lally J, Hatta H, Bonen A. Testosteroneincreases lactate transport, monocarboxylate transporter (MCT) 1and MCT4 in rat skeletal muscle. J Physiol . 2006;577:43343.
29. Fanelli A, Grollman EF, Wang D, Philp NJ. MCT1 and itsaccessory protein CD147 are differentially regulated by TSH inrat thyroid cells. Am J Physiol Endocrinol Metab . 2003;285:E12239.
30. Fishbein WN, Merezhinskaya N, Foellmer JW. Relative distribu-tion of three major lactate transporters in frozen human tissues andtheir localization in unfixed skeletal muscle. Muscle Nerve.2002;26:10112.
31. Garcia CK, Goldstein JL, Pathak RK, Anderson RG, Brown MS.Molecular characterization of a membrane transporter for lactate, pyruvate, and other monocarboxylates: implications for the Coricycle. Cell. 1994;76:86573.
32. Gertz EW, Wisneski JA, Stanley WC, Neese RA. Myocardialsubstrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J Clin Invest . 1988;82:201725.
33. Gladden LB. Lactate metabolism: a new paradigm for the thirdmillennium. J Physiol . 2004;558:530.
34. Halestrap AP, Price NT. The proton-linked monocarboxylatetransporter (MCT) family: structure, function and regulation. Biochem J . 1999;343:28199.
35. Hashimoto T, Hussien R, Brooks GA. Colocalization of MCT1,
CD147, and LDH in mitochondrial inner membrane of L6 musclecells: evidence of a mitochondrial lactate oxidation complex. Am J Physiol Endocrinol Metab . 2006;290:E123744.
36. Hashimoto T, Hussien R, Oommen S, Gohil K, Brooks GA.Lactate sensitive transcription factor network in L6 cells:activation of MCT1 and mitochondrial biogenesis. FASEB J.2007;21(10):260212.
37. Hashimoto T, Kambara N, Nohara R, Yazawa M, Taguchi S.Expression of MHC-beta and MCT1 in cardiac muscle after exercise training in myocardial-infarcted rats. J Appl Physiol .2004;97:84351.
38. Hashimoto T, Masuda S, Taguchi S, Brooks GA. Immunohisto-chemical analysis of MCT1, MCT2 and MCT4 expression in rat plantaris muscle. J Physiol . 2005;567:1219.
39. Henderson GC, Horning MA, Lehman SL, Wolfel EE, Bergman
BC, Brooks GA. Pyruvate shuttling during rest and exercise before and after endurance training in men. J Appl Physiol .2004;97:31725.
40. Holloszy JO, Booth FW. Biochemical adaptations to enduranceexercise in muscle. Annu Rev Physiol . 1976;38:27391.
41. Jobsis FF, Stainsby WN. Oxidation of NADH during contractionsof circulated mammalian skeletal muscle. Respir Physiol . 1968;4:292300.
42. Johannsson E, Lunde PK, Heddle C, et al. Upregulation of thecardiac monocarboxylate transporter MCT1 in a rat model of congestive heart failure. Circulation. 2001;104:72934.
43. Jouaville LF, Fellmann N, Coudert J, Clottes E. Skeletal muscleexpression of LDH and monocarboxylate transporters ingrowing rats submitted to protein malnutrition. Eur J Nutr . 2006;45:35562.
44. Kirk P, Wilson MC, Heddle C, Brown MH, Barclay AN,Halestrap AP. CD147 is tightly associated with lactate trans- porters MCT1 and MCT4 and facilitates their cell surfaceexpression. EMBO J . 2000;19:3896904.
45. Kline ES, Brandt RB, Laux JE, et al. Localization of L-lactatedehydrogenase in mitochondria. Arch Biochem Biophys . 1986;246:67380.
46. Laughlin MR, Taylor J, Chesnick AS, DeGroot M, Balaban RS.Pyruvate and lactate metabolism in the in vivo dog heart. Am J Physiol . 1993;264:H206879.
47. Manoharan C, Wilson MC, Sessions RB, Halestrap AP. The roleof charged residues in the transmembrane helices of monocarbox-ylate transporter 1 and its ancillary protein basigin in determining plasma membrane expression and catalytic activity. Mol Membr Biol . 2006;23:48698.
48. McClelland GB, Brooks GA. Changes in MCT 1, MCT 4, andLDH expression are tissue specific in rats after long-termhypobaric hypoxia. J Appl Physiol . 2002;92:157384.
49. McClelland GB, Khanna S, Gonzalez GF, Butz CE, Brooks GA.Peroxisomal membrane monocarboxylate transporters: evidencefor a redox shuttle system? Biochem Biophys Res Commun . 2003;304:1305.
50. McGill JK, Beal MF. PGC-1alpha, a new therapeutic target inHuntington ` s disease? Cell. 2006;127:4658.
51. Mootha VK, Bunkenborg J, Olsen JV, et al. Integrated analysis of protein composition, tissue diversity, and gene regulation inmouse mitochondria. Cell. 2003;115:62940.
THE INTRACELLULAR LACTATE SHUTTLE Medicine & Science in Sports & Exercise d 493
BA SI C S CI EN CE S
-
8/14/2019 2008 Rev Mitoc Lactate ( Cajigal)
9/9
52. Nakai M, Chen L, Nowak RA. Tissue distribution of basigin andmonocarboxylate transporter 1 in the adult male mouse: a studyusing the wild-type and basigin gene knockout mice. Anat Rec A Discov Mol Cell Evol Biol . 2006;288:52735.
53. Ongwijitwat S, Wong-Riley MT. Is nuclear respiratory factor 2 a master transcriptional coordinator for all ten nuclear-encoded cyto-chrome c oxidase subunits in neurons? Gene. 2005;360:6577.
54. Pellerin L, Magistretti PJ. Glutamate uptake into astrocytesstimulates aerobic glycolysis: a mechanism coupling neuronalactivity to glucose utilization. Proc Natl Acad Sci U. S. A .
1994;91:106259.55. Philp NJ, Ochrietor JD, Rudoy C, Muramatsu T, Linser PJ. Loss
of MCT1, MCT3, and MCT4 expression in the retinal pigment epithelium and neural retina of the 5A11/basigin-null mouse. Invest Ophthalmol Vis Sci . 2003;44:130511.
56. Pierre K, Pellerin L, Debernardi R, Riederer BM, Magistretti PJ.Cell-specific localization of monocarboxylate transporters, MCT1and MCT2, in the adult mouse brain revealed by doubleimmunohistochemical labeling and confocal microscopy. Neuro- science. 2000;100:61727.
57. Pilegaard H, Domino K, Noland T, et al. Effect of high-intensityexercise training on lactate/H+ transport capacity in humanskeletal muscle. Am J Physiol . 1999;276:E25561.
58. Pilegaard H, Terzis G, Halestrap A, Juel C. Distribution of thelactate/H+ transporter isoforms MCT1 and MCT4 in human
skeletal muscle. Am J Physiol . 1999;276:E8438.59. Ponsot E, Zoll J, N ` Guessan B, et al. Mitochondrial tissue
specificity of substrates utilization in rat cardiac and skeletalmuscles. J Cell Physiol . 2005;203:47986.
60. Rasmussen UF, Rasmussen HN, Krustrup P, Quistorff B, Saltin B,Bangsbo J. Aerobic metabolism of human quadriceps muscle: invivo data parallel measurements on isolated mitochondria. Am J Physiol Endocrinol Metab . 2001;280:E3017.
61. Rasmussen HN, van Hall G, Rasmussen UF. Lactate dehydroge-nase is not a mitochondrial enzyme in human and mouse vastuslateralis muscle. J Physiol . 2002;541:57580.
62. Richardson RS, Noyszewski EA, Leigh JS, Wagner PD. Lactateefflux from exercising human skeletal muscle: role of intracellular PO2. J Appl Physiol . 1998;85:62734.
63. Roth DA, Brooks GA. Lactate and pyruvate transport is
dominated by a pH gradient-sensitive carrier in rat skeletal musclesarcolemmal vesicles. Arch Biochem Biophys . 1990;279:38694.
64. Roth DA, Brooks GA. Lactate transport is mediated by a membrane-bound carrier in rat skeletal muscle sarcolemmalvesicles. Arch Biochem Biophys . 1990;279:37785.
65. Sahlin K, Fernstrom M, Svensson M, Tonkonogi M. No evidenceof an intracellular lactate shuttle in rat skeletal muscle. J Physiol .2002;541:56974.
66. Schurr A. Lactate: the ultimate cerebral oxidative energysubstrate? J Cereb Blood Flow Metab . 2006;26:14252.
67. St-Pierre J, Drori S, Uldry M, et al. Suppression of reactiveoxygen species and neurodegeneration by the PGC-1 transcrip-
tional coactivators. Cell. 2006;127:397408.68. Stellingwerff T, Leblanc PJ, Hollidge MG, Heigenhauser GJ,
Spriet LL. Hyperoxia decreases muscle glycogenolysis, lactate production, and lactate efflux during steady-state exercise. Am J Physiol Endocrinol Metab . 2006;290:E118090.
69. Taylor SW, Fahy E, Zhang B, et al. Characterization of the humanheart mitochondrial proteome. Nat Biotechnol . 2003;21:2816.
70. Watt PW, MacLennan PA, Hundal HS, Kuret CM, Rennie MJ.L(+)-lactate transport in perfused rat skeletal muscle: kineticcharacteristics and sensitivity to pH and transport inhibitors. Biochim Biophys Acta. 1988;944:21322.
71. Wilson MC, Jackson VN, Heddle C, et al. Lactic acid efflux fromwhite skeletal muscle is catalyzed by the monocarboxylatetransporter isoform MCT3. J Biol Chem . 1998;273:159206.
72. Wilson MC, Meredith D, Fox JE, Manoharan C, Davies AJ,
Halestrap AP. Basigin (CD147) is the target for organomercurialinhibition of monocarboxylate transporter isoforms 1 and 4: theancillary protein for the insensitive MCT2 is EMBIGIN (gp70). J Biol Chem . 2005;280:2721321.
73. Wilson MC, Meredith D, Halestrap AP. Fluorescence resonanceenergy transfer studies on the interaction between the lactatetransporter MCT1 and CD147 provide information on thetopology and stoichiometry of the complex in situ. J Biol Chem .2002;277:366672.
74. Yoshida Y, Holloway GP, Ljubicic V, et al. Negligible direct lactate oxidation in subsarcolemmal and intermyofibrillar mito-chondria obtained from red and white rat skeletal muscle. J Physiol. 2007;582:131735.
75. Zhao C, Wilson MC, Schuit F, Halestrap AP, Rutter GA.Expression and distribution of lactate/monocarboxylate trans-
porter isoforms in pancreatic islets and the exocrine pancreas. Diabetes. 2001;50:3616.
http://www.acsm-msse.org494 Official Journal of the American College of Sports Medicine
B A S I C S C I E N C E S