







Languages
Pages
Legal


DIPARTIMENTO DI BIOLOGIA ANIMALE E DELL’UOMO UNIVERSITÀ DI ROMA “LA SAPIENZA”
CORSO DI DOTTORATO IN BIOLOGIA ANIMALE
XXI CICLO (2005 – 2008)
TESI DI DOTTORATO DI RICERCA USO DI STRUMENTI
IN UNA POPOLAZIONE DI CEBUS LIBIDINOSUS ALLO STATO SELVATICO IN PIAUÍ, BRASILE
PhD Dissertation TOOL USE
IN A WILD POPULATION OF CEBUS LIBIDINOSUS IN PIAUI, BRAZIL
Noemi Spagnoletti
Roma, 2009


SCUOLA DI DOTTORATO IN BIOLOGIA ANIMALE DIPARTIMENTO DI BIOLOGIA ANIMALE E DELL’UOMO
UNIVERSITÀ DI ROMA “LA SAPIENZA”
XXI CICLO (2005-2008)
USO DI STRUMENTI IN UNA POPOLAZIONE DI CEBUS LIBIDINOSUS ALLO STATO SELVATICO IN PIAUÍ, BRASILE
TOOL USE IN A WILD POPULATION OF CEBUS LIBIDINOSUS IN PIAUI, BRAZIL
Noemi Spagnoletti
Docente guida: Dott.ssa Elisabetta Visalberghi (ISTC, CNR, Roma) Relatore interno: Prof. Carlo Utzeri (Università di Roma “La Sapienza”)
Roma, 2009


Ai miei genitori Alle mie famiglie, italiana e brasiliana


An anthropoid ape, if he could take a dispassionate view
of his own case, would admit that though he could form an
artful plan to plunder a garden—though he could use
stones for fighting or for breaking nuts, yet the thought of
fashioning a stone into a tool was quite beyond his scope.
Charles Darwin, The Descent of Man, 1871
�


INDICE
RIASSUNTO.................................................................................................................................. 1
ABSTRACT ................................................................................................................................... 3
1. INTRODUZIONE....................................................................................................................... 5
1.1 Uso di strumenti .................................................................................................................... 5 1.2 Uso di strumenti nei primati non umani ................................................................................. 6 1.3 Biologia del genere Cebus ..................................................................................................... 9
1.3.1 L’uso di strumenti nel genere Cebus ............................................................................. 11 1.4 Fattori che determinano l’uso di strumenti ........................................................................... 13 1.5 Fattori ecologici e uso di strumenti ...................................................................................... 14
2. OBIETTIVI DELLA RICERCA ............................................................................................... 17
2.1 L’uso di strumenti in Cebus libidinosus ............................................................................... 17 2.2 Ipotesi di ricerca e predizioni............................................................................................... 19 2.3 Obiettivi della ricerca .......................................................................................................... 20
3. AREA DI STUDIO................................................................................................................... 23
3.1 Boa Vista ............................................................................................................................ 23 3.2 Il Progetto EthoCebus.......................................................................................................... 26
4. METODI E MATERIALI ......................................................................................................... 27
4.1 I cebi di Boa Vista ............................................................................................................... 27 4.2 Osservazioni comportamentali............................................................................................. 30
4.2.1 Abituazione di animali selvatici alla presenza umana.................................................... 30 4.2.2 Registrazione dei pattern di attività............................................................................... 30
4.2.2.1 Analisi Statistiche .................................................................................................. 32 4.2.3 Registrazione dell’uso di strumenti ............................................................................... 33
4.2.3.1 Analisi Statistiche .................................................................................................. 35 4.3 Indici di disponibilità alimentare e dati climatici.................................................................. 38
4.3.1 Disponibilità di frutta.................................................................................................... 38 4.3.2 Disponibilità di invertebrati .......................................................................................... 38 4.3.3 Produttività delle palme ................................................................................................ 39 4.3.4 Dati climatici ................................................................................................................ 40
4.3.4.1 Analisi Statistiche .................................................................................................. 40
5. RISULTATI.............................................................................................................................. 41
5.1 Disponibilità delle risorse .................................................................................................... 41 5.2 Pattern di attività................................................................................................................. 44
5.2.1 Variabilità tra gruppi nei pattern di attività ................................................................... 44 5.2.2 Differenze tra classi di età e sesso nei pattern di attività................................................ 46 5.2.3 Variazione stagionale nei pattern di attività .................................................................. 48 5.2.4 Scelta del substrato ....................................................................................................... 49 5.2.5 Differenze tra classi di età e sesso nella scelta del substrato .......................................... 52
5.3 Uso di strumenti .................................................................................................................. 53 5.3.1 Confronto tra gruppi ..................................................................................................... 55 5.3.2 Relazione con dati climatici e abbondanza di risorse trofiche........................................ 57 5.3.3 Variazione stagionale.................................................................................................... 58 5.3.4 Confronti tra classi di età e sesso .................................................................................. 60

5.3.5 Efficenza nell’uso di strumenti ..................................................................................... 62 5.3.6 Uso dei materiali........................................................................................................... 65
6. DISCUSSIONE......................................................................................................................... 67
6.1 Disponibilità delle risorse .................................................................................................... 67 6.2 Pattern di attività................................................................................................................. 68
6.2.1 Variabilità tra gruppi e tra classi di età e sesso .............................................................. 68 6.2.2 Variazione stagionale.................................................................................................... 70 6.2.3 Scelta del substrato ....................................................................................................... 71 6.2.4 Differenze tra classi di età e sesso nell’uso del substrato ............................................... 72
6.3 Uso di strumenti .................................................................................................................. 74 6.3.1 Confronto tra gruppi ..................................................................................................... 74 6.3.2 Variazione stagionale.................................................................................................... 75 6.3.3 Confronti tra classi di età e sesso .................................................................................. 76 6.3.4 Efficenza e uso dei materiali ......................................................................................... 77
7. CONCLUSIONI ....................................................................................................................... 79
8. RINGRAZIAMENTI ................................................................................................................ 83
9. BIBLIOGRAFIA ...................................................................................................................... 87
APPENDICE 1 ............................................................................................................................. 99
APPENDICE 2 ........................................................................................................................... 101

1
RIASSUNTO
Lo scopo di questa tesi di dottorato è studiare l’uso di strumenti di due gruppi di Cebus
libidinosus che vivono allo stato selvatico nella regione di Boa Vista (Piauí, Brasile) ed in
particolare di descrivere l’utilizzo di sassi e incudini per rompere frutti dal guscio duro nelle varie
classi di età e sesso e determinare la frequenza di uso di strumenti in relazione a cambiamenti
stagionali nella disponibilità di risorse alimentari.
Il primo obiettivo della ricerca è stato quello di verificare la validità di due ipotesi alternative
proposte per spiegare la presenza di uso di strumenti. Secondo l’ipotesi della necessità, l’uso di
strumenti (che permette l’accesso a risorse alimentari altrimenti non disponibili) deve avvenire con
maggiore frequenza quando le risorse sono scarse (Boesch & Boesch-Achermann, 2000; Fox et al.,
2004; Moura e Lee, 2004). Secondo l’ipotesi dell’opportunità ambientale (Beck, 1980; Fox et al.,
2004) il comportamento di uso di strumenti è favorito da opportune condizioni locali.
Il secondo obiettivo della ricerca è stato determinare le differenze individuali nell’uso di
strumenti tra classi di età e sesso e testare l’ipotesi da noi formulata che, al contrario di ciò che è
stato osservato negli scimpanzé (Boesch & Boesch, 1984, Lonsdorf et al., 2004), nei cebi a causa
dello sforzo energetico molto elevato richiesto per rompere le noci e del dimorfismo sessuale, questo
comportamento sia più frequente nei maschi che nelle femmine.
I risultati hanno mostrato che l’uso di percussori nella popolazione di cebi esaminata è abituale,
ovverosia è mostrato dalla maggior parte degli individui e durante la maggior parte dell’anno
(secondo la definizione di Whiten et al., 1999). Inoltre, può essere definito tradizionale (Fragaszy &
Perry, 2003) in quanto nonostante che strumenti e frutti dal guscio duro siano presenti in molti altri
siti (anche se non in tutti) dove i cebi sono stati studiati, l’uso di percussori è stato osservato solo in
un numero molto limitato di popolazioni di Cebus libidinosus (Ottoni & Izar, 2008), ed è trasmesso
in generazione in generazione grazie ad influenze sociali (Resende et al., 2008; Silva, 2008).
L’assenza di una marcata differenza fra disponibilità delle risorse nella stagione secca e nella
stagione piovosa non permette di valutare l’ipotesi della necessità. Tuttavia l’ambiente semi-arido
di Boa Vista non sembra essere così povero di risorse in confronto ad altri ambienti in cui i cebi
vivono, come ad esempio la Foresta Atlantica (Izar, 2004; Nakai, 2007), da averli potuti spingere a
mettere a punto tecniche di uso di strumenti per aver accesso a risorse fondamentali per la loro
sopravvivenza. I nostri risultati sono in accordo con le predizioni dell’ipotesi dell’opportunità
ambientale (Beck, 1980; Fox et al., 2004) secondo cui l’emergenza di comportamenti innovativi –
tra cui l’uso di strumenti – è favorita da opportune condizioni ambientali. A Boa Vista sono presenti
sia sassi percussori sia superfici idonee per schiacciare le noci (Visalberghi et al., 2007) e i risultati

2
di questo studio mettono in evidenza che sebbene i cebi usino strumenti tutto l’anno, la frequenza
d’uso riflette la disponibilità delle noci e di altri frutti incapsulati.
Infine nei cebi, come da noi ipotizzato, le femmine usano strumenti meno frequentemente dei
maschi nonostante il loro elevato fabbisogno energetico dovuto ad allattamento e gestazione
(Clutton-Brock, 1977). E’ possibile che usare percussori sia così dispendioso a livello energetico da
non risultare vantaggioso per le femmine, la cui massa corporea e forza sono inferiori a quelle dei
maschi (Liu et al. 2009).

3
ABSTRACT
This study investigates the tool use behaviour in two wild groups of bearded capuchin monkeys
(Cebus libidinosus) living in the Fazenda Boa Vista (Piauí, Brazil) by determining the frequency of
tool use in relation to seasonal changes in food availability and by investigating the use of stone-
hammers and anvils to crack palm nuts and other encased fruits in relation to the age and sex of the
individuals.
Our first goal, was to test two alternative hypotheses which have been proposed to explain tool
use in primate populations: the necessity and the opportunity hypotheses. According to the first
hypothesis (Fox et al., 2004), since tool use allows the access to food otherwise not available, it
should be more frequent when food resources are scarce (Boesch & Boesch-Achermann, 2000; Fox
et al., 2004; Moura e Lee, 2004). In contrast, the second hypothesis (Beck, 1980; Fox et al., 2004)
predicts that favourable local conditions enhance the emergence and the maintenance of innovative
behaviours, including tool use. Our second goal was to determine the frequencies of tool use in the
different sex-age classes in order to verify whether, as we predict on the basis of the high sexual
dimorphism and the energetic effort required to crack open nuts, male capuchins perform this
behaviour more frequently than females.
The study area (13 km2 ) is located at Fazenda Boa Vista in the north-eastern Brazilian state of
Piauí (9º 00’ S; 45º 00’ W), 21 km north-west of the town of Gilbués. The physical geography of
the field site is a sandy plain placed approximately 420 m above sea level. The climate is seasonally
dry (annual rainfall 1156 mm; total rainfall during dry season: April to September 230 mm; data
from 1971–1990, source: Embrapa).
This study was conducted on two well-habituated wild groups of C. libidinosus that were
followed from June 2006 to July 2007. Activity budget were recorded by group scan sampling and
the occurrence of tool use was scored by focal animal sampling (Martin & Bateson, 1993).
The result shows that tool use is a habitual behaviour for the capuchins (sensu Whiten et al.,
1999) since it is enacted repeatedly and regularly by several individuals of a group. Moreover, tool
use can be defined as traditional (as defined by Fragaszy & Perry, 2003) since (i) the use of stones
as percussor has been observed only in a limited number of populations of C. libidinosus (Ottoni &
Izar, 2008), even if hammers and encased fruits are present in many other study sites and (ii) this
behaviour appears to be transmitted via social influences (Resende et al., 2008; Silva, 2008).
Food resources in Boa Vista do not appear more scarce than in other sites where capuchins live
in which food abundance was evaluated with similar methodologies (i.e., Atlantic Forest, Izar,
2004; see also Nakai, 2007); consequently, access to resources fundamental for survival does not

4
seem to have caused the use of tools in our groups. The lack of a marked seasonality in food
availability did not allow to further test the necessity hypothesis (Fox et al., 2004; Moura e Lee,
2004). Our results are in agreement with the predictions of the opportunity hypothesis (Beck, 1980;
Fox et al., 2004) since capuchins use of tools reflects the availability of nuts.
As we expected, female capuchins use tools less frequently. This is in sharp contrast with what
has been described for nut cracking in chimpanzees (Boesch & Boesch, 1984, Lonsdorf et al.,
2004), and what is expected on the basis of the high energy requirements due to lactation and
gestation (Clutton-Brock, 1977). Since using tools for capuchins is a strenuous activity, it is
possible that it requires energetic efforts too elevated for females, whose body weight and force are
lower than males (Liu et al., 2009), to be beneficial.

5
1. INTRODUZIONE
1.1 Uso di strumenti
L’uso di strumenti è stato a lungo ritenuto un’acquisizione fondamentale nel processo di
ominazione, che ha contraddistinto il genere Homo permettendone lo sviluppo dell’intelligenza
(Isaac 1978; Washburn, 1978; Leakey 1980). L’uso di strumenti ha permesso all’uomo di
diversificare il proprio stile di vita e di sfruttare risorse non accessibili ad altre specie animali.
Utilizzare uno strumento permette di avere accesso a maggiori risorse e risolvere importanti
problemi ecologici, come ad esempio ottenere cibo e costruirsi un riparo. Servirsi di uno strumento
è dunque generalmente considerato più vantaggioso rispetto ad affrontare la stessa situazione senza
farne uso (Visalberghi & Fragaszy, 2006).
La definizione di uso di strumenti maggiormente adottata dai biologi è quella proposta da Beck
(1980) secondo la quale un animale utilizza uno strumento quando usa un oggetto esterno come
estensione funzionale del proprio corpo (bocca, becco, mano o artiglio) per il conseguimento di uno
scopo immediato. E’importante però anche sottolineare la differenza tra un comportamento
esplorativo e quello di uso di strumento vero e proprio; nel primo caso infatti un individuo può
arrivare alla scoperta casuale di un oggetto come strumento, mentre nel secondo caso il soggetto è
intenzionato ad usare uno strumento per raggiungere uno scopo ben preciso.
Fino ad oggi molti autori hanno descritto l’uso di strumenti nel regno animale e diversi studi
riguardano gli uccelli. Il caso più noto è quello del fringuello-picchio delle Galapagos (Cactospiza
pallida) che usa ramoscelli, spine di cactus o peduncoli di foglie per catturare insetti nei buchi degli
alberi o nelle fessure dove non può arrivare con il becco (Eibl-Eibesfeldt, 1961; Tebbich et al.,
2002). Il capovaccaio (Neophron percnopterus) invece, per aprire le uova di struzzo vi fa cadere
sopra un sasso tenuto nel becco (van Lawick-Goodall 1966, 1970), e sempre tra gli uccelli, anche i
corvi della Nuova Caledonia (Corvus moneduloides) fanno uso di bastoncini ad uncino e di steli per
cacciare insetti nei buchi dei tronchi (Hunt, 1996, 2000; Hunt & Gray, 2004).
Molte osservazioni riguardano anche i mammiferi. Hall & Schaller (1964) descrivono il
comportamento delle lontre marine (Enhydra lutris) della California, che spaccano il guscio di
alcuni molluschi battendoli su pietre piatte che hanno sistemato sul loro torace, mentre nuotano in
posizione dorsale. Da osservazioni di animali in cattività e studi in natura, sia gli elefanti asiatici
(Elephus maximus) sia gli africani (Loxondonta africana) sono in grado, grazie alla loro proboscide,
di afferrare e lanciare tronchi e sassi in contesti di aggressione, di usare pezzi di legno per grattarsi

6
parti del corpo e di spezzare rami e usarli per scacciare insetti parassiti (Beck, 1980; Chevalier-
Skolnikoff & Liska, 1993; Hart et al., 2001). Infine, tra i cetacei, i delfini (Tursiops sp.) della Baia
di Shark in Australia rivestono il loro rostro con delle spugne marine con cui riescono a stanare
alcuni pesci che vivono nei fondali (Parapercis sp.) (Mann et al, 2008; Smolker et al. 1997).
Tuttavia, l’uso di strumenti si manifesta in maniera alquanto rigida e solo in alcuni contesti
sia negli uccelli che nei mammiferi. Nei primati non-umani invece il comportamento di uso di
strumenti è più flessibile per forma e più ricco per funzioni (Tomasello & Call, 1997).
1.2 Uso di strumenti nei primati non umani
I primati non umani hanno cervelli con aree corticali particolarmente sviluppate ed elevate
capacità cognitive (Dunbar, 1992).
Le mani possiedono una morfologia che permette attività di foraggiamento particolari, con presa
ed estrazione di una notevole varietà di alimenti. Nonostante la destrezza manipolativa sia diversa
da specie a specie, gli adattamenti cognitivi e morfologici permettono una manipolazione degli
oggetti più flessibile nei Primati che negli altri ordini (Tomasello & Call, 1997).
Le proscimmie non usano strumenti, nonostante l’aye-aye (Daubentonia) mostri capacità
estrattive molto peculiari (Quinn & Wilson, 2004). Questa specie possiede adattamenti morfologici
specializzati per l’alimentazione con dita molto allungate, ricurve e sottili. Il dito medio della mano,
particolarmente lungo, viene usato sia per catturare larve e insetti, presenti sotto la corteccia o nelle
fessure degli alberi, sia per estrarre le sostanze liquide (Erickson, 1995).
Tra i primati non antropomorfi dell’Africa e dell’Asia le osservazioni di uso di strumenti sono
alquanto rare e sporadiche, fatta eccezione per i pata (Erythrocebus patas), i macachi (Macaca sp.)
e i babbuini (genere Papio) osservati lanciare oggetti durante comportamenti aggressivi, sia in
condizioni di cattività sia in ambienti naturali (per una rassegna vedi Beck, 1980).
Recentemente alcuni macachi (Macaca fascicularis) che vivono nel Parco Nazionale di Leam
Son a sud della Tailandia sono stati osservati usare dei sassi per colpire ostriche, gasteropodi e
bivalvi (Malaivijitnond, 2007). I babbuini neri (Papio ursinus) allo stato libero usano bastoncini per
stanare insetti (Kortland & Kooij, 1963) e sassi per colpire frutti dal guscio duro (Marais, 1969)
mentre in cattività sono in grado di usare strumenti per raggiungere oggetti fuori dalla loro portata
Beck, 1980). Studi più recenti hanno descritto nei macachi giapponesi (Macaca fuscata) l’attività di
manipolazione delle pietre (stone-handling), una forma di gioco nel quale le pietre vengono battute
contro superfici dure, battute una contro l’altra o raccolte e impilate una sull’altra (Huffmann, 1984,
Huffmann & Quiatt, 1986). Il comportamento, non direttamente riconducibile alla sopravvivenza

7
degli animali, sembra avere un ruolo importante nello sviluppo motorio dei cuccioli e nel rallentare
il processo di invecchiamento delle funzioni cognitive degli individui più anziani (Nahallage &
Hoffman, 2007).
In cattività tutte le quattro specie di scimmie antropomorfe imparano spontaneamente ad usare
strumenti e quando risolvono questo tipo di compiti hanno prestazioni simili (Tomasello & Call,
1997), ciononostante in natura l’uso di strumenti è stato osservato solo in alcune di esse.
Tra le antropomorfe lo scimpanzé è la specie che mostra la maggior complessità e variabilità
nell’usare strumenti (McGrew, 1992; Whiten et al., 1999). Le prime documentazioni risalgono agli
anni ’60, quando Jane Goodall osservò e riportò per la prima volta negli scimpanzé (Pan
troglodytes) di Gombe in Tanzania, il comportamento di raccolta delle formiche scavatrici (ant
dipping) (Dorylus nigricans) che vivono in colonie affollate in nidi sotterranei da cui escono alla
ricerca di cibo (Goodall, 1963). In seguito McGrew (1974) riferisce il comportamento più
dettagliatamente, descrivendo che lo scimpanzé, una volta localizzato il nido delle formiche, estrae
manate di terra, provocando così una intensa reazione aggressiva da parte delle formiche soldato. A
questo punto la scimmia sceglie un ramo e lo modifica per infilarlo nel nido sotterraneo e poi lo
estrae rapidamente riuscendo in questo modo a catturare e mangiare le diverse centinaia di formiche
che si sono accalcate attorno al bastoncino.
Oltre alla raccolta delle formiche gli scimpanzé si dedicano alla “pesca” di termiti
(Macrotermes sp.) all’interno dei loro nidi (termite fishing), un’attività che consiste nell’ infilare nel
termitaio fili d’erba e ramoscelli. Non appena le termiti soldato afferrano la sonda con le mandibole,
gli scimpanzé la estraggono dal cunicolo e mangiano direttamente le termiti che vi sono rimaste
attaccate (van Lawick-Goodall, 1970; McGrew, 1992). Gli scimpanzé usano anche foglie come
spugne per assorbire acqua altrimenti irraggiungibile con la bocca (van Lawick-Goodall, 1970), e
alcune popolazioni aprono noci di palma con percussori di pietra o di legno. Qest’ultimo
comportamento è stato osservato solo in popolazioni ad est del fiume Sassandra in Costa d’Avorio
(Boesch & Boesch, 1983; Matsuzawa, 1994). Inoltre in Sierra Leone, Alp (1997) scrive di
scimpanzé che si servono di frasche per appoggiarsi o sedersi e proteggersi così dalle spine mentre
mangiano le foglie e i fiori negli alberi di Ceiba petranda. Recentemente Pruetz & Bertolani (2007)
hanno osservato che scimpanzé di savana catturano proscimmie notturne (Galago senegalensis) che
dormono nelle cavità degli alberi infilzandole con bastoni. Gli autori riferiscono inoltre, che gli
scimpanzé modificano questi bastoni per renderli idonei ad essere infilati nelle cavità degli alberi.
Nonostante i gorilla (Gorilla gorilla) in cattività siano capaci di risolvere molti esperimenti di
uso di strumenti (Tomasello & Call, 1997), la prima osservazione in natura risale solo a pochi anni
fa. Breuer e colleghi (2005) hanno osservato una femmina adulta di gorilla (G. g. gorilla), nella

8
foresta inondata del Parco Nazionale di Nouabalé-Ndoki in Repubblica del Congo, adoperare un
bastone come sostegno per attraversare un fiume, e una femmina adulta di un altro gruppo staccare
un ramo e usarlo per aumentare la stabilità mentre mangiava in una salina. Nei gorilla l’uso di
strumenti potrebbe essere messo in atto non tanto per estrarre il cibo, ma piuttosto per risolvere
problemi di altro tipo, come ad esempio attraversare uno specchio d’acqua in cui potrebbero
affogare se fosse troppo profondo.
Anche per l’orango (Pongo pygmaeus), nonostante questa specie possieda le capacità cognitive
per realizzare e usare strumenti (Park & Gibson, 1977), la maggior parte delle osservazioni si
riferiscono ad animali tenuti in cattività o in centri di riabilitazione (Byrne, 1995; Russon &
Galdikas, 1993), mentre gli episodi di uso di strumenti in oranghi selvatici sono molto rari e recenti
(Fox & bin’ Muhammad, 2002). Anche se agli inizi degli anni ’80 la Galdikas (1982 a,b) osservò
negli oranghi del Borneo l’uso di foglie per coprirsi dalla pioggia e per pulirsi, è solo verso la metà
degli anni ’90 che van Schaik e colleghi osservarono sistematicamente l’uso di strumenti negli
oranghi di Sumatra (P.p. abelii) in Indonesia. Il comportamento osservato consiste nello sbucciare
bastoncini con entrambe le mani ed estrarre insetti e miele dalle cavità degli alberi o nell’estrazione
dei semi di Neesia (van Schaik et al., 1996; Fox et al., 1999). Altri due aneddoti sono stati descritti
nella stessa popolazione di oranghi: un individuo adulto si è servito di un ramo come gancio per
raggiungere un altro albero e un individuo giovane ha ammassato delle foglie sotto i piedi e sulle
mani per spostarsi sugli alberi spinosi della specie Erythrina (Fox e bin’ Muhammad, 2002). In ogni
modo è ancora poco chiaro se alcuni di questi comportamenti siano da considerarsi comuni (sensu
McGrew, 1992) oppure occasionali e limitati a pochi individui della popolazione.
Infine le osservazioni sull’uso di strumenti da parte di bonobo (Pan paniscus) in condizioni
naturali sono quasi assenti (Ingmanson, 1996), nonostante la vicinanza filogenetica con gli
scimpanzé. Anche questa specie in laboratorio usa strumenti con successo paragonabile a quello
delle altre antropomorfe (Savage-Rumbaugh & Lewin, 1994, Visalberghi et al., 1995, Schick et al.,
1999).

9
1.3 Biologia del genere Cebus
Flessibilità, opportunismo e adattabilità sono le caratteristiche del successo delle scimmie del
genere Cebus. Di corporatura robusta e taglia media, i cebi hanno un moderato dimorfismo
sessuale. In cattività il peso medio varia dai 2,5 (per i giovani e le femmine) ai 5 kg (per i maschi
adulti). Questi primati si distinguono tra le scimmie del Nuovo Mondo per avere (1) un alto livello
di encefalizzazione (alto valore del rapporto tra dimensione del cervello e dimensione del corpo),
(2) una durata della vita mediamente lunga (fino a 55 anni in cattività), (3) un lungo periodo di
svezzamento, e (4) un alto grado di tolleranza sociale tra gli individui (vedi Fragaszy, Visalberghi &
Fedigan, 2004a, per una rassegna completa del genere).
Tra le scimmie del Nuovo Mondo il genere Cebus ha un’ampia distribuzione geografica. Lo si
trova infatti in quasi tutta la regione neotropicale, dal Costa Rica fino in Argentina ad est delle
Ande, e dal Peru fino alle coste atlantiche del Brasile. Tale successo adattativo è il risultato
dell’elevata flessibilità e plasticità comportamentale di questo genere. Le otto specie in cui è stato
recentemente suddiviso (C. apella, C. nigritus, C. libidinosus, C. xanthosternos, C.capucinus, C.
olivaceus, C. kaapori, C. albifrons) (Groves, 2001), vivono in ambienti molto differenti, dalle
foreste pluviali a quelle tropicali, dalle regioni continentali semi-aride alle foreste costiere, dalle
aree umide ricoperte da mangrovie fino a quelle coltivate e alle zone suburbane degradate. I cebi
sono una specie opportunista dall’alimentazione tipicamente onnivora. La loro dieta infatti,
principalmente a base di frutta, viene arricchita da invertebrati (insetti, molluschi, vermi, larve e
pupe di insetti) e da altre parti vegetali (foglie, steli, fiori, radici) (Figura 1.1). Il consumo di queste
risorse alternative aumenta nei periodi di scarsità alimentare o nelle popolazioni che vivono in
ambienti meno ricchi di frutta (Terborgh, 1983; Brown et al., 1986; Brown & Zunino, 1990;
Spironello, 1991; Galetti & Pedroni, 1994; Peres 1994; Zhang, 1995). Occasionalmente i cebi
catturano piccoli vertebrati come uccelli e roditori (Verderane et al., 2007; Ferreira et al., 2002;
Resende et al., 2002; Galetti, 1990), e sono noti anche casi di predazione di primati di piccole
dimensioni (Sampaio & Ferrari, 2005).
Ciò che contraddistingue i cebi da altri primati sudamericani con dieta simile è il tipo di
comportamento alimentare. I cebi infatti sono definiti “foraggiatori estrattivi e distruttivi”, in quanto
il repertorio di comportamenti legati alla manipolazione del cibo include una varietà di azioni che
richiedono un certo sforzo come strappare, mordere, spezzare, battere, scavare, infilare, raschiare, e
perché mostrano un grande interesse per i cibi che devono essere estratti. Grazie a queste attitudini i
cebi ottengono ottimi risultati in problemi che richiedono una manipolazione complessa degli

10
oggetti (Westergaard & Fragaszy, 1987; Visalberghi, 1990) e sono, tra le scimmie del Nuovo
Mondo, le uniche ad usare strumenti sia in cattività sia in condizioni naturali.
Figura 1.1 Maschio adulto di C. libidinosus intento ad estrarre le larve all’interno di un nido di vespe (Foto N. Spagnoletti). Figure 1.1 Adult male of C. libidinosus extractingt larvae from a wasp nest (Photograph: N. Spagnoletti).
Figura 1.2 Boa Vista, Piauí, Brasile. Femmina di C. libidinosus che usa un sasso come percussore per aprire una noce di palma mentre trasporta il suo cucciolo di un mese (Foto N. Spagnoletti). Figure 1.2 Boa Vista, Piauí, Brazil. Female of C. libidinosus using a stone as hammer to crack-open a palm nuts while carrying her one month old infant (Photograph: N. Spagnoletti).

11
1.3.1 L’uso di strumenti nel genere Cebus Le scimmie Platirrine del genere Cebus sono le uniche a mostrare abilità nell’uso di strumenti
paragonabili a quelle degli scimpanzé (Visalberghi & McGrew, 1997). In cattività grazie alle ben
sviluppate capacità manipolative, i cebi imparano spontaneamente a risolvere una grande varietà di
compiti che necessitano l’uso di strumenti, dimostrandosi abili nel manipolare oggetti e in grado di
risolvere compiti come inserire bastoncini dentro tubi per estrarre una ricompensa, selezionare
oggetti appropriati per tirare a sé del cibo o aprire frutti con guscio battendoci sopra un oggetto
adeguato (Fragaszy & Adams-Curtis, 1991; Westergaard & Suomi, 1994; Visalberghi et al. 1995;
per una rassegna degli esperimenti di uso di strumenti si rimanda a Fragaszy et al., 2004a). Questo
tipo di uso di strumenti è definito di “primo ordine”, perchè mette in relazione l’uso di un solo
oggetto esterno al corpo per ottenere l’obiettivo preposto (estrarre del cibo, portarlo a sé, liberarlo
dal guscio). Nel caso delle noci o di altri frutti chiusi da guscio duro, quando il terreno (la superficie
sperimentale) è relativamente soffice, ma sono presenti superfici dure che possono servire come
incudini e oggetti adatti per colpire le noci, le scimmie devono produrre una relazione di “secondo
ordine” per riuscire ad aprire il frutto desiderato. In questo caso infatti è necessario prima di tutto
posizionare la noce su una superficie idonea come incudine (prima relazione) e poi colpire il frutto
con un oggetto duro (seconda relazione) controllando allo stesso tempo che la noce rimanga
sull’incudine dopo averla colpita (Matsuzawa et al., 2001).
In cattività, a circa 6 mesi i cebi iniziano a battere oggetti contro le superfici, e alcuni individui a
circa due anni imparano la corretta relazione spaziale degli oggetti che permette di avere successo
nei compiti richiesti (Anderson, 1990). E’ interessante notare il livello di variabilità individuale che
presenta questo comportamento: in genere solamente una piccola proporzione degli individui dello
stesso gruppo usa strumenti per risolvere compiti sperimentali, come aprire delle noci o infilare
bastoncini per estrarre cibo inaccessibile (Visalberghi, 1987; Fragaszy & Visalberghi, 1989).
Rispetto alle sopraccitate performance di cebi in condizioni di cattività, fino a qualche anno fa
l’uso sistematico di strumenti nelle popolazioni selvatiche si limitava a osservazioni piuttosto rare.
Agli inizi degli anni ’90, la letteratura riguardante l’utilizzo di strumenti era basata su osservazioni
sporadiche e poco chiare, inerenti a contesti di foraggiamento. Boinski (1988) in Costa Rica ha
osservato un individuo di C. capucinus attaccare un serpente velenoso (Bothrops asper) colpendolo
con un ramo e la Chevalier-Skolnikoff (1990) sempre in Costa Rica, ha osservato un giovane
maschio infilare un bastoncino in un buco, presumibilmente per estrarre insetti. Durante un
sopralluogo in canoa lungo un mangrovieto nello Stato del Maranhão (Brasile), Fernandes (1991)
vede un maschio adulto di C. apella che apre un’ostrica battendoci sopra un altro oggetto,
probabilmente la conchiglia di un'altra ostrica. Phillips (1998) osserva alcuni individui di C.

12
albifrons usare foglie per estrarre l’acqua da una cavità di un albero, comportamento questo, simile
allo sponging-water-with-leaves osservato da Goodall (1986) negli scimpanzé.
Tecniche di processamento del cibo alquanto complesse avvengono quando cibo incapsulato
deve essere estratto, come avviene per il midollo dalle palme Euterpe edulis da parte di C. nigritus
in foresta atlantica (Taira et al., 2002), per le larve dai fusti di Bambù in C. olivaceus (Gunst et al.,
2008) e per l’endosperma dalle noci di palma Astrocaryum chambira in C. apella della foresta
pluviale colombiana (Izawa & Mizuno, 1977). In quest’ultimo caso i cebi adottano diverse strategie
secondo lo stato di maturazione del frutto. Quando il frutto è acerbo, ma al suo interno sono già
presenti sia il succo sia il latte di cocco, le scimmie raccolgono le noci e le battono solitamente
contro i tronchi di Bambusa guadua. Dopo numerosi colpi, la buccia esterna (esocarpo) si spacca e i
cebi sbucciandola con i denti o con le mani accedono al poro germinale dal quale bevono il succo
del cocco. Successivamente continuano a battere la noce contro il tronco fino a che anche il latte più
denso fuoriesce dal poro germinale talvolta appiccicandosi al tronco dal quale poi viene leccato.
Quando invece il frutto è più maturo, il latte è solidificato e la noce è più coriacea, i cebi la battono
con forza contro una superficie dura (ad es. un tronco), fino a rompere i rivestimenti
dell’endosperma (esocarpo, mesocarpo, endocarpo). A quel punto con i denti e/o con le dita i cebi
estraggono l’endosperma solidificato (Izawa & Mizuno, 1977). Nella stessa popolazione di C.
apella in Colombia alcuni individui riuscivano ad aprire le noci dell’Astrocaryum chambira
battendone una contro l’altra (Struhsaker & Leland, 1977).
Nonostante le forme di manipolazione sopra descritte dimostrino la grande abilità manipolativa
anche per i cebi in natura, nessuno di questi comportamenti rientra nella definizione di uso di
strumenti data da Beck (paragrafo 1.1). Secondo Visalberghi (1993) la differente frequenza con cui
l’uso di strumenti è stata osservata nei cebi in cattività e in natura, e le differenze riscontrate tra cebi
e scimmie antropomorfe, dipendono da una serie di fattori. Innanzitutto, i cebi in natura non sono
ampiamente studiati come le scimmie antropomorfe (in particolare gli scimpanzé). Inoltre il loro
stile di vita più arboricolo rispetto agli scimpanzé potrebbe limitare le loro opportunità di usare
strumenti e rendere più difficoltoso il compito degli osservatori: attività che avvengono a terra,
come l’uso di strumenti riportato negli scimpanzé, sono più facilmente osservabili rispetto a quelle
che avvengono nel folto degli alberi (Fragaszy et al, 2004a). Infine, sugli alberi gli arti anteriori
sono spesso adoperati per sorreggersi e gli strumenti sono meno disponibili e più difficili da
manipolare che a terra; è più facile battere una noce su un’incudine a terra che su un ramo
orizzontale di un albero e in quest’ultimo caso si corre facilmente il rischio di perdere noce, o
strumento, nel corso dell’azione. Tuttavia, la vita arboricola non preclude necessariamente l’utilizzo

13
di strumenti (Fragaszy et al., 2004a), dato che sia gli scimpanzé sia gli oranghi usano strumenti
sugli alberi (van Schaik et al., 1996; Boesch & Boesch-Achermann, 2000; van Schaik, 2003).
Fino ad oggi, tra le scimmie del genere Cebus, C. libidinosus è l’unica osservata fare uso
sistematico di strumenti in natura. I comportamenti descritti sono di 3 tipi: uso di strumenti (1) per
aprire noci e semi racchiusi da gusci duri in una maniera simile agli scimpanzé della Costa d’Avorio
(Fragaszy et al., 2004b; Waga et al., 2006), (2) per scavare nel terreno alla ricerca di tuberi e radici,
e (3) per sondare con bastoncini le cavità di alberi o di anfratti rocciosi per estrarre insetti, miele o
acqua (Moura & Lee 2004; Mannu & Ottoni, 2009).
1.4 Fattori che determinano l’uso di strumenti
La rottura di noci con incudine e percussore è un uso di strumenti particolarmente complesso
dal punto di vista cognitivo poiché richiede l’apprezzamento di due relazioni spaziali, una tra noce e
incudine l’altra tra sasso e noce (Matsuzawa et al., 2001; Fragaszy et al., 2004a). Fino alle recenti
osservazioni compiute sui cebi, questo comportamento era stato riportato in natura solo nello
scimpanzé, e in particolare nella sottospecie Pan troglodytes verus che vive nell’Africa occidentale
ad ovest del fiume Sassandra-Izo (Sugiyama, 1993; Boesch & Boesch-Achermann, 2000;
Matsuzawa et al., 2001) ma non nelle popolazioni che vivono ad est dello stesso fiume o altrove nel
continente africano, pur non mancando in quest’area incudini, percussori e frutti duri da rompere
(Whiten et al., 1999).
L’uso di incudini e percussori rende accessibile una risorsa altrimenti inaccessibile e durante la
stagione in cui le risorse alimentari sono scarse comporta un immediato vantaggio per la
sopravvivenza (Boesch & Boesch-Achermann, 2000). Sulla base dei risultati del progetto di ricerca
a lungo termine sugli scimpanzè del Parco di Taï in Costa D’Avorio, Boesch & Boesch-Achermann
(2000) affermano che la varietà di comportamenti di uso di strumenti osservato in popolazioni
naturali dipende dalle condizioni ecologiche in cui gli animali vivono: secondo questi autori quando
il cibo è scarso, e ottenerne a sufficienza richiede tecniche di foraggiamento complesse, il repertorio
di uso di strumenti risulta più ampio. Pertanto, la chiave esplicativa delle differenze, talvolta anche
piuttosto marcate, nel numero di diverse tipologie di uso di strumenti, potrebbe risiedere
nell’ecologia delle diverse popolazioni, e in particolare nell’abbondanza di risorse alimentari
disponibili (Yamakoshi, 1998; Whiten, 1999; Boesch & Boesch-Achermann, 2000). Anche Moura e
Lee (2004) interpretano l’uso di strumenti per scavare ed estrarre tuberi dal terreno, o per avere
accesso a cibo altrimenti inaccessibile, come una risposta alle condizioni ecologiche di scarsità
alimentare che spingono Cebus libidinosus a usare strategie alternative per sopravvivere.

14
Van Schaik, Deaner e Merrill (1999) ritengono invece che condizioni ambientali appropriate
sono necessarie, ma non sufficienti, per spiegare la distribuzione dell’uso di strumenti nei primati in
natura, e sottolineano la necessità di identificare anche le caratteristiche socio-ecologiche che
favoriscono l’emergenza di questo comportamento. Nel favorire l’emergenza e la diffusione
dell’uso di strumenti in popolazioni di primati non umani, giocano un ruolo importante i fattori
come una spiccata abilità manipolativa, la capacità di imitazione e di innovazione, un elevato livello
di tolleranza sociale, l’interesse per cibi il cui accesso necessita estrazione (che ovviamente devono
essere presenti nell’ambiente) e le influenze sociali.
L’unica specie di primati del Nuovo Mondo che possiede le caratteristiche sopra descritte, e per
la quale è possibile verificare l’ipotesi di van Schaik e collaboratori (1999) è Cebus libidinosus.
1.5 Fattori ecologici e uso di strumenti
Esistono 2 principali ipotesi che hanno tentato di spiegare come i fattori ecologici possano
influenzare la frequenza e l’emergenza dell’uso di strumenti nei primati non-umani:
(1) L’ipotesi della necessità (Fox et al., 2004), secondo cui la scarsità di risorse primarie può
forzare alcuni membri di un gruppo ad esplorare nuovi cibi che richiedono la messa a punto di
nuove tecniche di trattamento del cibo (food processing), incluso l’uso di strumenti. Sebbene non
esistano evidenze a tale proposito, sappiamo che la variazione stagionale nell’abbondanza di risorse
primarie è inversamente correlato con il comportamento di rottura delle noci di palma osservato
negli scimpanzè di Bossou (Yamakoshi, 1998). Questa ipotesi è stata anche utilizzata da Moura &
Lee (2004) per spiegare il comportamento di uso di strumenti dei cebi del Parco Nazionale di Serra
da Capivara nel Piauí. Secondo questi autori l’ambiente di caatinga, in cui le risorse alimentari sono
molto scarse soprattutto durante la stagione secca, è il fattore chiave che ha spinto i cebi all’utilizzo
di strumenti per accedere a frutti dal guscio duro e per scavare ed estrarre tuberi dal terreno.
(2) L’ipotesi delle opportunità offerte dall’ambiente (Beck, 1980; Fox et al., 2004) secondo cui
l’emergenza di comportamenti innovativi sono favoriti da opportune condizioni ambientali.
Secondo Leica et al. (2008) è la disponibilità di pietre che ha favorito l’emergenza del
comportamento di manipolazione delle pietre (stone-handling) in alcuni gruppi di macachi
giapponesi (ma non in altri) che potrebbe a lungo termine favorire l’uso di strumenti. La rottura di
noci con percussori osservata negli scimpanzé della foresta di Taï potrebbe essere dovuta alla
presenza di sassi e di superfici idonee per schiacciare le noci (es. le radici piatte), e alla disponibilità
delle noci (Boesch & Boesch, 1984). Tuttavia queste condizioni esistono anche in altri siti in cui gli
scimpanzé sono stati studiati per decenni, senza che sia mai stato osservato questo comportamento
di uso di strumenti.

15
Infine alcuni autori (Beck, 1980; Visalberghi, 1987; Visalberghi et al., 2005) hanno suggerito
che per l’acquisizione dell’uso di strumenti da parte dei cebi, potrebbe essere fondamentale il
trascorrere parte del tempo a terra dove le opportunità di scoprire l’uso di strumenti sono maggiori
che sugli alberi. A terra infatti le possibilità di entrare in contatto con sassi e di esplorarne le
potenzialità sono maggiori. Questi autori fanno notare anche che i gruppi di cebi in cui è stato
osservato uso di strumenti, siano essi in cattività o in condizioni naturali, trascorrono molto tempo a
terra e che per quelli in cattività non esistono periodi in cui il cibo è particolarmente scarso, durante
i quali, secondo l’ipotesi di Moura & Lee (2004), i cebi dovrebbero scoprire l’uso di strumenti. Il
grado di terrestrialità delle popolazioni dipende dal tipo di vegetazione in cui vivono gli animali e
dalla distribuzione delle risorse. In biomi secchi, come la caatinga e il cerrado, dove il substrato
arboreo è basso e lo spazio tra gli alberi relativamente grande, è possibile, anche se mancano dati
precisi a riguardo, che gli animali trascorrano più tempo a terra rispetto a quanto fanno le specie che
vivono nelle foreste tropicali.

16

17
2. OBIETTIVI DELLA RICERCA
2.1 L’uso di strumenti in Cebus libidinosus
La specie Cebus libidinosus (Spix, 1823), presa in esame in questo lavoro, ha il pelo castano
chiaro con tonalità fino al giallo che varia nelle diverse parti del corpo. Questa specie fa parte del
gruppo di cebi “con i cornetti” in quanto gli adulti, specialmente le femmine, hanno sulla testa due
ciuffi di pelo scuro che si sviluppano con la maturità sessuale e aumentano di dimensione con l’età.
Le scimmie di questa specie occupano la parte centrale della distribuzione geografica del genere
Cebus, e vivono sia in zone di foresta Atlantica, sia in ambienti semi-aridi di Caatinga e Cerrado
(Silva Júnior, 2001). In particolare le troviamo dalle regioni del Nordest del Brasile (a nord del
fiume São Francisco) alle zone centrali del Brasile, fino al Pantanal del Mato Grosso (Fragaszy, et
al., 2004a, Figura 1.3).
Figura 2.1 Distribuzione di Cebus libidinosus (da Fragaszy,Visalberghi & Fedigan, 2004, p. 21). Figure 2.1 Distribution of Cebus libidinosus (see in Fragaszy,Visalberghi & Fedigan, 2004, p. 21).
Gran parte delle osservazioni sull’uso di strumenti da parte di popolazioni selvatiche di cebi, si
riferiscono a gruppi di C. libidinosus che vivono in zone di cerrado e caatinga, ambienti semi-aridi
simili alle savane tropicali e alle foreste secche, sottoposti a condizioni climatiche difficili per il
prolungato periodo di siccità. Le prime evidenze indirette di uso di pietre per aprire noci di palma
risalgono al 1997. Durante una perlustrazione nello Stato di Paraíba (Brasile), che aveva lo scopo di
valutare lo stato di salute della popolazione locale di cebi, furono rinvenute su una superficie piatta

18
e rocciosa delle noci rotte e un sasso. Dopo aver udito dei rumori simili a quelli di una pietra battuta
contro una superficie dura e più tardi incontrato un gruppo di cebi, Langguth & Alonso (1997)
ipotizzano che siano stati i cebi ad aver usato le rocce come incudine e le pietre come martelli per
schiacciare le noci. L’uso spontaneo di pietre per aprire noci di Syagrus e aver accesso sia
all’endosperma sia alle larve parassite delle noci, è stato anche descritto in seguito in gruppi di C.
apella semi-liberi (Rocha et al., 1998; Ottoni & Mannu, 2001).
Le prime osservazioni di tale comportamento in natura sono state descritte piuttosto
recentemente in C. libidinosus in ambienti semi-aridi di caatinga e cerrado (Fragaszy et al., 2004b;
Moura & Lee, 2004; Waga et al., 2006). A Boa Vista, nello Stato del Piauí in Brasile, è stato
osservato un gruppo di cebi utilizzare sassi e incudini (di legno o pietra) per rompere noci di palma
dal guscio molto duro (Fragaszy et al., 2004b) (Figura 1.2) e sono stati descritti numerosi “siti
incudine” abitualmente utilizzati da gruppi di cebi che abitano nella zona (Visalberghi et al., 2007).
L’accesso a cibi incapsulati per mezzo di pietre è stato anche osservato, insieme ad altre forme di
uso di strumenti, nel Parco Nazionale di Serra da Capivara, località distante circa 400 Km da Boa
Vista. Qui gruppi di C. libidinosus usano sassi per scavare nel terreno alla ricerca di tuberi e per
tagliare pezzi di legno alla ricerca di insetti e larve (Moura & Lee, 2004; Mannu & Ottoni, 2009).
Alcuni individui usano anche bastoncini come sonde per cercare artropodi, esplorare nidi di insetti
sociali e estrarre miele dall’interno dei tronchi. In molti di questi episodi, prima di inserire il
bastoncino, i cebi lo modificano ripulendolo dalle foglie e assottigliandone la punta (Mannu &
Ottoni, 2009).
L’uso di pietre per aprire noci e frutti dal guscio duro è stato osservato anche nel cerrado del
Parco Nazionale di Brasilia. Nella zona del parco accessibile al pubblico (Agua Mineral) le scimmie
hanno la possibilità di interagire con i visitatori e approfittare di cibo di origine antropica. Qui i cebi
rompono le noci di Syagrus romanzoffiana utilizzando come incudine il pavimento di mattonelle e
usando sassi o mattonelle rotte come percussori (Sabbatini, 2006). Inoltre nelle zone più interne del
parco, i giovani usano strumenti per aprire frutti di Hymenea courbaril e Acrocomia aculeata,
analogamente a quanto osservato a Boa Vista (Waga et al., 2006).
Infine recentemente sono state pubblicate evidenze indirette (interviste a locali e ritrovamenti di
siti incudine secondo la definizione di Visalberghi et al., 2007) di uso di percussori e incudini per
aprire noci, in C. libidinosus che vive in ambiente di caatinga a nord del fiume San Francisco
(Alagoas, Brasile), e individuati siti di rottura a sud del fiume San Francisco in zone di caatinga e di
cerrado dove è presente C. xanthosternos (Canale et al., 2009).

19
In Appendice 1 viene riportata una mappa con i siti per i quali vi è evidenza diretta e indiretta
del comportamento di uso di strumenti nelle 4 specie di cebi dai cornetti (C. apella, C. libidinosus,
C. nigritus, C. xanthosternos) recentemente pubblicata da Ottoni & Izar (2008).
2.2 Ipotesi di ricerca e predizioni
Il primo scopo di questa ricerca è di indagare la relazione fra abbondanza delle risorse ed
utilizzo di strumenti ed in particolare di verificare l’ipotesi della necessità secondo la quale questo
comportamento tenda ad essere più frequente quando le risorse alimentari sono inferiori e l’ipotesi
dell’opportunità ambientale (vedi oltre). Nel nostro caso la forte stagionalità climatica presente
nell’aria di studio ed il fatto che uno dei gruppi di cebi studiati fosse approvvigionato creavano le
condizioni ideali per la verifica della prima ipotesi.
All’inizio della raccolta dati, due gruppi di Cebus libidinosus, erano abituati alla presenza dei
ricercatori. Il gruppo Chicao riceveva da alcuni anni un supplemento di cibo e acqua, mentre il
gruppo Zangado non lo ha mai ricevuto. Questa differenza ci ha offerto l’opportunità di comparare
il comportamento di due gruppi che vivono nello stesso ambiente di cui uno parzialmente
approvvigionato dall’uomo e dunque potenzialmente sottoposto a disponibilità di risorse alimentari
alternative.
Inoltre l’ambiente semi-arido di Boa Vista, dove si è svolta la nostra ricerca, si trova al confine
tra le vegetazioni di cerrado e caatinga, e presenta un clima fortemente stagionale, con una stagione
piovosa che va da ottobre a maggio e una stagione secca da giugno a settembre. Pertanto il tipo di
ambiente e la disponibilità di risorse alimentari nel corso dell’anno dovrebbero influenzare il modo
con cui gli animali usano le risorse incluso l’uso di strumenti.
Secondo l’ipotesi della necessità (Fox et al., 2004; Moura e Lee, 2004), quando le risorse sono
scarse il repertorio di uso di strumenti è maggiore o di più tipi (Boesch & Boesch-Achermann,
2000). In base a tale ipotesi ci aspettiamo che a Boa Vista durante la stagione in cui le altre risorse
alimentari scarseggiano: (1) entrambi i gruppi di cebi usino di più gli strumenti e (2) che ciò
avvenga in misura maggiore per il gruppo che non riceve cibo supplementare.
Al contrario, secondo l’ipotesi dell’opportunità ambientale (Fox et al., 2004; Beck, 1980),
condizioni locali opportune, come la presenza di pietre e di frutti incapsulati, favoriscono
comportamenti innovativi come l’uso di strumenti. In base a tale ipotesi ci aspettiamo che a Boa
Vista (3) il comportamento di uso di strumenti da parte di entrambi i gruppi di cebi rifletta la
disponibilità delle noci di palma e di altri frutti incapsulati.

20
Secondo l’ipotesi della terrestrialità (Visalberghi, 1987; Visalberghi et al., 2005), il tempo
trascorso a terra è il fattore chiave per l’emergere dell’uso di strumenti nei cebi, poiché aumenta la
probabilità individuale di manipolare e combinare sassi, incudini e noci. A conferma di questa
ipotesi ci aspettiamo che (4) i cebi di Boa Vista trascorrano in proporzione maggior tempo a terra
rispetto alle popolazioni di cebi che vivono in località dove non è stato osservato il comportamento
di uso di strumenti. �
Il secondo scopo della ricerca è di indagare le differenze fra classi di età e sesso nei budget di
attività e nell’uso di strumenti. Secondo van Schaik et al. (2003) il tasso di uso di strumenti varia in
relazione al sesso e all’età degli individui. La presenza nei due gruppi di individui di ambo i sessi e
di differenti età ci permette di determinare la frequenza di uso di strumenti in relazione a queste
variabili. Inoltre la determinazione dei budget di attività nelle varie classi di età/sesso ci permette di
comparare i cebi da noi studiati con quelli di altre popolazioni che non usano strumenti.
Dalle ricerche condotte da Boesch & Boesch (1984; 2000) sull’uso di percussori da parte di
scimpanzé risulta che gli adulti usano strumenti più frequentemente e che ottengono maggior
successo dei giovani. Inoltre le femmine adulte usano strumenti più frequentemente e con maggiore
abilità dei maschi adulti. E’ possibile che (1) il comportamento dei cebi sia analogo a quello degli
scimpanzé (Ipotesi 1), o che (2) l’elevata resistenza alla frattura delle noci (Visalberghi et al., 2008)
e il peso dei percussori (in media 1 kg, Visalberghi et al., 2007) rendano lo sforzo di sollevare il
sasso e colpire la noce maggiore per i giovani e le femmine adulte (le cui dimensioni corporee sono
inferiori a quelle dei maschi adulti (Ford and Davies, 1992) e che di conseguenza questi ultimi
usino strumenti con maggiore frequenza (Ipotesi 2).
2.3 Obiettivi della ricerca
Lo scopo di questo progetto è determinare la frequenza dell’uso di strumenti nella popolazione
selvatica di Cebus libidinosus a Boa Vista in relazione ai cambiamenti stagionali nella disponibilità
di risorse alimentari e descrivere il comportamento di uso di strumenti nelle varie classi di età e
sesso. In particolare gli obiettivi di questo studio sono:
1) esaminare i pattern di attività (mensili e annuale), incluso l’uso di strumenti e la scelta
del substrato (arboreo/non arboreo) in cui svolgere tali attività, in due gruppi di cebi
che vivono nello stesso habitat, ma di cui uno solo riceve cibo supplementare;

21
1a) verificare la validità dell’ipotesi della necessità (Fox et al., 2004; Moura e Lee, 2004,
Boesch & Boesch-Achermann, 2000), secondo la quale quando le risorse sono scarse
la frequenza di uso di strumenti è maggiore e/o include più tipi di uso di strumenti;
1b) discutere sulla base dei nostri dati e di quelli disponibili in letteratura per il genere Cebus
la validità dell’ipotesi dell’opportunità ambientale (Fox et al., 2004; Beck, 1980),
secondo la quale condizioni locali opportune favoriscono comportamenti innovativi
come l’uso di strumenti, tra cui il livello di terrestrialità proposto da Visalberghi
(1987) per il genere Cebus.
2) verificare possibili relazioni tra pattern di attività e variazione stagionale di dati climatici
e risorse trofiche;
3) analizzare i pattern di attività, incluso l’uso di strumenti e la selezione tra substrato
arboreo/non arboreo, tra le classi di età e sesso;
4) comparare il successo e l’efficenza nell’uso di strumenti tra le classi di età e sesso;

22

23
3. AREA DI STUDIO
3.1 Boa Vista
Boa Vista è una regione che si estende per circa 110 km2 (Hinely, 2006) all’interno del
municipio di Gilbués (9º 00’ Sud; 45º 00’ Ovest), nel sud-est dello stato del Piauí in Brasile (Figura
3.1). L’area di studio, di circa 13 km2, si trova all’interno di questa regione e comprende la Fazenda
di proprietà della famiglia de Oliveira. Dal 2005 la Fazenda de Oliveira è sede del progetto di
ricerca EthoCebus, nato dalla collaborazione dell’Università di San Paolo (USP-BR) e l’Università
della Georgia (UGA-USA) con l’Istituto di Scienze e Tecnologie della Cognizione di Roma (ISTC-
IT).
Figura 3.1 L’area di studio. A destra immagine satellitare in cui sono visibili i morros al centro delle zone pianeggianti. I nomi indicano la parte bassa delle pareti dei morros.
Figure 3.1 Study area. On the right, satellite image showing the morros in the center of the valley. The names indicate the bottom part of the morros.
La regione si trova in una zona di ecotono tra le vegetazioni cerrado e caatinga, due biomi tipici
delle zone semiaride del Brasile. Molto simile ad una savana alberata, il cerrado è per estensione il
secondo bioma brasiliano dopo l’Amazzonia ed è caratterizzato da un mosaico di formazioni
Boa Vista (9°S, 45°O)
Gilbués
Brasile
Piauí

24
vegetazionali tra cui erbacee perenni con arbusti di 2-3 m e campi aperti con alberi che possono
raggiungere i 30 m (de Souza Aguiar et al., 2004). Inoltre sono presenti corsi d’acqua permanenti e
torrenti che si prosciugano durante la stagione secca. Il territorio della regione è pianeggiante,
situato a circa 460 m s.l.m e localmente interrotto da rilievi (morros) costituiti da roccia
sedimentaria che si innalzano fino a 100 m e oltre (Hinely, 2006, figura 3.2). La copertura degli
alberi non è mai troppo densa e la loro altezza non supera i 12-15 m. La parte pianeggiante è
costituita dalla chapada caratterizzata dalla predominanza di alberi di media altezza come
Eschweilera nana (nome comune sapucaia), Caryocar brasiliense (pequi) e Hymenaea courbaril
(Jatobá) e dalla presenza di palme. La maggior parte delle specie di palme (Astrocaryum campestre,
Tucum; Attalea sp., catulè e catulì; Syagrus sp., patì; Orbygnia sp., piassava, e altre non ancora
identificate) mostra speciali adattamenti per evitare l’eccessiva traspirazione come un tronco corto e
sotterraneo e foglie e caschi di frutta molto vicini al suolo (Figura 3.3). Inoltre vi sono zone umide
chiamate brejo caratterizzate da una maggiore diversità di alberi che possono formare foreste a
galleria e dove sono presenti piccoli stagni e corsi d’acqua circondati dagli alberi della palma
Mauritia flexuosa (buriti). Nei morros di arenaria si possono distinguere due tipi di fisionomie
vegetazionali: lungo i costoni si trovano arbusti e alberi di media e piccola dimensione come la
Parkia pendula (faveira) e alcune specie della famiglia delle Euphorbiaceae (es. mandioca brava),
mentre sulla cima dei morros la vegetazione è prevalentemente erbacea e arbustiva con
Bromeliaceae e Cactaceae.
Figura 3.2 Panorama dell’area di studio; i morros si innalzano con ripide falesie dalla valle (foto di N. Spagnoletti).
Figure 3.2 Landscaoe in the study area; the morros steeply raised with cliff up from the valley (Photograph: N. Spagnoletti).
Sulla base delle precipitazioni è possibile definire due stagioni: una secca da maggio a settembre
(con precipitazioni medie mensili di 10 mm) e una piovosa da ottobre a aprile (con precipitazioni
medie mensili di 155 mm) (fonte: Embrapa, banca dati climatici del Brasile).
Fra i molti rettili presenti vanno ricordati Tupinambis sp. (teiù), Bothrops sp. (jararaca), Boa
constrictor (jibóia) e Micrurus sp. (cobra-coral). Tra gli uccelli Anhima cornuta (anhuma), Ara
chloropterus (arara-vermelha), Sarcoramphus papa (urubu-rei) e Dryocopus lineatus (pica-pau-de-

25
bandabranca) e tra i mammiferi Puma concolor (onça parda), Tamandua tetradactyla (tamanduá-
mirim) e Euphractus sexcinctus (tatu-peba). Tra i primati, oltre Cebus libidinosus (macaco-prego) ci
sono Alouatta caraya (bujìo) e Callithrix jacchus (saguì).
I cebi che vivono in questa regione si alimentano dei frutti di palma e usano strumenti per
accedere all’endosperma racchiuso in un endocarpo molto duro (Visalberghi et al., 2008, Figura
3.3). Inoltre i cebi si alimentano del mesocarpo della noce dopo avere sbucciato con i denti
l’esocarpo, e quando il frutto è immaturo sfruttano la polpa delle noci e l’endosperma sotto forma di
acqua di cocco. Occasionalmente possono cibarsi delle larve di coleottero che parassita queste noci.
Figura 3.3 A) pianta di palma della specie catulè (Attalea barreirensis). Si noti l’assenza del fusto e il casco di noci al livello del suolo. B) noce di catulè. Quando il frutto non è ancora maturo i cebi asportano con i denti l’esocarpo e l’endocarpo e bevono l’acqua contenuta al suo interno. C) sezione longitudinale di noce di palma (foto Noemi Spagnoletti, illustrazione Gloria Sabbatini).
Figure 3.3 A) example of catulè palm (Attalea barreirensis).Note that this species of palm is trunk less then palm nuts grown directly on the ground. B) walnut of catulè.Capuchins pick unripe fruits, remove exocarp and endocarp using their teeth and drink water inside. C) longitudinal section of palm nut (photographs: Noemi Spagnoletti; illustration: Gloria Sabbatini).
Nella fazenda della famiglia de Oliveira è presente un capanno di avvistamento (esconderijo)
posizionato di fronte ad alcuni “siti-incudine” frequentemente utilizzati dai cebi del gruppo Chicao.
All’esconderijo sono presenti varie pietre utilizzate come “percussori” e un tronco con diversi
blocchi di arenite utilizzati come “incudini” per aprire le noci di palma (per dettagli sulle incudini e
sui siti di rottura delle noci vedi Visalberghi et al., 2007). Dal 2001 sono stati distribuiti quasi
giornalmente frutta, verdura e noci di palme di fronte al capanno così da poter attirare le scimmie.

26
Ciò doveva servire per un piano di eco-turismo che prevedeva la collaborazione della famiglia de
Oliveira con la Fondazione BioBrasil e la Tropical Nature Inc.; il programma non è poi veramente
decollato ed è terminato nel 2007.
3.2 Il Progetto EthoCebus
Il presente progetto di dottorato si inserisce nel programma di ricerca internazionale EthoCebus
(http://www.ip.usp.br/ebottoni/EthoCebus/echome.html) nato in seguito alla prime osservazione di
uso di strumenti da parte dei cebi di Boa Vista (Fragaszy et al., 2004b). I suoi fondatori sono
Dorothy Fragaszy (Professore di psicologia all’Università della Georgia, USA), Patricia Izar e
Eduardo Ottoni (Professori al Dipartimento di Psicologia dell’Università di San Paolo, Brasile) e
Elisabetta Visalberghi (Dirigente di ricerca del Centro Primati dell’Istituto di Scienze e Tecnologie
della Cognizione, CNR di Roma). Dal 2005, grazie alla partecipazione di diversi studenti,
EthoCebus studia l’uso di strumenti attraverso la raccolta di dati demografici, ecologici e
comportamentali. Gli studi in corso riguardano gli aspetti ontogenetici (da Silva, 2008), sociali,
cognitivi (Visalberghi et al., 2009a,b) e biomeccanici (Liu et al., 2009) dell’uso di strumenti, e le
caratteristiche geografiche dell’habitat (Hinely et al., 2007). Inoltre sono stati effettuati censimenti
della fauna locale e dei siti incudini presenti nella zona (Visalberghi et al., 2007), e dall’inizio delle
ricerche vengono monitorate mensilmente le risorse alimentari.

27
4. METODI E MATERIALI
La raccolta dati di 12 mesi (giugno 2006-maggio 2007) è avvenuta dopo due sopralluoghi
nell’area di studio (giugno 2005 e febbraio-marzo 2006), mirati a verificare la fattibilità del progetto
di ricerca, iniziare l’abituazione dei cebi alla presenza dei ricercatori, formare assistenti e guide
locali e mettere a punto la metodologia per la raccolta dei dati comportamentali e ecologici.
4.1 I cebi di Boa Vista
Le osservazioni comportamentali sono state fatte su due gruppi di Cebus libidinosus, il “gruppo
Chicao” e il “gruppo Zangado” così chiamati dal nome del maschio dominante (alfa) presenti in
ciascun gruppo all’inizio della raccolta dati. Ogni gruppo è stato seguito dall’alba al tramonto per
circa 7-10 giorni al mese, per un totale di 12 mesi, e 1709 ore di osservazione diretta. L’ampiezza
del territorio dei due gruppi è di circa 5-7 km2 con un’area di sovrapposizione di circa 3 km2 (37 %)
(in Appendice 2 si riporta un’immagine con la dimensione dell’home range dei due gruppi e l’area
di sovrapposizione).
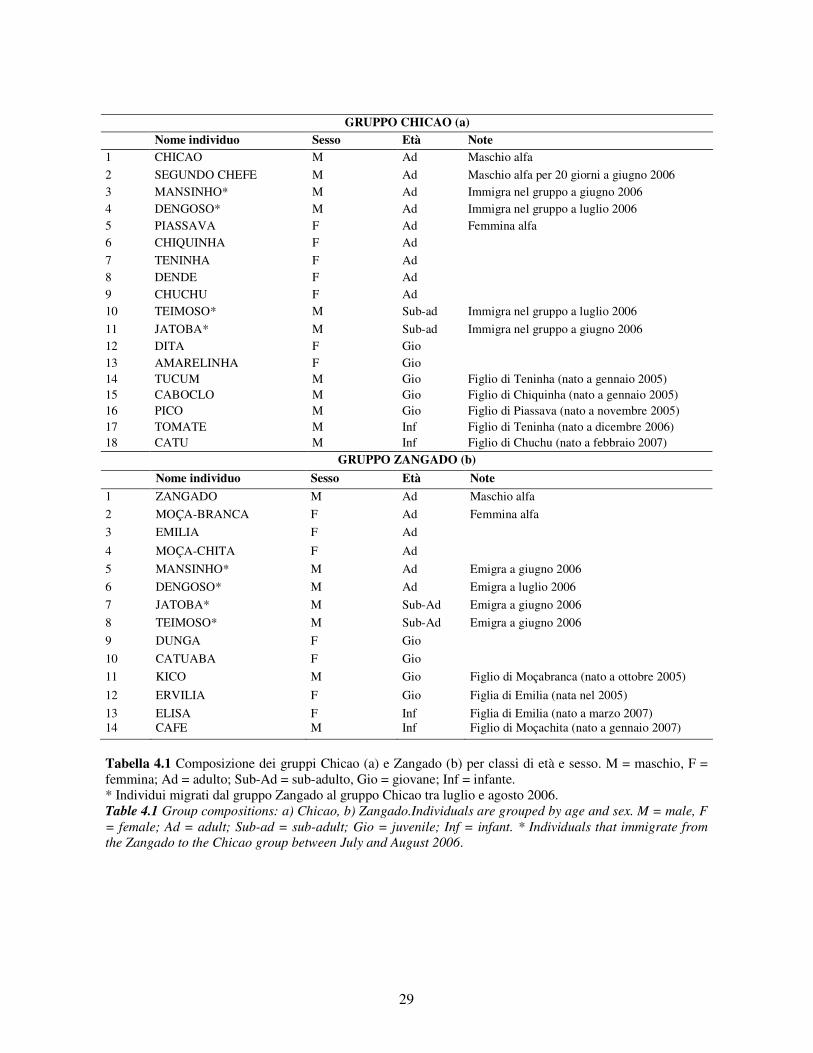
In Tabella 4.1 viene riportata la composizione dei due gruppi e il sesso e l’età stimata di ciascun
individuo. Analogamente a quanto fatto da Izawa (1980) abbiamo considerato le seguenti classi di
età.
• Infanti: individui dalla nascita fino allo svezzamento. Sebbene esista una grande variabilità
nell’età dello svezzamento, anche all’interno di uno stesso gruppo, lo svezzamento per il
genere Cebus in natura avviene a 16 ai 20 mesi (Fragaszy et al., 2004a). Gli individui di
questa età sono facilmente riconoscibili in base alle dimensioni e alla vicinanza con la
madre.
• Giovani: individui immaturi dallo svezzamento fino ai 3 anni per i maschi e fino al primo
estro (5-7 anni) per le femmine.
• Sub-adulti: maschi che non hanno ancora pienamente sviluppato le caratteristiche fisiche
dell’adulto in quanto pur avendone le dimensioni, non hanno i tipici ciuffi sulla testa. Età
stimata da 5 a 7 anni.
• Adulti: maschi e femmine che presentano tutte le caratteristiche fisiche tipiche degli adulti
quali ciuffi di pelo sulla testa ben sviluppate e basette ben accentuate nei maschi. In questa
categoria sono incluse le femmine che hanno avuto il primo estro e/o il primo piccolo. Età
stimata di circa 7 anni per i maschi, e 5 anni per le femmine.
Durante i mesi di studio la dimensione dei gruppi è variata (Chicao N = 12-18; Zangado N = 8-
12) a causa di differenti fattori quali: nascite (3 piccoli nel gruppo Chicao e 2 in quello Zangado),

28
emigrazione ed immigrazione (2 maschi adulti e 2 maschi sub-adulti sono emigrati dal gruppo
Zangado a quello di Chicao tra giugno e luglio 2006; a ottobre 2006 un maschio adulto sconosciuto
è arrivato nel gruppo Zangado rimanendovi quasi un mese intero).
Il gruppo Chicao è stato quotidianamente rifornito di cibo supplementare in prossimità di un
capanno di avvistamento (esconderijo). Tra le 7:00 e le 8:00 a.m. gli veniva distribuita frutta o
verdura (banane, anguria, mango, mais e canna da zucchero) per un totale di circa 3082 ± 908 Kcal
al giorno, circa 197 ± 56 Kcal al giorno per individuo (fonte INRAN: Istituto Nazionale di Ricerca
per gli Alimenti e la Nutrizione http://www.inran.it/).
29
GRUPPO CHICAO (a) Nome individuo Sesso Età Note 1 CHICAO M Ad Maschio alfa 2 SEGUNDO CHEFE M Ad Maschio alfa per 20 giorni a giugno 2006 3 MANSINHO* M Ad Immigra nel gruppo a giugno 2006 4 DENGOSO* M Ad Immigra nel gruppo a luglio 2006 5 PIASSAVA F Ad Femmina alfa 6 CHIQUINHA F Ad 7 TENINHA F Ad 8 DENDE F Ad 9 CHUCHU F Ad 10 TEIMOSO* M Sub-ad Immigra nel gruppo a luglio 2006 11 JATOBA* M Sub-ad Immigra nel gruppo a giugno 2006 12 DITA F Gio 13 AMARELINHA F Gio 14 TUCUM M Gio Figlio di Teninha (nato a gennaio 2005) 15 CABOCLO M Gio Figlio di Chiquinha (nato a gennaio 2005) 16 PICO M Gio Figlio di Piassava (nato a novembre 2005) 17 TOMATE M Inf Figlio di Teninha (nato a dicembre 2006) 18 CATU M Inf Figlio di Chuchu (nato a febbraio 2007)
GRUPPO ZANGADO (b) Nome individuo Sesso Età Note 1 ZANGADO M Ad Maschio alfa 2 MOÇA-BRANCA F Ad Femmina alfa 3 EMILIA F Ad
4 MOÇA-CHITA F Ad 5 MANSINHO* M Ad Emigra a giugno 2006 6 DENGOSO* M Ad Emigra a luglio 2006 7 JATOBA* M Sub-Ad Emigra a giugno 2006 8 TEIMOSO* M Sub-Ad Emigra a giugno 2006 9 DUNGA F Gio 10 CATUABA F Gio 11 KICO M Gio Figlio di Moçabranca (nato a ottobre 2005) 12 ERVILIA F Gio Figlia di Emilia (nata nel 2005) 13 ELISA F Inf Figlia di Emilia (nato a marzo 2007) 14 CAFE M Inf Figlio di Moçachita (nato a gennaio 2007)
Tabella 4.1 Composizione dei gruppi Chicao (a) e Zangado (b) per classi di età e sesso. M = maschio, F = femmina; Ad = adulto; Sub-Ad = sub-adulto, Gio = giovane; Inf = infante. * Individui migrati dal gruppo Zangado al gruppo Chicao tra luglio e agosto 2006. Table 4.1 Group compositions: a) Chicao, b) Zangado.Individuals are grouped by age and sex. M = male, F = female; Ad = adult; Sub-ad = sub-adult; Gio = juvenile; Inf = infant. * Individuals that immigrate from the Zangado to the Chicao group between July and August 2006.

30
4.2 Osservazioni comportamentali
4.2.1 Abituazione di animali selvatici alla presenza umana L’abituazione è il processo attraverso il quale animali selvatici imparano ad accettare
gradualmente la presenza dell’uomo considerandolo parte neutrale del proprio ambiente naturale
(Tutin & Fernandez, 1991). Come già verificato durante il lavoro di tesi (Spagnoletti, 2005),
l’abituazione è un passo fondamentale per poi procedere alle osservazioni di campo. Consiste
nell’avvicinare gruppi di animali selvatici e nel mantenere con loro un contatto a distanza.
Gradualmente, dopo le prime reazioni di paura, aggressione e curiosità, gli animali tendono ad
ignorare la presenza dell’uomo lasciandosi avvicinare e permettendo così di studiare il loro
comportamento nel loro ambiente naturale (Graziani, 2000).
Nell’area di studio, da settembre 2006, è iniziato il procedimento di abituazione di due gruppi di
cebi ad opera degli assistenti Mauro de Oliveira e Marino Junior de Oliveira. Dopo circa 6 mesi, e
comunque all’inizio della raccolta dati, i cebi si lasciavano avvicinare anche a pochi metri (3-10 m).
Ad eccezione delle volte in cui la visibilità era scarsa, a causa della vegetazione fitta lungo i pendii
delle falesie o all’interno dei canaloni, e delle volte in cui i gruppi si spostavano in cima alle
montagne, è stato possibile seguire gli animali durante tutto l’arco della giornata riuscendo ad
identificare gli individui e registrarne il comportamento. Ciò è stato fondamentale per consentirci di
osservare da vicino l’uso di strumenti.
4.2.2 Registrazione dei pattern di attività Il gruppo di cebi veniva raggiunto al mattino nel luogo dove era stato lasciato la sera precedente
e quindi seguito durante l’intera giornata. Le registrazioni comportamentali sono state effettuate
utilizzando un campionamento a scansione (group scan sampling, Martin & Bateson, 1993) di 10
minuti, seguito da un intervallo di 10 minuti, per un totale di tre scansioni per gruppo ogni ora. Per
ciascun individuo in vista durante lo scan sono state registrate i seguenti dati e comportamenti:
1. Giorno, ora, numero di scan.
2. Identità del soggetto.
3. Ambiente: brejo (pianura stagionalmente acquitrinosa), chapada (pianura secca), en-
costa (zona di transizione tra pianura e morro), grota (canalone), morro (morro vero e
proprio), top-morro (cima del morro; per la descrizione delle vegetazioni paragrafo 3.1 e
Figura 4.1).
4. Substrato: 1) arboreo: basso (0-3 m), medio (3,1-6 m), alto (> 6,1 m); 2) non arboreo:
suolo; roccia; caverna; incudine di arenaria, incudine di legno. Per il gruppo Chicao è

31
stato considerato anche il substrato “esconderijo” quando gli individui si trovavano sul
tetto del capanno di avvistamento.
5. Attività (vedi comportamenti e relative definizioni in Tabella 4.2). Per le attività di
alimentazione e di foraggiamento è stata indicata la parte dell’alimento coinvolta (frutta,
fiori, foglie, semi, radici, endosperma) e il tipo di risorsa (invertebrati, vertebrati).
Quando possibile sono state identificate le specie animali e vegetali mangiate.
6. Posizione geo-referenziata del gruppo (coordinate registrate con GPS).
I dati analizzati sono stati raccolti da me e da Michele P. Verderane, con l’assistenza di Jozemar
e Arizomar da Silva Oliveira. Le mie registrazioni sono state riportate su un foglio elettronico Excel
(Microsoft Office) con un computer palmare (HP iPaQ hx2400).
Tabella 4.2 Categorie comportamentali considerate. Table 4.2 Behavioral categories sampled in this study.
Comportamenti
Descrizione
Riposo Qualsiasi tipo di inattività, anche rimanere fermi, seduti o in posizione sdraiata, o dormire
Movimento Spostarsi nell’ambiente con qualsiasi tipo di locomozione.
Alimentazione Ingerire e masticare alimenti, o bere
Foraggiamento Ispezionare l’ambiente alla ricerca di cibo. Include la manipolazione, l’esplorazione e il trasporto del cibo
Uso di strumenti Usare sassi come percussori su incudini di legno o pietra per aprire semi e frutti con guscio
Attività sociali Pulizia del pelo, gioco, vocalizzazioni, corteggiamento, copula, allattamento, aggressioni e minacce
Altre attività
Tutti i comportamenti non inclusi nelle precedenti categorie comportamentali (ad es. interazioni tra gruppi o con specie diverse)

32
4.2.2.1 Analisi Statistiche Per le analisi dei dati normalmente distribuiti ho usato test parametrici. Per quelli non
normalmente distribuiti ho eseguito la trasformazione in arcoseno e ripetuto il test di normalità sui
residui quando possibile. Se anche dopo tale trasformazione i dati non sono risultati normalmente
distribuiti (Lilliefors test, p < 0,05), ho usato la statistica non parametrica.
Al fine di non sovrastimare alcune attività a scapito di altre, ho calcolato il time budget per
ciascun gruppo come suggerito da Clutton-Brock (1974). Questo metodo è considerato il più
appropriato quando il numero di individui osservati varia molto tra i singoli scan (Clutton-Brock,
1974). Inoltre, per ridurre il potenziale errore dovuto al differente numero di scan registrati al mese,
ho escluso dall’analisi quei giorni in cui il numero di scan è stato minore della metà della media
degli scan registrati durante i 12 mesi. A tale scopo ho calcolato la frequenza di ogni attività per
singolo scan in proporzione al numero totale di individui (# ID) osservati nelle differenti attività per
quello scan. Per esempio se nello scan n°1: 3 individui (ID) foraggiano, 2 ID riposano e 1 ID usa
strumenti, dato che il numero totale di ID registrati è uguale a 6, la frequenza dell’attività di
foraggiamento per quello scan è di 3/6 (corrispondente a 0,5), la frequenza dell’attività di riposo è
di 2/6 (corrispondente a 0,33) e la frequenza dell’attività di uso di strumenti è di 1/6 (corrispondente
a 0,17). Ciò fatto ho usato ogni singolo scan come punto indipendente per le successive analisi.
Quindi ho calcolato: 1) i pattern di attività giornaliera (distribuzione giornaliera delle attività)
sommando le proporzioni delle attività nei singoli scan e dividendoli per il totale degli scan
registrati per quel giorno; 2) i pattern di attività mensile (distribuzione mensile delle attività)
facendo per ciascun mese la media delle frequenze giornaliere sul totale dei giorni di osservazione
per quel mese; 3) i pattern di attività annuale (distribuzione annuale delle attività) facendo la media
dei pattern mensili. Per il gruppo Chicao, al fine di valutare la proporzione di tempo dedicata
all’uso di strumenti in condizioni naturali, ho escluso da questa analisi i giorni in cui sono state
collocate noci e/o cibo supplementari al capanno di avvistamento per attirarli e poterli fare
osservare dai visitatori o da altri ricercatori.
I budget di attività tra i due gruppi sono stati confrontati sulla base dei pattern mensili e
stagionali. In quest’ultimo caso ho paragonato la stagione piovosa di 5 mesi e quella secca di 7 mesi
(vedi paragrafo 3.1 per dettagli sul clima e sulla suddivisione dell’anno in stagioni sulla base di
Embrapa, banca dati climatici del Brasile).
I confronti tra le attività dei due gruppi sono stati fatti utilizzando il t-test per dati appaiati,
controllando la variazione delle differenti categorie comportamentali rispetto ai mesi. Per le
categorie “uso di strumenti” e “attività sociali”, che non seguono la distribuzione normale, ho usato
il test non parametrico di Wilcoxon per dati appaiati rispetto ai mesi. Per esaminare le relazioni tra

33
le proporzioni mensili di tempo che ciascun gruppo spende nelle singole categorie e la variazione
mensile della disponibilità di risorse (frutta, invertebrati, noci di palma), la precipitazione mensile e
la media delle temperature massime e minime, ho usato il test di correlazione di Spearman.
Per ciascun gruppo ho confrontato i budget di attività nelle due stagioni usando l’ANOVA a un
criterio di classificazione, usando come variabile indipendente la stagione (secca e piovosa).
Per confrontare la variabilità dei pattern di attività tra classi di età e sesso sono stati considerati
gli individui giovani (maschi e femmine), le femmine adulte e i maschi adulti di entrambi i gruppi.
Per comparare le classi di età e sesso sono stati esaminati insieme gli individui dei due gruppi e
applicato l’ANOVA a un criterio di classificazione, usando come variabile indipendente la classe di
età/sesso. Il test di Tukey HSD è stato usato per i confronti post-hoc tra medie mensili delle attività e classi
di età/sesso.
I confronti tra substrati sono stati fatti in base alle categorie “arboreo/non arboreo” definite in
4.2.2. Un ulteriore confronto è stato effettuato più in dettaglio tra le sottocategorie: incudine,
caverna, suolo e roccia per il substrato non arboreo; alberi ad altezze basse, medie e alte per il
substrato arboreo. Per i confronti tra i substrati usati (arboreo/non arboreo), ho usato il test non
parametrico di Wilcoxon per dati appaiati rispetto ai mesi. Per comparare il tempo trascorso nei
differenti substrati nelle stagioni ho applicato il test non parametrico di Mann-Whitney. Per le
differenze tra la scelta del substrato e le classi di età e sesso (maschi adulti, femmine adulte,
giovani) è stato applicato l'ANOVA per ranghi di Kruskal-Wallis considerando il tempo trascorso
sui vari substrati come variabile dipendente.
4.2.3 Registrazione dell’uso di strumenti I cebi sono soliti aprire le noci in particolari luoghi definiti siti incudine (anvil site),
caratterizzati da una superficie di pietra o di legno (incudine), un percussore di pietra e spesso dalla
presenza di gusci rotti di noci e/o di altri frutti (per una descrizione dettagliata dei siti incudine a
Boa Vista vedi Visalberghi et al. 2007).
Gli episodi di uso di strumenti sono stati registrati ad libitum con i metodi del campionamento
focale (focal animal sampling, Martin & Bateson, 1993) e di tutte le occorrenze per eventi rari (all
occurences, Altmann, 1974). Un episodio di uso di strumenti ha inizio dal momento in cui il
soggetto (S) è vicino o sull’incudine con almeno una noce (o parte di una noce, o altro frutto duro) e
termina quando se ne allontana. Ogni qualvolta S inizia a rompere una nuova noce, inizia un nuovo
episodio.
Per ogni episodio focale sono stati registrati i seguenti dati:
1. Data (giorno, mese, anno).

34
2. Identità del soggetto.
3. Latenza: ora di inizio e fine dell’episodio e se l’episodio era stato interamente o
parzialmente osservato, in quanto l’osservatore non aveva potuto registrarlo dall’inizio.
Annotata inoltre la latenza di soluzione, ovvero quando S apre la noce/frutto.
4. Ogni 10 secondi, con un campionamento istantaneo (instanteous sampling, Martin &
Bateson, 1993) venivano registrati i seguenti comportamenti: uso di strumenti (S colpisce
con un sasso percussore il frutto); attività legate all’alimentazione (S cerca la noce, cerca il
sasso, tenta di aprire la noce con i denti, toglie la parte esterna sbattendola, esplorandola,
annusandola, facendo tapping, o estraendone con le dita il contenuto); alimentazione vera e
propria (S mangia il contenuto della noce o la polpa esterna); altre attività (interazioni
aggressive, sottrazione o spartizione del cibo, altri comportamenti). Ogni 10 secondi
venivano registrati il numero di colpi dati con il percussore alla noce, la posizione dove S ha
mangiato (incudine, albero o altro luogo fuori dall’incudine) e la presenza di altri individui
sulla stessa incudine.
5. Al termine di ogni episodio o quando possibile, è stato annotato il tipo di alimento coinvolto
(specie di noce, frutto, seme) e il numero di alimenti trasportati all’incudine. Inoltre è stato
registrato se l’alimento veniva aperto e se l’uso di strumenti era stato di tipo “efficente”
(quando S usa nel modo corretto gli strumenti e apre almeno una noce), “adeguato” quando
S usa nel modo corretto gli strumenti ma la noce non si apre per sua inefficienza o per altri
fattori esterni, “inadeguato” quando S non usa gli strumenti nel modo corretto (Ottoni et al.,
2005).
6. Descrizione dell’incudine: tipo di incudine (di legno, di pietra, in caverna); vegetazione in
cui si trova (chapada, en-costa, grota, brejo, morro, top-morro, per la descrizione delle
vegetazioni vedi paragrafo 3.1 e Figura 4.1); copertura (alta, media, bassa); tipo di
percussore (quarzite, arenite, siltite); peso del martello; posizione GPS.
7. Trasporto: distanza in metri del trasporto del cibo; distanza in metri del trasporto del
martello.
8. Altro: uso contemporaneo di strumenti su altre incudini e altre annotazioni rilevanti.
I sassi percussori sono stati pesati con una bilancia (Pesola AG, Switzerland) e per il gruppo Chicao
è stato annotato se l’uso di strumenti avveniva con noci offerte al capanno di avvistamento. Per
evitare di sovrastimare l’uso spontaneo di strumenti queste registrazioni sono state considerate
esclusivamente nelle analisi dell’efficenza di uso di strumenti (Paragrafo 5.3.5).
In aggiunta alle registrazioni focali, sono stati annotati tutti gli episodi di uso di strumenti con il
metodo di tutte le occorrenze (all occurences, Martin & Bateson, 1993). Poiché l’uso di strumenti

35
per mezzo di sassi e incudini è un’attività percepibile anche indirettamente (rumore dei colpi del
percussore sull’incudine e rottura dei gusci di noce), sono stati registrati quando possibile, tutti gli
episodi fuori dalla vista dell’osservatore. Ho personalmente raccolto tutti i dati sull’uso di strumenti
per il gruppo Chicao e le mie osservazioni per il gruppo Zangado sono state integrate con i dati
raccolti da Darvin R. da Silva, Jozemar e Arizomar da Silva Oliveira. Le mie registrazioni sono
state riportate su un foglio elettronico Excel (Microsoft Office) con un computer palmare (HP iPaQ
hx2400).
4.2.3.1 Analisi Statistiche I dati sull’uso di strumenti non seguono una distribuzione normale, pertanto sono stati analizzati
mediante test non parametrici. Dal momento che la dimensione dei gruppi è differente (Paragrafo
4.1), non tutti gli individui usano strumenti, e il numero di individui che usa strumenti può variare
nei mesi, ho ricavato il tasso di uso di strumenti per gruppo (numero di episodi mensili di uso di
strumenti/totale di ore di osservazione mensili) e il tasso di uso di strumenti riferito agli individui
che usano strumenti in quel mese (numero di osservazioni mensili di uso di strumenti per
individuo/ore di osservazione nel mese). Nel calcolo del tasso mensile per individuo che usa
strumenti sono state considerate sia le osservazioni complete sia quelle incomplete, ed esclusi gli
episodi in cui non è stato possibile identificare l’individuo e quelli per aprire le noci distribuite al
capanno di avvistamento.
Per confrontare i tassi mensili di uso di strumenti tra i due gruppi, ho usato il test non
parametrico di Mann-Whitney per campioni indipendenti. Con il test di correlazione di Spearman
ho esaminato la relazione tra la variazioni mensile dell’uso di strumenti e i parametri ecologici
(temperatura, precipitazioni, umidità relativa, disponibilità di frutta e di invertebrati, disponibilità di
noci catulè e di noci piassava).
I cebi di Boa Vista usano strumenti per aprire sia noci di palma (Tucum, Astrocaryum
campestre; Catulè e Catulì, Attalea sp.; Patì, Syagrus sp.; Piassava, Orbygnia sp.) sia altri semi
racchiusi da guscio duro (fruta-danta, fam. Icacinaceae; caju, fam. Anacardiaceae; caroba, fam.
Bignoniaceae; mandioca brava, fam. Euphorbiaceae). Aprire questi frutti non richiede tanta forza
quanto quella necessaria per aprire le noci di palma (Visalberghi et al., 2008), per questo ho
suddiviso il comportamento di uso di strumenti in due categorie alimentari: (1) uso di strumenti per
aprire noci di palma (frutti ad alta resistenza di frattura), (2) uso di strumenti per aprire frutti diversi
dalle noci di palma (frutti a bassa resistenza di frattura). L’analisi delle differenze stagionali
nell’uso di strumenti per aprire le diverse categorie alimentari è stata fatta confrontando tra le
stagioni il tasso di uso di strumenti per ogni individuo con il test di Wilcoxon per dati appaiati.

36
Per confrontare i tassi di uso di strumenti finalizzati ad aprire noci, altri frutti e ambedue le
categorie insieme nella varie classi di età e sesso ho applicato l'ANOVA per ranghi di Kruskal-
Wallis e il test di Mann-Whitney per i confronti tra coppie di classi.
Per esaminare l’efficenza nell’uso di strumenti ho utilizzato tutti i dati raccolti da me (focal e all
occurrence) relativi agli episodi completi per i quali è stato possibile identificare il soggetto e
estrapolare la latenza al successo, il numero di colpi, il tipo di uso di strumenti (efficente, adeguato,
inadeguato), l’alimento e il numero di noci (o frutti) portati all’incudine. In questa analisi ho incluso
anche gli episodi per aprire le noci distribuite al capanno di avvistamento.
Nelle analisi del successo di apertura le noci di palma sono state suddivise in base alla loro
durezza (vedi Visalberghi et al., 2008) in noci ad alta resistenza di frattura (noci piassava e catulì) e
in noci a bassa resistenza di frattura (noci tucum e catulè). Nella comparazione tra le classi di età e
sesso i giovani sono stati eclusi dalle analisi statistiche a causa dei pochi individui con episodi di
uso di strumenti completi (solamente 2 giovani contro 12 adulti) e il loro comportamento è stato
riportato in maniera descrittiva. Il successo (numero di episodi efficenti/numero totale degli episodi)
è stato determinato per le noci ad alta resistenza di frattura e per quelle a bassa resistenza di frattura
in termini di percentuale di successo, numero di colpi necessari per aprire la noce e durata
dell’episodio. I confronti del successo di apertura tra le categorie di cibo (noci ad alta resistenza di
frattura/noci a bassa resistenza di frattura/altri tipi di frutti) sono stati fatti per gli individui adulti
con il test di Wilcoxon per dati appaiati e i confronti tra maschi adulti e femmine adulte sono stati
fatti con il test di Mann-Whitney. In quest’ultimo confronto la categoria altri frutti è stata esclusa
per il numero esiguo di episodi completi.
Nelle analisi del tipo di materiale usato come strumento (incudini e sassi percussori) ho
analizzato esclusivamente gli episodi in cui sono stati presi i dati riguardanti le incudini ed è stato
possibile identificare e pesare il sasso percussore. Anche in questo caso è stata fatta una distinzione
tra i sassi percussori per aprire le noci e quelli usati per aprire altri frutti e i dati sono stati analizzati
separatamente. Inoltre non sono stati considerati gli episodi in cui i cebi hanno usato come
percussore una noce e nel calcolo della media dei percussori è stato escluso un episodio in cui una
femmina ha aperto due noci tucum con un sasso che pesava 30 g. I confronti tra il peso dei
percussori usati dai maschi e dalle femmine sono stati fatti tramite il t-test per dati indipendenti
(confronto tra classi di sesso). I confronti all’interno della stessa classe di sesso tra il peso dei
percussori usati per aprire noci di differente resistenza sono stati fatti con un t-test per dati
dipendenti. In queste comparazioni sono stati esclusi gli episodi in cui non è stato possibile
identificare la noce.

37
I confronti tra il materiale del percussore usato per aprire le noci e gli altri frutti è stato fatto con
un test del �2 tra i 3 tipi di categorie di cibi (noci ad alta resistenza di frattura, noci a bassa resistenza
di frattura, altri frutti) e le due categorie di materiali dei martelli (friabili, non friabili).

38
4.3 Indici di disponibilità alimentare e dati climatici
4.3.1 Disponibilità di frutta La frutta disponibile per gli animali nell’area di studio è stata stimata mediante 100 trappole a
caduta (diametro 22 cm) distribuite lungo un transetto lineare di 3 km che attraversa i cinque
ambienti principali di Boa Vista (Paragrafo 3.1 e Figura 4.1). Le trappole sono state collocate sul
tronco di alberi distanti gli uni dagli altri 30 m, ad una altezza di circa 150 cm dal terreno per
renderle meno accessibili agli animali da pascolo. A distanza di 15 giorni il contenuto delle trappole
è stato raccolto e pesato con una bilancia elettronica e il peso (g) dei frutti disponibili per area
campione (m2) è stato stimato su base mensile.
E’ importante sottolineare che la stima del peso di frutti disponibili per gli animali attraverso il
metodo delle trappole a caduta non è altro che un indice di variazione di produttività nel corso del
tempo, e non una misura del peso dei frutti che sono realmente disponibili per gli animali (Izar,
2004). Con il nostro metodo infatti sono stati pesati solamente i frutti caduti in trappole a 150 cm
dal suolo e non mangiati da animali frugivori (scimmie incluse). In ogni caso tali indici sono stati
utilizzati esclusivamente per verificare se esiste una variazione nell’offerta di frutta disponibile per
gli animali durante la ricerca. La stima mensile della disponibilità di biomassa di frutta per unità di
area campionata è stata convertita in kg/ha per permettere di confrontare quest’area di studio con
altri siti di ricerca (Terborgh 1983, Izar, 2004).
4.3.2 Disponibilità di invertebrati La disponibilità mensile di biomassa di invertebrati nell’area di studio è stata stimata mediante
trappole a caduta, la più indicata per valutare l’abbondanza degli insetti che vivono in superfice nei
terreni con bassa vegetazione. Cento trappole a caduta (contenitori di plastica del diametro di 9,5
cm) sono state interrate ad una distanza di circa 30 m le une dalle altre lungo il transetto lineare di 3
km che attraversa i cinque principali ambienti che caratterizzano l’area di studio (Paragrafo 3.1 e
Figura 4.1). Il contenuto delle trappole è stato raccolto ogni quindici giorni, messo ad essiccare e
pesato con una bilancia elettronica. La stima della biomassa mensile di invertebrati (g) per unità di
area campionata (m2) è stata poi convertita in kg/ha.
Come sottolineato per la disponibilità della frutta (Paragrafo 4.3.1) attraverso il metodo delle
trappole si ottiene un indice di variazione di abbondanza degli invertebrati nel corso del tempo e
non una misura del peso degli invertebrati che sono realmente disponibili per gli animali. Nel nostro
caso infatti sono stati pesati solamente gli invertebrati che vivono sulla superfice del terreno.

39
morro
brejo chapada en costa
top-morro
10 –
50 m
morro
brejo chapada en costa
top-morro
10 –
50 m
morro
brejo chapada en costa
top-morro
10 –
50 m
morro
brejo chapada en costa
top-morro
10 –
50 m
Figura 4.1 Parte del transetto lineare di 3 km che attraversa 5 tipi di vegetazioni: la pianura stagionalmente acquitrinosa (brejo), la pianura sabbiosa (chapada), la zona di transizione tra la pianura e il morro (en costa), il morro vero e proprio (morro) e la cima del morro (top-morro). (Illustrazione Fabio R.D. Andrade). Figure 4.1 Part of the linear transect (3Km) that goes through 5 different types of vegetation: seasonally march plain (brejo), sandy flat plain (chapada), transition zone between plain and morro (en costa), the morro (morro) and the top of the morro (top-morro)(Illustration: Fabio R.D. Andrade).
4.3.3 Produttività delle palme Nell’area di studio sono presenti diverse specie di palme appartenenti ai generi Astrocaryum,
Attalea, Orbynia e altre non ancora identificate (Fragaszy et al., 2004b). I loro frutti sono una
risorsa alimentare importante per i cebi poiché sia la polpa sia l’endosperma hanno un elevato
valore nutrizionale (EthoCebus dati non pubblicati). La maggior parte di queste palme presenta un
tronco corto e sotterraneo con foglie e caschi di frutta che emergono dal suolo. Per stimare la
produttività delle palme sono state marcate e identificate 266 palme lungo un transetto lineare di 3
km (Figura 4.1). Quattro piante di palma sono state selezionate (una a Nord, una a Sud, una ad Est e
una ad Ovest) per ogni localizzazione distante circa 30 m l’una dall’altra. Partendo da queste
localizzazioni sul transetto sono state marcate le palme adulte che si incontravano ad una distanza
minima di 3 metri e massima di 10 con nastri e targhette metalliche che ne riportavano il nome della
specie e la direzione. Una volta al mese ogni palma marcata è stata monitorata e la presenza/assenza
di infiorescenze, frutti non ancora maturi e frutti maturi è stata registrata. La densità delle diverse
specie di palme presenti a Boa Vista è stata stimata mediante la conta diretta di piante di palma
(divise per specie) osservate all’interno di quadrati di lato 100 m (un ettaro). Al fine di ottenere la
densità delle palme relativa a ciascuna fisionomia vegetazionale sono stati realizzati 4 quadrati, uno
per ogni ambiente presente nell’area di studio (brejo/chapada, en-costa, morro/top-morro,
chapada). In questo lavoro sono state considerate le specie catulè (Attalea sp.) e piassava (Orbygnia
sp.), le più abbondanti nell’area di studio (dati EthoCebus non pubblicati). Ogni mese, dal numero

40
di palme con frutti maturi registrato lungo il transetto è stata stimata la produttività delle palme in
base alla loro densità per unità di area campione (ha).
4.3.4 Dati climatici Alla stazione di ricerca ogni giorno alle 19:00 sono stati registrati con un termometro elettronico
(Hygro-Thermometer Clock model 445702", EXTECH instruments) la temperatura massima e
minima e l’umidità relativa massima e minima. Con un pluviometro (All Weather Rain Gauge,
Productive Alternatives) posizionato lontano dalla copertura della vegetazione sono state registrate
le precipitazioni.
4.3.4.1 Analisi Statistiche Il test di correlazione di Spearman è stato usato per esaminare la relazione tra disponibilità
mensile delle risorse e media delle precipitazioni mensili; la media della temperatura massima e
minima mensile; media dell’umidità relativa massima e minima mensile. Il test non parametrico di
Mann-Whitney è stato utilizzato per comparare i parametri ecologici (la media mensile delle
precipitazioni, temperatura, umidità, abbondanza delle risorse alimentari) nelle due stagioni (secca e
piovosa).
Tutti i test statistici sono a 2 code, il livello di probabilità di 0,05. Per i confronti multipli è stata
applicata la correzione di Bonferroni (Sokal & Rohlf, 1995). Le analisi sono state realizzate con il
programma Statistica (versione 7, StatSoft, Inc.).

41
5. RISULTATI
I dati stagionali dei dati climatici e degli indici di disponibilità delle risorse sono riportati in
Tabella 5.1. A Boa Vista da giugno 2006 a maggio 2007 sono stati registrati (± DS) in media 97 ±
119 mm di precipitazioni al mese per un totale di 1162 mm. La media annuale delle temperature
massime è stata 32,3 ± 1,7 °C e la media annuale delle temperature minime registrate è stata 21,2 ±
1,7 °C. La media annuale dell’umidità relativa massima registrata è stata 84,2 ± 9,3 % e la media
annuale dell’umidità relativa minima registrata è stata 46,7 ± 11,1 %. In base ai nostri risultati e in
accordo con Embrapa (Banca dati climatici del Brasile) sono state definite due stagioni: la stagione
secca da maggio a settembre (N = 5); la stagione piovosa da ottobre a aprile (N = 7).
5.1 Disponibilità delle risorse
L’abbondanza delle risorse trofiche stimate tramite l’indice di disponibilità di frutta e di
invertebrati è molto variabile nel corso dell’anno (Figura 5.1) e non differisce significativamente tra
la stagione secca e piovosa (Tabella 5.1). Inoltre le variazioni mensili di biomassa di frutta e
invertebrati non sono correlate né alle precipitazioni (frutta: Spearman rs = 0,04, N = 12, p = 0,89;
invertebrati Spearman rs = 0,5, N = 12, p = 0,1), né alle temperature massime (frutta: Spearman rs =
-0,03, N = 12, p = 0,91; invertebrati Spearman rs = 0,52, N = 12, p = 0,08), né alle temperature
minime (frutta: Spearman rs = 0,31, N = 12, p = 0,33; invertebrati Spearman rs = 0,5, N = 12, p =
0,09), né alle umidità relative massime (frutta: Spearman rs = 0,24, N = 12, p = 0,46; invertebrati
Spearman rs = 0,08, N = 12, p = 0,81) né alle umidità relative minime (frutta: Spearman rs = 0,02, N
= 12, p = 0,46; invertebrati Spearman rs = 0,16, N = 12, p = 0,61).
Le noci sono disponibili durante tutto l’anno(Figura 5.2). La disponibilità di noci piassava non
varia fra stagioni (Tabella 5.1 e Figura 5.2a), mentre quella di noci catulè è maggiore nella stagione
secca rispetto a quella piovosa (Tabella 5.1) e tale abbondanza risulta negativamente correlata alle
precipitazioni (Spearman rs = -0,88, N = 12, p < 0,001; Figura 5.2a-b).
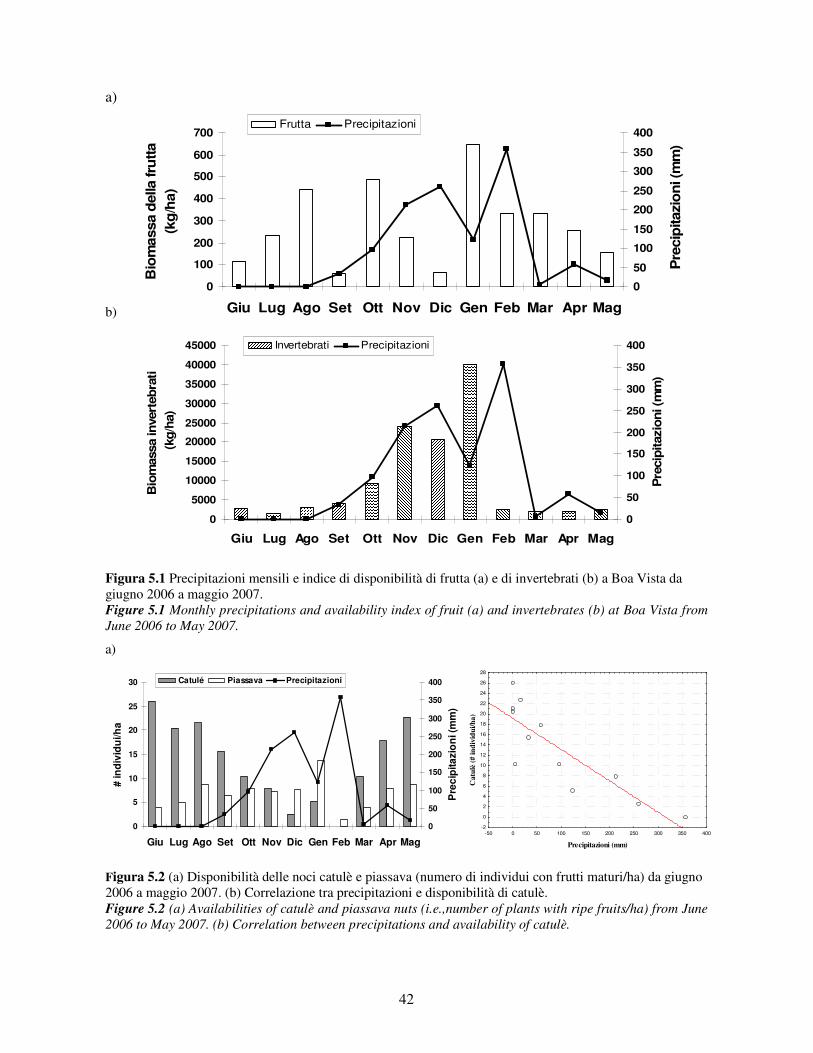
42
a)
b)
Figura 5.1 Precipitazioni mensili e indice di disponibilità di frutta (a) e di invertebrati (b) a Boa Vista da giugno 2006 a maggio 2007. Figure 5.1 Monthly precipitations and availability index of fruit (a) and invertebrates (b) at Boa Vista from June 2006 to May 2007.
a)
Figura 5.2 (a) Disponibilità delle noci catulè e piassava (numero di individui con frutti maturi/ha) da giugno 2006 a maggio 2007. (b) Correlazione tra precipitazioni e disponibilità di catulè. Figure 5.2 (a) Availabilities of catulè and piassava nuts (i.e.,number of plants with ripe fruits/ha) from June 2006 to May 2007. (b) Correlation between precipitations and availability of catulè.
0
100
200
300
400
500
600
700
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
Bio
mas
sa d
ella
fru
tta
(kg
/ha)
0
50
100
150
200
250
300
350
400
Pre
cip
itazi
oni
(mm
)
Frutta Precipitazioni
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
Bio
mas
sa in
vert
ebra
ti (k
g/ha
)
0
50
100
150
200
250
300
350
400
Pre
cipi
tazi
oni (
mm
)
Invertebrati Precipitazioni
-50 0 50 100 150 200 250 300 350 400
Precipitazioni (mm)
-2
0
2
4
6
8
10
12
14
16
18
20
22
24
26
28
Cat
ulè
(# in
divi
dui/h
a)
0
5
10
15
20
25
30
Giu Lug Ago Set Ott Nov Dic Gen Feb Mar Apr Mag
# in
divi
dui/h
a
0
50
100
150
200
250
300
350
400
Pre
cipi
tazi
oni (
mm
)
Catulé Piassava Precipitazioni
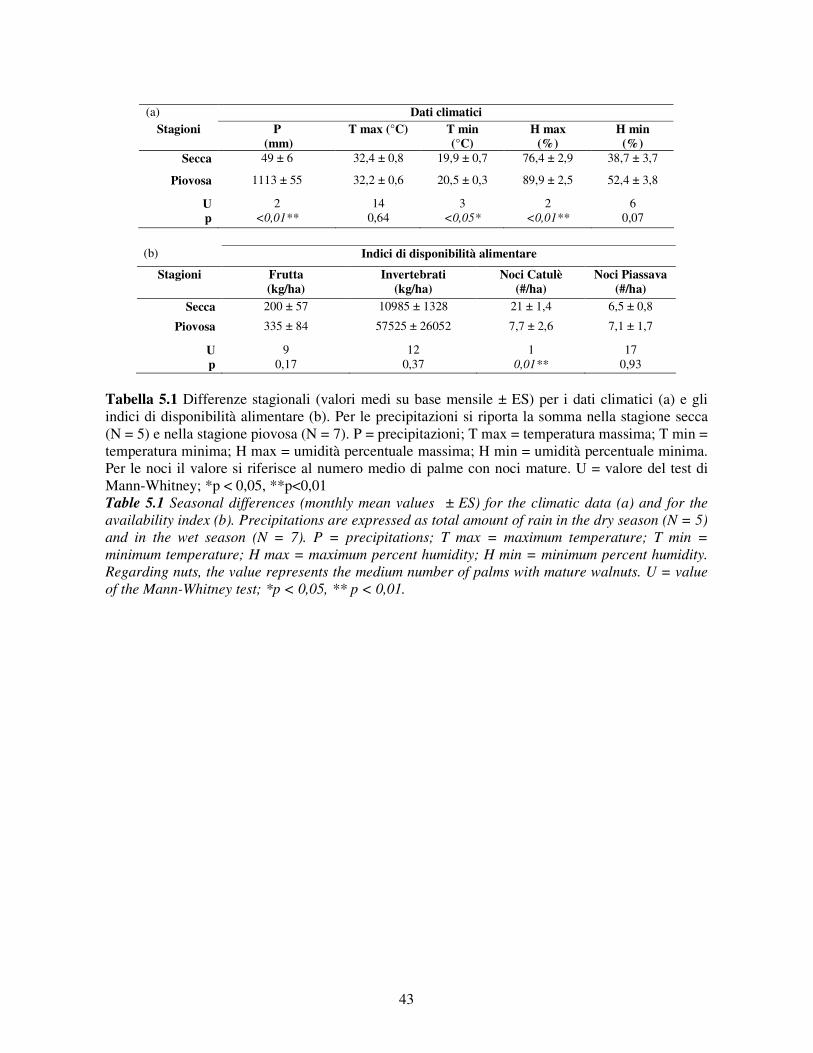
43
(a) Dati climatici
Stagioni P (mm)
T max (°C) T min (°C)
H max (%)
H min (%)
Secca 49 ± 6 32,4 ± 0,8 19,9 ± 0,7 76,4 ± 2,9 38,7 ± 3,7
Piovosa 1113 ± 55 32,2 ± 0,6 20,5 ± 0,3 89,9 ± 2,5 52,4 ± 3,8
U 2 14 3 2 6 p <0,01** 0,64 <0,05* <0,01** 0,07
(b) Indici di disponibilità alimentare
Stagioni Frutta (kg/ha)
Invertebrati (kg/ha)
Noci Catulè (#/ha)
Noci Piassava (#/ha)
Secca 200 ± 57 10985 ± 1328 21 ± 1,4 6,5 ± 0,8
Piovosa 335 ± 84 57525 ± 26052 7,7 ± 2,6 7,1 ± 1,7
U 9 12 1 17 p 0,17 0,37 0,01** 0,93
Tabella 5.1 Differenze stagionali (valori medi su base mensile ± ES) per i dati climatici (a) e gli indici di disponibilità alimentare (b). Per le precipitazioni si riporta la somma nella stagione secca (N = 5) e nella stagione piovosa (N = 7). P = precipitazioni; T max = temperatura massima; T min = temperatura minima; H max = umidità percentuale massima; H min = umidità percentuale minima. Per le noci il valore si riferisce al numero medio di palme con noci mature. U = valore del test di Mann-Whitney; *p < 0,05, **p<0,01 Table 5.1 Seasonal differences (monthly mean values ± ES) for the climatic data (a) and for the availability index (b). Precipitations are expressed as total amount of rain in the dry season (N = 5) and in the wet season (N = 7). P = precipitations; T max = maximum temperature; T min = minimum temperature; H max = maximum percent humidity; H min = minimum percent humidity. Regarding nuts, the value represents the medium number of palms with mature walnuts. U = value of the Mann-Whitney test; *p < 0,05, ** p < 0,01.
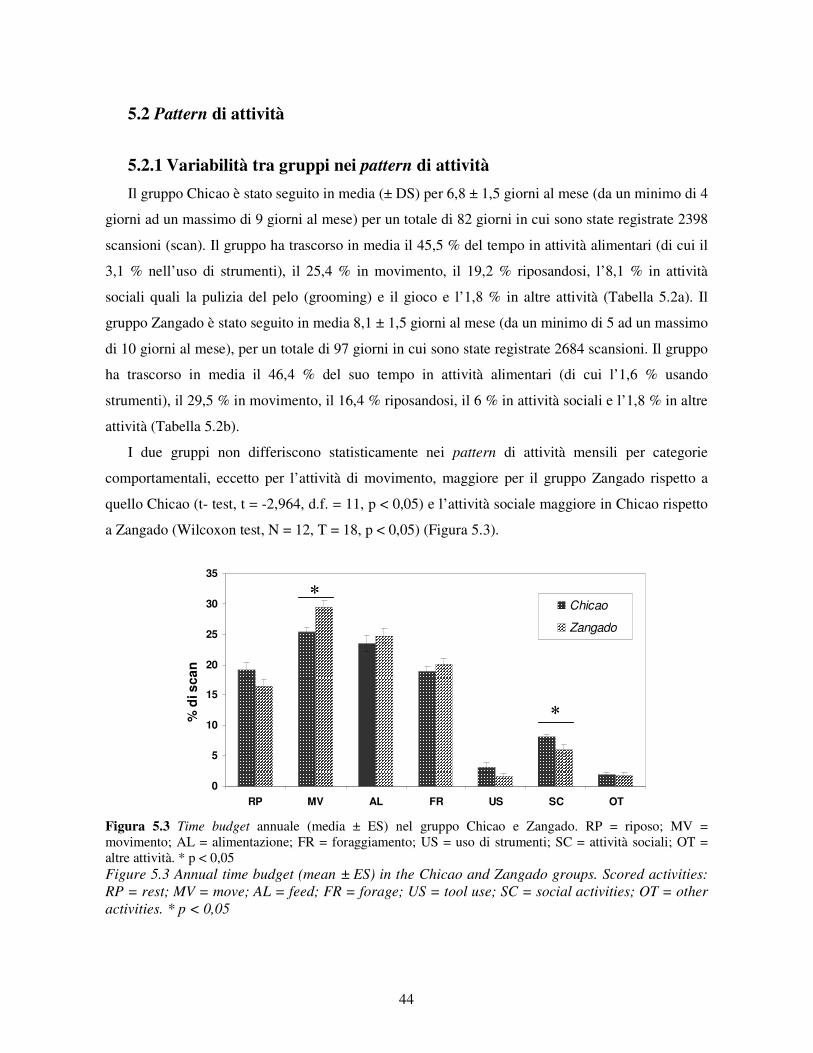
44
5.2 Pattern di attività
5.2.1 Variabilità tra gruppi nei pattern di attività Il gruppo Chicao è stato seguito in media (± DS) per 6,8 ± 1,5 giorni al mese (da un minimo di 4
giorni ad un massimo di 9 giorni al mese) per un totale di 82 giorni in cui sono state registrate 2398
scansioni (scan). Il gruppo ha trascorso in media il 45,5 % del tempo in attività alimentari (di cui il
3,1 % nell’uso di strumenti), il 25,4 % in movimento, il 19,2 % riposandosi, l’8,1 % in attività
sociali quali la pulizia del pelo (grooming) e il gioco e l’1,8 % in altre attività (Tabella 5.2a). Il
gruppo Zangado è stato seguito in media 8,1 ± 1,5 giorni al mese (da un minimo di 5 ad un massimo
di 10 giorni al mese), per un totale di 97 giorni in cui sono state registrate 2684 scansioni. Il gruppo
ha trascorso in media il 46,4 % del suo tempo in attività alimentari (di cui l’1,6 % usando
strumenti), il 29,5 % in movimento, il 16,4 % riposandosi, il 6 % in attività sociali e l’1,8 % in altre
attività (Tabella 5.2b).
I due gruppi non differiscono statisticamente nei pattern di attività mensili per categorie
comportamentali, eccetto per l’attività di movimento, maggiore per il gruppo Zangado rispetto a
quello Chicao (t- test, t = -2,964, d.f. = 11, p < 0,05) e l’attività sociale maggiore in Chicao rispetto
a Zangado (Wilcoxon test, N = 12, T = 18, p < 0,05) (Figura 5.3).
Figura 5.3 Time budget annuale (media ± ES) nel gruppo Chicao e Zangado. RP = riposo; MV = movimento; AL = alimentazione; FR = foraggiamento; US = uso di strumenti; SC = attività sociali; OT = altre attività. * p < 0,05 Figure 5.3 Annual time budget (mean ± ES) in the Chicao and Zangado groups. Scored activities: RP = rest; MV = move; AL = feed; FR = forage; US = tool use; SC = social activities; OT = other activities. * p < 0,05
0
5
10
15
20
25
30
35
RP MV AL FR US SC OT
% d
i sca
n
Chicao
Zangado
*
*
0
5
10
15
20
25
30
35
RP MV AL FR US SC OT
% d
i sca
n
Chicao
Zangado
**
**
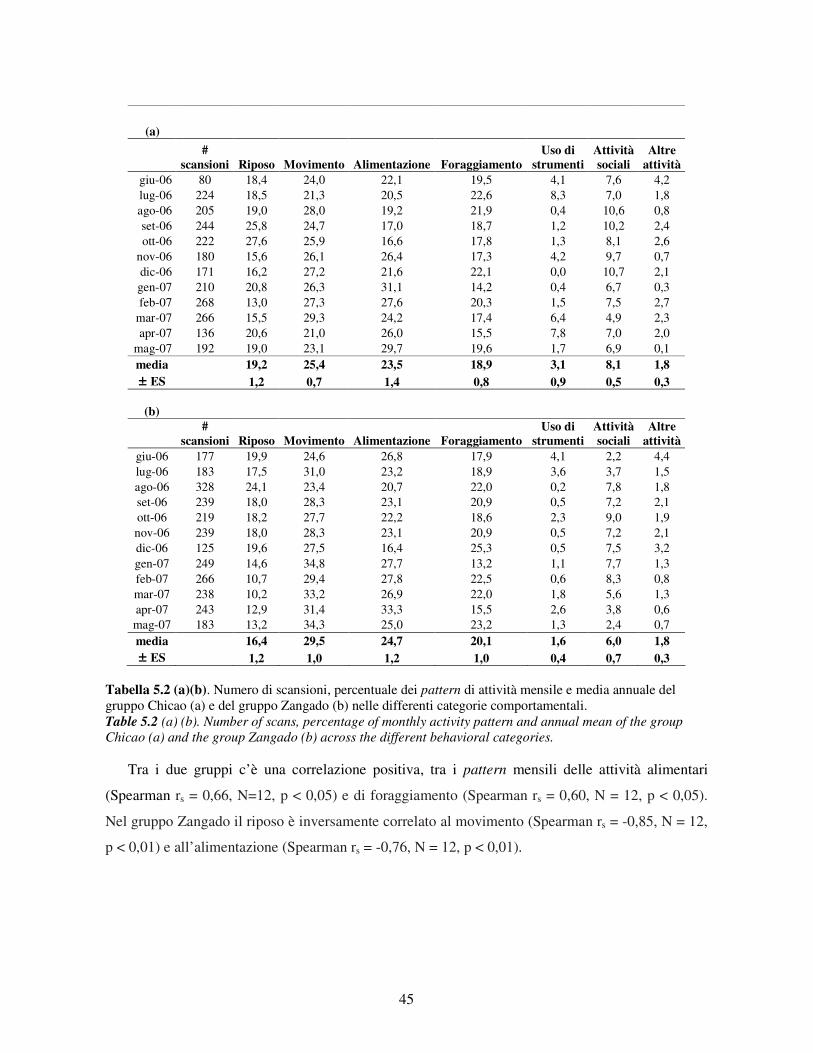
45
(a)
#
scansioni Riposo Movimento Alimentazione Foraggiamento Uso di
strumenti Attività sociali
Altre attività
giu-06 80 18,4 24,0 22,1 19,5 4,1 7,6 4,2 lug-06 224 18,5 21,3 20,5 22,6 8,3 7,0 1,8 ago-06 205 19,0 28,0 19,2 21,9 0,4 10,6 0,8 set-06 244 25,8 24,7 17,0 18,7 1,2 10,2 2,4 ott-06 222 27,6 25,9 16,6 17,8 1,3 8,1 2,6
nov-06 180 15,6 26,1 26,4 17,3 4,2 9,7 0,7 dic-06 171 16,2 27,2 21,6 22,1 0,0 10,7 2,1 gen-07 210 20,8 26,3 31,1 14,2 0,4 6,7 0,3 feb-07 268 13,0 27,3 27,6 20,3 1,5 7,5 2,7
mar-07 266 15,5 29,3 24,2 17,4 6,4 4,9 2,3 apr-07 136 20,6 21,0 26,0 15,5 7,8 7,0 2,0
mag-07 192 19,0 23,1 29,7 19,6 1,7 6,9 0,1 media 19,2 25,4 23,5 18,9 3,1 8,1 1,8 ± ES 1,2 0,7 1,4 0,8 0,9 0,5 0,3
(b)
#
scansioni Riposo Movimento Alimentazione Foraggiamento Uso di
strumenti Attività sociali
Altre attività
giu-06 177 19,9 24,6 26,8 17,9 4,1 2,2 4,4 lug-06 183 17,5 31,0 23,2 18,9 3,6 3,7 1,5 ago-06 328 24,1 23,4 20,7 22,0 0,2 7,8 1,8 set-06 239 18,0 28,3 23,1 20,9 0,5 7,2 2,1 ott-06 219 18,2 27,7 22,2 18,6 2,3 9,0 1,9 nov-06 239 18,0 28,3 23,1 20,9 0,5 7,2 2,1 dic-06 125 19,6 27,5 16,4 25,3 0,5 7,5 3,2 gen-07 249 14,6 34,8 27,7 13,2 1,1 7,7 1,3 feb-07 266 10,7 29,4 27,8 22,5 0,6 8,3 0,8 mar-07 238 10,2 33,2 26,9 22,0 1,8 5,6 1,3 apr-07 243 12,9 31,4 33,3 15,5 2,6 3,8 0,6 mag-07 183 13,2 34,3 25,0 23,2 1,3 2,4 0,7 media 16,4 29,5 24,7 20,1 1,6 6,0 1,8 ± ES 1,2 1,0 1,2 1,0 0,4 0,7 0,3
Tabella 5.2 (a)(b). Numero di scansioni, percentuale dei pattern di attività mensile e media annuale del gruppo Chicao (a) e del gruppo Zangado (b) nelle differenti categorie comportamentali. Table 5.2 (a) (b). Number of scans, percentage of monthly activity pattern and annual mean of the group Chicao (a) and the group Zangado (b) across the different behavioral categories.
Tra i due gruppi c’è una correlazione positiva, tra i pattern mensili delle attività alimentari
(Spearman rs = 0,66, N=12, p < 0,05) e di foraggiamento (Spearman rs = 0,60, N = 12, p < 0,05).
Nel gruppo Zangado il riposo è inversamente correlato al movimento (Spearman rs = -0,85, N = 12,
p < 0,01) e all’alimentazione (Spearman rs = -0,76, N = 12, p < 0,01).
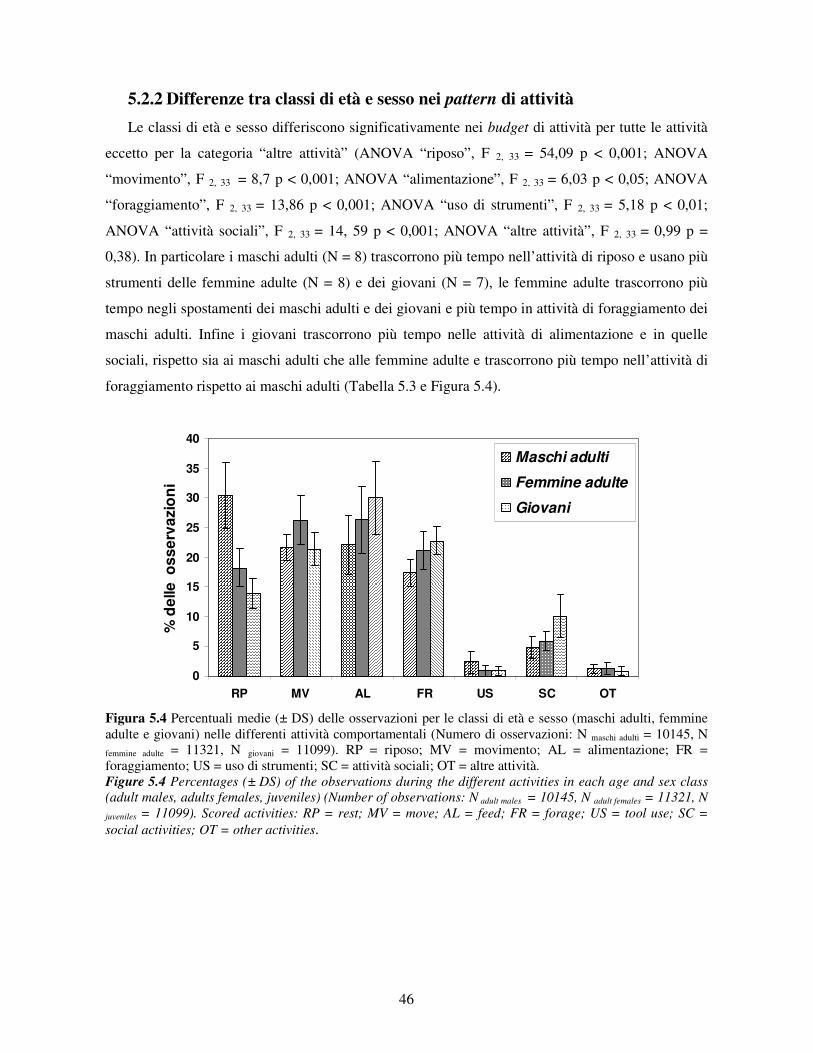
46
5.2.2 Differenze tra classi di età e sesso nei pattern di attività Le classi di età e sesso differiscono significativamente nei budget di attività per tutte le attività
eccetto per la categoria “altre attività” (ANOVA “riposo”, F 2, 33 = 54,09 p < 0,001; ANOVA
“movimento”, F 2, 33 = 8,7 p < 0,001; ANOVA “alimentazione”, F 2, 33 = 6,03 p < 0,05; ANOVA
“foraggiamento”, F 2, 33 = 13,86 p < 0,001; ANOVA “uso di strumenti”, F 2, 33 = 5,18 p < 0,01;
ANOVA “attività sociali”, F 2, 33 = 14, 59 p < 0,001; ANOVA “altre attività”, F 2, 33 = 0,99 p =
0,38). In particolare i maschi adulti (N = 8) trascorrono più tempo nell’attività di riposo e usano più
strumenti delle femmine adulte (N = 8) e dei giovani (N = 7), le femmine adulte trascorrono più
tempo negli spostamenti dei maschi adulti e dei giovani e più tempo in attività di foraggiamento dei
maschi adulti. Infine i giovani trascorrono più tempo nelle attività di alimentazione e in quelle
sociali, rispetto sia ai maschi adulti che alle femmine adulte e trascorrono più tempo nell’attività di
foraggiamento rispetto ai maschi adulti (Tabella 5.3 e Figura 5.4).
Figura 5.4 Percentuali medie (± DS) delle osservazioni per le classi di età e sesso (maschi adulti, femmine adulte e giovani) nelle differenti attività comportamentali (Numero di osservazioni: N maschi adulti = 10145, N femmine adulte = 11321, N giovani = 11099). RP = riposo; MV = movimento; AL = alimentazione; FR = foraggiamento; US = uso di strumenti; SC = attività sociali; OT = altre attività. Figure 5.4 Percentages (± DS) of the observations during the different activities in each age and sex class (adult males, adults females, juveniles) (Number of observations: N adult males = 10145, N adult females = 11321, N juveniles = 11099). Scored activities: RP = rest; MV = move; AL = feed; FR = forage; US = tool use; SC = social activities; OT = other activities.
0
5
10
15
20
25
30
35
40
RP MV AL FR US SC OT
% d
elle
oss
erva
zion
i
Maschi adulti
Femmine adulte
Giovani
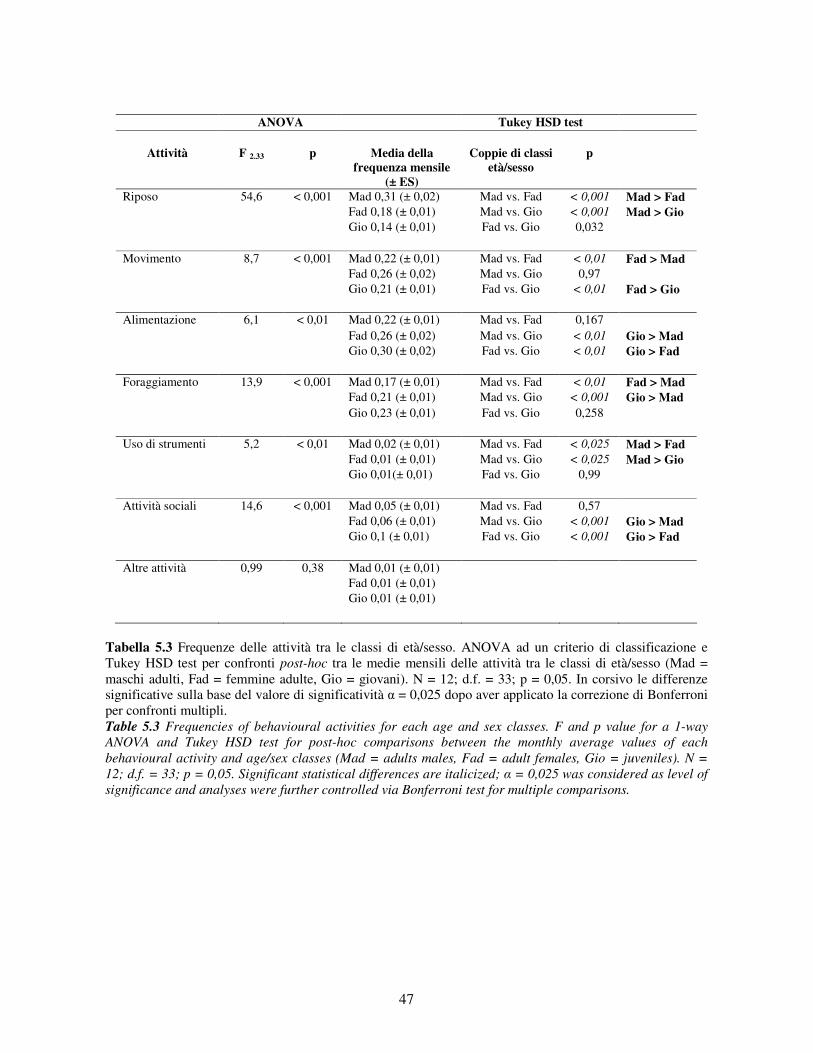
47
ANOVA Tukey HSD test
Attività
F 2.33
p
Media della frequenza mensile
(± ES)
Coppie di classi
età/sesso
p
Riposo 54,6 < 0,001 Mad 0,31 (± 0,02) Mad vs. Fad < 0,001 Mad > Fad Fad 0,18 (± 0,01) Mad vs. Gio < 0,001 Mad > Gio Gio 0,14 (± 0,01) Fad vs. Gio 0,032 Movimento 8,7 < 0,001 Mad 0,22 (± 0,01) Mad vs. Fad < 0,01 Fad > Mad Fad 0,26 (± 0,02) Mad vs. Gio 0,97 Gio 0,21 (± 0,01) Fad vs. Gio < 0,01 Fad > Gio Alimentazione 6,1 < 0,01 Mad 0,22 (± 0,01) Mad vs. Fad 0,167 Fad 0,26 (± 0,02) Mad vs. Gio < 0,01 Gio > Mad Gio 0,30 (± 0,02) Fad vs. Gio < 0,01 Gio > Fad Foraggiamento 13,9 < 0,001 Mad 0,17 (± 0,01) Mad vs. Fad < 0,01 Fad > Mad Fad 0,21 (± 0,01) Mad vs. Gio < 0,001 Gio > Mad Gio 0,23 (± 0,01) Fad vs. Gio 0,258 Uso di strumenti 5,2 < 0,01 Mad 0,02 (± 0,01) Mad vs. Fad < 0,025 Mad > Fad Fad 0,01 (± 0,01) Mad vs. Gio < 0,025 Mad > Gio Gio 0,01(± 0,01) Fad vs. Gio 0,99 Attività sociali 14,6 < 0,001 Mad 0,05 (± 0,01) Mad vs. Fad 0,57 Fad 0,06 (± 0,01) Mad vs. Gio < 0,001 Gio > Mad Gio 0,1 (± 0,01) Fad vs. Gio < 0,001 Gio > Fad Altre attività 0,99 0,38 Mad 0,01 (± 0,01) Fad 0,01 (± 0,01) Gio 0,01 (± 0,01)
Tabella 5.3 Frequenze delle attività tra le classi di età/sesso. ANOVA ad un criterio di classificazione e Tukey HSD test per confronti post-hoc tra le medie mensili delle attività tra le classi di età/sesso (Mad = maschi adulti, Fad = femmine adulte, Gio = giovani). N = 12; d.f. = 33; p = 0,05. In corsivo le differenze significative sulla base del valore di significatività � = 0,025 dopo aver applicato la correzione di Bonferroni per confronti multipli. Table 5.3 Frequencies of behavioural activities for each age and sex classes. F and p value for a 1-way ANOVA and Tukey HSD test for post-hoc comparisons between the monthly average values of each behavioural activity and age/sex classes (Mad = adults males, Fad = adult females, Gio = juveniles). N = 12; d.f. = 33; p = 0,05. Significant statistical differences are italicized; � = 0,025 was considered as level of significance and analyses were further controlled via Bonferroni test for multiple comparisons.
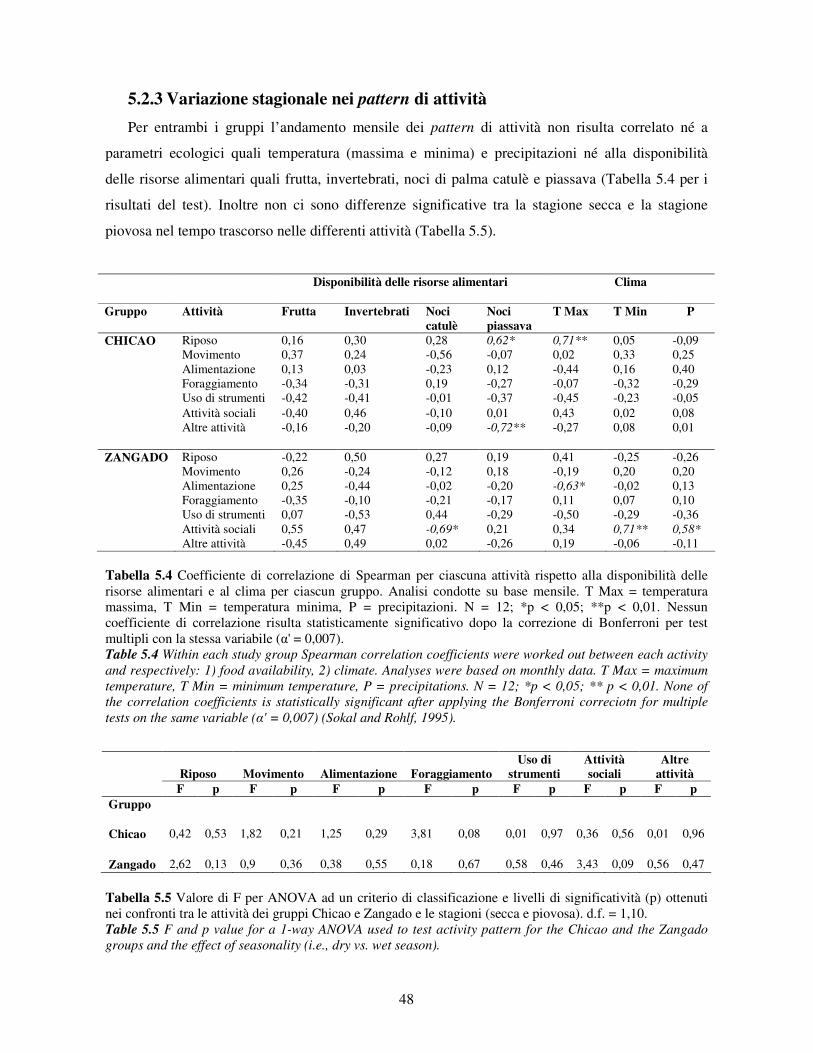
48
5.2.3 Variazione stagionale nei pattern di attività Per entrambi i gruppi l’andamento mensile dei pattern di attività non risulta correlato né a
parametri ecologici quali temperatura (massima e minima) e precipitazioni né alla disponibilità
delle risorse alimentari quali frutta, invertebrati, noci di palma catulè e piassava (Tabella 5.4 per i
risultati del test). Inoltre non ci sono differenze significative tra la stagione secca e la stagione
piovosa nel tempo trascorso nelle differenti attività (Tabella 5.5).
Disponibilità delle risorse alimentari
Clima
Gruppo Attività Frutta Invertebrati Noci catulè
Noci piassava
T Max T Min P
CHICAO Riposo 0,16 0,30 0,28 0,62* 0,71** 0,05 -0,09 Movimento 0,37 0,24 -0,56 -0,07 0,02 0,33 0,25 Alimentazione 0,13 0,03 -0,23 0,12 -0,44 0,16 0,40 Foraggiamento -0,34 -0,31 0,19 -0,27 -0,07 -0,32 -0,29 Uso di strumenti -0,42 -0,41 -0,01 -0,37 -0,45 -0,23 -0,05 Attività sociali -0,40 0,46 -0,10 0,01 0,43 0,02 0,08 Altre attività -0,16 -0,20 -0,09 -0,72** -0,27 0,08 0,01 ZANGADO Riposo -0,22 0,50 0,27 0,19 0,41 -0,25 -0,26 Movimento 0,26 -0,24 -0,12 0,18 -0,19 0,20 0,20 Alimentazione 0,25 -0,44 -0,02 -0,20 -0,63* -0,02 0,13 Foraggiamento -0,35 -0,10 -0,21 -0,17 0,11 0,07 0,10 Uso di strumenti 0,07 -0,53 0,44 -0,29 -0,50 -0,29 -0,36 Attività sociali 0,55 0,47 -0,69* 0,21 0,34 0,71** 0,58* Altre attività -0,45 0,49 0,02 -0,26 0,19 -0,06 -0,11 Tabella 5.4 Coefficiente di correlazione di Spearman per ciascuna attività rispetto alla disponibilità delle risorse alimentari e al clima per ciascun gruppo. Analisi condotte su base mensile. T Max = temperatura massima, T Min = temperatura minima, P = precipitazioni. N = 12; *p < 0,05; **p < 0,01. Nessun coefficiente di correlazione risulta statisticamente significativo dopo la correzione di Bonferroni per test multipli con la stessa variabile (�' = 0,007). Table 5.4 Within each study group Spearman correlation coefficients were worked out between each activity and respectively: 1) food availability, 2) climate. Analyses were based on monthly data. T Max = maximum temperature, T Min = minimum temperature, P = precipitations. N = 12; *p < 0,05; ** p < 0,01. None of the correlation coefficients is statistically significant after applying the Bonferroni correciotn for multiple tests on the same variable (�' = 0,007) (Sokal and Rohlf, 1995).
Riposo
Movimento
Alimentazione
Foraggiamento
Uso di strumenti
Attività sociali
Altre attività
F p F p F p F p F p F p F p Gruppo Chicao 0,42 0,53 1,82 0,21 1,25 0,29 3,81 0,08 0,01 0,97 0,36 0,56 0,01 0,96 Zangado 2,62 0,13 0,9 0,36 0,38 0,55 0,18 0,67 0,58 0,46 3,43 0,09 0,56 0,47 Tabella 5.5 Valore di F per ANOVA ad un criterio di classificazione e livelli di significatività (p) ottenuti nei confronti tra le attività dei gruppi Chicao e Zangado e le stagioni (secca e piovosa). d.f. = 1,10. Table 5.5 F and p value for a 1-way ANOVA used to test activity pattern for the Chicao and the Zangado groups and the effect of seasonality (i.e., dry vs. wet season).
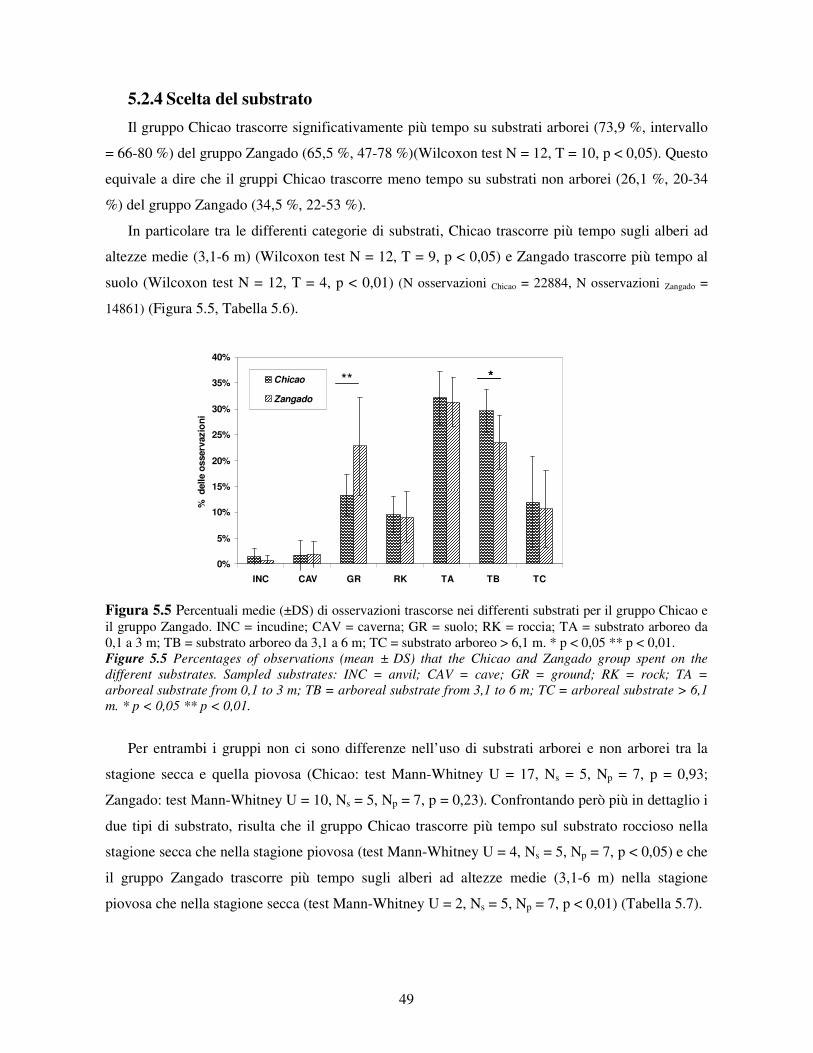
49
5.2.4 Scelta del substrato Il gruppo Chicao trascorre significativamente più tempo su substrati arborei (73,9 %, intervallo
= 66-80 %) del gruppo Zangado (65,5 %, 47-78 %)(Wilcoxon test N = 12, T = 10, p < 0,05). Questo
equivale a dire che il gruppi Chicao trascorre meno tempo su substrati non arborei (26,1 %, 20-34
%) del gruppo Zangado (34,5 %, 22-53 %).
In particolare tra le differenti categorie di substrati, Chicao trascorre più tempo sugli alberi ad
altezze medie (3,1-6 m) (Wilcoxon test N = 12, T = 9, p < 0,05) e Zangado trascorre più tempo al
suolo (Wilcoxon test N = 12, T = 4, p < 0,01) (N osservazioni Chicao = 22884, N osservazioni Zangado =
14861) (Figura 5.5, Tabella 5.6).
Figura 5.5 Percentuali medie (±DS) di osservazioni trascorse nei differenti substrati per il gruppo Chicao e il gruppo Zangado. INC = incudine; CAV = caverna; GR = suolo; RK = roccia; TA = substrato arboreo da 0,1 a 3 m; TB = substrato arboreo da 3,1 a 6 m; TC = substrato arboreo > 6,1 m. * p < 0,05 ** p < 0,01. Figure 5.5 Percentages of observations (mean ± DS) that the Chicao and Zangado group spent on the different substrates. Sampled substrates: INC = anvil; CAV = cave; GR = ground; RK = rock; TA = arboreal substrate from 0,1 to 3 m; TB = arboreal substrate from 3,1 to 6 m; TC = arboreal substrate > 6,1 m. * p < 0,05 ** p < 0,01.
Per entrambi i gruppi non ci sono differenze nell’uso di substrati arborei e non arborei tra la
stagione secca e quella piovosa (Chicao: test Mann-Whitney U = 17, Ns = 5, Np = 7, p = 0,93;
Zangado: test Mann-Whitney U = 10, Ns = 5, Np = 7, p = 0,23). Confrontando però più in dettaglio i
due tipi di substrato, risulta che il gruppo Chicao trascorre più tempo sul substrato roccioso nella
stagione secca che nella stagione piovosa (test Mann-Whitney U = 4, Ns = 5, Np = 7, p < 0,05) e che
il gruppo Zangado trascorre più tempo sugli alberi ad altezze medie (3,1-6 m) nella stagione
piovosa che nella stagione secca (test Mann-Whitney U = 2, Ns = 5, Np = 7, p < 0,01) (Tabella 5.7).
0%
5%
10%
15%
20%
25%
30%
35%
40%
INC CAV GR RK TA TB TC
% d
elle
oss
erva
zion
i
Chicao
Zangado
***
0%
5%
10%
15%
20%
25%
30%
35%
40%
INC CAV GR RK TA TB TC
% d
elle
oss
erva
zion
i
Chicao
Zangado
*
0%
5%
10%
15%
20%
25%
30%
35%
40%
INC CAV GR RK TA TB TC
% d
elle
oss
erva
zion
i
Chicao
Zangado
****
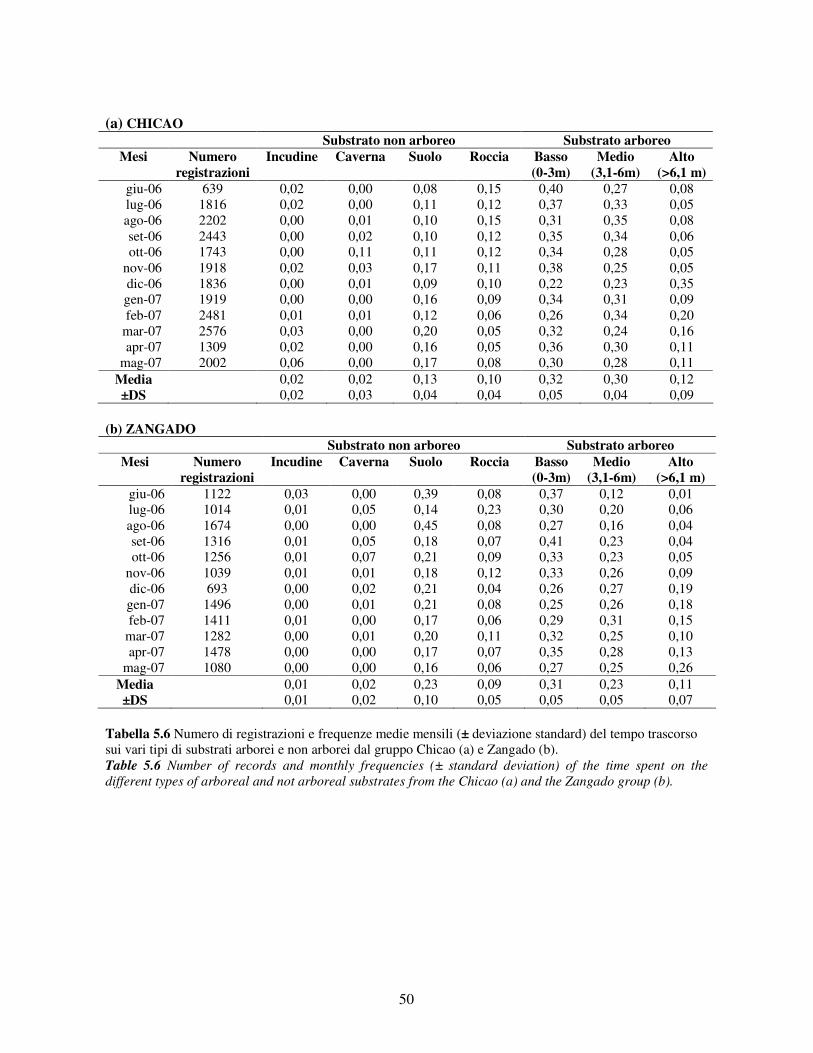
50
(a) CHICAO Substrato non arboreo Substrato arboreo
Mesi Numero registrazioni
Incudine Caverna Suolo Roccia Basso (0-3m)
Medio (3,1-6m)
Alto (>6,1 m)
giu-06 639 0,02 0,00 0,08 0,15 0,40 0,27 0,08 lug-06 1816 0,02 0,00 0,11 0,12 0,37 0,33 0,05 ago-06 2202 0,00 0,01 0,10 0,15 0,31 0,35 0,08 set-06 2443 0,00 0,02 0,10 0,12 0,35 0,34 0,06 ott-06 1743 0,00 0,11 0,11 0,12 0,34 0,28 0,05
nov-06 1918 0,02 0,03 0,17 0,11 0,38 0,25 0,05 dic-06 1836 0,00 0,01 0,09 0,10 0,22 0,23 0,35 gen-07 1919 0,00 0,00 0,16 0,09 0,34 0,31 0,09 feb-07 2481 0,01 0,01 0,12 0,06 0,26 0,34 0,20
mar-07 2576 0,03 0,00 0,20 0,05 0,32 0,24 0,16 apr-07 1309 0,02 0,00 0,16 0,05 0,36 0,30 0,11
mag-07 2002 0,06 0,00 0,17 0,08 0,30 0,28 0,11 Media 0,02 0,02 0,13 0,10 0,32 0,30 0,12 ±DS 0,02 0,03 0,04 0,04 0,05 0,04 0,09
(b) ZANGADO Substrato non arboreo Substrato arboreo
Mesi Numero registrazioni
Incudine Caverna Suolo Roccia Basso (0-3m)
Medio (3,1-6m)
Alto (>6,1 m)
giu-06 1122 0,03 0,00 0,39 0,08 0,37 0,12 0,01 lug-06 1014 0,01 0,05 0,14 0,23 0,30 0,20 0,06 ago-06 1674 0,00 0,00 0,45 0,08 0,27 0,16 0,04 set-06 1316 0,01 0,05 0,18 0,07 0,41 0,23 0,04 ott-06 1256 0,01 0,07 0,21 0,09 0,33 0,23 0,05
nov-06 1039 0,01 0,01 0,18 0,12 0,33 0,26 0,09 dic-06 693 0,00 0,02 0,21 0,04 0,26 0,27 0,19 gen-07 1496 0,00 0,01 0,21 0,08 0,25 0,26 0,18 feb-07 1411 0,01 0,00 0,17 0,06 0,29 0,31 0,15
mar-07 1282 0,00 0,01 0,20 0,11 0,32 0,25 0,10 apr-07 1478 0,00 0,00 0,17 0,07 0,35 0,28 0,13
mag-07 1080 0,00 0,00 0,16 0,06 0,27 0,25 0,26 Media 0,01 0,02 0,23 0,09 0,31 0,23 0,11 ±DS 0,01 0,02 0,10 0,05 0,05 0,05 0,07
Tabella 5.6 Numero di registrazioni e frequenze medie mensili (± deviazione standard) del tempo trascorso sui vari tipi di substrati arborei e non arborei dal gruppo Chicao (a) e Zangado (b). Table 5.6 Number of records and monthly frequencies (± standard deviation) of the time spent on the different types of arboreal and not arboreal substrates from the Chicao (a) and the Zangado group (b).

51
Substrato non arboreo Substrato arboreo
Incudine Caverna Suolo Roccia Basso (0-3m)
Medio (3,1-6m)
Alto (>6,1m)
Stagioni C Z C Z C Z C Z C Z C Z C Z Secca (n=5) 2,0 1,0 0,8 2,1 11,0 26,4 12,3 10,5 34,5 32,4 31,5 19,2 7,8 8,5 Piovosa (n=7) 1,2 0,6 2,2 1,9 14,4 19,2 8,1 8,4 31,7 30,5 27,9 26,6 14,5 12,8 Mann-Whitney (U)
15
13
15
12
9
17
4
16
13
14
9
2
11
8
p
0,75
0,53
0,75
0,43
0,20
1
0,03*
0,88
0,53
0,64
0,20
0,01*
0,34
0,15
Tabella 5.7 Percentuale di tempo trascorso dal gruppo Chicao (C) e dal gruppo Zangado (Z) su substrati non arborei (incudine, caverna, suolo, roccia) e su substrati arborei (basso, medio, alto) nelle differenti stagioni. Test di Mann-Whitney per confronto tra stagioni e tempo trascorso sui substrati arboreo/non arboreo * p < 0,05. Table 5.7 Percent of time spent from the Chicao (C) and the Zangado group (Z) on both no-arboreal substrates (anvil, cave, ground, rock) and arboreal substrates (low, medium, high) during the different seasons. Mann-Whitney test for comparison between seasons and time spent on arboreal/no-arboreal substrates. * p < 0,05.
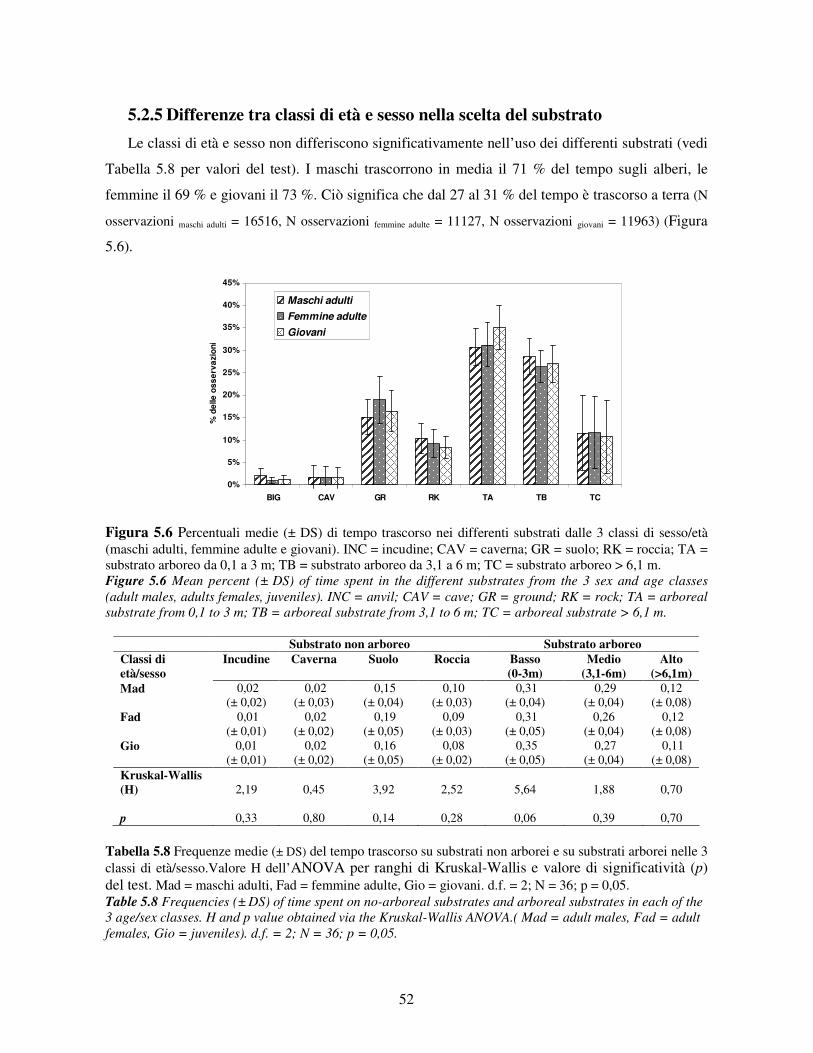
52
5.2.5 Differenze tra classi di età e sesso nella scelta del substrato Le classi di età e sesso non differiscono significativamente nell’uso dei differenti substrati (vedi
Tabella 5.8 per valori del test). I maschi trascorrono in media il 71 % del tempo sugli alberi, le
femmine il 69 % e giovani il 73 %. Ciò significa che dal 27 al 31 % del tempo è trascorso a terra (N
osservazioni maschi adulti = 16516, N osservazioni femmine adulte = 11127, N osservazioni giovani = 11963) (Figura
5.6).
0%
5%
10%
15%
20%
25%
30%
35%
40%
45%
BIG CAV GR RK TA TB TC
% d
elle
oss
erva
zion
i
Maschi adultiFemmine adulteGiovani
Figura 5.6 Percentuali medie (± DS) di tempo trascorso nei differenti substrati dalle 3 classi di sesso/età (maschi adulti, femmine adulte e giovani). INC = incudine; CAV = caverna; GR = suolo; RK = roccia; TA = substrato arboreo da 0,1 a 3 m; TB = substrato arboreo da 3,1 a 6 m; TC = substrato arboreo > 6,1 m. Figure 5.6 Mean percent (± DS) of time spent in the different substrates from the 3 sex and age classes (adult males, adults females, juveniles). INC = anvil; CAV = cave; GR = ground; RK = rock; TA = arboreal substrate from 0,1 to 3 m; TB = arboreal substrate from 3,1 to 6 m; TC = arboreal substrate > 6,1 m.
Substrato non arboreo Substrato arboreo Classi di età/sesso
Incudine Caverna Suolo Roccia Basso (0-3m)
Medio (3,1-6m)
Alto (>6,1m)
Mad 0,02 (± 0,02)
0,02 (± 0,03)
0,15 (± 0,04)
0,10 (± 0,03)
0,31 (± 0,04)
0,29 (± 0,04)
0,12 (± 0,08)
Fad 0,01 (± 0,01)
0,02 (± 0,02)
0,19 (± 0,05)
0,09 (± 0,03)
0,31 (± 0,05)
0,26 (± 0,04)
0,12 (± 0,08)
Gio 0,01 (± 0,01)
0,02 (± 0,02)
0,16 (± 0,05)
0,08 (± 0,02)
0,35 (± 0,05)
0,27 (± 0,04)
0,11 (± 0,08)
Kruskal-Wallis (H)
2,19
0,45
3,92
2,52
5,64
1,88
0,70
p 0,33 0,80 0,14 0,28 0,06 0,39 0,70 Tabella 5.8 Frequenze medie (± DS) del tempo trascorso su substrati non arborei e su substrati arborei nelle 3 classi di età/sesso.Valore H dell’ANOVA per ranghi di Kruskal-Wallis e valore di significatività (p) del test. Mad = maschi adulti, Fad = femmine adulte, Gio = giovani. d.f. = 2; N = 36; p = 0,05. Table 5.8 Frequencies (± DS) of time spent on no-arboreal substrates and arboreal substrates in each of the 3 age/sex classes. H and p value obtained via the Kruskal-Wallis ANOVA.( Mad = adult males, Fad = adult females, Gio = juveniles). d.f. = 2; N = 36; p = 0,05.

53
5.3 Uso di strumenti
Nei 12 mesi di campionamento sono stati registrati 734 episodi di uso di strumenti per il gruppo
Chicao su un totale di 705 ore di osservazione, e 889 episodi di uso di strumenti per il gruppo
Zangado su un totale di 1004 ore di osservazione (Tabella 5.9). L’uso di strumenti è avvenuto in
tutti i mesi dell’anno con una frequenza media mensile (± deviazione standard) di episodi per ora di
1,10 (± 0,66) per il gruppo Chicao e di 0,81 (± 0,57) per il gruppo Zangado. E’ interessante notare
che per ambedue i gruppi esiste una grande variabilità mensile nel tasso di uso di strumenti e che i
valori minimi si riscontrano durante la stagione piovosa (mesi di dicembre e gennaio) mentre i
valori massimi si registrano nella stagione secca (mesi di maggio, giugno e luglio). Sulla base delle
frequenze di uso di strumenti per individuo (Tabella 5.9), e assumendo che i cebi siano attivi per
una decina di ore al giorno, è possibile stimare che l’uso di strumenti avvenga con una frequenza di
circa 1 episodio al giorno per individuo.
I cebi hanno usato percussori sia per aprire noci di palma della specie catulè (Attalea
barreirensis), piassava (Orbignya sp.), catulì (Attalea sp.) e tucum (Astrocaryum campestre) sia per
aprire altri frutti e semi (fruta-danta, fam. Icacinaceae; caju, fam. Anacardiaceae; caroba, fam.
Bignoniaceae; mandioca brava, fam. Euphorbiaceae). Inoltre in un’occasione, Zangado ha usato un
sasso per pestare la base di una foglia di Mucambira (fam. Bromeliaceae), e in un’altra occasione
una femmina adulta (Chuchu) ha usato una noce per scavare nel terreno con l’aiuto di una noce.
Dei 28 individui appartenenti ai gruppi, 22 hanno usato strumenti in maniera efficente o
adeguata (vedi 4.2.3 per definizioni) in un totale di 1565 episodi (Tabella 5.10). I 4 piccoli di meno
di un anno non hanno mai usato o tentato di usare strumenti; inoltre anche una femmina giovane
(Amarelinha) non ha mai usato o tentato di usare strumenti. Una femmina adulta (Chiquinha) in 6
episodi ha trasportato una noce tucum (Atrocaryum campestre) all’incudine e posizionato il
percussore sull’incudine poi, al posto di battere il percussore sulla noce, ha battuto ripetutamente la
noce sul percussore mostrando un uso inadeguato dello strumento. Grazie alla sua insistenza e alla
relativamente bassa resistenza della noce tucum (Visalberghi et al., 2008) in 2 episodi Chiquinha è
riuscita ad aprire la noce impiegandoci in media 175 ± 163 sec (N = 2).
In 227 episodi abbiamo anche registrato se uno o più individui usavano strumenti allo stesso
tempo. Il 23,8 % degli episodi (23,3 % quando si aprono le noci e 0,5 % quando si aprono altri
frutti) avviene con almeno due individui che allo stesso tempo usano strumenti su incudini
differenti, mentre il 76,2 % degli episodi (68,7 % per aprire le noci e 7,5 % per aprire altri frutti)
avviene in assenza di altri individui che usano strumenti. Pertanto, si può affermare che in circa un
quarto degli episodi di uso di strumenti l’attività è sincronizzata.
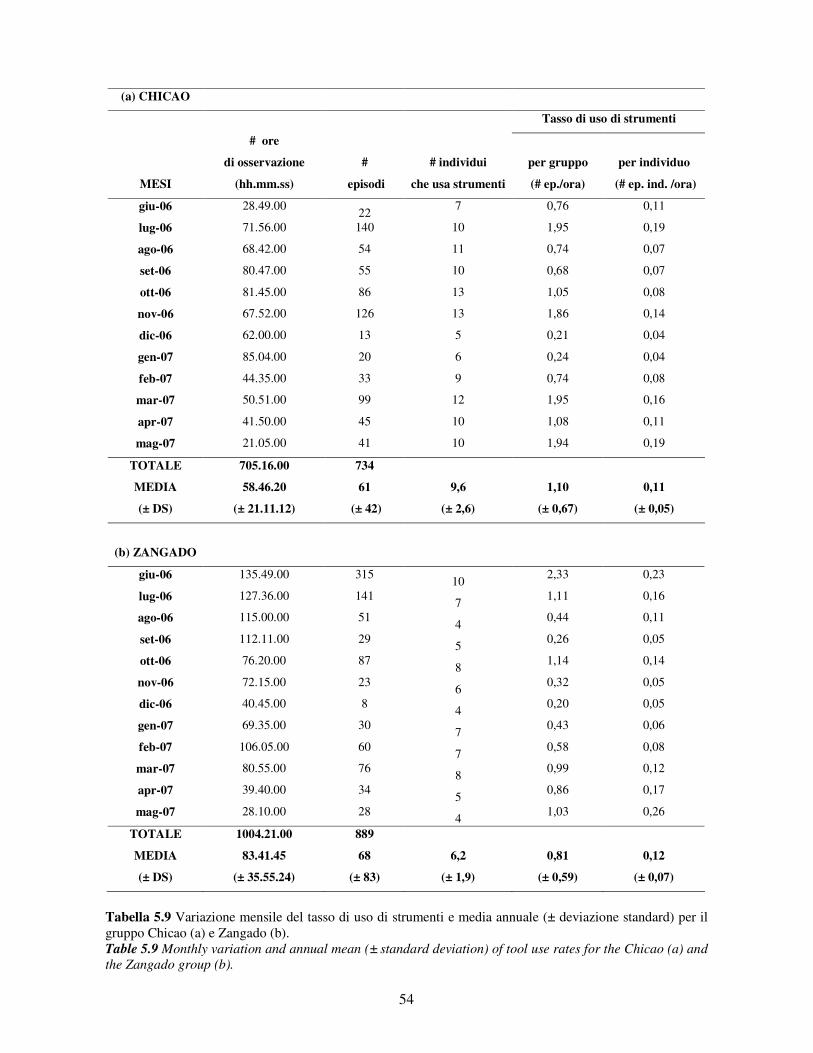
54
(a) CHICAO
Tasso di uso di strumenti
MESI
# ore
di osservazione
(hh.mm.ss)
#
episodi
# individui
che usa strumenti
per gruppo
(# ep./ora)
per individuo
(# ep. ind. /ora)
giu-06 28.49.00 22 7 0,76 0,11
lug-06 71.56.00 140 10 1,95 0,19
ago-06 68.42.00 54 11 0,74 0,07
set-06 80.47.00 55 10 0,68 0,07
ott-06 81.45.00 86 13 1,05 0,08
nov-06 67.52.00 126 13 1,86 0,14
dic-06 62.00.00 13 5 0,21 0,04
gen-07 85.04.00 20 6 0,24 0,04
feb-07 44.35.00 33 9 0,74 0,08
mar-07 50.51.00 99 12 1,95 0,16
apr-07 41.50.00 45 10 1,08 0,11
mag-07 21.05.00 41 10 1,94 0,19
TOTALE 705.16.00 734
MEDIA
(± DS)
58.46.20
(± 21.11.12)
61
(± 42)
9,6
(± 2,6)
1,10
(± 0,67)
0,11
(± 0,05)
(b) ZANGADO
giu-06 135.49.00 315 10 2,33 0,23
lug-06 127.36.00 141 7 1,11 0,16
ago-06 115.00.00 51 4 0,44 0,11
set-06 112.11.00 29 5 0,26 0,05
ott-06 76.20.00 87 8 1,14 0,14
nov-06 72.15.00 23 6 0,32 0,05
dic-06 40.45.00 8 4 0,20 0,05
gen-07 69.35.00 30 7 0,43 0,06
feb-07 106.05.00 60 7 0,58 0,08
mar-07 80.55.00 76 8 0,99 0,12
apr-07 39.40.00 34 5 0,86 0,17
mag-07 28.10.00 28 4 1,03 0,26
TOTALE 1004.21.00 889
MEDIA
(± DS)
83.41.45
(± 35.55.24)
68
(± 83)
6,2
(± 1,9)
0,81
(± 0,59)
0,12
(± 0,07)
Tabella 5.9 Variazione mensile del tasso di uso di strumenti e media annuale (± deviazione standard) per il gruppo Chicao (a) e Zangado (b). Table 5.9 Monthly variation and annual mean (± standard deviation) of tool use rates for the Chicao (a) and the Zangado group (b).
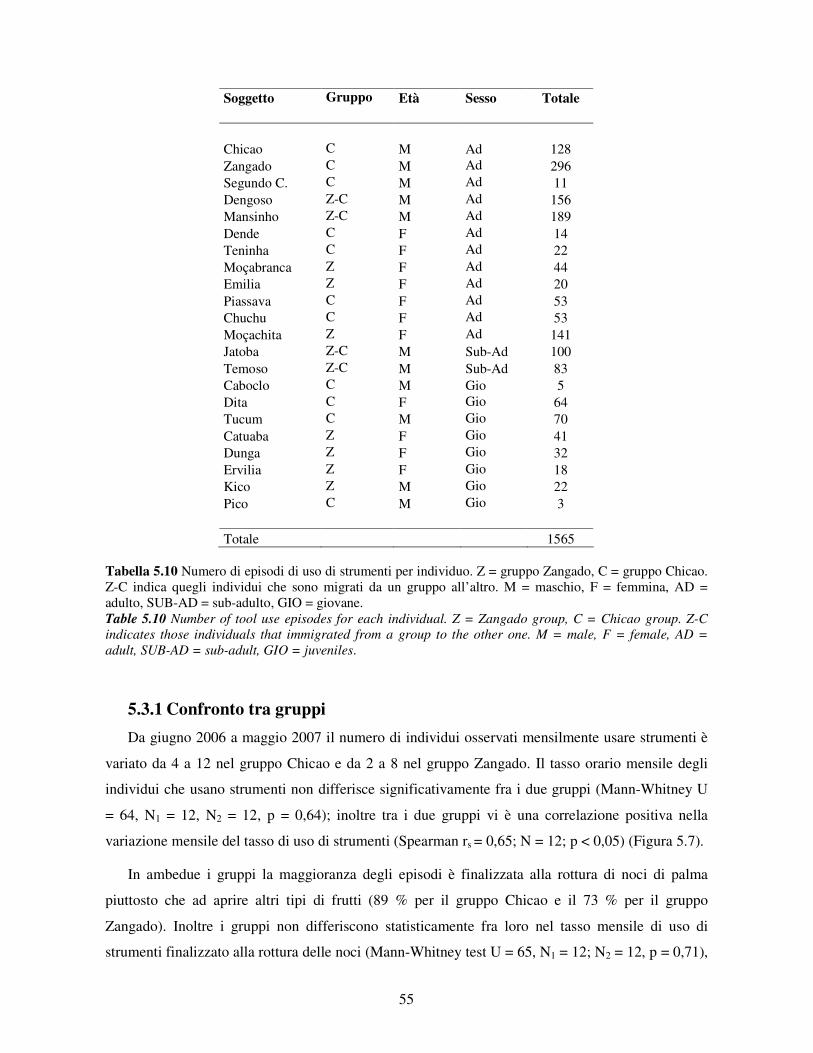
55
Soggetto Gruppo Età Sesso Totale Chicao C M Ad 128 Zangado C M Ad 296 Segundo C. C M Ad 11 Dengoso Z-C M Ad 156 Mansinho Z-C M Ad 189 Dende C F Ad 14 Teninha C F Ad 22 Moçabranca Z F Ad 44 Emilia Z F Ad 20 Piassava C F Ad 53 Chuchu C F Ad 53 Moçachita Z F Ad 141 Jatoba Z-C M Sub-Ad 100 Temoso Z-C M Sub-Ad 83 Caboclo C M Gio 5 Dita C F Gio 64 Tucum C M Gio 70 Catuaba Z F Gio 41 Dunga Z F Gio 32 Ervilia Z F Gio 18 Kico Z M Gio 22 Pico C M Gio 3 Totale 1565
Tabella 5.10 Numero di episodi di uso di strumenti per individuo. Z = gruppo Zangado, C = gruppo Chicao. Z-C indica quegli individui che sono migrati da un gruppo all’altro. M = maschio, F = femmina, AD = adulto, SUB-AD = sub-adulto, GIO = giovane. Table 5.10 Number of tool use episodes for each individual. Z = Zangado group, C = Chicao group. Z-C indicates those individuals that immigrated from a group to the other one. M = male, F = female, AD = adult, SUB-AD = sub-adult, GIO = juveniles.
5.3.1 Confronto tra gruppi Da giugno 2006 a maggio 2007 il numero di individui osservati mensilmente usare strumenti è
variato da 4 a 12 nel gruppo Chicao e da 2 a 8 nel gruppo Zangado. Il tasso orario mensile degli
individui che usano strumenti non differisce significativamente fra i due gruppi (Mann-Whitney U
= 64, N1 = 12, N2 = 12, p = 0,64); inoltre tra i due gruppi vi è una correlazione positiva nella
variazione mensile del tasso di uso di strumenti (Spearman rs = 0,65; N = 12; p < 0,05) (Figura 5.7).
In ambedue i gruppi la maggioranza degli episodi è finalizzata alla rottura di noci di palma
piuttosto che ad aprire altri tipi di frutti (89 % per il gruppo Chicao e il 73 % per il gruppo
Zangado). Inoltre i gruppi non differiscono statisticamente fra loro nel tasso mensile di uso di
strumenti finalizzato alla rottura delle noci (Mann-Whitney test U = 65, N1 = 12; N2 = 12, p = 0,71),
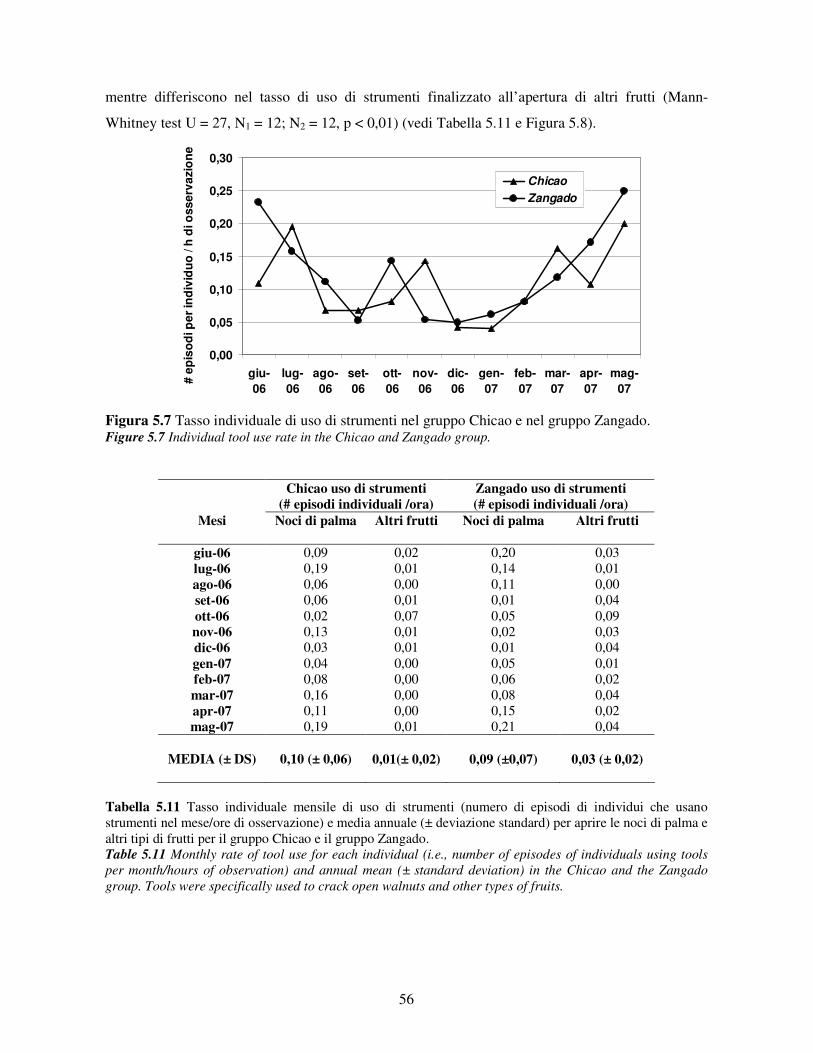
56
mentre differiscono nel tasso di uso di strumenti finalizzato all’apertura di altri frutti (Mann-
Whitney test U = 27, N1 = 12; N2 = 12, p < 0,01) (vedi Tabella 5.11 e Figura 5.8).
0,00
0,05
0,10
0,15
0,20
0,25
0,30
giu-06
lug-06
ago-06
set-06
ott-06
nov-06
dic-06
gen-07
feb-07
mar-07
apr-07
mag-07
# ep
isod
i per
ind
ivid
uo /
h d
i oss
erva
zio
ne
Chicao Zangado
Figura 5.7 Tasso individuale di uso di strumenti nel gruppo Chicao e nel gruppo Zangado. Figure 5.7 Individual tool use rate in the Chicao and Zangado group.
Chicao uso di strumenti (# episodi individuali /ora)
Zangado uso di strumenti (# episodi individuali /ora)
Mesi
Noci di palma Altri frutti Noci di palma Altri frutti
giu-06 0,09 0,02 0,20 0,03 lug-06 0,19 0,01 0,14 0,01 ago-06 0,06 0,00 0,11 0,00 set-06 0,06 0,01 0,01 0,04 ott-06 0,02 0,07 0,05 0,09 nov-06 0,13 0,01 0,02 0,03 dic-06 0,03 0,01 0,01 0,04 gen-07 0,04 0,00 0,05 0,01 feb-07 0,08 0,00 0,06 0,02 mar-07 0,16 0,00 0,08 0,04 apr-07 0,11 0,00 0,15 0,02 mag-07 0,19 0,01 0,21 0,04
MEDIA (± DS)
0,10 (± 0,06)
0,01(± 0,02)
0,09 (±0,07)
0,03 (± 0,02)
Tabella 5.11 Tasso individuale mensile di uso di strumenti (numero di episodi di individui che usano strumenti nel mese/ore di osservazione) e media annuale (± deviazione standard) per aprire le noci di palma e altri tipi di frutti per il gruppo Chicao e il gruppo Zangado. Table 5.11 Monthly rate of tool use for each individual (i.e., number of episodes of individuals using tools per month/hours of observation) and annual mean (± standard deviation) in the Chicao and the Zangado group. Tools were specifically used to crack open walnuts and other types of fruits.

57
Figura 5.8 Percentuale mensile di uso di strumenti nei due gruppi finalizzato all’apertura di noci di palma e di altri frutti. Figure 5.8 Monthly percent of tool use aimed to crack open walnuts and other fruit items in the two study groups.
5.3.2 Relazione con dati climatici e abbondanza di risorse trofiche Per il gruppo Chicao i tassi mensili di uso di strumenti (di gruppo, individuale, per aprire noci di
palma, per aprire altri frutti) non sono correlati significativamente né alla disponibilità delle risorse
alimentari (frutta, invertebrati, noci catulè e noci passava), né al clima (temperatura massima e minima,
precipitazioni) (Tabella 5.12).
Per il gruppo Zangado, il tasso di uso di strumenti individuale e quello individuale per aprire noci di
palma sono correlati positivamente alla disponibilità di noci catulè (tasso individuale: Spearman rs = 0,74;
N = 12; p < 0,01; tasso individuale di apertura noci: Spearman rs = 0,76; N = 12; p < 0,01) (Tabella
5.12 e Figura 5.9) indicando che i cebi di Zangado usano più strumenti quando le noci catulè sono più
disponibili.
0
5
10
15
20
25
30
g-06 l-06 a-06 s-06 o-06 n-06 d-06 g-07 f-07 m-07 a-07 m-07
kg/h
a
0
0,05
0,1
0,15
0,2
0,25
# episodi /h di osservazione
noce Catulè Zangado uso di strumenti
Figura 5.9 Tasso individuale di uso di strumenti per il gruppo Zangado e disponibilità delle noci catulè. Figure 5.9 Individual tool use rate for the Zangado group and availability of the catulè nuts.
Chicao
0%
20%
40%
60%
80%
100%
giu-06
lug-06
ago-
06
set-0
6ott-
06
nov-06
dic-06
gen-0
7
feb-0
7
mar-0
7
apr-0
7
mag-07
Uso
di s
trum
enti
Noci di palma Altri frutti
Zangado
0%
20%
40%
60%
80%
100%
giu-0
6
lug-06
ago-0
6
set-0
6ott-
06
nov-06
dic-06
gen-0
7
feb-0
7
mar
-07
apr-0
7
mag
-07
Uso
di s
trum
enti
Noci di palma Altri frutti
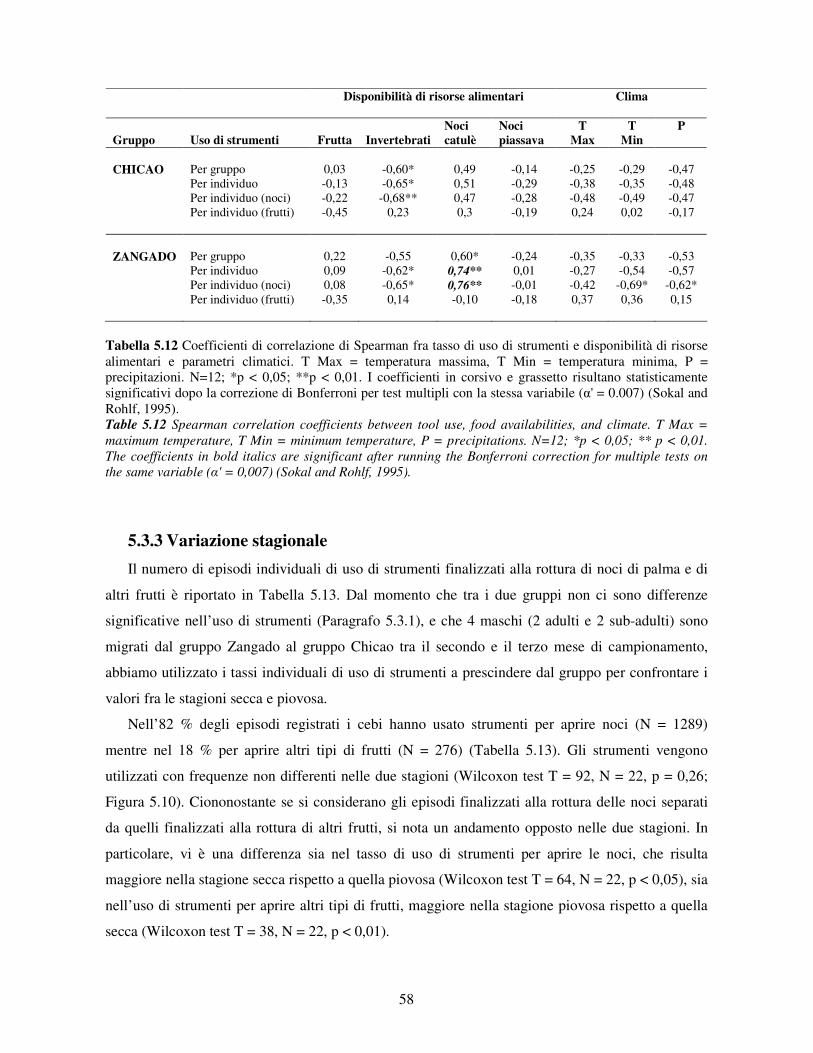
58
Disponibilità di risorse alimentari
Clima
Gruppo
Uso di strumenti
Frutta
Invertebrati
Noci catulè
Noci piassava
T Max
T Min
P
CHICAO Per gruppo 0,03 -0,60* 0,49 -0,14 -0,25 -0,29 -0,47 Per individuo -0,13 -0,65* 0,51 -0,29 -0,38 -0,35 -0,48 Per individuo (noci) -0,22 -0,68** 0,47 -0,28 -0,48 -0,49 -0,47 Per individuo (frutti) -0,45 0,23 0,3 -0,19 0,24 0,02 -0,17 ZANGADO Per gruppo 0,22 -0,55 0,60* -0,24 -0,35 -0,33 -0,53 Per individuo 0,09 -0,62* 0,74** 0,01 -0,27 -0,54 -0,57 Per individuo (noci) 0,08 -0,65* 0,76** -0,01 -0,42 -0,69* -0,62* Per individuo (frutti) -0,35 0,14 -0,10 -0,18 0,37 0,36 0,15
Tabella 5.12 Coefficienti di correlazione di Spearman fra tasso di uso di strumenti e disponibilità di risorse alimentari e parametri climatici. T Max = temperatura massima, T Min = temperatura minima, P = precipitazioni. N=12; *p < 0,05; **p < 0,01. I coefficienti in corsivo e grassetto risultano statisticamente significativi dopo la correzione di Bonferroni per test multipli con la stessa variabile (�' = 0.007) (Sokal and Rohlf, 1995). Table 5.12 Spearman correlation coefficients between tool use, food availabilities, and climate. T Max = maximum temperature, T Min = minimum temperature, P = precipitations. N=12; *p < 0,05; ** p < 0,01. The coefficients in bold italics are significant after running the Bonferroni correction for multiple tests on the same variable (�' = 0,007) (Sokal and Rohlf, 1995).
5.3.3 Variazione stagionale Il numero di episodi individuali di uso di strumenti finalizzati alla rottura di noci di palma e di
altri frutti è riportato in Tabella 5.13. Dal momento che tra i due gruppi non ci sono differenze
significative nell’uso di strumenti (Paragrafo 5.3.1), e che 4 maschi (2 adulti e 2 sub-adulti) sono
migrati dal gruppo Zangado al gruppo Chicao tra il secondo e il terzo mese di campionamento,
abbiamo utilizzato i tassi individuali di uso di strumenti a prescindere dal gruppo per confrontare i
valori fra le stagioni secca e piovosa.
Nell’82 % degli episodi registrati i cebi hanno usato strumenti per aprire noci (N = 1289)
mentre nel 18 % per aprire altri tipi di frutti (N = 276) (Tabella 5.13). Gli strumenti vengono
utilizzati con frequenze non differenti nelle due stagioni (Wilcoxon test T = 92, N = 22, p = 0,26;
Figura 5.10). Ciononostante se si considerano gli episodi finalizzati alla rottura delle noci separati
da quelli finalizzati alla rottura di altri frutti, si nota un andamento opposto nelle due stagioni. In
particolare, vi è una differenza sia nel tasso di uso di strumenti per aprire le noci, che risulta
maggiore nella stagione secca rispetto a quella piovosa (Wilcoxon test T = 64, N = 22, p < 0,05), sia
nell’uso di strumenti per aprire altri tipi di frutti, maggiore nella stagione piovosa rispetto a quella
secca (Wilcoxon test T = 38, N = 22, p < 0,01).
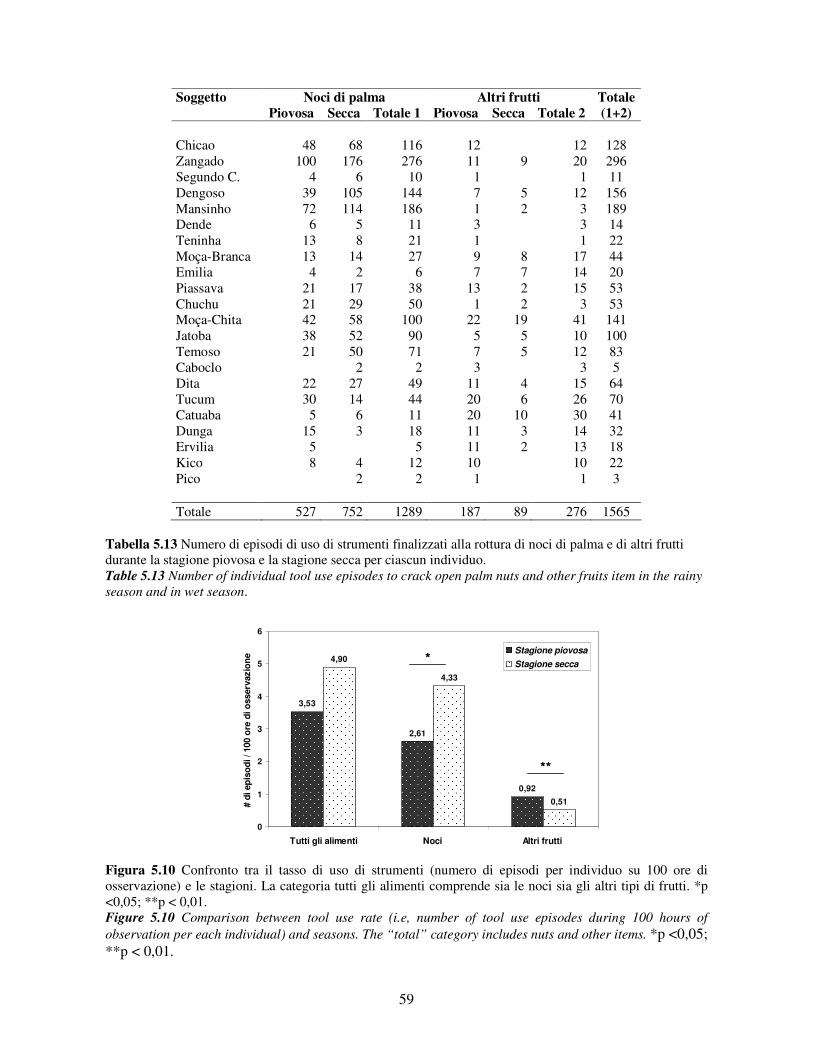
59
Soggetto Noci di palma Altri frutti Totale Piovosa Secca Totale 1 Piovosa Secca Totale 2 (1+2) Chicao 48 68 116 12 12 128 Zangado 100 176 276 11 9 20 296 Segundo C. 4 6 10 1 1 11 Dengoso 39 105 144 7 5 12 156 Mansinho 72 114 186 1 2 3 189 Dende 6 5 11 3 3 14 Teninha 13 8 21 1 1 22 Moça-Branca 13 14 27 9 8 17 44 Emilia 4 2 6 7 7 14 20 Piassava 21 17 38 13 2 15 53 Chuchu 21 29 50 1 2 3 53 Moça-Chita 42 58 100 22 19 41 141 Jatoba 38 52 90 5 5 10 100 Temoso 21 50 71 7 5 12 83 Caboclo 2 2 3 3 5 Dita 22 27 49 11 4 15 64 Tucum 30 14 44 20 6 26 70 Catuaba 5 6 11 20 10 30 41 Dunga 15 3 18 11 3 14 32 Ervilia 5 5 11 2 13 18 Kico 8 4 12 10 10 22 Pico 2 2 1 1 3 Totale 527 752 1289 187 89 276 1565
Tabella 5.13 Numero di episodi di uso di strumenti finalizzati alla rottura di noci di palma e di altri frutti durante la stagione piovosa e la stagione secca per ciascun individuo. Table 5.13 Number of individual tool use episodes to crack open palm nuts and other fruits item in the rainy season and in wet season.
3,53
2,61
0,92
4,90
4,33
0,51
0
1
2
3
4
5
6
Tutti gli alimenti Noci Altri frutti
# di
epi
sodi
/ 10
0 or
e di
oss
erva
zion
e Stagione piovosaStagione secca
**
*
3,53
2,61
0,92
4,90
4,33
0,51
0
1
2
3
4
5
6
Tutti gli alimenti Noci Altri frutti
# di
epi
sodi
/ 10
0 or
e di
oss
erva
zion
e Stagione piovosaStagione secca
**
3,53
2,61
0,92
4,90
4,33
0,51
0
1
2
3
4
5
6
Tutti gli alimenti Noci Altri frutti
# di
epi
sodi
/ 10
0 or
e di
oss
erva
zion
e Stagione piovosaStagione secca
****
**
Figura 5.10 Confronto tra il tasso di uso di strumenti (numero di episodi per individuo su 100 ore di osservazione) e le stagioni. La categoria tutti gli alimenti comprende sia le noci sia gli altri tipi di frutti. *p <0,05; **p < 0,01. Figure 5.10 Comparison between tool use rate (i.e, number of tool use episodes during 100 hours of observation per each individual) and seasons. The “total” category includes nuts and other items. *p <0,05; **p < 0,01.
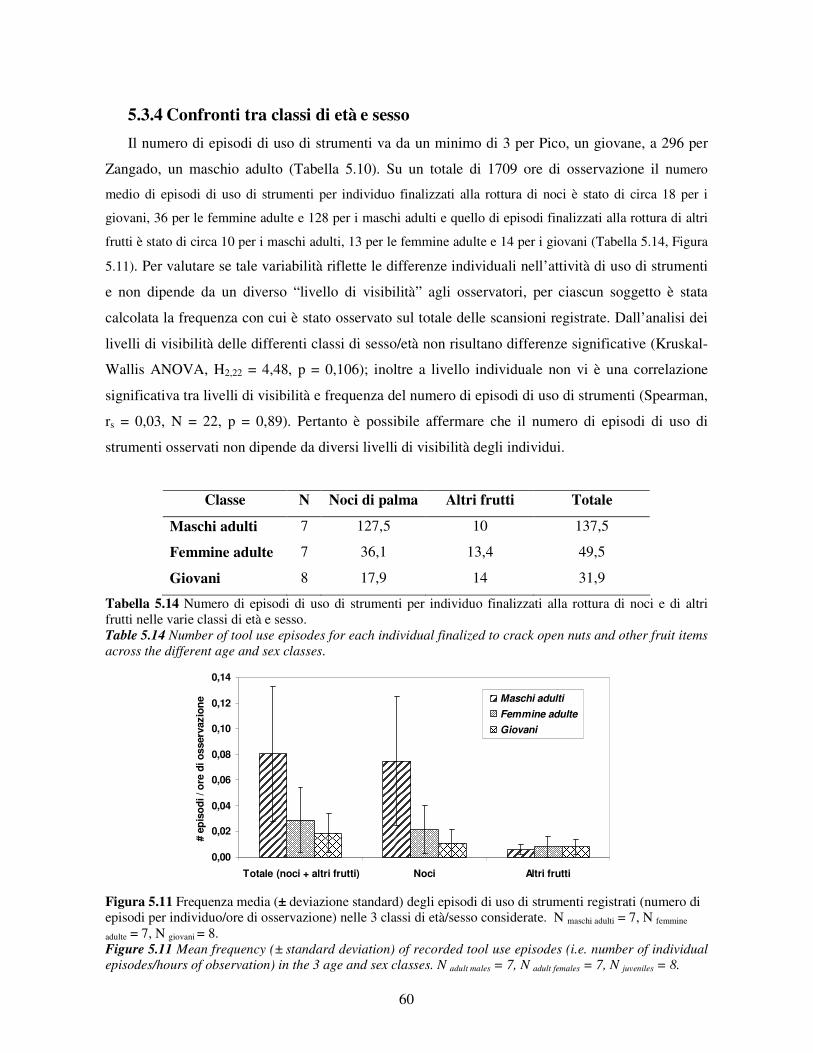
60
5.3.4 Confronti tra classi di età e sesso Il numero di episodi di uso di strumenti va da un minimo di 3 per Pico, un giovane, a 296 per
Zangado, un maschio adulto (Tabella 5.10). Su un totale di 1709 ore di osservazione il numero
medio di episodi di uso di strumenti per individuo finalizzati alla rottura di noci è stato di circa 18 per i
giovani, 36 per le femmine adulte e 128 per i maschi adulti e quello di episodi finalizzati alla rottura di altri
frutti è stato di circa 10 per i maschi adulti, 13 per le femmine adulte e 14 per i giovani (Tabella 5.14, Figura
5.11). Per valutare se tale variabilità riflette le differenze individuali nell’attività di uso di strumenti
e non dipende da un diverso “livello di visibilità” agli osservatori, per ciascun soggetto è stata
calcolata la frequenza con cui è stato osservato sul totale delle scansioni registrate. Dall’analisi dei
livelli di visibilità delle differenti classi di sesso/età non risultano differenze significative (Kruskal-
Wallis ANOVA, H2,22 = 4,48, p = 0,106); inoltre a livello individuale non vi è una correlazione
significativa tra livelli di visibilità e frequenza del numero di episodi di uso di strumenti (Spearman,
rs = 0,03, N = 22, p = 0,89). Pertanto è possibile affermare che il numero di episodi di uso di
strumenti osservati non dipende da diversi livelli di visibilità degli individui.
Classe N Noci di palma Altri frutti Totale
Maschi adulti 7 127,5 10 137,5
Femmine adulte 7 36,1 13,4 49,5
Giovani 8 17,9 14 31,9
Tabella 5.14 Numero di episodi di uso di strumenti per individuo finalizzati alla rottura di noci e di altri frutti nelle varie classi di età e sesso. Table 5.14 Number of tool use episodes for each individual finalized to crack open nuts and other fruit items across the different age and sex classes.
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
Totale (noci + altri frutti) Noci Altri frutti
# ep
isod
i / o
re d
i oss
erva
zion
e Maschi adultiFemmine adulteGiovani
Figura 5.11 Frequenza media (± deviazione standard) degli episodi di uso di strumenti registrati (numero di episodi per individuo/ore di osservazione) nelle 3 classi di età/sesso considerate. N maschi adulti = 7, N femmine
adulte = 7, N giovani = 8. Figure 5.11 Mean frequency (± standard deviation) of recorded tool use episodes (i.e. number of individual episodes/hours of observation) in the 3 age and sex classes. N adult males = 7, N adult females = 7, N juveniles = 8.
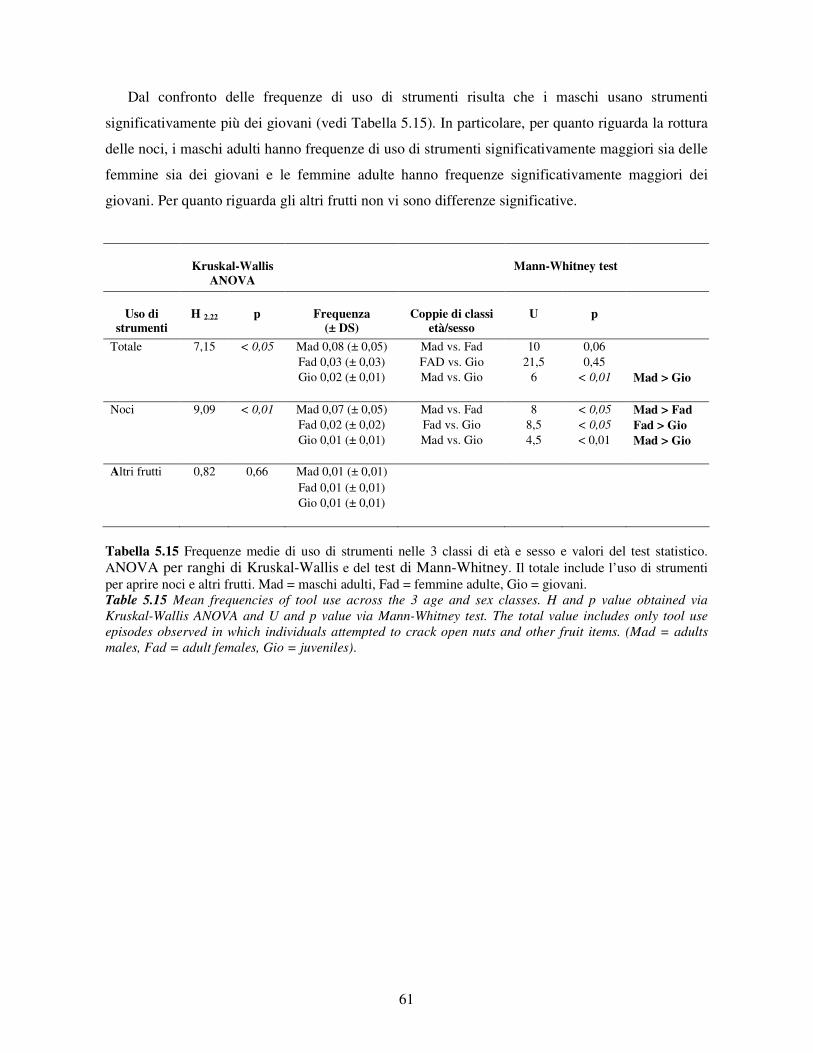
61
Dal confronto delle frequenze di uso di strumenti risulta che i maschi usano strumenti
significativamente più dei giovani (vedi Tabella 5.15). In particolare, per quanto riguarda la rottura
delle noci, i maschi adulti hanno frequenze di uso di strumenti significativamente maggiori sia delle
femmine sia dei giovani e le femmine adulte hanno frequenze significativamente maggiori dei
giovani. Per quanto riguarda gli altri frutti non vi sono differenze significative.
Kruskal-Wallis
ANOVA
Mann-Whitney test
Uso di
strumenti
H 2.22
p
Frequenza
(± DS)
Coppie di classi
età/sesso
U
p
Totale 7,15 < 0,05 Mad 0,08 (± 0,05) Mad vs. Fad 10 0,06 Fad 0,03 (± 0,03) FAD vs. Gio 21,5 0,45 Gio 0,02 (± 0,01) Mad vs. Gio 6 < 0,01 Mad > Gio Noci 9,09 < 0,01 Mad 0,07 (± 0,05) Mad vs. Fad 8 < 0,05 Mad > Fad Fad 0,02 (± 0,02) Fad vs. Gio 8,5 < 0,05 Fad > Gio Gio 0,01 (± 0,01) Mad vs. Gio 4,5 < 0,01 Mad > Gio Altri frutti 0,82 0,66 Mad 0,01 (± 0,01) Fad 0,01 (± 0,01) Gio 0,01 (± 0,01)
Tabella 5.15 Frequenze medie di uso di strumenti nelle 3 classi di età e sesso e valori del test statistico. ANOVA per ranghi di Kruskal-Wallis e del test di Mann-Whitney. Il totale include l’uso di strumenti per aprire noci e altri frutti. Mad = maschi adulti, Fad = femmine adulte, Gio = giovani. Table 5.15 Mean frequencies of tool use across the 3 age and sex classes. H and p value obtained via Kruskal-Wallis ANOVA and U and p value via Mann-Whitney test. The total value includes only tool use episodes observed in which individuals attempted to crack open nuts and other fruit items. (Mad = adults males, Fad = adult females, Gio = juveniles).
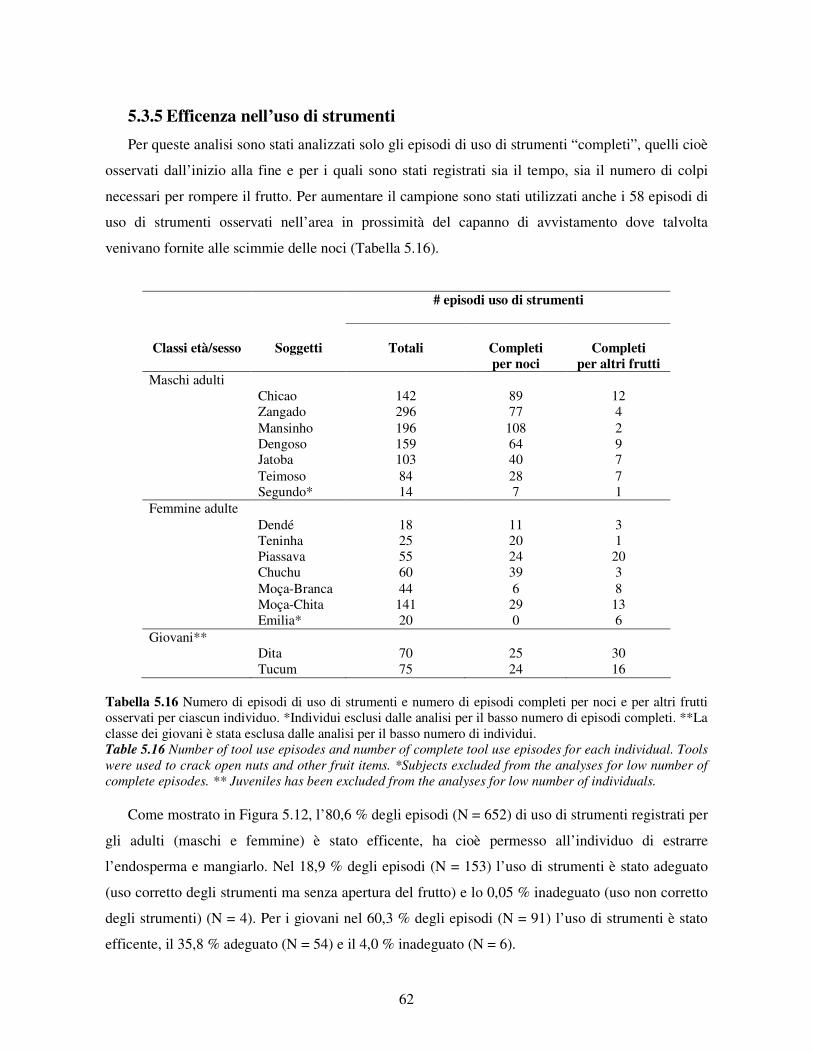
62
5.3.5 Efficenza nell’uso di strumenti Per queste analisi sono stati analizzati solo gli episodi di uso di strumenti “completi”, quelli cioè
osservati dall’inizio alla fine e per i quali sono stati registrati sia il tempo, sia il numero di colpi
necessari per rompere il frutto. Per aumentare il campione sono stati utilizzati anche i 58 episodi di
uso di strumenti osservati nell’area in prossimità del capanno di avvistamento dove talvolta
venivano fornite alle scimmie delle noci (Tabella 5.16).
# episodi uso di strumenti
Classi età/sesso
Soggetti
Totali
Completi per noci
Completi
per altri frutti Maschi adulti Chicao 142 89 12 Zangado 296 77 4 Mansinho 196 108 2 Dengoso 159 64 9 Jatoba 103 40 7 Teimoso 84 28 7 Segundo* 14 7 1 Femmine adulte Dendé 18 11 3 Teninha 25 20 1 Piassava 55 24 20 Chuchu 60 39 3 Moça-Branca 44 6 8 Moça-Chita 141 29 13 Emilia* 20 0 6 Giovani** Dita 70 25 30 Tucum 75 24 16
Tabella 5.16 Numero di episodi di uso di strumenti e numero di episodi completi per noci e per altri frutti osservati per ciascun individuo. *Individui esclusi dalle analisi per il basso numero di episodi completi. **La classe dei giovani è stata esclusa dalle analisi per il basso numero di individui. Table 5.16 Number of tool use episodes and number of complete tool use episodes for each individual. Tools were used to crack open nuts and other fruit items. *Subjects excluded from the analyses for low number of complete episodes. ** Juveniles has been excluded from the analyses for low number of individuals.
Come mostrato in Figura 5.12, l’80,6 % degli episodi (N = 652) di uso di strumenti registrati per
gli adulti (maschi e femmine) è stato efficente, ha cioè permesso all’individuo di estrarre
l’endosperma e mangiarlo. Nel 18,9 % degli episodi (N = 153) l’uso di strumenti è stato adeguato
(uso corretto degli strumenti ma senza apertura del frutto) e lo 0,05 % inadeguato (uso non corretto
degli strumenti) (N = 4). Per i giovani nel 60,3 % degli episodi (N = 91) l’uso di strumenti è stato
efficente, il 35,8 % adeguato (N = 54) e il 4,0 % inadeguato (N = 6).
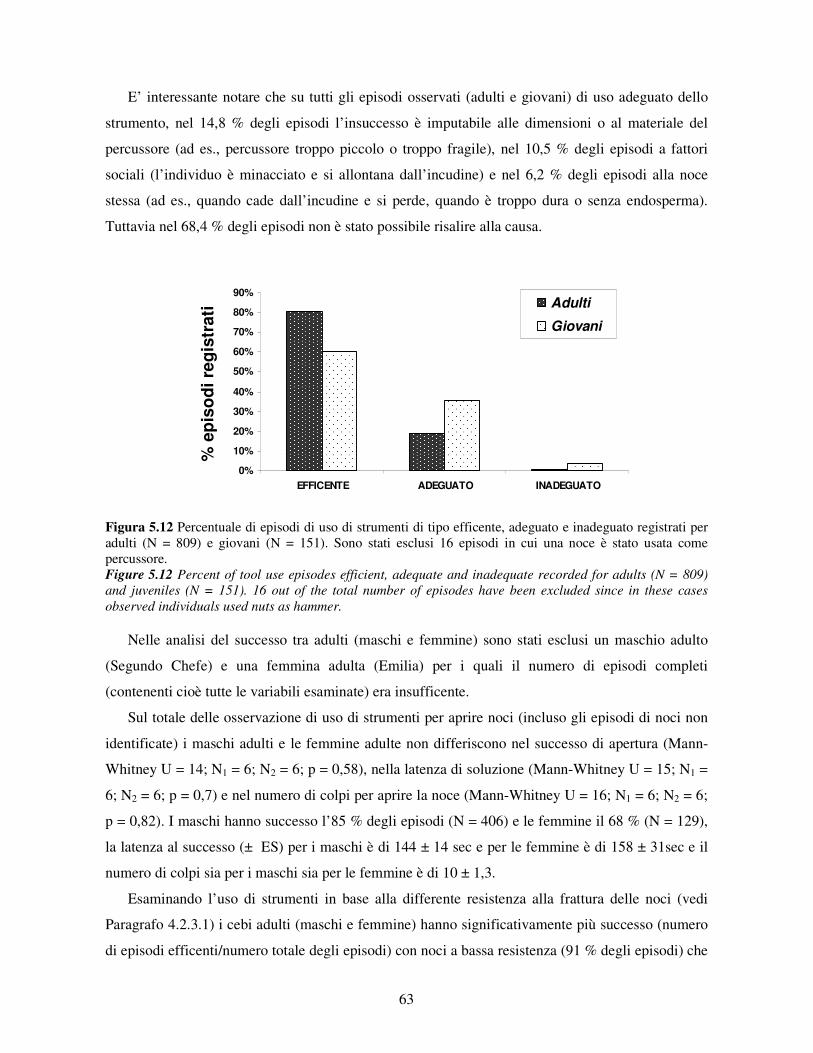
63
E’ interessante notare che su tutti gli episodi osservati (adulti e giovani) di uso adeguato dello
strumento, nel 14,8 % degli episodi l’insuccesso è imputabile alle dimensioni o al materiale del
percussore (ad es., percussore troppo piccolo o troppo fragile), nel 10,5 % degli episodi a fattori
sociali (l’individuo è minacciato e si allontana dall’incudine) e nel 6,2 % degli episodi alla noce
stessa (ad es., quando cade dall’incudine e si perde, quando è troppo dura o senza endosperma).
Tuttavia nel 68,4 % degli episodi non è stato possibile risalire alla causa.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
EFFICENTE ADEGUATO INADEGUATO
% e
piso
di r
egis
trat
i Adulti
Giovani
Figura 5.12 Percentuale di episodi di uso di strumenti di tipo efficente, adeguato e inadeguato registrati per adulti (N = 809) e giovani (N = 151). Sono stati esclusi 16 episodi in cui una noce è stato usata come percussore. Figure 5.12 Percent of tool use episodes efficient, adequate and inadequate recorded for adults (N = 809) and juveniles (N = 151). 16 out of the total number of episodes have been excluded since in these cases observed individuals used nuts as hammer.
Nelle analisi del successo tra adulti (maschi e femmine) sono stati esclusi un maschio adulto
(Segundo Chefe) e una femmina adulta (Emilia) per i quali il numero di episodi completi
(contenenti cioè tutte le variabili esaminate) era insufficente.
Sul totale delle osservazione di uso di strumenti per aprire noci (incluso gli episodi di noci non
identificate) i maschi adulti e le femmine adulte non differiscono nel successo di apertura (Mann-
Whitney U = 14; N1 = 6; N2 = 6; p = 0,58), nella latenza di soluzione (Mann-Whitney U = 15; N1 =
6; N2 = 6; p = 0,7) e nel numero di colpi per aprire la noce (Mann-Whitney U = 16; N1 = 6; N2 = 6;
p = 0,82). I maschi hanno successo l’85 % degli episodi (N = 406) e le femmine il 68 % (N = 129),
la latenza al successo (± ES) per i maschi è di 144 ± 14 sec e per le femmine è di 158 ± 31sec e il
numero di colpi sia per i maschi sia per le femmine è di 10 ± 1,3.
Esaminando l’uso di strumenti in base alla differente resistenza alla frattura delle noci (vedi
Paragrafo 4.2.3.1) i cebi adulti (maschi e femmine) hanno significativamente più successo (numero
di episodi efficenti/numero totale degli episodi) con noci a bassa resistenza (91 % degli episodi) che
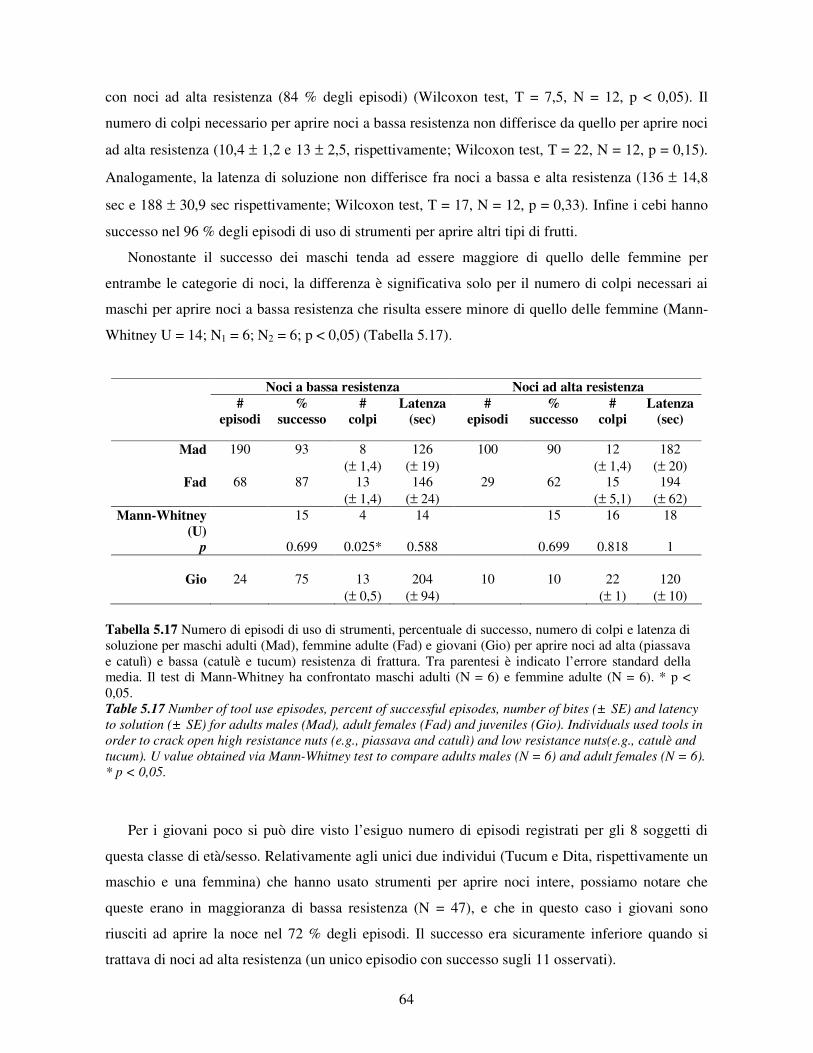
64
con noci ad alta resistenza (84 % degli episodi) (Wilcoxon test, T = 7,5, N = 12, p < 0,05). Il
numero di colpi necessario per aprire noci a bassa resistenza non differisce da quello per aprire noci
ad alta resistenza (10,4 ± 1,2 e 13 ± 2,5, rispettivamente; Wilcoxon test, T = 22, N = 12, p = 0,15).
Analogamente, la latenza di soluzione non differisce fra noci a bassa e alta resistenza (136 ± 14,8
sec e 188 ± 30,9 sec rispettivamente; Wilcoxon test, T = 17, N = 12, p = 0,33). Infine i cebi hanno
successo nel 96 % degli episodi di uso di strumenti per aprire altri tipi di frutti.
Nonostante il successo dei maschi tenda ad essere maggiore di quello delle femmine per
entrambe le categorie di noci, la differenza è significativa solo per il numero di colpi necessari ai
maschi per aprire noci a bassa resistenza che risulta essere minore di quello delle femmine (Mann-
Whitney U = 14; N1 = 6; N2 = 6; p < 0,05) (Tabella 5.17).
Noci a bassa resistenza Noci ad alta resistenza #
episodi %
successo #
colpi Latenza
(sec) #
episodi %
successo #
colpi Latenza
(sec)
Mad 190 93 8 (± 1,4)
126 (± 19)
100 90 12 (± 1,4)
182 (± 20)
Fad 68 87 13 (± 1,4)
146 (± 24)
29 62 15 (± 5,1)
194 (± 62)
Mann-Whitney (U)
15 4 14 15 16 18
p 0.699 0.025* 0.588 0.699 0.818 1
Gio 24 75 13 (± 0,5)
204 (± 94)
10 10 22 (± 1)
120 (± 10)
Tabella 5.17 Numero di episodi di uso di strumenti, percentuale di successo, numero di colpi e latenza di soluzione per maschi adulti (Mad), femmine adulte (Fad) e giovani (Gio) per aprire noci ad alta (piassava e catulì) e bassa (catulè e tucum) resistenza di frattura. Tra parentesi è indicato l’errore standard della media. Il test di Mann-Whitney ha confrontato maschi adulti (N = 6) e femmine adulte (N = 6). * p < 0,05. Table 5.17 Number of tool use episodes, percent of successful episodes, number of bites (± SE) and latency to solution (± SE) for adults males (Mad), adult females (Fad) and juveniles (Gio). Individuals used tools in order to crack open high resistance nuts (e.g., piassava and catulì) and low resistance nuts(e.g., catulè and tucum). U value obtained via Mann-Whitney test to compare adults males (N = 6) and adult females (N = 6). * p < 0,05.
Per i giovani poco si può dire visto l’esiguo numero di episodi registrati per gli 8 soggetti di
questa classe di età/sesso. Relativamente agli unici due individui (Tucum e Dita, rispettivamente un
maschio e una femmina) che hanno usato strumenti per aprire noci intere, possiamo notare che
queste erano in maggioranza di bassa resistenza (N = 47), e che in questo caso i giovani sono
riusciti ad aprire la noce nel 72 % degli episodi. Il successo era sicuramente inferiore quando si
trattava di noci ad alta resistenza (un unico episodio con successo sugli 11 osservati).
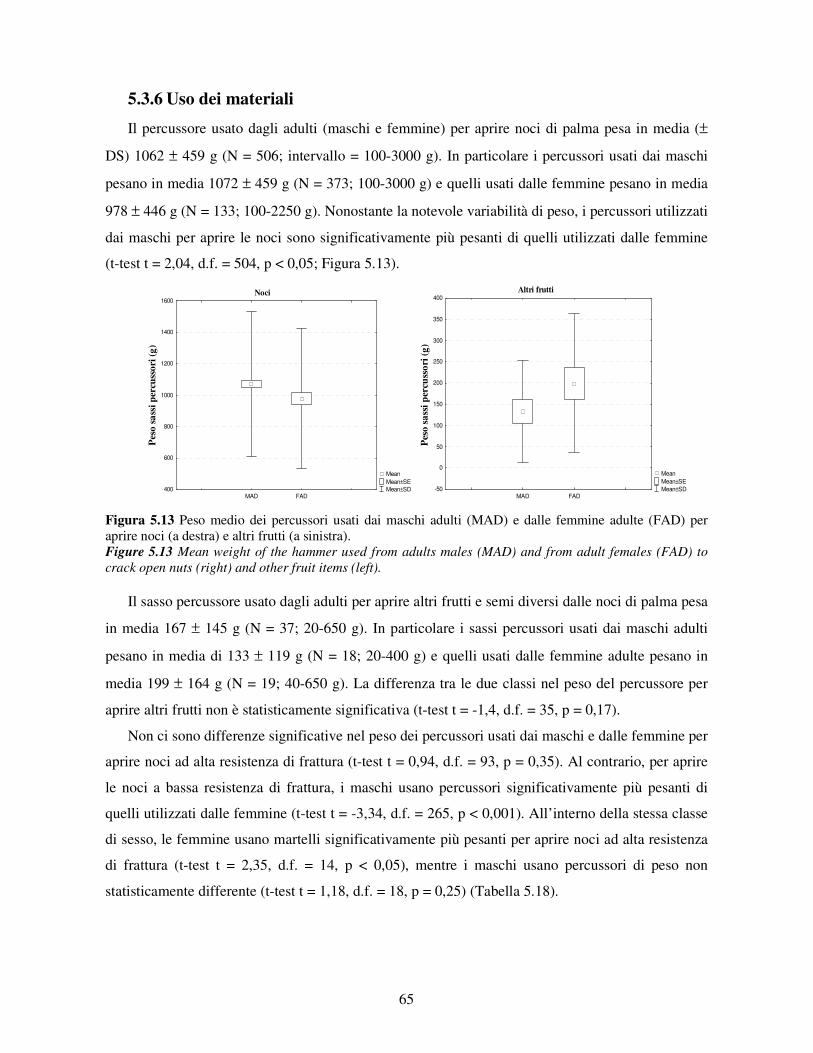
65
5.3.6 Uso dei materiali
Il percussore usato dagli adulti (maschi e femmine) per aprire noci di palma pesa in media (±
DS) 1062 ± 459 g (N = 506; intervallo = 100-3000 g). In particolare i percussori usati dai maschi
pesano in media 1072 ± 459 g (N = 373; 100-3000 g) e quelli usati dalle femmine pesano in media
978 ± 446 g (N = 133; 100-2250 g). Nonostante la notevole variabilità di peso, i percussori utilizzati
dai maschi per aprire le noci sono significativamente più pesanti di quelli utilizzati dalle femmine
(t-test t = 2,04, d.f. = 504, p < 0,05; Figura 5.13).
Noci
Mean Mean±SE Mean±SD
MAD FAD400
600
800
1000
1200
1400
1600
Peso
sass
i per
cuss
ori (
g)
Altri frutti
Mean Mean±SE Mean±SD
MAD FAD-50
0
50
100
150
200
250
300
350
400
Peso
sass
i per
cuss
ori (
g)
Figura 5.13 Peso medio dei percussori usati dai maschi adulti (MAD) e dalle femmine adulte (FAD) per aprire noci (a destra) e altri frutti (a sinistra). Figure 5.13 Mean weight of the hammer used from adults males (MAD) and from adult females (FAD) to crack open nuts (right) and other fruit items (left).
Il sasso percussore usato dagli adulti per aprire altri frutti e semi diversi dalle noci di palma pesa
in media 167 ± 145 g (N = 37; 20-650 g). In particolare i sassi percussori usati dai maschi adulti
pesano in media di 133 ± 119 g (N = 18; 20-400 g) e quelli usati dalle femmine adulte pesano in
media 199 ± 164 g (N = 19; 40-650 g). La differenza tra le due classi nel peso del percussore per
aprire altri frutti non è statisticamente significativa (t-test t = -1,4, d.f. = 35, p = 0,17).
Non ci sono differenze significative nel peso dei percussori usati dai maschi e dalle femmine per
aprire noci ad alta resistenza di frattura (t-test t = 0,94, d.f. = 93, p = 0,35). Al contrario, per aprire
le noci a bassa resistenza di frattura, i maschi usano percussori significativamente più pesanti di
quelli utilizzati dalle femmine (t-test t = -3,34, d.f. = 265, p < 0,001). All’interno della stessa classe
di sesso, le femmine usano martelli significativamente più pesanti per aprire noci ad alta resistenza
di frattura (t-test t = 2,35, d.f. = 14, p < 0,05), mentre i maschi usano percussori di peso non
statisticamente differente (t-test t = 1,18, d.f. = 18, p = 0,25) (Tabella 5.18).
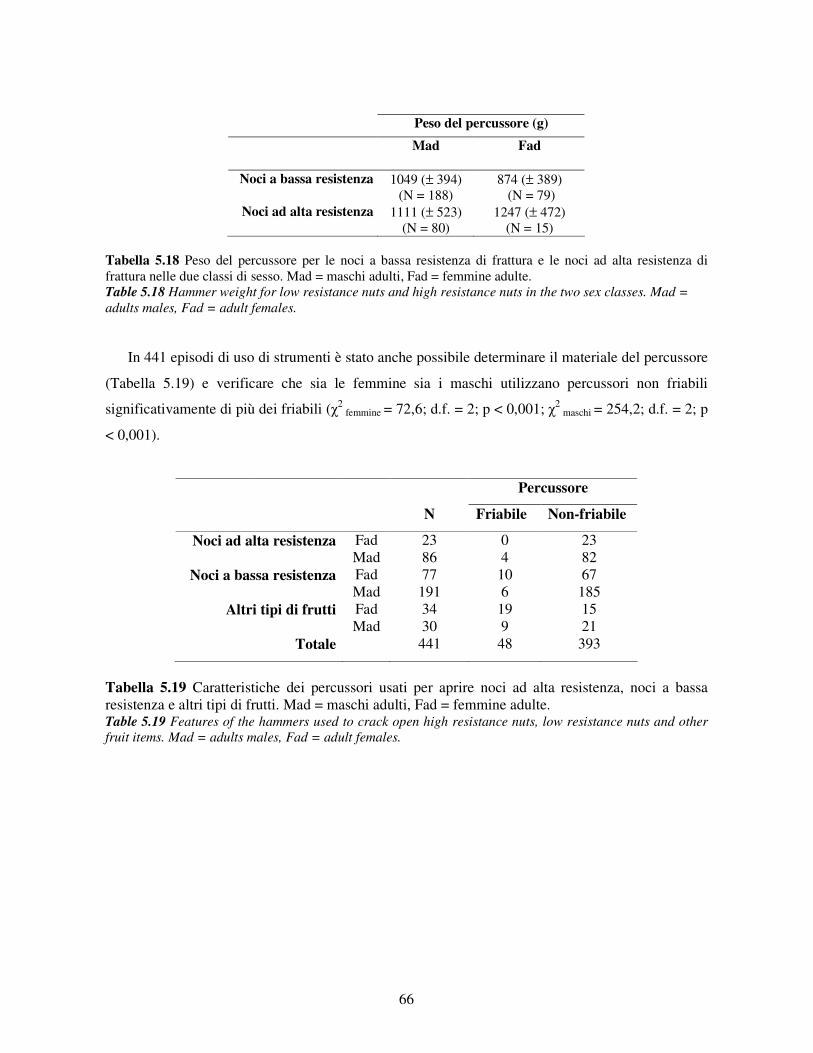
66
Peso del percussore (g) Mad Fad
Noci a bassa resistenza 1049 (± 394) (N = 188)
874 (± 389) (N = 79)
Noci ad alta resistenza 1111 (± 523) (N = 80)
1247 (± 472) (N = 15)
Tabella 5.18 Peso del percussore per le noci a bassa resistenza di frattura e le noci ad alta resistenza di frattura nelle due classi di sesso. Mad = maschi adulti, Fad = femmine adulte. Table 5.18 Hammer weight for low resistance nuts and high resistance nuts in the two sex classes. Mad = adults males, Fad = adult females.
In 441 episodi di uso di strumenti è stato anche possibile determinare il materiale del percussore
(Tabella 5.19) e verificare che sia le femmine sia i maschi utilizzano percussori non friabili
significativamente di più dei friabili (�2 femmine
= 72,6; d.f. = 2; p < 0,001; �2 maschi
= 254,2; d.f. = 2; p
< 0,001).
Percussore
N Friabile Non-friabile
Fad 23 0 23 Noci ad alta resistenza Mad 86 4 82 Fad 77 10 67 Noci a bassa resistenza Mad 191 6 185 Fad 34 19 15 Altri tipi di frutti Mad 30 9 21
Totale 441 48 393
Tabella 5.19 Caratteristiche dei percussori usati per aprire noci ad alta resistenza, noci a bassa resistenza e altri tipi di frutti. Mad = maschi adulti, Fad = femmine adulte. Table 5.19 Features of the hammers used to crack open high resistance nuts, low resistance nuts and other fruit items. Mad = adults males, Fad = adult females.

67
6. DISCUSSIONE
6.1 Disponibilità delle risorse
I parametri climatici (precipitazioni, temperatura, umidità) da noi rilevati ben si accordano con
quelli forniti dall’Embrapa (la Banca di dati climatici del governo brasiliano). A Boa Vista vi è una
marcata variazione stagionale determinata dalla persistente mancanza di precipitazioni durante
molti mesi. In base a ciò è possibile definire una stagione secca (di circa 7 mesi con precipitazioni
medie mensili di 10 mm) e una stagione piovosa (di circa 5 mesi con precipitazioni medie mensili
di 155 mm).
Nella fascia tropicale, alcuni ambienti occupati dal genere Cebus, come la foresta atlantica e la
foresta tropicale primaria, hanno una diversità ambientale relativamente alta e la disponibilità delle
risorse alimentari e in particolare di frutta, sono influenzate da una forte variazione stagionale
(Galetti & Pedroni, 1994; Zang, 1995; Nakai, 2007). Al contrario di ciò che ci aspettavamo, gli
indici di abbondanza delle risorse di frutta e di invertebrati nel corso dell’anno ottenuti con le
trappole a caduta (Izar, 2004) non hanno permesso di evidenziare una differenza fra stagioni, anche
se è stata rilevata una grande variazione mensile nell’abbondaza di queste risorse. Anche altri studi
basati sulla fenologia delle piante e la loro produttività hanno mostrato che in habitat simili a quello
di Boa Vista la produttività non differisce tra stagioni. Infatti, nelle foreste secche tropicali come la
caatinga, nonostante la diversità ambientale sia più bassa rispetto alle foreste umide (Murpho &
Lugo, 1986), alcuni alimenti, soprattutto i semi a dispersione zoocora, rimangono piuttosto
disponibili durante tutto l’anno (Moura, 2004). I dati fenologici da noi raccolti per le palme
mostrano invece che il catulé (Attalea sp.) fruttifica maggiormente nella stagione secca rispetto a
quella piovosa, mentre la piassava (Orbignya sp.) fruttifica durante tutto l’anno.
Pertanto, sebbene Boa Vista sia durante la stagione secca un ambiente arido, caldo e inospitale,
non sembrerebbe che ciò comporti una scarsità di risorse alimentari come spesso supposto (Moura,
2004; Fragaszy et al., 2004b). Inoltre grazie alla grande flessibilità comportamentale, i cebi
sfruttano le risorse disponibili e le zone umide e fresche (come le caverne o il letto sabbioso e
ombreggiato di fiumi in secca) per ripararsi nei momenti di massima calura.

68
6.2 Pattern di attività
6.2.1 Variabilità tra gruppi e tra classi di età e sesso Le percentuali di tempo trascorse nelle diverse attività per entrambi i gruppi (Tabella 6.1) non si
discostano molto dai valori riportati per popolazioni della stessa specie che vivono in ambienti
semi-aridi come la caatinga del Parco Nazionale di Serra da Capivara (Moura, 2004b). Pattern di
attività simili si ritrovano anche in gruppi di cebi che vivono in foreste tropicali semidecidue o
secche sottoposte a variazioni stagionali nella disponibilità delle risorse (Robinson 1981, 1986;
Rose, 1994; Miller, 1997). In tutte queste popolazioni i cebi trascorrono quasi il 45 % del tempo in
attività alimentari (incluso la ricerca del cibo), il 30 % in attività di movimento, il 20 % in attività di
riposo e il restante 5 % in attività sociali. Infine le popolazioni che abitano in foreste umide tropicali
mostrano pattern di attività simili, ma trascorrono più tempo in attività finalizzate all’alimentazione
(più del 60 %) a discapito delle attività sociali e di riposo (Terborgh, 1983).
I pattern mensili delle attività alimentari (mangiare e ricercare il cibo) hanno lo stesso
andamento nei due gruppi. Ciò sta ad indicare che il cibo supplementare ricevuto da Chicao non
influenza il suo comportamento alimentare. Inoltre i budget di attività di Chicao e di Zangado sono
abbastanza simili e ciò conferma che le attività di gruppi che ricevono cibo supplementare non sono
molto diverse da quelle di gruppi che non le ricevono (Ferreira et al., 2008). Le uniche differenze
riscontrate sono l’attività di movimento, maggiore nel gruppo Zangado, e l’attività sociale,
maggiore nel gruppo Chicao. In questo caso le differenze possono essere imputabili al supplemento
di cibo fornito al gruppo Chicao, che rende la necessità di spostarsi alla ricerca di cibo meno
impellente che per il gruppo Zangado. Il gruppo Chicao infatti, dopo aver ricevuto la sua razione
giornaliera di alimenti, tende a rimanere nell’area dell’esconderijo per riposarsi, pulirsi il pelo
reciprocamente (grooming) e giocare.
I confronti dei pattern di attività tra le classi di sesso/età mostrano che esistono differenze tra
maschi adulti, femmine adulte e giovani. I maschi adulti trascorrono tra il 13 % e il 17 % del tempo
in più in attività di riposo rispetto alle femmine e ai giovani. Inoltre per il 2 % del tempo usano
strumenti, mentre le femmine e i giovani lo fanno solo nell’1 %. Le femmine adulte trascorrono tra
il 4 % e il 5 % in più del loro tempo spostandosi rispetto ai maschi adulti e ai giovani. I giovani
invece trascorrono più tempo degli adulti sia in attività alimentari (4-8 % in più rispetto alle
femmine e ai maschi) sia in attività sociali (4-5 % in più rispetto alle femmine e ai maschi). Infine le
femmine adulte e i giovani trascorrono il 4-6 % di tempo in più alla ricerca di cibo di quanto non
facciano i maschi adulti. Anche in C. olivaceus, che vive nelle foreste semidecidue secche tropicali
dove è forte la differenza stagionale nelle risorse (Robinson, 1986), i giovani e le femmine
trascorrono più tempo dei maschi in attività alimentari e di foraggiamento.

69
Specie Habitat MV
(%)
RP
(%)
AL
(%)
FR
(%)
SC
(%)
Fonte
C. libidinosus Ecotono tra
cerrado/caatinga
(savana/foresta secca)
25 19 23 22** 8 Questo studio, gruppo Chicao
(BV, PI, Brasile)
C. libidinosus Ecotono tra
cerrado/caatinga
(savana/foresta secca)
29,5 16 25 22** 6
Questo studio, gruppo
Zangado
(BV, PI, Brasile)
C. libidinosus Cerrado
(savana) 41 15 17 17 8
Sabbatini (2008)
(PNB, DF, Brasile)
C. libidinosus Caatinga
(foresta secca) 37 ~ 5,5 20 22 ~ 6,5
Moura (2004)
(PNSC, PI, Brasile)
C. olivaceus Llanos (prateria umida)
alternata a foresta a
galleria
~ 20 ~ 10 ~ 62* ~ 8 Robinson (1986, 1981)
(Venezuela)
C. olivaceus Foresta semidecidua
secca tropicale 22 25 17 19 2
Miller (1997)
(Venezuela)
C. capucinus Foresta tropicale secca ~ 18 ~ 20 ~ 57* ~ 5
Rose (1994)
(SRNP, Costa Rica)
C. apella Foresta umida tropicale 21 12 16 50 < 1
Terborgh (1983)
(MNP, Peru)
C. albifrons Foresta umida tropicale 21 18 22 39 ~ 0
Terborgh (1983)
(Peru)
Tabella 6.1 Percentuale dei budget di attività in alcune popolazioni selvatiche di C. libidinosus e di altre specie del genere Cebus osservate in differenti habitat. MV = movimento, RP = riposo, AL = alimentazione, FR = foraggiamento, SC = attività sociali. In alcuni studi il totale delle attività non è pari a 100 perchè in tabella non è riportata la categoria “altre attività”. * il valore include attività di alimentazione e di foraggiamento; ** il valore include le attività di uso di strumenti. Table 6.1 Percent of activity budget in wild populations of C. libidinosus and other species among Cebus observed in different habitat. (sampled activity: MV = movement, RP = rest, AL = feeding, FR = foraging, SC = social activities). For some studies the total percent of the activities differs from 100, because the category “other activities” was excluded from this analysis. * the value includes foraging and feeding activity; ** the value includes tool use activities.
Il fatto che i maschi riposino di più e le femmine e i giovani siano più attivi nei comportamenti
alimentari è in contrasto con le teorie ecologiche secondo cui le richieste energetiche sono
proporzionali alla dimensione del corpo, per cui gli animali di maggiori dimensioni dovrebbero
trascorrere più tempo nelle attività di foraggiamento (McNab, 1983) e suggerisce che i maschi
abbiano una strategia alimentare più efficente, avendo accesso ad alimenti di maggiore qualità che
richiedono tempi minori di processamento (Fragaszy, 1986).
Le differenze sessuali, soprattutto per quanto riguarda il tempo dedicato al foraggiamento sono
riconducibili al dimorfismo sessuale e alle differenti richieste energetiche (Rose, 1994). I maschi

70
del genere Cebus sono circa il 25-30 % più grandi delle femmine (Fragaszy et al., 2004a,
EthoCebus dati non pubblicati), per cui per soddisfare i loro bisogni energetici dovrebbero
trascorrere più tempo alla ricerca di cibo (o mangiando). Le femmine d’altra parte, visto il maggiore
bisogno energetico relativo ai costi dell’allattamento e/o della gestazione (delle 8 femmine adulte
monitorate durante lo studio 2 allattavano e 4 hanno partorito), sarebbero spinte a trascorrere più
tempo nelle attività alimentari (Fedigan, 1993, Rose, 1994). Allo stesso modo i giovani che
necessitano di energie per lo sviluppo e la crescita (Gunst et al., 2008). I risultati di questo studio
sarebbero a favore della seconda ipotesi in cui sono le femmine e i giovani a trascorrere più tempo
nelle attività di foraggiamento rispetto ai maschi e in accordo con i risultati di alcune popolazioni di
C. olivaceus e C. capucinus (Fragaszy, 1990; Rose, 1994). E’ interessante riportare che nella
popolazione di C. olivaceus studiata da Robinson (1986) la risposta energetica delle femmine
sembra essere legata alla produttività stagionale delle risorse: le femmine gestanti aumentano lo
sforzo di foraggiamento durante la stagione secca (quando ci sono meno risorse), al contrario di
quanto fanno le femmine che allattano che trascorrono meno tempo foraggiando durante la stagione
piovosa ricca di risorse (Robinson, 1986). Risultati così diversi sottolineano ancora una volta la
grande flessibilità e plasticità comportamentale nel genere Cebus.
A Boa Vista, studi futuri saranno necessari a verificare le ipotesi sulle differenze sessuali e la
competizione intraspecifica in termini di dieta e di comportamento alimentare, per verificare le
risposte individuali e gli adattamenti di C. libidinosus che vive negli ambienti semi-aridi di caatinga
e cerrado.
6.2.2 Variazione stagionale I budget di attività di entrambi i gruppi non differiscono tra stagione secca e piovosa. D’altra
parte i risultati di questo studio hanno messo in evidenza che a Boa Vista nelle due stagioni definite
in base alle precipitazioni, l’abbondanza di risorse alimentari (specialmente la frutta) varia molto tra
i mesi ma non differisce fra stagioni. Ciononostante i cebi tendono a trascorrere maggior tempo
riposandosi nei mesi in cui la temperatura è più elevata, sfruttando, come già accennato, i luoghi
umidi e freschi quali le caverne, i letti sabbiosi dei fiumi in secca e le zone ombreggiate dei
canaloni. L’assenza di differenze stagionali nei budget delle attività alimentari, potrebbe riflettere
l’assenza di stagioni nella disponibilità delle risorse. Ricerche sul comportamento alimentare dei
cebi, incluso lo studio dettagliato della dieta, della distribuzione delle risorse e della loro fenologia
potranno meglio rendere conto di come questi cebi si siano adattati all’ambiente di Boa Vista.
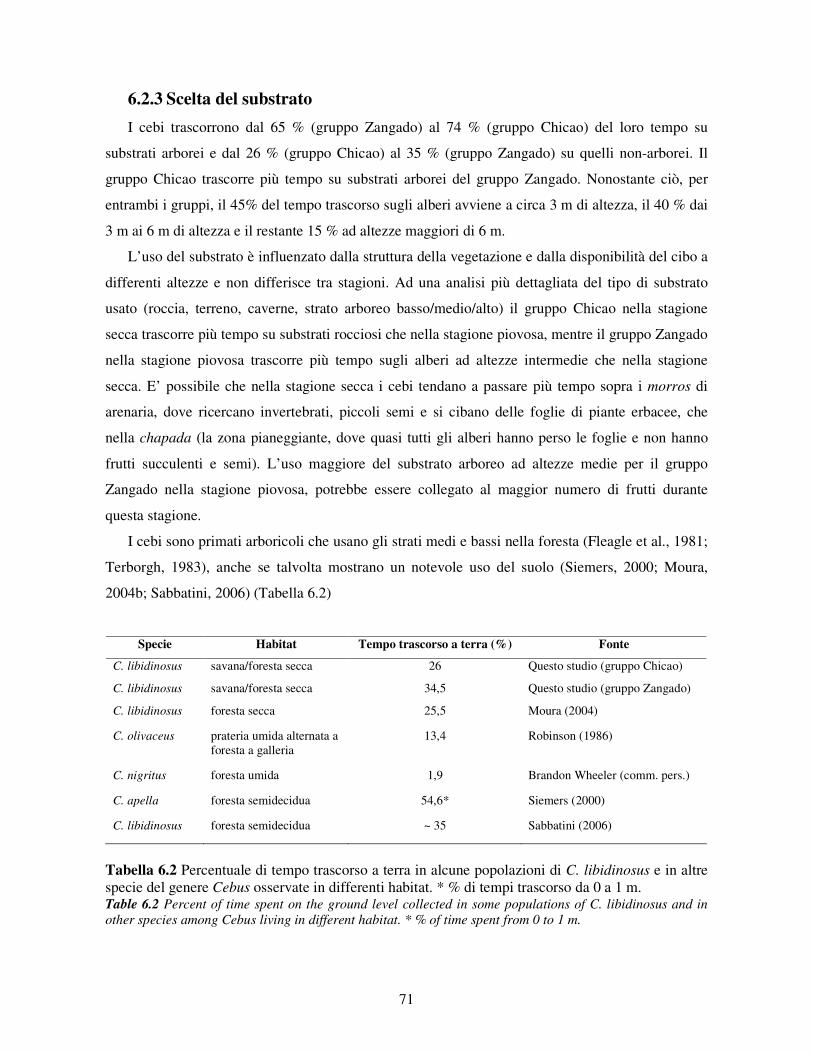
71
6.2.3 Scelta del substrato I cebi trascorrono dal 65 % (gruppo Zangado) al 74 % (gruppo Chicao) del loro tempo su
substrati arborei e dal 26 % (gruppo Chicao) al 35 % (gruppo Zangado) su quelli non-arborei. Il
gruppo Chicao trascorre più tempo su substrati arborei del gruppo Zangado. Nonostante ciò, per
entrambi i gruppi, il 45% del tempo trascorso sugli alberi avviene a circa 3 m di altezza, il 40 % dai
3 m ai 6 m di altezza e il restante 15 % ad altezze maggiori di 6 m.
L’uso del substrato è influenzato dalla struttura della vegetazione e dalla disponibilità del cibo a
differenti altezze e non differisce tra stagioni. Ad una analisi più dettagliata del tipo di substrato
usato (roccia, terreno, caverne, strato arboreo basso/medio/alto) il gruppo Chicao nella stagione
secca trascorre più tempo su substrati rocciosi che nella stagione piovosa, mentre il gruppo Zangado
nella stagione piovosa trascorre più tempo sugli alberi ad altezze intermedie che nella stagione
secca. E’ possibile che nella stagione secca i cebi tendano a passare più tempo sopra i morros di
arenaria, dove ricercano invertebrati, piccoli semi e si cibano delle foglie di piante erbacee, che
nella chapada (la zona pianeggiante, dove quasi tutti gli alberi hanno perso le foglie e non hanno
frutti succulenti e semi). L’uso maggiore del substrato arboreo ad altezze medie per il gruppo
Zangado nella stagione piovosa, potrebbe essere collegato al maggior numero di frutti durante
questa stagione.
I cebi sono primati arboricoli che usano gli strati medi e bassi nella foresta (Fleagle et al., 1981;
Terborgh, 1983), anche se talvolta mostrano un notevole uso del suolo (Siemers, 2000; Moura,
2004b; Sabbatini, 2006) (Tabella 6.2)
Specie Habitat Tempo trascorso a terra (%) Fonte
C. libidinosus savana/foresta secca 26 Questo studio (gruppo Chicao)
C. libidinosus savana/foresta secca 34,5 Questo studio (gruppo Zangado)
C. libidinosus foresta secca 25,5 Moura (2004)
C. olivaceus prateria umida alternata a foresta a galleria
13,4 Robinson (1986)
C. nigritus foresta umida 1,9 Brandon Wheeler (comm. pers.)
C. apella foresta semidecidua 54,6* Siemers (2000)
C. libidinosus foresta semidecidua ~ 35 Sabbatini (2006)
Tabella 6.2 Percentuale di tempo trascorso a terra in alcune popolazioni di C. libidinosus e in altre specie del genere Cebus osservate in differenti habitat. * % di tempi trascorso da 0 a 1 m. Table 6.2 Percent of time spent on the ground level collected in some populations of C. libidinosus and in other species among Cebus living in different habitat. * % of time spent from 0 to 1 m.

72
Per i cebi studiati in zone disturbate dalla presenza dell’uomo l’uso maggiore del suolo e degli
strati bassi è imputabile alla ricerca di alimenti di origine antropica (resti e rifiuti di cibo) e alla loro
maggiore disponibilità a terra o ad altezze relativamente basse (Siemers, 2000; Sabbatini, 2006).
Purtroppo la maggior parte degli studi sull’uso dello spazio di cebi in foreste umide o semidecidue
tropicali, non riporta il tempo trascorso a terra dai cebi (Fleagle et al., 1981, Terborgh, 1983), anche
se viene sottolineata la tendenza di alcune specie (C. apella, C. olivaceous) a vivere negli strati
medio-bassi della foresta. Cionostante, l’uso del suolo sembrerebbe maggiore in quelle popolazioni
che abitano zone semi-aride rispetto a quelle che vivono nelle foreste tropicali, anche se nuovi dati
serviranno a confermare queste prime osservazioni. Ad esempio nel Parco Nazionale di Iguazù in
Argentina, in un ambiente di foresta tropicale secondaria, i cebi trascorrono a terra solamente l’1,9
% del loro tempo (Brandon Wheeler, comunicazione personale) e più del 60 % sui substrati arborei
ad altezze maggiori di 6 m.
I risultati del nostro studio sono simili a quelli riportati per i cebi di Serra da Capivara da Moura
(2004), dove i cebi trascorrono il 22,5 % del loro tempo a terra. Serra da Capivara si trova in un
ambiente di foresta secca, dove sono presenti rilievi rocciosi simili a quelli di Boa Vista, pertanto il
grado maggiore di terrestrialità potrebbe essere legato al tempo trascorso dai cebi sui morros e
quindi dipendere dalla geomorfologia dell’ambiente in cui la popolazione vive. Nelle aree visitate
dai cebi, i morros costituiscono un ambiente molto importante, dove gli animali trovano riparo e
trascorrono molto tempo alla ricerca di cibo. In particolare, tra gli anfratti rocciosi, i cebi riescono a
catturare piccoli vertebrati, soprattutto rettili (Verderane et al., 2007). A ridosso di questi rilievi la
vegetazione è costituita da alberi di bassa statura e da arbusti, ma può essere del tutto assente in
altre zone. Pertanto il contatto con il terreno o il substrato roccioso è molto familiare e regolarmente
i cebi scalano le falesie rocciose per cercare cibo lungo i pendii o avere accesso all’altro lato della
valle. Infine ricordiamo che l’uso di strumenti avviene soprattutto su incudini rocciose e che i siti
incudine rilevati si trovano quasi sempre sui morros o comunque nelle loro vicinanze (Visalberghi
et al., 2007). Ricerche future sull’uso dell’habitat associato alla ricerca del cibo e alla dieta di questi
gruppi, saranno in grado di chiarire l’importanza che rivestono i morros di arenaria per i cebi di Boa
Vista.
6.2.4 Differenze tra classi di età e sesso nell’uso del substrato Non ci sono differenze significative nell’uso del suolo tra classi di età e sesso. In media i due
gruppi di cebi esaminati trascorrono a terra il 30 % del loro tempo e il 70 % sugli alberi. Le
differenze sessuali nella posizione spaziale degli individui all’interno del gruppo sono spiegate in
termini di dimorfismo sessuale (i maschi sono più grandi e pesanti e per questo si alimentano sugli

73
alberi ad altezze minori) e/o di rischio di predazione, maggiore per gli animali che si trovano a terra
o sugli strati più alti degli alberi (Chapman, 1986).
Il fatto che a Boa Vista i cebi passino molto tempo al suolo e che le classi di età/sesso non usino
in maniera differente gli strati arborei e il suolo, suggerisce che il rischio di predazione sia alquanto
basso e/o che i cebi si sentano a proprio agio al suolo come sugli alberi.
Occasionalmente, durante la raccolta dati, ci è capitato di vedere tracce di coguaro (Puma
concolor) come anche di volpi (Lycalopex vetulus) nelle aree visitate dai gruppi e lungo il transetto,
tuttavia non abbiamo mai assistito alla predazione di un cebo. Né eventi di questo tipo sono stati
osservati dai 4 altri studenti che per anni hanno osservato i nostri due gruppi. E’ da notare che oltre
a felidi e canidi, i cebi sono prede potenziali sia di rettili, sia di rapaci. Quando i cebi avvertono un
pericolo, emettono una vocalizzazione d’allarme tipica (allarm-call) se avvistano i rapaci sopra la
copertura degli alberi, o si mostrano minacciosi (mobbing) se scoprono la presenza di rettili, come
osservato durante un incontro con un Boa constrictor.
Considerato il tipo di ambiente e la facilità con cui è possibile avvicinare i cebi, sarebbe molto
interessante esaminare in dettaglio il modo con cui gli animali si dispongono sia rispetto al gruppo
sia rispetto alle classi di età/sesso, per valutare come è stato fatto in altri studi, se la posizione
spaziale influenza le strategie di foraggiamento e il rischio di predazione (Robinson, 1981,
Fragaszy, 1990; Rose, 1994).

74
6.3 Uso di strumenti
6.3.1 Confronto tra gruppi I cebi utilizzano strumenti durante tutti i mesi dell’anno e lo fanno più frequentemente per aprire
noci che per aprire altri frutti. Questa differenza potrebbe essere imputabile al fatto che mentre le
noci di palma sono impossibili da aprire senza strumenti, data l’elevata resistenza alla frattura del
loro guscio legnoso (Visalberghi et al., 2008), almeno alcuni degli altri frutti possono essere aperti
sbattendoli con energia su superfici dure (per frutti analoghi rotti con quest’ultima tecnica vedi
Izawa & Mizuno, 1977; Gunst et al., 2008).
Le nostre osservazioni dimostrano che i cebi dedicano alla rottura di noci e di altri frutti con gli
strumenti dal 1,6 % (gruppo Zangado) al 3,1 % (gruppo Chicao) del tempo. Ciò corrisponde al 3,4
% (gruppo Zangado) e al 6,8 % (gruppo Chicao) del tempo trascorso in attività di ricerca e consumo
di cibo. Le frequenze con cui gli individui dei due gruppi usano strumenti non sono statisticamente
differenti e il tasso di uso di strumenti non sembra influenzato dal ricevere o meno un supplemento
di cibo. Pertanto sebbene i cebi del gruppo Zangado si spostino maggiormente di quelli del gruppo
Chicao presumibilmente alla ricerca di cibo, non sembrano usare strumenti perché hanno meno
risorse alimentari a disposizione.
Secondo van Schaik et al. (1999) l’uso abituale di strumenti che si osserva negli scimpanzè e
negli oranghi è il frutto di caratteristiche biologiche quali: (1) un tipo di foraggiamento estrattivo
unito ad un certo livello di abilità manipolativa; (2) capacità cognitive che prevedono una certa
forma di apprendimento; (3) organizzazioni sociali che permettono agli individui di stare
spazialmente vicini ai “modelli” in grado di eseguire tali comportamenti. Infatti le informazioni
sono più facilmente acquisite quando gli individui si trovano vicini gli uni agli altri durante il
manifestarsi del comportamento stesso (Coussi-Korbel & Fragaszy, 1995).
Nonostante il genere Cebus sia filogeneticamente separato dalle antropomorfe da circa 40
milioni di anni (Jones at al., 1992), esistono numerose convergenze fra antropomorfe, soprattutto
scimpanzé, e cebi (Visalberghi & McGrew, 1997); alcune di queste riguardano anche le
caratteristiche biologiche che van Schaik et al. (1999) ritengono importanti per lo sviluppo di uso
abituale di strumenti: anche i cebi presentano comportamenti di tipo estrattivo, sono molto abili nel
manipolare oggetti e nel combinarli gli uni con gli altri, e in molti compiti cognitivi hanno
prestazioni simili a quelle degli scimpanzé.
Inoltre i cebi presentano elevati livelli di tolleranza interindividuale. Durante le attività di
foraggiamento e di rottura delle noci gli adulti tollerano la vicinanza degli individui più giovani. A
Boa Vista più della metà degli episodi di uso di strumenti avviene in presenza di un osservatore,

75
attratto all’incudine dalla speranza di poter ottenere frammenti sparsi dell’endosperma della noce
aperta (Silva, 2008). La possibilità di stare vicino al “modello” che sa usare strumenti e di ottenere
un rinforzo positivo come quello del cibo, facilita l’opportunità di processi di apprendimento
socialmente mediati (Resende, 2004).
6.3.2 Variazione stagionale Il fatto che i cebi usino strumenti durante tutto l’anno e che la gran parte degli individui, eccetto
quelli più giovani, usi strumenti, permette di affermare che si tratta di un comportamento abituale
nella popolazione di Boa Vista.
Il tasso di uso di strumenti varia durante i mesi, ma non risulta maggiore nella stagione secca
rispetto a quella piovosa. Tuttavia, i frutti per i quali vengono usati strumenti sono diversi nelle due
stagioni. Entrambi i gruppi usano strumenti per aprire noci più nella stagione secca (quando le noci
catulè sono maggiormente disponibili) che nella stagione piovosa, mentre usano strumenti per
aprire altri frutti più nella stagione piovosa che in quella secca. Tra questi frutti il caju, che fruttifica
agli inizi della stagione piovosa, è stato quello maggiormente sfruttato dai cebi. La buccia di questo
frutto contiene sostanze tossiche potenzialmente non commestibili e molto pericolose se vengono a
contatto con le mucose. Pertanto la rottura del guscio per mezzo di percussori consente ai cebi di
accedere al seme evitando di entrare in contatto con tali sostanze.
Il fatto che l’attività di uso di strumenti avvenga simultaneamente nel 23 % degli episodi,
mostra che una volta trovata la fonte di cibo (come ad esempio un albero di caju o una palma con
un casco di noci) più individui cercano di sfruttarla, analogamente a quanto avviene quando i cebi
incontrano un albero con frutti (Janson, 1986; Izar, 2004). Quest’uso simultaneo di strumenti può
però avvenire solo quando esistono più percussori e un sito incudine spazioso o più siti incudini. A
Boa Vista solo il 12 % dei siti incudine possiede più di un percussore (Visalberghi et al., 2007),
pertanto spesso la risorsa incudine/percussore viene monopolizzata dagli individui dominanti, in
particolare dal maschio alfa mentre gli individui subordinati rimangono nelle vicinanze ad aspettare
il proprio turno. Queste attese si sono verificate soprattutto quando il gruppo Chicao incontrava
nella chapada un casco di noci a bassa resistenza di frattura, come i tucum; in queste occasioni, in
cui era presente un'unica incudine, maschio dominante, femmine e giovani si sono succeduti
all’incudine. Ciò mostra che non solo la disponibilità ma anche l’abbondanza di incudini e i
percussori è un fattore chiave per il manifestarsi dell’attività di uso di strumenti.

76
6.3.3 Confronti tra classi di età e sesso
Fatta eccezione per i piccoli di età inferiore all’anno, quasi tutti gli individui hanno usato
strumenti (92 % su titti gli individui di entrambi i gruppi). La variabilità individuale nell’uso di
strumenti è molto alta, anche se i maschi adulti usano strumenti quasi 3 volte più delle femmine
adulte e oltre 4 volte più dei giovani. In generale gli adulti usano strumenti per aprire noci più dei
giovani, e i maschi adulti li usano più delle femmine adulte. Le differenze sessuali trovate sono del
tutto simili a quelle descritte per i cebi che usano percussori in condizioni di semi-libertà (Mannu,
2002; Resende, 2004) e per i cebi di Serra da Capivara che usano percussori per aprire frutti e
scavare (digging) nel terreno alla ricerca di tuberi e usano bastoncini per sondare (probing) le cavità
di alberi (Moura, 2004; Mannu & Ottoni, 2009).
Questi risultati sono in netto contrasto con quelli trovati per gli scimpanzé, nei quali le femmine
usano strumenti più dei maschi sia nell’attività di pesca delle termiti con bastoncini (Uehara, 1986;
McGrew, 1992) sia quando aprono le noci di palma con percussori (Boesch & Boesch-Achermann,
2000). Negli scimpanzé le differenze sessuali nell’uso di strumenti vengono attribuite al maggior
fabbisogno alimentare delle femmine sia durante la gravidanza sia durante il periodo di allattamento
dei piccoli (McGrew, 1992) e alla minore socialità di queste rispetto ai maschi; questi ultimi infatti
preferiscono trascorrere il loro tempo con altri maschi adulti piuttosto che rompere noci da soli
(Boesch & Boesch, 1984).
Le differenze sessuali messe in evidenza dal nostro studio non sono compatibili con l’ipotesi
che le femmine necessitino di un maggior apporto calorico dato che usano strumenti con frequenze
decisamente inferiori a quelle dei maschi nonostante durante il periodo di studio molte di queste
femmine fossero incinte o avessero piccoli da allattare (vedi Figura 1.2). Un’ipotesi che necessita
ulteriori verifiche è quella che le femmine in condizioni naturali, così come avviene in laboratorio
(Visalberghi, 1988) abbiano una minore tendenza dei maschi ad esibire comportamenti in cui
oggetti vengono battuti su una superficie. Poiché usare percussori produce rumore udibile a
distanza, è anche possibile che le femmine mostrino questo comportamento con minore frequenza
dei maschi per evitare di attirare l’attenzione di predatori (Fragaszy, 1990). In una popolazione di C.
apella nel Suriname, sebbene i maschi e le femmine siano ugualmente capaci di sbattere frutti dal
guscio duro contro superfici dure, le femmine esibiscono questa attività con minore frequenza dei
maschi (Boinski et al., 2000). Pertanto è possibile che una maggiore tendenza ad evitare rischi
dovuta alla loro maggiore vulnerabilità ai predatori, possa determinare la marcata differenza fra
sessi riscontrata sia da Boinski et al. (2000), sia nel nostro studio.
L’ipotesi della vulnerabilità alla predazione prevede altresì che le femmine trascorrono più
tempo sui substrati arborei piuttosto che a terra, ma ciò non si verifica per i cebi di Boa Vista.

77
Anche a Serra da Capivara le femmine e i maschi trascorrano lo stesso tempo a terra e l’uso di
strumenti è più frequente nei maschi che nelle femmine (Moura, 2004). Per questo autore tale
differenza è da ricondurre alla monopolizzazione delle risorse da parte dei maschi e ai costi
energetici dell’attività di uso di strumenti troppo alti per le femmine. A questo proposito va
ricordato che lo sforzo energetico che fa un maschio per aprire una noce è sicuramente inferiore a
quello che fa una femmina visto che il peso corporeo dei maschi è maggiore. Solo studi ad hoc
condotti in laboratorio per misurare il consumo di ossigeno in condizioni di sforzo nei due sessi
potranno dare indicazioni attendibili a tale proposito.
6.3.4 Efficenza e uso dei materiali Quando usano strumenti, gli adulti 4 volte su 5 aprono la noce mangiandone il contenuto,
mentre 1 volta su 5 non riescono ad aprirla, nonostante l’uso degli strumenti sia adeguato. In parte
le cause del mancato successo di apertura sono imputabili al materiale del percussore (percussori
friabili che si rompono quando vengono usati per aprire le noci) e in parte a fattori legati alla
dominanza (gli individui subordinati ricevono minacce da parte dei dominanti che tendono a
monopolizzare l’incudine). Un episodio di uso di strumenti dura circa 2 minuti per aprire le noci a
bassa resistenza di frattura e circa 3 minuti per aprire le noci ad alta resistenza di frattura. Maschi e
femmine adulte hanno un successo di apertura simile, anche se i maschi danno meno colpi alle noci
per aprirle. Ciò è dovuto probabilmente alla maggiore forza dei maschi rispetto alle femmine.
Nei giovani, nonostante il nostro campione si basi su pochi individui, l’efficenza sembra essere
minore degli adulti (i giovani aprono l’alimento 3 volte su 5), aumentano gli episodi in cui le noci
non si aprono (quasi 2 volte su 5) e c’è una piccola percentuale di episodi (4 %) in cui i cebi
combinano gli strumenti in maniera inadeguata.
Per i cebi, aprire le noci di palma è un’attività molto costosa dal punto di vista energetico in
quanto le noci sono molto difficili da aprire a causa del loro guscio legnoso. I risultati di questo
studio ci dicono che il peso medio del percussore usato per aprire le noci è di circa 1 kg, un peso
che conferma quello stimato precedentemente con il metodo indiretto del censimento delle incudini
(Visalberghi et al., 2007). Esiste una grande variabilità individuale nel peso dei percussori usati: il
percussore più pesante usato dai maschi è stato 3 kg, mentre il percussore più pesante usato dalle
femmine è stato 2,2 kg. Tale differenza si può ricondurre al dimorfismo sessuale: i maschi sono più
grandi e robusti delle femmine e pesano circa 3 kg, mentre le femmine sono più piccole e pesano
circa 2,3 kg (EthoCebus dati non pubblicati). Da analisi di laboratorio è stato stimato che la forza
necessaria per aprire le noci va da 5,5 a 11,5 kN (Visalberghi et al., 2008), e che la durezza della
noce piassava, è 10 volte quella di una noce comune Junglans regia (Schrauf, 2006). Pertanto per

78
aprire queste noci i cebi usano percussori che pesano molto in proporzione al loro peso (dal 30 al 50
%) e inoltre l’apertura della noce richiede non solo forza ma anche abilità tecniche (Liu et al.,
2009).
In termini di materiale del percussore, sia i maschi sia le femmine usano percussori meno
friabili rispetto a quelli friabili che rischiano di rompersi quando vengono battuti contro una
superfice dura. E’ interessante notare che le femmine quando aprono le noci a maggiore resistenza
di frattura usano percussori che pesano in media il 30 % in più dei percussori che usano per aprire
noci a bassa resistenza di frattura, mentre i maschi usano percussori di peso simile per aprire
entrambe le noci. Probabilmente i maschi non sono selettivi, in termini di peso, in quanto possono
contare sulla loro dimensione e forza, mentre le femmine scelgono un percussore più pesante per
aprire le noci più dure.
Che i cebi siano in grado di selezionare i percussori quando li trasportano all’incudine e/o
quando li usano, è stato recentemente confermato da Visalberghi e collaboratori (2009a,b). A Boa
Vista la densità di percussori idonei (di materiale non friabile) è bassa (17,4 per ettaro) e nel 3,4 %
degli episodi osservati, i cebi trasportano i percussori ai siti incudine. Quando aprono le noci, i cebi
usano percussori non friabili e pesanti (930 g il peso medio) e li trasportano per circa 3 m, mentre
quando aprono altri frutti, usano percussori più leggeri (30 g il peso medio) che trasportano per
circa 5,5 m (Visalberghi et al., 2009a).
Infine Visalberghi e collaboratori (2009b) hanno dimostrato tramite esperimenti sul campo che i
cebi operano una selezione dei percussori in maniera funzionale per peso e per materiale quando
devono aprire una noce. I cebi scelgono i sassi resistenti e non quelli friabili, quelli grandi e pesanti
e non quelli più piccoli e leggeri che si frantumerebbero nell'urto con la noce. A dimostrare che la
selezione viene fatta in base alle caratteristiche funzionali (il peso) per aprire la noce, i cebi hanno
scelto il percussore giusto, quello più pesante, anche quando sono stati esposti a prove
controintuitive in cui gli è stata presentata la scelta tra un sasso grande ma leggero e un sasso
piccolo ma pesante (Visalberghi et al. 2009b).

79
7. CONCLUSIONI
Negli ultimi anni, confrontando le osservazione relative a molti anni di studio provenienti da
differenti siti, è stata messa in luce la grande variabilità comportamentale tra popolazioni della
stessa specie. Quando è stato possibile escludere che la presenza di certi comportamenti in alcune
popolazioni e non in altre potesse essere dovuta a fattori genetici e/o ecologici è stato possibile
affermare che si trattava di tradizioni comportamentali (Fragaszy & Perry, 2003). In questo modo è
emerso che scimpanzé e oranghi, e altri mammiferi come i delfini (Mann & Sargeant, 2003), hanno
comportamenti tradizionali – molti dei quali si riferiscono all’uso di strumenti – non riconducibili a
differenze genetiche o ambientali, ma frutto di innovazioni e apprendimento sociale (McGrew et al.,
1997; Whiten et al., 2001; van Schaik et al., 2003a).
Il comportamento di uso di percussori documentato a Boa Vista attraverso questo lavoro di tesi,
è abituale (vedi la definizione di Whiten et al., 1999), ovverosia è mostrato dalla maggior parte
degli individui e durante la maggior parte dell’anno e presenta tutte le caratteristiche per essere
definito tradizionale (Fragaszy & Perry, 2003). Infatti, nonostante strumenti e frutti dal guscio duro
siano presenti in molti siti (anche se non in tutti) dove i cebi sono stati studiati, questo
comportamento è presente in un numero molto limitato di popolazioni di Cebus (Ottoni & Izar,
2008), e se presente viene mantenuto grazie ad influenze sociali (Resende et al., 2008; Silva, 2008).
Il primo scopo di questo studio era di valutare la variabilità nell’uso di strumenti tra due gruppi
di cebi, sulla base di ipotesi ecologiche. L’ipotesi della necessità (Fox et al., 2004) sostiene che è la
scarsità delle risorse primarie a favorire l’esplorazione di cibi nuovi e soluzioni tecniche differenti
come l’uso di strumenti. Secondo questa ipotesi (1) i cebi avrebbero dovuto usare più
frequentemente gli strumenti durante la stagione in cui il cibo era scarso e (2) il gruppo che non
riceveva cibo supplementare avrebbe dovuto usare gli strumenti più frequentemente del gruppo che
lo riceveva. Purtroppo l’assenza di una marcata differenza fra disponibilità delle risorse nella
stagione secca e nella stagione piovosa non ci permette di valutare la prima di queste predizioni.
Tuttavia l’ambiente semi-arido di Boa Vista non sembra essere così povero di risorse in confronto
ad altri ambienti in cui i cebi vivono, come ad esempio la Foresta Atlantica (Izar, 2004; Nakai,
2007), da averli potuti spingere a mettere a punto tecniche di uso di strumenti per aver accesso a
risorse fondamentali per la loro sopravvivenza. Inoltre i risultati da noi ottenuti non confermano la
seconda predizione secondo la quale le frequenze di uso di strumenti dovrebbero differire tra i due
gruppi che differiscono per necessità alimentari, in quanto solo uno di essi riceve cibo
supplementare.

80
I nostri risultati sono invece in accordo con le predizioni dell’ipotesi dell’opportunità ambientale
(Beck, 1980; Fox et al., 2004) secondo cui l’emergenza di comportamenti innovativi – tra cui l’uso
di strumenti – sono favoriti da opportune condizioni ambientali. A Boa Vista sono presenti sia sassi
percussori sia superfici idonee per schiacciare le noci (Visalberghi et al., 2007) e i nostri risultati
dimostrano che la disponibilità di risorse alimentari non sembra essere particolarmente scarsa e,
come già detto in precedenza, che le frequenze di uso di strumenti nei due gruppi sono simili.
Inoltre, secondo l’ipotesi dell’opportunità ambientale, ci aspettavamo che a Boa Vista (3) il
comportamento di uso di strumenti da parte di entrambi i gruppi riflettesse la disponibilità delle
noci e di altri frutti incapsulati. I risultati di questo studio mettono in evidenza che sebbene i cebi
usino strumenti tutto l’anno, la frequenza d’uso rispecchia l’abbondanza di frutti che necessitano di
questa tecnica per essere aperti; ciò è evidente nel caso delle noci catulè e per i frutti del caju. E’
interessante notare che per aprire noci sono necessari strumenti idonei, ma che questi non sempre
sono disponibili e/o abbondanti, pertanto la disponibilità di incudini e percussori opportuni potrebbe
essere considerata tra i fattori ecologici che limitano l’uso di strumenti in alcune popolazioni di cebi
ma non in altre. Analogamente Leca et al. (2008) ritengono che fattori ecologici abbiano impedito
ai macachi del Giappone di trasformare la manipolazione di pietre (stone-handling) in qualcosa di
finalizzato ad ottenere cibo altrimenti inaccessibile.
Infine l’elevato tempo trascorso a terra dai cebi di Boa Vista è in accordo con l’ipotesi (4)
secondo la quale le maggiori opportunità di esplorare le proprietà di noci e percussori offerte da un
elevato tasso di terrestrialità potrebbero favorire, così come avviene in condizioni di cattività,
l’emergenza del comportamento di uso di strumenti (Visalberghi, 1987; Visalberghi et al., 2005).
Fino ad oggi le popolazioni di cebi che usano abitualmente strumenti sono state osservate in habitat
simili (Moura, 2004a; Fragaszy 2004b; Ottoni & Izar, 2008) dove gli animali trascorrono
relativamente più tempo a terra anche a causa della geomorfologia dell’ambiente (bassa vegetazione
e presenza di rilievi rocciosi).
Il grado di terrestrialità riscontrato a Boa Vista e nelle altre popolazioni selvatiche che usano
strumenti, come del resto quello di gruppi mantenuti in condizioni di cattività o semi-libertà, è
molto elevato rispetto a quello riportato per popolazioni in cui l’uso di strumenti non è stato
osservato. Il fatto che l’uso di strumenti occupi solo una minima parte del tempo trascorso a terra,
rende l’ipotesi che i cebi abbiano imparato ad usare strumenti perché già svolgevano altre attività a
terra, più parsimoniosa e quindi scientificamente preferibile, di quella opposta secondo la quale i
cebi abbiano iniziato a svolgere altre attività a terra dopo aver acquisito questo comportamento.
Il secondo scopo di questo studio era valutare le differenze nell’uso di strumenti tra le classi di
età e sesso. Le differenze riscontrate sono simili a quanto osservato da Boesch & Boesch (1984)

81
negli scimpanzé in cui gli adulti usano più strumenti e hanno più successo dei giovani (Ipotesi 1).
Nei cebi però, al contrario di quanto riportato per gli scimpanzé, le femmine usano strumenti meno
frequentemente dei maschi, in accordo con quanto atteso (Ipotesi 2). Questo risultato è opposto a
quello atteso dall’ipotesi del fabbisogno energetico (Clutton-Brock, 1977) secondo cui durante
l’allattamento e la gestazione le femmine hanno maggiori richieste energetiche, pertanto dovrebbero
usare strumenti più dei maschi per ottenere cibi nutrienti e ricchi di lipidi e proteine come le noci o
gli invertebrati. Al contrario, per i cebi di Boa Vista sembrerebbe che usare percussori sia così
dispendioso a livello energetico da non risultare vantaggioso per le femmine, la cui massa corporea
e dimensioni sono inferiori a quelle dei maschi.
Infine i nostri risultati confermano che i cebi selezionano gli strumenti in maniera funzionale,
secondo il peso e il materiale del percussore, come riportato da Visalberghi et al. (2009a,b) e che
operano una scelta del percussore anche in base al tipo di alimento da aprire. In tale scelta
sembrerebbe che i maschi siano più generalisti delle femmine in termini di peso del percussore e
tale differenza potrebbe essere riconducibile al dimorfismo sessuale, per cui le femmine, meno forti
e pesanti dei maschi, ottimizzano il dispendio energetico usando percussori più pesati solo quando
devono aprire frutti più duri.
In conclusione i risultati di questo studio mettono in risalto che in condizioni non sperimentali
l’uso abituale di strumenti non è un comportamento unico del genere Homo o di alcune scimmie
antropomorfe, ma che fa parte del repertorio comportamentale anche del genere Cebus. Da un punto
di vista evolutivo le ricerche sull’uso di strumenti nei cebi sono interessanti per gli studi
comparativi dell’evoluzione della tecnologia dei primi ominidi. La paleoantropologia ha talvolta
sviluppato modelli del comportamento umano ispirati al comportamento dei primati non-umani, e
specialmente degli scimpanzé, filogeneticamente più vicini all’uomo (Standford, 1999). Ad
esempio mettendo in relazione la dieta, la distribuzione del cibo e le caratteristiche del sistema
sociale delle antropomorfe con i dati paleoclimatici e paleobiologici si è tentato di ricostruire il tipo
di habitat dei primi ominidi, e la distribuzione di piante e specie animali che potrebbero aver fatto
parte della loro dieta (Gollop & Foley, 2001). Da modelli di questo tipo risulta che la dieta delle
australopitecine comprendeva frutti carnosi, noci, semi e tuberi (Ungar, 2004). Inoltre si ritiene che
alcuni di questi alimenti venissero estratti con l’uso di pietre (Alba et al., 2003), sebbene questi
percussori siano stati poco studiati perché difficilmente distinguibili da sassi dello stesso materiale
ma non usati a tale scopo.
In questo contesto, la descrizione dettagliata dei percussori utilizzati dai cebi e dagli scimpanzé
può servire per meglio riconoscere analoghi strumenti utilizzati dai nostri antenati. Inoltre, la
possibilità di studiare il comportamento di uso di percussori da parte di scimmie altamente

82
encefalizzate, parzialmente terrestri e che abitano zone simili alle savane, che selezionano gli
strumenti idonei per estrarre cibo non altrimenti raggiungibile, e che li trasportano e li usano in
posizione bipede, possono fornire una nuova prospettiva per interpretare l’evoluzione tecnologica e
cognitiva della nostra specie (Ottoni & Izar, 2008).

83
8. RINGRAZIAMENTI Questa ricerca è stata finanziata dall’Università “La Sapienza” di Roma, dalla Scuola di Dottorato
in Biologia Animale e dell’Uomo, dall’Istituto di Scienze e Tecnologie della Cognizione del CNR
di Roma e dal Centro studi Etologici Ethoikos. Ringrazio il progetto EthoCebus per il supporto
tecnico e logistico presso la stazione di ricerca di Boa Vista, il CNPq e il FAPESP per aver fornito il
salario agli assistenti in Brasile e quindi reso possibile le mie ricerche sul campo.
Vorrei ringraziare tutte le persone che hanno contribuito a rendere possibile questo lavoro, sia in
Italia sia in Brasile.
Il mio docente guida, la Dr. Elisabetta Visalberghi, per la professionalità, l’interesse e l’entusiasmo
con cui ha seguito tutte le fasi del lavoro, nonché per la fiducia che in me ha riposto. Devo a lei la
mia crescita scientifica; grazie anche per avermi insegnato cosa vuol dire fare ricerca in un Paese
come l’Italia.
Il mio relatore interno, Prof. Carlo Utzeri, per la disponibilità sia durante le mie visite in Italia sia
nella fase finale della tesi.
Il mio contro-relatore, Dr. Gabriele Schino per le preziose revisioni della tesi.
Il Prof. Luigi Boitani per la disponibilità e l’incoraggiamento sin dalle prime fasi del dottorato.
Ringrazio inoltre il Collegio dei Docenti della Scuola di Dottorato in Biologia Animale per aver
compreso la mia prolungata assenza, per l’interesse mostrato nella mia materia di studio e per aver
contribuito alla revisione della tesi.
Per il supporto ricevuto dal primo giorno a Boa Vista, ringrazio infinitamente il team di ricercatori
EthoCebus. Innanzitutto la Prof. Dorothy Fragaszy che con pazienza mi ha accompagnata nei primi
giorni in foresta; grazie per avermi introdotta con gentilezza e competenza nel mondo dei cebi.
Per l’ospitalità, l’amicizia e gli insegnamenti di come si realizza una ricerca sul campo, tutta la mia
stima va alla Prof. Patrizia Izar; grazie per i preziosi consigli, l’aiuto e la solidarietà, sia a Boa Vista
sia durante l’analisi dei dati e la stesura della tesi.
Non sarei mai potuta rimanere 18 mesi in Brasile se non avessi ricevuto il visto da ricercatore;
ringrazio il Prof. Eduardo Ottoni per aver avuto la pazienza di seguire la pratica fin dall’inizio e per
la disponibilità che mi ha sempre dimostrato.

84
Ringrazio gli studenti Quing Liu, Adam Hinely e Luca Mantovani per la collaborazione ma
soprattutto i miei indimenticabili amici del sertão con i quali ho condiviso le levatacce alle 4 e
mezza, i chilometri nella foresta, il caldo, la polvere, il cielo stellato e le scalate di luoghi incredibili
come la cima dei morros. Michele P. Verderane per la collaborazione, l’amicizia e il supporto
morale. E. Darvin R. da Silva per l’amicizia, la serietà nel lavoro e la curiosità con cui mi ha
insegnato a guardare la natura. Jozemar e Arizomar da Silva Olivera, amici e assistenti di campo
che mi hanno guidata sulle tracce dei cebi e protetta in situazioni pericolose. Sono certa che se il
sudore è evaporato, le rumorose risate echeggiano ancora tra le valli di Boa Vista.
A Boa Vista sono stata ospite di una famiglia straordinaria, per composizione e dolcezza. Sono loro
che hanno reso possibile la mia permanenza in Brasile, e a loro va tutta la mia riconoscenza e
affetto. Mauro, Maria, Marcos, M. Junior, Marina, Mara, Martina, Marcio e Chico. Grazie per
avermi accolta nella vostra famiglia ancor prima che imparassi a comunicare nella vostra lingua.
Grazie a Mauro per gli insegnamenti che mi ha trasmesso con passione e per l’impegno nella
conservazione della sua terra. Per l’amore, la pazienza, l’incoraggiamento, ma soprattutto la sua
forza, non smetterò mai di ringraziare Maria. Grazie inoltre per i semplici e gustosi piatti con cui ci
hai accolto dopo lunghe giornate di lavoro. Per l’allegria, la vita sana, le cene, la dolcezza e la
spontaneità, ringrazio tutti i ragazzi della famiglia M, siete stati per me dei compagni e fratelli
indimenticabili.
Nelle visite a Gilbués ho avuto il piacere di incontrare Dona Penina. Grazie per la semplicità e
generosità con cui mi hai aperto quando ho bussato, per le tue rughe da 72enne e per i racconti
davanti ad un cafezinho. Grazie inoltre a Hildebrando Penteado per l’ospitalità mai negata e le
lunghe chiacchierate sull’uscio di casa.
Per la simpatia e scambio di informazioni ringrazio agli studenti e i ricercatori dell’Istituto di
Psicologia dell’Università di San Paolo, Briseida, Andrea, Camila, Erika, Tiago, Caio e la Prof.
Emma Otta. Sono molto grata anche a João e Patrizia per avermi sempre offerto una deliziosa
ospitalità.
Raone Beltrão Mendes per la sua amicizia: gli amici sono sempre amici non importa la distanza.
Ringrazio i ricercatori, gli studenti, i tecnici e i guardiani del Centro Primati dell’ISTC del CNR di
Roma con i quali ho condiviso l’ultimo anno del dottorato. Per lo scambio di informazioni, la

85
competenza scientifica, l’aiuto, i consigli durante l’analisi dei dati e le pause durante il lavoro
ringrazio Eugenia, Martina, Alessandra, Valentina, Gloria, Elsa e Tommaso. Il geologo Andrea
Evangelista con cui ho avuto il piacere di lavorare per 6 mesi; grazie per la disponibilità e
collaborazione, sei stato fondamentale per la riuscita di questo lavoro. Grazie inoltre alla Dr. Ilaria
Agostini per i preziosi consigli statistici, la solidarietà e l’amicizia anche dal Sudamerica. Paco
Bertolani per avermi spedito tanto materiale cartografico e Giulia Sirianni per le correzioni della
tesi. Ringrazio inoltre Barbara e Brandon Wheeler per la solidarietà, lo scambio di dati e la
revisione dell’inglese nel testo.
Un grazie speciale va ai miei amici italiani, a quelli che ho lasciato partendo e a quelli che ho
ritrovato tornando. Simon, per aver partecipato alle mie vicende e per continuare a farlo. Vincenzo,
apparso il primo giorno che è iniziata questa avventura e diventato in seguito un amico prezioso.
Grazie per aver condiviso le paure, le difficoltà e le soddisfazioni delle diverse fasi di questa
ricerca. Un grazie meritatissimo a Stefano e Vincenzo per le leccornie che sono riusciti a spedirmi e
per le lunghe lettere. Insieme a loro ringrazio le mie amiche più care: Vittoria, Lucia, Daria,
Valeria, Livia e Marta per essermi state sempre vicino. Non saprò mai come ringraziare Valentina e
Gamal, che sono venuti a trovarmi in Brasile: non c’è cosa più bella di condividere le proprie
esperienze con chi vuoi bene. Grazie per l’amicizia, siete dei compagni di viaggio impareggiabili.
Michele, per la curiosità e l’interesse nei miei racconti, l’amore, la comprensione e soprattutto la
pazienza dimostrata nei mesi di studio e di stesura della tesi. Grazie anche per la revisione di alcuni
capitoli.
Infine, ma non per questo ultimi, un grazie infinito va ai miei cari genitori e ai miei fratelli Paolo e
Gionata ai quali dedico questo lavoro. A migliaia di chilometri di distanza e senza alcun mezzo di
comunicazione, siete riusciti a starmi vicino con amore incondizionato. In quei lunghi mesi a sud
dell’equatore, così come nel periodo trascorso a Roma, non mi è mai mancato il vostro appoggio e
ho fatto tesoro di consigli e perplessità. Con la speranza di farvi conoscere Boa Vista. Ringrazio
inoltre il resto della famiglia e nonna Ofelia per le parole di conforto e l’incoraggiamento.
L’esperienza brasiliana mi ha dato l’opportunità di conoscere una parte della mia famiglia emigrata
mezzo secolo fa in Brasile. Ringrazio di cuore zia Lucia, zio Donato, Mariassunta, Alfonso e
Simone, per l’ospitalità e per l’affetto con cui mi avete accudito nelle settimane trascorse a San
Paolo. Grazie per l’esempio di coraggio e di enorme sacrificio di una vita lontana dalla propria terra
natale.

86
Grazie infine ai cebi di Chicao e Zangado, esempio e invidia di come si può vivere bene in un
ambiente tanto arido.
Andai nei boschi perché desideravo vivere con saggezza,
per affrontare solo i fatti essenziali della vita,
e per vedere se non fossi capace di imparare quanto essa aveva da insegnarmi.
Queste montagne spoglie e questi boschi silenziosi sono la mia bussola nella tempesta...
... sono il simbolo dell'essenzialità che ognuno di noi dovrebbe imparare a costruire
prima di tutto dentro di sé,
in qualunque luogo della terra ci si trovi.
(H.D. Thoreau, Walden, La vita nei boschi, 1854)

87
9. BIBLIOGRAFIA
Adams-Curtis, L. & Fragaszy, D. M. (1995). Influence of a skilled model on the behavior of
conspecific observers in tufted capuchin monkeys (Cebus apella). American Journal of
Primatology 37:65-71.
Aguiar, L.M. de S., Machado, B.R., Marino, J.F. (2004). A diversidade Biologia do Cerrado. In:
Aguiar, L.M.de S., Camargo, A.J.A. Cerrado - ecologia e caracterização. Brasilia: Embrapa
Informação Técnologica.
Alba, D.M., Moyà-Solà, S., Köhler, M. (2003). Morphological affinities of the Australopithecus
afarensis hand on the basis of manual proportions and relative thumb length. Journal of
Human Evolution 44:225–254.
Anderson, J. R. (1990). Use of objects as hammers to open nuts by capuchin monkeys (Cebus
apella). Folia Primatologica 54:138-145.
Alp, R. (1997). “Stepping-sticks” and “seat-sticks”: new types of tools used by wild chimpanzees
(Pan troglodytes) in Sierra Leone. American Journal of Primatology 41:45–52.
Altmann, J. (1974). Observational study of behaviour: sampling methods. Behaviour 49: 227-267.
Beck, B. (1980). Animal Tool Behavior: The Use and Manufacture of Tools by Animals. New
York: Garland Press. Edizione italiana Torino Boringhieri 1986, pag. 17.
Boesch, C., Boesch, H. (1984). Possible causes of sex differences in the use of natural hammers by
wild chimpanzees. Journal of Human Evolution 13:415–440.
Boesch, C.H. & Boesch-Achermann (2000). The Chimpanzees of the Taï Forest. Oxford University
Press, Oxford.
Boinski, S. (1988). Use of a club by wild white-faced capuchin (Cebus capucinus) to attack a
venomous snake (Bothrops asper). American Journal of Primatology 14:177-179.
Boinski, S., Quatrone, R.P. and Swartz, H. (2000). Substrate and tool use by brown capuchins in
Suriname: ecological contexts and cognitive bases. Am. Anthropol. 102: 741-761.
Breuer, T., Ndoundou-Hockemba, M., Fishlock, V. (2005). First Observation of tool use in wild
gorillas. PLoS Biology 3(11):e380.
Brown, A.D. & Zunino, G.E. (1990). Dietary variability in Cebus apella in extreme habitats:
Evidence for adaptability. Folia Primatologica 54: 187-195.
Brown, A.D., Chalukian, S.C., Malmierca, L.M., Colillas, O.J. (1986). Habitat structure and feeding
behavior of Cebus apella (Cebidae) in El Rey National Park, Argentina. In: Current

88
Perspectives in Primate Social Dynamics. D.M. Taub, F.A. King, eds. Van Nostrand
Reinhold Company, New York.
Byrne, R. (1995). The thinking ape: evolutionary origins of intelligence. Oxford: Oxford University
Press.
Canale, G.R., Guidorizzi, C.E., Kierulff, M.C.M., Gatto, C. (2009). First Record of Tool Use by
Wild Populations of the Yellow-Breasted Capuchin Monkey (Cebus xanthosternos) and
New Records for the Bearded Capuchin (Cebus libidinosus). American Journal of
Primatology 71:1–7.
Carpenter, A. (1887). Monkeys opening oysters. Nature 36:53
Chapman, C.A. (1986). Boa constrictor predation and group response in white-faced Cebus
monkeys. Biotropica 18(2):171–172.
Chevalier-Skolnikoff, S. (1990). Tool use by wild Cebus monkeys at Santa Rosa National Park,
Costa Rica. Primates 31(3):375-383.
Chevalier-Skolnikoff, S. & Liska, J. (1993). Tool use by wild and captive elephants. Animal
Behaviour 46:209–219.
Clutton-Brock, T.H. (1974). Activity patterns of red colobus (Colobus badius tephrosceles). Folia
Primatologica 21:161-187.
Clutton-Brock, T.H. (1977). Some Aspects of Intraspecific Variation in Feeding and Ranging
Behaviour in Primates. In T.H. Clutton-Brock, editor, Primate Ecology, New York:
Academic Press.
Coussi-Korbel, S., Fragaszy, D.M. (1995). On the relation between social dynamics and social
learning. Animal Behaviour 50:1441–1553.
Dunbar, R.I.M. (1992). Neocortex size as a constraint on group size in primates. Journal of Human
Evolution 20:469-493.
Eibl-Eibesfeldt, I. (1961). U¨ ber den Werkzeuggebrauch des Spechtfinken Camarhynchus pallidus
(Scalter und Salvin). Z. f. Tierpsychol. 18:343–346.
Erickson, C. (1995). Feeding Sites for Extractive Foragaing by the Aye-Aye. American Journal of
Primatology 35(3):235-240.
Fedigan, L. (1993). Sex differences and intersexual reation in adul white-faced capuchins (Cebus
capucinus). International Journal of Primatology 14: 853-877.
Fernandes, M.E.B. (1991). Tool use and predation of oysters (Crassostrea rhizophorae) by the
tufted capuchin, Cebus apella apella, in brackish water mangrove swamp. Primates 32(4):
529-531.

89
Ferreira, R.G., Resende B.D., Mannu, M., Ottoni, E.B., and Izar, P. (2002). Bird predation and
prey-transference in Brown Capuchin Monkeys (Cebus apella). Neotropical Primates 10:
84-89.
Ferreira, R.G., Lee, P.C., Izar, P. (2008). Food Competition in a Semi-Free-Ranging Cebus apella
Group. Folia Primatologica 79:463-475
Fleagle, J.G., Mittermeier, R.A. and Skopec, A.L., 1981, Differential habitat use by Cebus apella
and Saimiri sciureus in Central Surinam. Primates 22: 361-367.
Ford, S.M. and Davies, L. (1992). Systematic and body size: implication for feeding adaptations in
New World monkeys. American Journal of Physical Anthropology 88: 415-468.
Fox, E.A., Sitompul, A.F., van Schaik, C.P. (1999). Intelligent tool use in wild Sumatran
orangutans. In: Parker, S.T., Mitchell, R.W., Miles, H.L. editors. The mentalities of gorillas
and orangutans. Cambridge, UK: Cambridge University Press. p 99–116.
Fox, E.A. & bin’Muhammad, I. (2002). Brief Communication: New tool use by wild Sumatran
orangutans (Pongo pigmaeus abelii). American Journal of Physical Anthropology 119:186-
188.
Fox, E.A. , van Schaik, C.P., Sitompul, A., Wright, D.N. (2004). Intra and interpopulational
differences in orangutan (Pongo pygmaeus) activity and diet: implications for the invention
of tool use. American Journal of Physical Anthropology 125:162–174.
Fragaszy, D. M. (1986). Time budgets and foraging behavior in wedge-capped capuchins (Cebus
olivaceus): age and sex differences. pp. 159-174. In: Taub, D. and King, F. (Eds.) Current
perspectives in primate social dynamics. New York, van Nostrand Reinhold.
Fragaszy, D.M., (1990). Sex and age differences in the organization of behaviour in wedge-capped
capuchin. Behavioral Ecology 1, 81-94
Fragaszy, D.M. & Visalberghi, E. (1989). Social influences on the acquisition of tool-using
behaviors in tufted capuchin monkeys (Cebus apella). Journal of Comparative Psychology
103:159-170.
Fragaszy, D.M. & Adams-Curtis, L.E. (1991). Generative aspects of manipulation in tufted
capuchin monkeys (Cebus apella). Journal of Comparative Psychology. 105(4):387-397.
Fragaszy, D.M., Perry, S. (2003). Towards a biology of traditions. In: Fragaszy, D.M., Perry, S.,
editors. The biology of tradition: models and evidence. Cambridge: Cambridge University
Press. p 1–32.
Fragaszy, D.M., Visalberghi, E., Fedigan, L.M. (2004a). The Complete Capuchin: The Biology of
the Genus Cebus. Cambridge University Press: Cambridge, UK.

90
Fragaszy, D.M., Izar, P., Visalberghi, E., Ottoni, E. B., Oliveira, M. G. (2004b). Wild capuchin
monkeys (Cebus libidinosus) use anvils and stone pounding tools. American Journal of
Primatology 64 (4): 359-366.
Galetti, M. & Pedroni, F. (1994). Seasonal diet of capuchin monkeys (Cebus apella) in a semi-
deciduous forest in south-east Brazil. Journal of Tropical Ecology 10(1): 27-39.
Galleti, M. (1990). Predation on the squirrel, Sciurus aestuanus by capuchin monkeys, and Cebus
apella. Mammalia 54:152-154
Galdikas, B.M.F. (1982a). An unusual instance of tool-use among wild orang-utans in Tanjung
Puting Reserve, Indonesian Borneo. Primates 23:138–139.
Galdikas, B.M.F. (1982b). Orangutan tool-use at Tanjung Puting Reserve, Central Indonesian
Borneo (Kalimantan Tengah). Journal of Human Evolution 10:19–33.
Gollop P, Foley RA. 2001. Evolutionary geography, habitat availability and species distributions
among early African hominids. In: Harcourt C, editor. Monkey, apes and humans: an
evolutionary perspective. London: Westview Press. p 40–58.
Goodall, J. (1963). Feeding behaviour of wild chimpanzees: a preliminary report. Symp Zool. Soc.
Lond. 10:39-47.
Goodall, J. (1986). The chimpanzee of Gombe: pattern of behaviour. Cambridge, Harvard
University Press.
Graziani, G. Abituazione alla presenza umana di un gruppo di gorilla occidentali (Gorilla gorilla
gorilla), nel parco di Dzanga-Shanga, Repubblica Centrafricana. Tesi di laurea in Scienze
Biologiche, Università di Roma “La Sapienza”, a.a. 1999/2000.
Groves, C.P. (2001). Primate taxonomy. Washington, DC: Smithsonian Institution Press.
Gunther, M.M. & Boesch, C. (1993). Energetic cost of nut-cracking behaviour in wild chimpanzees.
In: Preuschoft, H. and Chivers, D.J. (Eds.). Pp.109-129. Hands of Primates. Springer-
Verlag, Wien.
Gunst, N., Boinski, S., and Fragaszy, D.M. (2008). Acquisition of foraging competence in wild
brown capuchins (Cebus apella), with special reference to conspecifics’ foraging artefacts as
an indirect social influence. Behavior 145, 195–229.
Hamilton, W.J. III, Buskirk, R.E., Buskirk, W.H. (1978). Environmental determinants of object
manipulation by chacma baboons (Papio ursinus) in two Southern African environments.
Journal of Human Evolution 7:205–216.
Hall, K.R.L., Schaller, G.B. (1964). Tool-using behavior of the California sea otter. Journal of
Mammalogy 45:287–298.

91
Hart, B.L., Hart, L.A., McCoy, M., Sarath, C.R. (2001). Cognitive behaviour in Asian elephants:
use and modification of branches for fly switching. Animal Behaviour 62: 839–847.
Henschel, J.R. (1995). Tool use by spiders: stone selection and placement by corolla spiders
Ariadna (Segestriidae) of the Namib desert. Ethology, 101:187-199.
Hinley, A.J. (2006). Gis-based habitat modelling related to bearded capuchin monkey tool use.
Master of Science, UGA, Athens (GA).
Huffman, M.A. (1984). Stone-play of Macaca fuscata in Arashiyama B troop: transmission of a
non-adaptive behavior. Journal of Human Evolution 13:725–735.
Huffman, M.A., Quiatt, D. (1986). Stone handling by Japanese macaques (Macaca fuscata):
implications for tool use of stone. Primates 27:427–437.
Hunt, G.R. & Gray, R. D. (2004). Direct observations of pandanus-tool manufacture and use by a
New Caledonian crow (Corvus moneduloides). Animal Cognition. 7:114-120.
Hunt, G.R. (2000). Tool Use by the New Caledonian Crow Corvus moneduloides to obtain
Cerambycidae from Dead Wood. EMU 100:109-114.
Hunt, G.R. (1996). Manufacture and use of hook-tools by New Caledonian crows. Nature 379:
249-251.
Ingmanson, E.J. (1996). Tool-using behavior in wild Pan paniscus: social and ecological
considerations. In: Russon, A.E., Bard, K.A., Parker, S.T., editors. Reaching into thought:
the minds of the great apes. Cambridge: Cambridge University Press. p 190–210.
Isaac, G. (1978). The food sharing behaviour of protohuman hominids. Scientific American 238:90-
108.
Izar, P. (2004). Female social relationships of Cebus apella nigritus in a southeastern Atlantic
forest: an analysis through ecological models of primate social evolution. Behaviour 141:
71-99.
Izawa, K. (1980). Social behavior of the wild black-capped capuchin (Cebus apella). Primates 21:
443-467.
Izawa, K. & Mizuno, A. (1977). Palm-fruit cracking behavior of wild black-capped capuchin
(Cebus apella). Primates 18:773–792.
Janson, C.H. (1986). The mating system as a determinant of social evolition in capuchin monkeys
(Cebus). In J.G. Else & P.C. Lee (eds.): Primate Ecology and Conservation. Cambridge,
England: Cambridge University Press, pp. 169-179.
Jones, S., Martin, R., and Pilbeam, D. (1992). The Cambridge Encyclopedia of Human Evolution,
Cambridge University Press, Cambridge.

92
Kortland, A. & Kooij, M. (1963). Protohominid behaviour in primates (preliminary
communication). Symp. Zool. Soc. Lond. 10:61-88.
Langguth, A. & Alonso, C. (1997). Capuchin monkeys in the Caatinga: Tool use and food habits
during drought. Neotropical Primates 5(3):77-78.
Leakey, R. (1980) The Making of Mankind. London, Book Club Associates.
Leca, J.B., Gunst, N. &Huffman, M.A. (2008). Of stones and monkeys: Testing ecological
constraints on SH, a behavioral tradition in Japanese macaques. American Journal of
Physical Anthropology 135:233–244.
Leighty, K.A., Byrne, G., Fragaszy, D.M., Visalberghi, E. (2004). Twinning in Tufted Capuchins
(Cebus apella): Rate, Survivorship, and Weight Gain. Folia Primatologica 75(1):14.
Liu, Q. (2007). Kinematics and energetic of nut-cracking in wild capuchin monkeys (Cebus
libidinosus) in Piauí, Brazil. Master’ thesis, UGA, Athens (GA).
Liu, Q., Simpson, K., Izar, P., Ottoni, E., Visalberghi, E., Fragaszy, E. (2009). Kinematics and
energetics of nut-cracking in wild capuchin monkeys (Cebus libidinosus) in Piauí, Brazil.
American Journal of Physical Anthropology 138(2):210-220.
Lonsdorf, E. V., Pusey, A. E. & Eberly, L. (2004). Sex differences in learning in chimpanzees.
Nature 428, 715–-716.
Manaivijitnond, S., Lekprayoon, C., Tandavanittj, N., Panha S., Cheewatham, C. and Hamada., Y.
(2007). Stone-Tool Usage by Thai Long-Tailed Macaques (Macaca fascicularis). American
Journal of Primatology 69:227–233.
Mann, J. & Sargeant, B.L. (2003). Like mother, liker calf: the ontogeny of foraging tradition in wild
Indian ocean bottlenose dolphins (Tursiops sp.). In: Fragaszy D.M., Perry S., editors. The
biology of tradition: Models and evidence. Cambridge: Cambridge University Press. Pag
237-266.
Mann, J., Sargeant, B.L., Watson-Capps, J.J., Gibson, Q.A., Heithaus, M.R., Connor, R.C.,
Patterson, E. (2008). Why do dolphins carry sponges? PLoS ONE 3(12): e3868.
Mannu, M. (2002). O uso espontâneo de ferramentas por macacos-prego (Cebus apella) em
condições de semi-liberdade: descrição e demografia. Dissertação de mestrado, Instituto de
Psicologia da Universidade de São Paulo.
Mannu, M. & Ottoni, E.B. (2009). The Enhanced Tool-Kit of Two Groups of Wild Bearded
Capuchin Monkeys in the Caatinga: Tool Making, Associative Use, and Secondary Tools.
Am. J. Primatol. 71:242–251.
Marais, E. (1969). The soul of the ape. New York: Atheneum.

93
Martin, P. & Bateson, P. (1993). Measuring behaviour - an introductory guide. Cambridge
University Press, Cambridge, 2nd ed.
Matsuzawa, T. (1994). Field experiments on use of stone tools in the wild, in: Wrangham, R.W.,
McGrew, W.C., de Waal, F.B.M., Heltne, P.G., Marquardt, L.A. (Eds.), Chimpanzee
cultures. Harvard University Press, Cambridge, pp. 351–370.
Matsuzawa, T., Biro, D., Humle, T., Inoue-Nakamura, N., Tonooka, R. and Yamakoshi, G. (2001).
Emergence of culture in wild chimpanzees: education by master-apprenticeship. In Primate
Origins of Human Cognition and Behaviour, ed. Matsuzawa. Tokyo: Springer-Verlag.
McGrew, W.C. (1974). Tool use by wild chimpanzees in feeding upon driver ant. Journal of
Human Evolution 3:501-508.
McGrew, W.C. (1992). Chimpanzee material culture. Cambridge: Cambridge University Press.
McGrew, W.C. & Marchant, L.F. (1997). Using the tools at hand: manual laterality and elementary
technology in Cebus spp. and Pan spp. International Journal of Primatology 18:787-810.
McGrew, W.C., Ham, R.M., White, L.J.T., Tutin, C.E.G., Fernandez, M. (1997). Why don’t
chimpanzees in Gabon crack nuts? International Journal of Primatology 18:353-374.
McNab, B.K., (1983). Ecological and behavioral consequences of adaptation to various food
resources. In J.F. Eisenberg and D. Kleiman, ed. Advances in the study of mammalian
behaviour. American Society of Mammologist Special publication 7:664-697.
Moura, A.C.A (2004). The capuchin monkey and the caatinga dry forest: a hard life in a harsh
habitat. PhD dissertation, University of Cambridge, Cambridge (UK).
Moura, A.C.A. & Lee, P. (2004). Capuchin stone tool use in caatinga dry forest. Science 306
(5703):1909-1910.
Murphy, P. G. and Lugo, A.E. (1986). Ecology of tropical dry forests. Ann. Rev. Ecol. Syst. 17: 67-
88.
Nahallage, C.A.D., Huffman, M.A. (2007). Age-specific functions of stone handling, a solitary-
object play behavior, in Japanese macaques (Macaca fuscata). American Journal
Primatology 69:1-15.
Nakai, E. S. (2007). Fissão-fusão em Cebus nigritus: flexibilidade social como estratégia de
ocupação em ambientes limitantes. Dissertação (Mestrado) - Instituto de Psicologia,
Universidade de São Paulo, São Paulo.
Ottoni, E.B. & Mannu, M. (2001). Semifree-ranging tufted capuchins (Cebus apella) spontaneously
use tools to crack open nuts. International Journal Primatology 22:347-358.

94
Ottoni, E.B., de Resende, B.D., Izar, P. (2005).Watching the best nutcracking: what capuchin
monkeys (Cebus apella) know about other’s tool-using skills. Animal Cognition 24:215-
219.
Ottoni, E.B, & Izar, P (2008). Capuchin monkey tool use: overview and implication. Evolutionary
Anthropology 17:171-178
Parker, S.T., Gibson, K.R. (1977). Object manipulation, tool use and sensorimotor intelligence as
feeding adaptations in cebus monkeys and great apes. Journal of Human Evolution 6:623–
641.
Peres, C.A. (1994). Primate responses to phenological changes in an Amazonian terra firme forest.
Biotropica 26(1):98-112.
Pickering, T. (2007). The optimisation of palm-nut (Attaleya sp.) cracking in wild bearded capuchin
monkeys (Cebus libidinosus). Bachelor's Honors Thesis UGA, Athens (GA).
Post, D.G., Hausfater, G., McCuskey, S.A. (1980). Feeding behavior of yellow baboons (Papio
cynocephalus): relationship to age, gender and dominance rank. Folia Primatologica
34:170–195.
Pruetz, J.D. & Bertolani, P. (2007). Savanna Chimpanzees, Pan troglodytes verus, hunt with tools.
Current Biology 17:412–417.
Quinn, A., D., Wilson (2004). Daubentonia madagascariensis. Mammalian Species 710: 1-6.
Resende, B.D., Greco, V.L.G., Ottoni, E.B., and Izar, P. (2002). Some Observations on the
Predation of Small Mammals by Tufted Capuchin Monkeys (Cebus apella). Neotropical
Primates 11:103-104.
Resende, B.D. (2004). Ontogenia de comportamentos manipulativos em um grupo de macacos-
prego (Cebus apella) em situação de semi-liberdade. Tese de Doutorado apresentado ao
Instituto de Psicologia da Universidade de São Paulo.
Resende, B.D., Ottoni, E.B, Fragaszy D.M. (2008). Ontogeny of manipulative behavior and nut-
cracking in young tufted capuchin monkeys Cebus apella): a perception-action perspective.
Developmental science 11(6):828-40.
Rocha, V. J., Reis, N. R. & Sekiama, M. L. (1998). Uso de ferramentas por Cebus apella
(Linnaeus)(Primates, Cebidae) para obtenção de larvas de coleóptera que parasitam
sementes de Syagrus romanzoffianum (Cham.) Glassm. (Arecaceae). Revista brasileira de
Zoologia 15(4):945-950.
Robinson, J. G. (1986). Seasonal variation in use of time and space by the wedge-capped capuchin
monkey, Cebus olivaceus: implications for foraging theory. Smithson. Contrib. Zool. 431:
1:60.

95
Robinson, J. G. (1981). Spatial Structure in Foraging Groups of Wedge-capped Capuchin Monkeys,
Cebus nigrivittatus. Animal Behaviour 29:1036-1056.
Russon, A. E. & Galdikas, B. M. F. (1993). Imitation in free-ranging rehabilitant orangutans
(Pongo pygmaeus). Journal of Comparative Psychology 107 (2):147-161.
Sabbatini, G., Stammati, M., Tavares, M.C.H., Visalberghi, E. (2008). Behavioral flexibility of a
group of bearded capuchin monkeys (Cebus libidinosus) in the National Park of Brasília
(Brazil): consequences of cohabitation with visitors. Brazilian Journal of Biology
68(4):685-693.
Sabbatini, G. (2006). Plasticità del comportamento alimentare di Cebus libidinosus nel Parco
Nazionale di Brasília (Brasile). Tesi di dottorato (XVIII ciclo) in Biologia Animale e
dell’Uomo, Sapienza Università di Roma.
Sampaio, D.T., Ferrari, S.F. (2005). Predation of an infant titi monkey (Callicebus moloch) by a
tufted capuchin (Cebus apella). Folia Primatologica 76:113-115.
Savage-Rumbaugh, S. & Lewin, R. (1994). Kanzi, the ape at the brink of human mind. New York:
John Wiley & Sons.
Schick, K.D., Toth, N., Garufi, G., Savage-Rumbaugh, S., Rumbaugh, D. and Sevcik, R. (1999).
Continuing Investigations into the Stone Tool-making and Tool-using Capabilities of a
Bonobo (Pan paniscus). Journal of Archaeological Science 26(7):821-832.
Schrauf, C. (2006). Tool selection in a nut-cracking task by capuchin monkeys (Cebus apella).
Unpublished master`s thesis, University of Vienna, Austria.
Siemers, B.M. (2000). Seasonal variation in food resources and forest strata use by brown capuchin
monkeys (Cebus apella) in a disturbed forest fragment. Folia Primatologica 71:181-193
Silva, E.D.R. (2008). Escolha de alvos coespecificos na observação do uso de ferramenta por
macaco-prego (Cebus libidinosus) selvagens. Tese de Mestrado em Psicologia
Experimental, Universidade de São Paulo.
Silva Junior, J. S. (2001). Especiação em macacos-prego e caiararas, gênero Cebus Erxleben, 1777
(Primates, Cebidae). Rio de Janeiro, 377 p. Tese (Doutorado em Genética), Curso de Pós-
graduação em Genética, UFRJ.
Smolker, R.A., Richards, A., Connor, R., Mann, J., Berggren, P. (1997). Sponge-carrying by Indian
Ocean bottlenose dolphins: possible tool-use by a delphinid. Ethology 103:454-465.
Sokal, R.R., Rohlf, F.J. (1995). Biometry. New York: W. H. Freeman & Co.
Spironelo, W.R. (1991). Importância dos frutos de palmeiras (Palmae) na dieta de um grupo de
Cebus apella (Cebidae, Primates) na Amazônia Central. In: A Primatologia no Brasil, vol.
3.

96
Spagnoletti, N. (2005). Ritmi di attività in un gruppo di gorilla di pianura occidentali (Gorilla
gorilla gorilla) nel Parco Nazionale di Dzanga-Ndoki, Repubblica Centrafricana. Tesi di
laurea, Sapienza Università di Roma.
Stanford, C.B. (1999). The hunting apes: meat eating and the origins of human behavior. Princeton:
Princeton University Press.
Struhsaker, T. T. & Leland, L. (1977). Palm-nut smashing by Cebus a. apella in Colômbia.
Biotropica 9:124-126.
Sugiyama, Y. (1993). Local variation of tools and tool use among wild chimpanzee population. In
The Use of Tools by Human and Non-Human Primates, ed. A. Berthelet and J. Chavillon,
pp. 175-187. Oxford: Claredon Press.
Taira, J.T., Verderane, M.P., Ottoni, E.B., Izar, P. (2002). Exploração das palmeiras Euterpe edulis
e Archontophoenix cunninghamiana por duas populaçoes de macacos-prego (Cebus apella).
Anais de Etologia 20:326.
Terborgh, J. (1983). Five New World monkeys. Princeton University Press, Princeton.
Tebbich, S., Taborsky, M., Fessl, B., & Dvorak, M. (2002). The ecology of tool-use in the
woodpecker finch (Cactospiza pallida). Ecology Letters 5: 656-664.
Tomasello, M. & Call, J. (1997). Primate Cognition. Oxford University Press, Oxford.
Tutin, C.E.G., Fernandez, M. (1991). Responses of wild chimpanzees and gorillas to the arrival of
primatologists: behaviour observed during habituation. In: Box, H.O. ed., Primate responses
to environmental change, p. 187-197. London: Chapman and Hall.
Uehara, S. (1986). Sex and group differences in feeding on animals by wild chimpanzees in the
Mahale Mountains National Park, Tanzania. Primates 27(1):1-113.
Ungar, P.S. (2004). Dental topography and diets of Australopithecus afarensis and early Homo.
Journal of Human Evolution 46:605–622.
van Lawick-Goodall, J. (1970). Tool-using in primates and other vertebrates. In: Advances in the
study of behaviour 3, 195-249
van Lawick-Goodall, J. & van Lawick-Goodall, H. (1966). Use of tools by the Egyptian vulture,
Neophron percnopterus. Nature 212:1468–1469.
van Schaik, C.P., Fox, E.A. & Sitompul, A.F. (1996). Manufacture and of tools in wild Sumatran
orangutans. Naturwissenschaften 83:186-188.
van Schaik, C.P., Deaner, R., Merrill, M. (1999). The conditions for tool use in primates:
implications for the evolution of material culture. Journal of Human Evolution 36, 719.

97
van Schaik, C.P., Ancrenaz, M., Borgen, G., Galdikas, B., Knott, C.D., Singleton, I., Suzuki, A.,
Utami, S.S. & Merrill, M. (2003a). Orangutan cultures and the evolution of material culture.
Science 299:102-105.
van Schaik, C.P., Fox, E.A., Fechtman, L.T. (2003b). Individual variation in the rate of use of tree-
hole tools among wild orangutans: implications for hominin evolution. Journal of Human
Evolution 44:11–23.
Verderane, M. P., Ramos Da Silva, E. D., Spagnoletti, N., Fragaszy, D. M., Visalberghi, E., Ottoni,
E. B., Izar, P. (2007). Predação de Vertebrados por macacos-prego selvagens (Cebus
libidinosus) em área de écotono Cerrado/Caatinga no Piauí. In: XII Congresso Brasileiro de
Primatologia, Belo Horizonte p. 130.
Visalberghi, E. (1987). Acquisition of nut-cracking behaviour by 2 capuchin monkeys (Cebus
apella). Folia Primatologica 49:168-181.
Visalberghi, E. (1988). Responsivenesse to objects in two social group of tufted capuchin monkeys
(Cebus apella). American Journal Of Primatology 15:168-181.
Visalberghi, E. (1993). Tool use in a South American monkey species: An overview of the
characteristics and limits of tool use in Cebus apella. In A. Berthelet & J. Chavaillon (Eds.),
The use of tools by human and non-human primates (pp. 118-131). Oxford: Claredon Press.
Visalberghi, E. & Trinca, L. (1989). Tool use in capuchin monkeys: distinguishing between
performing and understanding. Primates 30:511-521.
Visalberghi, E. (1990). Tool use in Cebus. Folia Primatologica 54:146–154.
Visalberghi, E., Fragaszy, D. M. & Savage-Rumbaugh, S. (1995). Performance in a tool using task
by common chimpanzee (Pan troglodytes), bonobos (Pan paniscus), an orangutan (Pongo
pygmaeus) and capuchin monkeys (Cebus apella). Journal of Comparative Psychology 109
(1):52-60.
Visalberghi, E. & McGrew, W.C. (1997). Cebus meets Pan. International Journal of Primatology
18:677–681.
Visalberghi, E., Fragaszy, D.M., Izar, P., Ottoni, E.B. (2005). Terrestriality and tool use. Science
308:951–952.
Visalberghi, E. & Fragaszy, D. (2006). What is challenging about tool use? The capuchin's
perspective. In: Comparative Cognition: Experimental Explorations Of Animal Intelligence.
Wasserman, E.A., Zentall, T.R., Editors. New York: Oxford Univ. Press. 529-552.
Visalberghi, E., Fragaszy, D., Ottoni, E., Izar, P., de Oliveira, M.G., Andrade, F.R.D. (2007).
Characteristics of hammer stones and anvils used by wild bearded capuchin monkeys

98
(Cebus libidinosus) to crack open palm nuts. American Journal of Physical Anthropology
132:426-444.
Visalberghi, E., Sabbatini, G., Spagnoletti, N., Andrade, F.R.D., Ottoni, E., Izar, P., Fragaszy, D.
(2008). Physical properties of palm fruits processed with tools by wild bearded capuchins
(Cebus libidinosus). American Journal Of Primatology 70(9): 884-891.
Visalberghi, E., Spagnoletti, N., da Silva, E. D. R., de Andrade, F. R., Ottoni, E., Izar, P., Fragaszy,
D. (2009a). Transport of hammer tools and nuts by wild capuchin monkeys. Primates
50(2):95-104.
Visalberghi, E., Addessi, E., Spagnoletti, N., Truppa, V., Ottoni, E., Izar, P. & Fragaszy, D.
(2009b). Selection of effective stone tools by wild capuchin monkeys. Current Biology
19:213-217.
Waga, I.C., Dacier, A.K., Pinga, P.S., Tavares, M.C.H. (2006). Spontaneous tool use by wild
capuchin monkeys (Cebus libidinosus) in the cerrado. Folia Primatol 77:337–344.
Washburn, S.L. (1978). The evolution of man. Scientific American 239:146-54.
Westergaard, G.C. & Fragaszy, D.M. (1987). The manufacture and use of tools by capuchin
monkeys (Cebus apella). Journal of Comparative Psychology 101:159–168.
Westergaard, G. C. & Suomi, S. J. (1994). A simple stone-tool technology in monkeys. Journal of
Human Evolution 27:399-404.
Whiten, A., Goodall, J., McGrew, W. C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C .E. G.,
Wrangham, R. W. & Boesch, C. (1999). Cultures in chimpanzees. Nature 399:682-685.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C.E.G.,
Wrangham, R.W., Boesch, C. (2001). Charting cultural variation in chimpanzees. Behaviour
138:1481–1516.
Zang, S.Y. (1995). Activity and ranging patterns in relation to fruit utilization by Brown Capuchins
(Cebus apella) in French Guiana. International Journal of Primatology 16(3):489-507.

99
APPENDICE 1 Figura A.1. Mappa di distribuzione dell’uso spontaneo di strumenti nel genere Cebus. Sono incluse le popolazioni
selvatiche che fanno uso sistematico di strumenti, i gruppi in condizioni di semi-libertà, le popolazioni dove sono stati
registrati episodi occasionali e indiretti e quelli in cui non è mai stato osservato tale comportamento (riadattato da
Ottoni & Izar, 2008).
Figure A.1 Distribution map of reports on spontaneous tool use by wild and semifree tufted capuchin monkeys (Ottoni
& Izar, 2008).
Legenda figura 5.14: ���� Nessun caso documentato su studi di lunga durata (fatta eccezione per evidenze di episodi di manipolazione
complessa di oggetti in contesti alimentari) A: Caratinga Biological Station, MG, Brasile [1]; B: Carlos Botelho State Park, SP, Brasile [2]; C: Iguazù National Park, Argentina [3]; D: El Rey National Park, Argentina [4]; E: Cocha Cashu Biological Station, Peru [5]; F: Urucu River, AM, Brasile [6]; G: La Macarena, Colombia [7,8]; H: BDFF Project, AM, Brazil [9]; I: Raleighvallen, Suriname [10]; J: Station des Nouragues, French Guiana [11].
� Evidenze indirette in gruppi selvatici (presenza di siti di rottura) o osservazioni sporadiche di uso di strumenti per aprire frutti incapsulati. K: Canelatiua, MA, Brazil [12]; L: Alto Paraiso, GO, Brasile [13]; M: Serra da Mesa, GO, Brasile [14]; N: Desterro do Malta, Paraiba, Brasile [15]; O: Martins, Rio Grande do Norte, Brasile [16]; P: Belo Monte, Alagoas, Brasile [17]; Q: Varzelandia/ Montes Claros, Minas Gerais, Brasile [17]; R: Itiuba/Ibotirama/Contendas do Sincorà, Bahia, Brasile [17]; S: Terra Ronca National Park, GO, Brasile [18]; T: Minaçu, GO, Brasile [18]; U: Mara Rosa, GO, Brasile [18]; V: Peixe, TO, Brasile [18].
� Uso abitudinario di pietre per aprire noci in popolazioni selvatiche o in semi-libertà. Quadrato piccolo: gruppi in semi-libertà in parchi urbani. a: Tieté Ecological Park, Sao Paulo, SP, Brasile [19]; b: Jaraguà State Park, Sao

100
Paulo, SP, Brasile [20]; c: Arthur Thomas Mun. Park, Londrina, PR, Brasile [21]; d: Agua Mineral Park, Brasilia, DF, Brasile [22]. Quadrato grande: popolazioni selvatiche. W: Boa Vista, PI, Brasile [23] e X: Brasilia National Park, Brasile [24]. Gruppi selvatici che usano strumenti per aprire frutti, raccogliere il cibo o si servono di bastoncini in maniera esplorativa (a volte le due cose sono combinate). Y: Serra da Capivara, Pl, Brasile [25] [26]. A-D: Cebus nigritus; E-K: C. apella; L-P, S-Y: C. libidinosus; Q-R: C. xanthosternos; a-d: popolazioni ibride o incerte (vivono in parchi urbani).
Referenze bibliografiche per la figura 5.14: [1] Rımoli J. 2001. Ecologia de macacos-prego (Cebus apella nigritus, Goldfuss, 1809) na Estacao Biologica de Caratinga (MG): implicaçoes para a conservaçao de fragmentos de Mata Atlantica. Tesi di dottorato non pubblicata. Federal University of Parà, Brasile. [2] Izar P. 2004. Female social relationships of Cebus apella nigritus in Southeastern Atlantic Forest: an analysis through ecological models of primate social evolution. Behaviour 141:71–99. [3] Di Bitetti MS. 2001. Home-range use by the tufted capuchin monkey (Cebus apella nigritus) in a subtropical rainforest of Argentina. Journal Zool Lond 253:33–45. [4] Brown AD, Zunino GE. 1990. Dietary variability in Cebus apella in extreme habitats: evidence for adaptability. Folia Primatologica 54:187–195. [5] Terborgh J. 1983. Five New World monkeys. Princeton: Princeton University Press. [6] Peres CA. 1994. Primate responses to phenological changes in an Amazonian terra firme forest. Biotropica 26:98–112. [7] Izawa K, Mizuno A. 1977. Palm-fruit cracking behavior of wild black-capped capuchin (Cebus apella). Primates 18:773–792. [8] Struhsaker TT, Leland L. 1977. Palm-nut smashing by Cebus a. apella in Colombia. Biotropica 9:124–126. [9] Spironello WR. 2001. The brown capuchin monkey (Cebus apella). Ecology and home range requirements in Central Amazonia. In: Bierregaard RO, Gascon C Jr, Lovejoy TE, Mesquita R, editors. Lessons from Amazonia: the ecology and conservation of a fragmented forest. New Haven: Yale University Press. p 271–283. [10] Boinski S. Comunicazione personale agli autori. [11] Zhang S. 1995. Activity and ranging patterns in relation to fruit utilization by brown capuchin monkeys (Cebus apella). International Journal of Primatology 16:489–507. [12] Fernandes MEB. 1991. Tool use and predation of oysters (Crassostrea rhizophorae) by the tufted capuchin, Cebus apella apella, in brackish water mangrove swamp. Primates 32:529–531. [13] Galvao O. Comunicazione personale agli autori [14] Souza CA, Mendes FDC, Silva JN Jr. 2002. Utilizaçao de ferramentas por Cebus apella libidinosus de dispersao livre. X Congresso Brasileiro de Primatologia. [15] Langguth A. & Alonso C. 1997. Capuchin monkeys in the Caatinga: tool use and food habits during drought. Neotropical Primates 5:77–78. [16] Ferreira RG. Comunicazione personale agli autori. [17] Canale, G.R., Guidorizzi, C.E., Kierulff, M.C.M., Gatto, C. (2009). First Record of Tool Use by Wild Populations of the Yellow-Breasted Capuchin Monkey (Cebus xanthosternos) and New Records for the Bearded Capuchin (Cebus libidinosus). American Journal of Primatology 71:1–7. [18] Mendes FD, Ottoni EB, Izar P, Cardoso R, Villar D, Marchezan R. In preparazione. [19] Ottoni EB, Mannu M. 2001. Semifree-ranging tufted capuchin monkeys (Cebus apella) spontaneously use tools to crack open nuts. International Journal of Primatology 22:347–358. [20] Silva EDR, Resende BD, Ottoni EB. 2005. Tecnicas de manipulaçao de alimentos utilizadas pelo grupo livre de macacos-prego (Cebus apella) do Parque Estadual do Jaraguà: um estudo preliminar. Res Cong Bras Primatol 11:164. [21] Rocha VJ, Reis NR, Sekiama ML. 1998. Uso de ferramentas por Cebus apella (Linnaeus) (Primates, Cebidae) para a obtençao de larvas de Coleoptera que parasitam de sementes de Syagrus romanzoffianum (Cam.) Glassm. (Arecaceae). Rev Bras Zool 15:945–950. [22] Visalberghi E. Comunicazione personale agli autori. [23] Fragaszy D, Izar P, Visalberghi E, Ottoni EB, Oliveira M. 2004. Wild capuchin monkeys use anvils and stone pounding tools. American Journal of Primatology 64:359–366. [24] Waga IC, Dacier AK, Pinga PS, Tavares MCH. 2006. Spontaneous tool use by wild capuchin monkeys (Cebus libidinosus) in the cerrado. Folia Primatologica 77:337–344. [25] Moura ACA, Lee P. 2004. Capuchin stone tool use in Caatinga dry forest. Science 306:1909. [26] Mannu M, Ottoni EB. (2009)The enhanced toolkit of two groups of wild bearded capuchin monkeys in the Caatinga: tool making, associative use, and secondary tools. American Journal of Primatology 71:242–251.

101
APPENDICE 2
Figura A.2 Home range del gruppo Chicao e del gruppo Zangado da giugno 2006 a maggio 2007. La stima
dell’area è stata ottenuta con il metodo del Minimo Poligono Convesso (MCP). Home range Chicao = 6,9
km2 (N punti = 1439, N giorni = 92); Home range Zangado = 4,9 km2 (N punti = 1919, N giorni =
105). Le aree dei due gruppi si sovrappongono per 3 km2 (37,5%) (Dati EthoCebus non pubblicati).
Figure A.2 Chicao and Zangado home range from June 2006 to May 2007. The estimated area has been
obtained by Minimum Convex Poligon (MCP). Home range Chicao group = 6,9 km2 (N recorded point
=1439, N follow day = 92; Home range Zangado group = 4,9 km2 (N recorded point = 1919, N
follow day = 105) The areas of the two groups are overlapped of 3 km2 (37,5%)(EthoCebus
unpublished data).
Top Related