
© 2009 Melissa Dawn Lingis -...
134
1 REGULATION OF THE MATERNAL HYPOTHALAMIC-PITUITARY-ADRENAL AXIS DURING PREGNANCY IN THE EWE: RELATIVE ROLES OF THE MINERALOCORTICOID RECEPTOR AND THE SEROTONERGIC SYSTEM By MELISSA DAWN LINGIS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2009
Transcript of © 2009 Melissa Dawn Lingis -...

1
REGULATION OF THE MATERNAL HYPOTHALAMIC-PITUITARY-ADRENAL AXIS DURING PREGNANCY IN THE EWE: RELATIVE ROLES OF THE
MINERALOCORTICOID RECEPTOR AND THE SEROTONERGIC SYSTEM
By
MELISSA DAWN LINGIS
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2009

2
© 2009 Melissa Dawn Lingis

3
To my husband, Rob, for his constant support and encouragement To my parents, Dave and Pam Landen, for their unconditional love
And to EACH of them for always believing in me For my son, Matthew, may even your dreams exceed your expectations

4
ACKNOWLEDGMENTS
I would first like to thank all of the members of my committee: Dr. Charles Wood, Dr.
Joanna Peris, Dr. Dorette Ellis, and Dr. Colin Sumners for their suggestions and guidance both in
my pursuit of this degree and on aspects of this research. A special thank you goes to Dr.
Maureen Keller-Wood, my committee chairperson, for her mentorship and unwavering
confidence in my abilities. For me, Dr. Keller-Wood has served as a wonderful role model as a
woman in the field of scientific research who still maintains the ability to be an involved mother
in the lives of her three children. As a future mentor myself, I hope to have inherited her energy,
enthusiasm, and encouraging attitude.
Within the College of Pharmacy, there are several people I would like to acknowledge.
Special thanks and congratulations go to my classmate and colleague, Dr. Chinki Bhatia. I
always enjoyed our chats over lunch and I am truly grateful for her support and friendship,
especially as we struggled through our qualifying exams together. I would also like to thank Dr.
Elaine Sumners for her mentorship, contributions to this research, and friendship. Through her
impeccable research ethic and technical abilities, she has served as an invaluable resource. Many
thanks also go to Dr. Yun-Ju He for his skillful work in various plasma hormone assays which
contributed to the data within this manuscript. Additionally, I would like to thank past members
of the department who played key roles in shaping me as a scientist. Thanks go to Dr. Marcela
von Reitzenstein, a former post-doctoral associate in our lab and Mrs. Krista Koehler, a former
technician, for encouraging me to pursue this degree. Special thanks go to Ms. Monique
Sutherland, who volunteered in the lab as an undergraduate student, for her friendship and
contributions to this research. I would also like to thank all of the undergraduate and pharmacy
students who have volunteered and contributed in any way to the data presented here.

5
Most importantly, I would like to express my appreciation for my wonderfully supportive
family. My husband, Rob, always supported me even through the most difficult times and
provided gentle, loving nudges to keep me from giving up. His outstanding academic and career
accomplishments continue to encourage me every day. I cannot thank him enough for the
sacrifices he has made in order to provide for our family and to enable me to finish this
dissertation in a timely manner. Finally, I would like to thank my parents for their never-ending
encouragement and confidence in me. It is impossible to put into words exactly how
appreciative I am for all they have given me. I know that I never would have accomplished this
much without their guidance, love, and understanding throughout my entire career at the
University of Florida.

6
TABLE OF CONTENTS page
ACKNOWLEDGMENTS ...............................................................................................................4
LIST OF TABLES ...........................................................................................................................9
LIST OF FIGURES .......................................................................................................................10
ABSTRACT ...................................................................................................................................12
CHAPTER
1 INTRODUCTION ..................................................................................................................14
Specific Aim 1: Relative Role of the Mineralocorticoid Receptor in Regulation of HPA Axis Activation During Pregnancy in the Ewe ...................................................................15
Specific Aim 2: Relative Role of Serotonergic System Responsivity in Regulation of HPA Axis Activation During Pregnancy in the Ewe ..........................................................16
Specific Aim 3: Relative Hypothalamic Expression of Genes Related to HPA Axis Regulation and the Serotonergic System ............................................................................17
2 REVIEW OF LITERATURE .................................................................................................18
Hypothalamic-Pituitary-Adrenal (HPA) Axis ........................................................................18 Role of Corticosteroids in Normal Physiology ...............................................................19 Negative Feedback Regulation ........................................................................................23 Role of Corticosteroids in Pregnancy and Parturition .....................................................25
Serotonergic System ...............................................................................................................28 Impact of the Serotonin System on HPA Axis Activity ..................................................31 Serotonergic System on Regulation of Food Intake ........................................................33
Evidence for Influence of Ovarian Hormones ........................................................................35
3 ROLE OF MINERALOCORTICOID RECEPTORS IN REGULATION OF CORTISOL, ALDOSTERONE, ELECTROLYTES, AND BLOOD PRESSURE IN PREGNANCY ........................................................................................................................40
Introduction .............................................................................................................................40 Materials and Methods ...........................................................................................................43
Animals ............................................................................................................................43 Surgical Protocol .............................................................................................................43 Experimental Protocol .....................................................................................................44 Plasma Hormone Determination .....................................................................................45 Plasma Volume Determination ........................................................................................45 Data Analysis ...................................................................................................................46
Results .....................................................................................................................................46 Plasma Hormone Levels ..................................................................................................46

7
Adrenocorticotropic hormone (ACTH) ....................................................................46 Cortisol .....................................................................................................................47 Aldosterone ..............................................................................................................48 Angiotensin II ...........................................................................................................48
Mean Arterial Blood Pressure .........................................................................................48 Hematocrit (%) and Plasma Solute (Total Protein, Potassium, and Sodium)
Concentrations .............................................................................................................49 Plasma Volume ................................................................................................................50
Discussion ...............................................................................................................................50
4 RELATIVE SEROTONERGIC ACTIVITY/RESPONSIVITY DURING PREGNANCY ........................................................................................................................64
Introduction .............................................................................................................................64 Materials and Methods ...........................................................................................................66
Animals ............................................................................................................................66 Surgical Protocol .............................................................................................................67 Experimental Protocol .....................................................................................................68 Daily Food Intake ............................................................................................................70 Plasma Hormone Determination .....................................................................................70 Euthanasia and Tissue Recovery .....................................................................................70 Data Analysis ...................................................................................................................71
Results .....................................................................................................................................71 Study I: HPA Axis Responses to Acute, Icv Fluoxetine .................................................71
Plasma ACTH ..........................................................................................................71 Plasma cortisol .........................................................................................................72
Study II: HPA Axis Responses to Subchronic, Icv Fluoxetine .......................................73 Plasma ACTH ..........................................................................................................73 Plasma cortisol .........................................................................................................73 Mean arterial pressure, hematocrit (%) and plasma solute (total protein,
potassium, and sodium) concentrations ................................................................74 Daily food intake ......................................................................................................74
Discussion ...............................................................................................................................75 Study I: HPA Axis Responses to Acute, Icv Fluoxetine .................................................75 Study II: HPA Axis Responses to Subchronic, Icv Fluoxetine .......................................77 Summary ..........................................................................................................................80
5 HYPOTHALAMIC EXPRESSION OF GENES RELATED TO HPA AXIS REGULATION AND THE SEROTONERGIC SYSTEM IN EWES ...................................90
Introduction .............................................................................................................................90 Materials and Methods ...........................................................................................................92
Euthanasia and Tissue Recovery .....................................................................................92 RNA Extraction and Quantification ................................................................................93 Reverse Transcription and Real-Time Quantitative PCR ...............................................93 Data Analysis ...................................................................................................................94
Results .....................................................................................................................................95

8
Glucocorticoid Receptor (GR) ........................................................................................95 Mineralocorticoid Receptor (MR) ...................................................................................95 Corticotropin-Releasing Hormone (CRH) ......................................................................95 Arginine Vasopressin (AVP) ...........................................................................................96 5-HT1A Receptor ..............................................................................................................96 5-HT2A Receptor ..............................................................................................................96 Serotonin Reuptake Transporter (SERT) ........................................................................96 Proopiomelanocortin (POMC) ........................................................................................97
Discussion ...............................................................................................................................97
6 SUMMARY ..........................................................................................................................110
LIST OF REFERENCES .............................................................................................................116
BIOGRAPHICAL SKETCH .......................................................................................................133

9
LIST OF TABLES
Table page 5-1 Probe and primer sequences for qPCR. ...........................................................................101

10
LIST OF FIGURES
Figure page 2-1 A general model of HPA axis regulation. ..........................................................................38
2-2 Basic regulatory circuitry for food intake ..........................................................................39
3-1 Plasma ACTH concentrations during canrenoate or vehicle infusion ...............................56
3-2 Plasma cortisol concentrations during canrenoate or vehicle infusion ..............................57
3-3 Plasma aldosterone concentrations during canrenoate or vehicle infusion .......................58
3-4 Plasma angiotensin II concentrations during canrenoate or vehicle infusion ....................59
3-5 Mean arterial pressure during canrenoate or vehicle infusion. ..........................................60
3-6 Hematocrit and plasma solute concentrations ....................................................................61
3-7 Linear regression of plasma potassium and plasma aldosterone during canrenoate infusion ..............................................................................................................................62
3-8 Plasma volume after 4 hours of vehicle or canrenoate infusion ........................................63
4-1 Study I: Plasma ACTH following acute icv fluoxetine or vehicle ....................................83
4-2 Study I: Plasma cortisol following acute icv fluoxetine or vehicle ...................................84
4-3 Study II: Plasma ACTH response to subchronic icv fluoxetine or vehicle .......................85
4-4 Study II: Plasma cortisol response to subchronic icv fluoxetine or vehicle ......................86
4-5 Study II: Mean arterial pressure during subchronic icv fluoxetine or vehicle. ..................87
4-6 Study II: Hematocrit and plasma solute concentrations during subchronic icv fluoxetine or vehicle. .........................................................................................................88
4-7 Study II: Daily food intake during subchronic icv fluoxetine or vehicle ..........................89
5-1 Relative hypothalamic glucocorticoid receptor (GR) mRNA expression ........................102
5-2 Relative hypothalamic mineralocorticoid receptor (MR) mRNA expression .................103
5-3 Relative hypothalamic corticotropin-releasing hormone (CRH) mRNA expression .......104
5-4 Relative hypothalamic arginine vasopressin (AVP) mRNA expression ..........................105
5-5 Relative hypothalamic 5-HT1A receptor mRNA expression ............................................106

11
5-6 Relative hypothalamic 5-HT2A receptor mRNA expression ............................................107
5-7 Relative hypothalamic serotonin reuptake transporter (SERT) mRNA expression .........108
5-8 Relative hypothalamic proopiomelanocortin (POMC) mRNA expression ......................109
6-1 Proposed circuitry for regulation of HPA axis ................................................................115

12
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
REGULATION OF THE MATERNAL HYPOTHALAMIC-PITUITARY-ADRENAL AXIS
DURING PREGNANCY IN THE EWE: RELATIVE ROLES OF THE MINERALOCORTICOID RECEPTOR AND THE SEROTONERGIC SYSTEM
By
Melissa Dawn Lingis
December 2009 Chair: Maureen Keller-Wood Major: Pharmaceutical Sciences
Studies in our laboratory and others have shown that cortisol, a corticosteroid important for
maternal hemodynamic changes and fetal homeostasis is basally regulated at higher levels in the
pregnant state than in the non-pregnant state. Our lab is testing two broad hypotheses to, at least
in part, explain this increase in activity during pregnancy: (1) alterations to the axis’ basal
negative feedback via mineralocorticoid receptors (MR) or (2) alterations in the serotonergic
system, a stimulatory central input to the axis.
To study the relative role of MR during pregnancy, I studied the effect of an acute
intravenous (iv) infusion of the MR antagonist, canrenoate on plasma adrenocorticotropic
hormone (ACTH), cortisol, aldosterone and angiotensin II as well as electrolyte balance and
blood pressure in pregnant and non-pregnant ewes. These two groups demonstrated a
differential time course for the stimulation of ACTH and cortisol in response to intravenous
mineralocorticoid blockade, suggestive of alterations in central MR-mediated regulation of the
axis while also supporting a role for MR in electrolyte balance and volume expansion in
pregnancy. To study possible alterations in the serotonergic system, I compared ACTH and
cortisol responses to a high-dose, acute intracerebroventricular (icv) injection of the selective

13
serotonin reuptake inhibitor (SSRI), fluoxetine (FLX) and the responses to low-dose, subchronic
icv infusion of fluoxetine between nonpregnant and pregnant ewes. Opposite to our hypothesis,
serotonergic responsivity may be blunted in pregnancy providing evidence that an upregulation
of this system may not be responsible for the increases in basal plasma ACTH and cortisol that
occur during pregnancy. The differential HPA axis responses to both mineralocorticoid receptor
blockade and serotonergic system stimulation were however suggestive of alterations in the roles
of these two systems in basal HPA axis activity regulation during pregnancy. Analyses of
relative hypothalamic mRNA expression of pertinent HPA axis and serotonergic system genes
did not demonstrate any differences at the gene level. Continued research to completely
characterize regulation of the HPA axis will prove beneficial to the clinical sector, both in its
importance to pregnancy and to nearly all aspects of human physiology and homeostasis.

14
CHAPTER 1 INTRODUCTION
The primary role of the hypothalamic-pituitary-adrenal (HPA) axis is to aid in the
adaptation to a variety of stressors through the production of the adrenal corticosteroid
hormones. One of the main focuses of our lab is to investigate alterations in the regulation of
basal maternal HPA axis activity that arise during pregnancy. Studies in humans and other
animal models such as the sheep have demonstrated upregulation of the maternal HPA axis
activity and therefore cortisol, a key corticosteroid, in pregnancy (Carr et al. 1981, Bell et al.
1991, Keller-Wood 1998, Erickson et al. 2001, Sandman et al. 2006, Kirschbaum et al. 2009).
Our lab has shown that corticosteroids are important for maternal volume expansion, uterine
blood flow, and fetal homeostasis and that clamping cortisol to levels observed in the
nonpregnant ewe has dramatic impact on these factors and the growth rate of the fetus (Jensen et
al. 2002, Jensen et al. 2003, Jensen et al. 2005). Our lab is interested in understanding the
mechanisms controlling basal HPA axis activity during pregnancy using sheep as an alternative
model to human pregnancy (Keller-Wood et al. 1998). This research focuses on an
understudied, basic adaptive process of successful pregnancy. It is important that we fully
characterize the regulation of the HPA axis to ultimately provide beneficial information for
future investigation of innovative approaches to treat dysregulation of such processes and
therefore prevent the development of an adverse environment for the fetus and to protect the
health of the mother.
The following studies are aimed at comparing: (1) the relative role of the mineralocorticoid
receptor and (2) relative serotonergic responsivity between pregnant and non-pregnant sheep as
they relate to negative feedback maintenance of basal HPA axis activity. The end-points of the
following studies are plasma levels of relevant hormones, such as adrenocorticotropic hormone

15
(ACTH), cortisol, aldosterone, angiotensin II and progesterone as well as hemodynamic
parameters including blood pressure, electrolyte concentration, plasma protein, and hematocrit.
Additionally, I have compared relative hypothalamic mRNA expression of relevant HPA axis
related and serotonergic system-associated genes between pregnant, nonpregnant, and
postpartum ewes, including corticotropin-releasing hormone (CRH), arginine vasopressin
(AVP), mineralocorticoid receptor (MR), glucocorticoid receptor (GR), serotonin receptors (5-
HT1A and 5-HT2A), and the serotonin reuptake transporter (SERT). An additional investigation
of relative hypothalamic expression of proopiomelanocortin (POMC) mRNA was also
performed.
Specific Aim 1: Relative Role of the Mineralocorticoid Receptor in Regulation of HPA Axis Activation During Pregnancy in the Ewe
Based on a number of studies in both sheep and humans, our lab and others have theorized
that there is an alteration in ‘set-point’ of the negative feedback regulation of specifically basal
HPA axis activity that may at least in part explain the elevations of maternal plasma ACTH and
cortisol seen in pregnancy (Nolten & Rueckert 1981, Charnvises et al. 1985, Tropper et al. 1987,
Odagiri et al. 1988, Keller-Wood 1996, Keller-Wood 1998). Cortisol acts at two corticosteroid
receptors located at sites upstream and along the axis in order to inhibit its activity. Of these two
receptors, the higher affinity mineralocorticoid receptor (MR) is thought to be important for
regulating the axis when cortisol levels are low, at basal, non-stressed levels.
In order to test our lab’s overall hypothesis that progesterone, which is elevated during
pregnancy, may be influencing cortisol’s ability to act at MR along negative feedback sites of the
axis, I have administered the MR antagonist, canrenoate to both pregnant and nonpregnant ewes
in order to examine the relative role of MR during pregnancy on HPA axis activation, blood
pressure, and plasma solute regulation.

16
Specific Aim 2: Relative Role of Serotonergic System Responsivity in Regulation of HPA Axis Activation During Pregnancy in the Ewe
The serotonergic system is thought to be one of the stimulatory afferent systems to the
HPA axis (Calogero et al. 1990, Fuller 1996). Upon intravenous or oral administration of
serotonin, its precursors, or 5-HT receptor agonists, significant increases in plasma ACTH and
cortisol are observed in human subjects and rodent models (Kile & Turner 1985, Calogero et al.
1990, Contesse et al. 2000, Heisler et al. 2007). In the synapse, cessation of the serotonergic
signal is achieved mainly by the removal of serotonin from the synaptic cleft through an active
membrane transporter encoded for by the serotonin transporter (SERT) gene. Administration of
selective serotonin reuptake inhibitors (SSRI’s), which act on this transporter, is also associated
with a rise in both portal CRH and plasma ACTH (Bevan & Scanlon 1998). Meanwhile, several
studies have indicated a relationship between estrogen and progesterone with serotonin receptor
activity (for review see, Bethea et al 2002). In this way, it becomes increasingly apparent that
there may be changes in serotonergic responsivity during pregnancy and these changes might be
in part responsible for altered basal HPA axis activity.
The objective of this study was to determine if there are alterations in the serotonergic
component of basal HPA axis regulation during pregnancy. There were two parts to this
objective: (1) to compare HPA axis responses to an acute intracerebroventricular (icv) injection
of a selective serotonin reuptake inhibitor, fluoxetine (FLX) in pregnant ewes to the response in
the same ewes post-partum; and (2) to compare responses to icv administration of a lower, but
more chronic dose of fluoxetine in pregnant ewes to non-pregnant ewes. The use of a selective
serotonin reuptake inhibitor in these studies was designed to exploit the inherent serotonergic
activity in these ewes and allow us to observe any differences that might exist relating to
differential regulation of the HPA axis during pregnancy.

17
Specific Aim 3: Relative Hypothalamic Expression of Genes Related to HPA Axis Regulation and the Serotonergic System
For this portion of the dissertation work, we sought to determine whether changes at the
genes level in the hypothalamus might at least in part explain the elevations in ACTH and
cortisol during pregnancy in the ewe. We chose the hypothalamus as it is the point of integration
for all upstream inputs driving or inhibiting HPA axis activity. Therefore, the objective of this
study was to characterize relative mRNA expression levels of HPA axis- and serotonergic
system-relevant genes in the hypothalamus between nonpregnant, pregnant, and post-partum
ewes. Any difference found here might additionally provide insight for any differential HPA
axis responses that might occur between pregnant and nonpregnant or postpartum ewes in
response to MR antagonism or selective serotonin reuptake transporter inhibition discussed in the
preceding specific aims of this dissertation.

18
CHAPTER 2 REVIEW OF LITERATURE
Hypothalamic-Pituitary-Adrenal (HPA) Axis
As the name implies, the hypothalamic-pituitary-adrenal axis comprises the pathway of
communication that exists between certain areas of the hypothalamus, the anterior lobe of the
pituitary gland, and the cortices of the adrenal glands (Figure 2-1). The ‘messages’ that are sent
between these components are in the form of hormones. Within the paraventricular nucleus
(PVN) of the hypothalamus, a subset of neurons called parvocellular neurons synthesize and
secrete both corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP) (Chrousos
& Gold 1992, Chrousos 1992, Tsigos & Chrousos 1994). The axons of these neurons project to
the median eminence and synapse onto the hypophyseal portal system. It is through this path
that CRH is carried to the anterior pituitary where it acts primarily through CRH1 receptors
located on the plasma membrane of pituitary corticotropes. Within the pituitary,
adrenocorticotropic hormone (ACTH) is synthesized as it is cleaved from its pre-cursor
proopiomelanocortin (POMC), whose transcription is stimulated by CRH binding (Fukuda et al.
2003). ACTH is stored intracellularly and is secreted by elevation of cytosolic Ca2+
concentration which is induced by activation of voltage-gated calcium channels or intracellular
calcium release (Luini et al. 1985). Once released, the ACTH travels through the systemic
circulation toward its main target organs, the adrenal glands, or more specifically, the adrenal
cortices. The adrenal cortex is then stimulated to synthesize and secrete corticosteroids. The
two classes of corticosteroids, glucocorticoids and mineralocorticoids, are synthesized and
released from the zona fasciculata and zona glomerulosa of the adrenal cortex, respectively.
Increased corticosteroid synthesis occurs as ACTH binding causes an increase in cholesterol
processing, such as movement into the mitochondria and subsequent conversion to

19
pregnenolone. Pregnenolone ultimately gets shuttled back to the endoplasmic reticulum,
converted to progesterone or 17-hydroxypregnenolone, and after multiple hydroxylation steps
gets converted to aldosterone or cortisol, respectively (Nussey & Whitehead 2001b). In addition
to ACTH stimulation of the adrenal cortex, corticosteroid synthesis and release may also be
regulated by cytokines, angiotensin II, plasma potassium concentration, lipid mediators of
inflammation, and through autonomic innervation of the adrenal glands themselves.
Role of Corticosteroids in Normal Physiology
The primary role of the hypothalamic-pituitary-adrenal (HPA) axis is to aid in the
adaptation to a variety of stressors as a means to maintain homeostasis. This process is called
allostasis or re-establishing “stability through change” (Sterling & Eyer 1988). In the case of the
HPA axis, the process of adaptation is achieved as hormones, acting through receptors, cause
changes at the cellular level. The stressors which evoke such changes can come in many forms
but are mainly classified as either neurogenic (psychological) or systemic (physiological).
Examples of animal models for neurogenic stress are environmental stimuli such as restraint,
electrical footshock, and maternal separation. Systemic stressor models include immunological
challenges or disease states, hemorrhage, and pregnancy. Interestingly, various studies have
uncovered the fact that the stress response circuitry up until activation of the PVN of the
hypothalamus appears to be somewhat specific to the type of stressor the individual is
encountering.
The end result upon activation of the HPA axis is the production and release of
corticosteroids from the adrenal cortex. These hormones are involved in the regulation of a wide
variety of physiologic systems. Two types of corticosteroids are glucocorticoids and
mineralocorticoids. Glucocorticoids and mineralocorticoids classically exert their effects
through two intracellular receptors. These receptors belong to the super-family of nuclear

20
receptors and are located in the cytoplasm and are complexed with heat-shock proteins and
others while in the inactive state. Being lipophilic, the corticosteroids readily enter the cell and
bind the receptor, causing dissociation from these inactivating proteins. After subsequent
phosphorylation, the hormone-receptor complex translocates into the nucleus and dimerizes with
another corticosteroid-receptor complex. The dimer is then able to bind steroid-response
elements found in the promoter region of a variety of target genes, usually with the aid of other
transcription factors thereby stimulating or suppressing transcription of the targeted gene
(Nussey & Whitehead 2001d). Additionally, gene transcription can be influenced through
receptor-ligand complex interactions with other transcription factors (Diamond et al. 1990,
Yang-Yen et al. 1990).
Glucocorticoids come from the zona fasciculata of the adrenal cortex. It has been
established that these adrenal gland molecules are essential to life based on previous research
which showed that bilateral adrenalectomy will result in death if glucocorticoids are not
replaced. Endogenous glucocorticoids, like cortisol in humans and corticosterone in rodents, are
involved in regulating many processes that serve to mobilize energy in order to optimize the
‘fight-or-flight’ mechanism in times of stress. Glucocorticoids are therefore involved in
stimulation of gluconeogenesis, lipolysis and proteolysis to aid in production of fuel sources for
this stress response. In addition, glucocorticoids have been shown to inhibit or suppress innate
immune responses, bone and muscle growth, and reproductive function (McEwen & Stellar
1993). Cortisol is also essential for maintaining normal blood pressure by influencing
myocardial function and arteriolar sensitivity to both sympathetic nervous stimulation and to
angiotensin II. In the brain, glucocorticoids may be involved in cognition, memory and mood as
receptors have been found to be expressed in functionally pertinent regions including the

21
prefrontal cortex, hippocampus and amygdala. Many groups have provided evidence that
cortisol alters neuronal excitability and can induce cell death in certain brain regions, whereas in
other regions glucocorticoids are thought to play a protective role (for review see, McEwen
1994, Nussey & Whitehead 2001a).
Aside from fluctuations in glucocorticoid levels occurring in response to stress, there exists
a circadian rhythm to their basal release. This is most likely due to the connections from the
suprachiasmatic nucleus (SCN) to the paraventricular nuclei of the hypothalamus. The SCN
consists of various cell types containing genes whose expression is responsive to light-sensitive
input signals from the optic tract. CRH is released in a circadian-dependent, pulsatile fashion
from the parvocellular cells of the PVN (Hauger & Dautzenberg 2000). Lesions of the SCN in
rats result in the loss of corticosteroid cyclicity (Moore & Eichler 1972). In humans, the pattern
of diurnal cortisol release is such that the lowest concentration of cortisol is found at about
midnight, continues to rise until it peaks around 9 am, and then gradually declines throughout the
course of the day (Nussey & Whitehead 2001c). This diurnal release of cortisol is often altered
by stress as well as changes in lighting, feeding schedules, and activity (Charmandari et al.
2005). Logically, this pattern is reversed in rats in correlation with their nocturnal habits.
Interestingly, in humans, a recent study suggests that increasing demands for glucose by the
brain over the course of sleep is the main contributor to the early morning rise in cortisol
(Benedict et al. 2009). Similarly, in dogs, there is no distinct rhythm at all but rather pulsatile
release of cortisol in response to episodic secretion of ACTH that may occur in response to
feeding patterns or possibly to the mild stress associated with human interaction (Kemppainen &
Sartin 1984). Such ultradian rhythms have been observed in lactating dairy cows and sheep, as
well (Brinklow & Forbes 1984, Lefcourt et al. 1993).

22
Mineralocorticoids such as aldosterone are produced and released by the zona glomerulosa.
Aldosterone is released in response to activation of the renin-angiotensin-aldosterone system
(RAAS) which occurs in response to a fall in renal perfusion pressure, stimulation by renal
sympathetic nerve afferents, or a reduction in sodium chloride delivery to the macula densa.
Classically, these cues cause an increase in the production of the enzyme renin, which starts the
process of converting angiotensinogen to angiotensin II, a peptide known to stimulate the release
of aldosterone from the adrenal cortex. Additionally, according to most textbooks, aldosterone
release can also be stimulated by an increase in plasma potassium (K+) concentration detected
directly by the adrenal zona glomerulosa cells. Aldosterone, by binding within the cells along
the kidney tubule, can contribute to the regulation of fluid and electrolyte balance by stimulating
K+ excretion into the urine, sodium (Na+) reabsorption and the resultant water retention at both
the distal convoluted segment and collecting duct of the renal tubule. Specifically, it has been
shown that aldosterone activates the epithelial sodium channel (ENaC) by regulating the
abundance and apical distribution of the α-subunit in principal cells (Masilamani et al. 1999). As
discussed in a review by Staub and Verrey (2005), the expression of serum- and glucocorticoid-
induced kinase 1 (sgk1) is rapidly upregulated by aldosterone. This review focuses on work
done in numerous labs to provide evidence that sgk1 phosphorylates Nedd4-2, a ubiquitin-
protein ligase, and thus inhibits its ability to interact with ENaC. Without ubiquitylation, ENaC
maintains its cell surface expression and can therefore aid in Na+ reabsorption in this segment of
the renal tubule. At the same time, aldosterone has the ability to stimulate Na/K ATPase activity
and to increase the driving force for K+ excretion into the tubule lumen. Consequently,
aldosterone plays a major role in regulation of electrolytes and plasma volume and, therefore, in
regulation of blood pressure.

23
Negative Feedback Regulation
The discussion of HPA axis circuitry does not end at the resultant responses of the various
effector organs to corticosteroid binding. Glucocorticoids also play an important role in
regulating the basal activity of the HPA axis and in terminating the stress response by exerting
negative feedback at several sites along the circuit. If unchecked or dysregulated, a chronically
hyper- or hypoactive HPA axis can be damaging and/or exacerbate disease states. The negative
feedback of glucocorticoids on CRH and ACTH release limits the duration of total tissue
exposure to these molecules, thereby minimizing their catabolic, antireproductive, and
immunosuppressive effects. In a review by Charmandari et al (2005), some of the adverse
effects of a hyperactive HPA axis are discussed, such as melancholic depression and insulin
resistance; while a hypoactive axis can contribute to the pathogenesis of fibromyalgia, chronic
fatigue syndromes and susceptibility to autoimmune inflammatory disease.
Likely due to varying mechanisms, elevations in glucocorticoids may cause suppression of
ACTH by a fast (within seconds to minutes), intermediate (over period of 2 to 10 hours) or slow
(which occurs over a period of hours to days in response peak cortisol concentrations) negative
feedback effect (for review see, Keller-Wood & Dallman 1984). The two receptors for
corticosteroid binding, mentioned above, are found in varying concentrations all along the circuit
to exert negative feedback effects. The low-affinity glucocorticoid receptor (GR) binds at
elevated glucocorticoid levels that occur in times of stress; while the high-affinity
mineralocorticoid receptor (MR) binds at both basal and stress concentrations (Keller-Wood &
Dallman 1984, Reul & de Kloet 1985, Reul et al. 1987, Bradbury et al. 1991, de Kloet et al.
1993). MR affinity for corticosterone, the main glucocorticoid in rodents, is 10-fold higher than
that of GR. Therefore, central MR is almost completely occupied at basal corticosterone
concentrations, while GR only becomes substantially occupied during times of stress or at the

24
peak of the circadian cycle (Reul & de Kloet 1986, De Kloet et al. 1998). Central glucocorticoid
receptors have widespread distribution throughout the brain, but are most densely expressed in
areas that are have been shown to be involved with regulation of the stress response such as the
PVN of the hypothalamus, the corticotrophs of the anterior pituitary, and the hippocampus (Reul
& de Kloet 1986).
It is important to note, both aldosterone and corticosterone (in rodents) and cortisol (in
humans) bind MR with subnanomolar affinity, but the glucocorticoids circulate at 1000-fold
higher concentrations (100-fold higher even after considering the presence of corticosteroid
binding proteins in the plasma). Experiments done in vitro have shown that the aldosterone is 10
times more potent at stimulating transcriptional changes by binding to MR (Arriza et al. 1988).
However, there is marginal brain penetration of aldosterone due at least in part to multidrug
resistance protein (mdr1 or P-glycoprotein), a protein transporter at the blood-brain barrier which
pumps select substrates back into the blood (Ueda et al. 1992, Uhr et al. 2002, Parker et al.
2006). Therefore, the vast majority of central mineralocorticoid receptors are occupied by
glucocorticoids rather than aldosterone.
Mineralocorticoid receptors have been studied extensively within the central nervous
system of many rodent models (Reul & de Kloet 1985, Luttge & Rupp 1989, Funder 1996). In
the rodent brain, MR have been found primarily in the hippocampus and septum. In fact, regions
of the hippocampal formation express both receptors and this is not surprising as it is thought to
play an inhibitory role in regulation of the HPA axis through its indirect connections to the PVN,
such as those made through the septal nucleus of the stria terminalis (Sapolsky et al. 1986,
Jacobson & Sapolsky 1991, Herman & Cullinan 1997, Herman & Mueller 2006). It is thought
that hippocampal neuronal inputs activate inhibitory gamma-aminobutyric acid- (GABA-)

25
releasing neurons that project to CRH neurons in the PVN. Evidence in support of this
relationship, as discussed in a review by Carrasco and Van de Kar (2003), include the use of
synthetic mineralocorticoid receptor antagonist RU28318 to elevate baseline corticosterone
levels in rats which can be blocked by the GABAA receptor agonist alprazolam (Grottoli et al.
2002). Distribution and function of mineralocorticoid receptors at the hippocampal level has
recently been investigated in other species. Studies characterizing corticosteroid receptor
distribution in the primate brain, found that MR mRNA and protein levels were much higher in
the dentate gyrus (DG) and Cornu Ammonis (CA) of the hippocampus (Sanchez et al. 2000).
Additionally, electrical stimulation of the hippocampus in humans and cats produces a decrease
in the plasma levels of cortisol, supporting an inhibitory role of the hippocampus on HPA axis
activity in these species as well (Carrasco & Van de Kar 2003). Like the rodent and primate
models, sheep are also known to express both MR and GR within the main regulatory areas of
the HPA axis: the hypothalamus, hippocampus, and pituitary (Roesch & Keller-Wood 1999).
Role of Corticosteroids in Pregnancy and Parturition
Studies in our laboratory and others have shown that cortisol, a key corticosteroid, is
basally regulated at higher levels in the pregnant state than in the non-pregnant state (Carr et al.
1981, Bell et al. 1991, Keller-Wood 1998, Erickson et al. 2001, Sandman et al. 2006,
Kirschbaum et al. 2009). Additionally, there is a marked increase in plasma aldosterone by the
8th week of gestation in human pregnancy which continues to rise throughout pregnancy to levels
4 to 6 fold higher than nonpregnant levels (Watanabe et al. 1963). Our lab, specifically, has
shown that corticosteroids are important for maternal volume expansion, uterine blood flow, and
fetal homeostasis (Jensen et al. 2002, Jensen et al. 2003, Jensen et al. 2005). In these studies,
pregnant ewes were adrenalectomized at 112 days of gestation and were under-replaced for
either cortisol or aldosterone to levels observed in nonpregnant ewes. It was determined that

26
reducing either corticosteroid prevented the increase in maternal plasma volume that occurs from
120 days to 130 days of gestation. Several adaptive responses were observed in the fetuses of
these ewes such as reduced lung-liquid and urine production presumably to combat the effects
that maternal hypovolemia might have on their own plasma volume. Additionally, in these
pregnant ewes which were inadequately replaced after adrenalectomy, the normal increase in
uterine and placental blood flow that occurs as gestation progresses was attenuated.
Consequently, Jensen et al (2005) reported adverse effects on fetal development such as
impaired fetal somatic growth rate and arterial oxygen tension. In support of these findings, case
studies in humans have shown that pregnancies with untreated adrenal insufficiency are
associated with high maternal and fetal morbidity and mortality (Lindsay & Nieman 2006). At
the same time, a link has been proposed between rising cortisol and changes in maternal basal
metabolic rate and body weight during pregnancy (Damjanovic et al. 2009).
Also important to note, the HPA axis has been implicated for many years in fetal organ
development and in the initiation of parturition. In a review by Liggins (1994), the role of
cortisol as it relates to organ maturation in the fetus is discussed. Cortisol is responsible for
regulating important proteins in many organ systems that are necessary for adaption to the extra-
uterine environment. The fetal lung for example, must rapidly absorb the liquid that has filled
each lobe throughout gestation as well as produce phospholipid surfactant to prevent alveolar
collapse by reducing surface tension. In fact, Liggins and Howie in (1972) first introduced the
idea of giving antenatal glucocorticoids to mothers who were threatening premature labor in
order to prevent respiratory problems in the neonate. Another example of cortisol-mediated
organ maturation is increased production of tri-iodothyronine by the thyroid gland which occurs

27
to address the need for increased thermogenesis and the higher metabolic rate that is necessary
for adaption to the colder external air and increased energy expenditure of breathing.
The role of cortisol in the initiation of parturition is currently being investigated. The
placenta is known to be an extrahypothalamic cite of CRH production and is primarily
responsible for the increased plasma CRH in late gestation (Jones et al. 1989, Zoumakis et al.
1996). Plasma CRH concentrations peak during labor and immediately decline postpartum
(McLean et al. 1995). Contrary to the inhibitory effect on hypothalamic production of CRH,
cortisol is thought to stimulate placental CRH production (Gonen et al. 1992). The maturing
fetal HPA axis near term is thought to contribute to this increase in placental CRH as well and is
the basis for the ‘placental clock’ theory of the timing of parturition (McLean et al. 1995).
Additionally, norepinephrine, angiotensin II and vasopressin which may be increased during
times of stress can also stimulate CRH production by the placenta (Jones et al. 1989, Petraglia et
al. 1989, Petraglia et al. 1991). These findings have lead to the theory that maternal or fetal
stress can lead to premature delivery in humans. The role of CRH in parturition is thought to
include its ability to stimulate of prostaglandin production in placenta in vitro (Jones & Challis
1989, Jones & Challis 1990). The prostaglandins are then thought to increase myometrial
oxytocin receptor levels and gap junctions, thereby enhancing the myometrial response to
oxytocin (Neulen & Breckwoldt 1994, Grazul-Bilska et al. 1996) as well as influence
extracellular matrix remodeling in the cervix. In vitro and in isolated placental tissues,
prostaglandins stimulate CRH release which would indicate a positive feedback relationship
(Jones & Challis 1989, Jones & Challis 1990, Petraglia et al. 1991). Although controversial,
elevated plasma CRH concentration midgestation has been suggested as a marker for preterm
delivery (Wolfe et al. 1988, Warren et al. 1992, McLean et al. 1995, Lockwood et al. 1996).

28
Serotonergic System
I will now direct attention to the serotonergic system, which has previously been shown to
be associated with HPA axis activation. The interactions of these two systems are particularly
evident in pathological conditions such as major depression, which is characterized by
dysregulation of both systems (for review see, Lanfumey et al. 2008).
In the mid-nineteenth and early twentieth centuries, a substance was being isolated in
platelets that was capable of constricting vascular smooth muscle. Due to its "tonic" action and
locale in "serum," the substance was given the name serotonin. Serotonin is found in a variety of
organ systems such as the intestinal mucosa and serum. Approximately 1–2% of the body’s
serotonin is contained in the central nervous system, specifically in serotonergic neurons
(Lozeva-Thomas 2004). Within the central nervous system, serotonin serves as a
neurotransmitter which is likely involved in the regulation of feeding behavior, body weight, the
sleep–wake cycle, circadian rhythmicity, locomotion, and learning and memory (Jacobs &
Azmitia 1992, Jacobs & Fornal 1999, Cooper et al. 2003). Serotonin has also been implicated in
several pathological conditions such as migraine, obsessive-compulsive disorders, depression
and suicidal behavior, bipolar disorder, schizophrenia, narcolepsy, alcohol dependence, obesity,
and chronic fatigue syndrome (Jacobs & Azmitia 1992, Jacobs & Fornal 1999, Cooper et al.
2003).
Within the central nervous system, serotonergic cell bodies are primarily found in discrete
clusters or groups of cells along the midline of the brain stem while their axons, however,
innervate nearly every area of the central nervous system. These clusters of cell bodies were
named and classified by Olszewski and Baxter in 1954. The nuclei found in the more caudal
region of the brain stem are thought to be involved in modulating sensory and motor processing
of the spinal cord, whereas the cells of the rostral nuclei send long axons to the forebrain.

29
Immunohistochemical staining techniques for either 5-HT or tryptophan hydroxylase have been
utilized to further characterize serotonergic innervation in the various forebrain regions (Frazer
& Hensler 1999).
In the cytosol of serotonergic neurons, a two step process occurs to synthesize serotonin or
5-hydoxytryptamine (5-HT) from the precursor amino acid, l-tryptophan. The primary source of
the tryptophan is dietary protein. Facilitated transport is necessary to move l-tryptophan from
the blood into the brain. This process is regulated not only by the concentration of tryptophan,
but also by the presence of other amino acids that compete for this type of transport. Therefore,
lowering intake of tryptophan relative to these other amino acids can affect the amount of
serotonin synthesis. In fact this strategy has been used to help elucidate the importance of brain
5-HT. Once tryptophan has crossed the blood-brain barrier and entered a serotonergic cell body,
it is hydroxylated to 5-hydroxytryptophan (5-HTP) catalyzed by tryptophan hydroxylase, the
rate-limiting step in the synthesis of serotonin. Activation of this enzyme requires
phosphorylation and the accessory protein 14-3-3 (Ichimura et al. 1995, Kuhn et al. 1997). Next,
5-HTP undergoes a decarboxylation reaction which is catalyzed by l-amino acid decarboxylase
to form serotonin. As is the case for many other neurotransmitters, serotonin is then stored
predominantly in vesicles which release their contents in response to an action potential passing
down the axon of the neuron. Serotonergic activity is then determined by the processes of
serotonin synthesis, reuptake, neuronal activity, degradation by monoamine oxidase (MAO), and
pre- and post-synaptic receptor activation.
There are thought to be at least seven major families which are further classified into at
least 14 subtypes of serotonin receptors. Serotonin binding at these receptors can either directly
stimulate or inhibit the activity of the target cell, whereas the presence of these receptors on

30
GABA-ergic interneurons can allow for indirect modulation of their downstream target cells as
well (Uphouse 1997). The following information regarding the serotonin receptors investigated
in this dissertation has been summarized from a textbook chapter entitled ‘Serotonin’ by Frazer
and Hensler (1999). The 5-HT1 family consists of receptors that are negatively coupled to
adenylyl cyclase and therefore, receptor activation causes a decrease in cAMP which reduces the
amount of kinase activation. The 5-HT1A receptor is additionally coupled to the opening of K+
channels, which results in neuronal hyperpolarization. These receptors are coupled to both
effector systems at the nerve terminus, but in the dorsal raphe nucleus, 5-HT1A receptors are
coupled only to the opening of potassium channels. 5-HT1A receptors have been shown to be
present in high density in the hippocampus, septum, amygdala, hypothalamus and neocortex.
The next family, the 5-HT2 receptors are G-protein coupled receptors as well, stimulating
phosphoinositide hydrolysis through activation of phospholipase C. Contrary to 5-HT1A
receptors, activation of 5-HT2A receptors mediates neuronal depolarization due to closing of
potassium channels. This family of receptors is particularly concentrated in the frontal cortex, in
parts of the limbic system, and the PVN of the hypothalamus.
Cessation of the serotonin signal has also been characterized. The majority (80%) of the
serotonin released into the synaptic cleft is removed through an active membrane transporter on
the pre-synaptic neuron encoded for by the serotonin transporter (SERT) gene. If not actively
transporting serotonin, the SERT protein is phosphorylated and subsequently degraded
(Ramamoorthy & Blakely 1999). Deactivation of free serotonin both in the synapse and in the
nerve terminal also occurs via a reaction catalyzed by mitochondrial monoamine oxidase (MAO)
which converts 5-HT to 5-hydroxyindoleacetic acid (Lozeva-Thomas 2004).

31
Impact of the Serotonin System on HPA Axis Activity
Early studies had already established a spatial relationship between the serotonergic
systems and the HPA axis. Using immunocytochemistry techniques, researchers demonstrated
the existence of serotonergic synapses on CRH-containing neurons of the PVN (Liposits et al.
1987). The major brain stem source of serotonergic innervation to the PVN of the hypothalamus
is from the dorsal raphe nuclei (Azmitia & Segal 1978). More recently, in situ hybridization
studies have uncovered moderately dense populations of 5-HT2A/2C binding sites and 5-HT2A
transcripts within the hypothalamic PVN.
It was also shown in vivo by Tsagarakis et al (1989) that hypothalamic implants of
serotonin directly stimulate CRH release; while Calogero et al (1989) demonstrated the same
effect in hypothalamic cultures. A review by Carrasco and Van de Kar (2003) discusses a
number of studies in rodents have shown the stimulatory effects of serotonergic agents of plasma
ACTH and corticosterone (the primary glucocorticoid in these species). Additionally, upon
intravenous injection of serotonin or oral administration of serotonin or its precursors, tryptophan
or 5-HTP, a significant increase in plasma cortisol is observed in human subjects and other
animal models such as sheep (Kile & Turner 1985, Calogero et al. 1990, Fuller 1996, Broadbear
et al. 2004, Broadbear et al. 2005, Heisler et al. 2007). Drugs that are designed to potentiate the
actions of serotonin, such as selective serotonin reuptake inhibitors (SSRIs), are also associated
with a rise in both portal CRH and plasma ACTH (Bevan & Scanlon 1998). The association of
these two systems is even being explored in fish. One group of researchers in Sweden has been
characterizing the hypothalamic-pituitary-interrenal axis, which is analogous to the HPA axis.
The end-product of this axis is also cortisol; and a recent study showed that tryptophan-
supplemented fish without exposure to a stressor exhibit elevated cortisol levels (Lepage et al.
2002).

32
Several studies have focused on elucidating exactly which 5-HT receptor is responsible for
stimulation of the HPA axis. Gartside and Cowen (1990) found the serotonin precursor 5-HTP
given intraperitoneally (ip) to dose dependently increase plasma ACTH in the male rat. These
responses were attenuated by pretreatment with the non-selective 5-HT receptor antagonist and
also by a variety of selective 5-HT2 receptor antagonists. The 5-HT1 receptor antagonists used in
those studies failed to antagonize these responses suggesting that the increases in ACTH with ip
injection of 5-HTP in the male rat are mediated by 5-HT2 receptors. Another study which
utilized systemic administration of 5-HTP in conjunction with the 5-HT reuptake inhibitor
fluoxetine, found a 64% increase in CRH mRNA expression within the PVN and a 17% increase
in POMC mRNA within the anterior pituitary. In agreement with the previous study, this group
also found ACTH secretion to be elevated five-fold. Through their use of specific 5-HT receptor
subtype agonists and anti-CRH antiserum, this group have indicated a number of serotonin
receptors might be responsible for increases in CRH and POMC synthesis including 5-HT1A and
5-HT2A and that these effects are mediated by CRH (Jorgensen et al. 2002).
The hippocampus, which is thought to be a site for HPA axis regulation as previously
mentioned, receives dense serotonergic innervation from the raphe nuclei as well and the
involvement of this line of communication in HPA axis regulation is currently being explored.
In studies utilizing parachlorophenylalanine, a specific 5-HT synthesis inhibitor, Semont et al
(1999) detected a significant increase in the number of hippocampal MR-binding sites. Upon
injection of the precursor, 5-HTP, MR-binding site levels were restored down to control levels.
No change in number of GR-binding sites was detected; however, hippocampal GR mRNA
levels were reduced. These researchers propose that hippocampal MR synthesis is inhibited by
5-HT and that this effect is not mediated by changes in hormone secretion of HPA axis. In

33
contrast, the expression of both glucocorticoid and mineralocorticoid receptor mRNA in
hippocampal cells has been reported to be upregulated upon activation of 5-HT receptors in vitro
(Seckl & Fink 1991, Lai et al. 2003). Moreover, in studies using reserpine, a substance which
depletes all monoamines, a reduction in both MR and GR levels is seen in hippocampal cytosol
(Lowy 1990). Additionally, lesions of the serotonergic projections to the hippocampus of
adrenalectomized rats also results in decreased levels of MR and GR mRNA in sub-regions of
the hippocampus (Seckl et al. 1990).
Finally, it is important to note that the complexity of the relationship between the
serotonergic system and the HPA axis is further complicated by the fact that the serotonergic
neurons themselves are known to express corticosteroid receptors. This would therefore imply
that they have the potential to be regulated by circulating glucocorticoid, directly (Harfstrand et
al. 1986, Fuxe et al. 1987, Morimoto et al. 1996).
Serotonergic System on Regulation of Food Intake
Serotonin is thought to play a role in food intake regulation as evidenced by the
anorexogenic effects of serotonergic agents, such as selective serotonin reuptake inhibitors and
serotonin releasing agents (Heisler et al. 2003). A brief overview on basic central food intake
circuitry is necessary to understand the proposed involvement of serotonin in these pathways. A
vast array of gastrointestinal, pancreatic, and adipocytic hormones which are responsive to
physical and chemical cues regarding meal size and energy stores are known to centrally regulate
food intake (Kelley & Berridge 2002, Dhillo & Bloom 2004, Woods et al. 2006, Naslund &
Hellstrom 2007). These hormones act via the nucleus of the solitary tract (NTS) or directly at
brain stem as well as hypothalamic and corticolimbic nuclei that are known to regulate feeding
(Swanson 2000, Browning & Travagli 2006). One such area of the hypothalamus, the arcuate
nucleus (ARC), consists of two neuronal populations, proopiomelanocortin (POMC) neurons and

34
those which co-express agouti-related protein (AgRP) and neuropeptide Y (NPY). These
neurons are known to be responsive to the peripheral hormones discussed above; therefore the
ARC can subsequently integrate an array of signals governing food intake (for review see,
Cummings & Overduin 2007). Figure 2-2 depicts basic signaling circuitry for inhibiting food
intake mediated by the peripheral hormones ghrelin (from the stomach) and leptin (from
adipocytes) acting at the neuronal systems. Briefly, leptin stimulates POMC neurons to release
melanocortins (cleavage products of the precursor POMC) which then act on melanocortin
receptors and ultimately inhibit feeding (Cone 2006). Meanwhile, ghrelin activates the
AgRP/NPY neuronal population. AgRP is thought to be an inverse agonist at melanocortin
receptors and therefore its effects are orexogenic (Chen et al. 2004).
It has been postulated that serotonergic agents which decrease body weight and food intake
are doing so by acting directly on the neurons of the arcuate nucleus. Heisler and colleagues
(2002) demonstrated fos-like immunoreactivity (FOS-IR) induction, a marker of neuronal
activation, in ARC POMC neurons of rats that have been given anorectic doses of fenfluramine
which blocks 5-HT reuptake while also stimulating its release. Additionally, they reported
consistent depolarization of ARC POMC neurons upon application of fenfluramine to coronal
hypothalamic slices from transgenic mice with POMC promoter-controlled green fluorescent
protein expression. Furthermore, this group found that in melanocortin receptor (MCR)
knockout mice and in those that have been given the MCR antagonist SHU9119, fenfluramine
exhibited limited efficacy at reducing food intake and body weight.
However, the mechanisms of serotonergic involvement in feeding regulation are likely to
be more complicated than simply its action at the arcuate nucleus of the hypothalamus. As
discussed earlier, it is known that in mammals, divergent serotonergic axons arise from the raphe

35
nuclei, thereby innervating a large proportion of the forebrain structures, as well as other regions
of the brain stem and spinal cord. In fact, all brain nuclei implicated in energy balance
regulation receive serotonergic afferents (for review see, Tecott 2007).
Evidence for Influence of Ovarian Hormones
In addition to the connections and receptor populations discussed above, the hypothalamus
is also densely populated with receptors for the ovarian hormones, estrogen and progesterone
(Bethea et al. 1996). Additionally, other brain regions upstream of the hypothalamus that are
known to regulate HPA axis activity such as the hippocampus and dorsal raphe nuclei express
these receptors as well. Several studies have demonstrated elevations in basal HPA axis activity
during periods of the estrous cycle that are defined by higher levels of progesterone and estrogen
(Raps et al. 1971, Pollard et al. 1975, Ogle & Kitay 1977, Buckingham et al. 1978, Phillips &
Poolsanguan 1978, Carey et al. 1995). The rise in CRH mRNA expression in the PVN in the
afternoon of pro-estrus in the rat has been attributed to estradiol (Bohler et al. 1990), however
when estrogen is administered in a chronic low dose, CRH expression has been shown to
decrease (Dayas et al. 2000). Another group has shown that in primates, estrogen given in a
manner that mimics the preovulatory surge increases CRH expression in the PVN (Roy et al.
1999), but if given chronically, estrogen or progesterone or both will decrease CRH mRNA and
protein (Bethea & Centeno 2008). Estrogen and progesterone are both known to increase
throughout the course of gestation as well and therefore might be at least partially responsible for
changes in HPA axis regulatory mechanisms during pregnancy.
Numerous reports indicate that progesterone can act as an MR antagonist (Rupprecht et al.
1993) and that in the presence of progesterone (Carey et al. 1995 and unpublished data from our
lab) or during pregnancy (Roesch & Keller-Wood 1999), cytosolic availability of hippocampal
MR is increased. Taken together, these findings suggest that progesterone might interfere with

36
cortisol’s ability to negative feedback at hippocampal MR. Contradictory effects of estrogen on
MR have been found as well which include estradiol-induced decreases in hypothalamic and
hippocampal MR binding capacity and mRNA levels (Carey et al. 1995, Castren et al. 1995)
while other groups reported increases in these same regions (Ferrini & De Nicola 1991) or no
change to hippocampal binding capacity in response to longer exposure to estradiol (Burgess &
Handa 1992). Several groups have demonstrated that progesterone will attenuate the estradiol-
induced decreases in hippocampal MR binding capacity, but progesterone will have no effect on
hippocampal mRNA when given alone (Carey et al. 1995, Castren et al. 1995).
Estrogen has been shown to reduce GR binding and mRNA in the anterior pituitary,
hypothalamus, and hippocampus (Peiffer & Barden 1987, Turner 1990, Turner 1992, Burgess &
Handa 1993), but no changes in GR mRNA expression have been found across the estrous cycle
(Sliwowska et al. 2008). It has been proposed that downregulation requires long term exposure
to estrogen (Burgess & Handa 1993, Redei et al. 1994). Our lab has reported no effect of
pregnancy on hippocampal expression of MR or GR in the ewe (unpublished data by Yi Hua).
Additional work by Carey et al (1995) suggests that neither estrogen nor progesterone influences
GR binding in the hippocampus. However, progesterone has also been proposed to be a partial
agonist at GR since binding causes translocation, however it is not entirely clear whether the
necessary conformational change occurs that would allow for transcription factor loading
(Rupprecht et al. 1993, Nordeen et al. 1995).
Meanwhile, ovarian hormones have been reported to cause changes in several components
of the serotonergic system. A review by Bethea et al (2002) describes the large variability in
ovarian hormone influence on serotonin synthesis across species. Estrogen was reported to
increase SERT binding in the rat hypothalamus (Mendelson et al. 1993, McQueen et al. 1997)

37
but decrease binding in the hippocampus (Mendelson et al. 1993). In contrast, Pecins-Thompson
et al (1998) found that estrogen decreased SERT mRNA in the dorsal raphe in nonhuman
primates. Additionally, the same group found that long-term exposure to estrogen decreases
mRNA expression of the 5-HT1A autoreceptor in the dorsal raphe (Pecins-Thompson et al. 1998,
Pecins-Thompson & Bethea 1999). At the level of the hypothalamus, estrogen has been shown
to attenuate 5-HT1A-stimulated increases in ACTH and corticosterone, and this effect appears to
be related to estrogen’s ability to reduce levels of G-proteins that are known to mediate the
actions of this receptor (Raap et al. 2000), but according to Frankfurt et al (1994) the density of
5-HT1A receptors in the hypothalamus is not altered by estrogen. Estrogen has previously been
reported to increase 5-HT2A receptors several non-hypothalamic regions in rats (Biegon et al.
1983, Sumner & Fink 1995, Sumner & Fink 1997, Cyr et al. 1998, Osterlund & Hurd 1998), but
more recently was shown to have no effect on mRNA levels in the hypothalamus of nonhuman
primates (Gundlah et al. 1999). Undoubtedly, there is strong evidence for estrogen and
progesterone modulation of several components which mediate the actions of serotonin in many
brain regions and across many species; however, the directionality and sensitivity of the
responses are quite variable. This is likely due the variability in the receptor milieu and/or
intracellular machinery present in these systems.

38
Figure 2-1. A general model of HPA axis regulation. Corticosteroids negatively feedback at
each level of the axis via mineralocorticoid (MR) and glucocorticoid (GR) receptors; while other systems, including the hippocampus and serotonergic nuclei in the brain stem are also thought to regulate its activity.

39
Figure 2-2. Basic regulatory circuitry for food intake mediated by the arcuate nucleus (ARC) of
the hypothalamus. Leptin from adipocytes stimulates proopiomelanocortin (POMC) neurons which release melanocortins such as α-melanocyte stimulating hormone (α-MSH, a cleavage product of POMC) which then act on melanocortin receptors to ultimately inhibit feeding by stimulating anorexogenic peptides in the paraventricular nuclei (PVN) and inhibiting orexogenic peptides in the lateral hypothalamic area (LHA). Ghrelin from the stomach activates the agouti-related protein (AgRP)/neuropeptide Y (NPY) neuronal population. AgRP is thought to be an inverse agonist at melanocortin receptors and therefore its effects are orexogenic.

40
CHAPTER 3 ROLE OF MINERALOCORTICOID RECEPTORS IN REGULATION OF CORTISOL,
ALDOSTERONE, ELECTROLYTES, AND BLOOD PRESSURE IN PREGNANCY
Introduction
One of the main focuses of our lab is to investigate alterations in the regulation of basal,
maternal hypothalamic-pituitary-adrenal (HPA) axis activity that arise during pregnancy. In both
human (Carr et al. 1981, Erickson et al. 2001, Sandman et al. 2006, Kirschbaum et al. 2009) and
sheep (Bell et al. 1991) studies, basal plasma adrenocorticotropic hormone (ACTH) and cortisol
have been shown to increase during pregnancy. From previous studies in our lab, we understand
that this increase in corticosteroid secretion in pregnancy contributes to the maternal volume
expansion and increased uterine blood flow that must occur to protect the health of the mother
and to insure a supportive environment for the developing fetus (Jensen et al. 2002, Jensen et al.
2003, Jensen et al. 2005). Additional studies in our lab in adrenalectomized sheep have shown
that the concentration for intravenous cortisol replacement required to normalize basal plasma
ACTH is increased for the pregnant ewes and that supplementation with nonpregnant cortisol
levels increases hypotension-stimulated ACTH release (Keller-Wood 1998, Keller-Wood &
Wood 2008). On the other hand, in studies using adrenal intact ewes, ACTH feedback
suppression by raising plasma cortisol above resting levels is not different during pregnancy
(Keller-Wood 1996). It has therefore been theorized by this lab and others that there is an
alteration in ‘set-point’ of the negative feedback regulation of basal HPA axis activity that may
at least in part explain the elevations of maternal plasma ACTH and cortisol seen in pregnancy.
There are a variety of endogenous mechanisms that control the overall activity of the HPA
axis, both basally and in times of stress, none of which have been completely characterized. It is
now known that corticosteroids exert their actions, such as feedback inhibition, through binding
to intracellular receptors, known as glucocorticoid receptors (GR) and mineralocorticoid

41
receptors (MR). It has been postulated through work in several species that activation of central
and pituitary GR leads to feedback inhibition of stress-induced HPA axis activation, thereby
reducing ACTH and cortisol secretion; while activation of the higher affinity MR mainly
expressed in the hippocampus inhibits the activity of the HPA axis at basal corticosteroid levels
(Keller-Wood & Dallman 1984, Reul & de Kloet 1985, Reul et al. 1987, Bradbury et al. 1991,
de Kloet et al. 1993, Reul et al. 2000). For the purposes of studying alterations in basal HPA
axis regulation during pregnancy, this study aims to characterize the relative role of MR in the
pregnant ewe.
MR have been studied extensively within the central nervous system of many rodent
models (Reul & de Kloet 1985, Luttge & Rupp 1989, Funder 1996). In the rodent brain, MR
have been found primarily in the hippocampus and septum. In fact, regions of the hippocampal
formation express both receptors and this is not surprising as it is thought to play an inhibitory
role in regulation of the HPA axis through indirect connections to the PVN, such as those made
through the septal nucleus of the stria terminalis (Sapolsky et al. 1986, Jacobson & Sapolsky
1991, Herman & Cullinan 1997, Herman & Mueller 2006). More recent studies characterizing
corticosteroid receptor distribution in the primate brain, also found that MR mRNA and protein
levels were much higher in the dentate gyrus (DG) and cornu ammonis (CA) of the hippocampus
than other brain regions (Sanchez et al. 2000). In sheep, both receptors are expressed within the
main regulatory areas of the HPA axis: the hypothalamus, hippocampus, and pituitary (Roesch &
Keller-Wood 1999). Our lab has also shown that the hippocampus has increased cytosolic MR
availability in pregnant ewes compared to nonpregnant ewes and a tendency, although not
significant, toward increased MR availability in the hypothalamus and pituitary as well. These
findings suggest reduced MR activation despite the higher plasma cortisol levels that occur with

42
pregnancy (Roesch & Keller-Wood 1999). We therefore hypothesize that the importance of MR
in feedback inhibition is altered in the pregnant state, allowing HPA activity to be increased.
Furthermore, renal MR are classically known to be important in electrolyte balance and
plasma volume regulation through its effects at the distal nephron. Specifically, when a drop in
renal perfusion pressure, stimulation by renal sympathetic nerves, or a reduction in sodium
chloride delivery is detected at the macula densa, aldosterone release from the adrenal cortex is
triggered by angiotensin II via increased activity of the renin-angiotensin-aldosterone system.
Additionally, aldosterone release can be stimulated directly when increased plasma potassium
concentration is detected by the adrenal zona glomerulosa cells. MR binding by aldosterone
starts the chain of events to increase production of proteins that are involved in active sodium
(Na+) reabsorption by principal cells along the distal tubule and collecting duct. Meanwhile,
potassium excretion is increased due to the actions of aldosterone on both Na/K ATPase activity
and on apical conductance of K+.
It has been reported previously that spironolactone has a very low affinity for the
glucocorticoid receptor (GR) relative to MR (Couette et al. 1992, Rupprecht et al. 1993).
Canrenoate potassium, the active metabolite of spironolactone, penetrates the blood-brain barrier
and in humans has a half-life of 3.7 ± 1.2 hrs (Funder et al. 1974, Rothuizen et al. 1993).
Previous reports have shown that administration of MR antagonists in animal and human studies
elevates plasma cortisol (Dodt et al. 1993, Young et al. 1998, Arvat et al. 2001, Grottoli et al.
2002). In the current study, by infusing canrenoate intravenously, we expect to effectively
antagonize both central and renal MR as a means to characterize the relative importance of MR
actions in the regulation of adrenal hormone secretion, electrolytes, and blood pressure in
pregnancy in the ewe. We therefore hypothesize that if reduced central MR action is at least in

43
part responsible for elevated cortisol during pregnancy, and if hemodynamic changes such as
plasma volume and blood pressure that occur during pregnancy are at least in part mediated by
MR, then systemic MR antagonism will result in attenuated HPA axis stimulation and greater
hemodynamic challenges in pregnant ewes compared to nonpregnant ewes.
Materials and Methods
Animals
Animals were housed in climate controlled, individual pens located in the University of
Florida Animal Care Facility; all animal use was in accordance with the rules and regulations of
the Institutional Animal Care and Use Committee (IACUC) at the University of Florida. Mixed
western breed ewes, both nonpregnant (NP: n=6) and pregnant (P: n= 6, at 136 ± 3 days of
gestation; term is approximately 147 days), were used in this study.
Surgical Protocol
Before surgery, food was withheld from the ewe for 24 hours. All surgeries were
performed in the surgery suite of the Health Science Center, Animal Resources Department.
Animals were prepared for and underwent surgery under aseptic conditions. Animals were
induced with isoflurane inhalant and maintained using either isoflurane or halothane inhalant (1-
3% in oxygen).
Sterile bilateral polyvinyl catheters (inner diameter (ID): 0.050in, Tygon® Microbore
Tubing, Saint-Gobain Performance Plastics Corp., Akron, Ohio) were placed into the femoral
arteries and veins of both nonpregnant and pregnant ewes. A trocar device was then used to
direct the catheters to an exit site near the flank of the ewe. For a five day recovery period, the
animals received twice daily intramuscular injections of ampicillin (1g); body temperature was
also monitored twice daily. Post-operative care during the recovery period and through the
remainder of the study also consisted of daily exit site cleansing with a povodine iodine solution.

44
Experimental Protocol
At the end of the recovery period, each ewe was randomly assigned to one of two
treatment regimens: (1) intravenously (iv) infused with the MR antagonist canrenoate in saline
(CAN: 4 mg/kg bolus followed by 1 mg/kg/hr infusion for 4 hours) or (2) saline (VEH) at the
same infusion rate (total volume infused for both treatment groups was 40 ml over 4 hours).
Two days later, the experiment was repeated for each ewe using the other of the two treatments.
Because this study focuses on basal HPA axis activity, access to arterial and venous
catheters was achieved using methodology previously established by this lab that minimizes
human interaction with the ewes (Bell et al. 1991). Briefly, the catheters were externalized from
the pens via a swiveling duct system, prior to each experiment. Following an acclimation period
of 1 hour, it was then possible to re-enter the room quietly in order to access the catheters
remotely from just outside the pen. Venous catheters were attached to syringes that were
positioned into syringe pumps containing either saline or canrenoate in saline. Arterial catheters
were filled with heparinized saline at the start of the experiment and after each sample; and this
‘dead-space’ was removed just prior to the subsequent sample. Plasma was acquired from
samples taken just prior to the start of the infusion and then at every hour during the infusion by
collecting 8 ml of whole blood into tubes containing 400 µL 0.3M ethylenediaminetetraacetic
acid (EDTA) followed by centrifugation at 3000 x g for 20 min at 4°C. Plasma was transferred
to a clean tube and stored at −20oC for future analysis. At the time of the experiment, separate
1.5 ml whole blood samples collected into heparinized syringes were used for determination of
hematocrit and total protein as well as for determination of plasma sodium and potassium
concentrations using ion specific electrodes (AVL 9180 Electrolyte Analyzer, AVL/Roche
Diagnostics, Roswell, Georgia). Basal mean arterial blood pressure (MAP) was recorded
continuously for the 4 hours of infusion (except during the brief sampling periods) via pressure

45
transducers connected to an analog-to-digital conversion board (LabView, National Instruments,
Austin, Texas). Blood pressures were taken at a sampling rate of 60 Hz and later averaged into
5-min bins for statistical analysis.
Plasma Hormone Determination
Plasma cortisol was determined using ELISA (Oxford Biomedical Research, Oxford,
Michigan). Plasma ACTH was determined by radioimmunoassay (RIA) as previously described
using an antibody to 1-39 ACTH (Bell et al. 1991). Plasma progesterone and aldosterone were
determined by RIA using the 125I Coat-A-Count® progesterone and aldosterone kits (Siemens
Healthcare Diagnostics Inc., Deerfield, Illinois). Plasma Angiotensin II levels were determined
by RIA after extraction from plasma using acetone, a method previously used in this laboratory
(Pecins-Thompson & Keller-Wood 1997).
Plasma Volume Determination
At the end of each experiment, the total plasma volume was determined using the Evans
blue dye dilution method (Pecins-Thompson & Keller-Wood 1997). More specifically, after the
last sample of the experiment and prior to injection of Evans blue dye, 10 ml of whole blood was
collected into EDTA-treated tubes. The plasma from this sample was used as the ‘no-dye’
sample and also as the vehicle for the standard curve of the plasma volume assay.
Approximately 2.5-4 ml (15-25 mg in saline) was injected into the venous catheter, followed by
saline (enough to cover the length of the catheter dead-space). Starting at 10 min post-Evans
blue injection, 2 ml of whole arterial blood was collected into EDTA-treated tubes at 5 min
intervals until 45 min. Following centrifugation, the plasma was transferred to clean tubes.
Evans blue concentration in the plasma was determined by measuring the absorbance at 620 nm
in a Synergy HT Multi-Mode Microplate reader (Bio Tek Instruments, Inc., Winooski, Vermont)
in triplicate and extrapolating the concentration from the standard curve. Using the

46
concentration of Evans blue at each time-point for each animal, the plasma volume was then
determined using the Indicator Dilution Principle. This method utilizes the formula: plasma
volume (PV) = Vd/BW, where BW is body weight and Vd (volume of distribution in liters) is
defined as: [volume of dye injected X concentration of injected dye X 1000]/C0. C0 is the
extrapolated concentration of dye in plasma at Time 0.
Data Analysis
The effects of pregnancy and MR blockade on blood pressure and plasma levels of
cortisol, ACTH, aldosterone, angiotensin II, sodium, potassium, hematocrit, and total protein
over time were analyzed using a three-way repeated measures analysis of variance (RM
ANOVA) of the between-subjects effect of pregnancy status and the within-subjects effects of
treatment and time. Separate two-way RM ANOVA’s were then performed, where indicated, to
identify differences within a given treatment or group. The effects of pregnancy and MR
blockade on plasma volume were analyzed using a two-way RM ANOVA of the between-
subjects effect of pregnancy status and the within-subjects effect of treatment. Duncan’s
multiple range test was performed to determine the source(s) of significance for each two-way
RM ANOVA. Plasma progesterone was compared between pregnant and nonpregnant ewes
using the Student’s t-test. Data are expressed as mean ± standard error of the mean (SEM). A
value of p < 0.05 was considered significant.
Results
Plasma Hormone Levels
Adrenocorticotropic hormone (ACTH)
As expected, there was an overall effect of pregnancy on the plasma ACTH concentration
(Figure 3-1). Three way RM ANOVA indicated that ACTH concentrations in the pregnant ewes
were significantly greater than in the nonpregnant ewes during (p<0.05). The mean plasma

47
ACTH collapsed over the four hours of vehicle infusion was 55 ± 2 pg/ml in the nonpregnant
ewes and 76 ± 5 in the pregnant ewes. Three-way RM ANOVA revealed that the MR antagonist
significantly increased plasma ACTH concentrations, but the effect of treatment varied by group
(main effect of treatment, interaction of treatment and group, p<0.05). Two-way RM ANOVA
performed within nonpregnant ewes followed by Duncan’s Multiple range test suggested that
canrenoate infusion resulted in transiently increased plasma ACTH at the 1 hour time-point in
the nonpregnant ewes compared to 1 hour levels during saline infusion, however this was not
significant. Two-way RM ANOVA performed within pregnant ewes revealed a significant
effect of treatment and Duncan’s Multiple Range test demonstrated a significant increase at 2
and 4 hours of canrenoate infusion compared to the corresponding time points during saline
infusion.
Cortisol
Three-way RM ANOVA on plasma cortisol concentrations in these ewes, indicated
significant main effects of treatment and time and significant time by group, treatment by time,
and treatment by time by group interactions. Separate two-way RM ANOVA followed by
Duncan’s multiple range test performed on nonpregnant and pregnant ewes revealed that in
nonpregnant ewes, cortisol exhibited an initial, transient rise at 1 hour and then declined to levels
that were not different from levels measured at the corresponding time point during infusion of
saline (Figure 3-2). In contrast, during infusion of canrenoate in pregnant ewes, there was still
no increase in plasma cortisol by 1 hour, but cortisol was significantly increased compared to
saline by 2 hours of canrenoate infusion and continued to be elevated over the next two time-
points.

48
Aldosterone
The plasma aldosterone response to canrenoate infusion over time varied between pregnant
and nonpregnant ewes (Figure 3-3). Aldosterone levels in nonpregnant ewes either prior to
infusion of canrenoate or throughout the vehicle infusion were either at or below the lower limit
of detection for this assay (12.5 pg/ml). Three-way RM ANOVA demonstrated that there were
main effects of time, treatment, and pregnancy status on plasma levels of aldosterone (p<0.05).
There were also significant two- and three-way interactions (treatment by pregnancy status, time
by treatment, pregnancy status by time, and time by treatment by pregnancy status; p<0.05).
Upon separate two-way RM ANOVA followed by Duncan’s multiple range test within each
group, the effect of canrenoate infusion in nonpregnant ewes was determined to be significantly
increased by 3 hours; whereas canrenoate effects in pregnant ewes are significantly increased by
2 hours. Additionally, a separate analysis within the canrenoate-treated ewes revealed that the
aldosterone response to MR blockade was significantly greater for pregnant ewes than for
nonpregnant ewes from the 2 hour time point onward.
Angiotensin II
Three-way RM ANOVA revealed significant main effects of time and pregnancy status on
plasma angiotensin concentration (Figure 3-4). Overall, angiotensin II was significantly greater
in pregnant compared to nonpregnant ewes. Additionally, plasma angiotensin concentration
increases over time (three-way RM ANOVA: main effect of time, p<0.05).
Mean Arterial Blood Pressure
As expected, mean arterial blood pressure (MAP) was significantly lower in the pregnant
ewes during infusion of either saline or MR antagonist (Figure 3-5, p<0.05). The MAP values
collapsed over time for the vehicle-infused nonpregnant and pregnant ewes were 116 ± 4 and
102 ± 3 mmHg, respectively. Based on three-way RM ANOVA, the decrease in MAP in

49
response to canrenoate infusion was not significant for either group, however, there were time by
group as well as treatment by group by time interactions (p<0.05).
Hematocrit (%) and Plasma Solute (Total Protein, Potassium, and Sodium) Concentrations
There was a significant effect of infusion of either vehicle or canrenoate on hematocrit as it
significantly decreased in both groups over time (Figure 3-6A, main effect of time based on
three-way RM ANOVA, p<0.05). This effect was attenuated by infusion with canrenoate
(interaction of time and treatment, p<0.05). Plasma protein concentration was significantly
lower overall in the pregnant ewes (Figure 3-6B, main effect of group based on three-way RM
ANOVA, p<0.05). There was no effect of canrenoate on plasma protein concentrations in either
the pregnant or the nonpregnant ewes. There were no overall differences in plasma potassium or
sodium levels between pregnant and nonpregnant ewes. Plasma potassium (K+) increased in
both groups during canrenoate infusion (Figure 3-6C, three-way RM ANOVA: main effects of
time and treatment, interaction of time and group, p<0.05). Two-way RM ANOVA followed by
Duncan’s Multiple Range test performed on each group separately revealed that elevations in
potassium concentration due to canrenoate infusion were significantly greater than vehicle by 2
hours in the pregnant ewes versus 3 hours in the nonpregnant ewes. There was no effect of
canrenoate infusion on plasma sodium (Na+) concentration in ewes in either group (Figure 3-
6D).
When backward stepwise linear regression was performed to assess the role of angiotensin
II and K+ in driving plasma aldosterone during infusion of canrenoate, both were found to be
significant (p=0.009 and P<0.001, respectively). However, when the relationship between
plasma angiotensin II and plasma aldosterone was determined for nonpregnant and pregnant
ewes separately, no significant correlation was found in either group (NP: r=0.147, p=0.437; P:
r=0.305, p=0.101). However, when the relationship between plasma K+ and plasma aldosterone

50
was compared between pregnant and nonpregnant ewes, there appeared to be a difference in this
relationship between the two groups (Figure 3-7). Two-way ANOVA of the between-subjects
effects of pregnancy status and 0.3 mEq/L bins of plasma K+ on plasma aldosterone
concentration during canrenoate infusion revealed a significant effect of pregnancy status on the
aldosterone values at plasma K+ of greater than 4.8 mEq/L (main effects of group and binned K+
concentration as well as a two-way interaction, p<0.05).
Plasma Volume
Plasma volume tended to be higher in the pregnant ewes (44.3 ± 2.3 ml/kg in P ewes
versus 34.6 ±2.3 ml/kg in the NP ewes, p=0.062). Although the differences in plasma volume
measured at the end of canrenoate infusion did not reach significance, there appeared to be a
trend for MR blockade to decrease plasma volume in both groups with an apparent tendency
toward greater decreases in the pregnant ewes (Figure 3-8). The reduction in plasma volume
after treatment with the MR antagonist in the pregnant and nonpregnant ewes was 3.7 ml/kg and
1.8 ml/kg, respectively.
Discussion
The nonpregnant responses in our study are consistent with those found in human and rat
studies in which anti-mineralocorticoid challenge using MR antagonists such as spironolactone
or its active metabolite, canrenoate, causes elevated plasma cortisol concentrations. In humans,
as in the nonpregnant ewes in this study, plasma cortisol concentration was increased as soon as
one hour after administration of MR blockade using spironolactone or canrenoate (Deuschle et
al. 1998, Young et al. 1998, Arvat et al. 2001, Kellner et al. 2002, Wellhoener et al. 2004,
(Grottoli et al. 2002). The plasma ACTH concentrations in these ewes follow a similar time
course, having a tendency to be transiently elevated one hour after the start of treatment with the

51
MR antagonist canrenoate. It is possible that after this time point, we may be observing the
negative feedback effect of elevated cortisol at GR, the lower affinity receptor.
Interestingly, the response pattern of the plasma cortisol and plasma ACTH measured
hourly during infusion with the MR antagonist, canrenoate, differed between pregnant and
nonpregnant ewes, which supports our hypothesis that mineralocorticoid receptor regulation of
the hypothalamic-pituitary-adrenal axis differs between the pregnant and nonpregnant states. I
hypothesize that the differential early (0-2 hours) response pattern between pregnant and
nonpregnant ewes is due, at least in part, to differences in relative importance of central and/or
pituitary MR in negative feedback control of the axis because the ACTH response to canrenoate
infusion appears to exhibit a similar temporal pattern to the cortisol response. The hippocampus
is a prime suspect for future investigations of the mechanism(s) for this differential response.
Based on previous evidence, hippocampal and/or septal MR are thought to be the mediators of
HPA axis responses to canrenoate infusion as they are most densely expressed in the dentate
gyrus and cornu ammonis of the hippocampus as well as the septum and thought to confer the
inhibitory function of the hippocampus on HPA axis regulation (Reul et al. 2000). Further
evidence of the importance of hippocampal and septal MR in regulation of the HPA axis is
provided by the fact that MR blockade-induced HPA axis activation by canrenoate can be
blocked by the GABAA receptor agonist alprazolam in humans (Grottoli et al. 2002).
It is also known that throughout gestation in humans and sheep, plasma progesterone is
much higher than it would be in the nonpregnant state (Rosenthal et al. 1969, Johansson &
Jonasson 1971, Bell et al. 1991). Our current study supports this pattern since the mean plasma
progesterone levels for nonpregnant and pregnant ewes were, 5.3 ± 1.6 and 25.4 ± 2.6 pg/ml,
respectively (p<0.001). Previous reports from this lab showed that progesterone is a

52
physiological ligand for MR and that prior exposure to progesterone reduces the Kd of MR for
cortisol, suggesting that progesterone’s effects are antagonistic (Richards et al. 2003). Studies in
other labs have also shown that progesterone shows a high affinity for MR with only weak
transactivation activity (Rafestin-Oblin et al. 1992, Carey & de Kloet 1994, Funder & Myles
1996, Turner 1997). This suggests progesterone effectively antagonizes binding and subsequent
activation by cortisol, but may also serve as a weak agonist when cortisol levels are relatively
low (Keller-Wood & Wood 2008). It has therefore been theorized that the increase in
hippocampal cytosolic MR availability in pregnancy in the ewe may be due to binding by
progesterone which is thought to prevent transactivation of the receptor and localization of the
receptor to the nucleus as would occur with cortisol or aldosterone as ligands (Roesch & Keller-
Wood 1999). In fact, unpublished data from our lab supports this theory as hippocampal
cytosolic MR availability was also increased in nonpregnant ewes treated with progesterone. In
humans, prior MR antagonism in arginine vasopressin- (AVP-), corticotropin-releasing
hormone- (CRH-), and exercise-induced HPA activation exacerbated the elevation in plasma
cortisol, suggesting that prior blockade of MR activity can alter the set point for HPA axis
activation (Heuser et al. 2000, Arvat et al. 2001, Wellhoener et al. 2004). I theorize that the
differential response to MR blockade in pregnancy reflects the presence of the endogenous
antagonist, progesterone, specifically at MR in the hippocampus. This could explain the absence
of the early stimulatory effect in the pregnant ewes during canrenoate infusion. I hypothesize
that because more MR are available in the nonpregnant ewes due to relatively low progesterone
levels, MR blockade with canrenoate allows for greater increases in HPA axis activity in these
ewes. Whereas in the pregnant animals, the presence of high plasma progesterone could allow
endogenously-derived HPA axis disinhibition and therefore addition of an MR antagonist has

53
relatively no direct effect on HPA axis activity. In this way, it is therefore possible that
progesterone is at least in part responsible for an alteration in HPA axis negative feedback set
point, much like the pre-treatment with MR blockade did in the human studies.
The differential late (2-4 hours) response pattern for ACTH and cortisol between pregnant
and nonpregnant ewes likely reflects differences in relative importance of MR and the HPA axis
in regulation of blood pressure and volume that must occur with pregnancy in order to support
the health of the mother and fetus. From Figure 3-5, although not significant, there appears to be
a trend for mean arterial pressure to be reduced by canrenoate infusion for both groups.
Additionally, there were tendencies for differential patterns of responses in plasma volume,
protein, potassium, and hematocrit with either with vehicle infusion or with MR blockade.
These findings may not have reached significance simply due to the need for more animals in
each group; or because the dose we used caused changes in these endpoints that were subtle
enough to be overcome by other mechanisms. I propose that the increases in cortisol over time
with canrenoate infusion in the pregnant ewes may be an indirect response to decreases (although
not significant by our methods of detection) in blood pressure and plasma volume. Additionally,
the enhanced plasma aldosterone response to anti-mineralocorticoid challenge in pregnant ewes
further supports a role for corticosteroids in the hormonal regulation of volume expansion in the
pregnant state. Angiotensin II concentrations in the pregnant ewes were greater than those
detected in the nonpregnant ewes, but this was not influenced by canrenoate infusion.
Interestingly, linear regression analyses of either plasma potassium or angiotensin II versus
plasma aldosterone concentrations with canrenoate infusion suggested that although both factors
are significant determinants of plasma aldosterone, that a difference in responsiveness to plasma
potassium was likely to be responsible for the greater plasma aldosterone concentrations between

54
the pregnant ewes during canrenoate infusion. The increase in plasma potassium was higher in
pregnant ewes compared to nonpregnant ewes at the 3 hr time point based on two-way RM
ANOVA followed by Duncan’s Multiple Range test on canrenoate-treated ewes. This suggests
that the relative importance of MR in regulation of plasma potassium might be shifted during
pregnancy. The tendencies toward differences in these hemodynamic endpoints are however
consistent with the proposed theory that pregnancy is perceived by homeostatic mechanisms
which regulate plasma volume as an “underfilled” state (for review, see: Schrier & Durr 1987).
Likely, the greater increases in plasma ACTH, cortisol and aldosterone that we observed in the
pregnant ewes during the second half of the canrenoate infusion, reflect activation of such
mechanisms necessary to combat any decreases in plasma volume and MAP that might have
otherwise occurred in response to blockade of MR. These results also support previous findings
in our lab that the elevated basal corticosteroid concentration of pregnancy contributes to
maternal volume expansion and further suggests that these effects may be mediated at least in
part by the mineralocorticoid receptor, presumably at the kidney. However, direct action of
mineralocorticoids in the brain and the resulting cardiovascular effects have also been reported
(Chen et al. 1989, Gomez-Sanchez 1997). Small amounts of aldosterone delivered
intracerebroventricularly in rats produce a significant increase in arterial blood pressure; while
chronic icv infusion of an MR antagonist inhibited mineralocorticoid-induced hypertension in
rats (Chen et al. 1989, Gomez-Sanchez et al. 1990, Janiak et al. 1990).
Overall, these results support my hypothesis that plasma ACTH and cortisol would be
elevated in response to intravenous MR antagonism and that direct HPA axis responses would be
blunted in pregnant ewes relative to nonpregnant ewes. The corresponding differential response
pattern for both ACTH and cortisol release suggests differences in relative importance of central

55
and/or pituitary MR in negative feedback control of the axis. The differential HPA axis
responses during the second half of the infusion are likely reflective of the increase in relative
importance of corticosteroids in homeostatic mechanisms for maintenance of hemodynamic
endpoints such as maternal plasma volume and blood pressure.
This research was supported by an R01 grant from the NIH (DK38114) to Maureen Keller-
Wood. Some of this data was presented at the 89th Annual Meeting of the Endocrine Society,
2007.

56
Figure 3-1. Plasma ACTH concentrations during canrenoate or vehicle infusion. Three-way RM
ANOVA indicated main effects of treatment and group as well as an interaction between treatment and group (p<0.05). * Significantly different from values obtained in the same group during saline infusion at the corresponding time point, based on separate two-way RM ANOVA followed by Duncan’s multiple range test for each group (p<0.05). Data are represented as means ± SEM.

57
Figure 3-2. Plasma cortisol concentrations during canrenoate or vehicle infusion. Three-way
RM ANOVA on plasma cortisol concentrations indicated significant main effects of treatment and time and significant time by group, treatment by time, and treatment by time by group interactions. * Significantly different from values obtained in the same group during saline infusion at the corresponding time point, based on separate two-way RM ANOVA followed by Duncan’s multiple range test for each group (p<0.05). Data are represented as means for plasma cortisol concentration ± SEM.

58
Figure 3-3. Plasma aldosterone (pg/ml) concentrations during iv administration of saline (VEH)
or canrenoate (CAN) in (A) nonpregnant and (B) pregnant ewes. Overall, pregnant aldosterone levels were significantly greater than nonpregnant levels. * Indicates levels significantly different from those obtained from the same group during saline infusion at corresponding time point. ** Indicates significantly different from nonpregnant response to canrenoate infusion at corresponding time point. Data are represented as means ± SEM and considered significant at p<0.05. The dotted line indicates the lower limit of detection for the aldosterone assay at 12.5 pg/ml.

59
Figure 3-4. Plasma angiotensin II (pg/ml) during iv administration of saline (VEH) or
canrenoate (CAN). Three-way RM ANOVA revealed that angiotensin II concentrations were greater in pregnant ewes compared to nonpregnant ewes and overall, there was a significant increase over time (*p<0.05). There was no significant effect of canrenoate treatment in either group. Data are represented as means ± SEM.

60
Figure 3-5. Mean arterial pressure during canrenoate or vehicle infusion in (A) nonpregnant and
(B) pregnant ewes. Overall, MAP in pregnant ewes was significantly lower than in nonpregnant ewes (p<0.05). Values are means ± SEM of 5-minute bins of MAP data.

61
Figure 3-6. Hematocrit and plasma solute concentrations. The effect of intravenous vehicle
(VEH) infusion on percent hematocrit (A) was attenuated by canrenoate (CAN) infusion in both groups. Plasma protein (B) was significantly lower in pregnant (P) ewes with no effect of canrenoate in either group. Canrenoate infusion increased plasma potassium (C) in both nonpregnant (NP) and pregnant ewes. There were no significant differences in plasma sodium (D) in response to canrenoate infusion or due to pregnancy. Values are means ± SEM and considered significant at p<0.05.

62
Figure 3-7. Linear regression of plasma potassium and plasma aldosterone during canrenoate
infusion. The adrenal responsiveness to plasma potassium concentration in nonpregnant (NP, open circles) and pregnant (P, filled circles) ewes during canrenoate infusion appears to differ (slope of 36.4 ± 7.8 in NP compared to 140.1 ± 25.4 in P). For both groups, plasma aldosterone was significantly and positively correlated with plasma potassium concentration (NP: r=0.660, p<0.001; P: r=0.722, p<0.001).

63
Figure 3-8. Plasma volume measured at the end of 4 hr infusion of canrenoate (CAN) or vehicle
(VEH). Plasma volume tended to be higher in pregnant ewes (P: 44.3 ± 2.3 ml/kg versus NP: 34.6 ±2.3 kg/ml, three-way RM ANOVA, p=0.062). There is also an apparent trend for MR blockade to decrease plasma volume in both groups, however this was not significant. Values are means ± SEM.

64
CHAPTER 4 RELATIVE SEROTONERGIC ACTIVITY/RESPONSIVITY DURING PREGNANCY
Introduction
Regulation of the HPA axis is indeed multifaceted and therefore it is not likely that
alterations in any one control mechanism are solely responsible for its increased activity during
pregnancy. I will now switch the attention to a system that has been shown to be associated with
the HPA axis based on physical proximity, pharmacological characterization and in disease
states, the serotonergic system.
The serotonergic system is thought to be one of the upstream stimulatory inputs to the
HPA axis (Calogero et al. 1990, Fuller 1996). Serotonergic cell bodies are found in discrete
clusters or groups of cells along the midline of the brain stem while their axons innervate nearly
every area of the central nervous system. Serotonin, or 5-hydroxytryptamine (5-HT),
synthesized from tryptophan within the cytosol of these neurons, serves as a neurotransmitter
involved in the regulation of mood, feeding behavior, circadian rhythmicity, learning and
memory (Jacobs & Azmitia 1992, Jacobs & Fornal 1999, Cooper et al. 2003). Dysfunction of
the serotonergic system is thought to be a key factor in the pathogenesis of many types of
depression in humans, many of which are also thought to be associated with a dysregulation of
the HPA axis.
Histologically, it has been demonstrated that serotonergic axons synapse on CRH-
containing neurons in the rat PVN and more recently, several 5-HT receptor subtypes have been
shown to moderately populate key areas for HPA axis regulation (Liposits et al. 1987, Petrov et
al. 1994, Wright et al. 1995, Li et al. 1997). Calogero et al (1989) demonstrated in vitro that
serotonin directly stimulates CRH release in hypothalamic cultures. Tsagarakis et al (1989) also
demonstrated this effect in vivo in rats using hypothalamic implants of serotonin. In fact, an

65
extensive review by Carrasco and Van de Kar (2003) discusses numerous studies in rodents that
provide evidence that serotonergic agents have a stimulatory effect on the HPA axis hormones.
In humans and animal models including sheep, intravenous injection or oral administration of
serotonin, its precursors, or 5-HT receptor agonists significantly increase plasma ACTH and
cortisol (Kile & Turner 1985, Calogero et al. 1990, Fuller 1996, Broadbear et al. 2004,
Broadbear et al. 2005, Heisler et al. 2007). Cessation of the serotonergic signal is achieved
primarily (80%) by the removal of serotonin from the synaptic cleft through an active membrane
transporter encoded for by the serotonin transporter (SERT) gene. It is therefore not surprising
that selective serotonin reuptake inhibitors (SSRI’s), which act on this transporter, are associated
with a rise in both portal CRH and plasma ACTH (Bevan & Scanlon 1998). Recently, another
group utilized 5-HT receptor subtype agonists along with anti-CRH antiserum to propose that
HPA axis activation likely occurs via 5-HT1A, 5-HT2A, 5-HT2C and/or 5-HT1B receptors and that
the response is mediated by CRH in rats (Jorgensen et al. 2002).
The goal of this study was to determine whether there are alterations in the serotonergic
component of basal HPA axis regulation during pregnancy that may at least in part explain the
elevation in plasma ACTH and cortisol that must occur to support the health of the mother and
developing fetus. We hypothesized that pregnant ewes would demonstrate an increase in
serotonergic tone or serotonergic responsivity by exhibiting a more robust HPA axis response to
fluoxetine. There were two parts to this objective: (1) to compare the response to an acute
intracerebroventricular (icv) injection of a selective serotonin reuptake inhibitor, fluoxetine
(FLX) in pregnant ewes to the response in the same ewes postpartum; and (2) to compare the
response to subchronic icv administration of a sub-maximal (and more clinically relevant) dose
of fluoxetine in pregnant ewes to nonpregnant ewes. The selective serotonin reuptake inhibitor,

66
fluoxetine has little affinity for muscarinic, histaminic, serotonergic, or noradrenergic receptors
and is selective for serotonin reuptake without affecting norepinephrine reuptake (Stark et al.
1985). Use of fluoxetine will serve to exploit the inherent serotonergic activity in these ewes by
keeping the neurotransmitter in the synaptic cleft longer in order for it to exert its effects as well
as provide information about relative responsivity of the HPA axis to serotonin between pregnant
and nonpregnant or postpartum ewes. If our findings support our hypothesis, this alteration in
serotonergic tone during pregnancy could explain, at least in part, the increase in basal HPA axis
activity that is necessary for maternal and fetal health.
Materials and Methods
Animals
Animals were housed in climate controlled, individual pens located in the University of
Florida Animal Care Facility; all animal use was in accordance with the rules and regulations of
the Institutional Animal Care and Use Committee (IACUC) at the University of Florida.
For Study I - HPA axis responses to acute, icv fluoxetine:
Pregnant (n=6, between 134 and 137 days of gestation; term is approximately 147 days)
mixed, western breed ewes were used for this study. Animals were then allowed to deliver and
were studied again in the postpartum state (8 ± 3 days postpartum). Lambs were removed
promptly after parturition in order to avoid any interference from changes in hormones
associated with lactation.
For Study II - HPA axis responses to “subchronic” icv fluoxetine:
Pregnant (P, between 117 and 126 days of gestation) and nonpregnant (NP) mixed western
breed ewes were used for this study. Prior to surgery, animals were randomly assigned to
receive either fluoxetine (FLX), a selective serotonin reuptake inhibitor (NP FLX: n=6, P FLX:
n=7) or vehicle (NP VEH: n=5, P VEH: n=5). The duration of infusion utilized in this study is

67
referred to as ‘subchronic’ as the fluoxetine is infused for only 6 consecutive days. This duration
is shorter than that which produces changes in MR, GR, or CRH expression observed with more
chronic dosing (at least 2-4 weeks) as in therapeutic treatment for clinical depression (Brady et
al. 1992, Seckl & Fink 1992, Lai et al. 2003).
Surgical Protocol
Before surgery, food was withheld from the ewe for 24 hours. All surgeries were
performed in the surgery suite of the Health Science Center Animal Resources Department.
Animals were prepared for and underwent surgery under aseptic conditions. Animals were
induced with isoflurane and maintained using isoflurane or halothane inhalant (1-3% in oxygen).
For infusion of drug or vehicle, a sterile polyvinyl catheter (ID: 0.030 inch at icv tip
affixed to ID: 0.040 inch catheter prior to sterilization, Tygon® Microbore Tubing, Saint-Gobain
Performance Plastics Corp., Akron, Ohio) was placed into the lateral ventricle of the ewe.
Briefly, a small hole was placed in the skull using a sterilized portable rotary Dremel® (Robert
Bosch Tool Corp, Racine, Wisconsin) approximately 3 mm to the right of bregma, and a 21g
needle was lowered until cerebrospinal fluid flowed into the hub. The needle was then replaced
with the catheter, the hole in the skull around the catheter was filled with bone wax (CP Medical,
Portland, Oregon) and the catheter was secured to the skull using VetbondTM Tissue Adhesive
(3M™, St. Paul, Minnesota).
For Study I:
The free-end of the icv catheter was externalized, sutured in place and plugged using a
sterile 16 g brass nail. The external portion of the catheter was protected under Vetrap™ (3M™,
St. Paul, Minnesota) until icv access was needed on the day of the experiment. Additionally, at
the time of experiment, a jugular venous catheter was placed for blood sampling for hormone
levels.

68
For Study II:
The free-end (ID: 0.040 in) of the icv catheter was affixed to a subcutaneous Alzet®
osmotic pump (model 2ML2; 5 µl/h; Durect Corporation, Cupertino, California) which was filled
at the time of surgery with either vehicle (50:50, dimethyl sulfoxide (DMSO): saline or H2O) or
fluoxetine (5 mg/ml in vehicle) and positioned in a subcutaneous pocket created near the base of
the skull. The Alzet® pump is designed to pump at a rate of 120 ul/day (or 0.6 mg FLX/24 hrs).
The 0.040 inch portion of the catheter in this study was 28 cm in length in order to provide two
days of ‘vehicle dead-space’ before the contents from the Alzet® pump would begin reaching the
lateral ventricle. Sterile bilateral femoral arterial and venous polyvinyl catheters (ID: 0.050 in,
Tygon® Microbore Tubing, Saint-Gobain Performance Plastics Corp., Akron, Ohio) were also
placed as previously described for blood sampling and direct mean arterial pressure recording
(Bell et al. 1991). A trocar device was then used to direct the catheters to an exit site on left side
of the ewe.
In both studies, during the five days immediately following surgery, the animals received
twice daily intramuscular injections of ampicillin (1g); body temperature was also monitored
twice daily. Post-operative care also consisted of daily exit site cleansing with a povodine iodine
solution during the recovery period and for the duration of the study.
Experimental Protocol
Access to arterial and venous catheters was achieved using a method previously
established by this lab (Bell et al. 1991). Briefly, in order to minimize the stress of human
contact to properly measure basal HPA axis activity, the femoral catheters are externalized from
the pens via a swiveling duct system prior to each experiment, followed by an acclimation period
of about 1 hour. Arterial catheters were filled with heparinized saline at the start of the
experiment and after each sample; and this ‘dead-space’ was removed just prior to the

69
subsequent sample. Plasma was acquired by collecting 8 ml of whole blood into tubes
containing 400 µL 0.3M ethylenediaminetetraacetic acid (EDTA) followed by centrifugation at
3000g for 20 min at 4°C. Plasma was then removed and stored at −20oC for future analysis.
For Study I:
After 5 days of post-operative recovery, morning experiments were conducted. For each
experiment, baseline plasma was collected, followed by an injection of either 3.3 mg fluoxetine
or saline (total volume injected = 1 ml) directly into the icv catheter. Blood samples were
collected as described above in 10 minute intervals until 60 minutes post-injection. The
experiment was then repeated at least 2 days later, using the other of the two treatments. The
animals were then allowed to deliver and the experiments were repeated in the postpartum state.
For Study II:
On the morning of the second post-operative day, just before the contents of the Alzet®
pump have begun infusing into the brain (labeled as Day 0), baseline plasma samples were taken
via one of the arterial catheters for all groups. Additionally, separate 1.5 ml whole blood
samples were collected into heparinized syringes and used for determination of sodium and
potassium concentrations using ion specific electrodes (AVL 9180 Electrolyte Analyzer,
AVL/Roche Diagnostics, Roswell, Georgia), as well as for determination of hematocrit and total
protein. Basal mean arterial blood pressure (MAP) was recorded continuously for 40-60 minutes
after the hour long acclimation period via pressure transducers connected to an analog-to-digital
conversion board (LabView, National Instruments, Austin, Texas). Arterial pressure values were
collected at 30 Hz and mean arterial pressure values were analyzed from time periods with
minimal interference from the investigators or from changes in body position relative to the fixed
transducer height. Plasma samples were collected from the arterial catheter at the end of the

70
recording period for replication in the hormone assays. This protocol was repeated every other
day through the 8th post-operative day (or 6 continuous days of drug infusion, labeled as Day 6).
Daily Food Intake
For Study II only, daily food intake (g) was also measured by subtracting the weight of
remaining feed from the amount provided on the previous day in order to assess any differential
effects of fluoxetine and/or pregnancy on feeding habits. In order to avoid any acute effects of
surgery on feeding, food intake was not measured until Day 0 of infusion (or 2 post-operative
days) and was not measured on Day 6 of the infusion due to the time of euthanasia.
Plasma Hormone Determination
All plasma samples were analyzed for ACTH and cortisol concentrations. ACTH levels
for both studies were determined using a radioimmunoassay (RIA) previously described using an
antibody to 1-39 ACTH (Bell et al. 1991). For Study I, cortisol levels were determined by RIA
using the 125I Coat-A-Count® cortisol kit (Siemens Healthcare Diagnostics Inc., Deerfield,
Illinois). For Study II, cortisol was determined using an RIA method previously used in this
laboratory (Wood et al. 1993). Plasma progesterone was determined for both studies by RIA
using the 125I Coat-A-Count® progesterone kit (Siemens Healthcare Diagnostics Inc., Deerfield,
Illinois).
Euthanasia and Tissue Recovery
For Study II, all animals were euthanized with an intravenous injection of a pentobarbital/
phenytoin solution (15-20 ml; Euthasol®, Virbac AH, Fort Worth, Texas) using an indwelling
venous catheter. Ewes were sacrificed in their home pens to minimize stress during this
procedure. Immediately after the ewes were moved to the necropsy room, the carotid arteries
were catheterized for perfusion with ice-cold 10% DMSO/0.9% saline solution in order to
rapidly cool the forebrain and pituitary and slow the enzymatic degradation of mRNA and

71
protein that might occur during removal and dissection of the brain. Using RNase-free
instruments and collection vials, relevant brain structures (hippocampus and hypothalamus) and
other peripheral tissues including pituitary were quickly removed, immediately snap-frozen in
liquid nitrogen and stored at −80oC for future protein and mRNA quantification.
Data Analysis
For Study I, the effects of pregnancy and acute icv fluoxetine administration on plasma
levels of ACTH and cortisol were determined using three-way repeated measures analysis of
variance (RM ANOVA) with repeated measures over drug and time. For Study II, the effects of
pregnancy and subchronic, icv fluoxetine administration on blood pressure and plasma levels of
cortisol, ACTH, sodium, potassium, hematocrit, and total protein over time were analyzed using
a three-way RM ANOVA of the between-subjects effect of group and treatment and the within-
subjects effects of time. Pairwise multiple comparisons were performed using Duncan’s
Multiple Range test to determine the source(s) of significance for each RM ANOVA. Data are
expressed as mean ± standard error of the mean (SEM). A value of P < 0.05 was considered
significant.
Results
Study I: HPA Axis Responses to Acute, Icv Fluoxetine
Plasma ACTH
For both pregnant and postpartum ewes, there were significant increases in plasma ACTH
in response to the icv fluoxetine injection as well as a significant effect of time (main effects of
time and treatment; interaction of treatment and time; Figure 4-1, p<0.05). Three-way RM
ANOVA was unable to detect a significant difference between responses of the ewes during
pregnancy and during the postpartum period. Based on an apparent trend for postpartum
responses to be greater than during pregnancy, two-way RM ANOVA was performed followed

72
by Duncan’s multiple range test on either each group separately or within fluoxetine treated
ewes. These tests suggested differences in the time course of the ACTH response between the
pregnant and postpartum state. Within the pregnant ewes, the plasma ACTH concentration was
significantly elevated from 10-40 minutes after injection compared to saline infusion at the
corresponding time point and the peak ACTH response to icv fluoxetine occurred 20 min post-
injection. On the other hand, in the same ewes studied during the postpartum period, plasma
ACTH was increased at all post-injection time points and the peak plasma ACTH response was
reached 30 min post-injection. Additionally, after two-way RM ANOVA within fluoxetine
treated ewes, the Duncan’s test revealed that within both 30 min and 40 min post-injection, the
postpartum plasma ACTH responses were significantly greater than those produced during
pregnancy. Two-way RM ANOVA within vehicle-treated ewes only showed that there was a
tendency for ACTH concentrations to be greater during pregnancy (p=0.097), although this was
not significant.
Plasma cortisol
Plasma cortisol was statistically greater for these ewes during pregnancy (main effect by
three way RM ANOVA; p<0.05). For both pregnant and postpartum ewes, there was a
significant increase in plasma cortisol in response to the icv fluoxetine injection (main effects of
treatment and time; interaction of treatment and time; Figure 4-2, p<0.05) and cortisol
concentrations at or near maximum adrenal secretion for this species were achieved. There were
no significant differences in the cortisol responses to acute icv fluoxetine between ewes during
pregnancy or during the postpartum period. Based on ACTH concentrations, plasma cortisol
levels were not quantified in samples from 40 and 50 minutes post-injection as the cortisol
response was expected to remain at maximal levels across these time points.

73
Study II: HPA Axis Responses to Subchronic, Icv Fluoxetine
Plasma ACTH
Plasma ACTH was not different between pregnant and nonpregnant ewes, although the
mean plasma ACTH during vehicle infusion was 111 ± 22 pg/ml in the pregnant ewes and 88 ±
10 pg/ml in the nonpregnant ewes. Three-way RM ANOVA revealed an effect of fluoxetine
treatment over time (interaction of treatment and time, p<0.05), and the time course of the
plasma ACTH response to icv infusion of fluoxetine tended to differ between nonpregnant and
pregnant ewes (interaction of time, group, and treatment; Figure 4-3, p= 0.057). Two-way RM
ANOVA followed by Duncan’s Multiple Range tests performed separately on each group
suggested plasma ACTH on Day 2 of the fluoxetine infusion was transiently increased in the
pregnant ewes compared to vehicle-infused pregnant ewes on the same day; whereas,
nonpregnant ewes remained unchanged until Day 6 compared to earlier time points (Days 0, 2,
and 4) for the same ewes or compared to vehicle-treated ewes on the Day 6.
Plasma cortisol
Figure 4-4 illustrates the differential time course of the plasma cortisol response to icv
infusion of fluoxetine between nonpregnant and pregnant ewes (three-way RM ANOVA:
interaction of time, group, and treatment, p<0.05). Subsequent two-way RM ANOVA followed
by Duncan’s Multiple Range test within fluoxetine-treated ewes, revealed that plasma cortisol
concentrations in the pregnant ewes were significantly increased on Day 2 compared to Day 6 of
infusion (p<0.05), whereas plasma cortisol remain unchanged until Day 6 of fluoxetine infusion
in nonpregnant ewes. These results reflect the pattern observed for ACTH release. Plasma
cortisol was not different between pregnant and nonpregnant ewes, although the mean plasma
cortisol levels for vehicle-infused pregnant and nonpregnant ewes were 3.3 ± 0.5 ng/ml and 2.4 ±
0.8 ng/ml, respectively.

74
Mean arterial pressure, hematocrit (%) and plasma solute (total protein, potassium, and sodium) concentrations
Three-way RM ANOVA revealed no significant effects of fluoxetine infusion on mean
arterial pressure (Figure 4-5). There was no main effect of group on mean arterial pressure,
contrary to findings reported in Chapter 3 and throughout the literature. Figure 4-6 illustrates the
hematocrit, plasma protein and electrolyte (sodium and potassium) responses to subchronic icv
infusion of fluoxetine or vehicle. Three-way RM ANOVA revealed that hematocrit (panel A)
was not altered by treatment with fluoxetine, but that in nonpregnant ewes, hematocrit decreased
over time in both vehicle- and fluoxetine-treated ewes (main effect of time, interaction of time
and group, p<0.05). Plasma protein (panel B) concentrations were significantly altered by
fluoxetine over time and were lower in pregnant ewes (main effects of treatment and group,
interaction of time and treatment). There were no significant effects on plasma sodium
concentrations (panel C). Plasma potassium (panel D) concentrations were significantly altered
by fluoxetine, but this effect varied between groups over time (main effect of treatment,
interaction of time, treatment, and group, p<0.05), with a trend for decreasing plasma K+ over
time in the nonpregnant ewes and increasing plasma K+ in the pregnant ewes.
Daily food intake
Figure 4-7 illustrates daily food consumption from the first day of the experiment until one
day prior to euthanasia. Three-way RM ANOVA was unable to detect significant effects of
fluoxetine treatment in either group. Post-hoc Duncan’s Multiple Range test after two-way RM
ANOVA revealed significant increases in food intake in both groups of vehicle-treated ewes on
Days 3 and 5 as compared to Day 0 (two-way RM ANOVA followed by Duncan’s Multiple
Range test performed on vehicle-treated ewes; p<0.05). Additionally, similar analyses within in
each group suggested that in the fluoxetine-treated nonpregnant ewes, food intake was lower on

75
Day 2 and Day 4 relative to Day 0 of the infusion (p<0.05), whereas there were no significant
differences in food intake in the pregnant ewes during fluoxetine infusion.
Discussion
Study I: HPA Axis Responses to Acute, Icv Fluoxetine
The ACTH responses to infusion of acute selective serotonin reuptake inhibitor during
pregnancy and postpartum were not significantly different by three-way RM ANOVA. These
findings disprove our hypothesis that pregnant ewes would have greater ACTH responses to
treatment with a selective serotonin reuptake inhibitor. These results suggest that not only is
there is not an increase in serotonergic responsivity driving the elevation in basal ACTH levels
during pregnancy, but rather our results suggest that serotonergic responsivity of the HPA axis
during pregnancy may in fact be depressed, based on follow-up analysis using two-way RM
ANOVA on fluoxetine responses only.
One possible mechanism for this apparent difference might be an effect of chronically
elevated cortisol on the serotonergic system. It has been reported that manipulation of
corticosteroid levels via adrenalectomy or exogenous administration regulates the abundance of
post-synaptic 5-HT1A mRNA and protein in the hippocampus (Chalmers et al. 1993, Chalmers et
al. 1994, Kuroda et al. 1994, Briones-Aranda et al. 2008). The administration of corticosterone
in rats reverses the increases in 5-HT1A binding that occur following adrenalectomy (Mendelson
& McEwen 1992). Recently, Lee and colleagues (2009) demonstrated 5-HT2A receptors within
the PVN may be desensitized by chronic corticosterone administration. Together these results
suggest a possible mechanism for the attenuated response to acute icv fluoxetine that may occur
during pregnancy compared to the postpartum period in the ewe.
The fact that three-way RM ANOVA was unable to detect an overall difference in the
ACTH response to acute icv fluoxetine administration during pregnancy and postpartum, was

76
surprising given the obvious trend shown in Figure 4-1. However, individual assessment of
ACTH responses in each subject revealed that 2 out of the 6 ewes’ responses during pregnancy
more closely resembled their respective postpartum responses to fluoxetine. As discussed in
Chapter 2, it is possible that progesterone and estrogen which become elevated during pregnancy
are impacting serotonergic responsivity of the HPA axis through their effects on pre- and post-
synaptic receptors, reuptake transporters, and intracellular coupling with other proteins which
mediate serotonin’s downstream effects (Bethea et al. 2002). We measured plasma progesterone
concentrations in these ewes and found that these 2 ewes had plasma progesterone
concentrations of 4.4 and 5.8 pg/ml (in the range of concentrations found in the nonpregnant
ewes in Chapter 3 and in Study II of this chapter), while the average plasma progesterone in the
remaining 4 ewes during pregnancy was 16.3 ± 2.3 pg/ml. Interestingly, linear regression
analysis revealed a significant inverse relationship between plasma progesterone and peak
ACTH responses to icv fluoxetine (r=0.687, p<0.05). This would suggest that serotonin-
stimulated HPA axis activation is blunted in the presence of progesterone levels typically
measured during pregnancy. Additionally, it is known that estrogen, which was not measured in
these ewes but was undoubtedly elevated in pregnancy, has been shown to attenuate 5-HT1A-
stimulated increases in ACTH and corticosterone, and this effect appears to be related to
estrogen’s ability to reduce levels of G-proteins that are known to mediate the actions of this
receptor (Raap et al. 2000).
On the other hand, the fact that there were no differences in plasma cortisol responses to
acute icv fluoxetine injection between these reproductive states was not surprising. Maximal
adrenal secretion of corticosteroid occurs in most species in response to plasma ACTH

77
concentrations well below the values achieved in this study in response to fluoxetine injection
(Keller-Wood et al. 1983).
Study II: HPA Axis Responses to Subchronic, Icv Fluoxetine
Because our previous study produced near maximal secretion of ACTH and likely
maximal cortisol secretion in response to fluoxetine, we investigated central treatment with a
more chronic, but submaximal dose of fluoxetine. In this study, we also found that, contrary to
our hypothesis, there was not a sustained increase in serotonergic responsivity during pregnancy
as exploited by subchronic administration of a lower dose of fluoxetine. Although the time
course for the ACTH response to subchronic fluoxetine infusion had a tendency to differ
between the reproductive groups, more animals are needed to confirm this. However, there was
a significant difference in the time course of the plasma cortisol response to fluoxetine infusion
between the two groups, suggesting that the trends observed in the plasma ACTH responses are
not simply due to random sampling variability. The continuous presence of serotonin within the
synaptic cleft appears to affect the release of ACTH and cortisol in a different manner between
pregnant and nonpregnant ewes. Specifically, in nonpregnant ewes, subchronic, icv fluoxetine
was not effective at stimulating cortisol until the 6th day of continuous infusion, while in
pregnant ewes, stimulation occurred transiently at Day 2. I propose that pregnant ewes might
have more 5-HT2A receptors on parvocellular neurons in the PVN at the start of the study than
the nonpregnant ewes likely due to estrogen (Sumner & Fink 1995, Sumner & Fink 1997,
Osterlund & Hurd 1998), allowing for stimulation of the HPA axis on Day 2 in response to
elevations in the synaptic cleft by fluoxetine. This hypothesis was later refuted, at least at the
gene level, as discussed in Chapter 5 of this dissertation. Prolonged serotonin in the synaptic
cleft may then downregulate these receptors thereby reducing the effect of fluoxetine for the
remainder of the infusion. Meanwhile, the delayed effect observed in nonpregnant ewes, maybe

78
a result of downregulation of the 5-HT1A autoreceptors, although in humans, this is not thought
to occur before 2 weeks of chronic SSRI treatment. Overall, the subchronic, icv fluoxetine
infusion had milder effects on plasma ACTH and cortisol than the acute fluoxetine injection
given in Study I, and therefore this dose of fluoxetine may have been too low to produce
measureable differences between pregnant and nonpregnant ewes.
Unexpectedly, neither plasma cortisol nor ACTH was significantly greater in the pregnant
ewes in this study. I theorize this may be due to the relatively high basal levels on Day 0 in the
vehicle-treated nonpregnant ewes. Plasma cortisol in the vehicle-infused nonpregnant ewes was
higher on Day 0 than all other days, based on two-way RM ANOVA followed by Duncan’s
Multiple Range test within vehicle-treated ewes (p<0.05). Although not significant, there is an
apparent trend for ACTH to exhibit the same pattern in these ewes. Meanwhile, this pattern does
not occur in the vehicle-treated pregnant ewes. This data supports unpublished observations in
our lab that the excitable nature of this species is somewhat subdued during pregnancy in
response to human interaction and experimental procedures compared to nonpregnant ewes.
At the same time, due to unexpected fluctuations in peripheral hemodynamic endpoints in
this study, the HPA axis responses to either vehicle or fluoxetine infusion should be interpreted
with caution. Additionally, in some cases there were detectable differences in these variables
even on Day 0, when fluoxetine from the osmotic pump should not have yet begun to reach the
tip of the catheter. As expected hematocrit and plasma sodium concentrations were not altered
by treatment with fluoxetine. However, in nonpregnant ewes, hematocrit decreased over time in
either treatment group. Again, the elevated hematocrit levels at the beginning of the study are
likely the result of stress from surgery or human interaction. Like the cortisol and ACTH
concentrations, these levels appear to normalize over time. Based on three-way RM ANOVA,

79
plasma protein and plasma potassium were both significantly affected by fluoxetine. However,
two-way RM ANOVA followed by Duncan’s Multiple Range test performed within each group
suggested that the plasma protein effect of fluoxetine is only significant in the pregnant ewes.
As for plasma potassium, a similar analysis was unable to detect significant effects in either
group when analyzed separately, and suggested that fluoxetine only tended to have an effect over
time in the nonpregnant ewes (p=0.059 for interaction of treatment and time). In any case, the
differential ACTH and cortisol responses to subchronic fluoxetine cannot be attributed to
secondary effects from changes in blood pressure, hematocrit, plasma protein, or electrolyte
concentrations.
Throughout the literature, reductions in meal size and eating rate are often observed
following peripheral or central injections of serotonergic agents and these changes are consistent
with alterations in the mechanisms of satiety (Blundell 1986). Interestingly, despite the lack of
significance by three-way RM ANOVA, the food intake data are suggestive of time effects and
differences in response to icv fluoxetine between nonpregnant and pregnant ewes. Based on the
literature and apparent trends of the food intake data in this study, separate two-way RM
ANOVA’s were performed. Post-hoc Duncan’s tests revealed significant increases in food
intake in both groups of vehicle-treated ewes on Days 3 and 5 as compared to Day 0 (two-way
RM ANOVA followed by Duncan’s Multiple Range test performed on vehicle-treated ewes;
p<0.05). This is consistent with unpublished observations in our lab that include a typical
pattern of reduced feeding during the first few post-operative days, with a gradual recovery over
time. Typically, we allow animals to recover from surgery for a period of 5 days which includes
daily intramuscular prophylactic antibiotic injections and rectal temperature assessment before
beginning any experimentation, and by this time, the feeding effects of surgery have diminished.

80
In this study, however, the experimentation began on the second post-operative day, before full
recovery of feeding has occurred. We were therefore able to observe any effects that icv
infusion of fluoxetine might have on this feeding recovery period and whether or not these
effects differ between pregnant and nonpregnant ewes. Additionally, post-hoc Duncan’s
Multiple Range tests suggested that in the fluoxetine-treated nonpregnant ewes, food intake was
lower on Day 2 and Day 4 relative to Day 0 of the infusion (p<0.05), whereas there were no
significant differences in food intake in the pregnant ewes during fluoxetine infusion. The data
suggest that this food intake recovery period for the pregnant ewes may not have been affected
by fluoxetine administration, while significant reductions in food intake in the nonpregnant ewes
are evident by 4 days of fluoxetine infusion. These results suggest an alteration in central
serotonin-mediated satiety pathways during pregnancy. As discussed in Chapter 2, serotonin is
thought to play a role in regulation of food intake mediated, at least in part, through its effects at
the POMC neurons of the arcuate nucleus of the hypothalamus. Future investigations to
characterize the subtle differential responses to fluoxetine between nonpregnant and pregnant
ewes observed in this study should therefore include this pathway.
Summary
The exact mechanism of serotonergic stimulation of the HPA axis is not entirely
understood. These studies may also demonstrate variability in responses as they relate to
duration and dose of fluoxetine administration. The differential responses between nonpregnant
and pregnant ewes may be explained by differences at the gene level, the protein level, or post-
translational differences between pregnant and nonpregnant ewes in terms of serotonin-mediated
HPA axis activation. Additionally, these differences could be occurring at the hypothalamus,
hippocampus or the brain stem, although our lateral ventricular infusion of fluoxetine would be
expected to primarily target forebrain, hypothalamic or hippocampal, sites of serotonin action.

81
Ovarian hormones might be influencing the responses to fluoxetine in these ewes. Our data
suggest that progesterone may play a role in attenuating the CRH or ACTH response to serotonin
stimulation as it was shown to be inversely and significantly related to ACTH responses to
fluoxetine in Study I. Additionally, the presence of serotonergic receptors and reuptake
transporters in the hippocampus as well as its innervation by serotonergic neurons would suggest
that it is a likely site for future investigations to explain potential differences among the
reproductive states. Meanwhile, serotonergic neurons themselves are known to express
corticosteroid receptors and therefore posses the potential to be regulated by circulating
glucocorticoid, directly (Harfstrand et al. 1986, Fuxe et al. 1987, Morimoto et al. 1996).
Despite the fact that the acute findings disprove our hypothesis that pregnancy is a state of
elevated serotonergic tone which in turn would play a role in elevating basal HPA axis activity
during pregnancy, these studies have, however, uncovered potential differences in responsivity to
serotonergic agents between the pregnant and postpartum states in terms of HPA axis activation.
The findings of the subchronic study, on the other hand, were suggestive of increased
serotonergic responsivity in pregnant ewes as an increase in plasma cortisol concentration was
observed sooner in these animals compared to the nonpregnant ewes in response to icv fluoxetine
infusion. Aside from the vast difference in dose and duration of administration, the seemingly
contradictory findings of Study I and Study II might also be related to differences between
postpartum and nonpregnant ewes in terms of either basal HPA axis regulation or serotonin
system activity. However, previously, our lab has reported comparable findings in studies on
pregnant compared to nonpregnant and pregnant compared to postpartum ewes in terms of
alterations in HPA axis feedback. Additionally, the observation that subchronic fluoxetine
infusion had less of a hindering effect on post-operative feeding recovery in the pregnant ewes,

82
may provide evidence that central serotonin-mediated feeding regulation may be altered in
pregnancy. The major implications of these studies are that if differences similar to those
measured in this study are also evident in humans, these findings could suggest a need for
adjustment of treatment regimens for depression, anxiety, and/or HPA axis dysregulation as a
woman transitions between these reproductive states.
This research was supported by an R01 grant from the NIH (DK38114) to Maureen Keller-
Wood.

83
Figure 4-1. Study I: Plasma ACTH following acute icv fluoxetine (FLX) or vehicle (VEH).
Grey boxes indicate time points in which the increase in plasma ACTH following acute icv fluoxetine (3.3 mg) was significantly greater for the postpartum (PP) ewes than it was for the same ewes during pregnancy (P) based on two-way RM ANOVA when given fluoxetine, followed by Duncan’s Multiple Range test. ** Indicates significantly different from ACTH concentration during saline infusion at the corresponding time point for both pregnant and postpartum states. * Indicates significantly different from ACTH concentration during saline infusion at the corresponding time point for ewes in postpartum state only. Values are means ± SEM and considered significant at p<0.05.

84
Figure 4-2. Study I: Plasma cortisol following acute icv fluoxetine (FLX) or vehicle (VEH).
Plasma cortisol was significantly higher during pregnancy; and there was a significant increase in plasma cortisol following acute icv fluoxetine (3.3 mg) injection, both during pregnancy (P) and in the postpartum (PP) period (main effects of group, treatment, and time as well as a time and treatment interaction by three-way RM ANOVA, p<0.05). ** Indicates significant difference from vehicle-infusion for both groups at the corresponding time point for the corresponding group.

85
Figure 4-3. Study II: Plasma ACTH response to subchronic icv infusion of fluoxetine (FLX) or
vehicle (VEH). Three-way RM ANOVA indicated a significant interaction of treatment and time (p<0.05), and the time course of the plasma ACTH response to icv infusion of fluoxetine tended to vary between nonpregnant (NP) and pregnant (P) ewes (interaction of time, group, and treatment, p= 0.057). Two-way RM ANOVA followed by Duncan’s Multiple Range test revealed increased plasma ACTH on Day 2 of the infusion in fluoxetine-treated ewes compared to those given vehicle, whereas nonpregnant ewes remained unchanged until Day 6 of fluoxetine infusion compared to both vehicle-infused ewes and to all previous days of vehicle infusion. * Indicates significantly greater than vehicle-infused ewes of the same group on the corresponding day of infusion.

86
Figure 4-4. Study II: Plasma cortisol response to subchronic icv infusion of fluoxetine (FLX) or
vehicle (VEH). The time course of the plasma cortisol response to subchronic icv fluoxetine varies between nonpregnant (NP) and pregnant (P) ewes (three-way RM ANOVA: interaction of time, group, and treatment, p<0.05). In response to fluoxetine infusion, significant elevation of cortisol is observed on Day 6 for the nonpregnant ewes. * Indicates significantly greater than levels observed in vehicle-treated ewes at the same time point (two-way RM AVOVA followed by Duncan’s Multiple Range test within nonpregnant ewes, interaction of treatment and time, p<0.05).

87
Figure 4-5. Study II: Mean arterial pressure (MAP) during subchronic icv infusion of fluoxetine
(FLX) or vehicle (VEH) in (A) nonpregnant and (B) pregnant ewes. Mean arterial pressure did not significantly change in response to fluoxetine infusion, based on three-way RM ANOVA. Data are expressed as group means ± SEM and considered significant at p<0.05.

88
Figure 4-6. Study II: Hematocrit and plasma solute concentrations during subchronic icv
fluoxetine (FLX) or vehicle (VEH) infusion. Hematocrit (A) decreased over time in nonpregnant ewes and was not altered by treatment with fluoxetine in either group (main effect of time, interaction of time and group, p<0.05). Plasma protein (B) concentrations were significantly altered by fluoxetine over time and were lower in pregnant ewes (main effects of treatment and group, interaction of time and treatment). Plasma sodium (C) concentration was not different between groups or in response to fluoxetine infusion. Plasma potassium (D) concentrations were significantly altered by fluoxetine over time, but this effect varied between groups (main effect of treatment, interaction of time, treatment, and group). Values are group means ± SEM and considered significant at p<0.05 when analyzed by three-way RM ANOVA.

89
Figure 4-7. Study II: Daily food intake during subchronic icv fluoxetine or vehicle infusion.
There was a significant increase in food intake in both groups of vehicle-treated ewes on Days 3 and 5 as compared to Day 0 (* indicates significantly different from same group at Day 0 at p<0.05, based on two-way RM ANOVA followed by Duncan’s Multiple Range test within vehicle-treated ewes). Overall, there was no main effect of time on daily food intake in the fluoxetine treated ewes. However in the fluoxetine-treated nonpregnant ewes there was a significant decrease in food intake at Day 2 and Day 4 relative to that on Day 0, whereas there were no significant differences in food intake in the pregnant ewes during fluoxetine infusion (** indicates significantly different from Day 0 in same group at p<0.05, based on two-way RM ANOVA followed by Duncan’s Multiple Range test within nonpregnant ewes).

90
CHAPTER 5 HYPOTHALAMIC EXPRESSION OF GENES RELATED TO HPA AXIS REGULATION
AND THE SEROTONERGIC SYSTEM IN EWES
Introduction
In our lab, we use pregnant, nonpregnant, and postpartum ewes as our experimental model;
and one of the benefits of an in vivo model is the ability to assess relative gene expression in
intact physiological systems. Because the hypothalamus is the point of integration for all
upstream inputs driving or inhibiting hypothalamic-pituitary-adrenal (HPA) axis activity,
investigating relevant gene expression patterns in this region is a logical starting point.
Therefore, the objective of this study was to characterize relative mRNA expression levels of
HPA axis- and serotonergic system-relevant genes in the hypothalamus between nonpregnant,
pregnant, and post-partum ewes. Any differences found here may at least partially explain
differences in basal HPA axis activity during pregnancy. Additionally, we hope to at shed some
light on the possible mechanisms for the differential HPA axis responses activation between
pregnant and nonpregnant or postpartum ewes in response to MR antagonism or selective
serotonin reuptake transporter inhibition discussed in the preceding chapters of this dissertation.
It is known that HPA axis negative feedback is mediated by corticosteroid binding to the
low affinity, glucocorticoid receptor (GR) and the higher affinity, mineralocorticoid receptor
(MR) (Keller-Wood & Dallman 1984, Reul & de Kloet 1985, Reul et al. 1987, Bradbury et al.
1991, de Kloet et al. 1993). Due in part to the affinity profiles, MR are thought to be the major
steroid receptor involved in regulation of basal HPA axis activity. At the same time, axons of
the serotonergic system have been shown to project to several components of the HPA axis,
while several animal studies and human trials have demonstrated increases in plasma ACTH and
cortisol concentrations in response to administration of serotonin, its precursors, serotonin

91
receptor agonists, and selective serotonin reuptake inhibitors (for review, see: Carrasco & Van de
Kar 2003).
Our lab is interested in understanding the mechanisms for elevated basal plasma ACTH
and cortisol concentrations during pregnancy in the human. The overall hypothesis of the lab is
that ovarian steroid hormones which are elevated in pregnancy, such as progesterone and
estrogen, may play a role in modifying regulatory mechanisms of HPA axis activity. Classically,
steroid hormones such as cortisol, progesterone and estrogen are known to modulate gene
expression as their respective receptors are members of the nuclear receptor family.
We hypothesize that relative mRNA expression might be increased for either or both CRH
and AVP thereby driving basal HPA axis activity at a higher level. On the other hand, a
reduction in negative feedback by basal levels of cortisol in pregnancy might be occurring in a
system with reduced MR expression. With less MR being transcribed, ultimately fewer
receptors might be present which could mean reduced HPA axis inhibition by basal levels of
cortisol as demonstrated previously in our lab (Keller-Wood 1998). The results in Chapter 3
suggest a differential early response to 4 hours of intravenous MR antagonism by canrenoate
between pregnant and nonpregnant ewes, in which the early (<2 hours) response of the pregnant
ewes is blunted. This would also suggest that MR expression might be reduced during
pregnancy. Changes in GR expression patterns are not likely as pregnant ewes have normal
inhibition of ACTH in response to high levels of cortisol (Keller-Wood 1998).
Serotonergic responsivity was originally hypothesized to be elevated during pregnancy as a
possible mechanism for increased basal HPA axis activity; and we expected elevations in ovarian
hormones might decrease hypothalamic 5-HT1A receptor mRNA and increase 5-HT2A. This
hypothesis is based on studies in rats which describe such changes in response to ovarian steroids

92
in other brain regions (Sumner & Fink 1995, Sumner & Fink 1997, Osterlund & Hurd 1998).
Several groups have also suggest that glucocorticoids might tonically inhibit the expression of 5-
HT1A (Mendelson & McEwen 1992, Chalmers et al. 1993, Chalmers et al. 1994, Briones-Aranda
et al. 2008). On the other hand, the results of the acute fluoxetine study (Study I) discussed
earlier suggest that serotonergic responsivity is in fact depressed compared to the postpartum
state. We would therefore expect hypothalamic 5-HT1A might be elevated and/or 5-HT2A
expression levels might be reduced in pregnancy relative to the postpartum state. Based on
results of the subchronic study (Study II), the pregnant ewes might have increased 5-HT2A
relative to nonpregnant ewes. A reduction in SERT expression relative to postpartum ewes
might also explain the reduced response to the selective serotonin reuptake inhibitor in the
pregnant ewes in the acute study. The differential food intake effects of subchronic fluoxetine
administration between pregnant and nonpregnant ewes suggest that hypothalamic POMC
expression might be lower during pregnancy allowing for reduced inhibition by serotonergic
agents.
Materials and Methods
Euthanasia and Tissue Recovery
All animals used in these studies were euthanized with an intravenous injection of a
pentobarbital/ phenytoin solution (15-20 ml; Euthasol®, Virbac AH, Fort Worth, Texas) using an
indwelling venous catheter. Ewes were sacrificed in their home pens to minimize stress during
this procedure. Immediately after the ewes were moved to the necropsy room, the carotid
arteries were catheterized for perfusion with ice-cold 10% DMSO/0.9% saline solution. This
procedure cools the forebrain and pituitary and slows the enzymatic degradation of mRNA and
protein that might have otherwise occurred during removal and dissection of the brain. Relevant
brain structures (hippocampus and hypothalamus) and other tissues including pituitary were

93
removed quickly using baked, RNase-free instruments, placed into RNase-free containers,
immediately snap-frozen in liquid nitrogen and stored at −80oC for future protein and mRNA
quantification.
At the time of tissue collection, the hypothalamus was divided at the midline into right and
left halves and each piece was stored separately in the −80oC freezer. The relevant nuclei to HPA
axis activation (PVN) are present in each half as they are bilateral and found adjacent to the third
ventricle, which forms part of midline division of the hypothalamus. The following data
describe mRNA expression levels from one half of the hypothalamus, while the other half has
been saved for extraction and quantification of proteins.
RNA Extraction and Quantification
RNA was extracted from the hypothalami of nonpregnant, pregnant (between 130 and 146
days of gestation, term is approximately 147 days), and postpartum (4 ± 1 days postpartum)
ewes. Tissues were first homogenized in Trizol® (GIBCO/BRL, Grand Island, New York)
according to the manufacturer’s directions. Genomic DNA was removed using RNeasy Plus
Mini Kits (Qiagen Inc., Valencia, California). The absorbance of a diluted aliquot of each
sample was then measured at 260 and 280 nm for determination of RNA concentration and
purity using a spectrophotometer.
Reverse Transcription and Real-Time Quantitative PCR
Reverse transcription in a thermocycler for 10 minutes at 25 °C followed by 120 minutes at
37 °C was performed using a high capacity cDNA archive kit (Applied Biosystems; Foster City,
California), followed by storage at −20oC. Quantitative polymerase chain reaction (qPCR) was
then performed and analyzed using the Taqman® Universal PCR Master Mix and the ABI Prism
7000 Sequence Detection System according to the manufacturers’ instructions (Applied
Biosystems, Foster City, California). Each PCR reaction contains a total volume of 25 ul which

94
includes 2.5 μl of cDNA product from the reverse transcription step as the template and 12.5 ul
Taqman® Master Mix. Reactions were performed under the following conditions: initial
incubation at 95°C for 10 min, followed by 40 cycles of 95°C denaturation for 15 sec and 60°C
annealing for 1 min. The relative mRNA expression levels in the hypothalamus of the following
genes were investigated in this study: mineralocorticoid receptor (MR), glucocorticoid receptor
(GR), corticotropin-releasing hormone (CRH), arginine vasopressin (AVP), serotonin receptor
1A (5-HT1A), serotonin receptor 2A (5-HT2A), serotonin reuptake transporter (SERT), and
proopiomelanocortin (POMC). The template amount for each gene was 20 ng of cDNA, except
for MR which utilized 100 ng of cDNA in each reaction. The probe and primer sequences along
with their concentrations are listed in Table 5-1. All probe/primer sequences and concentrations
have been previously optimized and validated for use in quantitative PCR via prior experiments
in this lab. Each sample was analyzed in triplicate and all samples for each gene were analyzed
on the same 96-well plate. β-actin was also run for each sample as the housekeeping gene.
Data Analysis
One-way analysis of variance (ANOVA) was performed using the relative cycle threshold
(ΔCt) method (Livak & Schmittgen 2001). Briefly, the threshold concentration (Ct) was
determined from a log–linear plot of the PCR product signal versus the cycle number. Then ΔCt
was calculated as the mean Ct for a set of triplicates in a given sample for the housekeeping gene
(β-actin) minus the mean Ct for a set of triplicates in the same animal for the gene of interest.
One-way ANOVA was then performed using these values with p<0.05 as the criterion for
significance. Post-hoc analysis was performed using Duncan’s multiple range test. On data sets
that were not normally distributed and/or had unequal variance, Kruskal-Wallis one-way
ANOVA on ranks was performed. Data in figures are expressed as group means of fold change
relative to expression in the nonpregnant group (NP) ± standard error of the mean. Relative gene

95
expression or fold change for each gene of interest was calculated as 2(− ΔΔCt); where ΔΔCt was
calculated as ΔCt for a given animal minus the mean ΔCt for the nonpregnant group.
For most of the genes including β-actin, there were 7 nonpregnant, 10 pregnant, and 12
post-partum ewes included in the analysis. For 5-HT2A, an initial run using fewer animals (NP,
n=4; P, n=10; PP, n=10) indicated that differences among these groups were unlikely and was
therefore not run on the additional five samples. For both POMC and AVP, two of the
nonpregnant ewes demonstrated late amplification relative to the rest of this group (shifts of 4
and 7 cycles to the right for POMC and approximately 9 cycles for AVP) and were therefore left
out of the analysis. We hypothesize that due to the location of the nuclei expressing POMC and
AVP, that these areas may have been inadvertently excluded during dissection.
Results
Glucocorticoid Receptor (GR)
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of glucocorticoid receptor mRNA (p=0.825). The average
ΔCts for each group were 11.6 ± 0.20, 11.50 ± 0.28, and 11.39 ± 0.11, respectively. Relative
expression compared to nonpregnant ewes is depicted in Figure 5-1.
Mineralocorticoid Receptor (MR)
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of mineralocorticoid receptor mRNA (p=0.722). The
average ΔCts for each group were 5.29 ± 0.35, 5.77 ± 0.13, and 5.67 ± 0.12, respectively.
Relative expression compared to nonpregnant ewes is depicted in Figure 5-2.
Corticotropin-Releasing Hormone (CRH)
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of corticotropin-releasing hormone mRNA (p=0.782).

96
The average ΔCts for each group were 10.73 ± 0.54, 10.34 ± 0.47, and 10.38 ± 0.22,
respectively. Relative expression compared to nonpregnant ewes is depicted in Figure 5-3.
Arginine Vasopressin (AVP)
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of arginine vasopressin mRNA (p=0.209). The average
ΔCts for each group were -1.18 ± 0.53, -0.36 ± 0.34, and 0.27 ± 0.54, respectively. Relative
expression compared to nonpregnant ewes is depicted in Figure 5-4.
5-HT1A Receptor
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of 5-HT1A receptor mRNA (p=0.449). The average ΔCts
for each group were 7.41 ± 0.18, 7.70 ± 0.16, and 7.76 ± 0.20, respectively. Relative expression
compared to nonpregnant ewes is depicted in Figure 5-5.
5-HT2A Receptor
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of 5-HT2A receptor mRNA (p=0.471). The average ΔCts
for each group were 9.06 ± 0.30, 8.61 ± 0.20, and 8.77 ± 0.18, respectively. Relative expression
compared to nonpregnant ewes is depicted in Figure 5-6.
Serotonin Reuptake Transporter (SERT)
There were no significant differences in ΔCt between nonpregnant, pregnant, and
postpartum ewes for expression levels of serotonin reuptake transporter mRNA (p=0.832). The
average ΔCts for each group were 11.83 ± 0.30, 11.74 ± 0.16, and 11.71 ± 0.09, respectively.
Relative expression compared to nonpregnant ewes is depicted in Figure 5-7.

97
Proopiomelanocortin (POMC)
Expression of POMC mRNA was significantly lower in pregnant and postpartum ewes
compared to nonpregnant ewes as indicated by comparison of ΔCt across groups using one-way
ANOVA followed by Duncan’s Multiple Range test (p=0.036). The average ΔCts were 8.01 ±
0.41 for nonpregnant, 9.84 ± 0.32 for pregnant, and 9.77 ± 0.47 for postpartum ewes. Relative
expression (fold change) compared to nonpregnant ewes is depicted in Figure 5-8 (0.3 ± 0.1 for
pregnant and 0.5 ± 0.1 for postpartum ewes).
Discussion
Because the studies discussed previously in this dissertation have suggested differences in
HPA axis activation between these reproductive states, relative gene expression at the level of
the hypothalamus was investigated in order to elucidate a mechanism for these potential
differences. The specific objective of this study was to characterize any differences in relative
mRNA expression levels of HPA axis- and serotonergic system-relevant genes in the
hypothalamus of nonpregnant, pregnant, and post-partum ewes. We hypothesized that changes
at the transcriptional level could at least partially explain regulatory differences in basal HPA
axis activity between the pregnant and nonpregnant states.
More specifically, one possible mechanism for the increased basal ACTH and cortisol of
pregnancy and the blunted early HPA axis response to intravenous MR blockade in pregnant
ewes would be a decrease in hypothalamic MR expression. Contrary to our hypothesis,
however, there were no differences in nonpregnant, pregnant, and postpartum expression of
hypothalamic MR expression. There were also no detectable differences in hypothalamic GR,
CRH, or AVP gene expression among the reproductive groups studied here. On the other hand
these data are consistent with previous findings in our lab that have also shown no differences in
hippocampal MR or GR between pregnant and nonpregnant ewes (unpublished data by Yi Hua),

98
nor were there differences between adrenal intact and adrenalectomized ewes (Richards et al.
2003), indicating a lack of effect by ovarian or adrenal hormones on these genes in the
hippocampus. However, several studies in rats have suggested that hippocampal GR and MR are
negatively regulated by adrenal hormones (Tornello et al. 1982, Sapolsky et al. 1984, Sapolsky
& McEwen 1985, Herman et al. 1989, Reul et al. 1989, Kalman & Spencer 2002) and possibly
estrogen (Burgess & Handa 1993, Carey et al. 1995, Castren et al. 1995).
Extensive evidence indicates a stimulatory role for serotonin on HPA axis activity. We
originally hypothesized that increased serotonergic responsivity might at least in part explain the
increase in basal ACTH and cortisol associated with pregnancy. However, based on the reduced
responses to acute serotonin reuptake transporter blockade during pregnancy, we expected
reduced hypothalamic expression of 5-HT2A or SERT and/or increases in hypothalamic 5-HT1A
in pregnant ewes. Whereas, based on the influence of ovarian hormones on this system as
discussed in Chapter 2, we might have predicted increased expression of 5-HT2A and decreased
expression of 5-HT1A, but likely still a decrease in SERT expression. The prediction for 5-HT1A
expression would be similar if the elevated cortisol of pregnancy is considered; however, the
data is limited regarding glucocorticoid modulation of 5-HT2A or SERT expression. Our
hypotheses were disproved as none of the serotonergic genes investigated in this study were
significantly different between nonpregnant, pregnant or postpartum states.
As a result of the interesting effects of subchronic icv infusion of fluoxetine on food intake
discussed in Chapter 4, we investigated the possibility of differential hypothalamic expression of
proopiomelanocortin (POMC) between nonpregnant and pregnant ewes. The data suggested a
trend for fluoxetine to reduce feeding over time in the nonpregnant ewes, whereas there was no
effect of fluoxetine on daily food intake in the pregnant ewes. Serotonergic agents such as

99
fluoxetine are known to be anorexogenic in both animal models and in humans; and serotonin is
thought to play a role in POMC-mediated satiety mechanisms discussed in Chapter 2.
Specifically, POMC neurons in the arcuate nucleus of the hypothalamus, which are known to be
responsive to peripheral energy balance signals such as ghrelin and leptin, are thought to be
stimulated by serotonergic agents (Heisler et al. 2002). Upon activation, these neurons release
melanocortins (cleavage products of the precursor POMC) which then act on melanocortin
receptors to ultimately inhibit feeding (Cone 2006). Therefore, the reduced expression of POMC
mRNA in pregnant compared to nonpregnant ewes, might suggest a mechanism for increased
feeding during pregnancy and might at least partially explain the lack of an anorexogenic effect
of fluoxetine in these ewes.
Although relative expression of these genes was not shown to be altered in the
hypothalamus across these reproductive states for the majority of the genes analyzed in this
study, alterations at the gene level may still be occurring in other brain regions that are known to
influence HPA axis activity. For the purposes of this study, the hypothalamus was chosen as a
starting point for investigating possible differential patterns of expression across the reproductive
states as it is the site of convergence for all upstream brain regions which are thought to regulate
cortisol. The hypothalamus receives signals from both inhibitory and stimulatory sources; and
the resultant cortisol release is from an integration of these inputs. It is also known that MR, GR,
serotonin receptors and serotonin reuptake transporters are expressed in many other brain regions
besides the hypothalamus. It is therefore entirely possible that expression patterns of the genes
of interest to these studies may be altered during pregnancy in other brain regions such as the
hippocampus or at the level of the brain stem which houses the serotonergic cell bodies. I
propose that there may be increased expression of the 5-HT1A autoreceptor at the level the brain

100
stem as a possible mechanism for reduced serotonergic tone in the pregnant ewes which could
explain the differential response to acute icv fluoxetine.
Specifically, we know from previous reports that both MR and GR are expressed in the
hippocampus; a region thought to inhibit HPA axis activity and that this area also receives inputs
from serotonergic neurons. The 5-HT1A receptor, for example, is known to be one of the most
highly expressed serotonin receptors in the hippocampus (Hoyer et al. 1986, Pazos et al. 1987,
Chalmers & Watson 1991, Chalmers et al. 1993, Pucadyil et al. 2005). This region of the brain
is therefore a prime target for future gene expression studies.
Additionally, it is possible that changes in hypothalamic expression of these genes are
simply being diluted by not specifically isolating and analyzing the paraventricular nucleus from
these ewes. It is also entirely possible, however, that any changes in basal HPA axis regulation
involving these systems that may occur during pregnancy are taking place after gene
transcription. Such changes might include alterations in the regulation of translation or post-
translational differences. Studies have shown, for example that hormone-dependent
phosphorylation of steroid receptors imparts stability and therefore influences receptor levels
available for binding within the cytosol and subsequent transactivation (for review see, Weigel
1996). It is also important to note that the current study has, by no means, represented an
exhaustive investigation of all possible genes within the hypothalamus that are known be
involved in HPA axis regulation, nor can we disregard the possibility of yet undiscovered
systems that influence cortisol regulation that may be altered during pregnancy.
This research was supported by an R01 grant from the NIH (DK38114) to Maureen Keller-
Wood.

101
Table 5-1. Sequences of probe and primer sets for all genes analyzed in ovine hypothalamus
along with the respective concentrations added to each reaction.

102
Figure 5-1. Relative hypothalamic glucocorticoid receptor (GR) mRNA expression. No
significant differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

103
Figure 5-2. Relative hypothalamic mineralocorticoid receptor (MR) mRNA expression. No
significant differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

104
Figure 5-3. Relative hypothalamic corticotropin-releasing hormone (CRH) mRNA expression.
No significant differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

105
Figure 5-4. Relative hypothalamic arginine vasopressin (AVP) mRNA expression. No
significant differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

106
Figure 5-5. Relative hypothalamic 5-HT1A receptor mRNA expression. No significant
differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

107
Figure 5-6. Relative hypothalamic 5-HT2A receptor mRNA expression. No significant
differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

108
Figure 5-7. Relative hypothalamic serotonin reuptake transporter (SERT) mRNA expression.
No significant differences between nonpregnant (NP), pregnant (P), and postpartum (PP) ewes. Data are expressed as fold change from nonpregnant ewes ± SEM.

109
Figure 5-8. Relative hypothalamic proopiomelanocortin (POMC) mRNA expression. Based on
one-way ANOVA, followed by Duncan’s Multiple Range test, delta Ct was significantly greater in both pregnant (P) and postpartum (PP) ewes compared to nonpregnant (NP) ewes, suggesting reduced hypothalamic POMC mRNA expression during pregnancy and early postpartum. Data are expressed as fold change from nonpregnant ewes ± SEM. * Indicates significantly different from nonpregnant group (p<0.05).

110
CHAPTER 6 SUMMARY
The overall goal of the research being done in our lab is to investigate the mechanisms of
maternal adaptation to pregnancy. More specifically, we are interested in the regulation of the
hypothalamic-pituitary-adrenal (HPA) axis during pregnancy. In the literature, several systems
have been found be associated with HPA axis regulation in normal physiology, either in an
inhibitory or stimulatory manner. The studies contained in this manuscript focus on two of those
systems and are aimed at comparing: (1) the relative role of the mineralocorticoid receptor (MR)
and (2) relative serotonergic responsivity between pregnant and non-pregnant sheep as they
relate to negative feedback control of basal HPA axis activity. In order to investigate the
relationship of these systems to HPA axis regulation during pregnancy, we studied pregnant,
nonpregnant and postpartum ewes.
For the first study, a mineralocorticoid antagonist, canrenoate was given intravenously to
both pregnant and nonpregnant ewes continuously over a period of 4 hours. Blockade of this
receptor systemically, allowed us to determine the relative role of both central and peripheral
MR in the regulation of both basal HPA axis activity and hemodynamic changes that occur
during pregnancy. Peripheral MR are known to be important in normal physiology for
electrolyte and fluid balance via binding by the mineralocorticoid, aldosterone at the renal
tubule, while central MR are thought to be the major steroid receptor involved in regulation of
basal HPA axis activity. The hypothesis for this part of my dissertation work was that systemic
blockade of MR would stimulate ACTH and cortisol and this effect may be blunted in the
pregnant ewe, reflecting an inherent reduction in the ability of MR to participate in negative
feedback during pregnancy as an underlying hypothesis in our lab is that progesterone
antagonizes cortisol negative feedback at MR. If this inherent reduction is in fact present, it

111
could provide one mechanism for increased basal HPA axis activity during pregnancy. These
results did in fact support my hypothesis as the response pattern of the plasma cortisol and
ACTH measured hourly during infusion with the MR antagonist, canrenoate, differed between
pregnant and nonpregnant ewes. The cortisol response was blunted in pregnant ewes during the
first half of canrenoate infusion and was reflected in the pattern of ACTH release which is
therefore suggestive of central or pituitary MR-mediated effects. The responses during the
second half of the infusion support the ‘underfill’ hypothesis of pregnancy as homeostatic factors
are upregulated to a greater extent in pregnant ewes in response to the subtle hemodynamic
effects of canrenoate.
For the second study, a selective serotonin reuptake inhibitor (SSRI), fluoxetine was given
intracerebroventricularly either acutely or continuously over a period of 6 days to pregnant and
nonpregnant ewes. Blockade of the serotonin reuptake transporter, allows the neurotransmitter
serotonin to remain in the synaptic cleft for a longer period, thereby enhancing its effects.
Administration through this route allows us to avoid any peripheral effects of fluoxetine that
might secondarily influence HPA axis activity. The serotonergic system has been shown, in
numerous animal models including sheep, to increase plasma ACTH and cortisol concentrations.
The hypothesis for this part of my dissertation work was that if increased serotonergic tone is
partially responsible for elevation in HPA axis activity during pregnancy, than administration of
an SSRI would demonstrate this via enhanced SSRI-stimulated ACTH and cortisol release in
pregnant compared to nonpregnant or postpartum ewes. The results of the acute study disproved
our hypothesis, in directionality but may still have uncovered potential differences in
responsivity to serotonergic agents between the pregnant and postpartum states in terms of HPA
axis activation, whereas the results of our subchronic infusion study may support our hypothesis.

112
Additionally, the observation that fluoxetine infusion had less of an inhibitory effect on post-
operative feeding recovery in the pregnant ewes, may provide evidence that central serotonin-
mediated feeding regulation may also be altered in pregnancy. As we know from the literature,
most if not all of the regions involved in the serotonin-mediated stimulation of ACTH and
cortisol are highly abundant in both corticosteroid and ovarian hormone receptors allowing for
the possibility of modulation by these steroids. Previous reports have demonstrated highly
variable effects across brain region and across species which are further compounded by variable
effects based on the duration of exposure to the steroids.
Because the studies discussed previously in this manuscript have suggested differences in
HPA axis activation between these reproductive states, I then sought to investigate whether such
differences are occurring as a result of changes at the gene expression level. The objective of the
final study was to characterize any differences in relative mRNA expression levels of HPA axis-
and serotonergic system-relevant genes in the hypothalamus of nonpregnant, pregnant, and post-
partum ewes. We hypothesized that changes at the transcriptional level could help to explain
basal HPA axis regulation differences between the pregnant and nonpregnant states.
Specifically, we expected that we might see a decrease in hypothalamic MR expression.
Contrary to our hypothesis, however, there were no differences in nonpregnant, pregnant, and
postpartum expression of hypothalamic MR expression. This suggests that the differential
response to canrenoate as described in Chapter 3 is not due a decrease in hypothalamic MR
expression during pregnancy, but rather might support our lab’s alternative hypothesis that
progesterone is antagonizing cortisol negative feedback at MR. There were also no detectable
differences in hypothalamic GR, CRH, or AVP gene expression among the reproductive groups
studied here. Meanwhile, extensive evidence indicates a stimulatory role for serotonin on HPA

113
axis activity. We formulated multiple hypotheses based on evidence for glucocorticoid and
ovarian hormone modulation of serotonergic system-related genes from the literature.
Additionally, we hoped to explain the differential responses to fluoxetine that were described in
Chapter 4 through alterations in hypothalamic gene expression patterns. Our hypotheses were
disproved as none of the serotonergic genes investigated in this study were significantly different
between nonpregnant, pregnant or postpartum states. On the other hand, the differential daily
food intake response to subchronic fluoxetine between nonpregnant and pregnant ewes suggested
that POMC expression during pregnancy might be reduced, thereby causing a reduction in
downstream signally to satiety centers. In fact, hypothalamic POMC expression was
significantly reduced in pregnant ewes compared to nonpregnant ewes.
The overall conclusions of this work are that (1) the role of MR in mediating the negative
feedback effects of cortisol does, in fact, appear to be altered in pregnancy, (2) an increase in
serotonergic responsivity does not explain the increase in basal HPA axis activity during
pregnancy, and (3) relative MR, GR, CRH, AVP, 5-HT1A, 5-HT2A, and SERT expression were
not altered across the reproductive states investigated here, while POMC expression was reduced
during pregnancy.
Figure 6-1 is a representative model of HPA axis regulation as it relates to these studies,
based on the consensus from the literature. As we well know, the hypothalamus is responsible
for stimulating secretion of ACTH release by the anterior pituitary. These studies have
uncovered potential modulation of MR-mediated negative feedback and serotonergic system-
mediated stimulation of the HPA axis, which could be at the level of the hypothalamus, but may
also be occurring in upstream brain regions such as the hippocampus or brain stem.

114
Our lab has previously shown that corticosteroids are important for many changes that
must occur during pregnancy such as maternal volume expansion, uterine blood flow, and fetal
homeostasis which impact maternal and fetal health. It is also important to note that the lack of
global changes in expression of key HPA axis regulatory genes, which are typically known to be
altered in response to chronic stress, further indicates that pregnancy is not perceived as a
stressor. As an understudied, basic adaptive process of successful pregnancy, it is important that
we continue to characterize the regulation of the HPA axis. Due to the importance of
corticosteroids in regulating such processes, several systems are in place to ensure its tight
regulation. The elevations in basal plasma ACTH and cortisol that occur during pregnancy are,
therefore, undoubtedly the result of coordinated alterations in many of these systems and
therefore, alterations in regulation of the HPA axis by other mechanisms still remain to be
investigated.
Our understanding of HPA axis regulation both in normal physiology and in pregnancy is
still far from complete. Interestingly, however, if differences similar to those observed in these
studies are also evident in humans, the findings could suggest a need for adjustment of treatment
regimens in women for HPA axis dysregulation as well as depression and/or anxiety as a she
transitions between these reproductive states in order to protect the health of both the mother and
her developing fetus.
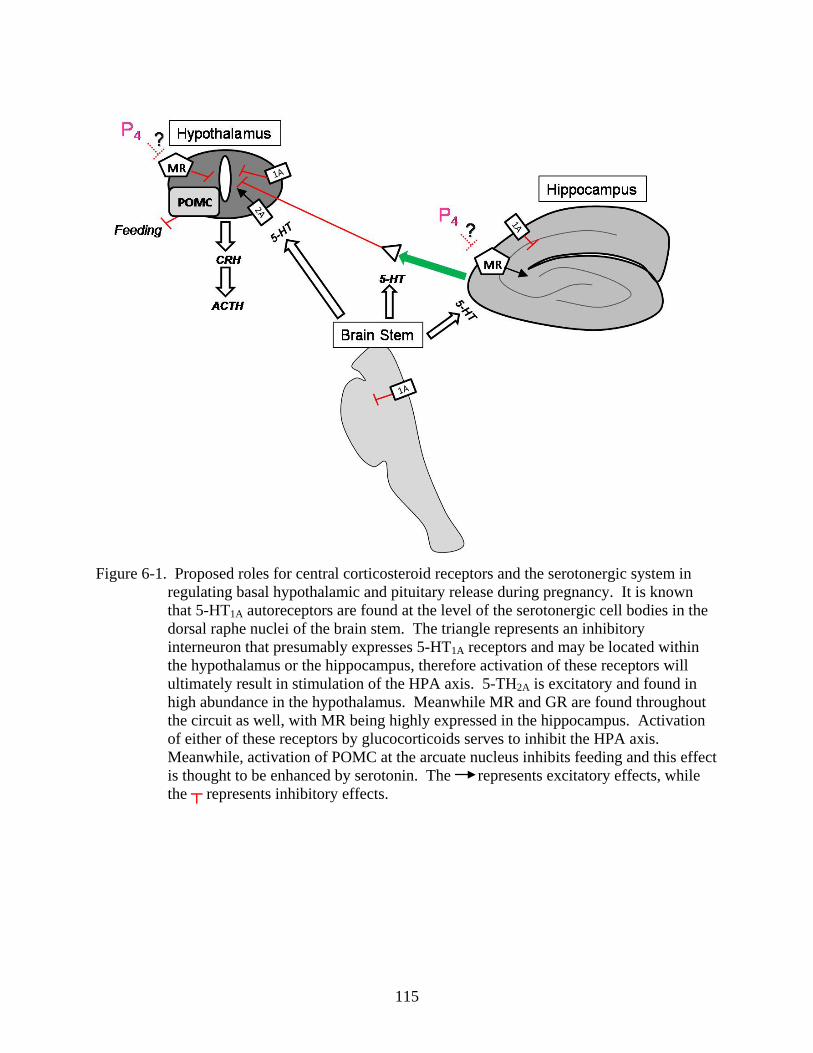
115
Figure 6-1. Proposed roles for central corticosteroid receptors and the serotonergic system in
regulating basal hypothalamic and pituitary release during pregnancy. It is known that 5-HT1A autoreceptors are found at the level of the serotonergic cell bodies in the dorsal raphe nuclei of the brain stem. The triangle represents an inhibitory interneuron that presumably expresses 5-HT1A receptors and may be located within the hypothalamus or the hippocampus, therefore activation of these receptors will ultimately result in stimulation of the HPA axis. 5-TH2A is excitatory and found in high abundance in the hypothalamus. Meanwhile MR and GR are found throughout the circuit as well, with MR being highly expressed in the hippocampus. Activation of either of these receptors by glucocorticoids serves to inhibit the HPA axis. Meanwhile, activation of POMC at the arcuate nucleus inhibits feeding and this effect is thought to be enhanced by serotonin. The represents excitatory effects, while the ┬ represents inhibitory effects.

116
LIST OF REFERENCES
Arriza JL, Simerly RB, Swanson LW & Evans RM 1988 The neuronal mineralocorticoid receptor as a mediator of glucocorticoid response. Neuron 1 887-900.
Arvat E, Maccagno B, Giordano R, Pellegrino M, Broglio F, Gianotti L, Maccario M, Camanni F & Ghigo E 2001 Mineralocorticoid receptor blockade by canrenoate increases both spontaneous and stimulated adrenal function in humans. The Journal of Clinical Endocrinology and Metabolism 86 3176-3181.
Azmitia EC & Segal M 1978 An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. The Journal of Comparative Neurology 179 641-667.
Bell ME, Wood CE & Keller-Wood M 1991 Influence of reproductive state on pituitary-adrenal activity in the ewe. Domestic Animal Endocrinology 8 245-254.
Benedict C, Kern W, Schmid SM, Schultes B, Born J & Hallschmid M 2009 Early morning rise in hypothalamic-pituitary-adrenal activity: A role for maintaining the brain's energy balance. Psychoneuroendocrinology 34 455-462.
Bethea CL, Brown NA & Kohama SG 1996 Steroid regulation of estrogen and progestin receptor messenger ribonucleic acid in monkey hypothalamus and pituitary. Endocrinology 137 4372-4383.
Bethea CL & Centeno ML 2008 Ovarian steroid treatment decreases corticotropin-releasing hormone (CRH) mRNA and protein in the hypothalamic paraventricular nucleus of ovariectomized monkeys. Neuropsychopharmacology 33 546-556.
Bethea CL, Lu NZ, Gundlah C & Streicher JM 2002 Diverse actions of ovarian steroids in the serotonin neural system. Frontiers in Neuroendocrinology 23 41-100.
Bevan JS & Scanlon MF 1998 Regulation of the hypothalamus and pituitary. In Clinical Endocrinology, p. 90, 2nd edn. A Grossman Ed. Oxford ; Malden, MA: Blackwell Science.
Biegon A, Reches A, Snyder L & McEwen BS 1983 Serotonergic and noradrenergic receptors in the rat brain: Modulation by chronic exposure to ovarian hormones. Life Sciences 32 2015-2021.
Blundell JE 1986 Serotonin manipulations and the structure of feeding behaviour. Appetite 7 Suppl 39-56.
Bohler HC,Jr, Zoeller RT, King JC, Rubin BS, Weber R & Merriam GR 1990 Corticotropin releasing hormone mRNA is elevated on the afternoon of proestrus in the parvocellular paraventricular nuclei of the female rat. Brain Research.Molecular Brain Research 8 259-262.

117
Bradbury MJ, Akana SF, Cascio CS, Levin N, Jacobson L & Dallman MF 1991 Regulation of basal ACTH secretion by corticosterone is mediated by both type I (MR) and type II (GR) receptors in rat brain. The Journal of Steroid Biochemistry and Molecular Biology 40 133-142.
Brady LS, Gold PW, Herkenham M, Lynn AB & Whitfield HJ,Jr 1992 The antidepressants fluoxetine, idazoxan and phenelzine alter corticotropin-releasing hormone and tyrosine hydroxylase mRNA levels in rat brain: Therapeutic implications. Brain Research 572 117-125.
Brinklow BR & Forbes JM 1984 The effect of short and skeleton long photoperiods on the plasma concentrations of prolactin and cortisol in sheep. Reproduction, Nutrition, Development 24 107-116.
Briones-Aranda A, Castillo-Salazar M & Picazo O 2008 Adrenalectomy modifies the hippocampal 5-HT(1A) receptors and the anxiolytic-like effect of 8-OH-DPAT in rats. Pharmacology, Biochemistry, and Behavior .
Broadbear JH, Hutton LC, Clarke IJ & Canny BJ 2004 Sex differences in the pituitary-adrenal response following acute antidepressant treatment in sheep. Psychopharmacology 171 450-457.
Broadbear JH, Pierce BN, Clarke IJ & Canny BJ 2005 Role of sex and sex steroids in mediating pituitary-adrenal responses to acute buspirone treatment in sheep. Journal of Neuroendocrinology 17 804-810.
Browning KN & Travagli RA 2006 Short-term receptor trafficking in the dorsal vagal complex: An overview. Autonomic Neuroscience : Basic & Clinical 126-127 2-8.
Buckingham JC, Dohler KD & Wilson CA 1978 Activity of the pituitary-adrenocortical system and thyroid gland during the oestrous cycle of the rat. The Journal of Endocrinology 78 359-366.
Burgess LH & Handa RJ 1992 Chronic estrogen-induced alterations in adrenocorticotropin and corticosterone secretion, and glucocorticoid receptor-mediated functions in female rats. Endocrinology 131 1261-1269.
Burgess LH & Handa RJ 1993 Hormonal regulation of androgen receptor mRNA in the brain and anterior pituitary gland of the male rat. Brain Research.Molecular Brain Research 19 31-38
Calogero AE, Bagdy G, Szemeredi K, Tartaglia ME, Gold PW & Chrousos GP 1990 Mechanisms of serotonin receptor agonist-induced activation of the hypothalamic-pituitary-adrenal axis in the rat. Endocrinology 126 1888-1894.

118
Calogero AE, Bernardini R, Margioris AN, Bagdy G, Gallucci WT, Munson PJ, Tamarkin L, Tomai TP, Brady L & Gold PW 1989 Effects of serotonergic agonists and antagonists on corticotropin-releasing hormone secretion by explanted rat hypothalami. Peptides 10 189-200.
Carey MP & de Kloet ER 1994 Interaction of progesterone with the hippocampal mineralocorticoid receptor. Annals of the New York Academy of Sciences 746 434-437.
Carey MP, Deterd CH, de Koning J, Helmerhorst F & de Kloet ER 1995 The influence of ovarian steroids on hypothalamic-pituitary-adrenal regulation in the female rat. The Journal of Endocrinology 144 311-321.
Carr BR, Parker CR,Jr, Madden JD, MacDonald PC & Porter JC 1981 Maternal plasma adrenocorticotropin and cortisol relationships throughout human pregnancy. American Journal of Obstetrics and Gynecology 139 416-422.
Carrasco GA & Van de Kar LD 2003 Neuroendocrine pharmacology of stress. European Journal of Pharmacology 463 235-272.
Castren M, Patchev VK, Almeida OF, Holsboer F, Trapp T & Castren E 1995 Regulation of rat mineralocorticoid receptor expression in neurons by progesterone. Endocrinology 136 3800-3806.
Chalmers DT, Kwak SP, Mansour A, Akil H & Watson SJ 1993 Corticosteroids regulate brain hippocampal 5-HT1A receptor mRNA expression. The Journal of Neuroscience 13 914-923.
Chalmers DT, Lopez JF, Vazquez DM, Akil H & Watson SJ 1994 Regulation of hippocampal 5-HT1A receptor gene expression by dexamethasone. Neuropsychopharmacology 10 215-222.
Chalmers DT & Watson SJ 1991 Comparative anatomical distribution of 5-HT1A receptor mRNA and 5-HT1A binding in rat brain--a combined in situ hybridisation/in vitro receptor autoradiographic study. Brain Research 561 51-60.
Charmandari E, Tsigos C & Chrousos G 2005 Endocrinology of the stress response. Annual Review of Physiology 67 259-284.
Charnvises S, Fencl MD, Osathanondh R, Zhu MG, Underwood R & Tulchinsky D 1985 Adrenal steroids in maternal and cord blood after dexamethasone administration at midterm. The Journal of Clinical Endocrinology and Metabolism 61 1220-1222.
Chen M, Lee J & Malvin RL 1989 Central administration of aldosterone increases blood pressure in rats. Clinical and Experimental Hypertension.Part A, Theory and Practice 11 459-472.

119
Chen HY, Trumbauer ME, Chen AS, Weingarth DT, Adams JR, Frazier EG, Shen Z, Marsh DJ, Feighner SD, Guan XM et al 2004 Orexigenic action of peripheral ghrelin is mediated by neuropeptide Y and agouti-related protein. Endocrinology 145 2607-2612.
Chrousos GP 1992 Regulation and dysregulation of the hypothalamic-pituitary-adrenal axis. the corticotropin-releasing hormone perspective. Endocrinology and Metabolism Clinics of North America 21 833-858.
Chrousos GP & Gold PW 1992 The concepts of stress and stress system disorders. overview of physical and behavioral homeostasis. The Journal of the American Medical Association 267 1244-1252.
Cone RD 2006 Studies on the physiological functions of the melanocortin system. Endocrine Reviews 27 736-749.
Contesse V, Lefebvre H, Lenglet S, Kuhn JM, Delarue C & Vaudry H 2000 Role of 5-HT in the regulation of the brain-pituitary-adrenal axis: Effects of 5-HT on adrenocortical cells. Canadian Journal of Physiology and Pharmacology 78 967-983.
Cooper JR, Bloom FE & Roth RH 2003 Serotonin (5-hydroxytryptamine), histamine, and adenosine. In The Biochemical Basis of Neuropharmacology, 8th edn. Oxford ; New York: Oxford University Press.
Couette B, Lombes M, Baulieu EE & Rafestin-Oblin ME 1992 Aldosterone antagonists destabilize the mineralocorticosteroid receptor. The Biochemical Journal 282 ( Pt 3) 697-702.
Cummings DE & Overduin J 2007 Gastrointestinal regulation of food intake. The Journal of Clinical Investigation 117 13-23.
Cyr M, Bosse R & Di Paolo T 1998 Gonadal hormones modulate 5-hydroxytryptamine2A receptors: Emphasis on the rat frontal cortex. Neuroscience 83 829-836.
Damjanovic SS, Stojic RV, Lalic NM, Jotic AZ, Macut DP, Ognjanovic SI, Petakov MS & Popovic BM 2009 Relationship between basal metabolic rate and cortisol secretion throughout pregnancy. Endocrine 35 262-268.
Dayas CV, Xu Y, Buller KM & Day TA 2000 Effects of chronic oestrogen replacement on stress-induced activation of hypothalamic-pituitary-adrenal axis control pathways. Journal of Neuroendocrinology 12 784-794.
de Kloet ER, Sutanto W, van den Berg DT, Carey MP, van Haarst AD, Hornsby CD, Meijer OC, Rots NY & Oitzl MS 1993 Brain mineralocorticoid receptor diversity: Functional implications. The Journal of Steroid Biochemistry and Molecular Biology 47 183-190.
de Kloet ER, Vreugdenhil E, Oitzl MS & Joels M 1998 Brain corticosteroid receptor balance in health and disease. Endocrine Reviews 19 269-301.

120
Deuschle M, Weber B, Colla M, Muller M, Kniest A & Heuser I 1998 Mineralocorticoid receptor also modulates basal activity of hypothalamus-pituitary-adrenocortical system in humans. Neuroendocrinology 68 355-360.
Dhillo WS & Bloom SR 2004 Gastrointestinal hormones and regulation of food intake. Hormone and Metabolic Research 36 846-851.
Diamond MI, Miner JN, Yoshinaga SK & Yamamoto KR 1990 Transcription factor interactions: Selectors of positive or negative regulation from a single DNA element. Science 249 1266-1272.
Dodt C, Kern W, Fehm HL & Born J 1993 Antimineralocorticoid canrenoate enhances secretory activity of the hypothalamus-pituitary-adrenocortical (HPA) axis in humans. Neuroendocrinology 58 570-574.
Erickson K, Thorsen P, Chrousos G, Grigoriadis DE, Khongsaly O, McGregor J & Schulkin J 2001 Preterm birth: Associated neuroendocrine, medical, and behavioral risk factors. The Journal of Clinical Endocrinology and Metabolism 86 2544-2552.
Ferrini M & De Nicola AF 1991 Estrogens up-regulate type I and type II glucocorticoid receptors in brain regions from ovariectomized rats. Life Sciences 48 2593-2601.
Frankfurt M, McKittrick CR, Mendelson SD & McEwen BS 1994 Effect of 5,7-dihydroxytryptamine, ovariectomy and gonadal steroids on serotonin receptor binding in rat brain. Neuroendocrinology 59 245-250.
Frazer A & Hensler JG 1999 Serotonin. In Basic Neurochemistry : Molecular, Cellular, and Medical Aspects, 6th edn. GJ Siegel, BW Agranoff, RW Albers, SK Fisher & MD Uhler Eds. Philadelphia, Pennsylvania: Lippincott-Raven Publishers.
Fukuda K, Uetsuki N, Uga H, Hashiguchi M, Sato M, Hisano T, Segawa H & Iwasaki Y 2003 Potentiation of proopiomelanocortin gene expression in cultured pituitary cells by benzodiazepines. Anesthesiology 98 1172-1177.
Fuller RW 1996 Serotonin receptors involved in regulation of pituitary-adrenocortical function in rats. Behavioural Brain Research 73 215-219.
Funder JW, Feldman D, Highland E & Edelman IS 1974 Molecular modifications of anti-aldosterone compounds: Effects on affinity of spirolactones for renal aldosterone receptors. Biochemical Pharmacology 23 1493-1501.
Funder J & Myles K 1996 Exclusion of corticosterone from epithelial mineralocorticoid receptors is insufficient for selectivity of aldosterone action: In vivo binding studies. Endocrinology 137 5264-5268.
Funder JW 1996 Mineralocorticoid receptors in the central nervous system. The Journal of Steroid Biochemistry and Molecular Biology 56 179-183.

121
Fuxe K, Cintra A, Agnati LF, Harfstrand A, Wikstrom AC, Okret S, Zoli M, Miller LS, Greene JL & Gustafsson JA 1987 Studies on the cellular localization and distribution of glucocorticoid receptor and estrogen receptor immunoreactivity in the central nervous system of the rat and their relationship to the monoaminergic and peptidergic neurons of the brain. Journal of Steroid Biochemistry 27 159-170.
Gartside SE & Cowen PJ 1990 Mediation of ACTH and prolactin responses to 5-HTP by 5-HT2 receptors. European Journal of Pharmacology 179 103-109.
Gomez-Sanchez EP 1997 Central hypertensive effects of aldosterone. Frontiers in Neuroendocrinology 18 440-462.
Gomez-Sanchez EP, Fort CM & Gomez-Sanchez CE 1990 Intracerebroventricular infusion of RU28318 blocks aldosterone-salt hypertension. The American Journal of Physiology 258 E482-4.
Gonen R, Perez R, David M, Dar H, Merksamer R & Sharf M 1992 The association between unexplained second-trimester maternal serum hCG elevation and pregnancy complications. Obstetrics and Gynecology 80 83-86.
Grazul-Bilska AT, Redmer DA, Johnson ML, Jablonka-Shariff A, Bilski JJ & Reynolds LP 1996 Gap junctional protein connexin 43 in bovine corpora lutea throughout the estrous cycle. Biology of Reproduction 54 1279-1287.
Grottoli S, Giordano R, Maccagno B, Pellegrino M, Ghigo E & Arvat E 2002 The stimulatory effect of canrenoate, a mineralocorticoid antagonist, on the activity of the hypothalamus-pituitary-adrenal axis is abolished by alprazolam, a benzodiazepine, in humans. The Journal of Clinical Endocrinology and Metabolism 87 4616-4620.
Gundlah C, Pecins-Thompson M, Schutzer WE & Bethea CL 1999 Ovarian steroid effects on serotonin 1A, 2A and 2C receptor mRNA in macaque hypothalamus. Brain Research.Molecular Brain Research 63 325-339.
Harfstrand A, Fuxe K, Cintra A, Agnati LF, Zini I, Wikstrom AC, Okret S, Yu ZY, Goldstein M & Steinbusch H 1986 Glucocorticoid receptor immunoreactivity in monoaminergic neurons of rat brain. Proceedings of the National Academy of Sciences of the United States of America 83 9779-9783.
Hauger RL & Dautzenberg FM 2000 Regulation of the stress response by corticotropin-releasing factor receptors. In Neuroendocrinology in Physiology and Medicine, p. 261-287PM Conn & ME Freeman Eds. Totowa, New Jersey: Humana Press.
Heisler LK, Cowley MA, Kishi T, Tecott LH, Fan W, Low MJ, Smart JL, Rubinstein M, Tatro JB, Zigman JM et al 2003 Central serotonin and melanocortin pathways regulating energy homeostasis. Annals of the New York Academy of Sciences 994 169-174.

122
Heisler LK, Cowley MA, Tecott LH, Fan W, Low MJ, Smart JL, Rubinstein M, Tatro JB, Marcus JN, Holstege H et al 2002 Activation of central melanocortin pathways by fenfluramine. Science 297 609-611.
Heisler LK, Pronchuk N, Nonogaki K, Zhou L, Raber J, Tung L, Yeo GS, O'Rahilly S, Colmers WF, Elmquist JK et al 2007 Serotonin activates the hypothalamic-pituitary-adrenal axis via serotonin 2C receptor stimulation. The Journal of Neuroscience 27 6956-6964.
Herman JP & Cullinan WE 1997 Neurocircuitry of stress: Central control of the hypothalamo-pituitary-adrenocortical axis. Trends in Neurosciences 20 78-84.
Herman JP & Mueller NK 2006 Role of the ventral subiculum in stress integration. Behavioural Brain Research 174 215-224.
Herman JP, Schafer MK, Young EA, Thompson R, Douglass J, Akil H & Watson SJ 1989 Evidence for hippocampal regulation of neuroendocrine neurons of the hypothalamo-pituitary-adrenocortical axis. The Journal of Neuroscience 9 3072-3082.
Heuser I, Deuschle M, Weber B, Stalla GK & Holsboer F 2000 Increased activity of the hypothalamus-pituitary-adrenal system after treatment with the mineralocorticoid receptor antagonist spironolactone. Psychoneuroendocrinology 25 513-518.
Hoyer D, Pazos A, Probst A & Palacios JM 1986 Serotonin receptors in the human brain. I. characterization and autoradiographic localization of 5-HT1A recognition sites. apparent absence of 5-HT1B recognition sites. Brain Research 376 85-96.
Ichimura T, Uchiyama J, Kunihiro O, Ito M, Horigome T, Omata S, Shinkai F, Kaji H & Isobe T 1995 Identification of the site of interaction of the 14-3-3 protein with phosphorylated tryptophan hydroxylase. The Journal of Biological Chemistry 270 28515-28518.
Jacobs BL & Azmitia EC 1992 Structure and function of the brain serotonin system. Physiological Reviews 72 165-229.
Jacobs BL & Fornal CA 1999 Activity of serotonergic neurons in behaving animals. Neuropsychopharmacology 21 9S-15S.
Jacobson L & Sapolsky R 1991 The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenocortical axis. Endocrine Reviews 12 118-134.
Janiak PC, Lewis SJ & Brody MJ 1990 Role of central mineralocorticoid binding sites in development of hypertension. The American Journal of Physiology 259 R1025-34.
Jensen E, Wood C & Keller-Wood M 2002 The normal increase in adrenal secretion during pregnancy contributes to maternal volume expansion and fetal homeostasis. Journal of the Society for Gynecologic Investigation 9 362-371.

123
Jensen E, Wood CE & Keller-Wood M 2005 Chronic alterations in ovine maternal corticosteroid levels influence uterine blood flow and placental and fetal growth. American Journal of Physiology.Regulatory, Integrative and Comparative Physiology 288 R54-61.
Jensen E, Wood CE & Keller-Wood M 2003 Alterations in maternal corticosteroid levels influence fetal urine and lung liquid production. Journal of the Society for Gynecologic Investigation 10 480-489.
Johansson ED & Jonasson LE 1971 Progesterone levels in amniotic fluid and plasma from women. I. levels during normal pregnancy. Acta Obstetricia Et Gynecologica Scandinavica 50 339-343.
Jones SA, Brooks AN & Challis JR 1989 Steroids modulate corticotropin-releasing hormone production in human fetal membranes and placenta. The Journal of Clinical Endocrinology and Metabolism 68 825-830.
Jones SA & Challis JR 1989 Local stimulation of prostaglandin production by corticotropin-releasing hormone in human fetal membranes and placenta. Biochemical and Biophysical Research Communications 159 192-199.
Jones SA & Challis JR 1990 Steroid, corticotrophin-releasing hormone, ACTH and prostaglandin interactions in the amnion and placenta of early pregnancy in man. The Journal of Endocrinology 125 153-159.
Jorgensen H, Knigge U, Kjaer A, Moller M & Warberg J 2002 Serotonergic stimulation of corticotropin-releasing hormone and pro-opiomelanocortin gene expression. Journal of Neuroendocrinology 14 788-795.
Kalman BA & Spencer RL 2002 Rapid corticosteroid-dependent regulation of mineralocorticoid receptor protein expression in rat brain. Endocrinology 143 4184-4195.
Keller-Wood M 1998 Evidence for reset of regulated cortisol in pregnancy: Studies in adrenalectomized ewes. The American Journal of Physiology 274 R145-51.
Keller-Wood M 1996 Inhibition of stimulated and basal ACTH by cortisol during ovine pregnancy. The American Journal of Physiology 271 R130-6.
Keller-Wood M, Cudd TA, Norman W, Caldwell SM & Wood CE 1998 Sheep model for study of maternal adrenal gland function during pregnancy. Laboratory Animal Science 48 507-512.
Keller-Wood ME & Dallman MF 1984 Corticosteroid inhibition of ACTH secretion. Endocrine Reviews 5 1-24.
Keller-Wood ME, Shinsako J & Dallman MF 1983 Integral as well as proportional adrenal responses to ACTH. The American Journal of Physiology 245 R53-9.

124
Keller-Wood M & Wood CE 2008 Regulation of maternal ACTH in ovine pregnancy: Does progesterone play a role? American Journal of Physiology.Endocrinology and Metabolism 295 E913-20.
Kelley AE & Berridge KC 2002 The neuroscience of natural rewards: Relevance to addictive drugs. The Journal of Neuroscience 22 3306-3311.
Kellner M, Baker DG, Yassouridis A, Bettinger S, Otte C, Naber D & Wiedemann K 2002 Mineralocorticoid receptor function in patients with posttraumatic stress disorder. The American Journal of Psychiatry 159 1938-1940.
Kemppainen RJ & Sartin JL 1984 Evidence for episodic but not circadian activity in plasma concentrations of adrenocorticotrophin, cortisol and thyroxine in dogs. The Journal of Endocrinology 103 219-226.
Kile JP & Turner BB 1985 Serotonergic and cholinergic interaction in the regulation of pituitary-adrenal function in rats. Experientia 41 1123-1127.
Kirschbaum C, Tietze A, Skoluda N & Dettenborn L 2009 Hair as a retrospective calendar of cortisol production—Increased cortisol incorporation into hair in the third trimester of pregnancy. Psychoneuroendocrinology 34 32-37.
Kuhn DM, Arthur R,Jr & States JC 1997 Phosphorylation and activation of brain tryptophan hydroxylase: Identification of serine-58 as a substrate site for protein kinase A. Journal of Neurochemistry 68 2220-2223.
Kuroda Y, Watanabe Y, Albeck DS, Hastings NB & McEwen BS 1994 Effects of adrenalectomy and type I or type II glucocorticoid receptor activation on 5-HT1A and 5-HT2 receptor binding and 5-HT transporter mRNA expression in rat brain. Brain Research 648 157-161.
Lai M, McCormick JA, Chapman KE, Kelly PA, Seckl JR & Yau JL 2003 Differential regulation of corticosteroid receptors by monoamine neurotransmitters and antidepressant drugs in primary hippocampal culture. Neuroscience 118 975-984.
Lanfumey L, Mongeau R, Cohen-Salmon C & Hamon M 2008 Corticosteroid-serotonin interactions in the neurobiological mechanisms of stress-related disorders. Neuroscience and Biobehavioral Reviews 32 1174-1184.
Lee TT, Redila VA, Hill MN & Gorzalka BB 2009 5-HT(2A) receptor mediated neuronal activation within the paraventricular nucleus of the hypothalamus is desensitized following prolonged glucocorticoid treatment. European Journal of Pharmacology 602 54-57.
Lefcourt AM, Bitman J, Kahl S & Wood DL 1993 Circadian and ultradian rhythms of peripheral cortisol concentrations in lactating dairy cows. Journal of Dairy Science 76 2607-2612.
Lepage O, Tottmar O & Winberg S 2002 Elevated dietary intake of L-tryptophan counteracts the stress-induced elevation of plasma cortisol in rainbow trout (oncorhynchus mykiss). The Journal of Experimental Biology 205 3679-3687.

125
Li Q, Battaglia G & Van de Kar LD 1997 Autoradiographic evidence for differential G-protein coupling of 5-HT1A receptors in rat brain: Lack of effect of repeated injections of fluoxetine. Brain Research 769 141-151.
Liggins GC 1994 The role of cortisol in preparing the fetus for birth. Reproduction, Fertility, and Development 6 141-150.
Liggins GC & Howie RN 1972 A controlled trial of antepartum glucocorticoid treatment for prevention of the respiratory distress syndrome in premature infants. Pediatrics 50 515-525.
Lindsay JR & Nieman LK 2006 Adrenal disorders in pregnancy. Endocrinology and Metabolism Clinics of North America 35 1-20, v.
Liposits Z, Phelix C & Paull WK 1987 Synaptic interaction of serotonergic axons and corticotropin releasing factor (CRF) synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. A light and electron microscopic immunocytochemical study. Histochemistry 86 541-549.
Livak KJ & Schmittgen TD 2001 Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 25 402-408.
Lockwood CJ, Radunovic N, Nastic D, Petkovic S, Aigner S & Berkowitz GS 1996 Corticotropin-releasing hormone and related pituitary-adrenal axis hormones in fetal and maternal blood during the second half of pregnancy. Journal of Perinatal Medicine 24 243-251.
Lowy MT 1990 Reserpine-induced decrease in type I and II corticosteroid receptors in neuronal and lymphoid tissues of adrenalectomized rats. Neuroendocrinology 51 190-196.
Lozeva-Thomas V 2004 Serotonin brain circuits with a focus on hepatic encephalopathy. Metabolic Brain Disease 19 413-420.
Luini A, Lewis D, Guild S, Corda D & Axelrod J 1985 Hormone secretagogues increase cytosolic calcium by increasing cAMP in corticotropin-secreting cells. Proceedings of the National Academy of Sciences of the United States of America 82 8034-8038.
Luttge WG & Rupp ME 1989 Differential up- and down-regulation of type I and type II receptors for adrenocorticosteroid hormones in mouse brain. Steroids 53 59-76.
Masilamani S, Kim GH, Mitchell C, Wade JB & Knepper MA 1999 Aldosterone-mediated regulation of ENaC alpha, beta, and gamma subunit proteins in rat kidney. The Journal of Clinical Investigation 104 R19-23.
McEwen BS 1994 Corticosteroids and hippocampal plasticity. Annals of the New York Academy of Sciences 746 134-42.

126
McEwen BS & Stellar E 1993 Stress and the individual. mechanisms leading to disease. Archives of Internal Medicine 153 2093-2101.
McLean M, Bisits A, Davies J, Woods R, Lowry P & Smith R 1995 A placental clock controlling the length of human pregnancy. Nature Medicine 1 460-463.
McQueen JK, Wilson H & Fink G 1997 Estradiol-17 beta increases serotonin transporter (SERT) mRNA levels and the density of SERT-binding sites in female rat brain. Brain Research.Molecular Brain Research 45 13-23.
Mendelson SD & McEwen BS 1992 Autoradiographic analyses of the effects of adrenalectomy and corticosterone on 5-HT1A and 5-HT1B receptors in the dorsal hippocampus and cortex of the rat. Neuroendocrinology 55 444-450.
Mendelson SD, McKittrick CR & McEwen BS 1993 Autoradiographic analyses of the effects of estradiol benzoate on [3H]paroxetine binding in the cerebral cortex and dorsal hippocampus of gonadectomized male and female rats. Brain Research 601 299-302.
Moore RY & Eichler VB 1972 Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Research 42 201-206.
Morimoto M, Morita N, Ozawa H, Yokoyama K & Kawata M 1996 Distribution of glucocorticoid receptor immunoreactivity and mRNA in the rat brain: An immunohistochemical and in situ hybridization study. Neuroscience Research 26 235-269.
Naslund E & Hellstrom PM 2007 Appetite signaling: From gut peptides and enteric nerves to brain. Physiology & Behavior 92 256-262.
Neulen J & Breckwoldt M 1994 Placental progesterone, prostaglandins and mechanisms leading to initiation of parturition in the human. Experimental and Clinical Endocrinology 102 195-202.
Nolten WE & Rueckert PA 1981 Elevated free cortisol index in pregnancy: Possible regulatory mechanisms. American Journal of Obstetrics and Gynecology 139 492-498.
Nordeen SK, Bona BJ, Beck CA, Edwards DP, Borror KC & DeFranco DB 1995 The two faces of a steroid antagonist: When an antagonist isn't. Steroids 60 97-104.
Nussey SS & Whitehead SA 2001a Actions of glucocorticoids and clinical features of cushing's syndrome. In Endocrinology: An Integrated Approach. Taylor & Francis. From http://www.ncbi.nlm.nih.gov/bookshelf.
Nussey SS & Whitehead SA 2001b Cholesterol and steroid synthesis in the adrenal cortex. In Endocrinology: An Integrated Approach. Taylor & Francis. From http://www.ncbi.nlm.nih.gov/bookshelf.
Nussey SS & Whitehead SA 2001c Feedback control of glucocorticoids. In Endocrinology: An Integrated Approach. Taylor & Francis. From http://www.ncbi.nlm.nih.gov/bookshelf.

127
Nussey SS & Whitehead SA 2001d Glucocorticoid receptors. In Endocrinology: An Integrated Approach. Taylor & Francis. From http://www.ncbi.nlm.nih.gov/bookshelf
Odagiri E, Ishiwatari N, Abe Y, Jibiki K, Adachi T, Demura R, Demura H & Shizume K 1988 Hypercortisolism and the resistance to dexamethasone suppression during gestation. Endocrinologia Japonica 35 685-690.
Ogle TF & Kitay JI 1977 Ovarian and adrenal steroids during pregnancy and the oestrous cycle in the rat. The Journal of Endocrinology 74 89-98.
Olszewski JD & Baxter D 1954 In Cytoarchitecture of the human brain stem, p.199, Karger, New York, NY.
Osterlund MK & Hurd YL 1998 Acute 17 beta-estradiol treatment down-regulates serotonin 5HT1A receptor mRNA expression in the limbic system of female rats. Brain Research.Molecular Brain Research 55 169-172.
Parker RB, Yates CR, Laizure SC & Weber KT 2006 P-glycoprotein modulates aldosterone plasma disposition and tissue uptake. Journal of Cardiovascular Pharmacology 47 55-59.
Pazos A, Probst A & Palacios JM 1987 Serotonin receptors in the human brain--III. autoradiographic mapping of serotonin-1 receptors. Neuroscience 21 97-122.
Pecins-Thompson M & Bethea CL 1999 Ovarian steroid regulation of serotonin-1A autoreceptor messenger RNA expression in the dorsal raphe of rhesus macaques. Neuroscience 89 267-277.
Pecins-Thompson M, Brown NA & Bethea CL 1998 Regulation of serotonin re-uptake transporter mRNA expression by ovarian steroids in rhesus macaques. Brain Research.Molecular Brain Research 53 120-129.
Pecins-Thompson M & Keller-Wood M 1997 Effects of progesterone on blood pressure, plasma volume, and responses to hypotension. The American Journal of Physiology 272 R377-85.
Peiffer A & Barden N 1987 Estrogen-induced decrease of glucocorticoid receptor messenger ribonucleic acid concentration in rat anterior pituitary gland. Molecular Endocrinology 1 435-440.
Petraglia F, Coukos G, Volpe A, Genazzani AR & Vale W 1991 Involvement of placental neurohormones in human parturition. Annals of the New York Academy of Sciences 622 331-340.
Petraglia F, Sutton S & Vale W 1989 Neurotransmitters and peptides modulate the release of immunoreactive corticotropin-releasing factor from cultured human placental cells. American Journal of Obstetrics and Gynecology 160 247-251.

128
Petrov T, Krukoff TL & Jhamandas JH 1994 Chemically defined collateral projections from the pons to the central nucleus of the amygdala and hypothalamic paraventricular nucleus in the rat. Cell and Tissue Research 277 289-295.
Phillips JG & Poolsanguan W 1978 A method to study temporal changes in adrenal activity in relation to sexual status in the female laboratory rat. The Journal of Endocrinology 77 283-291.
Pollard I, White BM, Bassett JR & Cairncross KD 1975 Plasma glucocorticoid elevation and desynchronization of the estrous cycle following unpredictable stress in the rat. Behavioral Biology 14 103-108.
Pucadyil TJ, Kalipatnapu S & Chattopadhyay A 2005 The serotonin1A receptor: A representative member of the serotonin receptor family. Cellular and Molecular Neurobiology 25 553-580.
Raap DK, DonCarlos L, Garcia F, Muma NA, Wolf WA, Battaglia G & Van de Kar LD 2000 Estrogen desensitizes 5-HT(1A) receptors and reduces levels of G(z), G(i1) and G(i3) proteins in the hypothalamus. Neuropharmacology 39 1823-1832.
Rafestin-Oblin ME, Lombes M, Couette B & Baulieu EE 1992 Differences between aldosterone and its antagonists in binding kinetics and ligand-induced hsp90 release from mineralocorticosteroid receptor. The Journal of Steroid Biochemistry and Molecular Biology 41 815-821.
Ramamoorthy S & Blakely RD 1999 Phosphorylation and sequestration of serotonin transporters differentially modulated by psychostimulants. Science 285 763-766.
Raps D, Barthe PL & Desaulles PA 1971 Plasma and adrenal corticosterone levels during the different phases of the sexual cycle in normal female rats. Experientia 27 339-340.
Redei E, Li L, Halasz I, McGivern RF & Aird F 1994 Fast glucocorticoid feedback inhibition of ACTH secretion in the ovariectomized rat: Effect of chronic estrogen and progesterone. Neuroendocrinology 60 113-123.
Reul JM & de Kloet ER 1986 Anatomical resolution of two types of corticosterone receptor sites in rat brain with in vitro autoradiography and computerized image analysis. Journal of Steroid Biochemistry 24 269-272.
Reul JM & de Kloet ER 1985 Two receptor systems for corticosterone in rat brain: Microdistribution and differential occupation. Endocrinology 117 2505-2511.
Reul JM, Gesing A, Droste S, Stec IS, Weber A, Bachmann C, Bilang-Bleuel A, Holsboer F & Linthorst AC 2000 The brain mineralocorticoid receptor: Greedy for ligand, mysterious in function. European Journal of Pharmacology 405 235-249.

129
Reul JM, Pearce PT, Funder JW & Krozowski ZS 1989 Type I and type II corticosteroid receptor gene expression in the rat: Effect of adrenalectomy and dexamethasone administration. Molecular Endocrinology 3 1674-1680.
Reul JM, van den Bosch FR & de Kloet ER 1987 Relative occupation of type-I and type-II corticosteroid receptors in rat brain following stress and dexamethasone treatment: Functional implications. The Journal of Endocrinology 115 459-467.
Richards EM, Hua Y & Keller-Wood M 2003 Pharmacology and physiology of ovine corticosteroid receptors. Neuroendocrinology 77 2-14.
Roesch DM & Keller-Wood M 1999 Differential effects of pregnancy on mineralocorticoid and glucocorticoid receptor availability and immunoreactivity in cortisol feedback sites. Neuroendocrinology 70 55-62.
Rosenthal HE, Slaunwhite WR,Jr & Sandberg AA 1969 Transcortin: A corticosteroid-binding protein of plasma. X. cortisol and progesterone interplay and unbound levels of these steroids in pregnancy. The Journal of Clinical Endocrinology and Metabolism 29 352-367.
Rothuizen J, Reul JM, van Sluijs FJ, Mol JA, Rijnberk A & de Kloet ER 1993 Increased neuroendocrine reactivity and decreased brain mineralocorticoid receptor-binding capacity in aged dogs. Endocrinology 132 161-168.
Roy BN, Reid RL & Van Vugt DA 1999 The effects of estrogen and progesterone on corticotropin-releasing hormone and arginine vasopressin messenger ribonucleic acid levels in the paraventricular nucleus and supraoptic nucleus of the rhesus monkey. Endocrinology 140 2191-2198.
Rupprecht R, Reul JM, van Steensel B, Spengler D, Soder M, Berning B, Holsboer F & Damm K 1993 Pharmacological and functional characterization of human mineralocorticoid and glucocorticoid receptor ligands. European Journal of Pharmacology 247 145-154.
Sanchez MM, Young LJ, Plotsky PM & Insel TR 2000 Distribution of corticosteroid receptors in the rhesus brain: Relative absence of glucocorticoid receptors in the hippocampal formation. The Journal of Neuroscience 20 4657-4668.
Sandman CA, Glynn L, Schetter CD, Wadhwa P, Garite T, Chicz-DeMet A & Hobel C 2006 Elevated maternal cortisol early in pregnancy predicts third trimester levels of placental corticotropin releasing hormone (CRH): Priming the placental clock. Peptides 27 1457-1463.
Sapolsky RM, Krey LC & McEwen BS 1984 Glucocorticoid-sensitive hippocampal neurons are involved in terminating the adrenocortical stress response. Proceedings of the National Academy of Sciences of the United States of America 81 6174-6177.
Sapolsky RM, Krey LC & McEwen BS 1986 The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocrine Reviews 7 284-301.

130
Sapolsky RM & McEwen BS 1985 Down-regulation of neural corticosterone receptors by corticosterone and dexamethasone. Brain Research 339 161-165.
Schrier RW & Durr JA 1987 Pregnancy: An overfill or underfill state. American Journal of Kidney Diseases 9 284-289.
Seckl JR, Dickson KL & Fink G 1990 Central 5,7-dihydroxytryptamine lesions decrease hippocampal glucocorticoid and mineralocorticoid receptor messenger ribonucleic acid expression. Journal of Neuroendocrinology 2 911-916.
Seckl JR & Fink G 1992 Antidepressants increase glucocorticoid and mineralocorticoid receptor mRNA expression in rat hippocampus in vivo. Neuroendocrinology 55 621-626.
Seckl JR & Fink G 1991 Use of in situ hybridization to investigate the regulation of hippocampal corticosteroid receptors by monoamines. The Journal of Steroid Biochemistry and Molecular Biology 40 685-688.
Semont A, Fache M, Ouafik L, Hery M, Faudon M & Hery F 1999 Effect of serotonin inhibition on glucocorticoid and mineralocorticoid expression in various brain structures. Neuroendocrinology 69 121-128.
Sliwowska JH, Lan N, Yamashita F, Halpert AG, Viau V & Weinberg J 2008 Effects of prenatal ethanol exposure on regulation of basal hypothalamic-pituitary-adrenal activity and hippocampal 5-HT1A receptor mRNA levels in female rats across the estrous cycle. Psychoneuroendocrinology 33 1111-1123.
Stark P, Fuller RW & Wong DT 1985 The pharmacologic profile of fluoxetine. The Journal of Clinical Psychiatry 46 7-13.
Staub O & Verrey F 2005 Impact of Nedd4 proteins and serum and glucocorticoid-induced kinases on epithelial na+ transport in the distal nephron. Journal of the American Society of Nephrology 16 3167-3174.
Sterling P & Eyer J 1988 Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress, Cognition and Health, p. 629S Fisher & JT Reason Eds. Chichester ; New York: John Wiley & Sons.
Sumner BE & Fink G 1995 Estrogen increases the density of 5-hydroxytryptamine(2A) receptors in cerebral cortex and nucleus accumbens in the female rat. The Journal of Steroid Biochemistry and Molecular Biology 54 15-20.
Sumner BE & Fink G 1997 The density of 5-hydoxytryptamine2A receptors in forebrain is increased at pro-oestrus in intact female rats. Neuroscience Letters 234 7-10.
Swanson LW 2000 Cerebral hemisphere regulation of motivated behavior. Brain Research 886 113-164.
Tecott LH 2007 Serotonin and the orchestration of energy balance. Cell Metabolism 6 352-361.

131
Tropper PJ, Goland RS, Wardlaw SL, Fox HE & Frantz AG 1987 Effects of betamethasone on maternal plasma corticotropin releasing factor, ACTH and cortisol during pregnancy. Journal of Perinatal Medicine 15 221-225.
Tornello S, Orti E, De Nicola AF, Rainbow TC & McEwen BS 1982 Regulation of glucocorticoid receptors in brain by corticosterone treatment of adrenalectomized rats. Neuroendocrinology 35 411-417.
Tsagarakis S, Navarra P, Rees LH, Besser M, Grossman A & Navara P 1989 Morphine directly modulates the release of stimulated corticotrophin-releasing factor-41 from rat hypothalamus in vitro. Endocrinology 124 2330-2335.
Tsigos C & Chrousos GP 1994 Physiology of the hypothalamic-pituitary-adrenal axis in health and dysregulation in psychiatric and autoimmune disorders. Endocrinology and Metabolism Clinics of North America 23 451-466.
Turner BB 1997 Influence of gonadal steroids on brain corticosteroid receptors: A minireview. Neurochemical Research 22 1375-1385.
Turner BB 1990 Sex difference in glucocorticoid binding in rat pituitary is estrogen dependent. Life Sciences 46 1399-1406.
Turner BB 1992 Sex differences in the binding of type I and type II corticosteroid receptors in rat hippocampus. Brain Research 581 229-236.
Ueda K, Okamura N, Hirai M, Tanigawara Y, Saeki T, Kioka N, Komano T & Hori R 1992 Human P-glycoprotein transports cortisol, aldosterone, and dexamethasone, but not progesterone. The Journal of Biological Chemistry 267 24248-24252.
Uhr M, Holsboer F & Muller MB 2002 Penetration of endogenous steroid hormones corticosterone, cortisol, aldosterone and progesterone into the brain is enhanced in mice deficient for both mdr1a and mdr1b P-glycoproteins. Journal of Neuroendocrinology 14 753-759.
Uphouse L 1997 Multiple serotonin receptors: Too many, not enough, or just the right number? Neuroscience and Biobehavioral Reviews 21 679-698.
Warren WB, Patrick SL & Goland RS 1992 Elevated maternal plasma corticotropin-releasing hormone levels in pregnancies complicated by preterm labor. American Journal of Obstetrics and Gynecology 166 1198-204; discussion 1204-7.
Watanabe M, Meeker CI, Gray MJ, Sims EA & Solomon S 1963 Secretion rate of aldosterone in normal pregnancy. The Journal of Clinical Investigation 42 1619-1631.
Weigel NL 1996 Steroid hormone receptors and their regulation by phosphorylation. The Biochemical Journal 319 ( Pt 3) 657-667.

132
Wellhoener P, Born J, Fehm HL & Dodt C 2004 Elevated resting and exercise-induced cortisol levels after mineralocorticoid receptor blockade with canrenoate in healthy humans. The Journal of Clinical Endocrinology and Metabolism 89 5048-5052.
Wolfe CD, Patel SP, Campbell EA, Linton EA, Anderson J, Lowry PJ & Jones MT 1988 Plasma corticotrophin-releasing factor (CRF) in normal pregnancy. British Journal of Obstetrics and Gynaecology 95 997-1002.
Wood CE, Cudd TA, Kane C & Engelke K 1993 Fetal ACTH and blood pressure responses to thromboxane mimetic U-46619. The American Journal of Physiology 265 R858-62.
Woods SC, Lutz TA, Geary N & Langhans W 2006 Pancreatic signals controlling food intake; insulin, glucagon and amylin. Philosophical Transactions of the Royal Society of London.Series B, Biological Sciences 361 1219-1235.
Wright DE, Seroogy KB, Lundgren KH, Davis BM & Jennes L 1995 Comparative localization of serotonin1A, 1C, and 2 receptor subtype mRNAs in rat brain. The Journal of Comparative Neurology 351 357-373.
Yang-Yen HF, Chambard JC, Sun YL, Smeal T, Schmidt TJ, Drouin J & Karin M 1990 Transcriptional interference between c-jun and the glucocorticoid receptor: Mutual inhibition of DNA binding due to direct protein-protein interaction. Cell 62 1205-1215.
Young EA, Lopez JF, Murphy-Weinberg V, Watson SJ & Akil H 1998 The role of mineralocorticoid receptors in hypothalamic-pituitary-adrenal axis regulation in humans. The Journal of Clinical Endocrinology and Metabolism 83 3339-3345.
Zoumakis E, Makrigiannakis A, Margioris A, Stournaras C & Gravanis A 1996 Corticotropin releasing hormone (CRH) in normal and pregnant uterus: Physiological implications. Frontiers in Bioscience : A Journal and Virtual Library 1 e1-8.

BIOGRAPHICAL SKETCH
Melissa Dawn Lingis was born Melissa Dawn Landen in 1976 in Fort Lauderdale, Florida
to David and Pamela Landen. Melissa developed a strong interest in science and critical thinking
as a member of the gifted program at Nova, a magnet school for Broward County, which she
attended from kindergarten through the twelfth grade. Immediately upon graduating high school,
Melissa began her undergraduate studies with the University of Florida during the summer of
1994. Melissa completed a Bachelor of Science degree in animal sciences in 1998 and began
applying to veterinary college to pursue her childhood dream of becoming a small animal
veterinarian.
As the end of her undergraduate studies was drawing near, Melissa began working as a
veterinary technician in several small animal practices both in Gainesville and in Fort
Lauderdale. Over time, Melissa slowly began to realize that she was not as enthusiastic about a
career in veterinary medicine as she had been as a child. After much consideration, she
eventually returned to the University of Florida in 2003 to attend graduate school as a means to
pursue a career in scientific research. In August 2004, Melissa completed a non-thesis Master of
Agriculture degree in animal sciences with a focus on reproductive biology, as this was one of
her major interests during her undergraduate studies.
While completing her master’s degree, she began working in the lab of Dr. Maureen
Keller-Wood as a laboratory technician. Melissa found the research exciting and enjoyed the
mentorship so much that she chose to extend her graduate school experience by adding to the
work being done in the lab. She was accepted into the doctorate program in the Department of
Pharmacodynamics and chose Dr. Keller-Wood as her major professor. Melissa’s dissertation
work focused on the relative roles of the mineralocorticoid receptor and the serotonergic system
in the regulation of basal maternal hypothalamic-pituitary-adrenal axis activity during pregnancy

in the ewe. Her work was funded by an R01 grant from the National Institutes of Health
(DK38114) awarded to Maureen Keller-Wood.
Melissa married Robert Lingis in 2006 and gave birth to their son, Matthew, in the summer
of 2008. Throughout the last year of her doctorate studies, she juggled motherhood, research,
and writing to the best of her abilities; ultimately receiving her PhD in August 2009. Melissa
looks forward to beginning post-doctoral work in the lab of Dr. Kirk Conrad in the College of
Medicine, also at the University of Florida. Here she will study the role of relaxin and its
receptor in cardiovascular and renal physiology. Following her post-doctoral work, Melissa
hopes to pursue a career in pre-clinical research at the industrial level and later return to
academia as a professor and mentor to students interested in a career in scientific research.


















