







Languages
Pages
Legal


Programmed Cell Death in Trichodesmium: Implications for Blooms
at Sea
Ilana Berman-Frank , 1. Faculty of Life Sciences. Bar Ilan University, Israel.

Co-authors…..
Xavier MariIRD – New Caledonia
Institute of Marine and Coastal Sciences, Rutgers University,USA
•Liti Haramaty
•Kay BidlePaul Falkowski

http://www.usc.edu/dept/LAS/biosci/tricho/trichohome/World_Tricho.htm
Trichodesmium spp.
Major pelagic diazotroph

•Contribution of Trichodesmium to:
• Global N2 fixation – 36% (Gallon 2001)
• Marine N2 fixation - 50-90% (Capone)
Trichodesmium spp.

Demise of Trichodesmium Blooms ?
Grazing by harpacticoidcopepods: Macrosetella(O’neil , J.Plankton Res. 1998).
Photo: JM O’neil
BacteriophageInfection and Lysis(Ohki, 1999)

0
100
200
300
400
500
600
0 5 10 15 20 25 30 35 40
days from inoculum
µg C
, NBUT:
Trichodesmium often displays rapid biomass crashes without presence of grazers or viruses
~50% drop in biomass in 2 days
WHY??

Pathways of death….
Modified from Danon et al. Plant Physiol. Biochem. 2000
ENVIRONMENTALSTRESS
ACCIDENTALDEATH
PHYSIOLOGICAL CELL
DEATH
NECROSIS APOPTOSISPCD
DNA smear, rupture of nuclear, organelle and
plasma membranes
Genetic program: caspases, DNA
fragmentation(ladder)
Morphology: cell, nucleus, apoptotic bodies,
membranes intact
DEV
ELOPMEN
T
grazing

OBJECTIVES
• DOES AN APOPTOTIC, AUTOCATALYTIC PROGRAMMED CELL DEATH (PCD) EXIST AND OPERATE IN TRICHODESMIUM ?
OBJECTIVE

APOPTOTIC, PCD CHARACTERISTICS
• MORPHOLOGICAL
• SYNTHESIS and ACTIVITY OF PROTEASES and NUCLEASES
• GENETIC POTENTIAL

0
200
400
600
0 10 20 30 40
days from culture initiation
µg C
0
200
400
600
0 10 20 30 40
days from culture initiation
µg C
thth
Vacuolization
Vacuolization
gv
gv
gv
cbpc
ld
1 mu
MORPHOLOGICAL EVIDENCE:
•Membranes intact
•Increased vacuolization.
• Less phycocyanin, carboxysomes, thylakoids.
•lipid droplets.
• Breaks between cells.
Berman-Frank et al. L&O 2004

MORPHOLOGICAL EVIDENCE:Membrane impermeable stains
SYTOX-green
Boiled cells - NECROSISCells under oxidative stress - APOPTOTIC

DNA fragmentationTerminal Deoxynucleotide Transferase
dUTP Nick End (TUNEL) labeling
A. C.B.
5 µm 5 µm 5 µm
Healthy cells – Intact DNA
3 h after H2O2 spike –
Negative control
Berman-Frank et al. L&O 2004

CASPASES and APOPTOSIS• Caspases (cysteine aspartate-specific proteases) contain
conserved QACXG pentapeptide with cysteine in the active site.
• Central role in initiation and induction of apoptosis -Caspase cascade: initiators, executioners (Effector).– NO activation of caspases found in NECROTIC
death!• Caspase 3: main executioner caspase. Activation
essential for DNA fragmentation, chromatin condensation and plasma membrane blebbing.
BIOCHEMICAL EVIDENCE:

Increase of caspase-3 immunoactivity during Trichodesmium sp. IMS101 bloom
progression.
Whole cell protein extracts were challenged with polyclonal antibodies to recombinant human caspase 3
Bloom decline
BIOCHEMICAL EVIDENCE:
Berman-Frank et al. L&O 2004

Dependence of DEVD cleavage on physiological status of Trichodesmium sp.
IMS101 .
0
500
1000
1500
2000
-2-1.5-1-0.500.5
Mortality rate
r = 0.968
r = 0.856
RFU
µµ µµg
Chla
-1
High DEVD cleavage
when growth is negative
Berman-Frank et al. L&O 2004
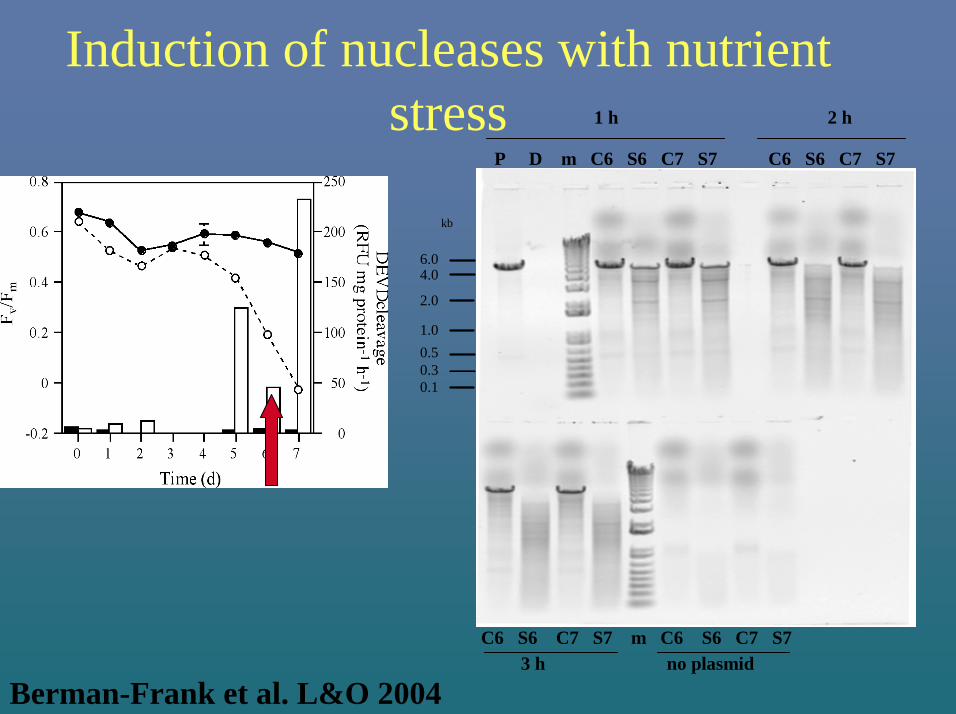
Induction of nucleases with nutrient stress
P D m C6 S6 C7 S7 C6 S6 C7 S7
1 h 2 h
C6 S6 C7 S7 m C6 S6 C7 S7 3 h no plasmid
6.0
2.0
1.00.50.30.1
4.0
kb
Berman-Frank et al. L&O 2004

Genetic potential.
• METACASPASES• Caspase-like proteins identified in plants,
fungi, unicellular protozoa and multiple bacterial species including Synechocystisand Anabaena (Uren et al., Molecular Cell6, 961-967)

Metacaspasesoverview
multiple protein sequence alignments –
(Bidle & Falkowski 2004)
Trichodesmium
humans

Key sequence similarities to yeast metacaspase Mca-1, a caspase-like enzyme that mediates PCD in aging cells . ZP_00070972 1 ----MRIEALVVGINEHVFEPG-----LNLKAPVKDAEAIAEMLEKYGNFHVQGLP---- ZP_00073861 1 MDKKNLRQALVVGINRYPLLKKKKLGDLNLKAAVKDAEAIANILEKYGKFRIQRLPSLPS ZP_00074337 1 --MTNQNIAITIGVQEYEFLTP-------LKYAANDAKKMRDFLLDEADFDDVFYLS--- ZP_00073436 1 ----MTNKAITIGVQKYQFFSP-------LKYAANDAEKMRNFLLEEAGFDEVLYYS--- ZP_00075051 139 -QPIKNRWAFLVGVNRYNDPGVH----K-LKFCVKDVQKLEKKLKELGYTVMCLHD---- ZP_00073782 1 -------YAVVIGVGDDLPITI-----------D-DATAIANVLGDLSRCAYP------- ZP_00073655 344 ----SHNYALLIGVGDYKYPDWS------LPVTVKDVQAIKSFLTNPDLCSYID------ ZP_00071249 29 ---MERRRALLIGITIFGEGLDN------FSPTLEDVQVMKKCLEEKGGFEVIQ------ YeastMCA-1 154 ---TGRRKALIIGINYIGSKNQ-------LRGCINDAHNIFNFLTNGYGYSSDD------ ZP_00070972 48 --KDYDETGRERFISDGFVYRKNLKIKVSNLFNPISKNEIPDVALFFFAGHGFVTTEGGV ZP_00073861 61 LPKNYDQEGTERFDPKGKVKINELQEAIINLFKPRKKNETPDVALLFFAGHGYVDEKGDI ZP_00074337 49 ------DNSPKINGASTRPTRSRLELVLEDEVKKLSLK-TGDNLWFFFSGHGHRENNN-- ZP_00073436 47 ------DYSPEINGDYTRPTRSNLEFLLENQFKEPFMG-IGDNFWFFFSGHGLRENG--- ZP_00075051 189 ------DLESDSR---RFPIHNNIEAELINLCNMVDVD---DLLLVYFACHGKLGRDE-- ZP_00073782 35 ------TDQVRLLTG-EKANRANVLSALSWLAETTGKD---DTAIVYFSGHGIEKPD--- ZP_00073655 388 ------DENHLRLLCNEQATKQNILNNINWLQEQAKND-PEATILVYYSGHGWLDKST-- ZP_00071249 74 ----------E-----ENLDRKDMEYAINKFFYESQKS---DTLLIYVSSHGITDAEG-- YeastMCA-1 198 ----IVILTDDQNDLVRVPTRANMIRAMQWLVKDAQPN---DSLFLHYSGHGGQTEDLDG
* ZP_00070972 106 REGFLVTSDV----QLKRDIYGISLSWLKDLLRQSPVKKQIVWLDCCFGGELLNSQ--- ZP_00073861 121 REGFLATSEA----HLSENVYGISLNWLKRLLQDSPVQEQIVWLDCCFSGEFLNFDRE- ZP_00074337 100 -IDYLIPIDG----HSNVERSGISVDYIIQQLQKSGADNIVLILDACRNKSDGGKGG-- ZP_00073436 97 -IDYLIPVDG----YKNVQKSGISVNYIIQQLQKCGADNVVLILDACRDEGDARRGGK- ZP_00075051 235 QPMLIVRDTR----LPTIEKTGLSLADIKKSMHSSKARRLVLTLDACHMGVETGRGID- ZP_00073782 82 -YYLMPYGYN----LENLEGTAIRGETFTECLRAIKTKKLLVLLDCCHAGGQADPKGII ZP_00073655 439 EKYYLIPHDTSPIKLQKTQKTALPATDFNNALQEISAQKLLVIIDSCHAQGMATAKE-- ZP_00071249 113 -TFYIATSETSYDRDTEIITGAIEASYFHRQIKNCTSNSQVWLLDLCYSGAFVRGYSK- YeastMCA-1 251 DEEDGMDDVIYPVDFETQGPIIDDEMHDIMVKPLQQGVRLTALFDSCHSGTVLDLPYT-
*
histidine- and cysteine-containing catalytic diadfound in true caspases (p20 subunit), paracaspases, metacaspases
PSI-BLAST Berman-Frank et al. L&O 2004

16S rRNA –Potential metacaspase inheritance
(Bidle & Falkowski 2004. Nature Rev. Microbiol.)
No metacaspasesfound (genes lost?)
Diazotrophs –bloom formers
Thermophiliccyanobact.
Purple non-sulphur

Conserved PCD domains detected in Trichodesmium
ZP_00070972ZP_00073861
Eukaryotic proteinsAdaptor/regulatory in signal transd., pre-mRNA processing
WD-40 domain
ZP_00072978Shared by plant resistance genes and regulators of cell death in animals
AP-ATPase,NB-ARC domain
ZP_00073901ZP_00073386ZP_00071486
Signal transductionPredicted NTPase(NACHT family)
ZP_00072870ZP_00073979ZP_00071776
Posttranslational modification, protein turnover, chaperones
DegQ trypsin-like serine proteaseHtrA-like protease
T. erythraeum protein sequence
Domain functionDomain description

Evidence for PCD in Trichodesmium:
•MORPHOLOGICAL: SYTOX, TEM, TUNEL
•PRESENCE OF PCD COMPONENTS: metacaspasesimilarity, DEATH Domains
•SYNTHESIS AND ACTIVITY OF CASPASE-LIKE PROTEINS:
• A. Westerns – human caspase-3
• B. Activity increased with stress and age of culture
• C. Activity inhibited by caspase-3 inhibitors

From Lab To Sea…..
• Project DIAPAZON –• New Caledonia

Objective
• DOES PCD OCCUR IN NATURAL POPULATIONS UNDER ENVIRONMENTAL STRESS?

Looking for Tricho….

Le vent chasse les Tricho….
Plan A: GONE WITH THE WIND….

Plan B: Simulations!

The Noumea experiments

Morphological Changes
3 h
23 h
8 h
Bottom of flask

0
2
4
6
8
10
12
Time from high light induction (h)
DEV
D R
FU µµ µµ
g pr
otei
n-1
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
µµ µµM le
uAM
C µµ µµ
g pr
otei
n-1
0 3 6 9 12 15 18 21 24
A
iiiiii
DEVD and General Proteolitic Activity

OXIDATIVE STRESS INDUCESAPOPTOSIS
(Chandra et al. Free Rad. Biol Med. 2000)
DEVD ACTIVITY
T. erythraeum
Buoyant trichomes
Sinking trichomes
0.3 ± 0.22 RFU µg protein-1
range (0.04 – 0.7) 17.1 ± 0.22 RFU µg protein-1

PCD and the Fate of TrichodesmiumBlooms ?

Transparent exopolymeric particles (TEP) PRODUCTION AND
BLOOM DECLINE• Source of organic matter for sedimentation and
biogeochemical recycling. • Sorption sites for dissolved organic and inorganic matter.• Attachment sites for bacteria.• Food source for microphageous protozoans, for particle
grazers such as copepods or for passive filter feeders such as tunicates,
• TEP can coagulate with other particles into marine snow aggregates, and enhance the vertical flux of major elements, and of adsorbed trace elements.

Hypothesis
• The production of TEP by Trichodesmium is positively coupled to
the induction of a PCD pathway.
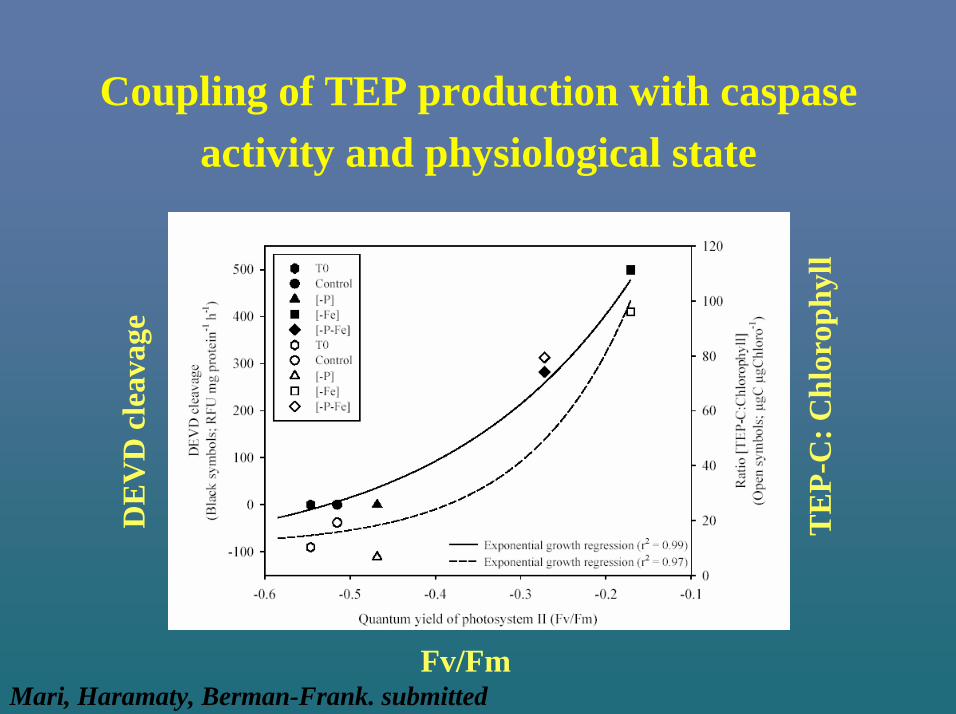
Coupling of TEP production with caspaseactivity and physiological state
Fv/Fm
DE
VD
cle
avag
e
TE
P-C
: Chl
orop
hyll
Mari, Haramaty, Berman-Frank. submitted

In natural and laboratory populations –coupling between PCD induction and TEP
production
With PCD induction:
1. Increase in caspaseactivity
2. Increase in TEP-C
3. Increase in size of TEP particles
Mari, Haramaty, Berman-Frank. submitted

TEP production in Trichodesmium
Mari, Haramaty, Berman-Frank. submitted
2-4 h 6-10 h
14-16 h

Change in TEP production with time
Mari, Haramaty, Berman-Frank. submitted
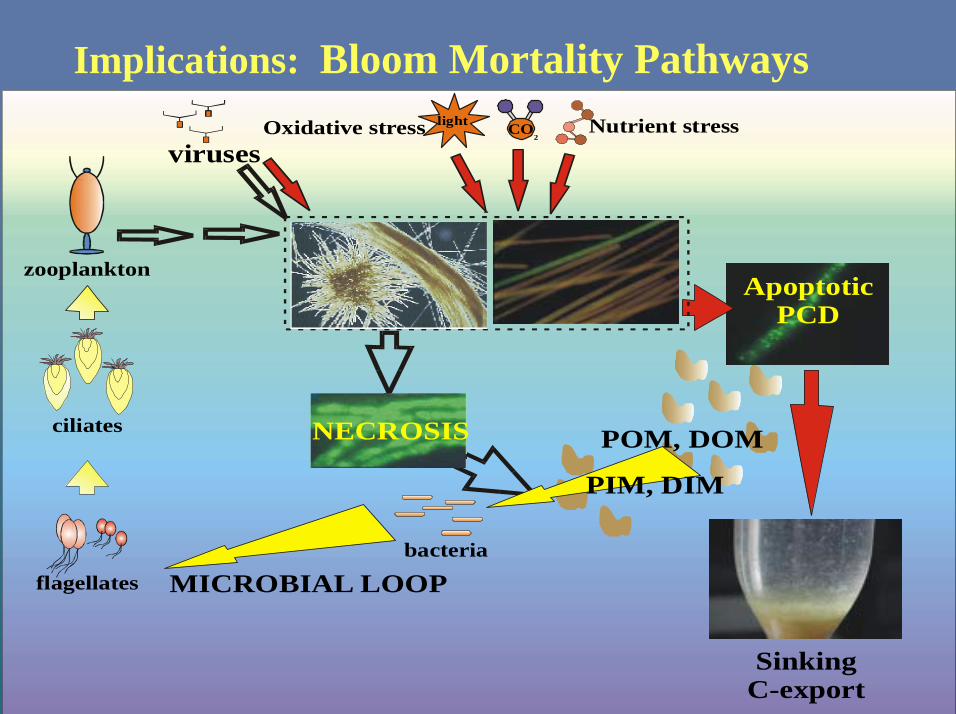
Implications: Bloom Mortality Pathways
ApoptoticPCD
bacteria
viruses
flagellates
zooplankton
ciliates
Oxidative stressCo2
light Nutrient stressCo2CO
2
SinkingC-export
POM, DOM
MICROBIAL LOOP
NECROSIS
PIM, DIM

Fate of Trichodesmium Blooms ?
Still looking!!!

Required research!!!Follow a natural bloom development
and crash (preferably in a nice, tropical paradise…).

Merci!!• C. DUPOUY,
• A. LE BOUTEILLER,
• THE WHOLE TEAM OF THE DIAPAZON PROGRAM, IRD-NOUVELLE CALEDONIE
• V. STAROVOYTOV: TEM analysis, RUTGERS U.
Top Related