







Languages
Pages
Legal


University of Groningen
The impact of nutrition on neuroinflammation in vitro and in vivoKurtys, Ewelina Anna
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2017
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Kurtys, E. A. (2017). The impact of nutrition on neuroinflammation in vitro and in vivo [Groningen]:University of Groningen
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 16-09-2018

Chapter 3
Anti-inflammatory effects
of rice bran components
Ewelina Kurtys1, Ulrich L. M. Eisel2, Robert J. J. Hageman3,
Martin J. Verkuyl3, Laus M. Broersen3,
Rudi A. J. O. Dierckx1, Erik F. J. de Vries1
1 Department of Nuclear Medicine and Molecular Imaging, University of Groningen,
University Medical Center Groningen,
PO Box 30.001, 9700 RB Groningen, The Netherlands. 2 Department of Molecular Neurobiology, Center for Life Sciences,
University of Groningen, Groningen, The Netherlands
3 Nutricia Research, Uppsalalaan 12, 3584 CT Utrecht, The Netherlands

Chapter 3
52
Abstract
Neuroinflammation has been implicated in the pathology of various psychiatric and
neurodegenerative disorders. Accumulating evidence suggests that food
components can modulate inflammatory processes and therefore it is hypothesized
that such nutrients exhibit therapeutic efficacy against these brain diseases.
Rice bran is often discarded as a waste product, although it contains a wide range
of potentially useful substances. Several rice bran components have been described
to possess anti-inflammatory properties. This review summarizes the evidence
supporting a modulatory effect of rice bran components on neuroinflammation. In
vitro studies with these rice bran components on immune cells and in vivo studies
on nutritional intervention in animal models of central and peripheral
inflammation are discussed in the context of the potential use of rice bran
components for prevention and treatment of brain diseases in which
neuroinflammation is involved.

Review on rice bran components
53
3
1. Introduction
Neuroinflammation is a natural response of the central nervous system (CNS) to
alterations in the environment and disturbances in homeostasis, for example due to
invasion of pathogens or neuronal damage. Astrocytes and microglia are two
important cell types in the CNS that are responsible for the maintenance of
homeostasis. Astrocytes are necessary for the trophic support of neurons and
microglia are the main immune cells in the brain and therefore are also called brain
macrophages [1]. During neuroinflammation, activated astrocytes and microglia
produce a wide range of inflammatory mediators, such as cytokines, chemokines,
reactive oxygen and nitrogen species (ROS and NOS) [1,2]. These
proinflammatory mediators help to destroy invading pathogens, to clear cell debris
and to repair damaged cells. However, when neuroinflammation becomes
persistent and excessive, it may also have a detrimental effect on brain functions
[1]. Evidence suggests that pro-inflammatory mediators, especially when
chronically produced, are involved in the development of brain diseases. Chronic
neuroinflammation has been observed in many psychiatric and neurodegenerative
diseases, including schizophrenia [3], depression [4], Parkinson’s disease and
Alzheimer’s disease [5–7]. It is currently is even believed that peripheral
inflammation can trigger brain diseases, such as Parkinson’s diseases, Alzheimer’s
disease [7–9], depression [10–12] and cognitive decline [13,14], via induction of
neuroinflammation.
Neuroinflammation appears to be an important factor in the etiology of brain
diseases and in disease progression. Therefore, neuroinflammation may present a
promising target for new treatment approaches against these diseases [15]. Since
epidemiological studies demonstrated that our diet can have an impact on
incidence of brain diseases in for which pathology neuroinflammation is involved
[16,17], there is a lot of interest in the investigation of specific nutrients that may be
responsible for this effect and therefore could be good candidates for future
nutrition-based therapies.

Chapter 3
54
2. Rice bran components
Rice is the major component of the diet for a large part of the human population
and provides more than 20% of the calories consumed by humans worldwide. Rice
bran, the outer layer of a whole brown rice kernel, consists of aleurone and
pericarp. It is often considered as a by-product of rice processing and is discarded
as waste or used as animal food. However, rice bran contains high amounts of
useful nutrients, including proteins, fats, dietary fibers, minerals, anti-oxidants and
phytochemicals [18,19]. Recent studies have shown that rice bran components like
phytosteryl pherulates and isoprenoids can exert anti-inflammatory effects, either
acting directly on the immune cells [20] or affecting inflammation indirectly via
modulation of gut microbiota [21]. These results may open the road towards the
design of an (adjuvant) therapeutic intervention with specific rice bran components
for brain disorders, for which effective treatment is currently not available. This
review therefore surveys the anti-inflammatory properties of several rice bran
components in the context of potential intervention for brain diseases associated
with neuroinflammation.
2.1 Phytosteryl pherulates and ferulic acid
2.1.1 Phytosteryl pherulates
Mode of action
Gamma oryzanol (-OZ) is a secondary plant metabolite from bran layers of grains
and is a mixture of at least 10 ferulic acid esters of phytosterols. -OZ is rich in anti-
oxidants and lipid lowering compounds [22–24]. Its ROS-scavenging properties
have been extensively investigated in vitro on lipids [24], tissue homogenates [25]
and LPS-stimulated macrophages [26]. The anti-inflammatory properties of -OZ
have been demonstrated in LPS-stimulated vascular endothelial cells (model for atherosclerosis) where it inhibited the expression of adhesion molecules through
inhibition of NF-B [27]. Interestingly, a recent in vivo study demonstrated that the
permeation of the -OZ components through the gut is very low, suggesting that
the effects of -OZ are probably indirect [28].

Review on rice bran components
55
3
Peripheral inflammation
In vivo, the anti-inflammatory properties of -OZ have been demonstrated in DSS-
induced colitis. Oral administration of -OZ inhibited inflammation in mice with
DSS-induced colitis and thus prevented disease progression [29]. The anti-
inflammatory and lipid-lowering properties of -OZ have also been demonstrated
in metabolic syndrome models, such as high fat and high fructose diet-induced
metabolic syndrome in rats [30], high fat diet in rats [22] and in Zucker rats [23].
Despite promising data from animal studies suggesting anti-inflammatory
properties of -OZ in the gut, potential effects on neuroinflammation, brain
function and behavior have not been shown yet.
2.1.2 Ferulic acid
Mode of action
One of the most extensively studied component of -OZ is ferulic acid (FA), a
phenolic derivative of cinnamic acid. FA is present in rice bran and several other
plants, such as whole grain, citrus fruit, banana, beet root, cabbage, spinach and
broccoli [31]. FA can also be isolated from arabinoxylans, fibers present in the
plant cell wall, which are also emerging as promising anti-inflammatory
compounds [32]. Studies on animal models of diseases in which inflammation is
involved have demonstrated neuroprotective and anti-oxidative effects of FA. In
the middle cerebral artery occlusion model of stroke in rats, administration of FA
decreased the infarct volume. This neuroprotective effect of FA was mediated by
activation of Akt signaling pathway, which plays a critical role in cell survival
signaling and restoring the anti-oxidant proteins peroxiredoxin-2 and thioredoxin
[33,34]. Further studies on the same model have demonstrated that FA
administration leads to inhibition of the activation of microglia, macrophages,
oxidative stress and apoptosis markers [35]. Likewise, FA showed neuroprotective
properties in aged rats by inhibiting cytokine release and upregulating the
MEK/ERK½ survival pathway in the hippocampus [36].
Peripheral inflammation
Several peripheral inflammation models were used to investigate the ability of FA
to modulate disease progression. In a model of high-fat diet-induced oxidative stress
in mice, FA alone or in combination with atorvastatin ameliorated oxidative stress

Chapter 3
56
and thus protected the liver by reducing lipid peroxidation, normalizing the hepatic
lipid profile and elevating hepatic antioxidant enzymes [37]. In a model of
streptozotocine-induced diabetes in rats, FA also decreased oxidative stress, the
release of pro-inflammatory cytokines and apoptosis of pancreatic beta cells and
consequently the normalization of blood glucose levels [31].
Neuroinflammation and behavior
Evidence also suggests a role of FA in the CNS, which could be due to the
modulation of neuroinflammation. In a model of gamma radiation-induced
neuroinflammation in mice, FA exhibited a neuroprotective and anti-
neuroinflammatory effect by increasing antioxidant enzymes (SOD, CAT) and by
partly reducing the levels of pro-inflammatory markers, such as NF-B, COX-2,
iNOS, TNFα and IL-6 [38]. An anti-depressant effect of FA was shown in several
preclinical investigations. Screening assays for new antidepressants with the tail
suspension test and the forced swim test in mice showed that acute administration
of FA has an anti-depressant effect through modulation of the serotoninergic
system. Moreover, sub-effective doses of FA demonstrated synergistic anti-
depressant effects with serotonergic drugs used in the clinic for the treatment of
depression (fluoxetine, paroxetine and sertraline) [39]. Other studies demonstrated
the involvement of anti-oxidative modulation in the anti-depressant effects of FA in
stress-induced depressive-like behavior. Amelioration of behavioral abnormalities
was accompanied with a decrease in stress-induced anti-oxidative enzymes (SOD,
CAT, GSH) [40]. Similar anti-depressant effects were shown in a reserpine-
induced pain and a depressive-like behavior model in mice, in which FA partly
reversed behavioral abnormalities, increased the nociceptive threshold and
decreased the inflammatory and apoptosis markers, NF-B and caspase 3 in the
prefrontal cortex [41]. Behavior improvement and anti-inflammatory effects due to
the FA administration was also demonstrated in animal models of
neurodegenerative diseases. For example, in a transgenic model of Alzheimer’s
disease, FA attenuated behavioral abnormalities and diseases-related pathology,
including the proinflammatory markers [42]. In chemically-induced Parkinson’s
disease models, FA attenuated behavioral abnormalities, suppressed
neuroinflammation and had a neuroprotective effect [43][43,44].

Review on rice bran components
57
3
Summarizing, these data from preclinical studies indicate that FA can have
beneficial effects on neuroinflammation, inflammation-related behavior and disease
progression. Therefore, this rice bran component might be considered as potential
candidates for further clinical investigation. Unfortunately, clinical studies with FA
are lacking so far.
2.2 Isoprenoids
Isoprenoids (terpenoids) are secondary metabolites of the mevalonate pathway, in
which cholesterol and isoprenoid lipids are produced. Isoprenoids are involved in
maintaining endotoxin tolerance. Some autoimmune diseases are caused by
mutations of enzymes in the mevalonate pathway. Isoprenoids, such as
geranylgeraniol (GGOH), farnesol (FOH) and geraniol (GOH), are produced
endogenously, but are also consumed via our diet, as they are present in several
fruits, vegetables and grains, including rice [45].
2.2.1 Geranylgeraniol
Mode of action
GGOH is believed to play an important role in regulation of inflammatory
responses. Recent studies have shown that sufficient production of GGOH is
necessary to prevent chronic inflammation due to repetitive exposure to a pro-
inflammatory stimulus (endotoxin tolerance). An in vitro study demonstrated that
chemical blocking of the mevalonate pathway caused mitochondrial dysfunction
and apoptosis in LPS-stimulated murine monocytes. The effect of inhibition of the
mevalonate pathway was reversed by treatment with GGOH [46]. In macrophage-
derived RAW cells, chemical suppression of mevalonate pathway caused an
excessive proinflammatory response, which could be reversed by the exogenous
administration of GGOH. Similar beneficial effects of GGOH were demonstrated
on human monocytes from patients with an impaired mevalonate pathway [47].
Also in peritoneal macrophages repeatedly stimulated in vitro with LPS, insufficient
GGOH production, caused by inhibition of 3-hydroxy-3-methyl-glutaryl-CoA
reductase, led to the excessive proinflammatory response [48].
Peripheral inflammation
The findings from in vitro models have been confirmed in animals. In a mouse
model of mevalonate pathway dysfunction, administration of GGOH reduced the

Chapter 3
58
levels of inflammatory markers, serum amyloid-A and peritoneal exudate cells [49].
In rats intraperitoneally injected with LPS, dietary supplementation with GGOH
reduced the plasma levels of inflammatory cytokines, attenuated the activation of
the pro-inflammatory transcription factor NF-B in the liver and prevented liver
damage [50].
Studies on the effect of GGOH on neuroinflammation and brain function are
currently lacking.
2.2.2 Farnesol
Peripheral inflammation
FOH was described to possess anti-inflammatory and anti-oxidative properties and
to exert beneficial effects on lung injury [51], metabolic disorders [52], colitis [53]
and allergic reactions [54]. In chemically-induced colitis in rats, FOH decreased
colonic mucosal damage by decreasing ROS, inflammation and apoptosis,
stimulating anti-oxidative enzymes and reducing colitis-related mucosal edema
[53]. However, the anti-inflammatory effects of FOH are not always beneficial for
human health. FOH produced by Candida albicans is involved in quorum sensing.
Thus, FOH can suppress the Th1 response in favor of an anti-inflammatory Th2
response and thus promote fungal growth and exacerbation of infection [55].
Neuroinflammation
Recent studies point towards beneficial effects of FOH on the CNS, as FOH was
suggested to be a modulator of neuroinflammation and pain. In a mouse model of
acrylamide-induced neurotoxicity, FOH supplementation caused behavioral
improvement (measured with gait performance, neuromuscular function and fine
motor coordination) and attenuation of inflammation as measured with astrocytes
and microglia staining in cortex, hippocampus and striatum [56]. Neroli oil
containing FOH was demonstrated to have central and peripheral nociceptive
effects in both rats and mice [57].
Although the available data suggest promising properties of isoprenoids on
inflammation and brain function, more studies in inflammation-related brain
diseases models are needed to determine the potential usefulness of isoprenoids as
modulator of neuroinflammation in clinical investigations.

Review on rice bran components
59
3
3. Nutrients combinations: the most effective way?
As follows from the above, several rice bran components are able to attenuate
inflammatory processes, not only in the gastrointestinal tract, but possibly also in
the brain. Dietary interventions with specific food components with potent anti-
inflammatory properties, such as rice bran components, may therefore be explored
as new therapeutic strategies against brain diseases. Yet, it can be expected that
each individual component is less effective than pharmaceutical drugs that are
specially designed to inhibit inflammation. However, food components can interact
with different pathways involved in the inflammatory process and therefore a
combination of several nutrients acting on convergent pathways may increase the
efficacy of dietary interventions. The first step to test possible synergistic or additive
anti-inflammatory effects of specific food components are in vitro experiments on
activated immune cells. So far, no studies have been published, in which the anti-
inflammatory properties of combinations of rice bran components have been
investigated. However, several studies have investigated combinations of other
nutrients on LPS-activated microglia or macrophages. For example, the
combination eicosapentaenoic acid and resveratrol showed higher potency to
inhibit LPS-induced NO release and proinflammatory genes expression than each
individual nutrients [58]. Likewise, a combination of flavonoids synergistically
inhibited the release of NO, TNF and PGE2 [59], whereas additive and
synergistic effects of vitamin D combined with the plant extract -sitosterol on the
release of NO, TNF-α , IFN-γ , IL-6, IL-10 and MCP-1 and on the activation of
NF-B were observed in murine J774A.1 macrophage cells stimulated with LPS
[60].
The concept of combined food components has also been investigated in vivo. For
example, a combination of vegetable components, containing polyphenols and
amino acids has been shown to improve spatial memory and modulate
inflammation in aged rats [61]. A similar approach could be applied to the
combinations of the rice bran components discussed in this review.
In vitro studies on single components may help to reveal the specific pathways that
rice bran components may affect. This information can be used to design studies
with multiple nutrient combinations acting on different pathways in order to induce
possible synergistic effects of these nutrients. Multi-nutrient dietary
supplementation targeting inflammation is an interesting concept for safe

Chapter 3
60
interventions in humans, given the low toxicity of common food supplements [62].
However, this compelling approach first needs to be tested in animal models of
diseases associated with neuroinflammation.
4. Concluding remarks
Rice bran components are emerging as potential anti-inflammatory agents. Some
of them have been demonstrated to act on peripheral and central inflammation
(figure 1). Although some promising results have been obtained in vitro and in vivo
(table 1 and 2), rice bran components still need to be further investigated in animal
models and subsequently in clinical trials in order to evaluate their potency to
prevent or cure brain diseases via modulation of neuroinflammation. To achieve
adequate efficacy, rice bran components may need to be combined with other anti-
inflammatory nutrients.
Better knowledge on nutritional intervention strategies can also contribute to the
development of lifestyle recommendations concerning healthy nutritional behavior,
which could be beneficial for general public.
Conflict of interest This study is part of the BrainMenu project and was
financially supported by the STW-Danone Partnership Program. J.M. Verkuyl and
L.M. Broersen are employees of Nutricia Research and therefore declare potential
conflicts of interest. All other authors report no financial interest or potential
conflicts of interest.
Figures and tables
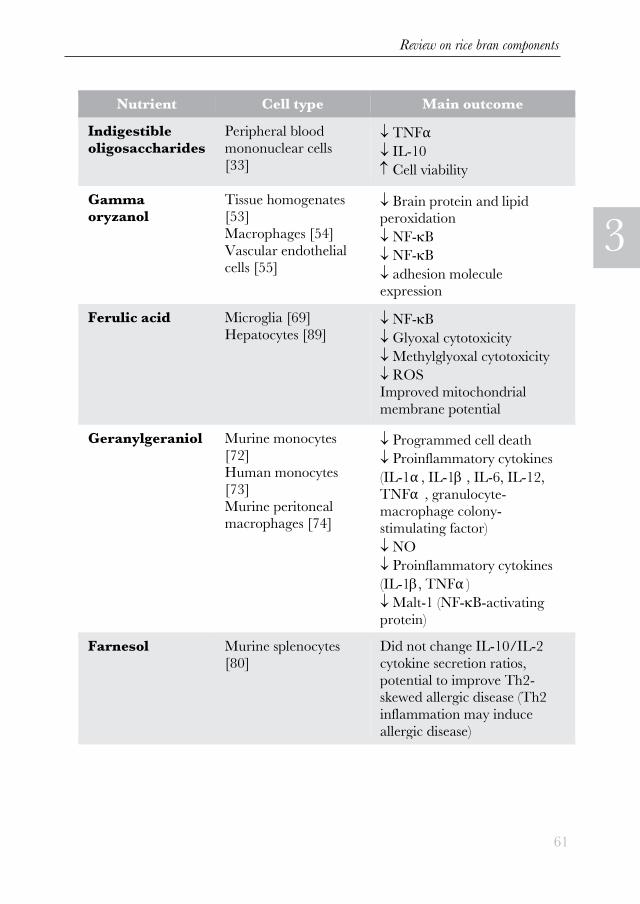
Table 1 The effects of nutrients on inflammation in cell models, divided according
to the cell type used.(on the next page)
Review on rice bran components
61
3
Nutrient Cell type Main outcome Indigestible
oligosaccharides Peripheral blood
mononuclear cells
[33]
TNFα IL-10 Cell viability
Gamma oryzanol
Tissue homogenates [53] Macrophages [54] Vascular endothelial cells [55]
Brain protein and lipid peroxidation NF-B NF-B adhesion molecule expression
Ferulic acid Microglia [69] Hepatocytes [89]
NF-B Glyoxal cytotoxicity Methylglyoxal cytotoxicity ROS Improved mitochondrial
membrane potential
Geranylgeraniol Murine monocytes
[72] Human monocytes
[73] Murine peritoneal macrophages [74]
Programmed cell death Proinflammatory cytokines
(IL-1α , IL-1 , IL-6, IL-12,
TNFα , granulocyte-macrophage colony-
stimulating factor) NO Proinflammatory cytokines
(IL-1, TNFα ) Malt-1 (NF-B-activating
protein) Farnesol Murine splenocytes
[80] Did not change IL-10/IL-2
cytokine secretion ratios,
potential to improve Th2-skewed allergic disease (Th2
inflammation may induce
allergic disease)

Chapter 3
62
Table 2 The effects of nutrients on inflammation and brain functions in animals
and humans, divided according to the model used. Abbreviations: BDNF - brain
derived neurotrophic factor, cPLA2 – calcium-dependent cytosolic phospholipase
A2, CSF – cerebrospinal fluid, DMH - 1,2-dimethylhydrazine, FST - forced swim
test, DSS – dextran sodium sulfate, MCAO - middle cerebral artery occlusion,
MPTP - 1-methyl-4 phenyl-1, 2, 3, 6-tetrahydropyridine, PBMCs – peripheral
blood mononuclear cells, TNBS - 2,4,6-trinitrobenzene sulfonic acid, TS - tail
suspension test (on the next pages).

Review on rice bran components
63
3

Chapter 3
64
Figure 1 A schematic overview of the potential role of nutrients described in this
review on the regulation of neuroinflammation.
References:
[1] Fischer R, Maier O. Interrelation of oxidative stress and inflammation in
neurodegenerative disease: role of TNF. Oxid Med Cell Longev 2015;2015:610813.
doi:10.1155/2015/610813.
[2] Kothur K, Wienholt L, Brilot F, Dale RC. CSF cytokines/chemokines as
biomarkers in neuroinflammatory CNS disorders: A systematic review. Cytokine
2016;77:227–37. doi:10.1016/j.cyto.2015.10.001.
[3] Doorduin J, de Vries EFJ, Willemsen ATM, de Groot JC, Dierckx RA, Klein HC.
Neuroinflammation in schizophrenia-related psychosis: a PET study. J Nucl Med
2009;50:1801–7. doi:10.2967/jnumed.109.066647.
[4] Haarman BCMB, Riemersma-Van der Lek RF, de Groot JC, Ruhé HGE, Klein
HC, Zandstra TE, et al. Neuroinflammation in bipolar disorder - A [(11)C]-(R)-
PK11195 positron emission tomography study. Brain Behav Immun 2014;40:219–
25. doi:10.1016/j.bbi.2014.03.016.
[5] Brites D, Fernandes A. Neuroinflammation and Depression: Microglia Activation,
Extracellular Microvesicles and microRNA Dysregulation. Front Cell Neurosci
2015;9:476. doi:10.3389/fncel.2015.00476.
[6] Bassani TB, Vital MABF, Rauh LK. Neuroinflammation in the pathophysiology of
Parkinson’s disease and therapeutic evidence of anti-inflammatory drugs. Arq

Review on rice bran components
65
3
Neuropsiquiatr 2015;73:616–23. doi:10.1590/0004-282X20150057.
[7] Lim SL, Rodriguez-Ortiz CJ, Kitazawa M. Infection, systemic inflammation, and
Alzheimer’s disease. Microbes Infect 2015;17:549–56.
doi:10.1016/j.micinf.2015.04.004.
[8] Sankowski R, Mader S, Valdés-Ferrer SI. Systemic inflammation and the brain:
novel roles of genetic, molecular, and environmental cues as drivers of
neurodegeneration. Front Cell Neurosci 2015;9:28. doi:10.3389/fncel.2015.00028.
[9] Collins LM, Toulouse A, Connor TJ, Nolan YM. Contributions of central and
systemic inflammation to the pathophysiology of Parkinson’s disease.
Neuropharmacology 2012;62:2154–68. doi:10.1016/j.neuropharm.2012.01.028.
[10] Häuser W, Janke K-H, Klump B, Hinz A. Anxiety and depression in patients with
inflammatory bowel disease. Inflamm Bowel Dis 2011;17:621–32.
doi:10.1002/ibd.21346.
[11] Panara a J, Yarur a J, Rieders B, Proksell S, Deshpande a R, Abreu MT, et al.
The incidence and risk factors for developing depression after being diagnosed with
inflammatory bowel disease: a cohort study. Aliment Pharmacol Ther
2014;39:802–10. doi:10.1111/apt.12669.
[12] Maes M. The cytokine hypothesis of depression: inflammation, oxidative &
nitrosative stress (IO&NS) and leaky gut as new targets for adjunctive treatments in
depression. Neuro Endocrinol Lett 2008;29:287–91.
[13] Hovens IB, Schoemaker RG, van der Zee E a., Absalom AR, Heineman E, van
Leeuwen BL. Postoperative cognitive dysfunction: Involvement of
neuroinflammation and neuronal functioning. Brain Behav Immun 2014;38:202–
10. doi:10.1016/j.bbi.2014.02.002.
[14] Hovens IB, Schoemaker RG, van der Zee EA, Heineman E, Izaks GJ, van
Leeuwen BL. Thinking through postoperative cognitive dysfunction: How to bridge
the gap between clinical and pre-clinical perspectives. Brain Behav Immun
2012;26:1169–79. doi:10.1016/j.bbi.2012.06.004 [doi].
[15] Smith JA, Das A, Ray SK, Banik NL. Role of pro-inflammatory cytokines released
from microglia in neurodegenerative diseases. Brain Res Bull 2012;87:10–20.
doi:10.1016/j.brainresbull.2011.10.004.
[16] Tsivgoulis G, Psaltopoulou T, Wadley VG, Alexandrov A V, Howard G,
Unverzagt FW, et al. Adherence to a Mediterranean diet and prediction of incident
stroke. Stroke 2015;46:780–5. doi:10.1161/STROKEAHA.114.007894.
[17] Pelletier A, Barul C, Féart C, Helmer C, Bernard C, Periot O, et al. Mediterranean
diet and preserved brain structural connectivity in older subjects. Alzheimers
Dement 2015;11:1023–31. doi:10.1016/j.jalz.2015.06.1888.
[18] Sharif MK, Butt MS, Anjum FM, Khan SH. Rice Bran: A Novel Functional
Ingredient. Crit Rev Food Sci Nutr 2013;54:807–16.
doi:10.1080/10408398.2011.608586.
[19] Friedman M. Rice brans, rice bran oils, and rice hulls: composition, food and
industrial uses, and bioactivities in humans, animals, and cells. J Agric Food Chem
2013;61:10626–41. doi:10.1021/jf403635v.
[20] Johnson RW. Feeding the beast: can microglia in the senescent brain be regulated
by diet? Brain Behav Immun 2015;43:1–8. doi:10.1016/j.bbi.2014.09.022.
[21] Jeurink P V., Van Esch BC a M, Rijnierse A, Garssen J, Knippels LMJ.

Chapter 3
66
Mechanisms underlying immune effects of dietary oligosaccharides. Am J Clin Nutr
2013;98:572–7. doi:10.3945/ajcn.112.038596.
[22] Son MJ, Rico CW, Nam SH, Kang MY. Effect of Oryzanol and Ferulic Acid on
the Glucose Metabolism of Mice Fed with a High-Fat Diet 2011:4–7.
doi:10.1111/j.1750-3841.2010.01907.x.
[23] Candiracci M, Ph D, Luisa M, Justo MD, Casta A, D RRP, et al. Rice bran
enzymatic extract – supplemented diets modulate adipose tissue in fl ammation
markers in Zucker rats 2014;30:466–72. doi:10.1016/j.nut.2013.09.016.
[24] Juliano C, Cossu M, Alamanni MC, Piu L. Antioxidant activity of gamma-
oryzanol : Mechanism of action and its effect on oxidative stability of
pharmaceutical oils 2005;299:146–54. doi:10.1016/j.ijpharm.2005.05.018.
[25] Parrado J. Prevention of brain protein and lipid oxidation elicited by a water-
soluble oryzanol enzymatic extract derived from rice bran 2003;314:307–14.
doi:10.1007/s00394-003-0424-4.
[26] Islam S, Yoshida H, Matsuki N, Ono K, Nagasaka R, Ushio H. Full Paper
Antioxidant , Free Radical – Scavenging , and NF- κ B – Inhibitory Activities of
Phytosteryl Ferulates : Structure – Activity Studies 2009;337:328–37.
doi:10.1254/jphs.09146FP.
[27] Sakai S, Murata T, Tsubosaka Y, Ushio H, Ozaki H. γ -Oryzanol Reduces
Adhesion Molecule Expression in Vascular Endothelial Cells via Suppression of
Nuclear Factor- κ B Activation 2012.
[28] Zhu D, Brambilla D, Leroux J-C, Nyström L. Permeation of steryl ferulates
through an in vitro intestinal barrier model. Mol Nutr Food Res 2015;59:1182–9.
doi:10.1002/mnfr.201400862.
[29] Islam MS, Murata T, Fujisawa M, Nagasaka R, Ushio H, Bari AM, et al. Anti-
inflammatory effects of phytosteryl ferulates in colitis induced by dextran sulphate
sodium in mice 2008:812–24. doi:10.1038/bjp.2008.137.
[30] Wang O, Liu J, Cheng Q, Guo X, Wang Y, Zhao L, et al. Effects of Ferulic Acid
and γ -Oryzanol on Metabolic Syndrome in Rats 2015:1–14.
doi:10.1371/journal.pone.0118135.
[31] Roy S, Metya SK, Sannigrahi S, Rahaman N, Ahmed F. Treatment with ferulic
acid to rats with streptozotocin-induced diabetes: Effects on oxidative stress, pro-
inflammatory cytokines, and apoptosis in the pancreatic ?? cell. Endocrine
2013;44:369–79. doi:10.1007/s12020-012-9868-8.
[32] Kamiya T, Shikano M, Tanaka M, Ozeki K, Ebi M, Katano T, et al. Therapeutic
Effects of Biobran, Modified Arabinoxylan Rice Bran, in Improving Symptoms of
Diarrhea Predominant or Mixed Type Irritable Bowel Syndrome: A Pilot,
Randomized Controlled Study. Evidence-Based Complement Altern Med
2014;2014:1–6. doi:10.1155/2014/828137.
[33] Koh P-O. Ferulic acid prevents the cerebral ischemic injury-induced decrease of
Akt and Bad phosphorylation. Neurosci Lett 2012;507:156–60.
doi:10.1016/j.neulet.2011.12.012.
[34] Sung J-H, Gim S-A, Koh P-O. Ferulic acid attenuates the cerebral ischemic injury-
induced decrease in peroxiredoxin-2 and thioredoxin expression. Neurosci Lett
2014;566:88–92. doi:10.1016/j.neulet.2014.02.040.
[35] Cheng CY, Su SY, Tang NY, Ho TY, Chiang SY, Hsieh CL. Ferulic acid provides

Review on rice bran components
67
3
neuroprotection against oxidative stress-related apoptosis after cerebral
ischemia/reperfusion injury by inhibiting ICAM-1 mRNA expression in rats. Brain
Res 2008;1209:136–50. doi:10.1016/j.brainres.2008.02.090.
[36] Jin Y, Yan E, Li X, Fan Y, Zhao Y, Liu Z, et al. Neuroprotective effect of sodium
ferulate and signal transduction mechanisms in the aged rat hippocampus. Acta
Pharmacol Sin 2008;29:1399–408. doi:10.1111/j.1745-7254.2008.00848.x.
[37] Swaraj Bandhu, Sikder K, A K, Das DK, Khan A, Das N, et al. Promising role of
ferulic acid, atorvastatin and their combination in ameliorating high fat diet-
induced stress in mice. Life Sci 2013;92:938–49. doi:10.1016/j.lfs.2013.03.015.
[38] Das U, Manna K, Sinha M, Datta S, Das DK, Chakraborty A, et al. Role of ferulic
acid in the amelioration of ionizing radiation induced inflammation: a murine
model. PLoS One 2014;9:e97599. doi:10.1371/journal.pone.0097599.
[39] Zeni ALB, Zomkowski ADE, Maraschin M, Rodrigues ALS, Tasca CI.
Involvement of PKA, CaMKII, PKC, MAPK/ERK and PI3K in the acute
antidepressant-like effect of ferulic acid in the tail suspension test. Pharmacol
Biochem Behav 2012;103:181–6. doi:10.1016/j.pbb.2012.08.020.
[40] Lenzi J, Rodrigues AF, Rós A de S, de Castro BB, de Lima DD, Magro DDD, et al.
Ferulic acid chronic treatment exerts antidepressant-like effect: role of antioxidant
defense system. Metab Brain Dis 2015. doi:10.1007/s11011-015-9725-6.
[41] Xu Y, Zhang L, Shao T, Ruan L, Wang L, Sun J, et al. Ferulic acid increases pain
threshold and ameliorates depression-like behaviors in reserpine-treated mice:
behavioral and neurobiological analyses. Metab Brain Dis 2013;28:571–83.
doi:10.1007/s11011-013-9404-4.
[42] Mori T, Koyama N, Guillot-Sestier M-V, Tan J, Town T. Ferulic acid is a
nutraceutical β -secretase modulator that improves behavioral impairment and
alzheimer-like pathology in transgenic mice. PLoS One 2013;8:e55774.
doi:10.1371/journal.pone.0055774.
[43] Kim B-W, Koppula S, Park S-Y, Kim Y-S, Park P-J, Lim J-H, et al. Attenuation of
neuroinflammatory responses and behavioral deficits by Ligusticum officinale
(Makino) Kitag in stimulated microglia and MPTP-induced mouse model of
Parkinson׳s disease. J Ethnopharmacol 2014;164:1–10.
doi:10.1016/j.jep.2014.11.004.
[44] Press D. Neuroprotective potential of ferulic acid in the rotenone model of
Parkinson ’ s disease 2015:5499–510.
[45] Muraguchi T, Okamoto K, Mitake M, Ogawa H, Shidoji Y. Polished rice as
natural sources of cancer-preventing geranylgeranoic acid. J Clin Biochem Nutr
2011;49:8–15. doi:10.3164/jcbn.10-110.
[46] Tricarico PM, Kleiner G, Valencic E, Campisciano G, Girardelli M, Crovella S, et
al. Block of the mevalonate pathway triggers oxidative and inflammatory molecular
mechanisms modulated by exogenous isoprenoid compounds. Int J Mol Sci
2014;15:6843–56. doi:10.3390/ijms15046843.
[47] Marcuzzi A, Tommasini A, Crovella S, Pontillo A. Natural isoprenoids inhibit LPS-
induced-production of cytokines and nitric oxide in aminobisphosphonate-treated
monocytes. Int Immunopharmacol 2010;10:639–42.
doi:10.1016/j.intimp.2010.03.008.
[48] Kim J, Lee JN, Ye J, Hao R, Debose-Boyd R, Ye J. Sufficient production of

Chapter 3
68
geranylgeraniol is required to maintain endotoxin tolerance in macrophages. J
Lipid Res 2013;54:3430–7. doi:10.1194/jlr.M042549.
[49] Marcuzzi A, Pontillo A, Leo L De, Tommasini A, Decorti G, Not T, et al. Natural
isoprenoids are able to reduce inflammation in a mouse model of mevalonate kinase
deficiency. Pediatr Res 2008;64:177–82. doi:10.1203/PDR.0b013e3181761870.
[50] Giriwono PE, Shirakawa H, Ohsaki Y, Hata S, Kuriyama H, Sato S, et al. Dietary
supplementation with geranylgeraniol suppresses lipopolysaccharide-induced
inflammation via inhibition of nuclear factor-κ B activation in rats. Eur J Nutr
2013;52:1191–9. doi:10.1007/s00394-012-0429-y.
[51] Qamar W, Sultana S. Farnesol ameliorates massive inflammation, oxidative stress
and lung injury induced by intratracheal instillation of cigarette smoke extract in
rats: An initial step in lung chemoprevention. Chem Biol Interact 2008;176:79–87.
doi:10.1016/j.cbi.2008.08.011.
[52] Goto T, Kim Y-I, Funakoshi K, Teraminami A, Uemura T, Hirai S, et al.
Farnesol, an isoprenoid, improves metabolic abnormalities in mice via both
PPARα -dependent and -independent pathways. Am J Physiol Endocrinol Metab
2011;301:E1022-32. doi:10.1152/ajpendo.00061.2011.
[53] Khan R, Sultana S. Farnesol attenuates 1,2-dimethylhydrazine induced oxidative
stress, inflammation and apoptotic responses in the colon of Wistar rats. Chem Biol
Interact 2011;192:193–200. doi:10.1016/j.cbi.2011.03.009.
[54] Ku CM, Lin JY. Anti-inflammatory effects of 27 selected terpenoid compounds
tested through modulating Th1/Th2 cytokine secretion profiles using murine
primary splenocytes. Food Chem 2013;141:1104–13.
doi:10.1016/j.foodchem.2013.04.044.
[55] Leonhardt I, Spielberg S, Weber M, Albrecht-eckardt D, Bläss M, Claus R, et al.
The Fungal Quorum-Sensing Molecule Farnesol Activates Innate Immune Cells
but Suppresses Cellular Adaptive Immunity. MBio 2015;6:1–14.
doi:10.1128/mBio.00143-15.Invited.
[56] Santhanasabapathy R, Vasudevan S, Anupriya K, Pabitha R, Sudhandiran G.
Farnesol quells oxidative stress, reactive gliosis and inflammation during
acrylamide-induced neurotoxicity: Behavioral and biochemical evidence.
Neuroscience 2015;308:212–27. doi:10.1016/j.neuroscience.2015.08.067.
[57] Khodabakhsh P, Shafaroodi H, Asgarpanah J. Analgesic and anti-inflammatory
activities of Citrus aurantium L. blossoms essential oil (neroli): involvement of the
nitric oxide/cyclic-guanosine monophosphate pathway. J Nat Med 2015:324–31.
doi:10.1007/s11418-015-0896-6.
[58] Pallarès V, Calay D, Cedó L, Castell-Auví A, Raes M, Pinent M, et al. Enhanced
anti-inflammatory effect of resveratrol and EPA in treated endotoxin-activated
RAW 264.7 macrophages. Br J Nutr 2012;108:1562–73.
doi:10.1017/S0007114511007057.
[59] Harasstani OA, Moin S, Tham CL, Liew CY, Ismail N, Rajajendram R, et al.
Flavonoid combinations cause synergistic inhibition of proinflammatory mediator
secretion from lipopolysaccharide-induced RAW 264.7 cells. Inflamm Res
2010;59:711–21. doi:10.1007/s00011-010-0182-8.
[60] Alappat L, Valerio M, Awad AB. Effect of vitamin D and β -sitosterol on immune
function of macrophages. Int Immunopharmacol 2010;10:1390–6.

Review on rice bran components
69
3
doi:10.1016/j.intimp.2010.08.003.
[61] Acosta S, Jernberg J, Sanberg CD, Sanberg PR, Small BJ, Gemma C, et al. NT-
020, a natural therapeutic approach to optimize spatial memory performance and
increase neural progenitor cell proliferation and decrease inflammation in the aged
rat. Rejuvenation Res 2010;13:581–8. doi:10.1089/rej.2009.1011.
[62] Di Lorenzo C, Ceschi A, Kupferschmidt H, Lüde S, De Souza Nascimento E, Dos
Santos A, et al. Adverse effects of plant food supplements and botanical
preparations: a systematic review with critical evaluation of causality. Br J Clin
Pharmacol 2015;79:578–92. doi:10.1111/bcp.12519.

Top Related