







Languages
Pages
Legal


The role of CXCR1 gene polymorphisms in bovine neutrophil
traits and mastitis
Joren Verbeke
Merelbeke, 2015


A problem is a chance for you to do your best.
(Duke Ellington, jazz musician)

The role of CXCR1 gene polymorphisms in bovine neutrophil traits and mastitis Joren Verbeke Cover: The coding sequence of the bovine CXCR1 gene, Clara Vermeersch Printing: University Press, Zelzate ISBN number: 9789058644060 Printing of this thesis was financially supported by Boehringer-Ingelheim, MSD and Vétoquinol.
The reception following the oral defense was supported by Milcobel.
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by Science and Technology in Flanders (IWT Vlaanderen) and performed at Ghent University.

The role of CXCR1 gene polymorphisms in bovine neutrophil
traits and mastitis
De rol van CXCR1 gen polymorfismen in boviene neutrofiel eigenschappen en
mastitis
(met een samenvatting in het Nederlands)
Proefschrift voorgedragen tot het behalen van de graad van Doctor in de Diergeneeskundige
Wetenschappen aan de Faculteit Diergeneeskunde, Universiteit Gent, 9 januari, 2015
door
Joren Verbeke
Vakgroep Voortplanting, Verloskunde en Bedrijfsdiergeneeskunde
Vakgroep Voeding, Genetica en Ethologie
Faculteit Diergeneeskunde
Universiteit Gent
Department of Reproduction, Obstetrics and Herd Health
Department of Nutrition, Genetics, and Ethology
Faculty of Veterinary Medicine
Ghent University


Promotors
Prof. dr. Sarne De Vliegher
Faculty of Veterinary Medicine, Ghent University, Belgium
Prof. dr. Luc Peelman
Faculty of Veterinary Medicine, Ghent University, Belgium
Members of the examination committee
Prof. em. dr. dr.h.c. Aart de Kruif
Chairman – Faculty of Veterinary Medicine, Ghent University, Belgium
Prof. dr. Christian Burvenich
Faculty of Veterinary Medicine, Ghent University, Belgium
Prof. dr. Eric Cox
Faculty of Veterinary Medicine, Ghent University, Belgium
Prof. dr. Johann Detilleux
Faculty of Veterinary Medicine, University of Liège, Belgium
Prof. dr. Christa Kühn
Leibniz Institute for Farm Animal Biology, Germany
Prof. dr. Filip Van Immerseel
Faculty of Veterinary Medicine, Ghent University, Belgium
Dr. Erwin Koenen
Coöperatie Rundveeverbetering, The Netherlands
Dr. Sofie Piepers
Faculty of Veterinary Medicine, Ghent University, Belgium
Dr. Mario Van Poucke
Faculty of Veterinary Medicine, Ghent University, Belgium


Table of contents
Chapter 1 General introduction 1 Chapter 2 Aims and outline of the thesis 45 Chapter 3 The role of CXCR1 gene polymorphisms in bovine neutrophil traits 51
Chapter 3.1 Association of CXCR1 polymorphisms with apoptosis, necrosis and concentration of milk neutrophils in early lactating dairy heifers
53
Chapter 3.2 Reactive oxygen species generation of bovine blood neutrophils with different CXCR1 (IL8RA) genotype following Interleukin-8 incubation
69
Chapter 4 The role of CXCR1 gene polymorphisms in subclinical mastitis 87
Chapter 4.1 Pathogen-group specific association between CXCR1 polymorphisms and subclinical mastitis in dairy heifers
89
Chapter 4.2 Somatic cell count and milk neutrophil viability of dairy heifers with specific CXCR1 genotypes following experimental intramammary infection with Staphylococcus chromogenes originating from milk
111
Chapter 5 The role of CXCR1 gene polymorphisms in clinical mastitis 123
Chapter 5.1 Pathogen-specific incidence rate of clinical mastitis in Flemish dairy herds, severity, and association with herd hygiene
125
Chapter 5.2 Associations between CXCR1 polymorphisms and pathogen-specific incidence rate of clinical mastitis, test-day somatic cell count and test-day milk yield
147
Chapter 6 Differential expression of CXCR1 and commonly used reference genes
in bovine milk somatic cells following experimental intramammary infection
177
Chapter 7 General discussion 197
Summary 223
Samenvatting 231
Curriculum vitae - Publications 239 Dankwoord – Acknowledgements 245


List of abbreviations
AUC Area under the curve C5a Complement factor 5a CAMP Christie, Atkins, Munch-Petersen CD11b Cluster of differentiation molecule 11b, a β2 integrin subunit CD18 Cluster of differentiation molecule 18, a β2 integrin subunit CFU Colony forming units CI Confidence interval Cq Quantification cycle CRV “Coöperatie Rundveeverbetering”, a cattle breeding organization CXCR1 Chemokine (C-X-C motif) receptor 1, Interleukin 8 receptor A (IL8RA) CXCR2 Chemokine (C-X-C motif) receptor 2, Interleukin 8 receptor B (IL8RB) dbSNP The Single Nucleotide Polymorphism Database DHI Dairy herd improvement DIM Days in milk dNTP Deoxynucleotide triphosphates IL-1 Interleukin 1, chemokine (C-X-C motif) ligand 1 (CXCL-1) IL-8 Interleukin 8, chemokine (C-X-C motif) ligand 8 (CXCL-8) IMI Intramammary infection IRCM Incidence rate of clinical mastitis LSM Least squares means MHC II Major histocompatibility complex class II NCBI National Center for Biotechnology Information NMC National Mastitis Council, a global organization for mastitis control and milk
quality NTP Nucleoside triphosphate OR Odds ratio OZP Opsonized zymosan particles PBS Phosphate-buffered saline PMA Phorbol 12-myristate 13-acetate QTL Quantitative trait loci rb Recombinant bovine rh Recombinant human RLU Relative light units ROS Reactive oxygen species RR Relative risk RT-qPCR Reverse transcription quantitative real-time polymerase chain reaction SCC Somatic cell count SCS Somatic cell score SNP Single nucleotide polymorphism TNF-α Tumor necrosis factor α


Chapter 1
General introduction

Chapter 1 General introduction
2
Chapter 1 General introduction
3
1. Mastitis
1.1 Introduction
Although the exact origin of lactation remains unclear, evolutionary advantages of milk
secretion are obvious. It allows mammals to supply offspring nutrients from diets that are
hard to capture or digest at an early stage in life (Oftedal, 2012). Domestication of wild cows
and consumption of cow milk were major steps in human development (Diamond, 2002). To
gain profitability and fulfil the growing demand of milk, many efforts have been done over
the last decades to increase the average daily milk yield per cow. The higher production per
cow was realized through intense genetic selection as well as improved cow nutrition and
management (Hansen, 2000). Of course, to meet those higher yields any cow should dispose
of a healthy and well-developed udder. However, udder health is continuously threatened
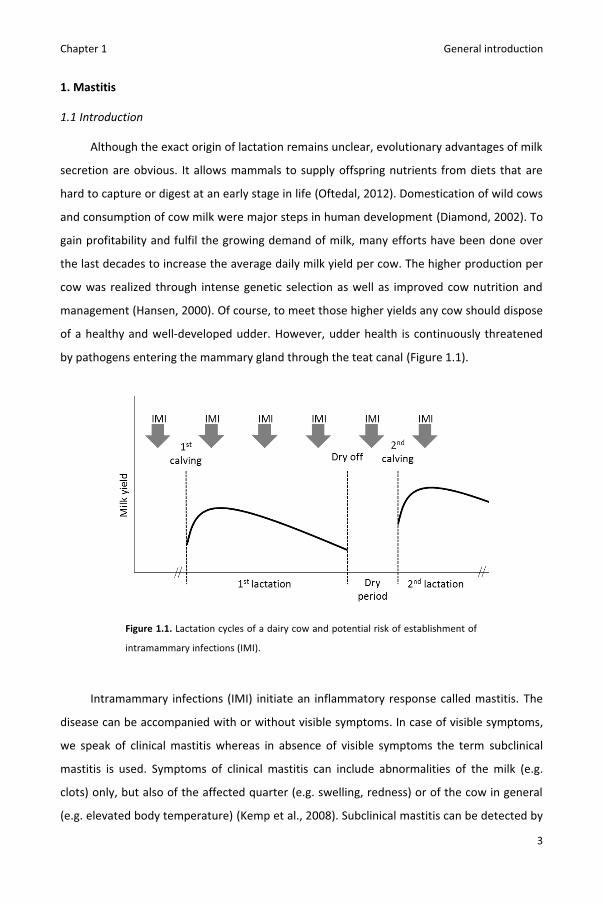
by pathogens entering the mammary gland through the teat canal (Figure 1.1).
Intramammary infections (IMI) initiate an inflammatory response called mastitis. The
disease can be accompanied with or without visible symptoms. In case of visible symptoms,
we speak of clinical mastitis whereas in absence of visible symptoms the term subclinical
mastitis is used. Symptoms of clinical mastitis can include abnormalities of the milk (e.g.
clots) only, but also of the affected quarter (e.g. swelling, redness) or of the cow in general
(e.g. elevated body temperature) (Kemp et al., 2008). Subclinical mastitis can be detected by
Figure 1.1. Lactation cycles of a dairy cow and potential risk of establishment of
intramammary infections (IMI).

Chapter 1 General introduction
4
an increase in the concentration of somatic cells in quarter or composite cow milk. The
somatic cell count (SCC) is typically expressed in cells per mL and is often transformed to
somatic cell score [SCS = LOG2(SCC/100,000) + 3] to obtain a normalized distribution
(Schukken et al., 2003). At the herd level, bulk milk SCC is measured to monitor the herd
prevalence of subclinical masitis and milk quality alongside other parameters (Schukken et
al., 2003).
Clinical and subclinical mastitis both cause a decrease in milk yield and quality (Ma et
al., 2000; Santos et al., 2003; Seegers et al., 2003). Additionally, mastitis increases
antimicrobial usage (Grave et al., 1999), hazard of culling (Beaudeau et al., 1995) and stress
for dairy producers (Lam et al., 2013). Furthermore, clinical mastitis affects animal welfare
by inflicting pain (Medrano-Galarza et al., 2012) (Figure 1.2).
For more than a century, research has been done on bovine mastitis (Delepine, 1910).
Initially, focus was put on lactating cows. More recent studies demonstrated that IMI also
occur in non-lactating cows in late gestation (dry cows) and prepartum dairy heifers (Bradley
and Green, 2004; De Vliegher et al., 2012) (Figure 1.1). Mastitis is a multifactorial disease
with pathogen, host and management factors influencing the prevalence and incidence (see
further).
Figure 1.2. Some adverse effects of mastitis
Chapter 1 General introduction
5
1.2 Pathogens involved
More than 100 different pathogens have been isolated from milk samples from the
mastitic bovine mammary gland. Intramammary infections are mainly caused by bacteria but
also algae, yeast and fungi have been associated with bovine mastitis (Watts, 1988). Mastitis
pathogens can be grouped based on their Gram staining characteristics (Gram-positive vs.
Gram-negative pathogens), the potential damage they cause to the host (major vs. minor
pathogens), or their epidemiology (contagious vs. environmental pathogens). Major
pathogens are considered to be more virulent, are more likely to cause clinical mastitis, and
result in more pronounced milk yield losses and higher SCC compared to minor pathogens
(Timms and Schultz, 1987). Contagious pathogens are often host-adapted and mainly spread
from cow to cow during milking whereas environmental pathogens are more adapted to
survive in the bovine environment (e.g. bedding) and are mainly transmitted from
environment to cow (Zadoks and Schukken, 2006) (Figure 1.3). Some mastitis pathogens
such as Streptococcus dysgalactiae show a mixed contagious-environmental epidemiology
(Zadoks et al., 2011).
Figure 1.3. Schematic representation of cow-to-cow transmission
(red) and environment-to-cow (blue) transmission of mastitis
pathogens.

Chapter 1 General introduction
6
Staphylococci and streptococci are the main causes of Gram-positive IMI (Watts,
1988). Staphylococcus aureus, a major contagious pathogen, was reported as the most
frequently isolated pathogen from clinical mastitis cases in Canada and Ireland (Keane et al.,
2013; Olde Riekerink et al., 2008) and one of the main causes of subclinical masitis in
Flanders (Piepers et al., 2007). In 2012, S. aureus was isolated from 10% of the samples of
clinical mastitis cases submitted to the Milk Control Centre Flanders (Lier, Belgium), the
largest milk laboratory in Flanders (Milk Control Centre Flanders, 2012). Virulence factors
such as biofilm formation and intracellular survival enable S. aureus to evade both the
immune system and antimicrobial agents, often resulting in chronic IMI and poor treatment
results (Bardiau et al., 2014; Bradley and Green, 2009; Melchior et al., 2006). In contrast to S.
aureus, the coagulase-negative staphylococci are regarded as minor pathogens. They are the
most frequently isolated pathogens from bovine milk samples (Piepers et al., 2007; Schukken
et al., 2009a) and the most common cause of heifer mastitis in Flanders and other regions
(De Vliegher et al., 2012; Fox, 2009). They mainly cause transient infections with little effect
on SCC and milk yield (Timms and Schultz, 1987). Heifers infected with coagulase-negative
staphylococci even outproduce non-infected herdmates (Piepers et al., 2010; Piepers et al.,
2013). Some coagulase-negative Staphylococcus species appear to be more host-adapted
whereas other species were mainly isolated from environmental habitats (De Visscher et al.,
2014).
Streptococcus uberis, Streptococcus dysgalactiae and Streptococcus agalactiae are the
most important streptococci associated with bovine mastitis and all three are regarded as
major pathogens (Watts, 1988). Streptococcus uberis is known as a typical environmental
pathogen, although cow-to-cow transmission has also been described (Zadoks et al., 2003).
Streptococcus uberis is a member of the group of esculin-positive cocci and not always
differentiated in routine laboratories from other esculin-positive cocci such as Enterococcus
spp. or Lactococcus spp. (e.g. Piepers et al., 2007). Esculin-positive cocci were isolated in 18%
of culture-positive milk samples of Flemish high SCC cows (Piepers et al., 2007).
Streptococcus uberis was the most frequently isolated pathogen from clinical mastitis cases
and was often cultured from subclinical masitis cases in a British study (Bradley et al., 2007).
It was isolated from 17% of the samples of clinical mastitis cases submitted to the Milk
Control Centre (Milk Control Centre Flanders, 2012). Streptococcus uberis is capable of
penetrating cells and often persists for several months in the udder (Matthews et al., 1994;

Chapter 1 General introduction
7
Schukken et al., 2011). Streptococcus dysgalactiae shows both cow to cow and environment
to cow transmission and is often cultured from clinical and subclinical mastitis cases (Olde
Riekerink et al., 2008; Whist et al., 2007). Streptococcus dysgalactiae was isolated in 4% of
culture-positive milk samples of Flemish high SCC cows (Piepers et al., 2007). Streptococcus
agalactiae, a typical contagious pathogen, used to be a major cause of clinical and subclinical
mastitis (Keefe, 1997) but is nowadays rarely isolated from milk samples in Flanders and
other developed regions (Barkema et al., 2009; Piepers et al., 2007; Sampimon et al., 2009).
The coliforms Escherichia coli and Klebsiella spp. are the most important causes of
Gram-negative IMI. Infections with these major pathogens are typically limited in time but
can cause severe clinical mastitis. However, chronic coliform IMI have also been described
(Dopfer et al., 1999). Coliforms were isolated in 2% of culture-positive milk samples of
Flemish high SCC cows (Piepers et al., 2007). Escherichia coli was found to be the most
common cause of clinical mastitis in large herds in Wisconsin (US) and isolated from 17% of
the samples of clinical mastitis cases submitted to the Milk Control Centre (Milk Control
Centre Flanders, 2012; Oliveira et al., 2013). Lipopolysaccharide or endotoxin present in the
cell wall causes an acute inflammatory response being responsible for most of the damage
to the cow (Hogan and Larry, 2003).
The distribution of mastitis-associated pathogens and bacteria on Flemish dairy herds
is summarized in Table 1.1.

Chapter 1 General introduction
8
Table 1.1. Distribution of mastitis-associated pathogens and bacteria on Flemish dairy herds
Level All cows1 High SCC cows2 Clinical mastitis3 n4 %tot.
5 n4 %tot.5 n4 %tot.
5 Sampled herds 1,087 ... 770 ... ... Sampled cows 44,677 ... 6,390 ... ... Sampled quarters/cases 178,668 ... 25,660 ... 4,468 ... Culture-positive cows 18,349 41.1 4,134 64.7 … Culture-positive quarters/cases 30,475 17.1 7,771 30.4 3,503 78.4 Staphylococcus aureus 5,607 3.1 1,946 7.6 433 9.7 Esculin-positive cocci 4,862 2.7 1,411 5.5 851 19.0 Streptococcus dysgalactiae 717 0.4 329 1.3 278 6.2 Streptococcus agalactiae 92 0.1 119 0.5 30 0.7 Non-aureus staphylococci 17,417 9.7 3,192 12.5 336 7.5 Coliforms 250 0.1 141 0.6 875 19.6 Contaminated samples6 1,029 0.6 415 1.6 515 11.5 Other 501 0.3 218 0.8 185 4.1
1Cows sampled during mandatory cross-sectional dairy herd screenings performed between 2000 and 2002
(Piepers et al., 2007). 2Cows with a geometric mean composite SCC ≥250,000 cells/mL sampled during mandatory cross-sectional
dairy herd screenings performed between 2000 and 2002 (Piepers et al., 2007).
3Clinical mastitis cases submitted to the Milk Control Centre Flanders in 2011 (Milk Control Centre Flanders,
2012). 4Absolute number of cows/quarters. 5Percentage of total cows/quarters. 6Isolation of 3 or more different pathogens.
1.3 Prevalence and incidence
Data on the occurrence of subclinical masitis in Flanders is available from a number of
sources using Dairy Herd Improvement (DHI) data including test-day SCC measurements,
complemented with bacteriological culture results (Table 1.1). The average bulk milk SCC
was 213,000 cells/ml in 2013 (Annual report, Milk Control Centre Flanders, Lier, Belgium,
2013). As well, individual cow SCC are monitored on 3,132 out of 5,260 (60%) Flemish dairy
herds through the DHI program organized by the Cattle Breeding Organization (CRV). Using a
threshold of 150,000 and 250,000 cells/ml for heifers and multiparous cows, respectively,
the cow prevalence of high SCC on Flemish dairy herds was estimated at 22.3 % in 2013 by
the CRV (K. Huijps, CRV, Alken, The Netherlands, personal communication). Using the same
source of information, approximately a third of the Flemish dairy heifers was shown to have
an elevated (>150,000 cells/ml) SCC in early lactation (De Vliegher et al., 2001). Data from

Chapter 1 General introduction
9
1,087 mandatory cross-sectional dairy herd screenings performed between 2000 and 2002
allowed an estimation of the pathogen-specific prevalence of subclinical mastitis (Piepers et
al., 2007): 40% of the sampled cows had at least one subclinically infected quarter.
As on most Flemish dairy herds clinical mastitis cases are not recorded and rarely
sampled for culture, the exact incidence of clinical mastitis remained unknown at the start of
this thesis, unlike a number of other countries (Barkema et al., 1998; Bradley et al., 2007;
Olde Riekerink et al., 2008). Incidence rates ranged from 5.5 quarter cases per 10,000 cow-
days at risk in French herds with a low bulk milk SCC to 12.9 quarter cases per 10,000 cow-
days at risk on randomly selected herds in England and Wales (Barkema et al., 1998;
Barnouin et al., 2005; Bradley et al., 2007; Olde Riekerink et al., 2008; Wolff et al., 2012).
1.4 Factors affecting prevalence and incidence
In brief, mastitis prevalence and incidence are influenced by infection pressure and
host resistance.
Mastitis pathogens are abundantly present on the bovine skin, milking machine unit
liners, bedding and numerous other extramammary habitats (De Visscher et al., 2014;
Piessens et al., 2011; Verbist et al., 2011; Zadoks et al., 2011). The higher the exposure to
mastitis pathogens, the more likely new IMI will occur. The total exposure to pathogens
likely to cause IMI is referred to as infection pressure. Not only the total number but also
virulence of the pathogens is of importance. For example, a few colony forming units (CFU)
of S. aureus are capable of causing IMI (Bannerman et al., 2004b) whereas more than 106
CFU S. chromogenes are required to invoke a long-lasting IMI (Simojoki et al., 2009).
Under similar infection pressure, some cows suffer from mastitis whereas other
remain healthy. To establish IMI, pathogens need to penetrate the teat canal and succeed in
surviving and multiplying in the mammary gland. The cow’s ability to hinder pathogens from
doing so is referred to as mastitis resistance, as opposed to mastitis susceptibility. In cows
with high mastitis resistance, pathogens are less likely to penetrate the teatcanal and more
effectively eliminated by the mammary gland immunity. Anatomical-physical barriers of the
teat canal form the first protection against pathogens. Between milkings and during the dry
period, the teat canal is (partially) sealed from the environment. Presence of (a) keratin
(plug) and contraction of the sphincter muscle in the teat end physically hinder pathogens to

Chapter 1 General introduction
10
enter the mammary gland. Keratin consists of a waxy material derived from the stratified
epithelium forming a physical barrier and having limited antibacterial characteristics (Hogan
et al., 1987). The anatomical-physical barriers function less in certain cows compared to
others. The keratin plug is not formed immediately after dry-off and already disappears
before calving making the first and last week of the dry period risk periods for new IMI
(Bradley and Green, 2004). Some cows fail to form a keratin plug in one or more quarters
making them more susceptible to IMI during the dry period (Dingwell et al., 2004). After
penetrating the teat canal, immune responses hinder pathogens to survive and multiply.
Some cows respond more effectively to pathogens compared to others (Burvenich et al.,
2003). The mammary gland immunity and factors affecting it are discussed in more detail in
the second part of this chapter.
Management factors can influence both infection pressure and host resistance
resulting in a lower or higher mastitis prevalence and incidence (Barkema et al., 1999;
Piepers et al., 2011). For example, cleaning the cubicles more frequently can reduce the
exposure to environmental pathogens and reduce clinical mastitis incidence (Schukken et al.,
1990). Malfunction of the milking machine can induce teat-end callosity interfering with the
anatomical-physical barriers of the teat canal and reducing host resistance (Neijenhuis et al.,
2001). Management factors such as nutrition influence mammary gland immunity
(O'Rourke, 2009) (see further).
1.5 Prevention and control
Several mastitis control programs have been designed all based on the same principle,
namely reducing the prevalence of IMI in a herd by lowering the incidence of new IMI and
shortening the duration of existing IMI (Bramley and Dodd, 1984). New IMI can be prevented
by reducing infectious pressure, increasing host resistance and prophylactic use of
antibiotics and teat sealants during the dry period. Duration of existing IMI can be shortened
by increasing host resistance to enhance the likelihood of spontaneous cure, therapeutic use
of antibiotics and culling of chronically infected animals (Bradley, 2002; Lam et al., 2013;
LeBlanc et al., 2006). Research in the 1960ies led to the development of the “Five-Point
Plan” including following elements: rapid identification and treatment of clinical mastitis,
blanket dry cow therapy, post milking teat disinfection, culling of chronically infected cows

Chapter 1 General introduction
11
and proper maintenance of the milking machine (Neave et al., 1969). Implementation of
these control measures caused a large reduction in the prevalence and incidence of clinical
and subclinical mastitis primarly caused by the contagious mastitis pathogens, the most
important pathogens at that time. Because the control measures were less successful
against environmental pathogens, an extended mastitis control program was suggested by
the National Mastitis Council (NMC, a global organization for mastitis control and milk
quality). This so-called “10-point mastitis control plan” included new elements such as
maintenance of a clean, dry, and comfortable environment (National Mastitis Council, 2014).
Herds with heifer mastitis problems, however, require a different practical approach. De
Vliegher et al. (2012) recently proposed a 10-point program to prevent and control heifer
mastitis including elements such as controlling cross-suckling in calves and young stock, fly
control, housing of primigravid heifers separate from multiparous cows and use of
prepartum antibiotic treatment in heifers under certain specific conditions.
2. Mammary gland immunity
2.1 Introduction
After penetrating the teat canal, pathogens are recognized by resident immune cells
and an immune response is initiated. In the early stage of infection, innate responses
combat invading pathogens with migration of leukocytes from blood to site of infection
being a key element (Sordillo et al., 1997). When the innate immunity fails to eliminate the
pathogen, the acquired immunity is activated relying on the recognition of antigens by
antibodies formed after previous contact (Rainard and Riollet, 2006). The innate and
acquired immunity closely interact and are downregulated by anti-inflammatory
mechanisms after successful removal of the pathogen (Aitken et al., 2011).
2.2 Innate immunity
The innate immunity is composed of a sensing arm present in healthy tissue and an
effector arm activated after recognition of the infection (Beutler, 2004). Bovine
macrophages and mammary epithelial cells recognize pathogens by binding of pathogen-
associated molecular patterns on specific pattern recognition receptors (Rainard and Riollet,
2006). The toll-like receptors are a major group of pattern recognition receptors binding

Chapter 1 General introduction
12
lipoteichoic acid of Gram-positive bacteria, Lipopolysaccharide of Gram-negative bacteria or
other pathogen-associated molecular patterns (Kawai and Akira, 2005). Ten functional toll
like receptors have been described in cows (Menzies and Ingham, 2006). Following
recognition, pathways are activated resulting in the expression and secretion of cytokines
and oxylipids (Sordillo and Streicher, 2002). Release of cytokines including interleukin 1 (IL-
1), interleukin 8 (Il-8) and tumor necrosis factor-α (TNF- α) can cause migration of leukocytes
from blood to site of infection, inflammation, induction of fever and synthesis of acute phase
proteins (Bannerman, 2009; Rainard and Riollet, 2006).
Within a few hours, the SCC can exceed 106 cells/mL with neutrophils being the
predominant cell type in early inflammation (Bannerman et al., 2004b). Passage of
neutrophils from blood to the tissue, referred to as diapedesis, is mediated by adhesion
molecules. Binding of L-selectin (CD62L) to its ligands slows down neutrophils allowing them
to roll on the endothelium. Next, binding of β2 integrins including CD11b/CD18 (heterodimer
with cluster of differentiation 11b and 18) on molecules of the endothelial surface causes
firm attachment and helps neutrophils to leave the bloodstream (Paape et al., 2003).
Following diapedesis, neutrophils migrate to the highest concentration of chemoattractants,
a process called chemotaxis. Barber and Yang (1998) demonstrated IL-8 to play a major role
in the chemotactic activities of bovine mastitic milk. Additionaly, IL-8 was found to enhance
expression of adhesion molecules such as CD11b and CD18 by bovine neutrophils
(McClenahan et al., 2000; Rambeaud et al., 2005). Other chemoattractants such as
complement factor 5a (C5a) and bacterial compounds also affect migration of bovine
neutrophils (Persson et al., 1993).
At the site of infection, resident and recruited neutrophils combat pathogens by
phagocytosis, generation of reactive oxygen species (ROS), release of antibacterial peptides
and formation of neutrophil extracellular traps (Lippolis et al., 2006; Paape et al., 2003;
Selsted et al., 1993). Phagocytosis is a process in which the neutrophil engulfs a pathogen to
form an internal vesicle called phagosome. Reactive oxygen species are unstable oxygen
metabolites with a strong antibacterial activity but that also damage milk synthesizing
tissues (Aitken et al., 2011). Diapedesis induces apoptosis and reduces the ROS capacity
(Smits et al., 1999; Van Oostveldt et al., 2002) explaining the lower viability and activity of
resident milk neutrophils compared to blood neutrophils (Mehrzad et al., 2009). In the early

Chapter 1 General introduction
13
stage of IMI, however, resident and recruited neutrophils are activated by cytokines released
by macrophages, neutrophils itself and other immune cells (Rainard and Riollet, 2006).
The innate immune response following IMI is represented graphically in Figure 1.4.
Figure 1.4. Simplified representation of the innate immune response following bacterial entrance into the
bovine mammary gland highlighting the role of interleukin 8. After binding of pathogen-associated molecular
patterns on pattern recognition receptors, resident immune cells release cytokines such as interleukin 8 (IL-8),
interleukin 1 (IL-1) and tumor necrosis factor α (TNF α). The release of cytokines is followed by migration of
neutrophils from blood to tissue being mediated by adhesion molecules. Binding of L-selectin to its ligands
slows down neutrophils allowing them to roll on the endothelium. Next, binding of β2 integrins on molecules of
the endothelial surface causes firm attachment and helps neutrophils to leave the bloodstream. Subsequently,
neutrophils migrate to the highest concentration of chemoattractants, a process called chemotaxis. At the site
of infection, resident and recruited neutrophils combat pathogens by phagocytosis and generation of reactive
oxygen species (ROS). Interleukin 8 upregulates adhesion molecule expression, is an important
chemoattractant and enhances the activity and viability of neutrophils.
IL-8
ROS
Blood vessel
Endothelium
Mammaryepithelium
Milk
Bacteria
Resident immune cell
Patternrecognitionreceptor
NeutrophilIL-1
TNF-α
L-selectinβ2 integrins

Chapter 1 General introduction
14
2.3 Adaptive immunity
Antigen presenting cells and lymphocytes are two cell types involved in the adaptive
immunity of the bovine mammary gland. Lymphocytes are divided in two functionally
different subgroups: T-lymphocytes or T-cells and B-lymphocytes or B-cells. Following
phagocytosis, antigen presenting cells process bacteria to antigens and present those
antigens in combination with major histocompatibility complex class II (MHC II) molecules on
their membrane. Among other cells, macrophages act as antigen presenting cells in the
bovine mammary gland (Fitzpatrick et al., 1992; Politis et al., 1992). After binding of the
antigen-MHC II complex on the T cell receptor, T helper cells (CD4+ lymphocytes) proliferate
and differentiate to effector T helper cells, memory T helper cells or regulatory T helper cells.
The effector T helper cells secrete cytokines that stimulate the innate immunity and activate
B-cells. For example, interferon gamma secreted by lymphocytes enhances phagocytosis
capacity and ROS generation of neutrophils in milk (Sordillo and Babiuk, 2004). Activated B-
cells proliferate and differentiate to antibody-producing plasma cells or memory B-cells.
Binding of antibodies on pathogens facilitates phagocytosis of bovine neutrophils and
macrophages (opsonization) (Howard et al., 1980; Sordillo et al., 1997). Memory T-cells and
memory B-cells allow for a stronger and faster response after repeated exposure to the
same antigen. Nevertheless, because specific T- and B-cells need to proliferate in the lymph
nodes, mounting an adaptive immune response takes several days. Immunological memory
in the bovine mammary gland is rather limited as was concluded from the fact that milk yield
loss and severity in repeated pathogen-group specific clinical mastitis cases were very similar
compared to the initial case (Schukken et al., 2009b).
2.4 Pathogen-specific host response
The previous paragraphs described the general concepts of the bovine mammary gland
immunity. However, different mastitis pathogens elicit different host responses. In the
following paragraphs, host responses following IMI by E. coli, S. aureus, S. uberis, and
coagulase-negative staphylococci are discussed in more detail.
Following penetration of the teat canal, E. coli can rapidly multiply in the milk releasing
lipopolysaccharides (Schukken et al., 2011). The released lipopolysaccharides bind, in
conjunction with CD14, on toll like receptor 4 of resident immune cells resulting in a fast

Chapter 1 General introduction
15
release of cytokines and oxylipids. Within a day, intramammary E. coli challenge induces an
increase of milk IL-1, TNF-α, IL-8 and C5a. Additionally, high concentration of milk bovine
serum albumin indicates disruption of the blood-milk barrier (Bannerman et al., 2004b). This
pronounced host response often leads to acute clinical symptoms. Death due to shock and
multiple organ failure are not uncommon (Burvenich et al., 2003). Escherichia coli mastitis is
characterized by a sharp increase in SCC and decrease in milk yield (de Haas et al., 2004;
Grohn et al., 2004). In most cases, bacteria are rapidly eliminated by the host response and
SCC and milk yield quickly restore to pre-infection levels (de Haas et al., 2004; Grohn et al.,
2004; Lago et al., 2011). However, due to damage to the udder tissue, not all cows recover
their potential yield (Grohn et al., 2004).
Among other pathogen-associated molecular patterns, lipoteichoic acid of S. aureus
binds on toll like receptor 2 and 6 (Rainard and Riollet, 2006). However, intramammary S.
aureus challenge induces only a moderate host response (Bannerman et al., 2004b; Petzl et
al., 2008). An increase in milk TNF-α and IL-8 remains absent whereas milk C5a, IL-1 and
bovine serum albumin show a delayed increase compared to E. coli challenge (Bannerman et
al., 2004b). Although S. aureus can cause severe cases of clinical mastitis (Sol et al., 2000),
clinical symptoms are rather rare (Schukken et al., 2011). Staphylococcus aureus mastitis is
characterized by a gradual, long lasting increase in SCC and decrease in milk yield (de Haas et
al., 2004; Grohn et al., 2004; Lago et al., 2011). As previously mentioned, S. aureus can evade
the immune response by biofilm formation and intracellular survival (Bardiau et al., 2014;
Bradley and Green, 2009; Melchior et al., 2006). As a result, chronic S. aureus IMI with
intermitting shedding patterns are common (Sears et al., 1990).
Although S. uberis shares several pathogen-associated molecular patterns with S.
aureus, S. uberis does not activate toll like receptor 2 (Farhat et al., 2008). Compared to E.
coli and S. aureus, intramammary S. uberis challenge induces an intermediate host response.
Increases in milk TNF-α, IL-1, IL-8, C5a and bovine serum albumin occur but only 24 h after
inoculation (Bannerman et al., 2004a; Rambeaud et al., 2003). Streptococcus uberis can
cause both subclinical and clinical mastitis but certain strains were found to be more
pathogenic compared to others (Leigh et al., 2010; Tomita et al., 2008). Intramammary
infections with S. uberis can induce a peak in SCC or long lasting SCC elevation (de Haas et
al., 2004). Milk production losses due to streptococcal clinical mastitis are similar to those
incurred by S. aureus clinical mastitis (Schukken et al., 2011).

Chapter 1 General introduction
16
Although differences between species exist (Simojoki et al., 2011), coagulase-negative
staphylococci induce in general a weak host response (Supré et al., 2011). Intramammary
challenge with S. chromogenes, S. simulans or S. epidermidis causes mild to moderate clinical
mastitis cases but very high inoculation doses were required to do so (Simojoki et al., 2009;
Simojoki et al., 2011). Increases in milk TNF-α, IL-1 and IL-8 were observed (Simojoki et al.,
2011). Because of the large number of coagulase-negative Staphylococcus species and their
differences in ecology and epidemiology (De Visscher et al., 2014; Vanderhaeghen et al.,
2014), more research is required to further unravel the interactions between cows and
coagulase-negative staphylococci.
2.5 Factors affecting mammary gland immunity
Many factors affect mammary gland immunity (Sordillo, 2005). Nutrition has an
influence. Lactation has a much higher energy requirement compared to late gestation. In
case of malnutrition during the transition period, cows can suffer from too severe negative
energy balance in the periparturient period and early lactation. Cows in too severe negative
energy balance mobilize more body fat than the liver is able to metabolize resulting in an
increase of ketone bodies in the blood referred to as ketosis. Ketosis is associated with an
increased risk of postpartum clinical mastitis, reduced blood neutrophil ROS generation and
more severe symptoms following intramammary E. coli challenge (Kremer et al., 1993;
Oltenacu and Ekesbo, 1994; Zerbe et al., 2000). Because of their antioxidant role, adequate
levels of selenium and vitamin E are required in the diet. Deficiencies of other
micronutrients including copper, zinc and vitamin A can also affect mammary gland
immunity (O'Rourke, 2009).
Mammary gland immunity differs within the lifetime of a cow. Incidence rate of clinical
mastitis (IRCM) is high in early lactation, lower in mid to late lactation and the lowest during
the non-lactating period (Barkema et al., 1998; McDougall et al., 2007). Intramammary
infection might occur during the non-lactating period but rarely becomes apparent before
parturition. However, the susceptibility to IMI differs within the dry period. Resistance to
new IMI is the highest in the middle phase of the dry period (involuted phase) due to
presence of a functional keratin plug and high concentrations of protective factors such as
lactoferrin and immunoglobulins (Bradley and Green, 2004). In early lactation, mammary

Chapter 1 General introduction
17
gland immunity is not yet at full strength and suppressed by stress, negative energy balance
or a combination of both (Sordillo, 2005). At the cellular level, blood neutrophils from early
lactating cows show a higher spontaneously induced apoptosis and lower capacity to
generate ROS, chemotaxis, diapedesis, phagocytosis of E. coli and bactericidal activity
against S. aureus compared to blood neutrophils from mid lactating cows (Dosogne et al.,
2001; Mehrzad et al., 2001; Stevens et al., 2011; Van Oostveldt et al., 2001). Furthermore,
resident milk neutrophils of early lactating cows show more necrosis and lower capacity to
generate ROS and bactericidal activity against S. aureus compared to resident milk
neutrophils of mid lactating cows (Dosogne et al., 2001; Mehrzad et al., 2001; Van Oostveldt
et al., 2001). In vitro differences in neutrophil viability and ROS generation can be linked to in
vivo mammary gland immunity. Higher viability and ROS generation of resident milk
neutrophils are associated with higher ROS generation and lower bacterial count and milk
losses following intramammary E. coli challenge (Mehrzad et al., 2004; Mehrzad et al., 2005).
Not only lactation stage but also parity relates to mammary gland immunity. Except for
the first days after calving, heifers have a lower IRCM compared with multiparous cows
(Barkema et al., 1998; McDougall et al., 2007). Heifers have a stronger mammary gland
immunity compared to multiparous cows. Blood and resident milk neutrophils from heifers
show higher phagocytosis and killing of S. aureus and capacity to generate ROS. Additionally,
resident milk neutrophils of heifers are more viable compared to resident milk neutrophils of
multiparous cows (Mehrzad et al., 2002; Mehrzad et al., 2009).
Under similar management and within the same lactation stage and parity, certain
cows are more likely to suffer from mastitis compared to others (De Vliegher et al., 2004;
Piepers et al., 2011) and the immune response following IMI by E. coli or other pathogens
differs remarkably between cows (Bannerman et al., 2008; Burvenich et al., 2003; Simojoki
et al., 2011). Efforts have been made to identify cows as so-called low-, moderate- or high
responders using dermal fibroblast models (Green et al., 2011; Kandasamy et al., 2011) or by
measuring cutaneous delayed type hypersensitivity and antibody production in response to
antigen administration (Heriazon et al., 2009; Hernandez et al., 2005; Thompson-Crispi and
Mallard, 2012). Also mammary gland immunity shows a large variation at the cow level. For
example, milk neutrophil apoptosis differs little between herds but largely between cows
within herds (Piepers et al., 2009).

Chapter 1 General introduction
18
Mastitis resistance, defined as the cow’s ability to hinder pathogens from causing IMI,
is partially hereditary (Rupp and Boichard, 2003). Certain breeds and cows within breeds are
genetically more susceptible to mastitis (Detilleux, 2009; Ramirez et al., 2014). The genetics
of mastitis susceptibility/resistance is discussed in more detail in the third part of this
chapter.
3. Genetics of mastitis resistance
3.1 Breeding for mastitis resistance
As mastitis resistance is genetically determined, selective breeding can be used to
improve udder health. To increase mastitis resistance, cows with the highest response
against invading pathogens are favorable. However, as previously mentioned, strong host
responses can also damage the udder tissue (Aitken et al., 2011). Hence, a certain amount of
tolerance towards the invading pathogen can be beneficial. Mastitis tolerance is defined as
the cow’s ability to limit the damage following IMI. The breeding goal should be a cow with
efficient immune responses inflicting little damage to itself.
In traditional selection, estimated breeding values are calculated based on phenotypic
records of relatives. The reliability of this estimated breeding values, being defined as the
squared correlation between true and estimated breeding value, depends on the number of
relatives tested and the heritability of the trait. The heritability of a trait equals the
proportion of additive genetic variation over phenotypic variation between animals. It varies
from 0 to 1 and can be estimated by comparing the variation in related and unrelated
animals. In genomic selection, genomic estimated breeding values are calculated based on
the DNA of an animal. Using a DNA chip, the genotype of the animal is determined at
thousands of single nucleotide polymorphisms (SNPs), locations in the genome that differ
between animals. The genomic estimated breeding value is estimated by comparing the
animal’s genotype at all the SNPs with that of the genotype of animals with reliable
estimated breeding values (Meuwissen et al., 2001). The major benefit of genomic selection
is that reliable breeding values can be estimated at birth. In traditional selection, accurate
estimation of breeding values requires at least 5 years to have sufficient records of offspring
(Schaeffer, 2006). Research demonstrated that the reliability of genomic estimated breeding
values can be as high as the reliability of estimated breeding values (Hayes et al., 2009).

Chapter 1 General introduction
19
Both in traditional and genomic selection, accurately measurable traits are required.
To select against mastitis, clinical mastitis forms an obvious trait. Several studies reported
heritability estimates for clinical mastitis. Depending on the way clinical mastitis was
measured, the observation period during lactation and the statistical methods used, figures
ranged around 0.05 (Detilleux, 2009). The low heritability indicates that mainly non-genetic
factors such as management determine the occurrence of clinical mastitis. Yet, direct
selection against clinical mastitis was demonstrated to reduce IRCM substantially
(Heringstad et al., 2003). Bacteriological culture results form a second potential trait
(Ouweltjes et al., 2008). However, both clinical mastitis records and culture results are not
available on a large scale in most countries. For this reason, indirect measures are used to
calculate estimated breeding values for udder health. Because its high genetic correlation
with clinical mastitis, its easy accessibility through DHI programs and relative high
heritability, SCC forms an excellent trait (Detilleux, 2009). By combining different SCC traits
such as SCC in early or late lactation and peak patterns in SCC, estimated breeding values for
clinical and subclinical mastitis can be calculated with high reliability (Windig et al., 2010).
Important to mention is that selection on one trait can have adverse effects on
another trait. For example, a positive genetic correlation exists between milk yield and
clinical mastitis in first lactation meaning that heifers with high genetic merit for milk yield
are genetically more likely to have clinical mastitis (Koeck et al., 2014; Koivula et al., 2005).
Indeed, genetic selection merely on milk production was shown to threaten udder health
(Heringstad et al., 2003; Ouweltjes et al., 2007).
3.2 Identifying genes associated with mastitis resistance
Due to its complexity, many genes are involved in mammary gland immunity.
However, polymorphisms in genes encoding proteins with an important role in the immune
response might alter mastitis susceptibility of cows significantly (Detilleux, 2009).
Identification of polymorphisms associated with mastitis resistance offers more insight in the
mammary gland immunity and can lead to the discovery of new therapies (Pighetti and
Elliott, 2011).
During meiosis, limited recombination takes place between SNPs closely located to
each other on the same chromosome. As a consequence, a certain genotype at one SNP

Chapter 1 General introduction
20
often co-occurs with a certain genotype at a SNP located close by, a phenomenon called
linkage disequilibrium. Due to linkage disequilibrium, not only causal SNPs altering
functionality or expression of the protein but also SNPs in the same region of the
chromosome will be associated with a trait. Latter regions are called quantitative trait loci
(QTL) when referring to quantitative traits and can be identified by genotyping cows with a
DNA chip and testing associations between all markers and the trait. In the final step of a
genome wide association study, public databases are consulted to analyze which genes are
located in or close to the QTL.
When thousands of associations are studied, a number will be significant merely by
chance. In genome wide association studies, methods are used to correct for the multiple
comparisons. A downside of these correction methods is that small effects are not detected
and relatively large sample sizes are required. In a candidate gene approach, polymorphisms
in one or a limited number of genes with an important function in the mammary gland
immunity are studied (Detilleux, 2009). Due to the limited number of comparisons, effects
can be detected in smaller sample sizes and more detailed phenotypic traits can be studied.
Measuring gene expression in healthy and infected quarters or cell lines forms a third
way to identify genes associated with mastitis resistance, besides genome wide association
and candidate gene studies. DNA microarrays and RNA sequencing can be used to identify
differential expressed genes among a large number of genes whereas reverse transcription
quantitative real-time PCR (RT-qPCR) can be used to study the mRNA expression of a limited
number of genes in more detail (Bionaz and Loor, 2007).
3.3 Quantitative trait loci and important genes for mastitis resistance
Genome-wide association studies using SNP arrays, whole-genome sequences or a
combination of both identified multiple regions in the bovine genome to be important for
mastitis (Abdel-Shafy et al., 2014; Sahana et al., 2014; Sodeland et al., 2009; Strillacci et al.,
2014). On all chromosomes, QTL for mastitis traits have been detected (Figure 1.5). Genes
such as chemokine (C-X-C motif) receptor 1 (CXCR1), chemokine (C-X-C motif) receptor 2
(CXCR2), IL-8, neuropeptide FF receptor 2 (NPFFR2) and vitamin D-binding protein precursor
(GC) were located closely to or within the QTL (Sahana et al., 2014; Sodeland et al., 2009).

Chapter 1 General introduction
21
Strong linkage disequilibrium, however, complicates the determination of causal genes
(Sahana et al., 2014).
Figure 1.5. Genomic location of some of the quantitative trait loci (QTL) for mastitis submitted to the Cattle QTL
Database (Hu et al., 2013) indicating the genetic complexicity of mastitis resistance. SCC and SCS stand for
somatic cell count and somatic cell score, respectively.
Genes encoding proteins involved in the recognition of pathogens, recruitment of
immune cells towards site of infection, elimination of the pathogens and resolution of the
inflammatory response form interesting study objects (Pighetti and Elliott, 2011). Using a
candidate gene approach, BoLa-DRB3 (MHC, class II, DRB3), CXCR1, NOD2 (nucleotide-
binding oligomerization domain containing 2 alias CARD15), MBL (mannan-binding lectin),
SPP1 (secreted phosphoprotein 1 alias osteopontin) TLR1 (toll like receptor 1) and TLR4 (toll
like receptor 4) were reported as important genes for mastitis resistance (Alain et al., 2009;
Pant et al., 2007; Rupp and Biochard, 2003; Russell et al., 2012; Sharma et al., 2006; Wang et

Chapter 1 General introduction
22
al., 2011; Youngerman et al., 2004). The candidate gene CXCR1 is further discussed in the
fourth part of this chapter.
4. The CXCR1 gene
4.1 Function of CXCR1
Interleukin 8 has a pleiotropic effect on bovine neutrophils. As previously mentioned,
IL-8 is a strong chemoattractant (Caswell et al., 1999). Furthermore, IL-8 delays apoptosis
and enhances β2 integrin expression and activity of neutrophils (McClenahan et al., 2000;
Mitchell et al., 2003; Rambeaud et al., 2006). It exerts its function by binding on CXCR1 and
CXCR2 [chemokine (C-X-C motif) receptor 1 and 2], also named IL8RA and IL8RB (Lahouassa
et al., 2008). Both are G-protein coupled receptors characterized by an extracellular N-
terminus, 3 extracellular loops, 7 transmembrane regions, 3 intracellular loops and an
intracellular C-terminus (Park et al., 2012). Research on human neutrophils indicates that
enhancement of ROS is mediated through CXCR1 and not through CXCR2 (Stillie et al., 2009).
Latter finding has yet to be tested in bovine neutrophils. Additionally, ligand specificity
differs between CXCR1 and CXCR2. Experiments with cells transfected with either bovine
CXCR1 or bovine CXCR2 showed that both receptors bind CXCL8 (IL-8) with high affinity but
CXCR2 also binds other chemokines such as CXCL2 (growth-related oncogen-β) (Lahouassa et
al., 2008). Because of the important role in the mammary gland immunity, CXCR1 and CXCR2
form interesting candidate genes for mastitis resistance.
4.2 Genomic location
In the initial studies (Rambeaud and Pighetti, 2005; Rambeaud et al., 2006;
Youngerman et al., 2004a; Youngerman et al., 2004b), CXCR1 was wrongly annotated as
CXCR2 (Pighetti and Rambeaud, 2006). The two genes lay on the second chromosome (BTA
2) approximately 21 kb separated from each other and on opposite strands. In vitro IL-8 and
CXCL2 stimulation of cells transfected with either of the two receptors demonstrated the
more centromeric gene (GenBank Gene ID: 100125580) to encode for CXCR1 (GenBank
NP_001098508.1) and the more telomeric gene (GenBank Gene ID: 782719) to encode for
CXCR2 (GenBank NP_001094755.1) (Figure 1.6) (Lahouassa et al., 2008).

Chapter 1 General introduction
23
Figure 1.6. The relative genomic locations and structure of bovine CXCR1 (GenBank Gene ID: 100125580) and
CXCR2 (GenBank Gene ID: 782719) according to GenBank. The sequences coding for the proteins (coding
sequences) are indicated in black. The 5’ and 3’ untranslated regions, transcribed to mRNA but not translated
to protein, are colored gray. Introns, not transcribed to mRNA, are colored white.
The amino acid sequence of bovine CXCR1 shares 84% identity with bovine CXCR2.
Mainly the first 50 amino acids, forming the N-terminal region and determining the IL-8
receptor subtype (Gayle et al., 1993), show differences (18% homology) (Lahouassa et al.,
2008). Bovine CXCR1 shares 70, 72 and 62 % amino acid sequence identity with rabbit,
human and mouse CXCR1, respectively (Lahouassa et al., 2008).
4.3 The CXCR1 polymorphisms
For more than a decade, bovine CXCR1 polymorphisms have been studied (Table 1.2)
(Figure 1.7). Initially, SNPs were named based on their position relative to a published
sequence (GenBank Accession No. U19947) and not based on their position relative to the
start codon (Pighetti et al., 2012). Pighetti et al. (2012) described 36 SNPs in the bovine
CXCR1 gene of which 5 cause a change in the CXCR1 amino acid sequence (Figure 1.7). The
SNP at position 365 relative to the start codon (c.365T>C) causes a valine to alanine switch at
position 122 (p.Val122Ala) in the first extracellular loop of the protein potentially altering
initial ligand binding and strength. Latter SNP was found to be in full linkage disequilibrium
with c.735C>G causing an amino acid change in the third intracellular loop (p.His245Glu)
potentially affecting G-protein binding and signal transduction. Single nucleotide
polymorphisms c.980A>G and c.995A>G cause changes in the C-terminus (p.Lys327Arg and
p.His332Arg) and might interfere with adaptin-2 binding and receptor internalization.
Additionally, SNP c.621G>A could introduce an early stop codon (Pighetti et al., 2012). A SNP
5’
CXCR2
3’
3’
5’CXCR1

Chapter 1 General introduction
24
located 1606 base pairs upstream the start codon (c.-1606T>A) might affect binding of
transcription factors and expression of the receptor (Leyva-Baca et al., 2008b).
Figure 1.7. Visual representation of bovine CXCR1. The amino acid sequence and positions of the single
nucleotide polymorphisms (SNP) identified by Pighetti et al. (2012) in the coding DNA sequence are noted.
Amino acids with a neutral, cationic and anionic charge are colored blue, green and orange, respectively.
Amino acids encoded by a codon with a non-synonymous and synonymous SNP have a red and yellow outline,
respectively. The structure is based on publications on human CXCR1 (Baggiolini et al., 1994; Damaj et al.,
1996). A putative disulfide bridge is indicated in purple.
4.4 CXCR1 polymorphisms and neutrophil traits
In vitro migration to recombinant human IL-8 (rhIL-8) was found to be higher in blood
neutrophils from c.735GG cows compared to blood neutrophils from c.735CC and c.735CG
cows (Rambeaud and Pighetti, 2005). Incubation with rhIL-8 increased the expression of
CD18 and CD11b on the neutrophils. However, c.735GG neutrophils showed a higher
upregulation of CD18 expression and tended to show a higher upregulation of CD11b
compared to c.735CC neutrophils. Besides an impaired migration, blood neutrophils from
c.735CC cows showed a lower PMA (phorbol 12-myristate 13-acetate) induced ROS

Chapter 1 General introduction
25
generation compared to blood neutrophils from c.735GG cows. Yet, rhIL-8 induced decrease
of apoptosis was higher in c.735CC blood neutrophils compared to c.735GG blood
neutrophils (Rambeaud et al., 2006). A radioligand binding assay demonstrated a tendency
towards higher binding capacity of IL-8 in c.735GG compared to c.735CC blood neutrophils.
Whether this was caused by higher affinity, higher expression of IL-8 receptors or a
combination of both was not clarified. Intracellular calcium release following IL-8 stimulation
was found to be higher in c.735GG compared to c.735CC neutrophils (Rambeaud and
Pighetti, 2007).
The abovementioned studies focused on in vitro associations between c.735C>G and
blood neutrophils of mid-lactating cows. Because neutrophil functionality and viability is
reduced in resident milk compared to blood neutrophils and in early compared to mid
lactation, results require confirmation in (milk) neutrophils of early lactating cows (Dosogne
et al., 2001; Mehrzad et al., 2001; Stevens et al., 2011; Van Oostveldt et al., 2001).
Additionally, associations with other CXCR1 SNP than c.735C>G yet have to be studied.
4.5 CXCR1 polymorphisms and mastitis
Youngerman et al. (2004) performed a cohort study in which 37 Holstein and 42 Jersey
cows were monitored for subclinical masitis by bacteriological culture of milk samples at
regular time intervals. In the Holstein population, c.735CC cows had a higher incidence of
chronic IMI compared to c.735CG and c.735GG cows but also had a lower SCC compared to
c.735CG cows. A complex relationship between CXCR1 polymorphism and SCC was
proposed: either, cows with genotypes associated with higher functionality of CXCR1
respond more efficiently against invading pathogens leading to less chronic IMI and lower
SCC or, these animals have higher SCC because of the higher response to IL-8 causing more
cells to migrate from blood to milk (Youngerman et al., 2004a). In an Irish study, c.735GG
cows tended to have a lower SCC compared to c.735CC and c.735CG cows but the
association could not be confirmed in a larger sire population (Beecher et al., 2010).
Similarly, c.735C>G was not associated with SCC in a Canadian population nor with
estimated breeding value for SCC in a Canadian and a German population (Galvao et al.,
2011; Goertz et al., 2009; Leyva-Baca et al., 2008b). Leyva-baca et al. (2008b) reported a
higher estimated breeding value for SCS in c.-1606AA sires compared to c.-1606TT sires,

Chapter 1 General introduction
26
although, this association could not be confirmed in a more recent study (Goertz et al.,
2009).
Polymorphisms located near CXCR1 and CXCR2 are highly associated with likelihood of
veterinary reported clinical mastitis (Sodeland et al., 2011). This association was confirmed
in a candidate gene study reporting a higher IRCM in c.735GG cows compared to c.735CC
and c.735CG cows (Galvao et al., 2011). Several research groups studied associations
between c.735C>G and milk yield. Two studies reported a lower milk yield in c.735GG cows
compared to c.735CG cows (Galvao et al., 2011; Youngerman et al., 2004a). However,
daughters of c.735GG sires have higher fat yields compared to daughters of c.735CG sires
(Beecher et al., 2010).
No association studies have been published on the non-synonymous SNP c.980A>G
and c.995A>G. Although immune responses strongly depend on the invading mastitis
pathogen (Bannerman, 2009; Schukken et al., 2011; Wellnitz and Bruckmaier, 2012),
pathogen(-group) specific associations between CXCR1 polymorphisms and mastitis
resistance have not yet been identified.
4.6 CXCR1 polymorphisms and gene expression
Up to date, two research groups published associations between CXCR1 polymorphism
and gene expression using reverse transcription quantitative real-time PCR (RT-qPCR).
Twenty-four hours after experimental infection with Streptococcus dysgalactiae, gene
expression of IL-8 and CXCR1 was found to be higher in milk somatic cells of c.735GG cows
compared to c.735CC cows (Beecher et al., 2012). Additionally, CXCR1 gene expression was
higher in c.-1606AA blood neutrophils compared to c.-1606TT blood neutrophils, both
before and after lipopolysaccharide challenge (Leyva-Baca et al., 2008a). Sufficient
experimental detail is required to evaluate and reproduce qPCR data (Bustin et al., 2009).
However, both articles lack information such as assessment of gDNA contamination,
validation of reference genes and qPCR validation (e.g. PCR efficiency). Although
normalization to a single reference gene can cause relative large errors (Vandesompele et
al., 2002), only one reference gene is mentioned. Technical articles on RT-qPCR are available
for bovine blood neutrophils but not for milk somatic cells of infected quarters (De Ketelaere
et al., 2006).

Chap
ter 1
Gene
ral i
ntro
duct
ion
27
Tabl
e 1.
2. A
ssoc
iatio
ns b
etw
een
singl
e nu
cleo
tide
poly
mor
phism
CXC
R1c.
735C
>G a
nd b
ovin
e ne
utro
phil
trai
ts a
nd m
astit
is
Sam
ple
Ph
enot
ype
Ge
noty
pe a
t c.7
35C>
G1
Refe
renc
e
CC
CG
GG
Bloo
d ne
utro
phils
of 3
0 co
ws
M
igra
tion
to IL
-82
-
- +
Ra
mbe
aud
and
Pigh
etti,
200
5
IL-8
indu
ced
upre
gula
tion
of C
D11b
exp
ress
ion3
-
-/+
+
IL
-8 in
duce
d up
regu
latio
n of
CD1
8 ex
pres
sion
-
-/+
+
Bloo
d ne
utro
phils
of 6
0 co
ws
IL
-8 in
duce
d de
crea
se in
apo
ptos
is
+
-/+
-
Ram
beau
d et
al.,
200
6
PMA4 in
duce
d RO
S5 gen
erat
ion
-
-/+
+
Bloo
d ne
utro
phils
of 1
0 co
ws
IL
-8 b
indi
ng c
apac
ity3
-
+
Ra
mbe
aud
and
Pigh
etti,
200
7 Bl
ood
neut
roph
ils o
f 36
cow
s
IL-8
indu
ced
intr
acel
lula
r cal
cium
rele
ase
-
+
Lact
atio
ns o
f 37
cow
s
Subc
linic
al m
astit
is in
cide
nce
+
- -
Yo
unge
rman
et a
l., 2
004a
Estim
ated
SCS
- +
-
Es
timat
ed 3
05-d
milk
yie
ld
+
+ -
Lact
atio
ns o
f 246
cow
s
Test
day
SCS
3
+ +
-
Beec
her e
t al.,
201
0 O
ffspr
ing
of 8
48 si
res
30
5-d
milk
fat y
ield
-
+
Lact
atio
ns o
f 350
cow
s
Inci
denc
e ra
te o
f clin
ical
mas
titis
- -
+
Galv
ao e
t al.,
201
1
Test
day
milk
yie
ld
-/
+ +
-
Milk
som
atic
cel
ls of
9 c
ows
Gene
ex
pres
sion
of
IL-8
an
d CX
CR1
follo
win
g St
rept
ococ
cus d
ysga
lact
iae
chal
leng
e
-
+
Beec
her e
t al.,
201
2 1 Ge
noty
pe g
roup
s with
a +
had
a si
gnifi
cant
hig
her p
heno
type
com
pare
d to
the
geno
type
gro
up w
ith a
-. G
enot
ype
grou
ps w
ith -/
+ sh
owed
an
inte
rmed
iate
phe
noty
pe.
2 Inte
rleuk
in 8
. 3 Di
ffere
nces
tend
ed to
be
signi
fican
t.
4 Phor
bol 1
2-m
yrist
ate
13-a
ceta
te.

Chap
ter 1
Gene
ral i
ntro
duct
ion
28
Figu
re 1
.8. F
rom
gen
otyp
e to
phe
noty
pe: c
urre
nt k
now
ledg
e on
CXC
R1 a
nd b
ovin
e ne
utro
phil
trai
ts a
nd m
astit
is. 1 Pi
ghet
ti et
al.
(201
2); 2 Ra
mbe
aud
and
Pigh
etti
(200
7);
3 Ram
beau
d et
al.
(200
5); 4 Ra
mbe
aud
and
Pigh
etti
(200
6); 5 Yo
unge
rman
et a
l. (2
004a
); 6 G
alva
o et
al.
(201
1); 7 Be
eche
r et a
l. (2
012)

Chapter 1 General introduction
29
5. References
Abdel-Shafy, H., R. H. Bortfeldt, J. Tetens, and G. A. Brockmann GA. 2014. Single nucleotide
polymorphism and haplotype effects associated with somatic cell score in German
Holstein cattle. Genet. Sel. Evol. 46:35.
Aitken, S. L., C. M. Corl, and L. M. Sordillo. 2011. Immunopathology of mastitis: insights into
disease recognition and resolution. J. Mammary Gland Biol. Neoplasia 16:291-304.
Alain K., N. A. Karrow, C. Thibault, J. St-Pierre, M. Lessard, and N. Bissonnette. 2009.
Osteopontin: an early innate immune marker of Escherichia coli mastitis harbors genetic
polymorphisms with possible links with resistance to mastitis. BMC Genomics 10:444.
Baggiolini, M., B. Dewald, and B. Moser. 1994. Interleukin-8 and related chemotactic
cytokines - CXC and CC chemokines. Adv. Immunol. 55:97-179.
Bannerman, D. D. 2009. Pathogen-dependent induction of cytokines and other soluble
inflammatory mediators during intramammary infection of dairy cows. J. Anim. Sci.
87:10-25.
Bannerman, D. D., M. J. Paape, J. P. Goff, K. Kimura, J. D. Lippolis, and J. C. Hope. 2004a.
Innate immune response to intramammary infection with Serratia marcescens and
Streptococcus uberis. Vet. Res. 35:681-700.
Bannerman, D. D., M. J. Paape, J. W. Lee, X. Zhao, J. C. Hope, and P. Rainard. 2004b.
Escherichia coli and Staphylococcus aureus elicit differential innate immune responses
following intramammary infection. Clin. Diagn. Lab. Immunol. 11:463-472.
Bannerman, D. D., A. C. Kauf, M. J. Paape, H. R. Springer, and J. P. Goff. 2008. Comparison of
Holstein and Jersey innate immune responses to Escherichia coli intramammary
infection. J. Dairy Sci. 91:2225-2235.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab. Immunol. 5:82-86.
Bardiau, M., J. Detilleux, F. Farnir, J. G. Mainil, and I. Ote. 2014. Associations between
properties linked with persistence in a collection of Staphylococcus aureus isolates from
bovine mastitis. Vet. Microbiol. 169:74-79.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, H. Wilmink, G. Benedictus, and A.
Brand. 1998. Incidence of clinical mastitis in dairy herds grouped in three categories by
bulk milk somatic cell counts. J Dairy Sci 81:411-419.

Chapter 1 General introduction
30
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, G. Benedictus, and A. Brand. 1999.
Management practices associated with the incidence rate of clinical mastitis. J. Dairy Sci.
82:1643-1654.
Barkema, H. W., M. J. Green, A. J. Bradley, and R. N. Zadoks. 2009. Invited review: The role of
contagious disease in udder health. J. Dairy Sci. 92:4717-4729.
Barnouin, J., S. Bord, S. Bazin, and M. Chassagne. 2005. Dairy management practices
associated with incidence rate of clinical mastitis in low somatic cell score herds in
France. J. Dairy Sci. 88:3700-3709.
Beaudeau, F., V. Ducrocq, C. Fourichon, and H. Seegers. 1995. Effect of disease on length of
productive life of French Holstein dairy cows assessed by survival analysis. J. Dairy Sci.
78:103-117.
Beecher, C., M. Daly, S. Childs, D. P. Berry, D. A. Magee, T. V. McCarthy, and L. Giblin. 2010.
Polymorphisms in bovine immune genes and their associations with somatic cell count
and milk production in dairy cattle. BMC Genet. 11:99.
Beecher, C., M. Daly, R. P. Ross, J. Flynn, T. V. McCarthy, and L. Giblin. 2012. Characterization
of the bovine innate immune response in milk somatic cells following intramammary
infection with Streptococcus dysgalactiae subspecies dysgalactiae. J. Dairy Sci. 95:5720-
5729.
Beutler, B. 2004. Innate immunity: an overview. Mol. Immunol. 40:845-859.
Bionaz, M., and J. J. Loor. 2007. Identification of reference genes for quantitative real-time
PCR in the bovine mammary gland during the lactation cycle. Physiol. Genomics 29:312-
319.
Bradley, A. J. 2002. Bovine mastitis: an evolving disease. Vet. J. 164:116-128.
Bradley, A. J., and M. J. Green. 2004. The importance of the nonlactating period in the
epidemiology of intramammary infection and strategies for prevention. Vet. Clin. North
Am.-Food Anim. Pract. 20:547-568.
Bradley, A. J., K. A. Leach, J. E. Breen, L. E. Green, and M. J. Green. 2007. Survey of the
incidence and aetiology of mastitis on dairy farms in England and Wales. Vet. Rec.
160:253-258.
Bradley, A. J., and M. J. Green. 2009. Factors affecting cure when treating bovine clinical
mastitis with cephalosporin-based intramammary preparations. J. Dairy Sci. 92:1941-
1953.

Chapter 1 General introduction
31
Bramley, A. J., and F. H. Dodd. 1984. Reviews of the progress of dairy science - mastitis
control - progress and prospects. J. Dairy Res. 51:481-512.
Burvenich, C., V. Van Merris, J. Mehrzad, A. Diez-Fraile, and L. Duchateau. 2003. Severity of
E. coli mastitis is mainly determined by cow factors. Vet. Res. 34:521-564.
Bustin, S. A., V. Benes, J. A. Garson, J. Hellemans, J. Huggett, M. Kubista, R. Mueller, T. Nolan,
M. W. Pfaffl, G. L. Shipley, J. Vandesompele, and C. T. Wittwer. 2009. The MIQE
guidelines: minimum information for publication of quantitative real-time PCR
experiments. Clin. Chem. 55:611-622.
Caswell, J. L., D. M. Middleton, and J. R. Gordon. 1999. Production and functional
characterization of recombinant bovine interleukin-8 as a specific neutrophil activator
and chemoattractant. Vet. Immunol. Immunopathol. 67:327-340.
Damaj, B. B., S. R. McColl, K. Neote, N. Songqing, K. T. Ogborn, C. A. Hebert, and P. H.
Naccache. 1996. Identification of G-protein binding sites of the human interleukin-8
receptors by functional mapping of the intracellular loops. FASEB J. 10:1426-1434.
de Haas, Y., R. F. Veerkamp, H. W. Barkema, Y. T. Grohn, and Y. H. Schukken. 2004.
Associations between pathogen-specific cases of clinical mastitis and somatic cell count
patterns. J. Dairy Sci. 87:95-105.
De Ketelaere, A., K. Goossens, L. Peelman, and C. Burvenich. 2006. Validation of internal
control genes for gene expression analysis in bovine polymorphonuclear leukocytes. J.
Dairy Sci. 89:4066-4069.
Delepine, S. 1910. Contribution to the study of the influences determining the prevalence of
bovine tuberculous mastitis. Proc. R. Soc. Med. 3 (Sect. Epidemiol. State Med.):217-256.
de Mol, R. M., and W. Ouweltjes. 2001. Detection model for mastitis in cows milked in an
automatic milking system. Prev. Vet. Med. 49:71-82.
Detilleux, J. C. 2009. Genetic factors affecting susceptibility to udder pathogens. Vet
Microbiol. 134:157-164.
De Visscher, A., K. Supré, F. Haesebrouck, R. N. Zadoks, V. Piessens, E. Van Coilli, S. Piepers,
and S. De Vliegher. 2014. Further evidence for the existence of environmental and host-
associated species of coagulase-negative staphylococci in dairy cattle. Vet. Microbiol.
172:466-474.

Chapter 1 General introduction
32
De Vliegher, S., H. Laevens, G. Opsomer, E. De Muelenaere, and A. de Kruif. 2001. Somatic
cell counts in dairy heifers during early lactation. Vlaams Diergeneeskundig Tijdschrift
70:212-215.
De Vliegher, S., H. Laevens, H. W. Barkema, I. R. Dohoo, H. Stryhn, G. Opsomer, and A. de
Kruif. 2004. Management practices and heifer characteristics associated with early
lactation somatic cell count of Belgian dairy heifers. J. Dairy Sci. 87:937-947.
De Vliegher, S., L. K. Fox, S. Piepers, S. McDougall, and H. W. Barkema. 2012. Invited review:
Mastitis in dairy heifers: nature of the disease, potential impact, prevention, and control.
J. Dairy Sci. 95:1025-1040.
Diamond, J. 2002. Evolution, consequences and future of plant and animal domestication.
Nature 418:700-707.
Dingwell, R. T., K. E. Leslie, Y. H. Schukken, J. M. Sargeant, L. L. Timms, T. F. Duffield, G. P.
Keefe, D. F. Kelton, K. D. Lissemore, and J. Conklin. 2004. Association of cow and quarter-
level factors at drying-off with new intramammary infections during the dry period. Prev.
Vet. Med. 63:75-89.
Dopfer, D., H. W. Barkema, T. J. G. M. Lam, Y. H. Schukken, and W. Gaastra. 1999. Recurrent
clinical mastitis caused by Escherichia coli in dairy cows. J. Dairy Sci. 82:80-85.
Dosogne, H., F. Vangroenweghe, B. Barrio, P. Rainard, and C. Burvenich. 2001. Decreased
number and bactericidal activity against Staphylococcus aureus of the resident cells in
milk of dairy cows during early lactation. J. Dairy Res. 68:539-549.
Ellis, T. N., and B. L. Beaman. 2004. Interferon-gamma activation of polymorphonuclear
neutrophil function. Immunology 112:2-12.
Farhat, K., K. S. Sauter, M. Brcic, J. Frey, A. J. Ulmer, and T. W. Jungi. 2008. The response of
HEK293 cells transfected with bovine TLR2 to established pathogen-associated molecular
patterns and to bacteria causing mastitis in cattle. Vet. Immunol. Immunopathol.
125:326-336.
Fitzpatrick, J. L., P. J. Cripps, A. W. Hill, P. W. Bland, and C. R. Stokes. 1992. MHC class-II
expression in the bovine mammary gland. Vet. Immunol. Immunopathol. 32:13-23.
Fox, L. K. 2009. Prevalence, incidence and risk factors of heifer mastitis. Vet. Microbiol.
134:82-88.

Chapter 1 General introduction
33
Galvao, K. N., G. M. Pighetti, S. H. Cheong, D. V. Nydam, and R. O. Gilbert. 2011. Association
between interleukin-8 receptor-alpha (CXCR1) polymorphism and disease incidence,
production, reproduction, and survival in Holstein cows. J Dairy Sci. 94:2083-2091.
Gayle, R. B., P. R. Sleath, S. Srinivason, C. W. Birks, K. S. Weerawarna, D. P. Cerretti, C. J.
Kozlosky, N. Nelson, T. Vanden Bos, and M. P. Beckmann. 1993. Importance of the amino
terminus of the interleukin-8 receptor in ligand interactions. J. Biol. Chem. 268, 7283–
7289.
Goertz, I., C. Baes, C. Weimann, N. Reinsch, and G. Erhardt. 2009. Association between single
nucleotide polymorphisms in the CXCR1 gene and somatic cell score in Holstein dairy
cattle. J. Dairy Sci. 92:4018-4022.
Grave, K., C. Greko, L. Nilsson, K. Odensvik, T. Mørk, and M. Rønning. 1999. The usage of
veterinary antibacterial drugs for mastitis in cattle in Norway and Sweden during 1990-
1997. Prev. Vet. Med. 42:45-55.
Green, B. B., S. Kandasamy, T. H. Elsasser, and D. E. Kerr. 2011. The use of dermal fibroblasts
as a predictive tool of the toll-like receptor 4 response pathway and its development in
Holstein heifers. J. Dairy Sci. 94:5502-5514.
Grohn, Y. T., D. J. Wilson, R. N. Gonzalez, J. A. Hertl, H. Schulte, G. Bennett, and Y. H.
Schukken. 2004. Effect of pathogen-specific clinical mastitis on milk yield in dairy cows. J.
Dairy Sci. 87:3358-3374.
Hansen, L. B. 2000. Consequences of selection for milk yield from a geneticist's viewpoint. J.
Dairy Sci. 83:1145-1150.
Hayes, B. J., P. J. Bowman, A. J. Chamberlain, and M. E. Goddard. 2009. Invited review:
Genomic selection in dairy cattle: Progress and challenges. J. Dairy Sci. 92:433-443.
Heriazon, A., J. A. Yager, W. Sears, and B. A. Mallard. 2009. Induction of delayed-type
hypersensitivity and interferon-gamma to Candida albicans and anti-hen-egg white
lysozyme antibody as phenotypic markers of enhanced bovine immune response. Vet.
Immunol. Immunopathol. 129:93-100.
Heringstad, B., G. Klemetsdal, and T. Steine. 2003. Selection responses for clinical mastitis
and protein yield in two Norwegian dairy cattle selection experiments. J. Dairy Sci.
86:2990-2999.

Chapter 1 General introduction
34
Hernandez, A., J. A. Yager, B. N. Wilkie, K. E. Leslie, and B. A. Mallard. 2005. Evaluation of
bovine cutaneous delayed-type hypersensitivity (DTH) to various test antigens and a
mitogen using several adjuvants. Vet. Immunol. Immunopathol. 104:45-58.
Hogan, J., and S. K. Larry. 2003. Coliform mastitis. Vet. Res. 34:507-519.
Hogan, J. S., J. W. Pankey, and A. H. Duthie. 1987. Growth-inhibition of mastitis pathogens by
long-chain fatty-acids. J. Dairy Sci. 70:927-934.
Howard, C. J., G. Taylor, and J. Brownlie. 1980. Surface-receptors for immunoglobulin on
bovine polymorphonuclear neutrophils and macrophages. Res. Vet. Sci. 29:128-130.
Hu, Z. L., C. A. Park, X. L. Wu, and J. M. Reecy. 2013. Animal QTLdb: an improved database
tool for livestock animal QTL/association data dissemination in the post-genome era.
Nucleic Acids Res. 41:D871-879.
Kandasamy, S., B. B. Green, A. L. Benjamin, and D. E. Kerr. 2011. Between-cow variation in
dermal fibroblast response to lipopolysaccharide reflected in resolution of inflammation
during Escherichia coli mastitis. J. Dairy Sci. 94:5963-5975.
Kawai, T., and S. Akira. 2005. Pathogen recognition with Toll-like receptors. Curr. Opin.
Immunol. 17:338-344.
Keane, O. M., K. E. Budd, J. Flynn, and F. Mccoy. 2013. Pathogen profile of clinical mastitis in
Irish milk-recording herds reveals a complex aetiology. Vet. Rec. 173:17.
Keefe, G. P. 1997. Streptococcus agalactiae mastitis: a review. Can. Vet. J. 38:429-437.
Kemp, M. H., A. M. Nolan, P. J. Cripps, and J. L. Fitzpatrick. 2008. Animal-based
measurements of the severity of mastitis in dairy cows. Vet. Rec. 163:175-179.
Koeck, A., S. Loker, F. Miglior, D. F. Kelton, J. Jamrozik, and F. S. Schenkel. 2014. Genetic
relationships of clinical mastitis, cystic ovaries, and lameness with milk yield and somatic
cell score in first-lactation Canadian Holsteins. J. Dairy Sci. 97:5806-5813.
Koivula, M., E. A. Mantysaari, E. Negussie, and T. Serenius. 2005. Genetic and phenotypic
relationships among milk yield and somatic cell count before and after clinical mastitis. J.
Dairy Sci. 88:827-833.
Kremer, W. D. J., E. N. Noordhuizenstassen, F. J. Grommers, Y. H. Schukken, R. Heeringa, A.
Brand, and C. Burvenich. 1993. Severity of experimental Escherichia coli mastitis in
ketonemic and nonketonemic dairy cows. J. Dairy Sci. 76:3428-3436.
Lago, A., S. M. Godden, R. Bey, P. L. Ruegg, and K. Leslie. 2011. The selective treatment of
clinical mastitis based on on-farm culture results: I. Effects on antibiotic use, milk

Chapter 1 General introduction
35
withholding time, and short-term clinical and bacteriological outcomes. J. Dairy Sci.
94:4441-4456.
Lahouassa, H., P. Rainard, A. Caraty, and C. Riollet. 2008. Identification and characterization
of a new interleukin-8 receptor in bovine species. Mol. Immunol. 45:1153-1164.
Lam, T. J. G. M., B. H. P. van den Borne, J. Jansen, K. Huijps, J. C. L. van Veersen, G. van
Schaik, and H. Hogeveen. 2013. Improving bovine udder health: A national mastitis
control program in the Netherlands. J. Dairy Sci. 96:1301-1311.
LeBlanc, S. J., K. D. Lissemore, D. F. Kelton, T. F. Duffield, and K. E. Leslie. 2006. Major
advances in disease prevention in dairy cattle. J. Dairy Sci. 89:1267-1279.
Leigh, J. A., S. A. Egan, P. N. Ward, T. R. Field, and T. J. Coffey. 2010. Sortase anchored
proteins of Streptococcus uberis play major roles in the pathogenesis of bovine mastitis
in dairy cattle. Vet. Res. 41:63.
Leyva-Baca, I., G. Pighetti, and N. A. Karrow. 2008a. Genotype-specific IL8RA gene expression
in bovine neutrophils in response to Escherichia coli lipopolysaccharide challenge. Anim.
Genet. 39:298-300.
Leyva-Baca, I., F. Schenkel, J. Martin, and N. A. Karrow. 2008b. Polymorphisms in the 5 '
upstream region of the CXCR1 chemokine receptor gene, and their association with
somatic cell score in Holstein cattle in Canada. J. Dairy Sci. 91:407-417.
Lippolis, J. D., T. A. Reinhardt, J. P. Goff, and R. L. Horst. 2006. Neutrophil extracellular trap
formation by bovine neutrophils is not inhibited by milk. Vet. Immunol. Immunopathol.
113:248-255.
Ma, Y., C. Ryan, D. M. Barbano, D. M. Galton, M. A. Rudan, and K. J. Boor. 2000. Effects of
somatic cell count on quality and shelf-life of pasteurized fluid milk. J. Dairy Sci. 83:264-
274.
Matthews, K. R., R. A. Almeida, and S. P. Oliver. 1994. Bovine mammary epithelial cell
invasion by Streptococcus uberis. Infect. Immun. 62:5641-5646.
McClenahan, D., J. Fagliari, O. Evanson, and D. Weiss. 2000. Role of inflammatory mediators
in priming, activation, and deformability of bovine neutrophils. Am. J. Vet. Res. 61:492-
498.
McDougall, S., K. E. Agnew, R. Cursons, X. X. Hou, and C. R. Compton. 2007. Parenteral
treatment of clinical mastitis with tylosin base or penethamate hydriodide in dairy cattle.
J. Dairy Sci. 90:779-789.

Chapter 1 General introduction
36
Medrano-Galarza, C., J. Gibbons, S. Wagner, A. M. de Passillé, and J. Rushen. 2012.
Behavioral changes in dairy cows with mastitis. J Dairy Sci. 95:6994-7002.
Mehrzad, J., H. Dosogne, E. Meyer, R. Heyneman, and C. Burvenich. 2001. Respiratory burst
activity of blood and milk neutrophils in dairy cows during different stages of lactation. J.
Dairy Res. 68:399-415.
Mehrzad, J., L. Duchateau, S. Pyorala, and C. Burvenich. 2002. Blood and milk neutrophil
chemiluminescence and viability in primiparous and pluriparous dairy cows during late
pregnancy, around parturition and early lactation. J. Dairy Sci. 85:3268-3276.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2004. Viability of milk neutrophils and severity
of bovine coliform mastitis. J. Dairy Sci. 87:4150-4162.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2005. High milk neutrophil chemiluminescence
limits the severity of bovine coliform mastitis. Vet. Res. 36:101-116.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2009. Phagocytic and bactericidal activity of
blood and milk-resident neutrophils against Staphylococcus aureus in primiparous and
multiparous cows during early lactation. Vet. Microbiol. 134:106-112.
Melchior, M. B., H. Vaarkamp, and J. Fink-Gremmels. 2006. Biofilms: A role in recurrent
mastitis infections? Vet.. J. 171:398-407.
Menzies, M., and A. Ingham, 2006. Identification and expression of Toll-like receptors 1-10 in
selected bovine and ovine tissues. Vet. Immunol. Immunopathol., 109: 23-30.
Meuwissen, T. H. E., B. J. Hayes, and M. E. Goddard. 2001. Prediction of total genetic value
using genome-wide dense marker maps. Genetics 157:1819-1829.
Milk Control Centre Flanders. 2012. Annual Report. Milk Control Centre Flanders, Lier,
Belgium.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
National Mastitis Council. 2014. Recommended mastitis control program. NMC INC.,
Madison, WI. http://www. nmconline. org/docs/NMCchecklistINT. pdf.
Neave, F. K., F. H. Dodd, R. G. Kingwill, and D. R. Westgarth. 1969. Control of mastitis in the
dairy herd by hygiene and management. J. Dairy Sci. 52:696-707.
Neijenhuis, F., H. W. Barkema, H. Hogeveen, and J. P. Noordhuizen. 2001. Relationship
between teat-end callosity and occurrence of clinical mastitis. J. Dairy Sci. 84:2664-2672.

Chapter 1 General introduction
37
Oftedal, O. T. 2012. The evolution of milk secretion and its ancient origins. Animal 6:355-368.
Ogorevc, J., T. Kunej, A. Razpet, and P. Dovc. 2009. Database of cattle candidate genes and
genetic markers for milk production and mastitis. Anim. Genet. 40:832-851.
Olde Riekerink, R. G. M., H. W. Barkema, D. F. Kelton, and D. T. Scholl. 2008. Incidence rate
of clinical mastitis on Canadian dairy farms. J. Dairy Sci. 91:1366-1377.
Oliveira, L., C. Hulland, and P. L. Ruegg. 2013. Characterization of clinical mastitis occurring in
cows on 50 large dairy herds in Wisconsin. J. Dairy Sci. 96:7538-7549.
Oltenacu, P. A., and I. Ekesbo. 1994. Epidemiologic study of clinical mastitis in dairy cattle.
Vet. Res. 25:208-212.
O'Rourke, D. 2009. Nutrition and udder health in dairy cows: a review. Irish Vet. J. 62:15-20.
Ouweltjes, W., B. Beerda, J. J. Windig, M. P. L. Calus, and R. F. Veerkamp. 2007. Effects of
management and genetics on udder health and milk composition in dairy cows. J. Dairy
Sci. 90:229-238.
Ouweltjes, W., J. J. Windig, G. de Jong, T. J. G. M. Lam, J. ten Napel, and Y. de Haas. 2008.
The use of data from sampling for bacteriology for genetic selection against clinical
mastitis. J. Dairy Sci. 91:4860-4870.
Paape, M. J., D. D. Bannerman, X. Zhao, and J. W. Lee. 2003. The bovine neutrophil:
Structure and function in blood and milk. Vet. Res. 34:597-627.
Pant, S.D., F. S. Schenkel, I. Leyva-Baca, B. S. Sharma , and N. A. 2007. Identification of single
nucleotide polymorphisms in bovine CARD15 and their associations with health and
production traits in Canadian Holsteins. BMC Genomics 8:421.
Park, S. H., B. B. Das, F. Casagrande, Y. Tian, H. J. Nothnagel, M. N. Chu, H. Kiefer, K. Maier, A.
A. De Angelis, F. M. Marassi, and S. J. Opella. 2012. Structure of the chemokine receptor
CXCR1 in phospholipid bilayers. Nature 491:779-783.
Persson, K., I. Larsson, and C. H. Sandgren. 1993. Effects of certain inflammatory mediators
on bovine neutrophil migration in vivo and in vitro. Vet. Immunol. Immunopathol. 37:99-
112.
Petzl, W., H. Zerbe, J. Gunther, W. Yang, H. M. Seyfert, G. Nurnberg, and H. J. Schuberth.
2008. Escherichia coli, but not Staphylococcus aureus triggers an early increased
expression of factors contributing to the innate immune defense in the udder of the cow.
Vet. Res. 39:18.

Chapter 1 General introduction
38
Piepers, S., L. De Meulemeester, A. de Kruif, G. Opsomer, H. W. Barkema, and S. De Vliegher.
2007. Prevalence and distribution of mastitis pathogens in subclinically infected dairy
cows in Flanders, Belgium. J. Dairy Res. 74:478-483.
Piepers, S., G. Opsomer, E. Meyer, K. Demeyere, H. W. Barkema, A. de Kruif, and S. De
Vliegher. 2009. Heifer and quarter characteristics associated with periparturient blood
and milk neutrophil apoptosis in healthy heifers and in heifers with subclinical mastitis. J.
Dairy Sci. 92:4330-4339.
Piepers, S., G. Opsomer, H. W. Barkema, A. de Kruif, and S. De Vliegher. 2010. Heifers
infected with coagulase-negative staphylococci in early lactation have fewer cases of
clinical mastitis and higher milk production in their first lactation than noninfected
heifers. J. Dairy Sci. 93:2014-2024.
Piepers, S., K. Peeters, G. Opsomer, H. W. Barkema, K. Frankena, and S. De Vliegher. 2011.
Pathogen group specific risk factors at herd, heifer and quarter levels for intramammary
infections in early lactating dairy heifers. Prev. Vet. Med. 99:91-101.
Piepers, S., Y. H. Schukken, P. Passchyn, and S. De Vliegher. 2013. The effect of
intramammary infection with coagulase-negative staphylococci in early lactating heifers
on milk yield throughout first lactation revisited. J. Dairy Sci. 96:5095-5105.
Piessens, V., E. Van Coillie, B. Verbist, K. Supré, G. Braem, A. Van Nuffel, L. De Vuyst, M.
Heyndrickx, and S. De Vliegher. 2011. Distribution of coagulase-negative Staphylococcus
species from milk and environment of dairy cows differs between herds. J. Dairy Sci.
94:2933-2944.
Pighetti, G. M., and M. Rambeaud. 2006. Genome conservation between the bovine and
human interleukin-8 receptor complex: Improper annotation of bovine interleukin-8
receptor b identified. Vet. Immunol. Immunopathol. 114:335-340.
Pighetti, G. M., and A. A. Elliott. 2011. Gene polymorphisms: the keys for marker assisted
selection and unraveling core regulatory pathways for mastitis resistance. J. Mammary
Gland Biol. Neoplasia. 16:421-432.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Politis, I., X. Zhao, B. W. Mcbride, and J. H. Burton. 1992. Function of bovine mammary
macrophages as antigen-presenting cells. Vet. Immunol. Immunopathol. 30:399-410.

Chapter 1 General introduction
39
Rainard, P., and C. Riollet. 2006. Innate immunity of the bovine mammary gland. Vet. Res.
37:369-400.
Rambeaud, M., R. A. Almeida, G. M. Pighetti, and S. P. Oliver. 2003. Dynamics of leukocytes
and cytokines during experimentally induced Streptococcus uberis mastitis. Vet.
Immunol. Immunopathol. 96:193-205.
Rambeaud, M., and G. M. Pighetti. 2005. Impaired neutrophil migration associated with
specific bovine CXCR2 genotypes. Infect. Immun. 73:4955-4959.
Rambeaud, M., R. Clift, and G. M. Pighetti. 2006. Association of a bovine CXCR2 gene
polymorphism with neutrophil survival and killing ability. Vet. Immunol. Immunopathol.
111:231-238.
Rambeaud, M., and G. M. Pighetti. 2007. Differential calcium signaling in dairy cows with
specific CXCR1 genotypes potentially related to interleukin-8 receptor functionality.
Immunogenetics 59:53-58.
Ramirez, N. F., G. Keefe, I. Dohoo, J. Sanchez, O. Arroyave, J. Ceron, M. Jaramillo, and L. G.
Palacio. 2014. Herd- and cow-level risk factors associated with subclinical mastitis in
dairy farms from the High Plains of the northern Antioquia, Colombia. J. Dairy Sci.
97:4141-4150.
Rupp, R., and D. Boichard. 2003. Genetics of resistance to mastitis in dairy cattle. Vet. Res.
34:671-688.
Russell, C. D., S. Widdison , J. A. Leigh, and T. J. Coffey. 2012. Identification of single
nucleotide polymorphisms in the bovine Toll-like receptor 1 gene and association with
health traits in cattle. Vet. Res. 43:17.
Sahana, G., B. Guldbrandtsen, B. Thomsen, L. E. Holm, F. Panitz, R. F. Brøndum, C. Bendixen,
and M. S. Lund. 2014. Genome-wide association study using high-density single
nucleotide polymorphism arrays and whole-genome sequences for clinical mastitis traits
in dairy cattle. J. Dairy Sci. 97: 7258–7275.
Sampimon, O., H. W. Barkema, I. Berends, J. Sol, and T. Lam. 2009. Prevalence of
intramammary infection in Dutch dairy herds. J. Dairy Res. 76:129-136.
Santos, M. V., Y. Ma, and D. M. Barbano. 2003. Effect of somatic cell count on proteolysis
and lipolysis in pasteurized fluid milk during shelf-life storage. J. Dairy Sci. 86:2491-2503.
Schaeffer, L. R. 2006. Strategy for applying genome-wide selection in dairy cattle. J. Anim.
Breed. Genet. 123:218-223.

Chapter 1 General introduction
40
Schukken, Y. H., F. J. Grommers, D. Vandegeer, H. N. Erb, and A. Brand. 1990. Risk factors for
clinical mastitis in herds with a low bulk milk somatic cell count .1. Data and risk factors
for all cases. J. Dairy Sci. 73:3463-3471.
Schukken, Y. H., D. J. Wilson, F. Welcome, L. Garrison-Tikofsky, and R. N. Gonzalez. 2003.
Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 34:579-596.
Schukken, Y. H., R. N. Gonzalez, L. L. Tikofsky, H. F. Schulte, C. G. Santisteban, F. L. Welcome,
G. J. Bennett, M. J. Zurakowski, and R. N. Zadoks. 2009a. CNS mastitis: nothing to worry
about? Vet. Microbiol. 134:9-14.
Schukken, Y. H., J. Hertl, D. Bar, G. J. Bennett, R. N. Gonzalez, B. J. Rauch, C. Santisteban, H. F.
Schulte, L. Tauer, F. L. Welcome, and Y. T. Grohn. 2009b. Effects of repeated Gram-
positive and Gram-negative clinical mastitis episodes on milk yield loss in Holstein dairy
cows. J. Dairy Sci. 92:3091-3105.
Schukken, Y. H., J. Gunther, J. Fitzpatrick, M. C. Fontaine, L. Goetze, O. Holst, J. Leigh, W.
Petzl, H. J. Schuberth, A. Sipka, D. G. E. Smith, R. Quesnell, J. Watts, R. Yancey, H. Zerbe,
A. Gurjar, R. N. Zadoks, and H. M. Seyfert. 2011. Host response patterns of
intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 144:270-289.
Sears, P. M., B. S. Smith, P. B. English, P. S. Herer, and R. N. Gonzalez. 1990. Shedding pattern
of Staphylococcus aureus from bovine intramammary infections. J. Dairy Sci. 73:2785-
2789.
Seegers, H., C. Fourichon, and F. Beaudeau. 2003. Production effects related to mastitis and
mastitis economics in dairy cattle herds. Vet. Res. 34:475-491.
Selsted, M. E., Y. Q. Tang, W. L. Morris, P. A. McGuire, M. J. Novotny, W. Smith, A. H.
Henschen, and J. S. Cullor. 1993. Purification, primary structures, and antibacterial
activities of beta-defensins, a new family of antimicrobial peptides from bovine
neutrophils. J. Biol. Chem. 268:6641-6648.
Sharma, B. S., I. Leyva, F. Schenkel, and N. A. Karrow. 2006. Association of toll-like receptor 4
polymorphisms with somatic cell score and lactation persistency in Holstein bulls. J. Dairy
Sci. 89:3626-3635.
Simojoki, H., T. Orro, S. Taponen, and S. Pyorala. 2009. Host response in bovine mastitis
experimentally induced with Staphylococcus chromogenes. Vet. Microbiol. 134:95-99.

Chapter 1 General introduction
41
Simojoki, H., T. Salomaki, S. Taponen, A. Iivanainen, and S. Pyorala. 2011. Innate immune
response in experimentally induced bovine intramammary infection with Staphylococcus
simulans and S. epidermidis. Vet. Res. 42:49.
Smits, E., C. Burvenich, A. J. Guidry, R. Heyneman, and A. Massart-Leen. 1999. Diapedesis
across mammary epithelium reduces phagocytic and oxidative burst of bovine
neutrophils. Vet. Immunol. Immunopathol. 68:169-176.
Sodeland, M., M. P. Kent, H. G. Olsen, M. A. Opsal, M. Svendsen, E. Sehested, B. J. Hayes, and
S. Lien. 2011. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20
in Norwegian Red cattle. Anim Genet. 42:457-465.
Sol, J., O. C. Sampimon, H. W. Barkema, and Y. H. Schukken. 2000. Factors associated with
cure after therapy of clinical mastitis caused by Staphylococcus aureus. J. Dairy Sci.
83:278-284.
Sordillo, L. M., and L. A. Babiuk LA. 1991. Modulation of bovine mammary neutrophil
function during the periparturient period following in vitro exposure to recombinant
bovine interferon gamma. Vet Immunol Immunopathol. 27:393-402.
Sordillo, L. M., K. Shafer-Weaver, and D. DeRosa. 1997. Immunobiology of the mammary
gland. J. Dairy Sci. 80:1851-1865.
Sordillo, L. M., and K. L. Streicher. 2002. Mammary gland immunity and mastitis
susceptibility. J. Mammary Gland Biol. Neoplasia 7:135-146.
Sordillo, L. M. 2005. Factors affecting mammary gland immunity and mastitis susceptibility.
Livest. Prod. Sci. 98:89-99.
Stevens, M. G. H., L. J. Peelman, B. De Spiegeleer, A. Pezeshki, G. R. Van De Walle, L.
Duchateau, and C. Burvenich. 2011. Differential gene expression of the toll-like receptor-
4 cascade and neutrophil function in early- and mid-lactating dairy cows. J. Dairy Sci.
94:1277-1288.
Stillie, R., S. M. Farooq, J. R. Gordon, and A. W. Stadnyk. 2009. The functional significance
behind expressing two IL-8 receptor types on PMN. J. Leukoc. Biol. 86:529-543.
Strillacci, M. G., E. Frigo, F. Schiavini, A. B. Samoré, F. Canavesi, M. Vevey, M. C. Cozzi, M.
Soller, E. Lipkin, and A. Bagnato. 2014. Genome-wide association study for somatic cell
score in Valdostana Red Pied cattle breed using pooled DNA. BMC Genet. 15:106.

Chapter 1 General introduction
42
Supré, K., F. Haesebrouck, R. N. Zadoks, M. Vaneechoutte, S. Piepers, and S. De Vliegher.
2011. Some coagulase-negative Staphylococcus species affect udder health more than
others. J. Dairy Sci. 94:2329-2340.
Thompson-Crispi, K. A., and B. A. Mallard. 2012. Type 1 and type 2 immune response profiles
of commercial dairy cows in 4 regions across Canada. Can. J. Vet. Res. 76:120-128.
Timms, L. L., and L. H. Schultz. 1987. Dynamics and significance of coagulase-negative
staphylococcal intramammary infections. J. Dairy Sci. 70:2648-2657.
Tomita, T., B. Meehan, N. Wongkattiya, J. Malmo, G. Pullinger, J. Leigh, and M. Deighton.
2008. Identification of Streptococcus uberis multilocus sequence types highly associated
with mastitis. Appl. Environ. Microbiol. 74:114-124.
Vanderhaeghen, W., S. Piepers, F. Leroy, E. Van Coillie, F. Haesebrouck, and S. De Vliegher.
2014. Invited review: effect, persistence, and virulence of coagulase-negative
Staphylococcus species associated with ruminant udder health. J. Dairy Sci. 97:5275-
5293.
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe, and F.
Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by
geometric averaging of multiple internal control genes. Genome Biol. 3: research0034.
Van Oostveldt, K., M. J. Paape, and C. Burvenich. 2002. Apoptosis of bovine neutrophils
following diapedesis through a monolayer of endothelial and mammary epithelial cells. J.
Dairy Sci. 85:139-147.
Van Oostveldt, K., F. Vangroenweghe, H. Dosogne, and C. Burvenich. 2001. Apoptosis and
necrosis of blood and milk polymorphonuclear leukocytes in early and midlactating
healthy cows. Vet. Res. 32:617-622.
Verbist, B., V. Piessens, A. Van Nuffel, L. De Vuyst, M. Heyndrickx, L. Herman, E. Van Coillie,
and S. De Vliegher. 2011. Sources other than unused sawdust can introduce Klebsiella
pneumoniae into dairy herds. J. Dairy Sci. 94:2832-2839.
Wang, C., M. Liu, Q. Li, Z. Ju, J. Huang, and J. Li. 2011. Three novel singlenucleotide
polymorphisms of MBL1 gene in Chinese native cattle and their associations with milk
performance traits. Vet. Immunol. Immunopathol. 139:229–236.
Watts, J. L. 1988. Etiological agents of bovine mastitis. Vet. Microbiol. 16:41-66.
Wellnitz, O., and R. M. Bruckmaier. 2012. The innate immune response of the bovine
mammary gland to bacterial infection. Vet. J. 192:148-152.

Chapter 1 General introduction
43
Whist, A. C., O. Osteras, and L. Solverod. 2007. Streptococcus dysgalactiae isolates at calving
and lactation performance within the same lactation. J. Dairy Sci. 90:766-778.
Windig, J. J., W. Ouweltjes, J. Ten Napel, G. de Jong, R. F. Veerkamp, and Y. de Haas. 2010.
Combining somatic cell count traits for optimal selection against mastitis. J. Dairy Sci.
93:1690-1701.
Wolff, C., M. Espetvedt, A. K. Lind, S. Rintakoski, A. Egenvall, A. Lindberg, and U. Emanuelson.
2012. Completeness of the disease recording systems for dairy cows in Denmark,
Finland, Norway and Sweden with special reference to clinical mastitis. BMC Vet. Res.
8:131.
Youngerman, S. M., A. M. Saxton, S. P. Oliver, and G. M. Pighetti. 2004a. Association of
CXCR2 polymorphisms with subclinical and clinical mastitis in dairy cattle. J. Dairy Sci.
87:2442-2448.
Youngerman, S. M., A. M. Saxton, and G. M. Pighetti. 2004b. Novel single nucleotide
polymorphisms and haplotypes within the bovine CXCR2 gene. Immunogenetics 56:355-
359.
Zadoks, R. N., B. E. Gillespie, H. W. Barkema, O. C. Sampimon, S. P. Oliver, and Y. H.
Schukken. 2003. Clinical, epidemiological and molecular characteristics of Streptococcus
uberis infections in dairy herds. Epidemiol. Infect. 130:335-349.
Zadoks, R. N., and Y. H. Schukken. 2006. Use of molecular epidemiology in veterinary
practice. Vet. Clin. North Am. Food Anim Pract. 22:229-261.
Zadoks, R. N., J. R. Middleton, S. McDougall, J. Katholm, and Y. H. Schukken. 2011. Molecular
epidemiology of mastitis pathogens of dairy cattle and comparative relevance to
humans. J. Mammary Gland Biol. Neoplasia 16:357-372.
Zerbe, H., N. Schneider, W. Leibold, T. Wensing, T. A. M. Kruip, and H. J. Schuberth. 2000.
Altered functional and immunophenotypical properties of neutrophilic granulocytes in
postpartum cows associated with fatty liver. Theriogenology 54:771-786.

44

45
Chapter 2
Aims and outline of the thesis

Chapter 2 Aims of the thesis
46

Chapter 2 Aims of the thesis
47
Mastitis, an inflammation of the mammary gland mostly caused by bacterial
intramammary infection, is one of the most common diseases in dairy cattle. Mastitis
resistance, defined as the cow’s ability to hinder pathogens from causing intramammary
infection, is partly genetically determined. Identification of genetic polymorphisms linked
with mastitis resistance allows for marker assisted selection for udder health. Neutrophils
play an important role in the mammary gland immunity. Binding of interleukin 8 on the
chemokine (C-X-C motif) receptor 1 (CXCR1) enhances migration, viability and activity of
neutrophils. Polymorphisms in the CXCR1 gene potentially alter expression or functionality
of the receptor causing impaired migration, viability or activity of neutrophils and variation
in mastitis resistance (Figure 2.1).
The general aim of this work is to increase our understanding of the role of CXCR1
gene polymorphisms in neutrophil traits (Chapter 3), subclinical mastitis (Chapter 4) and
clinical mastitis (Chapter 5).
Even before the first milking, a large proportion of heifers suffers from mastitis.
Previous research demonstrated milk neutrophil viability to differ among early lactating
dairy heifers. The aim of Chapter 3.1 is to analyze whether part of this variation is due to
CXCR1 polymorphisms. Additional to neutrophil viability, neutrophil activity might be
associated with CXCR1 polymorphisms. To test this hypothesis, reactive oxygen species
generation of blood neutrophils from early lactating heifers with different CXCR1 genotype is
studied in Chapter 3.2.
Subclinical heifer mastitis threatens the future performances of heifers, especially
when so called major mastitis pathogens (e.g. Staphylococcus aureus and Streptococcus
uberis) are involved. In contrast, the impact on udder health of minor pathogens (e.g.
coagulase-negative staphylococci) is limited. In Chapter 4.1, pathogen-group specific
associations between CXCR1 polymorphisms and subclinical heifer mastitis are studied. To
further study the role of CXCR1 polymorphisms in the host response, quarters of heifers with
different CXCR1 genotype were experimentally infected. Differences in somatic cell count
and milk neutrophil viability following experimental intramammary infection are analyzed
(Chapter 4.2).
Subclinical and clinical mastitis are two different traits. Unlike other regions in the
world, little is known about clinical mastitis on Flemish dairy herds. The aim of Chapter 5.1 is
to estimate the pathogen-specific incidence of clinical mastitis in Flanders. Because different

Chapter 2 Aims of the thesis
48
mastitis pathogens elicit different host responses, CXCR1 polymorphisms might influence
resistance against specific pathogens. In Chapter 5.2, pathogen-specific associations
between CXCR1 polymorphisms and incidence of clinical mastitis are tested.
As previously mentioned, CXCR1 polymorphisms potentially affect receptor expression
or functionality. In Chapter 6, associations between CXCR1 polymorphisms and CXCR1 gene
expression in milk somatic cells of healthy and experimentally infected quarters are studied.
Additionally, the impact of experimental infection on expression of CXCR1 and commonly
used reference genes is analyzed.
In Chapter 7, the overall results, future perspectives of CXCR1 research and potential
practical applications are discussed.

Chap
ter 2
Aim
s of t
he th
esis
49
Fi
gure
2.1
. Fro
m g
enot
ype
to p
heno
type
: aim
s and
out
line
of th
e th
esis.
1 Si
ngle
nuc
leot
ide
poly
mor
phism
s.
2 Reac
tive
oxyg
en sp
ecie
s.

50

51
Chapter 3
The role of CXCR1 gene polymorphisms in bovine
neutrophil traits

52

53
Chapter 3.1
Association of CXCR1 polymorphisms with apoptosis,
necrosis and concentration of milk neutrophils
in early lactating dairy heifers
J. Verbeke1, S. Piepers1, L. Peelman2, M. Van Poucke2, S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine,
Ghent University, Belgium
Research in Veterinary Science, 2014, 97: 55-59.

Chapter 3.1 The CXCR1 gene and neutrophil viability
54

Chapter 3.1 The CXCR1 gene and neutrophil viability
55
Abstract
Associations between polymorphisms in the candidate gene CXCR1, encoding the
chemokine (C-X-C motif) receptor 1, and udder health have been identified before. In the
present study, associations between the CXCR1 genotype (whole coding region) and
apoptosis, necrosis, and concentration of milk neutrophils of 292 quarters belonging to 73
early lactating dairy heifers were studied. In uninfected quarters, % milk neutrophil
apoptosis was higher in c.980GG heifers [least squares means (LSM) 27%] compared to
c.980AG heifers (LSM 16%), whereas in infected quarters, % milk neutrophil apoptosis was
higher in c.642GG heifers (LSM 29%) compared to c.642AG heifers (LSM 18%). Differences in
milk neutrophil concentration between infected and uninfected quarters were smaller in
c.980AG heifers than in c.980GG heifers. An association between the CXCR1 genotype and
necrosis of milk neutrophils could not be demonstrated. Results indicate that CXCR1
polymorphisms influence viability and concentration of milk neutrophils and provide a
foundation for future research.
Key words
Apoptosis, necrosis, CXCR1, dairy heifer, neutrophil

Chapter 3.1 The CXCR1 gene and neutrophil viability
56
1. Introduction
Innate immune responses are the predominant form of defense in the early stage of
intramammary infection (IMI) (Sordillo et al., 1997), with migration of neutrophils from the
blood to the site of infection being a key component (Paape et al., 2003). Both number and
viability of neutrophils that pathogens encounter are of importance (Mehrzad et al., 2004).
Apoptosis and necrosis impair the phagocytic and respiratory burst capacity of neutrophils
(Mehrzad et al., 2004; Van Oostveldt et al., 2002). Percentage milk neutrophil apoptosis and
milk neutrophil concentration differ among heifers (Piepers et al., 2009b) and are potentially
genetically determined. In this context, genes involved in the innate immunity of the
mammary gland form an interesting area of study. Recently, we reported 16 polymorphisms
in the coding region of CXCR1 and revealed an interesting association between
polymorphism c.980A>G (dbSNP ID: rs43323012) and the odds of IMI in early lactating
heifers: heifers with genotype c.980AG were less likely to have IMI due to major mastitis
pathogens compared to heifers with genotype c.980GG but were not likely to have less IMI
by coagulase-negative staphylococci, still considered as minor pathogens (Verbeke et al.,
2012). The CXCR1 gene actually encodes one of the two receptors present on neutrophils for
interleukin 8 (IL-8), named chemokine (C-X-C motif) receptor 1 (CXCR1). Interleukin 8 is
known as the main chemoattractant for neutrophils after intramammary invasion of a
pathogen (Barber and Yang, 1998) and has also been shown to increase activity (Mitchell et
al., 2003) and inhibits apoptosis of neutrophils (Kettritz et al., 1998). There is strong in vitro
evidence that polymorphism CXCR1 c.735C>G (dbSNP ID: rs208795699) affects the survival
and migration of bovine blood neutrophils in response to IL-8 (reviewed in Pighetti et al.,
2012). Polymorphisms in CXCR1 potentially alter the functionality of the receptor and might
hence be associated with neutrophil viability and concentration in milk, and could explain
our previous findings on susceptibility to IMI to some extent.
We aimed at analyzing whether or not variation in milk neutrophil apoptosis, milk
neutrophil necrosis, and milk neutrophil concentration in (quarters of) early lactating dairy
heifers can be due to CXCR1 polymorphisms.

Chapter 3.1 The CXCR1 gene and neutrophil viability
57
2. Materials and methods
2.1 Phenotypic data
Phenotypic records from 73 heifers from 19 commercial Flemish dairy herds (Piepers
et al., 2009b) were available. On average, four heifers per farm were included, ranging
between 1 and 7. Forty-six different sires and 72 different dams were noted, indicating that
the heifers were not closely related to each other. Apoptosis, necrosis, and concentration of
milk neutrophils were measured on quarter milk samples collected in early lactation
(between 1 and 4 DIM). To determine the IMI status, additional quarter milk samples (5 ml)
were aseptically collected between 1 and 4 DIM and between 5 and 8 DIM for bacteriological
culturing. The interval between both samplings was at least 3 days. Samples were taken just
before morning milking (Piepers et al., 2011; Verbeke et al., 2012).
The % milk neutrophil apoptosis and necrosis were measured flow cytometrically using
a double fluorescein isothiocyanate-annexin-V and propidium iodide staining. Milk
neutrophils and other milk cells were differentiated by a two-step fluorescent
immunolabeling as described before (Piepers et al., 2009a, b). Approximate concentration of
neutrophils in milk, further referred as milk neutrophil concentration, was quantified based
on the absolute number of milk neutrophil events as determined by flow cytometry, the
sample acquisition time (tacq, s), and the flow rate (FR, μL/s) (Piepers et al., 2009b). The IMI
status of each quarter was determined using bacteriological culture of two consecutive milk
samples as described before (Verbeke et al., 2012).
2.2 Genotypic data
Genotyping of the whole single-exon coding region (1083 bp) of bovine CXCR1 was
performed by sequencing as described by Verbeke et al. (2012). In brief, DNA was extracted
from blood samples (100 μl) using a proteinase K digestion method (Van Poucke et al., 2005)
and its concentration was estimated with a ND spectrophotometer (NanoDrop). A PCR mix
containing approximately 100 ng genomic DNA, 1.0 μl 10x FastStart Taq DNA Polymerase
Buffer (Roche Applied Science), dNTP Mix (0.2 mM each; BIOLINE), 0.25 μM forward primer
(Integrated DNA Technologies, 5'-TCCTTGATGAGAGTGATTTGGA-3'), 0.25 μM reverse primer
(Integrated DNA Technologies, 5'-TTGACATGGGACTGTGAACG-3') and 0.5 U Taq DNA
Polymerase (Roche Applied Science) was made.

Chapter 3.1 The CXCR1 gene and neutrophil viability
58
The PCR program consisted of an initiation step of 5 min at 95°C followed by 30
amplification cycles (denaturation for 30 s at 95°C, annealing for 30 s at 62°C and extension
for 75 s at 72°C) and a final 4 min elongation step at 72°C. All amplicons were sequenced by
direct sequencing with the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied
Biosystems) on a 16-capillary 3130xl DNA Analyzer (Applied Biosystems) according to the
manufacturer’s protocol. Sequencing reactions per sample were performed in duplicate
using the forward and reverse primers. Deviations from Hardy-Weinberg equilibrium of each
studied polymorphism were analyzed using chi-square goodness of fit tests.
2.3 Statistical analysis
Associations between phenotypic traits and genotype were analyzed for
polymorphisms c.642G>A, c.735C>G, c.816C>A and c.980A>G (dbSNP ID: rs109587131,
rs208795699, rs210501501, and rs43323012) only. All other polymorphisms were in full
linkage disequilibrium (correlation 100%) with one of these polymorphisms, and results
would be the same for that reason. In all models, statistical significance was assessed at P ≤
0.05 while a P-value between 0.05 and 0.10 was considered as a tendency toward
significance. Transformation were performed to obtain normalized distributions. Percentage
milk neutrophil apoptosis and necrosis were square-root transformed whereas milk
neutrophil concentration was log transformed.
The association between the different polymorphisms (predictor variables of main
interest) and the % milk neutrophil apoptosis and necrosis, and milk neutrophil
concentrations (outcome variables) were determined using similar linear mixed regression
models. Herd and heifer were included as random effects to correct for clustering of heifers
within herds and quarters within heifers, respectively (RANDOM statement) (PROC MIXED,
SAS 9.2, SAS Institute Inc., NC, US). The different models included IMI status (0 = uninfected
quarter versus 1 = infected quarter, all pathogens combined) and polymorphism c.642G>A,
c.735C>G, c.816C>A or c.980A>G, respectively, as categorical fixed effects. The interaction
between both variables was also tested. If the interaction term was significant, post-hoc
pairwise comparisons were performed to test differences between quarters with different
IMI statuses or genotypes. Post-hoc comparisons were adjusted according to Bonferroni.

Chapter 3.1 The CXCR1 gene and neutrophil viability
59
3. Results
3.1 Descriptive statistics
Participating herds consisted on average of 48 lactating cows with an average 365-d
milk production of 8,368 kg of milk. The average bulk milk SCC was 278,736 cells/ml between
2006 and 2007 (Piepers et al., 2009b).
The IMI status in early lactation could be determined for 245 out of 292 quarters
(84%). Two quarters showed signs of clinical mastitis between 1 and 4 DIM. One-hundred-
and-two quarters (42%) were uninfected. The large majority (91%) of the IMI were caused by
coagulase-negative staphylococci (n = 130). Of the remaining 13 infected quarters,
Corynebacterium bovis (n = 2), Staphylococcus aureus (n = 5), Streptococcus dysgalactiae (n =
1) and esculine-positive cocci (n = 5), respectively, were identified as the cause. The average
milk neutrophil necrosis was 29% (IQR 14 - 46%) for uninfected quarters, and 28% (IQR 17 –
43%) for infected quarters. The average milk neutrophil apoptosis was 23% (IQR 15 - 34%)
for uninfected quarters, and 22% (IQR 15 – 34%) for infected quarters. The average milk
neutrophil concentration was higher in infected quarters (log10 = 3.7; IQR 2.9 - 4.3)
compared to uninfected quarters (log10 = 3.3; IQR 2.9 – 3.8).
3.2 CXCR1 genotypes
All 16 polymorphisms (c.37A>T, c.38T>A, c.68G>A, c.291C>T, c.333T>C, c.337G>A,
c.365T>C, c.570G>A, c.642G>A, c.735C>G, c.816C>A, c.819G>A, c.980A>G, c.995A>G,
c.1008C>T, and c.1068G>A) described by Verbeke et al. (2012) were observed in this subset
of heifers. The studied population was in Hardy-Weinberg equilibrium (P > 0.05) for all
polymorphisms. Four (H1-H4, [GenBank: HM13954-7]) of the five haplotypes reported and
labeled (H1-H5) by Verbeke et al. (2012) were present (Table 3.1.1).

Chapter 3.1 The CXCR1 gene and neutrophil viability
60
Table 3.1.1. CXCR1 haplotype sequences detected in 73 Belgian dairy heifers
Label 371 38 68 291 333 337 365 570 642 735 816 819 980 995 1008 1068 Freq. Accession2
H1 A T G C T G T G G C C G A A C G 0.16 HM013954
H2 A T G T C A T G G C A G G G T G 0.29 HM013955
H3 T A A C C A C A A G C A G G C A 0.36 HM013956
H4 T A A C C A C A G G C A G G C A 0.19 HM013957
1Numbers indicate positions of the polymorphisms relative to the start codon. 2GenBank accession number.
3.3 Associations between CXCR1 polymorphisms and milk neutrophil apoptosis and necrosis
Neither CXCR1 polymorphisms nor IMI status were associated with % milk neutrophil
necrosis (P > 0.05) (Table 3.1.2). In contrast, % milk neutrophil apoptosis was significantly
associated with polymorphism c.980A>G (P = 0.02), and tended to be associated with
polymorphisms c.642G>A and c.816C>A (P = 0.10 and 0.08, respectively) (Table 3.1.2).
However, the association of genotype and % milk neutrophil apoptosis was modified by the
IMI status of quarters; the interaction terms between IMI status and c.642G>A, c.735C>G,
and c.980A>G were statistically significant (Table 3.1.2, Fig 3.1.1). Taking genotype, IMI
status and their interaction into account, pairwise post-hoc comparisons revealed two
significant differences. The % milk neutrophil apoptosis was significantly higher in infected
quarters of heifers expressing genotype c.642GG (back-transformed LSM: 29%) compared to
infected quarters of heifers expressing genotype c.642GA (back-transformed LSM: 18%)
(Bonferroni corrected P = 0.01) (Figure 3.1.1A). Additionally, % milk neutrophil apoptosis
tended to be higher in infected quarters of heifers expressing genotype c.735CC (back-
transformed LSM: 29%) compared to infected quarters of heifers expressing genotype
c.735CG (back-transformed LSM: 19%) (Bonferroni corrected P = 0.09) (Figure 3.1.1B).
Furthermore, percentage milk neutrophil apoptosis was significantly higher in uninfected
quarters of heifers expressing genotype c.980GG (back-transformed LSM: 26%) compared to
uninfected quarters of heifers expressing genotype c.980AG (back-transformed LSM: 17%)
(Bonferroni corrected P = 0.007) (Figure 3.1.1C).

Chapter 3.1 The CXCR1 gene and neutrophil viability
61
3.4 Associations between CXCR1 polymorphisms and milk neutrophil concentration
In all models, milk neutrophil concentration was significantly associated with IMI status
but not with CXCR1 polymorphisms (Table 3.1.2). However, the effect of IMI status tended to
depend on c.980A>G genotype (interaction term P = 0.08) (Table 3.1.2). Infected quarters
had significantly higher milk neutrophil concentration than uninfected quarters in heifers
with genotype c.980GG (P < 0.001, Bonferroni corrected P = 0.001) but not in heifers with
genotype c.980AG (P = 0.89) (Figure 3.1.1D).
Figure 3.1.1. % milk neutrophil apoptosis by CXCR1 polymorphisms c.642A>G, c.735C>G, and c.980A>G,
respectively (A-C). Log10 transformed milk neutrophil concentration by CXCR1 polymorphism c.980A>G (D).
Data are back-transformed LSM ± SE. Significant pairwise differences are indicated with letters (Bonferroni
corrected P ≤ 0.05). Genotype c.980AA was not observed in the study population.
0
5
10
15
20
25
30
35
c.642GG c.642GA c.642AA
% m
ilk n
eutr
ophi
l apo
ptos
is
Uninfected
Infected
0
5
10
15
20
25
30
35
c.735CC c.735CG c.735GG
% m
ilk n
eutr
ophi
l apo
ptos
is
Uninfected
Infected
0
5
10
15
20
25
30
35
c.980AG c.980GG
% m
ilk n
eutr
ophi
l apo
ptos
is
Uninfected
Infected
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
c.980AG c.980GG
Log1
0 tr
ansf
orm
ed m
ilk
neut
roph
il co
ncen
trat
ion
UninfectedInfected
a
b
a
b
b a
A B
C D
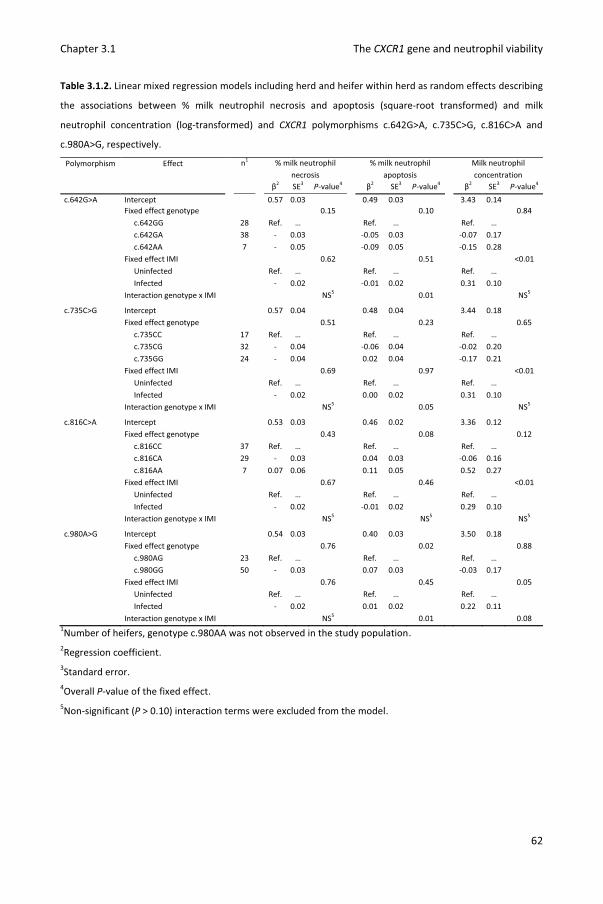
Chapter 3.1 The CXCR1 gene and neutrophil viability
62
Table 3.1.2. Linear mixed regression models including herd and heifer within herd as random effects describing
the associations between % milk neutrophil necrosis and apoptosis (square-root transformed) and milk
neutrophil concentration (log-transformed) and CXCR1 polymorphisms c.642G>A, c.735C>G, c.816C>A and
c.980A>G, respectively.
Polymorphism Effect n1 % milk neutrophil necrosis
% milk neutrophil apoptosis
Milk neutrophil concentration
β2 SE3 P-value4 β2 SE3 P-value4 β2 SE3 P-value4 c.642G>A Intercept 0.57 0.03 0.49 0.03 3.43 0.14 Fixed effect genotype 0.15 0.10 0.84
c.642GG 28 Ref. … Ref. … Ref. … c.642GA 38 - 0.03 -0.05 0.03 -0.07 0.17 c.642AA 7 - 0.05 -0.09 0.05 -0.15 0.28 Fixed effect IMI 0.62 0.51 <0.01 Uninfected Ref. … Ref. … Ref. … Infected - 0.02 -0.01 0.02 0.31 0.10 Interaction genotype x IMI NS5 0.01 NS5
c.735C>G Intercept 0.57 0.04 0.48 0.04 3.44 0.18 Fixed effect genotype 0.51 0.23 0.65 c.735CC 17 Ref. … Ref. … Ref. … c.735CG 32 - 0.04 -0.06 0.04 -0.02 0.20 c.735GG 24 - 0.04 0.02 0.04 -0.17 0.21 Fixed effect IMI 0.69 0.97 <0.01 Uninfected Ref. … Ref. … Ref. … Infected - 0.02 0.00 0.02 0.31 0.10 Interaction genotype x IMI NS5 0.05 NS5
c.816C>A Intercept 0.53 0.03 0.46 0.02 3.36 0.12 Fixed effect genotype 0.43 0.08 0.12 c.816CC 37 Ref. … Ref. … Ref. … c.816CA 29 - 0.03 0.04 0.03 -0.06 0.16 c.816AA 7 0.07 0.06 0.11 0.05 0.52 0.27 Fixed effect IMI 0.67 0.46 <0.01 Uninfected Ref. … Ref. … Ref. … Infected - 0.02 -0.01 0.02 0.29 0.10 Interaction genotype x IMI NS5 NS5 NS5
c.980A>G Intercept 0.54 0.03 0.40 0.03 3.50 0.18 Fixed effect genotype 0.76 0.02 0.88 c.980AG 23 Ref. … Ref. … Ref. … c.980GG 50 - 0.03 0.07 0.03 -0.03 0.17 Fixed effect IMI 0.76 0.45 0.05 Uninfected Ref. … Ref. … Ref. … Infected - 0.02 0.01 0.02 0.22 0.11 Interaction genotype x IMI NS5 0.01 0.08
1Number of heifers, genotype c.980AA was not observed in the study population. 2Regression coefficient. 3Standard error. 4Overall P-value of the fixed effect. 5Non-significant (P > 0.10) interaction terms were excluded from the model.

Chapter 3.1 The CXCR1 gene and neutrophil viability
63
4. Discussion
Because heifer mastitis is partly genetically determined (Wanner et al., 1998; Nash et
al., 2003) and innate immune responses provide the first line of defense against invading
pathogens (Paape et al., 2002), genes involved in these processes form interesting study
objects and potential candidate genes for selection. In this study, we analyzed in vivo
associations between polymorphisms in the coding region of CXCR1 and apoptosis, necrosis,
and concentration of milk neutrophils.
Polymorphisms in CXCR1 were associated with % milk neutrophil apoptosis but not
with % milk neutrophil necrosis. In infected quarters, % milk neutrophil apoptosis was higher
in c.642GG heifers compared to c.642GA heifers and tended to be higher in c.735CC heifers
compared to c.735CG heifers. The association with c.735C>G is in agreement with an in vitro
study performed by another research group (Pighetti et al. 2012; Rambeaud et al., 2006).
Rambeaud et al. (2006) observed a higher increase in survival in response to IL-8 for
neutrophils isolated from c.735CC cows compared to neutrophils isolated from c.735GG
cows. However, whether a low proportion of apoptotic neutrophils in infected quarters is
desirable can be debated. Although apoptosis impairs activity of neutrophils (Van Oostveldt
et al., 2002), it is vital for the resolution of the intramammary inflammation (Paape et al.,
2003). Also, stage of infection (acute versus chronic IMI) could have been different,
explaining the differences in % apoptosis between genotypes. In uninfected quarters, % milk
neutrophil apoptosis was higher in c.980GG heifers compared to c.980AG heifers. Apoptosis
impairs phagocytic and respiratory burst activity of neutrophils (Van Oostveldt et al., 2002)
and viability of resident milk neutrophils was shown to be negatively correlated with the
severity of experimentally induced IMI (Mehrzad et al., 2004). Based on the latter results, we
hypothesize that the higher proportion of apoptotic resident neutrophils in heifers with
genotype c.980GG is associated with a reduction in overall neutrophil functionality and
ability to combat invading mastitis pathogens. This could eventually explain the higher
likelihood of IMI with major pathogens that we identified before (Verbeke et al., 2012).
Although genotype c.980GG seems to be undesirable, allele frequency of c.980G was high in
the study population (0.78). The study population lacked c.980AA animals but no deviation
from Hardy-Weinberg equilibrium was detected. Associations with other traits (e.g. milk

Chapter 3.1 The CXCR1 gene and neutrophil viability
64
production) could explain the low allele frequency of c.980A and are currently being studied
in a larger population.
As expected, milk neutrophil concentration was higher in infected compared to
uninfected quarters. However, differences in milk neutrophil concentration between
infected and uninfected quarters were much smaller in c.980AG heifers than in c.980GG
heifers. Milk neutrophils are needed to combat invading pathogens but high concentrations
during IMI damage the mammary parenchyma as well (Paape et al., 2003). Consequently,
tissue damage following IMI might be lower in c.980AG heifers.
The lower % apoptosis in resident milk neutrophils and smaller difference in milk
neutrophil concentration between infected and uninfected quarters heifers might indicate
higher neutrophil functionality and a more efficient immune response against invading
pathogens in c.980AG heifers. These hypotheses could be tested by measuring functionality
of neutrophils (e.g. respiratory burst activity) expressing different c.980A>G genotype and by
experimentally infecting c.980AG and GG heifers to study differences in immune response.
Because different mastitis pathogens elicit different immune responses (Bannerman et
al., 2004; Schukken et al., 2011), not only IMI status but also pathogen-species should ideally
be accounted for in our analysis. However, due to the small number of major pathogen
infections this was unfeasible. To the best of our knowledge, functional effects of bovine
CXCR1 polymorphisms have yet to be analyzed. Amino acid changes potentially influence
receptor activity through ligand binding, G-protein related signaling, and/or receptor
internalization (reviewed in Pighetti et al., 2012). Additionally, (linked) polymorphisms could
affect gene expression (Leyva-Baca et al., 2007). Associations in this study could be explained
by functional effects of the studied polymorphisms or linkage with other causal
polymorphisms.
5. Conclusions
Apoptosis of resident milk neutrophils was associated with CXCR1 polymorphisms in
vivo. Additionally, difference in milk neutrophil concentration between infected and
uninfected quarters depended on CXCR1 genotype. Although exact mechanisms remain to
be determined, the current research suggests that CXCR1 polymorphisms affect the innate
immunity of the bovine mammary gland.

Chapter 3.1 The CXCR1 gene and neutrophil viability
65
6. Acknowledgements
This study was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen). The authors wish to thank the Cattle
Breeding Association (CRV) for access to data on the relationship among heifers.
7. References
Bannerman, D. D., M. J. Paape, J. W. Lee, X. Zhao, J. C. Hope, and P. Rainard. 2004.
Escherichia coli and Staphylococcus aureus elicit differential innate immune responses
following intramammary infection. Clin. Diagn. Lab. Immunol. 11:463-472.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab. Immunol. 5:82-86.
Kettritz, R., M. L. Gaido, H. Haller, F. C. Luft, C. J. Jennette, and R. J. Falk. 1998. Interleukin-8
delays spontaneous and tumor necrosis factor-alpha-mediated apoptosis of human
neutrophils. Kidney Int. 53: 84-91.
Leyva-Baca, I., G. Pighetti, and N. A. Karrow. 2008. Genotype-specific IL8RA gene expression
in bovine neutrophils in response to Escherichia coli lipopolysaccharide challenge. Anim.
Gen. 39:298-300.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2004. Viability of milk neutrophils and severity
of bovine coliform mastitis. J. Dairy Sci. 87:4150-4162.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
Nash, D. L., G. W. Rogers, J. B. Cooper, G. L. Hargrove, and J. F. Keown. 2003. Heritability of
intramammary infections at first parturition and relationships with sire transmitting
abilities for somatic cell score, udder type traits, productive life, and protein yield. J.
Dairy Sci. 86:2684-2695.
Paape, M.J., D. D. Bannerman, X. Zhao, J. W. Lee. 2003. The bovine neutrophil: Structure and
function in blood and milk. Vet. Res. 34:597-627.

Chapter 3.1 The CXCR1 gene and neutrophil viability
66
Paape, M. J., J. Mehrzad, X. Zhao, J. Detilleux, and C. Burvenich. 2002. Defense of the bovine
mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary Gland Biol.
Neoplasia. 7:109-121.
Piepers, S., S. De Vliegher, K. Demeyere, B. N. Lambrecht, A. de Kruif, E. Meyer, and G.
Opsomer. 2009a. Technical note: Flow cytometric identification of bovine milk
neutrophils and simultaneous quantification of their viability. J. Dairy Sci. 92:626-631.
Piepers, S., G. Opsomer, E. Meyer, K. Demeyere, H. W. Barkema, A. de Kruif, and S. De
Vliegher. 2009b. Heifer and quarter characteristics associated with periparturient blood
and milk neutrophil apoptosis in healthy heifers and in heifers with subclinical mastitis. J.
Dairy Sci. 92:4330-4339.
Piepers, S., K. Peeters, G. Opsomer, H. W. Barkema, K. Frankena, S. De Vliegher. 2011.
Pathogen group specific risk factors at herd, heifer and quarter levels for intramammary
infections in early lactating dairy heifers. Prev. Vet. Med. 99:91-101.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Rambeaud, M., R. Clift, and G. M. Pighetti. 2006. Association of a bovine CXCR2 gene
polymorphism with neutrophil survival and killing ability. Vet. Immunol. Immunopathol.
111:231-238.
Schukken, Y. H., J. Gunther, J. Fitzpatrick, M. C. Fontaine, L. Goetze, O. Holst, J. Leigh, W.
Petzl, H. J. Schuberth, A. Sipka, D. G. E. Smith, R. Quesnell, J. Watts, R. Yancey, H. Zerbe,
A. Gurjar, R. N. Zadoks, and H. M. Seyfert. 2011. Host-response patterns of
intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 144:270-289.
Sordillo, L. M., K. Shafer-Weaver, and D. DeRosa. 1997. Immunobiology of the mammary
gland. J. Dairy Sci. 80:1851-1865.
Van Oostveldt, K., M. J. Paape, H. Dosogne, and C. Burvenich. 2002. Effect of apoptosis on
phagocytosis, respiratory burst and CD18 adhesion receptor expression of bovine
neutrophils. Domest. Anim. Endocrinol. 22:37-50.
Van Poucke, M., J. Vandesompele, M. Mattheeuws, A. Van Zeveren, and L. J. Peelman. 2005.
A dual fluorescent multiprobe assay for prion protein genotyping in sheep. BMC Infect.
Dis. 5:13.

Chapter 3.1 The CXCR1 gene and neutrophil viability
67
Verbeke, J., S. Piepers, L. J. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-
group specific association between CXCR1 polymorphisms and subclinical mastitis in
dairy heifers. J. Dairy Res. 79:341-351.
Wanner, J. M., G. W. Rogers, M. E. Kehrli, J. B. Cooper. 1998. Intramammary infections in
primiparous Holsteins: heritabilities and comparisons of bovine leukocyte adhesion
deficiency carriers and noncarriers. J. Dairy Sci. 81: 3293-3299.

68

69
Chapter 3.2
Reactive oxygen species generation of bovine blood
neutrophils with different CXCR1 (IL8RA) genotype
following Interleukin-8 incubation
J. Verbeke1, X. Boulougouris2, C. Rogiers2, C. Burvenich2, L. Peelman3, B. De
Spiegeleer4, S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Comparative Physiology and Biometrics, Faculty of Veterinary Medicine,
Ghent University, Belgium 3Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine, Ghent
University, Belgium 4Department of Pharmaceutical Analysis, Faculty of Pharmaceutical Sciences, Ghent
University, Belgium
BMC Veterinary Research, submitted.

Chapter 3.2 The CXCR1 gene and neutrophil activity
70

Chapter 3.2 The CXCR1 gene and neutrophil activity
71
Abstract
Associations between polymorphisms in the bovine CXCR1 gene, encoding the
chemokine (C-X-C motif) receptor 1 (IL8RA), and udder health and milk neutrophil viability
have been described. In the present study, blood neutrophils were isolated from twenty
early lactating heifers with a different CXCR1 genotype at position 735 or 980, incubated
with different concentrations of recombinant bovine IL-8 (rbIL-8) for 2 or 6 h and stimulated
with phorbol 12-myristate 13-acetate (PMA) or opsonized zymosan particles (OZP).
Associations between CXCR1 genotype and the reactive oxygen species (ROS) generation,
measured by luminol chemiluminescence, were analyzed. Although single nucleotide
polymorphisms (SNPs) potentially affect CXCR1 functionality, SNPs c.735C>G and c.980A>G
were neither associated with ROS generation in general nor with the effect of rbIL-8
incubation on ROS generation. However, neutrophils incubated with rbIL-8 for 2 or 6 h
showed higher total PMA induced ROS generation but lower total OZP induced ROS
generation compared to neutrophils incubated without rbIL-8. Results indicate inhibitory
effects of IL-8 on ROS generation in bovine neutrophils and provide a foundation for future
research.
Key words
Bovine neutrophil, reactive oxygen species generation, CXCR1 polymorphism, interleukin 8

Chapter 3.2 The CXCR1 gene and neutrophil activity
72
1. Introduction
Neutrophils play a major role in the mammary gland immunity (Paape et al., 2002).
Constantly, neutrophils migrate from blood to milk (Paape et al., 2002). Diapedesis induces
apoptosis and reduces the reactive oxygen species (ROS) capacity (Smits et al., 1999; Van
Oostveldt et al., 2002). In case of intramammary infection, an immune response is initiated
attracting new neutrophils and activating neutrophils at the site of infection (Rainard and
Riollet, 2006). Within a couple of hours, milk neutrophil concentration climbs (Bannerman et
al., 2004; Mehrzad et al., 2001a). Activated neutrophils can kill mastitis pathogens by
generation of reactive oxygen species (ROS) (Heyneman et al., 1990; Mehrzad et al., 2005;
Mehrzad et al., 2009). Among other cytokines, interleukin 8 (IL-8) is locally produced by
macrophages, neutrophils and other immune cells (Boudjellab et al., 1998; Rainard and
Riollet, 2006; Sohn et al., 2007). The released IL-8 has a pleiotropic effect, as it was found to
enhance ROS generation (Daniels et al., 1992; Mitchell et al., 2003; Wozniak et al., 1993),
cause chemotaxis (Barber and Yang, 1998) and delay apoptosis (Kettritz et al., 1998) of
neutrophils. Studies using selective antagonists showed this activation to be mediated
through CXCR1 (IL8RA) and not through CXCR2 (IL8RB) in human neutrophils (Jones et al.,
1996; Jones et al., 1997).
Despite its important function, many polymorphisms were detected in the coding
region of the bovine CXCR1 gene (Pighetti et al., 2012; Verbeke et al., 2012). Single
nucleotide polymorphism (SNP) c.735C>G (dbSNP ID: rs208795699) causes an amino acid
change in the third intracellular loop (p.His245Glu) potentially affecting G-protein binding
and signal transduction. Furthermore, c.735C>G was found to be in full linkage
disequilibrium with SNPs c.37A>T (rs380621468), c.38T>A (rs110296731) and c.68G>A
(rs133273369) causing amino acid changes p.Ile13Tyr and p.Gly23Glu in the N-terminus of
CXCR1 known to have an important role in the first steps of binding IL-8 (Liou et al., 2014;
Verbeke et al., 2012). Associations between SNP c.735C>G and neutrophil functionality have
been studied: blood neutrophils with genotype c.735CC showed a lower intracellular calcium
release when stimulated with IL-8 and a reduced ROS generation in response to phorbol 12-
myristate 13-acetate (PMA) compared to neutrophils with genotype c.735GG (Pighetti and
Elliott, 2011). Single nucleotide polymorphisms c.980A>G (rs43323012) and c.995A>G
(rs43323013) cause changes in the C-terminus (p.Lys327Arg and p.His332Arg) and might

Chapter 3.2 The CXCR1 gene and neutrophil activity
73
interfere with adaptin-2 binding and receptor internalization (Pighetti et al., 2012). Recently,
we demonstrated a higher % milk neutrophil apoptosis in quarters from heifers with
genotype c.980GG compared to quarters from heifers with genotype c.980AG (Verbeke et
al., 2014a).
To further study effects of SNPs c.735C>G and c.980A>G on neutrophil functionality,
we isolated blood neutrophils from 20 early lactating heifers with different CXCR1 genotype
and analyzed their ROS production following IL-8 incubation and stimulation with PMA or
opsonized zymosan particles (OZP). We used blood neutrophils instead of milk neutrophils in
our model to have equal viability in all samples (Mehrzad et al., 2002). Freshly calved heifers
were sampled because neutrophil functionality is reduced in this stage of lactation (Mehrzad
et al., 2001b) and previous research indicated an association between SNP c.980A>G and
likelihood of intramammary infection by major pathogens in early lactating heifers (Verbeke
et al., 2012).
We report the results of an association study between CXCR1 SNPs c.735C>G and
c.980A>G and neutrophil ROS. Additionally, we discuss the unexpected effect rbIL-8 had on
neutrophil ROS depending on the stimulatory agent.
2. Materials and methods
2.1 Study design
The experiment has been approved by the ethical committee of the Faculty of
Veterinary Medicine, Ghent University (EC2013/190). Twenty Holstein heifers with different
CXCR1 genotype were included from 5 different commercial dairy herds. Selected heifers
were not siblings, had no history of diseases and all quarters were culture negative for major
mastitis pathogens. Within 24 h after calving, neutrophils were isolated from blood and
incubated with 0, 40 or 400 ng/ml recombinant bovine IL-8 (rbIL-8) for 2 and 6h. Next,
neutrophils were stimulated with PMA or OZP and ROS generation was measured by
chemiluminescence. Finally associations between ROS generation and genotype, incubation
time and rbIL-8 concentration were statistically analyzed. The sample size (n = 20) was based
on previous research demonstrating significant differences in ROS generation between 10
early and 10 mid lactating cows (Mehrzad et al., 2001b). The incubation times were
determined in a preliminary experiment in which blood neutrophils from 2 early lactating

Chapter 3.2 The CXCR1 gene and neutrophil activity
74
heifers were incubated for 2, 4, 6 and 18h with 0, 40 or 400 ng/ml rbIL-8. A differential count
of the isolated cells was performed to estimate the % neutrophils. Viability of neutrophils
was measured after isolation and after each incubation time by trypan blue exclusion.
2.2 Bacteriological culture
As mastitis can affect functionality of blood neutrophils (Mehrzad et al., 2001a),
aseptic quarter milk samples were collected at the time of blood sampling and
bacteriologically cultured. Ten μL of each sample was spread on blood-esculin and
MacConkey’s agar and incubated aerobically for 24-48 h at 37 °C. Bacteriological culture was
performed according to National Mastitis Council (NMC) guidelines (National Mastitis
Council, 1999). Four heifers were culture-positive in five quarters for major pathogens and
excluded from the analysis. Staphylococcus aureus, esculin-positive cocci and Escherichia coli
were isolated from 2 quarters of 1 heifers, 1 quarter of 1 heifer and 2 quarters of 2 heifers,
respectively.
2.3 CXCR1 genotype
To include heifers with common and rare CXCR1 genotypes (e.g. c.980AA), a sufficient
number of heifers were genotyped before calving. A blood sample was taken from 60
Holstein heifers belonging to 5 herds and having an expected calving date between January
and June 2014. Genotype at SNPs c.735C>G and c.980A>G was determined using a
fluorescent multiprobe PCR assay as previously described (Verbeke et al., 2014b). Efforts
were made to include sufficient heifers with genotype c.980AA or c.980AG. Of the 20 heifers
included in the final analysis, 7, 6 and 7 had genotype c.735CC, c.735CG and c.735GG,
respectively. Three, 5 and 12 had genotype c.980AA, c.980AG and c.980GG, respectively.
2.4 Reactive oxygen species assay
Seventy-five mL blood was collected from the coccygeal vein using 8 mL Vacutainer
tubes (Becton Dickinson, Erembodegem, Belgium) containing 150 μL of EDTA as
anticoagulant. Blood neutrophils were isolated within 1 h of collection by hypotonic lysis of
red blood cells and Histopaque 1077/1119 gradient (Sigma-Aldrich, Bornem, Belgium)

Chapter 3.2 The CXCR1 gene and neutrophil activity
75
centrifugation according to Siemens et al. (Siemens et al., 2007). Cell concentration was
measured in triplicate with a Bürker chamber.
Two hundred thousand blood neutrophils were suspended in 200 μL of 1x Hank's
balanced salt solution (Gibco, Life technologies, Carlsbad, CA) supplemented with 0, 40 or
400 ng recombinant bovine IL-8 (rbIL-8; Kingfisher Biotech, Saint Paul, MN) per mL and
incubated for 2 or 6 h at 37 °C in 2 mL test tubes. Next, blood neutrophils were pelleted by
centrifugation at 1000 x g for 5 min and resuspended in 120 μl 1x Hank's balanced salt
solution. Luminol (0.30 mmol/L Sigma-Aldrich) and PMA (100 ng/mL; Sigma-Aldrich) or OZP
(750 μg/mL) were added to a final volume of 200 μL. Zymosan A (Sigma-Aldrich) was
opsonized by washing the pellet with 60 and 30 mL 1 x phosphate-buffered saline (PBS;
Gibco) (centrifugation at 200 x g for 10 min) followed by 1 h incubation at 37 °C in 5 mL 1 x
PBS and 35 mL bovine serum and two additional washing steps with 30 mL 1x PBS
(centrifugation at 200 x g for 10 min). Bovine serum was collected from the coccygeal vein of
a healthy Holstein cow using 8 mL gel and clot activator tubes (Vacutest Kima, Piove di
Sacco, Italy). Reactions of blood neutrophils primed by rbIL-8 were performed in duplicate.
Chemiluminescence was measured every 60 sec for 90 min with a luminometer (TriStar² LB
942 Multidetection Microplate Reader, Berthold Technologies, Bad Wildbad, Germany) and
expressed in relative light units (RLU). Area under the curve (AUC) values (in 106 RLU * s)
were calculated to analyze the total ROS generation whereas peak values (RLUmax; in 103
RLU) and time of peak values (Tmax; in min) were saved in the dataset to study the kinetics
of ROS generation (Mehrzad et al., 2005).
2.5 Statistical analysis
Different linear mixed regression models (PROC MIXED, SAS 9.4, SAS Institute Inc.)
were fit for AUC, RLUmax and Tmax after stimulation with PMA or OZP (6 outcome variables)
and for SNP c.735C>G or c.980A>G (12 models in total). Heifer was added as random effect
to correct for clustering of multiple observations (6) per heifer (RANDOM statement). The
models included heifers’ genotype at position of the SNP, incubation (2 or 6 h) and rbIL-8 (0,
40 or 400 ng/ml) as categorical fixed effects. All two-way interactions were tested and kept
in the model if significant (P < 0.05).

Chapter 3.2 The CXCR1 gene and neutrophil activity
76
3. Results
3.1 Preliminary experiment
A differential count demonstrated 94.3% (± 0.1 %) of the isolated cells to be
neutrophils. The viability of the neutrophils was 100 % immediately after isolation and
decreased to 96 % (± 1 %), 96 % (± 2 %), 98 % (± 1 %) and 81 % (± 3 %) after 2, 4, 6 and 18 h
isolation, respectively. Differences in viability between neutrophils incubated with 0, 40 or
400 ng/mL rbIL-8 were small (data not shown). Because of the low viability and strongly
diminished ROS generation after 18 h incubation, neutrophils were incubated for 2 and 6 h
(Figure 3.2.1). As expected (Lieberman et al., 1996), chemiluminescence increased fast after
PMA stimulation with a clear peak and increased more gradually after OZP stimulation
(Figure 3.2.1).
3.2 Associations with ROS generation after PMA stimulation
All two-way interactions between fixed effects were non-significant and removed from
the models. Single nucleotide polymorphisms c.735C>G and c.980A>G were not associated
with AUC, RLUmax or Tmax (P > 0.05). Incubation was associated with AUC, RLUmax and
Tmax. Blood neutrophils incubated for 6 h showed higher AUC, RLUmax and Tmax values
compared to blood neutrophils incubated for 2 h (P < 0.05). Concentration of rbIL-8 was
associated with AUC and Tmax (P <0.01) and not with RLUmax (P = 0.17). Blood neutrophils
incubated with 40 or 400 ng/mL showed higher AUC and Tmax values compared to blood
neutrophils incubated without rbIL-8 (Table 3.2.1).
3.3 Associations with ROS generation after OZP stimulation
All two-way interactions between fixed effects were non-significant and removed from
the models. Single nucleotide polymorphisms c.735C>G and c.980A>G were not associated
with AUC, RLUmax or Tmax (P > 0.05). Incubation was associated with AUC and RLUmax and
not with Tmax. Blood neutrophils incubated for 6 h showed higher AUC and RLUmax values
compared to blood neutrophils incubated for 2 h (P < 0.05). Concentration of rbIL-8 was
associated with AUC and RLUmax (P <0.01) and not with Tmax (P = 0.89). Blood neutrophils
incubated with 40 or 400 ng/ml showed lower AUC and Tmax values compared to blood
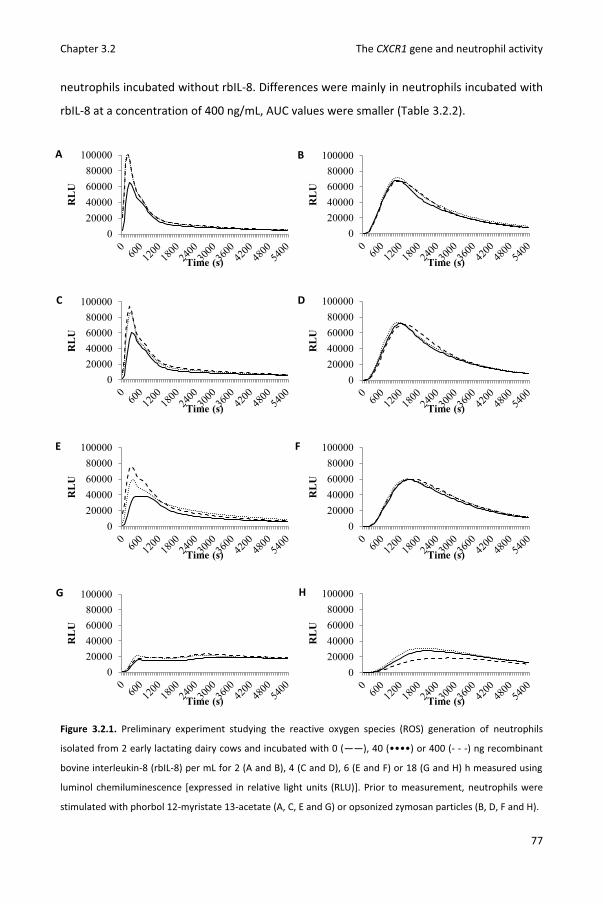
Chapter 3.2 The CXCR1 gene and neutrophil activity
77
neutrophils incubated without rbIL-8. Differences were mainly in neutrophils incubated with
rbIL-8 at a concentration of 400 ng/mL, AUC values were smaller (Table 3.2.2).
Figure 3.2.1. Preliminary experiment studying the reactive oxygen species (ROS) generation of neutrophils
isolated from 2 early lactating dairy cows and incubated with 0 (——), 40 (••••) or 400 (- - -) ng recombinant
bovine interleukin-8 (rbIL-8) per mL for 2 (A and B), 4 (C and D), 6 (E and F) or 18 (G and H) h measured using
luminol chemiluminescence [expressed in relative light units (RLU)]. Prior to measurement, neutrophils were
stimulated with phorbol 12-myristate 13-acetate (A, C, E and G) or opsonized zymosan particles (B, D, F and H).
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000R
LU
Time (s)
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000
RL
U
Time (s)
020000400006000080000
100000
RL
U
Time (s)
A B
C D
E F
G H
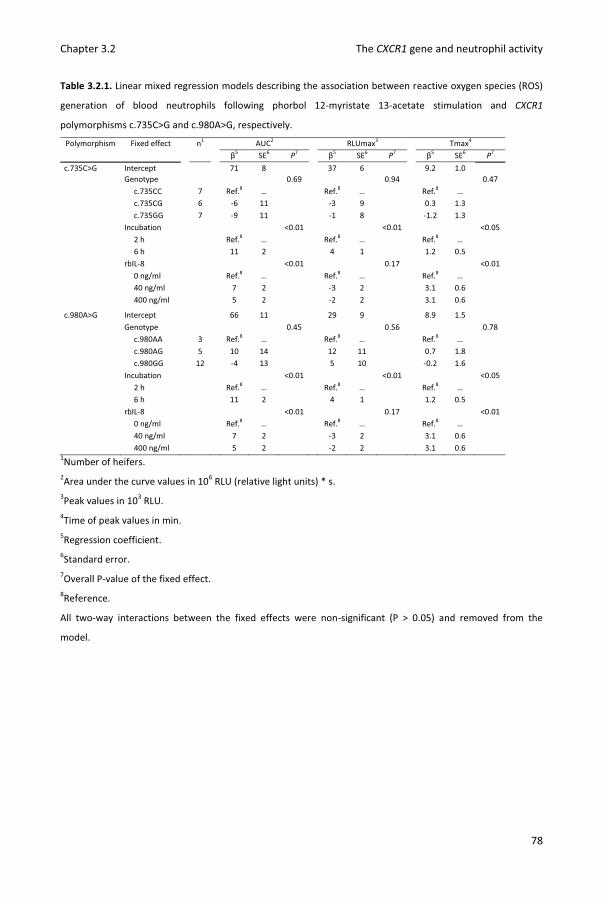
Chapter 3.2 The CXCR1 gene and neutrophil activity
78
Table 3.2.1. Linear mixed regression models describing the association between reactive oxygen species (ROS)
generation of blood neutrophils following phorbol 12-myristate 13-acetate stimulation and CXCR1
polymorphisms c.735C>G and c.980A>G, respectively.
Polymorphism Fixed effect n1 AUC2 RLUmax3 Tmax4 β5 SE6 P7 β5 SE6 P7 β5 SE6 P7
c.735C>G Intercept 71 8 37 6 9.2 1.0 Genotype 0.69 0.94 0.47
c.735CC 7 Ref.8 … Ref.8 … Ref.8 … c.735CG 6 -6 11 -3 9 0.3 1.3 c.735GG 7 -9 11 -1 8 -1.2 1.3 Incubation <0.01 <0.01 <0.05 2 h Ref.8 … Ref.8 … Ref.8 … 6 h 11 2 4 1 1.2 0.5 rbIL-8 <0.01 0.17 <0.01 0 ng/ml Ref.8 … Ref.8 … Ref.8 … 40 ng/ml 7 2 -3 2 3.1 0.6 400 ng/ml 5 2 -2 2 3.1 0.6
c.980A>G Intercept 66 11 29 9 8.9 1.5 Genotype 0.45 0.56 0.78 c.980AA 3 Ref.8 … Ref.8 … Ref.8 … c.980AG 5 10 14 12 11 0.7 1.8 c.980GG 12 -4 13 5 10 -0.2 1.6 Incubation <0.01 <0.01 <0.05 2 h Ref.8 … Ref.8 … Ref.8 … 6 h 11 2 4 1 1.2 0.5 rbIL-8 <0.01 0.17 <0.01 0 ng/ml Ref.8 … Ref.8 … Ref.8 … 40 ng/ml 7 2 -3 2 3.1 0.6 400 ng/ml 5 2 -2 2 3.1 0.6
1Number of heifers. 2Area under the curve values in 106 RLU (relative light units) * s. 3Peak values in 103 RLU. 4Time of peak values in min. 5Regression coefficient. 6Standard error. 7Overall P-value of the fixed effect. 8Reference.
All two-way interactions between the fixed effects were non-significant (P > 0.05) and removed from the
model.
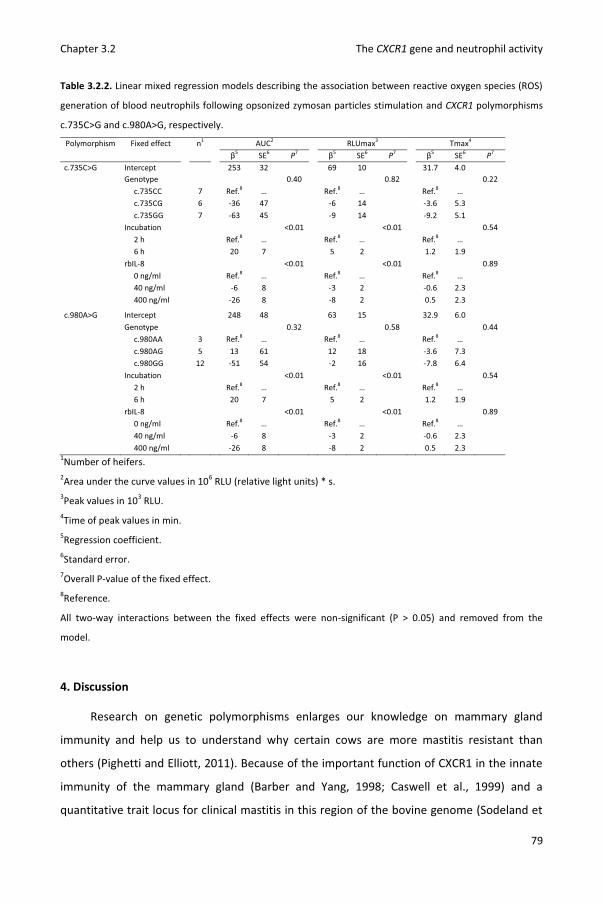
Chapter 3.2 The CXCR1 gene and neutrophil activity
79
Table 3.2.2. Linear mixed regression models describing the association between reactive oxygen species (ROS)
generation of blood neutrophils following opsonized zymosan particles stimulation and CXCR1 polymorphisms
c.735C>G and c.980A>G, respectively.
Polymorphism Fixed effect n1 AUC2 RLUmax3 Tmax4 β5 SE6 P7 β5 SE6 P7 β5 SE6 P7
c.735C>G Intercept 253 32 69 10 31.7 4.0 Genotype 0.40 0.82 0.22
c.735CC 7 Ref.8 … Ref.8 … Ref.8 … c.735CG 6 -36 47 -6 14 -3.6 5.3 c.735GG 7 -63 45 -9 14 -9.2 5.1 Incubation <0.01 <0.01 0.54 2 h Ref.8 … Ref.8 … Ref.8 … 6 h 20 7 5 2 1.2 1.9 rbIL-8 <0.01 <0.01 0.89 0 ng/ml Ref.8 … Ref.8 … Ref.8 … 40 ng/ml -6 8 -3 2 -0.6 2.3 400 ng/ml -26 8 -8 2 0.5 2.3
c.980A>G Intercept 248 48 63 15 32.9 6.0 Genotype 0.32 0.58 0.44 c.980AA 3 Ref.8 … Ref.8 … Ref.8 … c.980AG 5 13 61 12 18 -3.6 7.3 c.980GG 12 -51 54 -2 16 -7.8 6.4 Incubation <0.01 <0.01 0.54 2 h Ref.8 … Ref.8 … Ref.8 … 6 h 20 7 5 2 1.2 1.9 rbIL-8 <0.01 <0.01 0.89 0 ng/ml Ref.8 … Ref.8 … Ref.8 … 40 ng/ml -6 8 -3 2 -0.6 2.3 400 ng/ml -26 8 -8 2 0.5 2.3
1Number of heifers. 2Area under the curve values in 106 RLU (relative light units) * s. 3Peak values in 103 RLU. 4Time of peak values in min. 5Regression coefficient. 6Standard error. 7Overall P-value of the fixed effect. 8Reference.
All two-way interactions between the fixed effects were non-significant (P > 0.05) and removed from the
model.
4. Discussion
Research on genetic polymorphisms enlarges our knowledge on mammary gland
immunity and help us to understand why certain cows are more mastitis resistant than
others (Pighetti and Elliott, 2011). Because of the important function of CXCR1 in the innate
immunity of the mammary gland (Barber and Yang, 1998; Caswell et al., 1999) and a
quantitative trait locus for clinical mastitis in this region of the bovine genome (Sodeland et

Chapter 3.2 The CXCR1 gene and neutrophil activity
80
al., 2011), CXCR1 polymorphisms form interesting study objects. In this study, an in vitro
model was used to analyze the effect of CXCR1 SNP on neutrophil functionality in a sample
population of freshly calved heifers. Associations between CXCR1 genotype and neutrophil
ROS generation after rbIL-8 incubation and stimulation with PMA or OZP were studied in
detail.
The association between SNP c.735C>G and PMA induced ROS generation reported by
Rambeaud et al. (2006) could not be confirmed in our model. In contrast to the previously
demonstrated higher ROS generation (Rambeaud et al., 2006), we observed lower AUC
values in c.735GG neutrophils compared to c.735CC neutrophils. Additionally, no significant
interaction effects between c.735C>G and rbIL-8 concentration were observed. Based on
previous research (Verbeke et al., 2012; Verbeke et al., 2014b), we hypothesized a higher
ROS generation and response to rbIL-8 in c.980AG compared to c.980GG neutrophils.
However, neither c.980A>G nor the interaction between rbIL-8 concentration and c.980A>G
were associated with AUC, RLUmax or Tmax of PMA or OZP induced ROS generation in our
model. Despite potential effects on ligand binding, signal transduction and internalization of
CXCR1 (Pighetti et al., 2012), no interaction between CXCR1 SNPs and rbIL-8 incubation
could be demonstrated. Human CXCR1 but not hCXCR2 was found to be important for
activation of ROS (Stillie et al., 2009). Latter functional differences were partly attributed to
amino acid sequence differences in the C-terminus causing a faster receptor phosphorylation
and internalization of hCXCR2 compared to hCXCR1 (Nasser et al., 2005; Richardson et al.,
1998; Richardson et al., 2003). In contrast to hCXCR1 and hCXCR2, the C-terminus of both
bovine IL-8R show identical amino acid sequences (Lahouassa et al., 2008). Hence, activation
of neutrophils could be mediated by both receptors and functional effects caused by CXCR1
SNP might be compensated by a fully functional CXCR2 explaining similar rbIL-8 responses on
ROS across CXCR1 genotypes in our study.
Several studies demonstrated a priming effect of IL-8 on ROS generation in human
neutrophils (Daniels et al., 1992; Jones et al., 1996; Wozniak et al., 1993). Enhancement of
ROS generation already occurred after 5 min of IL-8 incubation (Daniels et al., 1992) and was
explained by intracellular calcium mobilization (Wozniak et al., 1993) and by activation of
phospholipase D (Jones et al., 1996), protein kinase C-ε (Nasser et al., 2005) and
phospholipase A2 (Daniels et al., 1992). To the best of our knowledge, Mitchell et al. (2003)
were the only to study the priming effect of IL-8 on bovine neutrophils. Intracellular ROS

Chapter 3.2 The CXCR1 gene and neutrophil activity
81
generation was measured by 2,7-Dichlorodihydrofluorecein diacetate (H2DCFDA) flow
cytometry. Incubation with rbIL-8 for 18h enhanced the Mannheimia haemolytica-induced
ROS generation whereas incubation for 30 or 60 min had little effect (Mitchell et al., 2003).
In our study, ROS generation was measured using luminol chemiluminescence. This assay
has the benefit that both intra- and extracellular ROS generation are measured and allows
for measurement over time (Rinaldi et al., 2007). Blood neutrophils were exposed to the
same concentrations of rbIL-8 as in the study of Mitchell et al. (2003) but for only 2 or 6 h.
We opted not to incubate for 18 h because our preliminary experiments showed a reduced
viability and a strongly diminished ROS generation after such a long period. Associations
were detected between rbIL-8 concentration and ROS generation indicating the presence of
a functional IL-8 receptor on the isolated blood neutrophils. In contrast to research on
human neutrophils (Daniels et al., 1992; Wozniak et al., 1993), IL-8 also had inhibitory effects
on neutrophil ROS generation in our model. Incubation with rbIL-8 had a positive effect on
the total PMA induced ROS generation but a negative effect on the total OZP induced ROS
generation. The stimulatory agent dependend effect could be explained by differences in the
pathways of ROS generation by PMA and OZP (Maridonneau-Parini et al., 1986). Incubation
with rbIL-8 might have simultaneously activated components of the pathway induced by
PMA (e.g. protein kinase C) while inhibiting components of the pathway induced by OZP (e.g.
calcium mobilization).
5. Conclusions
In conclusion, no differences in PMA or OZP induced ROS generation were detected in
blood neutrophils isolated from early lactating heifers with different CXCR1 c.735C>G and
c.980A>G genotypes. The inhibitory effects of rbIL-8 on neutrophil ROS generation suggest a
more complex interaction between IL-8 and ROS generation in bovine neutrophils than
initially expected and provides a foundation for future research.
6. Acknowledgements
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen). The authors wish to thank the
participating dairy producers and bovine practitioner Raf Deconinck.

Chapter 3.2 The CXCR1 gene and neutrophil activity
82
7. References
Bannerman, D. D., M. J. Paape, J. W. Lee, X. Zhao, J. C. Hope, and P. Rainard. 2004.
Escherichia coli and Staphylococcus aureus elicit differential innate immune responses
following intramammary infection. Clin. Diagn. Lab Immunol. 11(3):463-472.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab Immunol. 5(1):82-86.
Boudjellab, N., H. S. Chan-Tang, X. Li, and X. Zhao. 1998. Interleukin 8 response by bovine
mammary epithelial cells to lipopolysaccharide stimulation. Am. J. Vet. Res. 59:1563-
1567.
Caswell, J. L., D. M. Middleton, and J. R. Gordon. 1999. Production and functional
characterization of recombinant bovine interleukin-8 as a specific neutrophil activator
and chemoattractant. Vet. Immunol. Immunopathol. 67:327-340.
Daniels, R. H., M. J. Finnen, M. E. Hill, and J. M. Lackie. 1992. Recombinant human monocyte
Il-8 primes NADPH-oxidase and phospholipase-A2 activation in human neutrophils.
Immunology 75:157-163.
Heyneman, R., C. Burvenich, and R. Vercauteren. 1990. Interaction between the respiratory
burst activity of neutrophil leukocytes and experimentally induced Escherichia coli
mastitis in cows. J. Dairy Sci. 73:985-994.
Jones, S. A., M. Wolf, S. X. Qin, C. R. Mackay, and M. Baggiolini. 1996. Different functions for
the interleukin 8 receptors (IL-8R) of human neutrophil leukocytes: NADPH oxidase and
phospholipase D are activated through IL-8R1 but not IL-8R2. Proc. Natl. Acad. Sci. U. S.
A. 93:6682-6686.
Jones, S. A., B. Dewald, I. ClarkLewis, and M. Baggiolini. 1997. Chemokine antagonists that
discriminate between interleukin-8 receptors - Selective blockers of CXCR2. Journal of
Biological Chemistry 272:16166-16169.
Kettritz, R., M. L. Gaido, H. Haller, F. C. Luft, C. J. Jennette, and R. J. Falk. 1998. Interleukin-8
delays spontaneous and tumor necrosis factor-alpha-mediated apoptosis of human
neutrophils. Kidney Int. 53:84-91.
Lahouassa, H., P. Rainard, A. Caraty, and C. Riollet. 2008. Identification and characterization
of a new interleukin-8 receptor in bovine species. Mol. Immunol. 45:1153-1164.

Chapter 3.2 The CXCR1 gene and neutrophil activity
83
Lieberman, M. M., D. M. Sachanandani, and C. A. Pinney. 1996. Comparative study of
neutrophil activation by chemiluminescence and flow cytometry. Clin. Diagn. Lab
Immunol. 3:654-662.
Liou, J. W., F. T. Chang, Y. Chung, W. Y. Chen, W. B. Fischer, and H. J. Hsu. 2014. In Silico
analysis reveals sequential interactions and protein conformational changes during the
binding of chemokine CXCL-8 to its receptor CXCR1. Plos One 9.
Maridonneau-Parini, I., S. M. Tringale, and A. I. Tauber. 1986. Identification of distinct
activation pathways of the human neutrophil NADPH-oxidase. J. Immunol. 137:2925-
2929.
Mehrzad, J., H. Dosogne, E. Meyer, and C. Burvenich. 2001a. Local and systemic effects of
endotoxin mastitis on the chemiluminescence of milk and blood neutrophils in dairy
cows. Vet. Res. 32:131-144.
Mehrzad, J., H. Dosogne, E. Meyer, R. Heyneman, and C. Burvenich. 2001b. Respiratory burst
activity of blood and milk neutrophils in dairy cows during different stages of lactation. J.
Dairy Res. 68:399-415.
Mehrzad, J., L. Duchateau, S. Pyorala, and C. Burvenich. 2002. Blood and milk neutrophil
chemiluminescence and viability in primiparous and pluriparous dairy cows during late
pregnancy, around parturition and early lactation. J. Dairy Sci. 85:3268-3276.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2005. High milk neutrophil chemiluminescence
limits the severity of bovine coliform mastitis. Vet. Res. 36:101-116.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2009. Phagocytic and bactericidal activity of
blood and milk-resident neutrophils against Staphylococcus aureus in primiparous and
multiparous cows during early lactation. Vet. Microbiol. 134:106-112.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
Nasser, M. W., R. J. Marjoram, S. L. Brown, and R. M. Richardson. 2005. Cross-desensitization
among CXCR1, CXCR2, and CCR5: Role of protein kinase C-epsilon. J. Immunol. 174:6927-
6933.
National Mastitis Council. 1999. Laboratory Handbook on Bovine Mastitis. National Mastitis
Council. Inc., Madison, WI.

Chapter 3.2 The CXCR1 gene and neutrophil activity
84
Paape, M., J. Mehrzad, X. Zhao, J. Detilleux, and C. Burvenich. 2002. Defense of the bovine
mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary. Gland. Biol.
Neoplasia. 7:109-121.
Pighetti, G. M., and A. A. Elliott. 2011. Gene Polymorphisms: The keys for marker assisted
selection and unraveling core regulatory pathways for mastitis resistance. J. Mammary.
Gland. Biol. Neoplasia. 16:421-432.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Rainard, P., and C. Riollet. 2006. Innate immunity of the bovine mammary gland. Vet. Res.
37:369-400.
Rambeaud, M., R. Clift, and G. M. Pighetti. 2006. Association of a bovine CXCR2 gene
polymorphism with neutrophil survival and killing ability. Vet. Immunol. Immunopathol.
111:231-238.
Richardson, R. M., R. J. Marjoram, L. S. Barak, and R. Snyderman. 2003. Role of the
cytoplasmic tails of CXCR1 and CXCR2 in mediating leukocyte migration, activation, and
regulation. J. Immunol. 170:2904-2911.
Richardson, R. M., B. C. Pridgen, B. Haribabu, H. Ali, and R. Snyderman. 1998. Differential
cross-regulation of the human chemokine receptors CXCR1 and CXCR2 - Evidence for
time-dependent signal generation. J. Biol. Chem. 273:23830-23836.
Rinaldi, M., P. Moroni, M. J. Paape, and D. D. Bannerman. 2007. Evaluation of assays for the
measurement of bovine neutrophil reactive oxygen species. Vet. Immunol.
Immunopathol. 115:107-125.
Siemens, D. W., I. A. Schepetkin, L. N. Kirpotina, B. Lei, and M. T. Quinn. 2007. Neutrophil
isolation from nonhuman species. Methods Mol. Biol. 138:21-34.
Smits, E., C. Burvenich, A. J. Guidry, R. Heyneman, and A. Massart-Leen. 1999. Diapedesis
across mammary epithelium reduces phagocytic and oxidative burst of bovine
neutrophils. Vet. Immunol. Immunopathol. 68:169-176.
Sodeland, M., M. P. Kent, H. G. Olsen, M. A. Opsal, M. Svendsen, E. Sehested, B. J. Hayes, and
S. Lien. 2011. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20
in Norwegian Red cattle. Anim Genet. 42:457-465.

Chapter 3.2 The CXCR1 gene and neutrophil activity
85
Sohn, E. J., M. J. Paape, E. E. Connor, D. D. Bannerman, R. H. Fetterer, and R. R. Peters. 2007.
Bacterial lipopolysaccharide stimulates bovine neutrophil production of TNF-alpha, IL-1
beta, IL-12 and IFN-gamma. Vet. Res. 38:809-818.
Stillie, R., S. M. Farooq, J. R. Gordon, and A. W. Stadnyk. 2009. The functional significance
behind expressing two IL-8 receptor types on PMN. J. Leukoc. Biol. 86:529-543.
Van Oostveldt, K., M. J. Paape, H. Dosogne, and C. Burvenich. 2002. Effect of apoptosis on
phagocytosis, respiratory burst and CD18 adhesion receptor expression of bovine
neutrophils. Domest. Anim. Endocrinol. 22:37-50.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-group
specific association between CXCR1 polymorphisms and subclinical mastitis in dairy
heifers. J. Dairy Res. 79:341-351.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2014a. Association of
CXCR1 polymorphisms with apoptosis, necrosis and concentration of milk neutrophils in
early lactating dairy heifers. Res. Vet. Sci. 97: 55-59.
Verbeke, J., M. Van Poucke, L. Peelman, S. Piepers, and S. De Vliegher. 2014b. Associations
between CXCR1 polymorphisms and pathogen-specific clinical mastitis, test-day somatic
cell count and test-day milk yield. J. Dairy Sci. In press.
Wozniak, A., W. H. Betts, G. A. Murphy, and M. Rokicinski. 1993. Interleukin-8 primes human
neutrophils for enhanced superoxide anion production. Immunology 79:608-615.

86

87
Chapter 4
The role of CXCR1 gene polymorphisms in subclinical
mastitis

88

89
Chapter 4.1
Pathogen-group specific association between CXCR1
polymorphisms and subclinical mastitis
in dairy heifers
J. Verbeke1, S. Piepers1, L. Peelman2, M. Van Poucke2, S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine, Ghent
University, Belgium
Journal of Dairy Research, 2012, 79: 341-351.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
90

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
91
Abstract
The chemokine (C-X-C motif) receptor 1 (CXCR1) gene encodes the homonymous
receptor for interleukin 8 (IL8) on neutrophils. Binding causes migration from blood to milk,
activation and prolonged survival of neutrophils, a crucial process in the innate immune
defense of the bovine mammary gland against invading mastitis causing pathogens. The
main objective of this study was to screen the entire coding region of the CXCR1 gene for
polymorphisms and to analyze their association with udder health of dairy heifers. One
hundred forty Belgian Holstein heifers originating from 20 commercial dairy farms were
genotyped by DNA sequencing. Detailed phenotypic data on udder health was available
including quarter bacteriological culture results and somatic cell count (SCC) in early
lactation and composite milk SCC during first lactation. In total, 16 polymorphisms (including
8 missense mutations) were detected. Polymorphism c.980A>G was associated with
pathogen-group specific IMI: heifers with genotype AG were less likely to have an IMI due to
major mastitis pathogens compared to heifers with genotype GG but did not have less IMI by
coagulase-negative staphylococci, so-called minor pathogens. CXCR1 genotype was neither
associated with quarter SCC in early lactation nor with composite SCC during lactation.
Although mastitis susceptibility is influenced by many factors, some genetic polymorphisms
potentially have major effects on udder health of heifers, as was shown here. These results
trigger us to further study the relationship between CXCR1 polymorphisms and mastitis
susceptibility in both observational and experimental trials.
Key words
CXCR1 polymorphisms; heifer mastitis; neutrophil

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
92
1. Introduction
At calving, dairy heifers are expected to initiate a healthy and highly productive first
lactation. Still, ample evidence exists that a large proportion of heifers suffers from mastitis,
an inflammation of the mammary gland mostly caused by bacterial intramammary infection
(IMI), even before first milking (De Vliegher et al., 2012). This disease can be accompanied
with local or systemic symptoms (clinical heifer mastitis) or without visible symptoms
(subclinical heifer mastitis). The latter is mostly detected by an increase in the concentration
of somatic cells, mainly leukocytes, in milk (somatic cell count, SCC) (Fox, 2009). Both clinical
and subclinical heifer mastitis threaten the future performances of heifers and consequently
farm profitability (Huijps et al., 2009), especially when so-called major mastitis pathogens
such as Staphylococcus aureus and Streptococcus uberis are involved (Piepers et al., 2010). In
contrast, the impact on udder health of the so-called minor pathogens such as the
coagulase-negative staphylococci, is limited.
Seeking for better prevention and control of heifer mastitis, multiple research groups
have put efforts into the identification of risk factors potentially associated with this disease.
As a result, risk factors at the farm (De Vliegher et al., 2004; Svensson et al., 2006), heifer (De
Vliegher et al., 2004; Myllys and Rautala, 1995; Piepers et al., 2011; Waage et al., 2001), and
quarter level (Kromker and Friedrich, 2009; Piepers et al., 2011) have been identified. Still,
these risk factors only explain part of the variation in the prevalence and incidence of
mastitis in (early) lactating heifers. In particular the variation in udder health between
recently calved heifers housed on the same herd and thus managed under identical
conditions is still remarkably high (De Vliegher et al., 2004; Piepers et al., 2011), suggesting a
role for genetics of these animals.
Part of the variation in susceptibility to IMI between heifers is genetically determined
(Nash et al., 2003; Wanner et al., 1998). Thus, selection of heifers that are less susceptible
might offer a durable and sustainable prevention and control of heifer mastitis, avoiding
extensive antimicrobial treatment. Identification of genetic polymorphisms linked with
(heifer) mastitis resistance would allow evaluation of genetic make-up of male and female
breeding animals for udder health, even before their offspring is born. For this reason,
genetic markers associated with udder health are needed (Ogorevc et al., 2009). In a recent

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
93
genome-wide association study, polymorphisms located near CXCR1 and CXCR2 were highly
associated with occurrence of mastitis (Sodeland et al., 2011).
Migration of neutrophils from peripheral blood to the milk is a key event in the immune
defense of the mammary gland against invading bacteria (Paape et al., 2000). Interleukin 8
(IL-8) is the main chemoattractant in this process (Barber and Yang, 1998; Caswell et al.,
1999) and binds to two receptors, namely chemokine (C-X-C motif) receptor 1 (CXCR1) and
chemokine (C-X-C motif) receptor 2 (CXCR2). Besides inducing and mediating chemotaxis,
binding IL-8 causes prolonged survival (Kettritz et al., 1998) and increases activity (Mitchell
et al., 2003) of neutrophils. Polymorphisms in CXCR1 might affect expression or functionality
of the receptor causing variability in the innate immune responses subsequent to the
contact with a mastitis pathogen. Until recently, 5 single nucleotide polymorphisms (SNP)
have been identified (Grosse et al., 1999; Pighetti and Rambeaud, 2006; Youngerman et al.,
2004b) of which some seem to explain some udder health variability (Beecher et al., 2010;
Galvao et al., 2011; Youngerman et al., 2004a). Very recently, 5 additional SNP in the coding
region and 25 SNP in non-coding regions were reported (Pighetti et al., 2012). It is
reasonable to believe that CXCR1 polymorphisms might play a key role in the host resistance
of (periparturient) heifers as well and explain part of the variation in prevalence and
incidence of heifer mastitis. Still, these polymorphisms have yet not been studied as
potential factors associated with heifer mastitis.
The objectives of this study were to screen the entire coding region of CXCR1 for
polymorphisms and to analyze potential (pathogen-group specific) associations between
CXCR1 polymorphisms and subclinical mastitis phenotype (quarter milk SCC in early
lactation, IMI status in early lactation, and composite milk SCC during first lactation) in
Belgian dairy heifers.
2. Materials and methods
2.1 Herds and heifers
The database for the current research consisted of phenotypic records (Piepers et al.,
2010) combined with data on the CXCR1 genotype. In total, records and blood samples of
140 Holstein heifers were available. Heifers were housed on 20 commercial dairy farms. On
average, 7 heifers per farm were included, ranging between 3 and 10. The Cattle Breeding

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
94
Association (CRV, Arnhem, The Netherlands) was consulted for data on the relationship of
heifers. Records of all but 2 heifers could be retrieved. Seventy-seven different fathers, 136
different mothers, 339 different grandparents, and 550 different great-grandparents were
noted, indicating that the heifers are not closely related to each other.
2.2 Data collection and sampling
As described previously (Piepers et al., 2010), quarter milk samples were aseptically
collected twice, between 1 and 4 days in milk (DIM) and between 5 and 8 DIM, for
determination of the quarter milk SCC (30 mL) and bacteriological culturing (5 mL) at each
sampling. Composite SCC throughout first lactation was determined on milk samples
collected as part of the dairy herd improvement (DHI) program of the CRV on a four- to six-
weekly basis. Records from 14 until 285 DIM were used.
As a source of DNA, blood samples were taken from the tail vein of all heifers and stored
in EDTA vacutainer tubes (Terumo) at -20°C.Phenotypic records.
2.3 Phenotypic data
Quarter and composite milk SCC was determined at the Milk Control Centre Flanders
(Lier, Belgium) using a Fossomatic 5000 (Foss Electric).
Bacteriological culture was based on National Mastitis Council standards and performed
at the Milk Control Centre Flanders (Lier, Belgium) as described by Piepers et al. (2007). In
short, 10 μl of milk was spread on blood-esculin agar (Oxoid) and on MacConkey’s agar
(Oxoid). In order to grow bacteria, plates were incubated aerobically for 24-48 h at 36-38°C.
Identification of bacteria was done by Gram-staining and inspection of the colony
morphology. Catalase tests were performed to differentiate Gram-positive cocci in catalase-
positive or catalase-negative cocci. Staphylococci were identified as coagulase-negative
staphylococci or S. aureus by colony morphology, hemolysis patterns, and DNase tests.
Streptococci were differentiated in esculine-positive and esculine-negative streptococci
(Streptococcus agalactiae and Streptococcus dysgalactiae). Streptococcus agalactiae and S.
dysgalactiae were distinguished using the Christie, Atkins, Munch-Petersen (CAMP) test.
Gram-negative bacteria were not further differentiated. Corynebacterium bovis and
coagulase-negative staphylococci were categorized as minor pathogens, while S. aureus,

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
95
esculine-positive streptococci, S. dysgalactiae and S. agalactiae were categorized as major
pathogens. Other than the abovementioned Gram-positive bacteria and Gram-negative
bacteria were classified as “other pathogens” and excluded from the analyses. Eight quarters
yielded Gram-negative bacteria, one was culture-positive for Trueperella pyogenes, and one
was culture-positive for both latter pathogens. In order to study subclinical mastitis only,
quarters reported to be clinically infected between 1 and 8 DIM were excluded from the
analysis (in total 6 cases, see further). Intramammary infection status of each quarter was
determined on the outcome of bacteriological culture of the 2 consecutive milk samples
(collected between 1- 4 DIM, and 5-8 DIM, respectively) (Piepers et al., 2011). In short, a
quarter was considered non-infected if no pathogens could be isolated from both samples. A
infection with coagulase-negative staphylococci was defined as isolation of coagulase-
negative staphylococci at the first sampling and isolation of coagulase-negative
staphylococci or no pathogen at the second sampling. A Corynebacterium bovis-infection
was defined as isolation of C. bovis at the first sampling and isolation of C. bovis or no
pathogen at the second sampling. A quarter was considered as infected with a major
pathogen if a major pathogen was isolated at the first sampling and if the same pathogen, a
minor pathogen or no pathogen was isolated at the second sampling. All quarters that could
not be allocated to one of those classes were considered as missing values.
2.4 CXCR1 genotyping and polymorphism screening
DNA was extracted from blood samples (100 μl) using a proteinase K digestion method
(Van Poucke et al., 2005) and its concentration was estimated with a ND spectrophotometer
(NanoDrop).
In order to amplify the whole single-exon coding region (1083 bp) of bovine CXCR1, a
forward PCR primer (5'-TCCTTGATGAGAGTGATTTGGA-3') binding 76-55 bp upstream the
start codon and a reverse PCR primer (5'-TTGACATGGGACTGTGAACG-3') binding 61-80 bp
downstream the stop codon was designed using Primer3Plus (Untergasser et al., 2007)
based on the reference sequence of CXCR1 [GenBank: NM_001105038.1]. Regions forming
potential secondary structures were identified with Mfold (Zuker, 2003) and avoided.
Specificity of binding of both primers was analyzed using NCBI BLAST (Altschul et al., 1990). A
PCR mix containing approximately 100 ng genomic DNA, 1.0 μl 10x FastStart Taq DNA

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
96
Polymerase Buffer (Roche Applied Science), dNTP Mix (0.2 mM each; BIOLINE), 0.25 μM
forward primer (Integrated DNA Technologies), 0.25 μM reverse primer (Integrated DNA
Technologies) and 0.5 U Taq DNA Polymerase (Roche Applied Science) was made.
The PCR program consisted of an initiation step of 5 min at 95°C followed by 30
amplification cycles (denaturation for 30 s at 95°C, annealing for 30 s at 62°C and extension
for 75 s at 72°C) and a final 4 min elongation step at 72°C. All amplicons were sequenced by
direct sequencing with the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied
Biosystems) on a 16-capillary 3130xl DNA Analyzer (Applied Biosystems) according to the
manufacturer’s protocol. Sequencing reactions per sample were performed in duplicate
using the forward and reverse primers.
Sequences of the coding region of CXCR1 of all samples were compared with the
reference sequence and with each other to identify polymorphisms. Four haplotypes were
identified based on homozygous genotypes, present in a number of animals. The genotype
of all but 2 heterozygous heifers could be explained by a combination of these haplotypes
(see further). The haplotypes of these 2 heifers were identified by cloning. First, the
amplicon generated by PCR of either of both alleles was cloned into a pCR 2.1 vector using
TA Cloning Kit (Invitrogen). Next, the vector was transformed in DH5a Competent Cells
(Invitrogen). Plasmid DNA from single colonies (Birnboim and Doly, 1979) was sequenced for
the whole CXCR1 coding region as previously described. Finally, the haplotype of the non-
ligated allele was deducted from this sequence and the heifer’s genotype. Single nucleotide
polymorphisms and haplotype sequences were submitted to the NCBI dbSNP and GenBank
database, respectively (Table 1, Table 2).
2.5 Statistical analysis
Deviations from Hardy-Weinberg equilibrium of each studied polymorphism were
analyzed using chi-square goodness of fit tests. Chi-square were calculated to analyze
linkage association between loci.
Associations with phenotype were only analyzed for polymorphisms c.642G>A, c.735C>G,
c.816C>A and c.980A>G, as all other detected polymorphisms were in strong linkage
disequilibrium (correlation > 95%) with either one of these SNP, and results would be similar
for that reason.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
97
The associations between the different polymorphisms and quarter milk SCC in early
lactation were determined using a linear mixed regression model with herd, heifer and
quarter as random effects to correct for clustering of heifers within herds, quarters within
heifers, and the two observations (measured between 1-4 DIM and 5-8 DIM, respectively)
per quarter (PROC MIXED, SAS 9.2, SAS Institute Inc.). A natural logarithmic transformation
of quarter milk SCC was performed to obtain a normalized distribution. The different models
included “observation” (1 = between 1-4 DIM, 2 = between 5-8 DIM, respectively) and the
different heifers’ genotypes (c.642G>A, c.735C>G, c.816C>A and c.980A>G, respectively) at
the position of the polymorphism as categorical fixed effects (PROC MIXED, SAS 9.2, SAS
Institute Inc.). Models excluding and including IMI status (0 = non-infected quarter, 1 =
quarter infected with coagulase-negative staphylococci and 2 = quarter infected with S.
aureus, S. agalactiae, S. dysgalactiae and esculine-positive streptococci, respectively) as
fixed effect were compared and the interaction between IMI status and CXCR1 genotype
were tested as well in the models including IMI status.
The associations between the different polymorphisms and presence of IMI in early
lactation were determined using a logistic mixed regression model with herd and heifer as
random effects to correct for clustering of heifers within herds and quarters within heifers
(MlwiN 2.02, Centre for Multilevel Modeling, Bristol, UK). Different models were fit
separately for three different binary outcome variables: (1) “coagulase-negative
staphylococci IMI” (0 = non-infected quarter versus 1 = quarter infected with coagulase-
negative staphylococci), (2) “Major pathogen IMI” (0 = non-infected quarter versus 1 =
quarter infected with S. aureus, S. agalactiae, S. dysgalactiae and esculine-positive
streptococci), and (3) “all IMI” (0 = non-infected quarter versus 1 = quarter infected with
coagulase-negative staphylococci, C. bovis, S. aureus, S. agalactiae, S. dysgalactiae or
esculine-positive streptococci). All models included heifers’ genotype at the position of the
polymorphism (c.642G>A, c.735C>G, c.816C>A and c.980A>G, respectively) as a categorical
fixed effect. To allow convergence of the statistic models, records on rare genotypes
(c.980AA, see further) were omitted.
The associations between the different polymorphisms and test-day composite SCC were
determined using a linear mixed regression model with herd and heifer as random effects to
correct for clustering of heifers within herds and multiple observations per heifers (PROC
MIXED, SAS 9.2, SAS Institute Inc.). A first order autoregressive correlation structure (AR1)

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
98
was used to model the multiple milk recordings within heifer. A natural logarithmic
transformation of composite SCC was performed to obtain a normalized distribution. The
model included DIM as continuous fixed effect and heifers’ genotype (c.642G>A, c.735C>G,
c.816C>A and c.980A>G, respectively) at the position of the polymorphism as categorical
fixed effect. In all models, the quadratic term for DIM and the interaction between DIM and
genotype were tested.
Statistical significance for all tests was assessed at P 0.05.
3. Results
3.1 Phenotypic data
On average, herds counted 48 lactating cows [Interquartile range (IQR) 27 – 56]
producing 8344 kg milk/year (IQR 7693 - 9470 kg milk/year) and had a herd milk SCC of
280,000 cells/ml (IQR 193,000 - 343,000 cells/ml) during the study period. Of the 140 heifers
selected for analysis, two heifers had a non-functional quarter at calving. Six quarters were
reported to be clinically infected between 1 and 8 DIM. The latter 8 quarters were omitted
from the analysis.
Geometric mean of quarter milk SCC of all quarters declined from 336,000 cells/mL (IQR
102,000-1013,000) between 1 and 4 DIM to 110,000 cells/mL (IQR 35,000-253,000) between
5 and 8 DIM (P < 0.0001). Quarters infected with S. aureus, S. agalactiae, S. dysgalactiae or
esculine-positive streptococci had higher geometric mean quarter milk SCC (1,011,000
cells/ml, IQR 277,000 – 4,151,000) than healthy quarters (119,000 cells/ml, IQR 44,000 -
255,000)(P < 0.0001). The geometric mean SCC of quarters infected with coagulase-negative
staphylococci was intermediate (341,000 cells/ml, IQR 57,000 – 498,000) with respect to
healthy quarters and quarters infected with a major pathogen.
The IMI status could be determined for 442 out of 552 (80%) functional, nonclinical
quarters from the 140 heifers. In total, 46% of 442 quarters (n = 204) and 9% of the heifers
(n = 13) were non-infected. Approximately 54% of the quarters (n = 238) and 76% of the
heifers (n = 106) were subclinically infected, the vast majority of which were coagulase-
negative staphylococci infections (84%, n = 201). Infections by major pathogens were
detected in 35 quarters of 24 heifers (15% of IMI). Esculine-positive streptococci were the
most frequently isolated pathogens (n = 20), followed by S. aureus (n = 14). One quarter was

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
99
infected with S. agalactiae, two were infected with C. bovis. No IMI status could be
determined in 20% of the quarters (n = 110) because of the definitions in use. Data on these
quarters were considered as missing values in the analysis. The main reason of exclusion was
a culture-negative result between 1 and 4 DIM combined with a culture-positive result
between 5 and 8 DIM (n = 59).
During lactation, the composite SCC followed a quadratic curve (DIM²: P < 0.01). The
geometric mean composite SCC was 92,000 cells/ml (IQR 35,000-174,000) between 14 and
45 DIM, declined to 57,000 cells/ml (IQR 25,000-97,000) between 76 and 105 DIM and rose
again till 94,000 cells/ml (IQR 38,000-199,000) between 256 and 285 DIM.
3.2 Polymorphism screening
In total, 16 polymorphisms were detected. Ten have been described in literature before
(Pighetti et al., 2012) including 6 silent mutations (c.291C>T, c.570G>A, c.816C>A, c.819G>A,
c.1008C>T, c.1068G>A) and 4 missense mutations (c.365T>C, c.735C>G, c.980A>G,
c.995A>G). Two novel silent polymorphisms (c.333T>C, c.642G>A) and 4 novel missense
polymorphisms (c.37A>T, c.38T>A, c.68G>A, c.337G>A) were discovered. The missense
mutations cause amino acid changes at 7 positions in the protein (p.Ile13Tyr, p.Gly23Glu,
p.Val113Ile, p.Val122Ala, p.His245Glu, p.Lys327Arg, p.His332Arg) (Table 4.1.1). All studied
polymorphisms were in strong linkage disequilibrium (P < 0.05). In the investigated
population, polymorphisms c.37A>T, c.38T>A, c.68G>A, c.735C>G and c.819G>A were in
complete linkage (linkage group 1). The same was true for polymorphisms c.333T>C,
c.337G>A, c.980A>G and c.995A>G (linkage group 2), polymorphisms c.291C>T and c.816C>A
(linkage group 3), and for polymorphisms c.365T>C, c.570G>A and c.1068G>A (linkage group
4, Table 4.1.1). The studied population was in Hardy-Weinberg equilibrium (P > 0.05) for all
polymorphisms. Five haplotypes were detected. Sequences, and frequencies are shown in
Table 4.1.2.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
100
Table 4.1.1. Polymorphisms detected in the coding region of CXCR1 of 140 Holstein-Friesian heifers from 20 Belgian dairy farms
SNP1 Amino acid change
Genotype frequencies (n = 140)2 Linkage group 13
Linkage group 24
Observed by5
dbSNP6
Homozygous allele 17
Heterozygous Homozygous allele 2
c.37A>T p.Ile13Tyr 36 (26) 67 (48) 37 (26) 1 V ss525192449 c.38T>A p.Ile13Tyr 36 (26) 67 (48) 37 (26) 1 V ss525192453 c.68G>A p.Gly23Glu 36 (26) 67 (48) 37 (26) 1 V ss525192456 c.291C>T 64 (46) 63 (45) 13 (9) 3 3 V/P ss525192459 c.333T>C 3 (2) 42 (30) 95 (68) 2 V ss525192462 c.337G>A p.Val113Ile 3 (2) 42 (30) 95 (68) 2 V ss525192464 c.365T>C p.Val122Ala 35 (25) 67 (48) 38 (27) 1 4 V/P ss525192466 c.570G>A 35 (25) 67 (48) 38 (27) 1 4 V/P ss525192469 c.621G>A p.Trp207Stop 140 (100) 0 (0) 0 (0) 4 P c.642G>A 66 (47) 64 (46) 10 (7) 5 V ss525192472 c.735C>G p.His245Glu 36 (26) 67 (48) 37 (26) 1 1 V/P ss525192475 c.816C>A 64 (46) 63 (45) 13 (9) 3 3 V/P ss525192477 c.819G>A 36 (26) 67 (48) 37 (26) 1 1 V/P ss525192479 c.980A>G p.Lys327Arg 3 (2) 42 (30) 95 (68) 2 2 V/P ss525192482 c.995A>G p.His332Arg 3 (2) 42 (30) 95 (68) 2 2 V/P ss525192485 c.1008C>T 63 (45) 63 (45) 14 (10) 3 6 V/P ss525192488 c.1068G>A 35 (25) 67 (48) 38 (27) 1 4 V/P ss525192669 1Locations are relative to the start codon. 2Numbers of the heifers in the study expressing the given genotype and their relative percentage in
parentheses. 3,4Linkage groups as observed by Pighetti et al. (2012) and in our study, respectively 5Polymorphisms detected in this study (V), by Pighetti et al. (2012) (P) or in both studies (V/P) 6Submitted SNP (ss) number in the NCBI dbSNP database (http://www.ncbi.nlm.nih.gov/snp) 7Allele with nucleotide(s) equal to the nucleotide(s) in the reference sequence [GenBank:NM_001105038.1]
Table 4.1.2. CXCR1 haplotype sequences and frequencies for 140 Holstein Friesian dairy heifers from 20 Belgian
dairy farms
Label 371 38 68 291 333 337 365 570 642 735 816 819 980 995 1008 1068 Freq. Accession2
H1 A T G C T G T G G C C G A A C G 0.16 HM013954
H2 A T G T C A T G G C A G G G T G 0.29 HM013955
H3 T A A C C A C A A G C A G G C A 0.36 HM013956
H4 T A A C C A C A G G C A G G C A 0.19 HM013957
H5 A T G C C A C A G C C G G G T A 0.01 JX050220
1Numbers indicate positions of the polymorphisms relative to the start codon. 2GenBank accession number.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
101
3.3 Association between polymorphisms in CXCR1 and subclinical mastitis
Descriptive statistics on subclinical mastitis by CXCR1 polymorphisms are given in Table
4.1.3. Neither in the models including IMI status as fixed effect nor in the models excluding
IMI status, polymorphisms were significantly associated with qLnSCC (Table 4.1.4). None of
the studied polymorphisms were associated with “All IMI” or “coagulase-negative
staphylococci IMI” (P > 0.05). However, a significant association between polymorphism
c.980A>G and “major pathogen IMI” was detected: quarters of heifers with genotype GG on
position 980 were more likely to be infected with major pathogens compared to quarters
from heifers with genotype AG (P < 0.05, Table 4.1.5). None of the investigated
polymorphisms (c.642G>A, c.735C>G, c.816C>A or c.980A>G) were associated with
composite SCC (Data not shown).

Chap
ter 4
.1
Th
e CX
CR1
gene
and
subc
linic
al h
eife
r mas
titis
102
Tabl
e 4.
1.3.
Des
crip
tive
stat
istic
s on
subc
linic
al m
astit
is by
CXC
R1 p
olym
orph
isms c
.642
G>A
, c.7
35C>
G, c
.816
C>A
and
c.98
0A>G
, res
pect
ivel
y
SNP
Geno
type
n
heife
rs
n qu
arte
rs
Early
lact
atio
n (1
-8 D
IM)
Firs
t lac
tatio
n
qu
arte
r int
ram
amm
ary
infe
ctio
n st
atus
quar
ter S
CC (x
1000
cel
ls/m
l)
com
posit
e SC
C (x
1000
cel
ls/m
l)
non-
infe
cted
(%
)1
n co
agul
ase-
nega
tive
stap
hylo
cocc
i (%
)2
n m
ajor
pa
thog
en
(%)3
n C.
bov
is (%
)4
1-4
DIM
(IQ
R)5
5-8
DIM
(IQ
R)6
14
-285
DIM
(IQ
R)7
GG
66
20
5
86 (4
2)
104
(52)
14
(5)
1 (1
)
316
(103
-854
) 11
6 (4
1-22
4)
76
(33-
139)
c.
642G
>A
GA
64
201
98
(49)
83
(42)
19
(10)
1
(0)
38
3 (1
11-1
231)
10
3 (3
0-25
6)
77
(32-
146)
AA
10
36
20
(56)
14
(15)
2
(0)
0 (0
)
216
(65-
403)
12
2 (3
6-32
5)
10
4 (4
6-16
2)
CC
36
111
49 (4
4)
56 (5
0)
5 (5
) 1
(1)
309
(111
-713
) 11
1 (4
2-19
8)
77 (3
3-14
6)
c.73
5C>G
CG
67
21
6 10
3 (4
8)
91 (4
2)
21 (1
0)
1 (0
) 37
2 (1
06-1
266)
10
6 (3
0-28
9)
75 (3
3-13
1)
GG
37
115
52 (4
5)
54(4
7)
9 (8
) 0
(0)
303
(89-
854)
11
7 (3
6-25
1)
85 (3
5-17
1)
CC
64
19
8
95 (4
8)
90 (4
5)
13 (7
) 0
(0)
32
7 (1
02-8
68)
111
(34-
273)
82 (3
5-15
4)
c.81
6C>A
CA
63
20
6
99 (4
8)
89 (4
3)
17 (8
) 1
(0)
32
8 (9
1-11
14)
98 (3
5-19
9)
67
(29-
115)
AA
13
38
10
(26)
22
(58)
5
(13)
1
(3)
43
0 (1
35-1
113)
18
3 (4
1-46
2)
99
(49-
244)
AA
3 7
6 (8
6)
1 (1
4)
0 (0
) 0
(0)
448
(107
-119
3)
141
(56-
235)
45
(18-
92)
c.98
0A>G
AG
42
13
5 65
(48)
67
(50)
3
(2)
0 (0
) 28
8 (1
08-5
99)
86 (3
1-20
4)
74 (3
0-14
1)
GG
95
30
0
133
(44)
13
3 (4
4)
32 (1
1)
2 (1
)
356
(102
-121
9)
122
(36-
276)
81 (3
5-15
1)
1 No
path
ogen
s iso
late
d.
2 IMI c
ause
d by
coa
gula
se-n
egat
ive
stap
hylo
cocc
i .
3 IMI c
ause
d by
Sta
phyl
ococ
cus a
ureu
s, S
trep
toco
ccus
aga
lact
iae,
Str
epto
cocc
us d
ysga
lact
iae
or e
scul
ine-
posit
ive
stre
ptoc
occi
. 4 IM
I cau
sed
by C
oryn
ebac
teriu
m b
ovis.
5,
6G
eom
etric
mea
n qu
arte
r milk
SCC
bet
wee
n 1
and
4 DI
M a
nd b
etw
een
5 an
d 8
DIM
, res
pect
ivel
y.
7 Geom
etric
mea
n co
mpo
site
milk
SCC
det
erm
ined
at t
est-
day
on a
four
- to
six-w
eekl
y ba
sis.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
103
Table 4.1.4. Association between quarter SCC in early lactation and CXCR1 polymorphisms c.642G>A, c.735C>G,
c.816C>A and c.980A>G, respectively
Polymorphism Independent variable Models1 excluding IMI status Models1 including IMI status β2 SE3 LSM4 P-value5 β SE LSM P-value c.642G>A Intercept 4.50 0.36 4.82 0.35 Genotype GG ref. … 5.26 0.80 ref. … 5.57 0.46 GA 0.06 0.20 5.32 0.22 0.18 5.80 AA -0.20 0.39 5.06 0.00 0.34 5.58 Sampling 1-4 DIM ref. … 5.77 <0.01 ref. … 6.20 <0.01 5-8 DIM -1.12 0.06 4.66 -1.11 0.07 5.10 IMI status6 non-infected … … … ref. … 4.85 <0.01 coagulase-negative staphylococci … … … 0.43 0.12 5.28 Major pathogen … … … 1.98 0.21 6.83
c.735C>G Intercept 4.70 0.19 6.33 0.26 Genotype CC ref. … 5.22 0.93 ref. … 5.58 0.81 CG 0.09 0.23 5.31 0.14 0.22 5.71 GG 0.03 0.27 5.26 0.13 0.24 5.70 Sampling 1-4 DIM ref. … 5.82 <0.01 ref. … 6.22 <0.01 5-8 DIM -1.12 0.06 4.70 -1.11 0.07 5.11 IMI status6 non-infected … … … ref. … 4.87 <0.01 coagulase-negative staphylococci … … … 0.63 0.12 5.29
Major pathogen … … … 1.97 0.21 6.84
c.816C>A Intercept 5.05 0.31 6.42 0.35 Genotype CC ref. … 5.27 0.50 ref. … 5.75 0.61 CA -0.06 0.20 5.20 -0.17 0.18 5.58 AA 0.34 0.35 5.61 0.04 0.33 5.79 Sampling 1-4 DIM ref. … 5.92 <0.01 ref. … 6.26 <0.01 5-8 DIM -1.12 0.06 4.80 -1.11 0.07 5.16 IMI status6 non-infected … … … ref. … 4.91 <0.01 coagulase-negative staphylococci … … … 0.42 0.12 5.33 Major pathogen … … … 1.98 0.21 6.89
c.980A>G Intercept 4.79 0.12 6.42 0.35 Genotype AA ref. … 5.52 0.44 ref. … 5.52 0.95 AG -0.43 0.66 5.08 0.13 0.73 5.65 GG -0.17 0.68 5.34 0.17 0.72 5.69 Sampling 1-4 DIM ref. … 5.87 <0.01 ref. … 6.17 <0.01 5-8 DIM -1.12 0.06 4.76 -1.11 0.07 5.07 IMI status6 non-infected … … … ref. … 4.82 <0.01 coagulase-negative staphylococci … … … 0.42 0.12 5.24
Major pathogen … … … 1.97 0.21 6.80 1The linear mixed regression models included herd, heifer and quarter as random effect. A natural logarithmic
transformation of quarter SCC (qLnSCC) was performed to obtain a normalized distribution. 2Regression coefficien. 3Standard error. 4Least squares mean. 5Overall P-value of the fixed effect. 6Intramammary infection (IMI) status; IMI caused by coagulase-negative staphylococci and IMI caused by major
pathogens Staphylococcus aureus, Streptococcus agalactiae, Streptococcus dysgalactiae and esculine-positive
streptococci versus non-infected.
The interaction term between CXCR1 genotype and IMI was not significant and therefore excluded from the
models.

Chap
ter 4
.1
Th
e CX
CR1
gene
and
subc
linic
al h
eife
r mas
titis
104
Tabl
e 4.
1.5.
Ass
ocia
tion
betw
een
path
ogen
-gro
up sp
ecifi
c in
tram
amm
ary
infe
ctio
n an
d CX
CR1
poly
mor
phism
s c.6
42G
>A, c
.735
C>G,
c.8
16C>
A an
d c.
980A
>G, r
espe
ctiv
ely
Mod
el1
Inde
pend
ent
Depe
nden
t var
iabl
es
varia
ble
Coag
ulas
e-ne
gativ
e st
aphy
loco
cci I
MI2
M
ajor
pat
hoge
n IM
I3
All I
MI4
β5 SE
6 O
R7 95
% C
I8 P-
valu
e9 β
SE
OR
95%
CI
P-va
lue
β SE
O
R 95
% C
I P-
valu
e c.
642G
>A
Inte
rcep
t 0.
17
0.21
-1
.75
0.37
0.
31
0.20
Ge
noty
pe10
0.
417
0.
909
0.
448
GG
re
f. …
…
…
re
f. …
…
…
re
f. …
…
…
GA
-0
.34
0.28
0.
71
0.41
-1.2
3
0.
14
0.49
1.
15
0.44
-3.0
0
-0
.29
0.27
0.
75
0.44
-1.2
8
AA
-0
.42
0.51
0.
66
0.24
-1.7
8
-0
.22
0.92
0.
80
0.13
-4.8
6
-0
.48
0.51
0.
62
0.23
-1.6
6
c.73
5C>G
In
terc
ept
0.07
0.
27
-2.2
7 0.
57
0.18
0.
27
Geno
type
10
0.71
0 0.
500
0.91
3
CC
re
f. …
…
…
re
f. …
…
…
re
f. …
…
…
CG
-0
.20
0.33
0.
82
0.43
-1.5
5 0.
77
0.65
2.
15
0.60
-7.7
2 -0
.10
0.32
0.
91
0.48
-1.7
1
GG
0.
04
0.37
0.
73
0.51
-2.1
3 0.
63
0.74
1.
87
0.44
-7.8
9 0.
03
0.37
1.
03
0.50
-2.1
0
c.81
6C>A
In
terc
ept
-0.0
2 0.
21
-1.8
8 0.
37
0.08
0.
20
Geno
type
10
0.38
6
0.47
4
0.23
2
CC
ref.
…
…
…
ref.
…
…
…
ref.
…
…
…
CA
-0.1
0 0.
28
0.91
0.
53-1
.55
0.18
0.
50
1.20
0.
45-3
.20
-0.0
1 0.
27
0.99
0.
58-1
.69
AA
0.61
0.
51
1.84
0.
67-5
.03
1.08
0.
88
2.93
0.
52-1
6.44
0.
83
0.51
2.
29
0.85
-6.1
8
c.98
0A>G
In
terc
ept
0.00
0.
24
-3.0
0 0.
64
0.04
0.
24
Geno
type
10,1
0.97
5 0.
016
0.81
6
GA
re
f. …
…
…
re
f. …
…
…
re
f. …
…
…
GG
0.01
0.
29
1.01
0.
58-1
.76
1.65
0.
69
5.19
1.
35-1
9.92
0.
18
0.29
1.
20
0.69
-2.1
0
1 The
logi
stic
mix
ed re
gres
sion
mod
els i
nclu
ded
herd
, hei
fer a
nd q
uart
er a
s ran
dom
effe
ct. 2 In
tram
amm
ary
infe
ctio
n ca
used
by
coag
ulas
e-ne
gativ
e st
aphy
loco
cci v
ersu
s non
-
infe
cted
. 3 Intr
amam
mar
y in
fect
ion
caus
ed b
y St
aphy
loco
ccus
aur
eus,
Str
epto
cocc
us a
gala
ctia
e, S
trep
toco
ccus
dys
gala
ctia
e an
d es
culin
e-po
sitiv
e st
rept
ococ
ci v
ersu
s no
n-
infe
cted
. 4 Intr
amam
mar
y in
fect
ion
caus
ed b
y co
agul
ase-
nega
tive
stap
hylo
cocc
i, Co
ryne
bact
eriu
m b
ovis,
Sta
phyl
ococ
cus
aure
us, S
trep
toco
ccus
aga
lact
iae,
Str
epto
cocc
us
dysg
alac
tiae
and
escu
line-
posit
ive
stre
ptoc
occi
ver
sus
non-
infe
cted
. 5 Re
gres
sion
coef
ficie
nt.
6 Stan
dard
err
or.
7 Odd
s ra
tio.
8 95%
con
fiden
ce i
nter
val
of O
R, i
nter
vals
excl
udin
g “1
” ar
e m
arke
d in
bol
d. 9 P-
valu
e of
the
over
all f
ixed
effe
ct w
ith v
alue
s <0.
05 m
arke
d in
bol
d. 10
Geno
type
at t
he p
ositi
on o
f the
stud
ied
poly
mor
phism
. 11
To a
llow
con
verg
ence
of t
he st
atist
ic m
odel
s, re
cord
s on
the
rare
gen
otyp
e c.
980A
A w
ere
omitt
ed.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
105
4. Discussion
To improve prevention and control of heifer mastitis, identification of risk factors is
required. Because of the multifactorial nature of mastitis, not only pathogen-specific but
host-specific factors as well determine the outcome. Mastitis resistance, and host resistance
in general, are partly genetically determined (Wanner et al., 1998; Nash et al., 2003).
Although many genes influence mastitis traits, polymorphisms in a few genes with major
effects might alter mastitis susceptibility of cows significantly (Detilleux, 2009). Because of
the importance of the encoded protein in the innate immunity of the mammary gland
(Barber and Yang, 1998; Caswell et al., 1999), a QTL for mastitis in this region of the bovine
genome (Sodeland et al., 2011) and the demonstrated relevance of CXCR1 polymorphisms
for mastitis susceptibility (Beecher et al., 2010; Galvao et al., 2011; Leyva-Baca et al., 2008;
Youngerman et al., 2004a), we opted to study CXCR1 genotype as a potential risk (or
protective) factor for heifer mastitis. Novel polymorphisms and a pathogen-group specific
association between CXCR1 genotype and intramammary infection status of early lactating
heifers were detected.
Recently, Pighetti et al. (2012) performed a polymorphism screening on CXCR1 by
genotyping 88 Holstein dairy cows. Although investigating the coding region of CXCR1 in the
same breed, we observed some differences compared to their results. Ten common
polymorphisms were detected in the two study populations. Additionally, one missense
polymorphism (c.621G>A) was discovered in the American dairy population which was not
found in our Belgian dairy heifer population and 6 polymorphisms (c.37A>T, c._38T>A,
c.68G>A, c.333T>C, c.337G>A and c.642G>A) were only observed in our study. Consequently,
linkage groups of SNP and haplotypes differed slightly among both study populations. The
haplotypes labeled GCCA, GCAG and GGCG based on tag SNP c.621G>A, c.735C>G, c.816C>A
and c.980A>G present in the American study population are equal to the haplotypes H1, H2
and H4 of our study, respectively. Haplotype AGCG was only observed in the American study,
whereas haplotype H3 and H5 were only found in the Belgian study population.
Intramammary infection status at quarter level assessed by bacteriological culture is
usually not available on large scale, although it is a direct parameter for udder health.
Youngerman et al. (2004a) were the first and until now the only to report on the association
between CXCR1 polymorphisms and udder health determined by bacteriological culture.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
106
Subclinical mastitis was quantified as the percentage of observations positive on IMI per cow
and found to be associated with polymorphism c.735C>G. Pathogen-group specific
associations were not studied. As not all pathogens have the same impact on udder health
and milk production of early lactating heifers (Piepers et al., 2010), and as risk factors for IMI
with the different groups of pathogens differ as well (Piepers et al., 2011), it is important to
differentiate between them. We studied associations with both IMI by all pathogens and
pathogen-group specific IMI (IMI by coagulase-negative staphylococci and IMI by major
pathogens, respectively). This allowed us to identify an association between polymorphism
c.980A>G and IMI by major pathogens while no association was found with IMI by all
pathogens and IMI by coagulase-negative staphylococci, respectively. Polymorphism
c.980A>G (p.Lys327Arg) was in complete linkage disequilibrium with 2 other missense SNP
(c.337G>A, p.Val113Ile; c.995A>G, p.His332Arg) and one silent SNP (c.333T>C). The less
frequently detected allele c.980A seemed to offer protection compared to the more
frequent allele c.980G. Although c.980 may not be the causative mutation, the A allele codes
for lysine at amino acid position 327 which is conserved across species (Pighetti et al., 2012).
Although polymorphism in CXCR1 has been linked with mastitis susceptibility
(Youngerman et al., 2004a; Galvao et al., 2011), we and several other research groups failed
to demonstrate an association with SCC (Galvao et al., 2011) or estimated breeding values of
SCC (Goertz et al., 2009). As Youngerman et al. (2004a) proposed, the relationship between
CXCR1 and SCC might be more complex than expected. From one point of view, heifers with
genotype associated with more expression or higher functionality of the receptor could
respond more efficient to invading pathogens. Therefore, these heifers might have less
(chronic) IMI and subsequent elevation of SCC. From another point of view, geometric mean
SCC could be higher because of the higher response to IL8 causing more cells to migrate
from blood to milk. Additionally, as suggested by results in our study, associations between
genotype and udder health might be pathogen (-group) specific. Latter associations might
not be detected when using SCC data only.
The sample size of this study was rather small (140 heifers) and the number of infections
caused by major pathogens was limited, i.e. 32 quarters of heifers with genotype AG, 3
quarters of heifers with genotype GG. Whether a P-value below 0.05 is sufficient to claim
that associations are not observed by chance can be argued. Keeping in mind the important
function of the receptor and findings of other research (Youngerman et al., 2004a; Galvao et

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
107
al., 2011, Sodeland et al., 2011; Pighetti et al., 2012), it seems unlikely that heifers with
genotype GG had more infections by major pathogens compared to heifers with genotype
AG merely by coincidence. Nevertheless, confirmation of the findings in other observational
studies with larger populations and experimental studies would be helpful. The current
results increase the knowledge on the impact of CXCR1 polymorphisms on mastitis
susceptibility, complement what is already known and allow for designing further studies.
Information from previous (Galvao et al., 2011; Pighetti et al., 2012; Sodeland et al., 2011;
Youngerman et al., 2004a), current and future research will enable us to draw better
conclusions on the relationship between mutations in CXCR1 and mastitis susceptibility and
its potentials for genetic selection.
5. Conclusions
Sixteen polymorphisms including 8 missense were discovered in the coding region of
CXCR1. Because of the important function of CXCR1 in innate immune responses, mutations
in the encoding gene might alter disease resistance. In this study, an association between
subclinical mastitis of early lactating heifers and polymorphism c.980A>G was detected.
Allele A was found to be a protective factor for IMI by major pathogens. These results
indicate that selection against certain CXCR1 genotypes could offer possibilities for
prevention and control of heifer mastitis. The observed association stimulates us to further
unravel the impact of CXCR1 polymorphisms on innate immunity and pathogen-specific
resistance of the mammary gland.
6. Acknowledgements
This study was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen). The authors wish to thank Milk
Control Centre Flanders for bacteriological culture and quarter SCC analyses and the Cattle
Breeding Association (CRV) for access to data on composite SCC at test-day and relationship
among heifers. Technical assistance by Linda Impe, Ruben Van Gansbeke and Dominique
Vander Donckt was greatly appreciated.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
108
7. References
Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic Local Alignment
Search Tool. J Mol Biol. 215:403-410.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab. Immunol. 5:82-86.
Beecher, C., M. Daly, S. Childs, D. P. Berry, D. A. Magee, T. V. McCarthy, and L. Giblin. 2010.
Polymorphisms in bovine immune genes and their associations with somatic cell count
and milk production in dairy cattle. BMC Gen. 11:99.
Birnboim, H. C. 1983. A rapid alkaline extraction method for the isolation of plasmid DNA.
Methods Enzymol. 100:243-255.
Caswell, J. L., D. M. Middleton, and J. R. Gordon. 1999. Production and functional
characterization of recombinant bovine interleukin-8 as a specific neutrophil activator
and chemoattractant. Vet. Immunol. Immunopathol. 67:327-340.
De Vliegher, S., H. Laevens, H. W. Barkema, I. R. Dohoo, H. Stryhn, G. Opsomer, and A. de
Kruif. 2004. Management practices and heifer characteristics associated with early
lactation somatic cell count of Belgian dairy heifers. J. Dairy Sci. 87:937-947.
De Vliegher, S., L. K. Fox, S. Piepers, S. McDougall, and H. W. Barkema. 2012. Invited review:
Mastitis in dairy heifers: nature of the disease, potential impact, prevention, and control.
J. Dairy Sci. 95:1025-1040.
Detilleux, J. C. 2009. Genetic factors affecting susceptibility to udder pathogens. Vet
Microbiol. 134:157-164.
Fox, L. K. 2009. Prevalence, incidence and risk factors of heifer mastitis. Vet. Microbiol.
134:82-88.
Galvao, K. N., G. M. Pighetti, S. H. Cheong, D. V. Nydam, and R. O. Gilbert. 2011. Association
between interleukin-8 receptor-alpha (CXCR1) polymorphism and disease incidence,
production, reproduction, and survival in Holstein cows. J Dairy Sci. 94:2083-2091.
Goertz, I., C. Baes, C. Weimann, N. Reinsch, and G. Erhardt. 2009. Association between single
nucleotide polymorphisms in the CXCR1 gene and somatic cell score in Holstein dairy
cattle. J Dairy Sci. 92:4018-4022.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
109
Grosse, W. M., S. M. Kappes, W. W. Laegreid, J. W. Keele, C. G. Chitko-McKown, and M. P.
Heaton. 1999. Single nucleotide polymorphism (SNP) discovery and linkage mapping of
bovine cytokine genes. Mamm. Genome 10:1062-1069.
Huijps, K., S. De Vliegher, T. Lam, and H. Hogeveen. 2009. Cost estimation of heifer mastitis
in early lactation by stochastic modelling. Vet. Microbiol. 134:121-127.
Kettritz, R., M. L. Gaido, H. Haller, F. C. Luft, C. J. Jennette, and R. J. Falk. 1998. Interleukin-8
delays spontaneous and tumor necrosis factor-alpha-mediated apoptosis of human
neutrophils. Kidney Int. 53:84-91.
Kromker, V., and J. Friedrich. 2009. Teat canal closure in non-lactating heifers and its
association with udder health in the consecutive lactation. Vet. Microbiol. 134:100-105.
Leyva-Baca, I., F. Schenkel, J. Martin, and N. A. Karrow. 2008. Polymorphisms in the 5 '
upstream region of the CXCR1 chemokine receptor gene, and their association with
somatic cell score in Holstein cattle in Canada. J Dairy Sci. 91:407-417.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
Myllys, V., and H. Rautala. 1995. Characterization of clinical mastitis in primiparous heifers. J.
Dairy Sci. 78:538-545.
Ogorevc, J., T. Kunej, A. Razpet, and P. Dovc. 2009. Database of cattle candidate genes and
genetic markers for milk production and mastitis. Anim Genet. 40:832-851.
Paape, M. J., K. Shafer-Weaver, A. V. Capuco, K. Van Oostveldt, and C. Burvenich. 2000.
Immune surveillance of mammary tissue by phagocytic cells. Adv. Exp. Med. Biol.
480:259-277.
Piepers, S., L. De Meulemeester, A. de Kruif, G. Opsomer, H. W. Barkema, and S. De Vliegher.
2007. Prevalence and distribution of mastitis pathogens in subclinically infected dairy
cows in Flanders, Belgium. J. Dairy Res. 74:478-483.
Piepers, S., G. Opsomer, H. W. Barkema, A. de Kruif, and S. De Vliegher. 2010. Heifers
infected with coagulase-negative staphylococci in early lactation have fewer cases of
clinical mastitis and higher milk production in their first lactation than noninfected
heifers. J. Dairy Sci. 93:2014-2024.

Chapter 4.1 The CXCR1 gene and subclinical heifer mastitis
110
Piepers, S., K. Peeters, G. Opsomer, H. W. Barkema, K. Frankena, and S. De Vliegher. 2011.
Pathogen group specific risk factors at herd, heifer and quarter levels for intramammary
infections in early lactating dairy heifers. Prev. Vet Med. 99:91-101.
Pighetti, G. M., and M. Rambeaud. 2006. Genome conservation between the bovine and
human interleukin-8 receptor complex: Improper annotation of bovine interleukin-8
receptor b identified. Vet. Immunol. Immunopathol. 114:335-340.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Sodeland, M., M. P. Kent, H. G. Olsen, M. A. Opsal, M. Svendsen, E. Sehested, B. J. Hayes, and
S. Lien. 2011. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20
in Norwegian Red cattle. Anim. Genet. 42:457-465.
Svensson, C., A. K. Nyman, W. K. Persson, and U. Emanuelson. 2006. Effects of housing,
management, and health of dairy heifers on first-lactation udder health in southwest
Sweden. J. Dairy Sci. 89:1990-1999.
Untergasser, A., H. Nijveen, X. Rao, T. Bisseling, R. Geurts, and J. A. M. Leunissen. 2007.
Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 35:W71-W74.
Van Poucke, M., J. Vandesompele, M. Mattheeuws, A. Van Zeveren, and L. J. Peelman. 2005.
A dual fluorescent multiprobe assay for prion protein genotyping in sheep. BMC Infect.
Dis. 5:13.
Waage, S., S. A. Odegaard, A. Lund, S. Brattgjerd, and T. Rothe. 2001. Case-control study of
risk factors for clinical mastitis in postpartum dairy heifers. J. Dairy Sci. 84:392-399.
Wanner, J. M., G. W. Rogers, M. E. Kehrli, and J. B. Cooper. 1998. Intramammary infections in
primiparous Holsteins: heritabilities and comparisons of bovine leukocyte adhesion
deficiency carriers and noncarriers. J. Dairy Sci. 81:3293-3299.
Youngerman, S. M., A. M. Saxton, S. P. Oliver, and G. M. Pighetti. 2004a. Association of
CXCR2 polymorphisms with subclinical and clinical mastitis in dairy cattle. J. Dairy Sci.
87:2442-2448.
Youngerman, S. M., A. M. Saxton, and G. M. Pighetti. 2004b. Novel single nucleotide
polymorphisms and haplotypes within the bovine CXCR2 gene. Immunogenetics 56:355-
359.
Zuker, M. 2003. Mfold web server for nucleic acid folding and hybridization prediction.
Nucleic Acids Res. 31:3406-3415.

111
Chapter 4.2
Somatic cell count and milk neutrophil viability of
dairy heifers with specific CXCR1 genotypes following
experimental intramammary infection with
Staphylococcus chromogenes originating from milk
J. Verbeke1, K. Piccart1, S. Piepers1, M. Van Poucke2, L. Peelman2, A. De
Visscher1, S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine, Ghent
University, Belgium
The Veterinary Journal, revisions.

Chapter 4.2 The CXCR1 gene and experimental infection
112
Abstracts
Previous observational studies suggest an association between polymorphism c.980A>G
in the CXCR1 gene, encoding the chemokine (C-X-C motif) receptor 1, and the innate
immunity and infection status of the mammary gland. Mammary glands of eight Holstein
heifers were experimentally infected with a Staphylococcus chromogenes isolate originating
from a chronic intramammary infection (IMI) to study differences between CXCR1 genotypes
c.980AG and c.980GG.
Quarters from heifers with genotype c.980AG and c.980GG both became subclinically
infected but showed differences in the early stage of infection. Bacterial count at 18 h post
challenge tended to be higher in quarters from c.980AG heifers compared to c.980GG
heifers. Somatic cell count (SCC) was lower at 6 h post challenge and tended to be lower at 9
h post challenge in c.980AG heifers compared to c.980GG heifers. Yet, milk production
decreased similarly. Milk neutrophils of c.980AG heifers showed more apoptosis at 9 h post
challenge and tended to show more necrosis at 6, 9 and 12 h post challenge than c.980GG
heifers. Differences were less pronounced in the later stage (> 18 h) of infection. Results
demonstrate that CXCR1 polymorphism can influence SCC and milk neutrophil viability
following experimental IMI.
Keywords
CXCR1 polymorphism, Experimental intramammary challenge, Staphylococcus chromogenes

Chapter 4.2 The CXCR1 gene and experimental infection
113
1. Introduction
Mastitis incidence can be decreased by selective breeding for higher resistance to
pathogens (Detilleux, 2009; Windig et al., 2010). Identification of genetic polymorphisms
linked with mastitis resistance allows for marker-assisted selection for udder health
(Ogorevc et al., 2009; Pighetti and Elliott, 2011). Neutrophils play a major role in the
mammary gland immunity (Paape et al., 2002). Binding of interleukin-8 (IL-8) on chemokine
(C-X-C motif) receptor 1 (CXCR1) causes chemotaxis and delay of apoptosis of neutrophils
(Barber and Yang, 1998; Kettritz et al., 1998). Sixteen polymorphisms in the coding region of
CXCR1 potentially influence IL-8 binding and signal transduction (Pighetti et al., 2012;
Verbeke et al., 2012).
Polymorphism CXCR1 c.980A>G causes the amino acid change p.Lys327Arg in the C-
terminal region of the receptor known to be important for internalization and chemotaxis
(Richardson et al., 2003). Recently, we revealed that CXCR1 c.980AG heifers showed a lower
% apoptosis of resident milk neutrophils and were less likely to be infected with major
mastitis pathogens (e.g. Staphylococcus aureus and Streptococcus uberis) in early lactation
compared to CXCR1 c.980GG heifers (Verbeke et al., 2012; Verbeke et al., 2014). To gain
more insight in the effect of c.980A>G on mammary gland immunity, we compared the
quarter bacterial count, SCC, milk production (MP), % milk neutrophil apoptosis and % milk
neutrophil necrosis of 4 c.980AG and 4 c.980GG dairy heifers following an experimental
challenge with S. chromogenes originating from a chronic IMI.
2. Materials and methods
2.1 Test animals
The experiment has been approved by the ethical committee of the Faculty of Veterinary
Medicine, Ghent University (EC2012/73, 14 May 2012). A blood sample was taken from all
Holstein heifers (n = 20) at the Ghent University dairy herd (Biocenter Agri-Vet, Melle,
Belgium). The whole coding region of CXCR1 was genotyped by direct sequencing as
described by Verbeke et al. (2012). Four heifers with genotype c.980AG and four heifers with
genotype c.980GG were selected. Selected heifers were not siblings, had no history of
clinical mastitis or other diseases and were between 75 and 280 days in milk at the time of
the experiment. Because any existing IMI could bias results, heifers were treated 15 days

Chapter 4.2 The CXCR1 gene and experimental infection
114
before experimental infection with 200 mg cefalexin and 100,000 I.U. kanamycin
intramammary for 2 consecutive days (Ubrolexin, Boehringer Ingelheim) and with 10 g
penethamate hydriodide (Mamyzin, Boehringer Ingelheim) intramuscularly for three
consecutive days according the manufacturer’s product label. Eighteen and four days before
the experimental infection, duplicate quarter milk samples were taken. Ten microliter of milk
was spread on blood-aesculin and MacConkey’s agar and incubated aerobically for 24-48 h
at 37 °C. Bacteriological culture was performed according to National Mastitis Council (NMC)
guidelines (National Mastitis Council, 1999). All quarters of all heifers were free from major
pathogens 18 days before experimental infection and culture-negative four days before
experimental infection. Two days before challenge, heifers were moved from the free stall
barn to the tie stall barns and returned 80 h after the experimental infection.
2.2 Experimental challenge
Data were available from a larger experimental challenge study in which eight heifers
were inoculated with two different strains of S. chromogenes, a strain of Staphylococcus
fleurettii and sterile phosphate buffered saline (PBS) using a split-udder design (one strain or
PBS per individual quarter). One S. chromogenes strain (S. chromogenes IM) originated from
a chronically infected quarter (Piessens et al., 2011) whereas the other (S. chromogenes TA)
originated from a teat apex and was found to inhibit the growth of major pathogens in vitro
(De Vliegher et al., 2004). The S. fleurettii strain originated from sawdust (Piessens et al.,
2011). The bacterial count was determined by incubating tenfold serial dilutions of
representative frozen aliquots 18 h before inoculation. In total, 1 x 106 colony forming units
(CFU) in 5 mL PBS were inoculated using a catheter (Vygon, Ecouen, France). The optimal
infection dose to elicit subclinical mastitis was determined by challenging a reform cow with
different doses of S. chromogenes TA in each quarter (0.25, 0.5, 1 and 2 x 106 CFU). An
inflammatory response (SCC > 1,000 cells/μL) but no clinical signs were observed in the
quarter inoculated with 1 x 106 CFU. In the current study, only data from the quarters
inoculated with S. chromogenes IM was used because only bacterial count of S. chromogenes
IM increased between 4 and 6 h post challenge proving IMI.
2.3 Sampling

Chapter 4.2 The CXCR1 gene and experimental infection
115
Heifers were clinically examined before each sampling by a veterinarian (first, second or
third author). Duplicate quarter milk samples were taken 24, 12 and 1 h before inoculation
and four, six, nine, 12, 18, 24, 28, 32, 36, 48, 54, 60, 72, and 78 h after inoculation for
bacteriological culture. Ten microliters of milk was spread on blood-aesculin and
MacConkey’s agar and incubated aerobically for 24-48 h at 37 °C. Additionally, tenfold serial
dilutions in PBS were plated on tryptic soy agar (TSA) and incubated aerobically for 48 h at
37 °C to determine the bacterial count (CFU/ml milk). Coagulase-negative staphylococci
growing on TSA were identified to the species level by tRNA-intergenic spacer PCR combined
with capillary electrophoresis and rpoB sequencing as described before (Supré et al., 2009).
Twenty-four and one hour before and 4, 6, 9, 12, 18, 24, 28, 32, 36, 48, 54, 60, 72, and 78 h
after inoculation, quarter SCC was measured in duplicate using a DeLaval cell counter (DCC).
Heifers were machine milked twice daily and quarter milk production (kg) was registered 24
and 1 h before and 12, 24, 36, 48, 60, and 72 h after inoculation. Additional milk samples
were taken 24 and 1 h before and 6, 9, 12, 18, 24, 48, and 72 h after inoculation to monitor
milk neutrophil viability. The % milk neutrophil apoptosis and necrosis was measured
flowcytometrically as described before (Piepers et al., 2009).
2.4 Statistical methods
Data transformations were performed to obtain normalised distributions when needed.
Bacterial count was log-transformed [log(CFU/mL + 1)] whereas SCC (cells/μL), % milk
neutrophil apoptosis and % milk neutrophil necrosis were natural log-transformed. Data
were analysed similarly as Sipka et al. (2013). Different linear mixed regression models were
fit (PROC MIXED, SAS 9.4, SAS Institute) for bacterial count, SCC, milk production, % milk
neutrophil apoptosis and % milk neutrophil necrosis (outcome variables) with heifer as
random effect to correct for clustering of multiple observations per heifer (REPEATED
statement) and cows’ genotype (c.980AG or c.980GG), sampling moment and their
interaction as categorical fixed effects. Least square means of c.980AG and c.980GG heifers
were compared at each sampling moment (independent post-hoc tests). Statistical
significance was assessed at P ≤ 0.05 whereas a P-value between 0.05 and 0.10 was
considered as a tendency toward significance.

Chapter 4.2 The CXCR1 gene and experimental infection
116
3. Results
All milk samples taken 24, 12 and 1 h before inoculation remained culture negative. All
quarters became subclinically infected as no local or systemic signs were observed and the
isolates recovered from milk after inoculation were identified as the challenge species. No
pathogens other than coagulase-negative staphylococci were recovered. Bacteria were
rapidly eliminated in quarters from both genotype groups. Yet, bacterial counts tended to be
higher in quarters from c.980AG heifers compared to c.980GG heifers at 18 h post challenge
(P = 0.09) (Fig. 4.2.1A). Quarter milk SCC increased in both genotypes. Quarters from heifers
with genotype c.980GG had a higher SCC at 6 h post challenge (P < 0.05) and tended to have
a higher SCC 9 h post challenge (P = 0.06) compared to heifers with genotype c.980AG (Fig.
4.21B). Milk production following intramammary challenge decreased in all quarters . No
statistically significant differences were observed between genotype groups at any sampling
moment (Fig. 4.2.1C). Mean % milk neutrophil apoptosis was higher in c.980AG heifers
compared to c.980GG heifers at each sampling moment after inoculation with differences
being statistically significant at 9 h post challenge (P < 0.05) (Fig. 4.2.2A). Similar to
apoptosis, % milk neutrophil necrosis was higher in c.980AG heifers compared to c.980GG on
the day of inoculation. Differences tended to be statistically significant at 1 h before
inoculation (P = 0.06) and at 6 (P = 0.08), 9 (P = 0.07) and 12 h (P = 0.08) post challenge (Fig.
4.2.2B).

Chapter 4.2 The CXCR1 gene and experimental infection
117
A
B
C
Figure 4.2.1. Bacterial count (A), somatic cell count (B) and milk production (C) following experimental
intramammary challenge with Staphylococcus chromogenes in four CXCR1 c.980AG heifers and four CXCR1
c.980GG heifers. Data are least square means ± standard error. Capital and lower case letters indicate the
presence (P < 0.05) or tendency towards (P < 0.1) statistically significant differences between genotypes,
respectively.
a
b 00.5
11.5
22.5
33.5
4
-24 -18 -12 -6 0 6 12 18 24 30 36 42 48 54 60 66 72 78
Log
bact
eria
l cou
nt (C
FU/m
L)
Time relative to inoculation (h)
c.980AGc.980GG
B
b
A
a
0123456789
-24 -18 -12 -6 0 6 12 18 24 30 36 42 48 54 60 66 72 78Ln so
mat
ic c
ell c
ount
(ce
lls/μ
L)
Time relative to inoculation (h)
c.980AGc.980GG
0
0.5
1
1.5
2
2.5
3
3.5
4
-24 -18 -12 -6 0 6 12 18 24 30 36 42 48 54 60 66 72 78
Milk
yie
ld (
kg)
Time relative to inoculation (h)
c.980AGc.980GG

Chapter 4.2 The CXCR1 gene and experimental infection
118
A
B
Figure 4.2.2. Percentage milk neutrophil apoptosis (A) and necrosis (B) following experimental intramammary
challenge with Staphylococcus chromogenes in four CXCR1 c.980AG heifers and four CXCR1 c.980GG heifers.
Data are least square means ± standard error. Capital and lower case letters indicate the presence (P < 0.05) or
tendency towards (P < 0.1) statistically significant differences between genotypes, respectively.
4. Discussion
Experimental intramammary infection trials offer the possibility to study host response
differences in much detail. Based on our previous results (Verbeke et al., 2012; Verbeke et
al., 2014), we decided to challenge heifers and analyze differences in bacterial count,
somatic cell count, milk production and milk neutrophil viability between c.980AG and
c.980GG heifers.
During the early stage of infection, c.980AG heifers differed from c.980GG heifers. The
lower SCC at 6 h post challenge and tendency towards lower SCC at 9 h post challenge
A
B 0102030405060708090
100
-24 -18 -12 -6 0 6 12 18 24 30 36 42 48 54 60 66 72 78
Milk
neu
trop
hil a
popt
osis
(%)
Time relative to inoculation (h)
c.980AGc.980GG
a a a a
b b b b 0102030405060708090
100
-24 -18 -12 -6 0 6 12 18 24 30 36 42 48 54 60 66 72 78
Milk
neu
trop
hil n
ecro
sis (
%)
Time relative to inoculation (h)
c.980AGc.980GG

Chapter 4.2 The CXCR1 gene and experimental infection
119
suggest a slower milk leukocyte migration in quarters from c.980AG heifers compared to
c.980AG heifers. The latter finding corresponds well with in vitro research demonstrating an
association between CXCR1 genotype and neutrophil migration (Rambeaud and Pighetti,
2005). Genotype c.980AG was associated with a higher % milk neutrophil apoptosis in this
study but a lower % milk neutrophil apoptosis in a previous study (Verbeke et al., 2014).
However, previous results demonstrated differences mainly in uninfected quarters whereas
acutely infected quarters were studied in this research. Not only after but also 1 h before
inoculation, quarters from c.980AG heifers tended to have a higher % milk neutrophil
necrosis compared to c.980GG heifers. The lower viability might have impaired the
phagocytic and respiratory burst capacity of neutrophils in c.980AG heifers (Mehrzad et al.,
2004; Van Oostveldt et al., 2002). Although bacteria were rapidly eliminated in both
genotype groups, quarters from c.980AG heifers tended to have higher bacterial count at 18
h post challenge compared to c.980GG heifers. Nevertheless, the milk yield reduction was
similar.
Single nucleotide polymorphism c.980A>G causes an amino acid changes in the C-
terminus of the protein (p.Lys327Arg) potentially affecting adaptin-2 binding and receptor
internalization (Pighetti et al., 2012). The current research indicates c.980A>G influences the
host response following S. chromogenes infection and provides a foundation for future
research. Infection trials with other mastitis pathogens (e.g. S. aureus or S. uberis) or in vitro
studies focusing on the same and other SNP in CXCR1 could further elucidate the role of
CXCR1 genotype for mammary gland immunity and udder health.
5. Conclusions
In conclusion, both c.980AG and c.980GG heifers rapidly eliminated Staphylococcus
chromogenes following intramammary challenge. Yet, quarters from c.980GG heifers
showed a higher SCC and lower milk neutrophil viability during the early infection compared
to c.980AG heifers. Results demonstrate that CXCR1 polymorphism can influence SCC and
milk neutrophil viability following experimental IMI.

Chapter 4.2 The CXCR1 gene and experimental infection
120
6. Acknowledgements
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen). Technical assistance by Lars Hulpio,
Véronique Byn and Marnik De Norre was appreciated.
7. References
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab Immunol. 5:82-86.
Detilleux, J. C. 2009. Genetic factors affecting susceptibility to udder pathogens. Vet
Microbiol. 134:157-164.
De Vliegher, S., G. Opsomer, A. Vanrolleghem, L. A. Devriese, O. C. Sampimon, J. Sol, H. W.
Barkema, F. Haesebrouck, and A. de Kruif. 2004. In vitro growth inhibition of major
mastitis pathogens by Staphylococcus chromogenes originating from teat apices of dairy
heifers. Vet. Microbiol. 101:215-221.
Kettritz, R., M. L. Gaido, H. Haller, F. C. Luft, C. J. Jennette, and R. J. Falk. 1998. Interleukin-8
delays spontaneous and tumor necrosis factor-alpha-mediated apoptosis of human
neutrophils. Kidney Int. 53:84-91.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2004. Viability of milk neutrophils and severity
of bovine coliform mastitis. J. Dairy Sci. 87:4150-4162.
National Mastitis Council. 1999. Laboratory Handbook on Bovine Mastitis. National Mastitis
Council. Inc., Madison, WI.
Ogorevc, J., T. Kunej, A. Razpet, and P. Dovc. 2009. Database of cattle candidate genes and
genetic markers for milk production and mastitis. Anim Genet. 40:832-851.
Paape, M., J. Mehrzad, X. Zhao, J. Detilleux, and C. Burvenich. 2002. Defense of the bovine
mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary Gland Biol.
Neoplasia 7:109-121.
Piepers, S., S. De Vliegher, K. Demeyere, B. N. Lambrecht, A. de Kruif, E. Meyer, and G.
Opsomer. 2009. Technical note: Flow cytometric identification of bovine milk neutrophils
and simultaneous quantification of their viability. J. Dairy Sci. 92:626-631.

Chapter 4.2 The CXCR1 gene and experimental infection
121
Piessens, V., E. Van Coillie, B. Verbist, K. Supre, G. Braem, A. Van Nuffel, L. De Vuyst, M.
Heyndrickx, and S. De Vliegher. 2011. Distribution of coagulase-negative Staphylococcus
species from milk and environment of dairy cows differs between herds. J. Dairy Sci.
94:2933-2944.
Pighetti, G. M., and A. A. Elliott. 2011. Gene polymorphisms: the keys for marker assisted
selection and unraveling core regulatory pathways for mastitis resistance. J. Mammary.
Gland. Biol. Neoplasia. 16:421-432.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Veterinary Immunology and Immunopathology 145:464-470.
Rambeaud, M., and G. M. Pighetti. 2005. Impaired neutrophil migration associated with
specific bovine CXCR2 genotypes. Infect. Immun. 73:4955-4959.
Richardson, R. M., R. J. Marjoram, L. S. Barak, and R. Snyderman. 2003. Role of the
cytoplasmic tails of CXCR1 and CXCR2 in mediating leukocyte migration, activation, and
regulation. J. Immunol. 170:2904-2911.
Sipka, A., A. Gurjar, S. Klaessig, G. E. Duhamel, A. Skidmore, J. Swinkels, P. Cox, and Y.
Schukken. 2013. Prednisolone and cefapirin act synergistically in resolving experimental
Escherichia coli mastitis. J. Dairy Sci. 96:4406-4418.
Supré, K., S. De Vliegher, O. C. Sampimon, R. N. Zadoks, M. Vaneechoutte, M. Baele, E. De
Graef, S. Piepers, and F. Haesebrouck. 2009. Technical note: Use of transfer RNA-
intergenic spacer PCR combined with capillary electrophoresis to identify coagulase-
negative Staphylococcus species originating from bovine milk and teat apices. J. Dairy Sci.
92:3204-3210.
Van Oostveldt, K., M. J. Paape, H. Dosogne, and C. Burvenich. 2002. Effect of apoptosis on
phagocytosis, respiratory burst and CD18 adhesion receptor expression of bovine
neutrophils. Domest. Anim. Endocrinol. 22:37-50.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2014a. Association of
CXCR1 polymorphisms with apoptosis, necrosis and concentration of milk neutrophils in
early lactating dairy heifers. Res. Vet. Sci. 97: 55-59.
Verbeke, J., S. Piepers, L. J. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-
group specific association between CXCR1 polymorphisms and subclinical mastitis in
dairy heifers. J. Dairy Res. 79:341-351.

Chapter 4.2 The CXCR1 gene and experimental infection
122
Windig, J. J., W. Ouweltjes, J. Ten Napel, G. de Jong, R. F. Veerkamp, and Y. de Haas. 2010.
Combining somatic cell count traits for optimal selection against mastitis. J. Dairy Sci.
93:1690-1701.

123
Chapter 5
The role of CXCR1 gene polymorphisms in clinical mastitis

124

125
Chapter 5.1
Pathogen-specific incidence rate of clinical mastitis in
Flemish dairy herds, severity, and association with
herd hygiene
J. Verbeke1, S. Piepers1, K. Supré2, and S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Milk Control Centre Flanders, Belgium
Journal of Dairy Science, 2014, 97: 6926-6934.

Chapter 5.1 Clinical mastitis in Flanders
126
Abstract
A one-year survey on clinical mastitis was conducted on 50 randomly selected
commercial Flemish dairy herds to estimate the pathogen-specific incidence rate of clinical
mastitis (IRCM). The severity of the cases and the potential associations with herd hygiene
were studied. Participating producers sampled 845 cases and 692 dairy cows. The mean and
median IRCM was estimated at 7.4 and 5.3 quarter cases per 10,000 cow-days at risk,
respectively. A large between-herd variation was observed (range 0-21.3). In general, the
IRCM was lower in heifers compared to multiparous cows (2.9 vs. 11.0 quarter cases per
10,000 cow-days at risk). However, the overall IRCM in the first week after calving was
higher in heifers compared to cows (43.4 vs. 31.6 quarter cases per 10,000 cow-days at risk).
Streptococcus uberis (18.2% of the cases) and Escherichia coli (15.5%) were the most
frequently isolated pathogens and no growth was observed in 19.9% of the cases. The
majority of the cases (63.1%) were mild (only clots in milk). Moderate (hard quarter without
general signs) and severe symptoms (systemic illness) were observed in 29.9%, and 7.0% of
the cases, respectively. Isolation of E. coli (vs. any other culture result) was more likely in
moderate and severe cases compared to mild cases. Overall IRCM and E. coli IRCM were
higher in dirty compared to clean herds based on udder hygiene scores (9.0 and 1.7 vs. 6.0
and 0.6 quarter cases per 10,000 cow-days at risk, respectively). This study broadens the
knowledge on clinical mastitis in Flemish dairy herds and underlines the high risk of clinical
mastitis in early lactating heifers, the role of the so-called environmental pathogens, and
herd hygiene.
Keywords
Clinical mastitis, incidence rate, severity, herd hygiene

Chapter 5.1 Clinical mastitis in Flanders
127
1. Introduction
Mastitis is one of the most common diseases in dairy cattle. The inflammatory reaction
primarily occurs in response to bacterial intramammary infections (IMI) and impairs milk
quality (Ma et al., 2000; Santos et al., 2003). The disease is accompanied by clinical signs
(clinical mastitis) or presents itself without observable signs (subclinical mastitis).
Several countries have reported IRCM data, ranging from 5.5 quarter cases per 10,000
cow-days at risk in French herds with a low bulk milk SCC to 12.9 quarter cases per 10,000
cow-days at risk in randomly selected herds in England and Wales (Barkema et al., 1998;
Barnouin et al., 2005; Bradley et al., 2007; Olde Riekerink et al., 2008; Wolff et al., 2012). In
contrast to subclinical mastitis (Piepers et al., 2007), there is a general lack of information on
the occurrence of clinical mastitis in Flanders. As on most Flemish dairy herds cases are not
recorded and rarely sampled for culture, the exact incidence of clinical mastitis remains
unknown. Yet, according to an internet questionnaire performed by Passchyn and colleagues
(P. Passchyn, S. Piepers and S. De Vliegher, Ghent University, Ghent, Belgium, personal
communication) 300 dairy producers estimated that 46% of the Flemish dairy cows suffer at
least once per year from clinical mastitis. Because the majority of questioned producers
admitted not to keep disease records, the latter estimate might differ substantially from the
actual IRCM in Flanders.
In studies performed in the Netherlands, Canada and Ireland, Staphylococcus aureus was
the most frequently isolated pathogen isolated from clinical mastitis cases (Barkema et al.,
1998; Olde Riekerink et al., 2008; Keane et al., 2013) whereas Streptococcus uberis and
Escherichia coli were the most frequently isolated pathogens in studies performed in the UK
(Bradley et al., 2007) and the US (Oliviera et al., 2013). Because management systems differ
between regions (e.g. Olde Riekerink et al., 2008) and dairy farming and mastitis control
evolve over time (Bradley et al., 2002), regular clinical mastitis studies at regional or national
level remain indispensable for adapted mastitis prevention programs and development of
novel prevention and control tools. In 2012, S. uberis and E. coli were the most frequently
isolated pathogens from samples of clinical mastitis cases submitted to the Flemish Milk
Control Centre (MCC Vlaanderen, Lier, Belgium), the largest milk laboratory in Flanders
(Annual Report 2012). Although the included herds and cases are not randomly selected, the
results suggest these so-called environmental pathogens to be the main cause of clinical

Chapter 5.1 Clinical mastitis in Flanders
128
mastitis in Flanders. Reducing exposure of teats with manure and organic materials is pivotal
in the prevention of environmental mastitis (Smith and Hogan, 1993). Schreiner and Ruegg
(2003) developed a four-point scale to score udder hygiene and reported that cows with a
higher udder hygiene score (dirtier) had higher SCC values and were more likely to have
subclinical mastitis. Using the same scoring system, Breen et al. (2009) identified high udder
hygiene score as a cow-level risk factor for clinical mastitis in general and for E. coli clinical
mastitis in specific. It remains unclear whether hygiene issues can be detected as risk factors
for clinical mastitis in Flanders, Belgium, as well.
Signs of clinical mastitis range from abnormal milk to systemic illness with severity of
cases depending on both cow and pathogen factors (Burvenich et al., 2003; Bannerman et
al., 2004; Schukken et al., 2011). Oliveira et al. (2013) recently characterized clinical mastitis
on large dairy herds in Wisconsin (US) and reported that systemic signs of illness were more
likely to be observed in Gram-negative cases. Escherichia coli was isolated in 12.5%, 22.3%
and 48.3% of the mild, moderate and severe cases. To the best of our knowledge, no
scientific information on signs of clinical mastitis is available in Flanders, let alone a link with
the associated pathogens. Yet, as in other regions, “E. coli mastitis” is often used as a
synonym for severe clinical mastitis.
The primary objective of this study was to estimate the pathogen-specific IRCM in
Flanders, Belgium. Additionally, associations between the culture result and the reported
severity and between (pathogen-specific) IRCM and herd hygiene were studied.
2. Materials and methods
2.1 Herds and study design
Clinical mastitis was monitored at the cow level (experimental unit) on randomly selected
dairy herds (sampling unit) during one year. The sample size for this study was estimated at
42 herds, using following formula (Dohoo et al., 2003): n = = , where n =
number of herds needed to estimate the incidence rate, Zα/2 = 95th percentile of a standard
normal distribution, p = a priori estimate of the proportion set at 0.46 (P. Passchyn, S.
Piepers and S. De Vliegher, Ghent University, Ghent, Belgium, personal communication), q =
1 – p, and L = margin of error set at 0.15.

Chapter 5.1 Clinical mastitis in Flanders
129
To allow for noncompliance, 67 herds were randomly selected from the database of the
Flemish Milk Control Centre comprising all Flemish dairy producers (n = 5,261) using the
Excel RAND function (Excel 2010, Microsoft Corp., Redmond, WA). Thirteen, 4, 7, 19 and 24
herds were contacted in the provinces Antwerp, Flemish Brabant, Limburg, East Flanders
and West Flanders, respectively, matching the distribution of dairy herds over the five
Flemish provinces (19 %, 6 %, 10 %, 28 %, and 37%) (Federal Public Service Economy, Small
and Medium Enterprises, Self-Employed, and Energy, 2012). No inclusion criteria were
applied. Fifty-three producers agreed to participate in this study (response rate of 79%). The
main argument for non-compliance was lack of time (n = 7) followed by planned retirement
(n = 3). The 4 remaining producers gave no specific argument for not participating.
The selected dairy herds were visited at the beginning of the study between September
and October 2012. At that time, the study details were discussed with the producers and
herd veterinarians, a questionnaire was performed and observations were made (see
further). From then on, producers were asked to take a single sample from each quarter
showing signs of clinical mastitis during the 12 month study period. Signs of clinical mastitis
were defined as visible abnormalities in the udder or milk indicating udder inflammation and
were detected by examination of foremilk and the udder before milking. In herds with an
automated milking system, producers examined foremilk and the udder of cows with
changes in sensor data (electrical conductivity, color, and yield), reduced milking frequency
or visual abnormalities (e.g. swelling or redness of the udder). Thresholds were set by the
producer. Dry cows were monitored by visual observation. Sample materials were provided
and sampling procedures were explained as well as the importance of an aseptic sampling
procedure. Sampling date, cow identification, quarter position, and clinical signs (absence or
presence of clots in milk, a hard quarter or systemic illness) were recorded by the producer
as well. Cases were categorized as mild (only clots in milk), moderate (hard quarter but no
general signs) or severe (signs of systemic illness) similar to Pinzón-Sánchez and Ruegg
(2011). Samples were frozen on-farm for one to several weeks and collected by the herd
veterinarian. A courier of the Flemish Animal Health Service (Dierengezondheidszorg)
transported the samples from the veterinary practice to the Milk Control Centre Flanders
where bacteriological culture was performed. Three herds were omitted from the analysis
along the study period because producers admitted halfway the study they were not

Chapter 5.1 Clinical mastitis in Flanders
130
sampling all the cases, resulting in 50 herds and 4,133 cows to be included in the final
dataset.
Producers were motivated to sample each single case by making the culture results
available to them as soon as possible, by paying an incentive of 3€ per collected sample, by
performing a secondary herd visit between February and March 2013 to discuss the
preliminary results, and by contacting them at least once every 2 months by phone. Herd
veterinarians were actively involved in the project by sharing all culture results from all
herds they were associated with, inviting them to join both herd visits, and financially
supporting them to keep track of the herd administration. Both producers and herd
veterinarians were invited at a meeting in October 2013, after completion of the survey, to
discuss the results and received a summary with herd-specific results, as was promised at
the onset of the study. Both producers and vets were aware of this initiative at the start of
the study.
2.2 Herd hygiene
Udder hygiene was scored of 20 randomly selected lactating cows during both herd visits
performed by the first author as described by Schreiner and Ruegg (2003). In herds with less
than 20 lactating cows, all lactating cows were scored. The proportion of cows having udder
hygiene score 3 or 4 was calculated for each visit. Herds having an average proportion > 50%
over the 2 visits were categorized as dirty, other herds were categorized as clean.
2.3 Cow data
Cow-level records [calving date(s), parity and culling date] were retrieved from DHI
records for herds participating in the DHI program and from the identification and
registration system of the Animal Health Service Flanders (Drongen, Belgium) for other
herds.
2.4 Bacteriological culture
Bacteriological culture was based on National Mastitis Council guidelines (1999) and
performed at the Milk Control Centre Flanders. From each thawed sample, 10 μL of milk was
spread on blood-aesculin and MacConkey’s agar and incubated aerobically for 24-48 h at 37

Chapter 5.1 Clinical mastitis in Flanders
131
°C. Samples were considered to be culture-positive if one or more colonies were observed (≥
100 CFU/ml). Identification of bacteria was done by Gram-staining, inspection of the colony
morphology and biochemical testing. Catalase tests were performed to differentiate Gram-
positive cocci in catalase-positive or catalase-negative cocci. Staphylococci were identified as
Staphylococcus aureus or non-aureus staphylococci, referred to as Staphylococcus spp.
throughout this paper, by colony morphology, haemolysis patterns, and DNase tests. Isolates
of Streptococcus-Entercococcus group were differentiated in aesculin-positive and aesculin-
negative cocci. Streptococcus uberis was distinguished from other aesculin-positive cocci by
incubation in NaCl 6.5% medium and Bile Aesculin agar. Christie, Atkins, Munch-Petersen
(CAMP) tests were used to differentiate aesculin-negative cocci in Streptococcus agalactiae
and Streptococcus dysgalactiae. Gram-negative bacteria were identified by colony
morphology, lactose fermentation on MacConkey agar, incubation in SIM (Sulfide-Indole-
Motility) medium, and oxidase, TSI (Triple Sugar Iron), citrate and urease testing. The API 20
E® system (bioMérieux, Marcy l'Etoile, France) was used if the abovementioned tests failed
to identify the Gram-negative bacterium. Samples yielding 2 different bacterial species were
grouped as “mixed culture”, whereas samples yielding 3 or more different bacterial species
were considered to be contaminated.
2.5 Outcome variables
The cow IRCM was calculated by dividing the number of quarter cases by the days at risk
during the study and expressed as cases per 10,000 cow-days at risk. Samples taken from the
same cow within 2 weeks from a previous case (n = 22) were not considered new cases and
therefore excluded from the analysis (Barkema et al., 1998). The at-risk period for a cow
started at the beginning of the survey or at the date of first calving and ended at the end of
the survey or at the culling date. As dry cows can suffer from clinical mastitis (Scherpenzeel
et al., 2014) and clinical mastitis in early lactation may originate from IMI established in the
nonlactating period (Bradley and Green, 2004), dry periods were included in the at-risk
period. Overall IRCM (independent of the culture results), as well as pathogen-specific IRCM
(S. aureus, S. uberis, S. dysgalactiae, and E. coli, specifically) were calculated.
2.6 Data analyses

Chapter 5.1 Clinical mastitis in Flanders
132
The association between pathogen isolation and severity was tested on a dataset
including all clinical mastitis cases (n = 845). Five different logistic regression models (PROC
LOGISTIC, SAS 9.4, SAS Institute Inc., Cary, NC) were fit with isolation as the outcome
variable [(1) no growth vs. growth, (2) S. aureus isolation vs. any other culture result, (3) S.
uberis isolation vs. any other culture result, (4) S. dysgalactiae isolation vs. any other culture
result, and (5) E. coli isolation vs. any other culture result)] and severity (mild, moderate or
severe) as categorical fixed effect. Odds ratio’s (OR) with 95% confidence intervals (95% CI)
were calculated.
The associations between herd hygiene and the different outcome variables [(1) overall
IRCM, (2) S. aureus IRCM, (3) S. uberis IRCM, (4) S. dysgalactiae IRCM, and (5) E. coli IRCM]
were determined using 5 mixed Poisson regression models (PROC GLIMMIX, SAS 9.4). All
models contained herd as a random effect to correct for clustering of cows within herds, the
natural logarithm of the number of days at risk as an offset variable (Barkema et al., 1999)
and herd hygiene (dirty vs. clean) as a categorical fixed effect. For each outcome variable
rate ratio’s (RR) with 95% CI were calculated. Confounding by parity distribution in the herd
was tested by adding the proportion of heifers in the herd (days at risk heifers/ days at risk
total) to the model as a continuous fixed effect. Parity distribution was considered to act as a
confounder if the regression coefficients of herd hygiene underwent a relative change >25%.
No confounding was detected. Overdispersion was evaluated for each model by calculating a
dispersion parameter (Pearson χ²/df) (Dohoo et al., 2003). The power to demonstrate a
significant difference between dirty and clean herds (1-β, where β is the type II error rate)
was estimated for each model using following formula (Dohoo et al., 2003): P(z> ) where z
= the standard normal variable and Zβ = the calculated boundary value, and =
, where n = 25 (herds per group), λ1 = average count of cases in clean
herds, λ2 = average count of cases in dirty herds and Zα/2 = 95th percentile of a standard
normal distribution = 1.96.
Because clinical mastitis detection in herds with an automated milking system is
different, all analyses were repeated on a subset of data excluding the three herds with an
automated milking system. Because of the limited proportion of herds with an automated
milking system, only changes in regression coefficients were calculated and reported.

Chapter 5.1 Clinical mastitis in Flanders
133
3. Results
3.1 Herd characteristics
Thirty-two (64%) herds participated in the DHI program whereas 18 did not. The average
herd size was 60 lactating cows (range 16 to 240). All producers milked Holstein-Friesian
cows. Cows were housed in free stalls with cubicles in 34 herds (68%), in free stalls with
deep litter bedding in 7 herds (14%) and in tie-stalls in 9 herds (18%). Zero-grazing was
practiced in 10 herds (20%). Cows were milked using an automated milking system in 3 herds
(6%). Forty-one producers (82%) used individual paper towels for premilking treatment
whereas 6 producers used a cotton towel (12%). Eight producers (16%) used a foaming pre-
dip. Cows were sprayed in 12 herds (24%) and dipped in 29 herds (48%) after milking.
Postmilking teat disinfection was not practiced in 9 herds (18%). In all herds, blanket dry cow
therapy (DCT) was applied. Dry cow therapy with long-acting antimicrobial agents was
combined with an internal teat sealant on all cows in 8 herds (16%) and on some cows in 10
(20%) herds. Twenty-seven (54%) producers declared never to purchase cows/young stock
whereas 23 did. During the survey, participating herds had an average bulk milk SCC of
236,000 cells/ml (range 85,000 to 453,000 cells/ml). On average 47.9% of the cows per herd
had an udder hygiene score of 3 or 4 (range 15 to 77.5%). Twenty-seven herds (54%) were
categorized as clean (≤50% of cows having udder hygiene score 3 or 4), 23 as dirty (>50% of
cows having udder hygiene score 3 or 4). The average proportion heifers in a herd (days at
risk heifers/ days at risk total) was 23.8% (range 6.0 to 45.9%).
3.2 Clinical mastitis
In total, 845 clinical mastitis cases from 692 cows were sampled by the producers. During
the survey, 490 (77.7%), 111 (17.4%), 30 (4.7%), 5 (0.8%) and 3 (0.5%) cows suffered from 1,
2, 3, 4 and 5 cases, respectively. The population was 1,192,800 cow-days at risk. In total,
1032 heifers calved during the survey and 722 animals were culled.
The IRCM was estimated at 7.1 quarter cases per 10,000 cow-days at risk for the whole
population and was lower in heifers (2.9 quarter cases per 10,000 cow-days at risk)
compared to multiparous cows (11.0 quarter cases per 10,000 cow-days at risk). However,
the IRCM in the first week after calving was higher in heifers compared to cows (Figure
5.1.1).
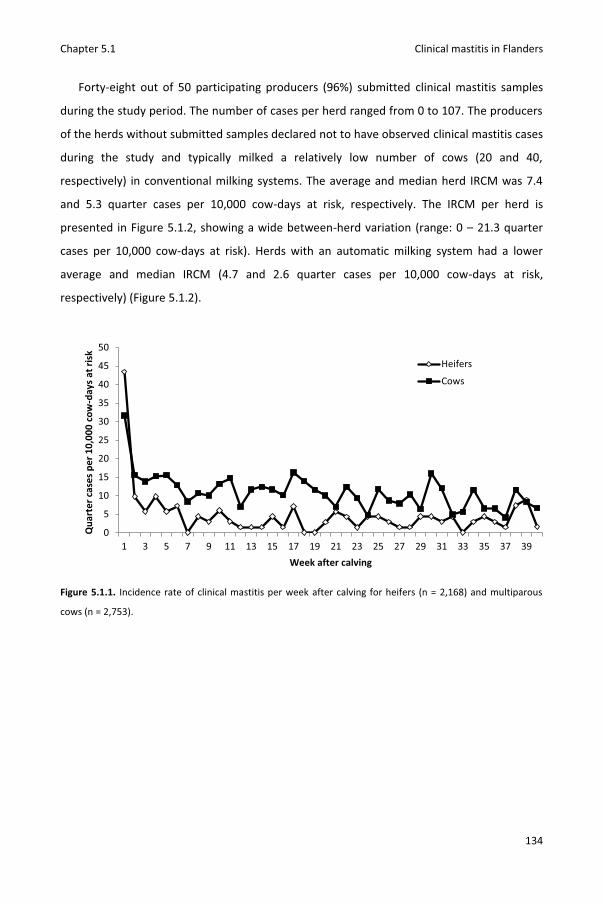
Chapter 5.1 Clinical mastitis in Flanders
134
Forty-eight out of 50 participating producers (96%) submitted clinical mastitis samples
during the study period. The number of cases per herd ranged from 0 to 107. The producers
of the herds without submitted samples declared not to have observed clinical mastitis cases
during the study and typically milked a relatively low number of cows (20 and 40,
respectively) in conventional milking systems. The average and median herd IRCM was 7.4
and 5.3 quarter cases per 10,000 cow-days at risk, respectively. The IRCM per herd is
presented in Figure 5.1.2, showing a wide between-herd variation (range: 0 – 21.3 quarter
cases per 10,000 cow-days at risk). Herds with an automatic milking system had a lower
average and median IRCM (4.7 and 2.6 quarter cases per 10,000 cow-days at risk,
respectively) (Figure 5.1.2).
Figure 5.1.1. Incidence rate of clinical mastitis per week after calving for heifers (n = 2,168) and multiparous
cows (n = 2,753).
0
5
10
15
20
25
30
35
40
45
50
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
Qua
rter
cas
es p
er 1
0,00
0 co
w-d
ays
at ri
sk
Week after calving
Heifers
Cows
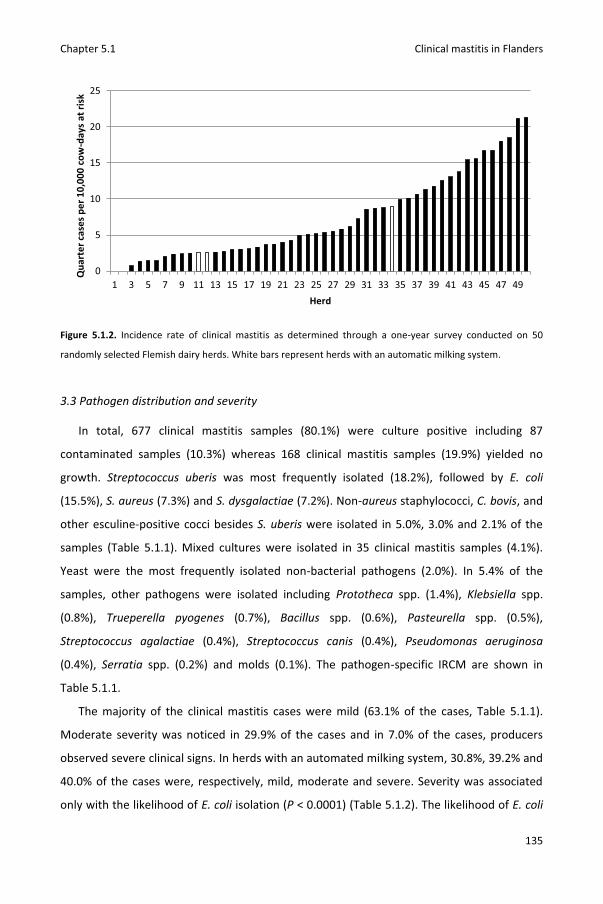
Chapter 5.1 Clinical mastitis in Flanders
135
Figure 5.1.2. Incidence rate of clinical mastitis as determined through a one-year survey conducted on 50
randomly selected Flemish dairy herds. White bars represent herds with an automatic milking system.
3.3 Pathogen distribution and severity
In total, 677 clinical mastitis samples (80.1%) were culture positive including 87
contaminated samples (10.3%) whereas 168 clinical mastitis samples (19.9%) yielded no
growth. Streptococcus uberis was most frequently isolated (18.2%), followed by E. coli
(15.5%), S. aureus (7.3%) and S. dysgalactiae (7.2%). Non-aureus staphylococci, C. bovis, and
other esculine-positive cocci besides S. uberis were isolated in 5.0%, 3.0% and 2.1% of the
samples (Table 5.1.1). Mixed cultures were isolated in 35 clinical mastitis samples (4.1%).
Yeast were the most frequently isolated non-bacterial pathogens (2.0%). In 5.4% of the
samples, other pathogens were isolated including Prototheca spp. (1.4%), Klebsiella spp.
(0.8%), Trueperella pyogenes (0.7%), Bacillus spp. (0.6%), Pasteurella spp. (0.5%),
Streptococcus agalactiae (0.4%), Streptococcus canis (0.4%), Pseudomonas aeruginosa
(0.4%), Serratia spp. (0.2%) and molds (0.1%). The pathogen-specific IRCM are shown in
Table 5.1.1.
The majority of the clinical mastitis cases were mild (63.1% of the cases, Table 5.1.1).
Moderate severity was noticed in 29.9% of the cases and in 7.0% of the cases, producers
observed severe clinical signs. In herds with an automated milking system, 30.8%, 39.2% and
40.0% of the cases were, respectively, mild, moderate and severe. Severity was associated
only with the likelihood of E. coli isolation (P < 0.0001) (Table 5.1.2). The likelihood of E. coli
0
5
10
15
20
25
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49
Qua
rter
cas
es p
er 1
0,00
0 co
w-d
ays
at ri
sk
Herd
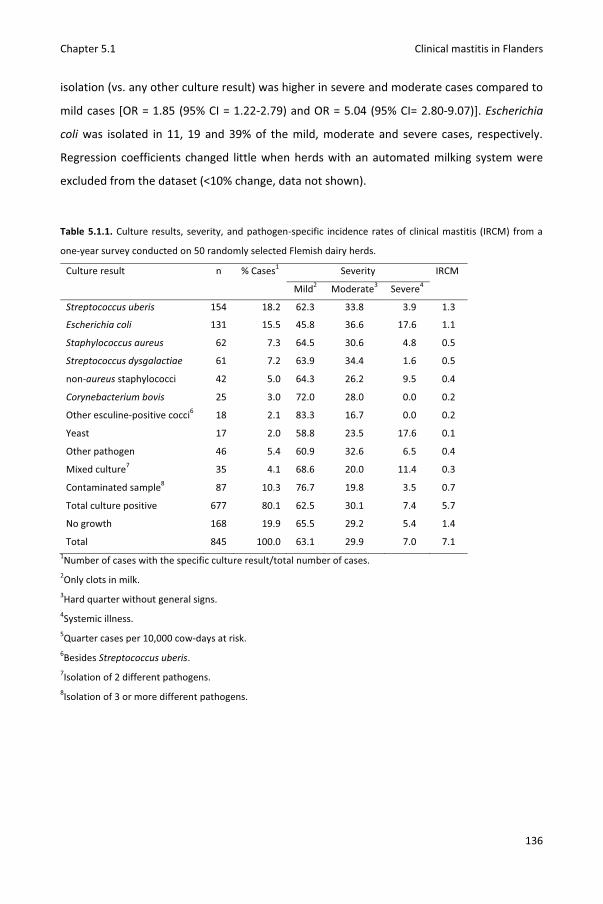
Chapter 5.1 Clinical mastitis in Flanders
136
isolation (vs. any other culture result) was higher in severe and moderate cases compared to
mild cases [OR = 1.85 (95% CI = 1.22-2.79) and OR = 5.04 (95% CI= 2.80-9.07)]. Escherichia
coli was isolated in 11, 19 and 39% of the mild, moderate and severe cases, respectively.
Regression coefficients changed little when herds with an automated milking system were
excluded from the dataset (<10% change, data not shown).
Table 5.1.1. Culture results, severity, and pathogen-specific incidence rates of clinical mastitis (IRCM) from a
one-year survey conducted on 50 randomly selected Flemish dairy herds.
Culture result n % Cases1 Severity IRCM
Mild2 Moderate3 Severe4
Streptococcus uberis 154 18.2 62.3 33.8 3.9 1.3
Escherichia coli 131 15.5 45.8 36.6 17.6 1.1
Staphylococcus aureus 62 7.3 64.5 30.6 4.8 0.5
Streptococcus dysgalactiae 61 7.2 63.9 34.4 1.6 0.5
non-aureus staphylococci 42 5.0 64.3 26.2 9.5 0.4
Corynebacterium bovis 25 3.0 72.0 28.0 0.0 0.2
Other esculine-positive cocci6 18 2.1 83.3 16.7 0.0 0.2
Yeast 17 2.0 58.8 23.5 17.6 0.1
Other pathogen 46 5.4 60.9 32.6 6.5 0.4
Mixed culture7 35 4.1 68.6 20.0 11.4 0.3
Contaminated sample8 87 10.3 76.7 19.8 3.5 0.7
Total culture positive 677 80.1 62.5 30.1 7.4 5.7
No growth 168 19.9 65.5 29.2 5.4 1.4
Total 845 100.0 63.1 29.9 7.0 7.1 1Number of cases with the specific culture result/total number of cases. 2Only clots in milk. 3Hard quarter without general signs. 4Systemic illness. 5Quarter cases per 10,000 cow-days at risk. 6Besides Streptococcus uberis. 7Isolation of 2 different pathogens. 8Isolation of 3 or more different pathogens.

Chapter 5.1 Clinical mastitis in Flanders
137
Table 5.1.2. Logistic regression models describing the association between pathogen isolation and severity of
clinical mastitis.
Pathogen isolation1 Severity2 β3 SE4 OR5 95%CI OR6 LSM7 P-value8
No growth Mild Ref. … … … 0.21 0.60
Moderat -0.08 0.19 0.9 0.63-1.34 0.19
Severe -0.37 0.38 0.6 0.33-1.45 0.15
Staphylococcus aureus Mild Ref. … … … 0.08 0.79
Moderat 0.00 0.29 1.0 0.57-1.77 0.08
Severe -0.42 0.62 0.6 0.20-2.20 0.05
Streptococcus uberis Mild Ref. … … … 0.18 0.18
Moderat 0.16 0.19 1.1 0.81-1.72 0.21
Severe -0.66 0.45 0.5 0.22-1.23 0.10
Streptococcus dysgalactiae Mild Ref. … … … 0.07 0.27
Moderat 0.14 0.28 1.1 0.66-1.99 0.08
Severe -1.52 1.02 3.3 0.03-1.62 0.02
Escherichia coli Mild Ref. … … … 0.11 <0.0001
Moderat 0.61 0.21 1.8 1.22-2.79 0.19
Severe 1.62 0.30 5.0 2.80-9.07 0.39 1Outcome variable with 1 = isolation of the pathogen and 0 = any other culture result. 2Mild = only clots in milk, moderate = hard quarter without general signs, or severe = systemic illness. 3Regression coefficient. 4Standard error. 5Odds ratio. 695% confidence interval around OR. 7Least Square Means: proportion of cases with the specific culture result compared to the total number of
cases with the specific severity. 8Overall P-value.
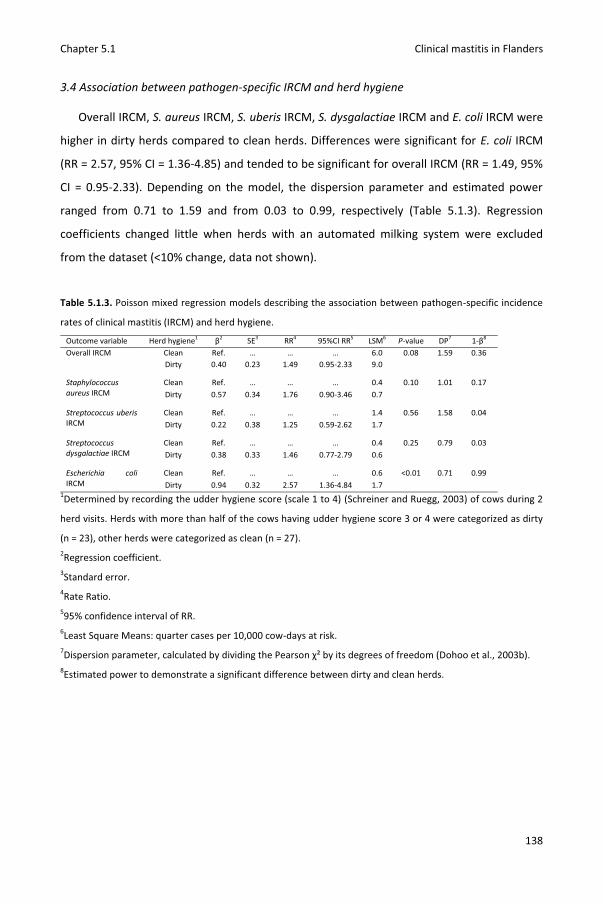
Chapter 5.1 Clinical mastitis in Flanders
138
3.4 Association between pathogen-specific IRCM and herd hygiene
Overall IRCM, S. aureus IRCM, S. uberis IRCM, S. dysgalactiae IRCM and E. coli IRCM were
higher in dirty herds compared to clean herds. Differences were significant for E. coli IRCM
(RR = 2.57, 95% CI = 1.36-4.85) and tended to be significant for overall IRCM (RR = 1.49, 95%
CI = 0.95-2.33). Depending on the model, the dispersion parameter and estimated power
ranged from 0.71 to 1.59 and from 0.03 to 0.99, respectively (Table 5.1.3). Regression
coefficients changed little when herds with an automated milking system were excluded
from the dataset (<10% change, data not shown).
Table 5.1.3. Poisson mixed regression models describing the association between pathogen-specific incidence
rates of clinical mastitis (IRCM) and herd hygiene.
Outcome variable Herd hygiene1 β2 SE3 RR4 95%CI RR5 LSM6 P-value DP7 1-β8 Overall IRCM Clean Ref. … … … 6.0 0.08 1.59 0.36 Dirty 0.40 0.23 1.49 0.95-2.33 9.0
Staphylococcus aureus IRCM
Clean Ref. … … … 0.4 0.10 1.01 0.17 Dirty 0.57 0.34 1.76 0.90-3.46 0.7
Streptococcus uberis IRCM
Clean Ref. … … … 1.4 0.56 1.58 0.04 Dirty 0.22 0.38 1.25 0.59-2.62 1.7
Streptococcus dysgalactiae IRCM
Clean Ref. … … … 0.4 0.25 0.79 0.03 Dirty 0.38 0.33 1.46 0.77-2.79 0.6
Escherichia coli IRCM
Clean Ref. … … … 0.6 <0.01 0.71 0.99 Dirty 0.94 0.32 2.57 1.36-4.84 1.7
1Determined by recording the udder hygiene score (scale 1 to 4) (Schreiner and Ruegg, 2003) of cows during 2
herd visits. Herds with more than half of the cows having udder hygiene score 3 or 4 were categorized as dirty
(n = 23), other herds were categorized as clean (n = 27). 2Regression coefficient. 3Standard error. 4Rate Ratio. 595% confidence interval of RR. 6Least Square Means: quarter cases per 10,000 cow-days at risk. 7Dispersion parameter, calculated by dividing the Pearson χ² by its degrees of freedom (Dohoo et al., 2003b). 8Estimated power to demonstrate a significant difference between dirty and clean herds.

Chapter 5.1 Clinical mastitis in Flanders
139
4. Discussion
To estimate the pathogen-specific IRCM in Flanders, 68 herds were randomly selected. A
fair response rate of 79% was achieved. Three of the initial 53 herds were excluded from the
analysis because producers admitted they did not sample all cases. Participation in the DHI
program was comparable between the study herds (64%) and all Flemish dairy herds (60%)
(K. Huijps, CRV, Alken, The Netherlands, personal communication), suggesting randomization
worked well. Still, the average herd size and bulk milk SCC were slightly higher in the study
herds (60 lactating cows and 236.000 cells/ml, respectively) compared to all Flemish dairy
herds [50 lactating cows (Federal Public Service Economy, Small and Medium Enterprises,
Self-Employed, and Energy, 2012) and 214.000 cells/ml (Annual Report 2013, Milk Control
Centre Flanders, Lier, Belgium), respectively], suggesting some selection bias. As high bulk
milk SCC was found to be associated with higher S. aureus and S. dysgalactiae IRCM and
lower E. coli IRCM (Barkema et al., 1999; Olde Riekerink et al., 2008) we might have
underestimated E. coli IRCM and overestimated S. aureus and S. dysgalactiae IRCM.
Similar to other research (Barkema et al., 1998; McDougall et al., 2007), heifers had a
lower IRCM in general but a higher IRCM in early lactation compared to multiparous cows.
Latter findings stress the importance of prevention and control of heifer mastitis (De
Vliegher et al., 2012) and the need to understand differences between pathogens associated
with this disease. The mean herd IRCM (expressed as quarter cases per 10,000 cow-days at
risk) was estimated considerably lower (7.4) than what could have been expected from a
recent internet questionnaire indicating 46% of the Flemish dairy cows to suffer at least once
per year from clinical mastitis (P. Passchyn, S. Piepers and S. De Vliegher, Ghent University,
Ghent, Belgium, personal communication). Based on the internet questionnaire, one would
expect at least 1380 cases (46%*60 lactating cows*50 herds), whereas we collected 845
samples. Because methodology and especially selection criteria differ between studies,
caution is required in comparing results between clinical mastitis surveys (Olde Riekerink et
al., 2008), especially as the producers participating in the aforementioned online
questionnaire estimated the IRCM in the lack of accurate data as the majority admitted they
were not keeping disease records. Yet, our figure differed little from figures estimated by
Barkema et al. (1998) in the Netherlands (7.6, 7.0 and 6.9 for herds with low, medium and
high bulk milk SCC, respectively) and Wolff et al. (2012) in Sweden (7.2). Incidence rate of

Chapter 5.1 Clinical mastitis in Flanders
140
clinical mastitis was estimated lower in France and Canada (5.5 and 6.4, respectively), but
higher in England and Wales (12.9), Norway (8.6), Finland (10.6) and Denmark (12.8)
(Barnouin et al., 2005; Bradley et al., 2007; Olde Riekerink et al., 2008; Wolff et al., 2012).
Herds were randomly selected in some studies (Barkema et al., 1998; Bradley et al., 2007;
Wolff et al., 2012) and conveniently selected in others (Barnouin et al., 2005; Olde Riekerink
et al., 2008). In contrast to the studies in which the animals were randomly selected as well,
no inclusion criteria were applied in our study, underlining the external validity of our data.
The pathogen distribution in this survey corresponded with those of Flemish clinical
mastitis samples submitted to the laboratories of both Milk Control Centre Flanders (n =
4,468) and the M-team (n = 329) (Annual Reports 2012, Milk Control Centre Flanders, Lier,
Belgium and M-team, Ghent University, Belgium). In the three datasets, E. coli and aesculin-
positive cocci including S. uberis were the most frequently isolated pathogens. Similar to
other regions in the world (Bradley et al., 2007; Oliveira et al., 2013), environmental
pathogens appear to be the most common cause of clinical mastitis in Flanders.
Staphylococcus aureus was less frequently isolated in our study (7.3% of the samples)
compared to clinical mastitis studies conducted in Canada (10.3% of the samples) (Olde
Riekerink et al., 2008) and Ireland (23% of the samples) (Keane et al., 2013), suggesting that
a different focus in clinical mastitis prevention is required per country. Both in this and in
other studies (Bradley et al., 2007; Olde Riekerink et al., 2008; Keane et al., 2013; Oliveira et
al., 2013), a relative large proportion of cases were culture-negative. Spontaneous cure, low
bacterial viability and pathogens not growing in standard culture media (Mycoplasma spp.)
might explain this high proportion (Taponen et al., 2009). Freezing and thawing of the
samples might have decreased the culture sensitivity of E. coli (Schukken et al., 1989) but
was required to allow convenient transportation of samples by the herd veterinarians. As
the sensitivity of detecting IMI using a single milk sample was found to be higher for S.
aureus compared to other pathogens (Dohoo et al., 2011) we might have overestimated the
S. aureus IRCM compared to the IRCM by other pathogens.
Clinical signs were mostly mild. However, we should mention that the clinical signs were
recorded by the producers who might have missed subtle signs of systemic illness (e.g. fever)
in some cases. Compared to a clinical mastitis study in large herds in Wisconsin (US) (Oliveira
et al., 2013), the proportion of moderate and severe cases was smaller (29.9 and 7% vs. 36.9
and 15.3%, respectively). Still, in the latter study, E. coli was more frequently isolated

Chapter 5.1 Clinical mastitis in Flanders
141
compared to our study (21.6 vs. 15.3% of the cases). Additionally, as producers were
rewarded per collected milk sample to keep them motivated, it is not unlikely that very mild
cases in our study were sampled. Nevertheless, cases might also have been missed.
Especially in dry cows and herds with an automated milking system where detection
intensity could potentially have been less resulting in some bias and an underestimation of
the true IRCM. Isolation of E. coli was more likely in moderate and severe cases compared to
mild cases. Still, E. coli could only be isolated in less than half of the severe cases. Using “E.
coli mastitis” as synonym for severe mastitis should for that reason be discouraged. Instead,
dairy producers and herd veterinarians should be informed that other pathogens besides E.
coli can cause severe clinical mastitis and that E. coli can also cause mild clinical mastitis
cases. Bacteriological culture of mild, moderate and severe cases is required to estimate the
herd pathogen distribution and a basis for an effective herd-specific mastitis treatment and
control plan.
The accuracy of measuring herd hygiene by scoring the udder hygiene of a limited
number of cows during two herd visits can be debated. Nevertheless, we observed a higher
overall and E. coli IRCM in dirty herds compared to clean herds which corresponds well with
British research demonstrating a higher risk of clinical mastitis in general and E. coli clinical
mastitis in specific in cows with dirty udders (Breen et al., 2009). However, and in contrast to
the British study, we cannot claim that dirty cows were more likely to have clinical mastitis
because hygiene was measured at the herd level potentially causing ecological bias (Dohoo
et al., 2003). Yet, evaluating and improving udder and herd hygiene could reduce IRCM in
Flanders. Interestingly, results in both studies demonstrated a much stronger association for
E. coli clinical mastitis compared to S. uberis clinical mastitis. Although the power to detect a
difference in S. uberis IRCM between dirty and clean herds in our study was very low and
although S. uberis is considered as an environmental pathogen, cow-to-cow transmission
might occur (Zadoks et al., 2003). We speculate that on a number of the herds included in
our study cow-adapted S. uberis strains were causing clinical mastitis. At least, our findings
indicate that improving hygiene will reduce the number of E. coli cases but will not have a
large impact on S. uberis IRCM.

Chapter 5.1 Clinical mastitis in Flanders
142
5. Conclusions
The mean and median IRCM in Flemish dairy herds was estimated at 7.4 and 5.3 quarter
cases/10,000 cow-days at risk and showed a high between-herd variation as indicated by the
wide range (0-21.3). The IRCM of heifers compared to multiparous cows was lower
throughout the entire lactation yet higher in early lactation. Streptococcus uberis and E. coli
were the most frequently isolated pathogens. Clinical signs were mild in most cases.
Isolation of E. coli was more likely in moderate and severe cases compared to mild cases.
Yet, less than half of the severe cases had E. coli as culture result. Overall and E. coli IRCM
were higher in dirty compared to clean herds.
6. Acknowledgements
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen), the Flemish cattle monitoring project
(‘Veepeiler Rund’), headed by the Flemish Animal Health Service (Dierengezonheidszorg-
Vlaanderen), as well as the Flemish Government (Departement Landbouw en Visserij,
Afdeling Duurzame Landbouwontwikkeling). The authors wish to thank the Cattle Breeding
Association (CRV) for access to DHI records, Koen Lommelen (Milk Control Centre Flanders)
for collecting bulk milk SCC data, and Jo Maris and Hans Van Loo (Dierengezondheidszorg-
Vlaanderen) for access to cow records of the identification and registration system. Last but
not least, we thank all participating producers and herd veterinarians.
7. References
Bannerman, D. D., M. J. Paape, J. W. Lee, X. Zhao, J. C. Hope, and P. Rainard. 2004.
Escherichia coli and Staphylococcus aureus elicit differential innate immune responses
following intramammary infection. Clin. Diagn. Lab. Immunol. 11:463-472.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, H. Wilmink, G. Benedictus, and A.
Brand. 1998. Incidence of clinical mastitis in dairy herds grouped in three categories by
bulk milk somatic cell counts. J. Dairy Sci. 81:411-419.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, G. Benedictus, and A. Brand. 1999.
Management practices associated with the incidence rate of clinical mastitis. J. Dairy Sci.
82:1643-1654.

Chapter 5.1 Clinical mastitis in Flanders
143
Barnouin, J., S. Bord, S. Bazin, and M. Chassagne. 2005. Dairy management practices
associated with incidence rate of clinical mastitis in low somatic cell score herds in
France. J. Dairy Sci. 88:3700-3709.
Bradley, A. J. 2002. Bovine Mastitis: An Evolving Disease. Vet. J. 164:116-128.
Bradley, A. J., and M. J. Green. 2004. The importance of the nonlactating period in the
epidemiology of intramammary infection and strategies for prevention. Vet. Clin. North
Am. Food Anim. Pract. 20:547-568.
Bradley, A. J., K. A. Leach, J. E. Breen, L. E. Green, and M. J. Green. 2007. Survey of the
incidence and aetiology of mastitis on dairy farms in England and Wales. Vet. Rec.
160:253-258.
Breen, J. E., M. J. Green, and A. J. Bradley. 2009. Quarter and cow risk factors associated with
the occurrence of clinical mastitis in dairy cows in the United Kingdom. J. Dairy Sci.
92:2551-2561.
Burvenich, C., V. Van Merris, J. Mehrzad, A. Diez-Fraile, and L. Duchateau. 2003. Severity of
E. coli mastitis is mainly determined by cow factors. Vet. Res. 34:521-564.
De Vliegher, S., L. K. Fox, S. Piepers, S. McDougall, and H. W. Barkema. 2012. Invited review:
Mastitis in dairy heifers: nature of the disease, potential impact, prevention, and control.
J. Dairy Sci. 95:1025-1040.
Dohoo, I., M. Wayne, and H. Stryhn. 2003. Sample-size determination. Pages 39-49,
Modelling Count and Rate Data. Pages 445-466, and Ecological and Group-level Studies.
Pages 773-788 in Veterinary Epidemiologic Research. I. Dohoo, W. Martin, and H. Stryhn,
ed. AVCInc., Charlottetown, Canada.
Dohoo, I., J. Smith, S. Andersen, D. F. Kelton, and S. Godden. 2011. Diagnosing
intramammary infections: Evaluation of definitions based on a single milk sample. J.
Dairy Sci. 94:250-261.
Federal Public Service Economy, Small and Medium Enterprises, Self-Employed, and Energy.
2011. Landbouwtelling 2010. Accessed Jul. 20, 2012.
http://statbel.fgov.be/nl/binaries/3.1.3.6.1.Landbouw%20definitief%20T%202010%2005
_tcm325-118744.xls
Keane, O. M., K. E. Budd, J. Flynn, and F. McCoy. 2013. Pathogen profile of clinical mastitis in
Irish milk-recording herds reveals a complex aetiology. Vet. Rec. 173:17.

Chapter 5.1 Clinical mastitis in Flanders
144
Ma, Y., C. Ryan, D. M. Barbano, D. M. Galton, M. A. Rudan, and K. J. Boor. 2000. Effects of
somatic cell count on quality and shelf-life of pasteurized fluid milk. J. Dairy Sci. 83:264-
274.
McDougall, S., K. E. Agnew, R. Cursons, X. X. Hou, and C. R. Compton. 2007. Parenteral
treatment of clinical mastitis with tylosin base or penethamate hydriodide in dairy cattle.
J. Dairy Sci. 90:779-789.
Milk Control Centre Flanders. 2012. Annual Report. Milk Control Centre Flanders, Lier,
Belgium.
Milk Control Centre Flanders. 2013. Annual Report. Milk Control Centre Flanders, Lier,
Belgium.
National Mastitis Council. 1999. Laboratory Handbook on Bovine Mastitis. Revised Edition.
National Mastitis Council. Inc., Madison, WI.
Oliveira, L., C. Hulland, and P. L. Ruegg. 2013. Characterization of clinical mastitis occurring in
cows on 50 large dairy herds in Wisconsin. J. Dairy Sci. 96:7538-7549.
Piepers, S., L. De Meulemeester, A. de Kruif, G. Opsomer, H. W. Barkema, and S. De Vliegher.
2007. Prevalence and distribution of mastitis pathogens in subclinically infected dairy
cows in Flanders, Belgium. J. Dairy Res. 74:478-483.
Olde Riekerink, R. G. M., H. W. Barkema, D. F. Kelton, and D. T. Scholl. 2008. Incidence rate
of clinical mastitis on Canadian dairy farms. J. Dairy Sci. 91:1366-1377.
Pinzón-Sánchez, C., and P. L. Ruegg. 2011. Risk factors associated with short-term post-
treatment outcomes of clinical mastitis. J. Dairy Sci. 94:3397-3410.
Santos, M. V., Y. Ma, and D. M. Barbano. 2003. Effect of somatic cell count on proteolysis
and lipolysis in pasteurized fluid milk during shelf-life storage. J. Dairy Sci. 86:2491-2503.
Scherpenzeel, C. G. M., I. E. M. den Uijl, G. van Schaik, R. G. M. Olde Riekerink, and T. J. G. M.
Lam. 2014. Evaluation of the use of dry cow antibiotics in low somatic count cows. J.
Dairy Sci. 97:3606-3614.
Schreiner, D. A., and P. L. Ruegg. 2003. Relationship between udder and leg hygiene scores
and subclinical mastitis. J. Dairy Sci. 86:3460-3465.
Schukken, Y. H., J. A. H. Smit, F. J. Grommers, D. Vandegeer, and A. Brand. 1989. Effect of
freezing on bacteriologic culturing of mastitis milk samples. J. Dairy Sci. 72:1900-1906.
Schukken, Y. H., J. Gunther, J. Fitzpatrick, M. C. Fontaine, L. Goetze, O. Holst, J. Leigh, W.
Petzl, H. J. Schuberth, A. Sipka, D. G. E. Smith, R. Quesnell, J. Watts, R. Yancey, H. Zerbe,

Chapter 5.1 Clinical mastitis in Flanders
145
A. Gurjar, R. N. Zadoks, and H. M. Seyfert. 2011. Host-response patterns of
intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 144:270-289.
Smith, K. L., and J. S. Hogan. 1993. Environmental Mastitis. Vet. Clin. North Am. Food Anim.
Pract. 9:489-498.
Taponen, S., L. Salmikivi, H. Simojoki, M. T. Koskinen, and S. Pyorala. 2009. Real-time
polymerase chain reaction-based identification of bacteria in milk samples from bovine
clinical mastitis with no growth in conventional culturing. J. Dairy Sci. 92:2610-2617.
Wolff, C., M. Espetvedt, A. K. Lind, S. Rintakoski, A. Egenvall, A. Lindberg, and U. Emanuelson.
2012. Completeness of the disease recording systems for dairy cows in Denmark,
Finland, Norway and Sweden with special reference to clinical mastitis. BMC Vet. Res.
8:131.
Zadoks, R. N., B. E. Gillespie, H. W. Barkema, O. C. Sampimon, S. P. Oliver, and Y. H.
Schukken. 2003. Clinical, epidemiological and molecular characteristics of Streptococcus
uberis infections in dairy herds. Epidemiol. Infect. 130:335-349.

146

147
Chapter 5.2
Associations between CXCR1 polymorphisms and
pathogen-specific incidence rate of clinical mastitis,
test-day somatic cell count and test-day milk yield
J. Verbeke1, M. Van Poucke2, L. Peelman2, S. Piepers1, and S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine, Ghent
University, Belgium
Journal of Dairy Science, 97: 7927-7939.

Chapter 5.2 The CXCR1 gene and clinical mastitis
148

Chapter 5.2 The CXCR1 gene and clinical mastitis
149
Abstract
The CXCR1 gene plays an important role in the innate immunity of the bovine mammary
gland. Associations between single nucleotide polymorphisms (SNPs) CXCR1c.735C>G and
c.980A>G and udder health have been identified before in small populations. A fluorescent
multiprobe PCR assay was designed specifically and validated to genotype both SNPs
simultaneously on a reliable and cost-effective manner. In total, 3,106 cows from 50
commercial Flemish dairy herds were genotyped using this assay. Associations between
genotype and detailed phenotypic data including pathogen-specific incidence rate of clinical
mastitis (IRCM), test-day somatic cell count (SCC) and test-day milk yield were analyzed.
Staphylococcus aureus IRCM tended to associate with SNP c.735C>G. Cows with genotype
c.735GG had lower S. aureus IRCM compared to cows with genotype c.735CC [rate ratio (RR)
= 0.35, 95% confidence interval (CI) 0.14-0.90]. Additionally, a parity-specific association
between S. aureus IRCM and SNP c.980A>G was detected. Heifers with genotype c.980GG
had a lower S. aureus IRCM compared to heifers with genotype c.980AG (RR = 0.15, 95% CI
0.04-0.56). Differences were less pronounced in multiparous cows. Associations between
CXCR1 genotype and SCC were not detected. However, milk yield was associated with SNP
c.735C>G. Cows with genotype c.735GG out-produced cows with genotype c.735CC by 0.8 kg
milk/day. Results provide a basis for further research on the relation between CXCR1
polymorphism and pathogen-specific mastitis resistance and milk yield.
Keywords
CXCR1 polymorphism, dairy cow, udder health, milk yield

Chapter 5.2 The CXCR1 gene and clinical mastitis
150
1. Introduction
Mastitis, an inflammation of the mammary gland mostly caused by bacterial
intramammary infection (IMI), impairs milk quality and production, and increases the culling
probability of dairy cows substantially (Beaudeau et al., 1995; Hortet et al., 1999; Ma et al.,
2000; Santos et al., 2003; Seegers et al., 2003). The disease can be accompanied by local or
systemic symptoms (clinical mastitis) or presents itself without visible symptoms (subclinical
mastitis ). In absence of clinical symptoms, mastitis can be detected by an increase in the
concentration of somatic cells in milk (Schukken et al., 2003).
Resistance to mastitis is genetically determined (Detilleux, 2009) making a decrease in
the incidence of mastitis by breeding a possibility (Windig et al., 2010). Identification of
genetic polymorphisms linked with mastitis resistance would allow evaluation of the genetic
make-up of male and female breeding animals for udder health, even before their offspring
is born (Ogorevc et al., 2009). Polymorphisms in genes involved in the recognition of
pathogens, recruitment of immune cells towards site of infection, elimination of the
pathogens, and resolution of the inflammatory response are potential genetic markers for
mastitis resistance (Pighetti and Elliott, 2011). In a recent genome-wide association study,
polymorphisms located near CXCR1 and CXCR2 were highly associated with occurrence of
clinical mastitis (Sodeland et al., 2011). The latter genes encode the two receptors present
on neutrophils for interleukin 8 (IL-8, CXCL-8), named chemokine (C-X-C motif) receptor 1
and 2 (CXCR1 and CXCR2). Binding of IL-8 causes chemotaxis, increases activity and inhibits
apoptosis of neutrophils (Barber and Yang, 1998; Kettritz et al., 1998; Mitchell et al., 2003).
Polymorphisms in the coding region of CXCR1 have been described (Pighetti et al., 2012;
Verbeke et al., 2012). Eight of the detected SNPs were non-synonymous. The most
frequently studied SNP located at position 735 relative to the start codon [c.735C>G
(rs208795699)] causes an amino acid change (p.His245Glu) in the third intracellular loop of
the receptor and was in full linkage disequilibrium with non-synonymous SNPs c.37A>T
(rs380621468), c.38T>A (rs110296731) and c.68G>A (rs133273369) causing changes in the
N-terminus (p.Ile13Tyr and p.Gly23Glu). Single nucleotide polymorphisms c.980A>G
(rs43323012) and c.995A>G (rs43323013) cause changes in the C-terminus (p.Lys327Arg and
p.His332Arg) and were in full linkage disequilibrium with SNP c.337G>A (rs207564206)
causing a change in the first extracellular loop (p.Val113Ile). The SNP c.365T>C

Chapter 5.2 The CXCR1 gene and clinical mastitis
151
(rs211042414) encodes for p.Val122Ala in the third transmembrane domain (Verbeke et al.,
2012). Amino acid changes potentially influence receptor activity through ligand binding, G-
protein related signaling, and/or receptor internalization (reviewed in Pighetti et al., 2012).
Functional effects have yet to be studied. Additionally, (linked) polymorphisms could affect
gene expression (Leyva-Baca et al., 2008a). Because of the results of the genome wide
association study and the important innate immune function of CXCR1, it is reasonable to
believe that CXCR1 polymorphisms alter mastitis resistance.
Youngerman et al. (2004) detected associations between SNP c.735C>G and the
incidence of subclinical masitis, somatic cell score (SCS), and milk yield. Holstein c.735CC
cows showed a higher incidence of subclinical masitis and milk yield compared to c.735GG
cows. Additionally, c.735GG cows had lower SCS compared to c.735CG cows. The association
with SCS was confirmed later in an Irish cow population (Beecher et al., 2010). Galvao et al.
(2011) detected lower milk yield and higher incidence rate of clinical mastitis (IRCM) in
c.735GG cows. Because innate immune responses heavily depend on the invading mastitis
pathogen (Bannerman et al., 2004; Schukken et al., 2011), associations between
polymorphisms in innate immune genes and udder health are ideally studied using
pathogen-specific data. Recently, we revealed a pathogen-group specific association
between SNP c.980A>G and the odds of IMI in early lactating heifers: c.980AG heifers were
less likely to have IMI due to major mastitis pathogens (e.g. Staphylococcus aureus and
Streptococcus uberis) compared to c.980GG heifers but were not less likely to have IMI by
coagulase-negative staphylococci still considered as minor pathogens (Verbeke et al., 2012).
In this study, associations between SNPs c.735C>G and c.980A>G, and pathogen-specific
IRCM, test-day SCC and test-day milk yield were studied using a fluorescent multiprobe PCR
assay that was specifically designed and validated to genotype a large number of cows in a
reliable, fast and cost-effective manner.
2. Materials and methods
2.1 Herds and animals
Sixty-seven Flemish dairy producers were invited to participate in a one-year cohort
study to estimate the pathogen-specific IRCM in Flanders (Verbeke et al., 2014). All
participating herds (n = 53) were visited at the beginning of the study in September or

Chapter 5.2 The CXCR1 gene and clinical mastitis
152
October 2012 by the first author. At that time, the study details were discussed with the
producers and herd veterinarians, and blood samples of all lactating heifers and cows were
taken. From then on, producers were asked to take a milk sample of each quarter showing
signs of clinical mastitis during the 12 month study period (see further). To be able to include
data of early lactating heifers, participating herds were revisited between February and
March 2013 also by the first author. At that time, heifers that had calved between both visits
were blood sampled. Data of 3 herds were omitted from the analysis along the study period
because producers stopped sampling cases halfway the study.
In total, 3,106 animals from 50 herds were blood sampled ranging between 18 and 250
per herd. Cow-level records [birth date, calving date(s), parity, culling date, sire and dam
(both when available)] were retrieved from dairy herd improvement (DHI) records for herds
participating in the DHI program organized by the Cattle Breeding Organization (CRV,
Oosterzele, Belgium) (n = 32) or from the identification and registration system of the Animal
Health Service Flanders (Dierengezondheidszorg Vlaanderen, Drongen, Belgium) for other
herds (n = 18).
2.2 Phenotypic records
Clinical Mastitis (full dataset, 50 herds, 3,106 cows). Producers were asked to take a
milk sample from each quarter showing signs of clinical mastitis during the 12 month survey
period. Signs of clinical mastitis were defined as visible abnormalities of the udder or milk
indicating udder inflammation. Sampling date and cow identification were recorded.
Samples were frozen on-farm and collected by the herd veterinarian. A courier of the Animal
Health Service Flanders transported the samples from the veterinary practice to the Milk
Control Centre Flanders (Lier, Belgium) where bacteriological culture was performed
according to National Mastitis Council (NMC) guidelines (National Mastitis Council, 1999).
From each thawed sample, 10 μL of milk was spread on blood-aesculin and MacConkey’s
agar and incubated aerobically for 24-48 h at 37 °C. Identification of bacteria was done by
Gram-staining, inspection of the colony morphology and biochemical testing. Catalase tests
were performed to differentiate Gram-positive cocci in catalase-positive or catalase-negative
cocci. Staphylococci were identified as Staphylococcus aureus or non-aureus staphylococci,
referred to as Staphylococcus spp. throughout this paper, by colony morphology, haemolysis

Chapter 5.2 The CXCR1 gene and clinical mastitis
153
patterns, and DNase tests. Streptococci were differentiated in aesculin-positive and aesculin-
negative streptococci (Streptococcus agalactiae and Streptococcus dysgalactiae).
Streptococcus uberis was distinguished from other aesculin-positive cocci by incubation in
NaCl 6.5% medium and Bile Aesculin agar. Christie, Atkins, Munch-Petersen (CAMP) tests
were used to differentiate Streptococcus agalactiae and Streptococcus dysgalactiae. Gram-
negative bacteria were identified by colony morphology, lactose fermentation on
MacConkey agar, incubation in SIM (Sulfide-Indole-Motility) medium, and oxidase, TSI (Triple
Sugar Iron), citrate and urease testing. The API 20 E® system (bioMérieux, Craponne, France)
was used if the abovementioned tests failed to identify the Gram-negative bacterium.
Samples yielding 2 different bacterial species were grouped as “mixed culture”, whereas
samples yielding 3 or more different bacterial species were considered to be contaminated.
The lactation-level IRCM was calculated by dividing the number of quarter cases by the
days at risk during the lactation within the study and expressed as cases per 10,000 cow-
days at risk. Samples taken from the same cow within 2 weeks from a previous case were
not considered new cases and therefore excluded from the analysis (Barkema et al., 1998).
The number of days at risk for each lactation was equal to the time between the start of the
study/ calving and the end of the study/ culling date/ the next calving minus 14d per case.
Overall IRCM (all cases), as well as pathogen-specific IRCM (S. aureus, S. uberis, S.
dysgalactiae and E. coli, specifically) were calculated (Verbeke et al., 2014).
Test-day Somatic Cell Count and Milk Yield (subset of data, 32 herds, 1,961 cows).
Composite milk SCC (SCC, cells per ml) and milk yield (kg of milk) at test-day per cow were
available from DHI records of all herds participating in the DHI program (n = 32 herds).
Records between 14 and 285 DIM from cows with at least 2 measurements per parity were
saved in a dataset.
2.3 CXCR1 genotyping
Genotype at SNPs c.735C>G and c.980A>G was determined using a multiprobe PCR assay
similar to the assay described by Van Poucke and colleagues (2005). DNA was extracted from
blood samples (100 μl) using a proteinase K digestion method (Van Poucke et al., 2005).
Three PCR primers and 4 dual-labelled ASO (Allele-Specific Oligonucleotide) probes were
designed and synthesized by Sigma-Aldrich (St. Louis, MO). Sequences, Tm [calculated using

Chapter 5.2 The CXCR1 gene and clinical mastitis
154
OligoAnalyzer 3.1 (Integrated DNA Technologies, Coralville, IO)], and binding positions are
presented in Table 5.2.1. Regions forming potential secondary structures or including repeat
sequences were identified with Mfold (Zuker, 2003) and RepeatMasker (Smit et al., 2010),
respectively, and avoided. The forward primer was designed to bind CXCR1 specifically.
Because of SNP *6C>A (db SNP ss 974294528), *7A>C (db SNP ss 974294529) and *10C>T
(db SNP ss 974294530), two reverse primers were used. Homology between CXCR1 and
CXCR2, CXCR1 SNP and binding positions of primers and probes are visualized in Figure 5.2.1.
In order to amplify a region including both SNPs specifically, a relatively large amplicon was
multiplied (523 bp).
Table 5.2.1. Primers and probes used in the real-time PCR assay for genotyping SNPs CXCR1c.735C>G and
CXCR1c.980A>G.
Primer/Probe Name Sequence (5’ → 3’)1 Length (bp) Tm (°C)2,3 avTm(°C)2,4 Binding position
Forward primer TCCGACCTAGTCTGCTA 17 59.6 N/A c.574→c.590
Reverse primer 1 CCGGTGTGGAGTCTC 15 59.2 N/A c.1082←*13
Reverse primer 2 CGATGGTGAGTCTCAGA 17 58.7 58.7 c.1079←*12
C735-probe HEX-CGGTGCTTGTGCCC-BHQ1 14 61.6 55.3 c.730←c.743
G735-probe FAM-CGGTGCTTCTGCCCC-BHQ1 15 64.0 54.2 c.729←c.743
A980-probe Cy5-CCATGATCTTGAGGAGTCC-BHQ2 19 60.3 56.5 c.970←c.988
G980-probe TexasRed-CCATGATCCTGAGGAGTCC-BHQ2 19 62.2 55.0 c.970←c.988 1Fluorescent labels and quenchers are in italic. SNPs are underlined. 2Melting temperatures were calculated using OligoAnalyzer 3.1, Integrated DNA Technologies (Coralville, CA).
3Melting temperature when annealed to CXCR1 for primers and to target variant for probes. 4Melting temperature when annealed to CXCR2 for primers and to allelic variant for probes.
The PCR was performed in the CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad,
Hercules, CA) with a 10 μl PCR mix containing approximately 100 ng genomic DNA, 1.0 μl
10×FastStart Taq DNA Polymerase Buffer (Roche Applied Science, Penzberg, Germany), dNTP
Mix (0.2 mM each; 5 PRIME), 0.5 μM forward primer (Integrated DNA Technologies), 0.5 μM
reverse primer 1 (Integrated DNA Technologies), 0.5 μM reverse primer 2 (Integrated DNA
Technologies), 0.6 μM C735-probe, 0.3 μM G735-probe, 0.4 μM A980-probe, 0.2 μM G980-
probe and 0.5 U Taq DNA Polymerase (Roche Applied Science). The real-time PCR program
consisted of an initiation step of 3’30” at 95°C, followed by 40 amplification cycles
(denaturation for 30” at 95°C, annealing-elongation for 40” at 60°C and detection of
fluorescent signals generated by cleavage of the dual-labeled probes).

Chapter 5.2 The CXCR1 gene and clinical mastitis
155
Quantification cycles (Cq values) for each probe were analyzed using CFX Manager™
Software v3.1 (Bio-Rad). In each run, positive controls with genotype c.735CC980AA,
c.735CG980AG, and c.735GG980GG and a no template control were included to assign
Baseline Cycles and Treshold position, and check for PCR contamination. Potential unspecific
amplification of CXCR2 was analyzed by digesting a PCR amplicon with restriction enzyme
BsrD1 (New England Biolabs, Ipswich, MA) according to the manufacturer’s protocol. In silico
analysis showed that BsrD1 cleaves the predicted CXCR1 region but not the homologous
CXCR2 region (NM_001101285.1) (Figure 5.2.1). The sample was run on ethidium bromide-
stained agarose (2%) gel (135 V, 40’). None of the original fragments was visible indicating
that no CXCR2 was multiplied. To validate the genotyping method, 26 DNA samples
genotyped by direct sequencing (Verbeke et al., 2012) were tested in a blind manner. At
least 3 samples of genotype c.735CC980AA, c.735CC980AG, c.735CC980GG, c.735CG980AG,
c.735CG980GG, and c.735GG980GG were included. Results were concordant for all 26
samples. Real-time amplification plots for all detected genotypes are presented in Figure
5.2.2.

Chapter 5.2 The CXCR1 gene and clinical mastitis
156
Figure 5.2.1. The CDS and 13 bp downstream the STOP codon of bovine CXCR1 as detected by DNA sequencing
(Verbeke et al., 2012) was aligned with bovine CXCR2 (AC_000159 REGION: 106913181..106914285) using
ClustalX2 (Larkin et al., 2007). The start codon, SNPs in CXCR1 and stop codon are indicated. Binding positions
of primers and probes used for CXCR1 genotyping (see Table 1) are marked with arrows.

Chapter 5.2 The CXCR1 gene and clinical mastitis
157
Figure 5.2.2. Amplification plots obtained with the fluorescent multiprobe PCR assay for bovine CXCR1
genotyping of seven cows with different genotype at SNPs CXCR1c.735C>G or CXCR1c.980A>G. Allele c.735C,
c.735G, c.980A and c.980G were detected using probes labelled with HEX, FAM, Cy5 and TexasRed,
respectively. Genotype c.735GG980AA and c.735GG980AG were not detected in the study population. RFU
stands for relative fluorescence units.

Chapter 5.2 The CXCR1 gene and clinical mastitis
158
2.4 Statistical analysis
Deviations from Hardy-Weinberg equilibrium were analyzed using chi-square goodness
of fit tests. Chi-square values were calculated to analyze linkage disequilibrium between loci.
The relative value of disequilibrium, d, was calculated as introduced by Kaplan and Weir
(1992).
In all models, statistical significance was assessed at P ≤ 0.05 whereas a P-value between
0.05 and 0.10 was considered as a tendency towards significance. Non-significant fixed
effects except genotype as variable of main interest were removed from the models.
Genotype c.735CC and c.980AA encoding the conserved amino acids p.245Glu and p.327Lys
(Pighetti et al., 2012) were set as reference groups in the models with c.735C>G and
c.980A>G, respectively. Other genotypes were compared with the reference (2 independent
post-hoc tests).
Clinical Mastitis. Associations between SNPs c.735C>G and c.980A>G and 5 different
outcome variables (1) overall IRCM, (2) S. aureus IRCM, (3) S. uberis IRCM, (4) S. dysgalactiae
IRCM and (5) E. coli IRCM were determined in a number of models using multilevel Poisson
regression (MlwiN 2.16, Centre for Multilevel Modeling, Bristol, UK). Herd and cow were
added as a random effect to correct for clustering of cows within herds and lactations within
cows. The models included the cows’ genotype (c.735C>G and c.980A>G, respectively) at the
position of the polymorphism, parity (1, 2, and ≥3), season of calving (January-March, April-
June, July-September, and October-December) and the interaction between genotype and
parity as categorical fixed effects and the natural logarithm of the number of days at risk as
an offset variable (Barkema et al., 1999). To avoid non-convergence of the statistical models,
records on rare genotypes (c.980AA, see further) were omitted in the pathogen-specific
models. In latter models, c.980AG was set as reference. Results are presented as rate ratio’s
(RR) with 95% confidence intervals (95% CI).
Test-day SCC and milk yield. Associations between SNP c.735C>G and c.980A>G
(predictor variables of main interest) and test-day SCC and milk yield (separate outcome
variables) were determined using 2 mixed regression models per outcome variable with
herd, cow and lactation as random effects to correct for clustering of cows within herds,
lactations within cows and multiple observations per lactation, respectively (MlwiN 2.16,
Centre for Multilevel Modeling, Bristol, UK). A natural logarithmic transformation of SCC

Chapter 5.2 The CXCR1 gene and clinical mastitis
159
(LnSCC) was performed to obtain a normalized distribution. The models included DIM and
the quadratic term for DIM as continuous fixed effects and the cows’ genotype (c.735C>G
and c.980A>G, respectively) at the position of the polymorphism, parity (1, 2, and ≥3),
season of calving (January-March, April-June, July-September, and October-December),
season of the test-day (January-March, April-June, July-September, and October-December)
and the interaction between genotype and parity as categorical fixed effects.
3. Results
3.1 Phenotypic data
The average herd size was 60 lactating cows (range 16 to 240). All producers milked
Holstein-Friesian cows. During the survey, participating herds had an average bulk milk SCC
of 236,000 cells/ml (range 85,000 to 453,000 cells/ml).
In total, 685 clinical mastitis cases from 534 cows were sampled by the producers. During
the survey, 421, 94, 18, 3 and 2 cows suffered from 1, 2, 3, 4 and 5 cases, respectively.
Seventy-nine percentage (n = 538) of the samples were culture positive including 10%
contaminated samples (n = 69). Streptococcus uberis was the most frequently isolated
pathogen (n = 121, 18% of all cases), followed by E. coli (n = 98, 14%), S. aureus (n= 55, 8%)
and S. dysgalactiae (n = 51, 7%). Two different pathogens were isolated in 35 clinical mastitis
samples (5%). Non-aureus staphylococci (n = 32, 5%), C. bovis (n = 17, 2%), yeast (n = 14,
2%), Prototheca spp. (n = 12, 2%), other esculine-positive cocci besides S. uberis (n = 9, 1%),
Klebsiella spp. (n = 7, 1%), Trueperella pyogenes (n = 5, 1%), Bacillus spp. (n = 3, <1%),
Pasteurella spp. (n = 3, <1%), Streptococcus canis (n = 3, <1%), Pseudomonas aeruginosa (n =
3, <1%), S. agalactiae (n = 2, <1%), Serratia spp. (n = 2, <1%) and molds (n = 1, <1%) were
isolated in the remaining samples. The overall, S. aureus, S. uberis, S. dysgalactiae, and E. coli
IRCM was calculated at 7.1, 0.5, 1.3, 0.5, and 1.1 quarter cases per 10,000 cow-days at risk,
respectively.
In herds participating in the DHI program, genotyped cows had a geometric mean SCC of
79,000 cells per ml (range: 2,000 – 9,590,000) and an average milk yield of 27.8 kg (range:
1.9 – 63.5) at test-day.

Chapter 5.2 The CXCR1 gene and clinical mastitis
160
3.2 Genotypic data
The sire and dam of 957 (31%) and 1,964 (63%) cows, respectively, were available from
DHI records. Among these cows, 460 different sires and 1,951 different dams were noted,
indicating that they were not closely related to each other.
All cows were genotyped using the multiprobe PCR assay. Frequencies of genotype
c.735CC, c.735CG, and c.735GG were 25%, 48%, and 27% in the total population.
Frequencies of genotype c.980AA, c.980AG, and c.980GG were 5%, 34%, and 61% in the total
population. Frequencies of c.735C>G and c.980A>G were not significantly different in the
subpopulation of cows with DHI records compared to the total population [P( > 0.45) =
0.80 and P( > 0.95) = 0.62, respectively]. Both SNPs were in Hardy-Weinberg equilibrium
(P > 0.05). In total, 7 combined genotypes were detected: c.735CC980AA (5%),
c.735CC980AG (12%), c.735CC980GG (8%), c.735CG980AA (<1%), c.735CG980AG (22%),
c.735CG980GG (26%), and c.735GG980GG (27%). Combined genotypes c.73GG980AA and
c.735GG980AG were not observed in the study population. The SNPs were in strong linkage
disequilibrium (P < 0.05, d = 0.65).
3.3 Associations with udder health and milk yield
Descriptive statistics on pathogen-specific IRCM, LnSCC and milk yield by SNPs c.735C>G
and c.980A>G are given in Table 5.2.2 and 5.2.3, respectively.
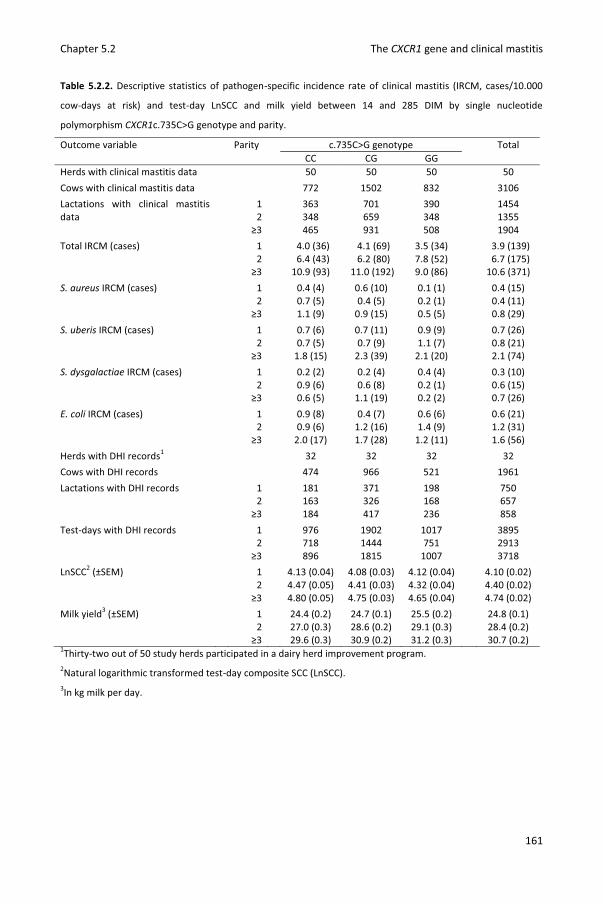
Chapter 5.2 The CXCR1 gene and clinical mastitis
161
Table 5.2.2. Descriptive statistics of pathogen-specific incidence rate of clinical mastitis (IRCM, cases/10.000
cow-days at risk) and test-day LnSCC and milk yield between 14 and 285 DIM by single nucleotide
polymorphism CXCR1c.735C>G genotype and parity.
Outcome variable Parity c.735C>G genotype Total CC CG GG Herds with clinical mastitis data 50 50 50 50 Cows with clinical mastitis data 772 1502 832 3106 Lactations with clinical mastitis data
1 363 701 390 1454 2 348 659 348 1355
≥3 465 931 508 1904 Total IRCM (cases) 1 4.0 (36) 4.1 (69) 3.5 (34) 3.9 (139) 2 6.4 (43) 6.2 (80) 7.8 (52) 6.7 (175) ≥3 10.9 (93) 11.0 (192) 9.0 (86) 10.6 (371) S. aureus IRCM (cases) 1 0.4 (4) 0.6 (10) 0.1 (1) 0.4 (15) 2 0.7 (5) 0.4 (5) 0.2 (1) 0.4 (11) ≥3 1.1 (9) 0.9 (15) 0.5 (5) 0.8 (29) S. uberis IRCM (cases) 1 0.7 (6) 0.7 (11) 0.9 (9) 0.7 (26) 2 0.7 (5) 0.7 (9) 1.1 (7) 0.8 (21) ≥3 1.8 (15) 2.3 (39) 2.1 (20) 2.1 (74) S. dysgalactiae IRCM (cases) 1 0.2 (2) 0.2 (4) 0.4 (4) 0.3 (10) 2 0.9 (6) 0.6 (8) 0.2 (1) 0.6 (15) ≥3 0.6 (5) 1.1 (19) 0.2 (2) 0.7 (26) E. coli IRCM (cases) 1 0.9 (8) 0.4 (7) 0.6 (6) 0.6 (21) 2 0.9 (6) 1.2 (16) 1.4 (9) 1.2 (31) ≥3 2.0 (17) 1.7 (28) 1.2 (11) 1.6 (56) Herds with DHI records1 32 32 32 32 Cows with DHI records 474 966 521 1961 Lactations with DHI records 1 181 371 198 750 2 163 326 168 657 ≥3 184 417 236 858 Test-days with DHI records 1 976 1902 1017 3895 2 718 1444 751 2913 ≥3 896 1815 1007 3718 LnSCC2 (±SEM) 1 4.13 (0.04) 4.08 (0.03) 4.12 (0.04) 4.10 (0.02) 2 4.47 (0.05) 4.41 (0.03) 4.32 (0.04) 4.40 (0.02) ≥3 4.80 (0.05) 4.75 (0.03) 4.65 (0.04) 4.74 (0.02) Milk yield3 (±SEM) 1 24.4 (0.2) 24.7 (0.1) 25.5 (0.2) 24.8 (0.1) 2 27.0 (0.3) 28.6 (0.2) 29.1 (0.3) 28.4 (0.2) ≥3 29.6 (0.3) 30.9 (0.2) 31.2 (0.3) 30.7 (0.2) 1Thirty-two out of 50 study herds participated in a dairy herd improvement program. 2Natural logarithmic transformed test-day composite SCC (LnSCC). 3In kg milk per day.

Chapter 5.2 The CXCR1 gene and clinical mastitis
162
Table 5.2.3. Descriptive statistics of pathogen-specific incidence rate of clinical mastitis (IRCM, cases/10.000
cow-days at risk) and test-day LnSCC and milk yield between 14 and 285 DIM by single nucleotide
polymorphism CXCR1c.980A>G genotype and parity.
Outcome variable Parity c.980A>G genotype Total AA AG GG Herds with clinical mastitis data 41 50 50 50 Cows with clinical mastitis data 149 1053 1904 3106 Lactations with clinical mastitis data
1 62 511 881 1454 2 64 453 838 1355
≥3 103 622 1179 1904 Total IRCM (cases) 1 3.8 (6) 4.3 (54) 3.7 (79) 3.9 (139) 2 5.0 (6) 5.6 (48) 7.3 (121) 6.7 (175) ≥3 7.7 (15) 12.0 (129) 10.0 (227) 10.6 (371) S. aureus IRCM (cases) 1 0.0 (0) 1.0 (12) 0.1 (3) 0.4 (15) 2 0.8 (1) 0.5 (4) 0.4 (6) 0.4 (11) ≥3 0.0 (0) 0.7 (8) 1.0 (21) 0.8 (29) S. uberis IRCM (cases) 1 0.0 (0) 0.6 (8) 0.9 (18) 0.7 (26) 2 0.8 (1) 0.5 (4) 1.0 (16) 0.8 (21) ≥3 2.6 (5) 2.1 (24) 2.1 (45) 2.1 (74) S. dysgalactiae IRCM (cases) 1 0.0 (0) 0.2 (3) 0.3 (7) 0.3 (10) 2 1.7 (2) 0.6 (5) 0.5 (8) 0.6 (15) ≥3 0.0 (0) 0.9 (10) 0.7 (16) 0.7 (26) E. coli IRCM (cases) 1 0.0 (0) 0.6 (8) 0.6 (13) 0.6 (21) 2 0.0 (0) 1.4 (12) 1.2 (19) 1.2 (31) ≥3 2.0 (4) 2.0 (22) 1.4 (30) 1.6 (56) Herds with DHI records1 28 32 32 32 Cows with DHI records 93 691 1177 1961 Lactations with DHI records 1 31 291 428 750 2 35 215 407 657 ≥3 43 294 521 858 Test-days with DHI records 1 163 1506 2226 3895 2 145 941 1827 2913 ≥3 200 1296 2222 3718 LnSCC2 (±SEM) 1 4.14 (0.09) 4.14 (0.03) 4.08 (0.02) 4.10 (0.02) 2 4.21 (0.10) 4.43 (0.04) 4.40 (0.03) 4.40 (0.02) ≥3 4.74 (0.10) 4.80 (0.04) 4.70 (0.03) 4.74 (0.02) Milk yield3 (±SEM) 1 23.8 (0.4) 24.8 (0.2) 25.0 (0.1) 24.8 (0.1) 2 27.7 (0.7) 27.9 (0.3) 28.6 (0.2) 28.4 (0.2) ≥3 30.4 (0.7) 30.6 (0.3) 30.8 (0.2) 30.7 (0.2) 1Thirty-two out of 50 study herds participated in a dairy herd improvement program. 2Natural logarithmic transformed test-day composite SCC (LnSCC). 3In kg milk per day.

Chapter 5.2 The CXCR1 gene and clinical mastitis
163
Incidence Rate of Clinical Mastitis. Season of calving was not associated with (pathogen-
specific) IRCM and therefore removed from all IRCM models. Overall, S. uberis, S.
dysgalactiae and E. coli IRCM were significantly associated with parity (P < 0.05) but not with
SNP c.735C>G, c.980A>G or the interaction between genotype and parity (Table 5.2.4 and
5.2.5). Results were different for S. aureus IRCM. In the model with c.735C>G, the interaction
between c.735C>G and parity was not significant. Parity was significant (P < 0.01) and
genotype tended to be significant (P = 0.08). Cows with genotype c.735GG had on average a
lower S. aureus IRCM compared to cows with genotype c.735CC (RR = 0.35, 95% CI 0.14-
0.90) (Table 5.2.4). In the model with c.980A>G, the interaction between genotype and
parity was significant (P < 0.05). Heifers with genotype c.980GG had on average a lower S.
aureus IRCM compared to heifers with genotype c.980AG (RR = 0.15, 95% CI 0.04-0.56). The
association between c.980A>G and S. aureus IRCM was less pronounced in cows with parity
2 and ≥3 (Table 5.2.5).

Chapter 5.2 The CXCR1 gene and clinical mastitis
164
Table 5.2.4. Multilevel Poisson regression models describing the associations between the pathogen-specific
incidence rate of clinical mastitis (IRCM) and SNP CXCR1c.735C>G.
IRCM Independent variable β2 SE3 RR4 95% CI RR5 P-value7 Overall Intercept -7.87 0.16
c.735C>G1 CC ref. … … … 0.73 CG 0.04 0.11 1.04 0.84-1.28 GG -0.05 0.13 0.95 0.75-1.22 Parity 1 ref. … … … <0.01 2 0.60 0.12 1.83 1.44-2.32 ≥3 1.05 0.11 2.86 2.30-3.56 Interaction NS
Staphylococcus aureus Intercept -9.83 0.37 c.735C>G1 CC ref. … … … 0.08 CG -0.16 0.34 0.85 0.44-1.65 GG -1.05 0.48 0.35 0.14-0.90 Parity 1 ref. … … … <0.05 2 0.01 0.42 1.00 0.44-1.65 ≥3 0.73 0.35 2.08 1.05-4.12 Interaction NS
Streptococcus uberis Intercept -9.55 0.37 c.735C>G1 CC ref. … … … 0.59 CG 0.20 0.23 1.22 0.78-1.89 GG 0.25 0.25 1.28 0.78-2.09 Parity 1 ref. … … … <0.01 2 0.07 0.28 1.07 0.62-1.85 ≥3 1.03 0.22 2.80 1.83-4.29 Interaction NS
Streptococcus dysgalactiae Intercept -10.45 0.42 c.735C>G1 CC ref. … … … 0.13 CG 0.20 0.36 1.22 0.61-2.44 GG -0.68 0.49 0.51 0.19-1.33 Parity 1 ref. … … … <0.05 2 0.69 0.42 1.99 0.87-4.57 ≥3 0.96 0.39 2.62 1.22-5.63 Interaction NS
Escherichia coli Intercept -9.63 0.34 c.735C>G1 CC ref. … … … 0.73 CG -0.20 0.27 0.82 0.48-1.39 . GG -0.20 0.31 0.82 0.45-1.51 Parity 1 ref. … … … 2 0.79 0.31 2.21 1.22-4.02 ≥3 1.03 0.29 2.81 1.60-4.96 Interaction NS
1Genotype at SNP CXCR1c.735C>G. 2Regression coefficient. 3Standard error. 4Rate Ratio. 595% confidence interval of RR. 7Overall P-value of the fixed effect.

Chapter 5.2 The CXCR1 gene and clinical mastitis
165
Table 5.2.5. Multilevel Poisson regression models describing the associations between the pathogen-specific
incidence rate of clinical mastitis (IRCM) and SNP CXCR1c.980A>G.
IRCM Independent variable β2 SE3 RR4 95% CI RR5 P-value7 Overall Intercept -8.17 0.26
c.980A>G8 AA ref. … … … 0.29 AG 0.36 0.23 1.43 0.91-2.27 GG 0.30 0.23 1.35 0.86-2.10 Parity 1 ref. … … … <0.01 2 0.61 0.12 1.83 1.45-2.33 ≥3 1.06 0.11 2.88 2.32-3.58 Interaction NS
Staphylococcus aureus Intercept -9.30 0.34 c.980A>G8 AG ref. … … … <0.01 GG -1.91 0.68 0.15 0.04-0.56 Parity 1 ref. … … … <0.05 2 -0.67 0.61 0.51 0.16-1.69 ≥3 -0.26 0.51 0.77 0.29-2.07 Interaction GG*2 1.63 0.95 5.08 0.79-32.82 <0.05 GG*≥3 2.23 0.82 9.29 1.87-46.26
Streptococcus uberis Intercept -9.44 0.28 c.980A>G8 AG ref. … … … 0.49 GG 0.13 0.19 1.14 0.78-1.66 Parity 1 ref. … … … <0.01 2 0.03 0.28 1.03 0.59-1.79 ≥3 1.01 0.22 2.75 1.78-4.23 Interaction NS
Streptococcus dysgalactiae Intercept -10.39 0.39 c.980A>G8 AG ref. … … … 0.77 GG -0.09 0.32 0.91 0.49-1.71 Parity 1 ref. … … … <0.05 2 0.57 0.44 1.78 0.75-4.19 ≥3 1.00 0.39 2.72 1.26-5.88 Interaction NS
Escherichia coli Intercept -9.56 0.32 c.980A>G8 AG ref. … … … 0.24 GG -0.27 0.23 0.76 0.48-1.21 Parity 1 ref. … … … <0.01 2 0.79 0.30 2.20 1.21-3.99 ≥3 0.97 0.29 2.64 1.49-4.65 Interaction NS
1Genotype at SNP CXCR1c.735C>G. 2Regression coefficient. 3Standard error. 4Rate Ratio. 595% confidence interval of RR. 7Overall P-value of the fixed effect. 8Genotype at SNP CXCR1c.980A>G. Cows with genotype c.980AA were excluded from the analysis to allow for
convergence.

Chapter 5.2 The CXCR1 gene and clinical mastitis
166
Somatic Cell Count. Season of calving, season of test-day, DIM² and the interaction
between genotype and parity were not associated with LnSCC and therefore removed from
both LnSCC models. Test-day LnSCC was significantly associated with parity (P < 0.01) and
DIM (P < 0.01) but not with genotype (Table 5.2.6).
Milk Yield. The interaction between genotype and parity was not associated with milk
yield and therefore removed from both milk yield models. Test-day milk yield was
significantly associated with parity, season of calving, season of test-day, DIM and DIM² (all P
< 0.01). Milk yield was associated with SNP c.735C>G (P < 0.05) but not with SNP c.980A>G
(P = 0.13). Cows with genotype c.735CG and genotype c.735GG produced on average 0.30
(SE 0.27) and 0.81 (SE 0.31) more kg milk per day compared to c.735CC cows (Table 5.2.6).
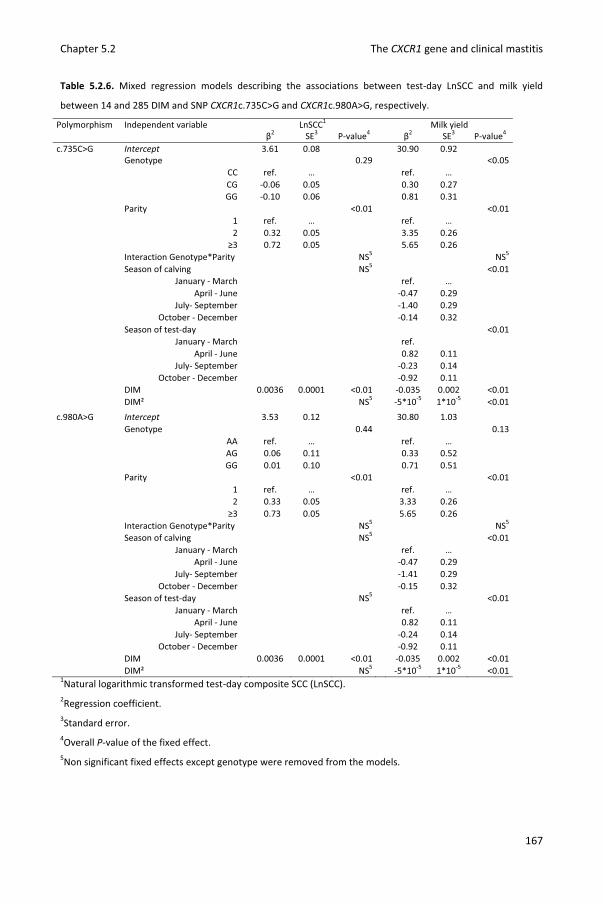
Chapter 5.2 The CXCR1 gene and clinical mastitis
167
Table 5.2.6. Mixed regression models describing the associations between test-day LnSCC and milk yield
between 14 and 285 DIM and SNP CXCR1c.735C>G and CXCR1c.980A>G, respectively.
Polymorphism Independent variable LnSCC1 Milk yield β2 SE3 P-value4 β2 SE3 P-value4 c.735C>G Intercept 3.61 0.08 30.90 0.92 Genotype 0.29 <0.05 CC ref. … ref. … CG -0.06 0.05 0.30 0.27 GG -0.10 0.06 0.81 0.31 Parity <0.01 <0.01 1 ref. … ref. … 2 0.32 0.05 3.35 0.26 ≥3 0.72 0.05 5.65 0.26 Interaction Genotype*Parity NS5 NS5 Season of calving NS5 <0.01 January - March ref. … April - June -0.47 0.29 July- September -1.40 0.29 October - December -0.14 0.32 Season of test-day <0.01 January - March ref. April - June 0.82 0.11 July- September -0.23 0.14 October - December -0.92 0.11 DIM 0.0036 0.0001 <0.01 -0.035 0.002 <0.01 DIM² NS5 -5*10-5 1*10-5 <0.01 c.980A>G Intercept 3.53 0.12 30.80 1.03
Genotype 0.44 0.13 AA ref. … ref. … AG 0.06 0.11 0.33 0.52 GG 0.01 0.10 0.71 0.51
Parity <0.01 <0.01 1 ref. … ref. … 2 0.33 0.05 3.33 0.26
≥3 0.73 0.05 5.65 0.26 Interaction Genotype*Parity NS5 NS5 Season of calving NS5 <0.01 January - March ref. … April - June -0.47 0.29 July- September -1.41 0.29 October - December -0.15 0.32 Season of test-day NS5 <0.01 January - March ref. … April - June 0.82 0.11 July- September -0.24 0.14 October - December -0.92 0.11 DIM 0.0036 0.0001 <0.01 -0.035 0.002 <0.01 DIM² NS5 -5*10-5 1*10-5 <0.01
1Natural logarithmic transformed test-day composite SCC (LnSCC). 2Regression coefficient. 3Standard error. 4Overall P-value of the fixed effect. 5Non significant fixed effects except genotype were removed from the models.

Chapter 5.2 The CXCR1 gene and clinical mastitis
168
4. Discussion
Several research groups studied associations between SNPs in CXCR1 and udder health
(Beecher et al., 2010; Galvao et al., 2011; Goertz et al., 2009; Leyva-Baca et al., 2008b;
Verbeke et al., 2012; Youngerman et al., 2004). However, we were the first to study
pathogen-specific data in a relative large population from multiple commercial dairy herds.
The 50 study herds were randomly selected. No inclusion criteria were applied in our study,
underlining the external validity of our results. A downside of this selection procedure was
the lack of DHI records and pedigree information in a large number of herds. Consequently,
we could not study test-day milk yield and LnSCC of all animals, nor account for additive
polygenic effects in the statistic models.
Different methods were used to genotype animals in previous studies. Sequencing allows
detection of new SNPs (Verbeke et al., 2012; Youngerman et al., 2004) but is relatively
expensive and time consuming. The PCR-RFLP assay used by Beecher et al. (2010), Galvao et
al. (2011), and Goertz et al. (2009) is cheaper and contains only 4 steps (DNA extraction, PCR,
restriction enzyme digestion, and gel electrophoresis). To further reduce the time required
per sample, we designed a fluorescent multiprobe PCR assay allowing us to genotype SNPs
c.735C>G and c.980A>G simultaneously in only 2 steps (DNA extraction and PCR). Using this
assay, we could process the relative large number of samples cost and time effectively.
Although the use of small amplicons (75-150 bp) is recommended (Livak, 1999), we used a
relative large amplicon to be able to genotype the SNPs simultaneously. Purification of DNA
and usage of special qPCR mixes generally result in lower Cq-values and higher RFU (relative
fluorescence units) values (own observations) but was not required in our assay, which
reduced the cost per sample. Comparison with sequence data demonstrated that results
were reliable and repeatable. The assay can be used in further research or in breeding
programs.
When studying associations between gene polymorphisms and mastitis resistance,
different udder health parameters can be measured (Pighetti and Elliott, 2011). By
monitoring clinical signs, clinical mastitis incidence can be estimated directly. In the Nordic
countries Denmark, Finland, Norway and Sweden, disease recording systems provide clinical
mastitis data on the majority of dairy herds (Wolff et al., 2012). In Belgium and other
countries, clinical mastitis data is not available on a larger scale. Yet, conducting this study

Chapter 5.2 The CXCR1 gene and clinical mastitis
169
on the same herds as a study estimating the IRCM in Flanders (Verbeke et al., 2014), enabled
us to analyze associations between SNPs in CXCR1 and IRCM in a relatively large population.
However, we could not confirm the overall higher IRCM in c.735GG cows compared to
c.735CG and CC cows observed by Galvao et al. (2011). Because different mastitis pathogens
elicit different immune responses (Bannerman et al., 2004; Schukken et al., 2011), we also
analyzed associations with pathogen-specific IRCM. This allowed us to detect a lower S.
aureus IRCM in c.735GG compared to c.735CC cows. Additionally, genotype c.980GG was
associated with lower S. aureus IRCM in heifers but not in multiparous cows. The interaction
between SNP c.980A>G and parity could be explained by differences in innate immunity
between heifers and multiparous cows. Phagocytosis and killing of S. aureus by milk
neutrophils was shown to be higher in heifers compared to multiparous cows (Mehrzad et
al., 2009). Although S. aureus IMI elicits little to no IL-8 response (Bannerman, 2009), SNPs in
the gene encoding its receptor were associated with S. aureus IRCM. However, we should
mention that the number of S. aureus cases was limited in our study and that we could not
account for additive polygenic effects in the statistic models. The association does not prove
causality and needs further substantiation, e.g. through conducting an experimental
challenge study in which a number of cows per genotype are infected with S. aureus.
Polymorphisms linked with SNPs c.735C>G and c.980A>G might cause differential immune
responses following S. aureus IMI and explain the differences in S. aureus IRCM, and could
be pictured in such an experiment. Additionally, it could elucidate, why genotype c.980AG
was associated with a lower S. aureus IRCM in this study yet with a higher likelihood of IMI
by major pathogens in a previous study (Verbeke et al., 2012). Hypothetically, heifers with
genotype c.980AG are more likely to develop clinical signs in case of S. aureus IMI without
being more susceptible to the pathogen. Considering S. aureus is worldwide a major cause of
clinical mastitis (Olde Riekerink et al., 2008; Keane et al., 2013; Oliveira et al., 2013)
associated with considerable milk losses (Hertl et al., 2014), more research on this topic is
warranted and justified.
Besides (pathogen-specific) IRCM, we also studied SCC at test-day. Similar to Galvao et al.
(2011), Goertz et al. (2009) and Verbeke et al. (2012), no associations between SCC and SNPs
in CXCR1 were detected. This does not necessarily indicate that the SNPs are not associated
with incidence of subclinical masitis as SCC is merely an indirect test for subclinical mastitis
that does not give information on the pathogens associated with the SCC elevations. Yet,

Chapter 5.2 The CXCR1 gene and clinical mastitis
170
using culture data, c.735C>G was found to be associated with overall incidence of subclinical
masitis (Youngerman et al., 2004) whereas c.980A>G was found to be associated with the
prevalence of subclinical masitis caused by major pathogens in early lactating heifers
(Verbeke et al., 2012). Culture data on a large number of subclinical masitis cases were not
collected in our study but would have allowed to test pathogen-specific associations
between SNPs in CXCR1 and subclinical masitis incidence as well.
Marker assisted selection against mastitis is a noble goal but would receive little
attention if it would decrease the genetic merit for production (Detilleux, 2009). Unfavorable
genetic correlations between clinical mastitis and milk yield traits have been described
(Koivula et al., 2004). Therefore, we also tested associations between SNPs c.735C>G and
c.980A>G and milk yield. Besides a lower S. aureus IRCM, c.735GG had higher milk yield
compared to c.735CC cows contradicting findings by Galvao et al. (2011). Whether the lower
S. aureus IRCM partly explains the higher milk yield is hard to test because the traits were
recorded in somewhat different populations.
As mentioned in the introduction, functional effects of CXCR1 SNP have yet to be
analyzed. As suggested by Pighetti et al. (2012), amino acid change p.His245Glu caused by
c.735C>G could affect G-protein binding and signal transduction whereas amino acid change
p.Lys327Arg caused by c.980A>G could interfere with adaptin-2 binding and receptor
internalization. In silico analysis showed an important role of the N-terminus of CXCR1 in the
first steps of binding IL-8 (Liou et al., 2014). Hence, amino acid changes p.Ile13Tyr and
p.Gly23Glu in this region caused by SNPs c.37A>T, c.38T>A and c.68G>A being in strong
linkage disequilibrium with c.735C>G could alter ligand binding. Latter and other processes
could affect receptor functionality and therefore mastitis resistance/susceptibility.
5. Conclusions
A fluorescent multiprobe PCR assay was designed, validated and used to genotype CXCR1
SNPs c.735C>G and c.980A>G in 3,106 dairy cows. Because of the important function of
CXCR1 in innate immune responses, mutations in the encoding gene might affect mastitis
resistance and milk yield. In this study, SNP c.735C>G tended to associate with S. aureus
IRCM and was associated with milk yield. Cows with genotype c.735GG had a lower S. aureus
IRCM and a higher milk yield compared to cows with genotype c.735CC. Additionally, SNP

Chapter 5.2 The CXCR1 gene and clinical mastitis
171
c.980A>G was associated with S. aureus IRCM in heifers. Genotype c.980GG resulted in
lower S. aureus IRCM compared to genotype c.980AG. The results provide a basis for further
research on the relation between CXCR1 polymorphism and pathogen-specific mastitis
resistance and milk yield.
6. Acknowledgements
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen) and by the Flemish cattle monitoring
project (‘Veepeiler Rund’), headed by the Flemish Animal Health Service
(Dierengezondheidszorg-Vlaanderen). The authors wish to thank the Cattle Breeding
Association (CRV) for access to DHI records and all participating dairy producers.
7. References
Bannerman, D. D., M. J. Paape, J. W. Lee, X. Zhao, J. C. Hope, and P. Rainard. 2004.
Escherichia coli and Staphylococcus aureus elicit differential innate immune responses
following intramammary infection. Clin. Diagn. Lab. Immunol. 11:463-472.
Bannerman, D. D. 2009. Pathogen-dependent induction of cytokines and other soluble
inflammatory mediators during intramammary infection of dairy cows. J. Anim Sci. 87:10-
25.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab. Immunol. 5:82-86.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, H. Wilmink, G. Benedictus, and A.
Brand. 1998. Incidence of clinical mastitis in dairy herds grouped in three categories by
bulk milk somatic cell counts. J. Dairy Sci. 81:411-419.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, G. Benedictus, and A. Brand. 1999.
Management practices associated with the incidence rate of clinical mastitis. J. Dairy Sci.
82:1643-1654.
Beaudeau, F., V. Ducrocq, C. Fourichon, and H. Seegers. 1995. Effect of disease on length of
productive life of French Holstein dairy cows assessed by survival analysis. J. Dairy Sci.
78:103-117.

Chapter 5.2 The CXCR1 gene and clinical mastitis
172
Beecher, C., M. Daly, S. Childs, D. P. Berry, D. A. Magee, T. V. McCarthy, and L. Giblin. 2010.
Polymorphisms in bovine immune genes and their associations with somatic cell count
and milk production in dairy cattle. BMC Genet. 11:99.
Detilleux, J. C. 2009. Genetic factors affecting susceptibility to udder pathogens. Vet.
Microbiol. 134:157-164.
Galvao, K. N., G. M. Pighetti, S. H. Cheong, D. V. Nydam, and R. O. Gilbert. 2011. Association
between interleukin-8 receptor-alpha (CXCR1) polymorphism and disease incidence,
production, reproduction, and survival in Holstein cows. J. Dairy Sci. 94:2083-2091.
Goertz, I., C. Baes, C. Weimann, N. Reinsch, and G. Erhardt. 2009. Association between single
nucleotide polymorphisms in the CXCR1 gene and somatic cell score in Holstein dairy
cattle. J. Dairy Sci. 92:4018-4022.
Hertl, J. A., Y. H. Schukken, F. L. Welcome, L. W. Tauer, and Y. T. Grohn. 2014. Pathogen-
specific effects on milk yield in repeated clinical mastitis episodes in Holstein dairy cows.
J. Dairy Sci. 97:1465-1480.
Hortet, P., F. Beaudeau, H. Seegers, and C. Fourichon. 1999. Reduction in milk yield
associated with somatic cell counts up to 600.000 cells/ml in French Holstein cows
without clinical mastitis. Livest. Prod. Sci. 61:33-42.
Kaplan, N., and B. S. Weir. 1992. Expected behavior of conditional linkage disequilibrium.
Am. J. Hum. Genet. 51: 333–343.
Keane, O. M., K. E. Budd, J. Flynn, and F. Mccoy. 2013. Pathogen profile of clinical mastitis in
Irish milk-recording herds reveals a complex aetiology. Vet. Rec. 173:17.
Kettritz, R., M. L. Gaido, H. Haller, F. C. Luft, C. J. Jennette, and R. J. Falk. 1998. Interleukin-8
delays spontaneous and tumor necrosis factor-alpha-mediated apoptosis of human
neutrophils. Kidney Int. 53:84-91.
Koivula, M., E. Negussie, and E. A. Mantysaari. 2004. Genetic parameters for test-day
somatic cell count at different lactation stages of Finnish dairy cattle. Livest. Prod. Sci.
90:145-157.
Larkin, M. A., G. Blackshields, N. P. Brown, R. Chenna, P. A. McGettigan, H. McWilliam, F.
Valentin, I. M. Wallace, A. Wilm, R. Lopez, J. D. Thompson, Gibson T. J., and D. G. Higgins.
2007. Clustal W and Clustal X version 2.0. Bioinformatics 23:2947-2948.

Chapter 5.2 The CXCR1 gene and clinical mastitis
173
Leyva-Baca, I., G. Pighetti, and N. A. Karrow. 2008a. Genotype-specific IL8RA gene expression
in bovine neutrophils in response to Escherichia coli lipopolysaccharide challenge. Anim.
Gen. 39, 298-300.
Leyva-Baca, I., F. Schenkel, J. Martin, and N. A. Karrow. 2008b. Polymorphisms in the 5 '
upstream region of the CXCR1 chemokine receptor gene, and their association with
somatic cell score in Holstein cattle in Canada. J. Dairy Sci. 91:407-417.
Liou, J. W., F. T. Chang, Y. Chung, W. Y. Chen, W. B. Fischer, and H. J. Hsu. 2014. In Silico
analysis reveals sequential interactions and protein conformational changes during the
binding of chemokine CXCL-8 to its receptor CXCR1. Plos One 9.
Livak, K. J. 1999. Allelic discrimination using fluorogenic probes and the 5 ' nuclease assay.
Genet. Anal. 14:143-149.
Ma, Y., C. Ryan, D. M. Barbano, D. M. Galton, M. A. Rudan, and K. J. Boor. 2000. Effects of
somatic cell count on quality and shelf-life of pasteurized fluid milk. J. Dairy Sci. 83:264-
274.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2009. Phagocytic and bactericidal activity of
blood and milk-resident neutrophils against Staphylococcus aureus in primiparous and
multiparous cows during early lactation. Vet. Microbiol. 134:106-112.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
National Mastitis Council. 1999. Laboratory Handbook on Bovine Mastitis. Revised Edition.
National Mastitis Council. Inc., Madison, WI.
Ogorevc, J., T. Kunej, A. Razpet, and P. Dovc. 2009. Database of cattle candidate genes and
genetic markers for milk production and mastitis. Anim. Genet. 40:832-851.
Oliveira, L., C. Hulland, and P. L. Ruegg. 2013. Characterization of clinical mastitis occurring in
cows on 50 large dairy herds in Wisconsin. J. Dairy Sci. 96:7538-7549.
Olde Riekerink, R. G. M., H. W. Barkema, D. F. Kelton, and D. T. Scholl. 2008. Incidence rate
of clinical mastitis on Canadian dairy farms. J. Dairy Sci. 91:1366-1377.
Pighetti, G. M., and A. A. Elliott. 2011. Gene polymorphisms: the keys for marker assisted
selection and unraveling core regulatory pathways for mastitis resistance. J. Mammary
Gland Biol. Neoplasia. 16:421-432.

Chapter 5.2 The CXCR1 gene and clinical mastitis
174
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Santos, M. V., Y. Ma, and D. M. Barbano. 2003. Effect of somatic cell count on proteolysis
and lipolysis in pasteurized fluid milk during shelf-life storage. J. Dairy Sci. 86:2491-2503.
Schukken, Y. H., D. J. Wilson, F. Welcome, L. Garrison-Tikofsky, and R. N. Gonzalez. 2003.
Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 34:579-596.
Schukken, Y. H., J. Gunther, J. Fitzpatrick, M. C. Fontaine, L. Goetze, O. Holst, J. Leigh, W.
Petzl, H. J. Schuberth, A. Sipka, D. G. E. Smith, R. Quesnell, J. Watts, R. Yancey, H. Zerbe,
A. Gurjar, R. N. Zadoks, and H. M. Seyfert. 2011. Host-response patterns of
intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 144:270-289.
Seegers, H., C. Fourichon, and F. Beaudeau. 2003. Production effects related to mastitis and
mastitis economics in dairy cattle herds. Vet. Res 34:475-491.
Smit, A. F. A., R. Hubley, and P. Green. 2010. RepeatMasker Open-3.0. Accessed January 18,
2013. http://www.repeatmasker.org.
Sodeland, M., M. P. Kent, H. G. Olsen, M. A. Opsal, M. Svendsen, E. Sehested, B. J. Hayes, and
S. Lien. 2011. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20
in Norwegian Red cattle. Anim. Genet. 42:457-465.
Van Poucke, M., J. Vandesompele, M. Mattheeuws, A. Van Zeveren, and L. J. Peelman. 2005.
A dual fluorescent multiprobe assay for prion protein genotyping in sheep. BMC Infect.
Dis. 5:13.
Verbeke, J., S. Piepers, L. J. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-
group specific association between CXCR1 polymorphisms and subclinical mastitis in
dairy heifers. J. Dairy Res. 79:341-351.
Verbeke, J., S. Piepers, K. Supré, and S. De Vliegher. 2014. Pathogen-specific incidence rate of
clinical mastitis in Flemish dairy herds, severity, and association with herd hygiene. J.
Dairy Sci. 97:6926-6934.
Windig, J. J., W. Ouweltjes, J. Ten Napel, G. de Jong, R. F. Veerkamp, and Y. de Haas. 2010.
Combining somatic cell count traits for optimal selection against mastitis. J. Dairy Sci.
93:1690-1701.
Wolff, C., M. Espetvedt, A. K. Lind, S. Rintakoski, A. Egenvall, A. Lindberg, and U. Emanuelson.
2012. Completeness of the disease recording systems for dairy cows in Denmark,

Chapter 5.2 The CXCR1 gene and clinical mastitis
175
Finland, Norway and Sweden with special reference to clinical mastitis. BMC Vet. Res.
8:131.
Youngerman, S. M., A. M. Saxton, S. P. Oliver, and G. M. Pighetti. 2004. Association of CXCR2
polymorphisms with subclinical and clinical mastitis in dairy cattle. J. Dairy Sci. 87:2442-
2448.
Zuker, M. 2003. Mfold web server for nucleic acid folding and hybridization prediction.
Nucleic Acids Res. 31:3406-3415.

176

177
Chapter 6
Differential expression of CXCR1 and commonly used
reference genes in bovine milk somatic cells following
experimental intramammary infection
J. Verbeke1, M. Van Poucke2, L. Peelman2, and S. De Vliegher1 1Department of Reproduction, Obstetrics, and Herd Health, Faculty of Veterinary Medicine,
Ghent University, Belgium 2Department of Nutrition, Genetics, and Ethology, Faculty of Veterinary Medicine, Ghent
University, Belgium
BMC Genetics, submitted.

Chapter 6 Gene expression in milk cells
178

Chapter 6 Gene expression in milk cells
179
Abstract
Chemokine (C-X-C motif) receptor 1 (CXCR1 or IL-8RA) plays an important role in the
bovine mammary gland immunity. Previous research indicated polymorphism c.980A>G in
the CXCR1 gene to influence milk neutrophils and mastitis resistance. In the present study,
four c.980AG heifers and four c.980GG heifers were experimentally infected with
Staphylococcus chromogenes. RNA was isolated from milk somatic cells one hour before and
12 hours after the experimental intramammary infection. Expression of CXCR1 and eight
candidate reference genes (ACTB, B2M, H2A, HPRT1, RPS15A, SDHA, UBC and YWHAZ) was
measured by reverse transcription quantitative real-time PCR (RT-qPCR). Differences in
relative CXCR1 expression between c.980AG heifers and c.980GG heifers were studied and
the effect of the experimental intramammary infection on relative expression of CXCR1 and
the candidate reference genes was analyzed. Relative expression of CXCR1 was not
associated with polymorphism c.980A>G but was significantly upregulated following the
experimental intramammary infection. Additionally, differential expression was detected for
B2M, H2A, HPRT1, SDHA and YWHAZ. This study reinforces the importance of CXCR1 in
mammary gland immunity and demonstrates the potential effect of experimental
intramammary infection on expression of candidate reference genes in milk somatic cells.
Keywords
CXCR1, RT-qPCR, milk somatic cell, experimental intramammary infection

Chapter 6 Gene expression in milk cells
180
1. Introduction
After invading the bovine mammary gland, pathogens can cause an intramammary
infection (IMI) followed by an inflammatory response called mastitis. Neutrophils migrating
from blood to milk play an important role in the mammary gland immunity (Paape et al.,
2002). Binding of cytokine interleukin-8 on chemokine (C-X-C motif) receptor 1 (CXCR1)
causes chemotaxis and enhances viability of bovine neutrophils (Barber and Yang, 1998;
Rambeaud et al., 2006). Despite its important function, many single nucleotide
polymorphisms (SNP) were detected in the CXCR1 gene (Pighetti et al., 2012; Verbeke et al.,
2012). Recently, we reported a higher milk neutrophil viability and lower likelihood of IMI by
major mastitis pathogens (e.g. Staphylococcus aureus and Streptococcus uberis) in heifers
with genotype CXCR1c.980AG compared to heifers with genotype CXCR1c.980AG (Verbeke
et al., 2012; Verbeke et al., 2014). Polymorphism c.980A>G causes the amino acid change
p.Lys327Arg in the C-terminal region of the receptor potentially influencing interleukin 8
signal transduction. However, phenotypical differences could also be explained by linkage
with SNPs in regulatory regions and an association between SNP c.980A>G and CXCR1 gene
expression. To test this hypothesis, we isolated RNA from milk somatic cells isolated before
and after an experimental infection with Staphylococcus chromogenes. Next, differences in
CXCR1 expression between c.980AG and c.980GG heifers were studied using reverse
transcription quantitative real-time PCR (RT-qPCR). Additionally, the influence of the
experimental intramammary infection on expression of CXCR1 and commonly used
reference genes was analyzed.
2. Materials and methods
2.1 Test animals
This experiment has been approved by the ethical committee of the Faculty of Veterinary
Medicine, Ghent University (EC2012/73). A blood sample was taken from all Holstein heifers
(n = 20) of the commercial dairy herd of Ghent University (Biocenter Agri-Vet, Melle,
Belgium). The whole coding region of CXCR1 was genotyped by direct sequencing as
described by Verbeke et al. (Verbeke et al., 2012). Four heifers with genotype c.980AG and 4
heifers with genotype c.980GG were selected. Selected heifers were not siblings, had no
history of clinical mastitis or other diseases, and were between 75 and 280 days in milk at

Chapter 6 Gene expression in milk cells
181
the time of the experiment. Duplicate milk samples were taken four days and one hour
before the experiment. Bacteriological culture was performed according to National Mastitis
Council (NMC) guidelines (National Mastitis Council, 1999). All quarters of all heifers were
culture-negative at that time.
2.2 Experimental infection
Each heifer was inoculated briefly after the morning milking (8 a.m.) with two different
strains of S. chromogenes, a strain of Staphylococcus fleurettii, and sterile phosphate
buffered saline (PBS) in a split-udder design (one strain or PBS per individual quarter) to
study differences in pathogenicity and immune response between coagulase-negative
staphylococci. One S. chromogenes (S. chromogenes IM) strain originated from a chronically
infected quarter (Piessens et al., 2011) whereas the other (S. chromogenes TA) originated
from the teat apex and was found to inhibit the growth of major pathogens in vitro (De
Vliegher et al., 2004). The S. fleurettii strain originated from sawdust (Piessens et al., 2011).
For each strain, 1*106 CFU in 5 mL PBS was inoculated using a sterile catheter (Vygon,
Ecouen, France). The bacterial count was determined by incubating a tenfold serial dilution
of a representative frozen aliquot 18 h before inoculation. Five mL of sterile PBS was
inoculated in the fourth quarter. For this research, additional milk samples of 600 mL were
taken 1 h before and 12 h after inoculation from quarters (to be) inoculated with PBS or S.
chromogenes IM (Figure 6.1). Cows were milked after sampling. Quarter SCC was measured
in duplicate using a DeLaval cell counter (DCC, DeLaval International AB, Tumba, Sweden).

Chapter 6 Gene expression in milk cells
182
Figure 6.1. Samples. Four CXCR1c.980AG heifers and 4four CXCR1c.980AG heifers were inoculated with PBS, a
Staphylococcus chromogenes strain isolated from a chronic intramammary infection (S. chromogenes IM), a
Staphylococcus chromogenes strain from a teat apex (S. chromogenes TA) and a Staphylococcus fleurettii strain
in a split-udder design to study differences between coagulase-negative staphylococci. For this research,
somatic cells were isolated from milk samples taken 1 h before and 12 h after inoculation from quarters (to be)
inoculated with PBS or S. chromogenes IM.
2.3 Milk somatic cell isolation
The milk was divided equally between three 400-mL centrifuge bottles, diluted 50%
(vol/vol) with cold PBS, and centrifuged at 1500 × g for 15 min at 4°C in a fixed angle rotor.
The supernatant was discarded. The three milk somatic cell pellets were resuspended in a
total of 40 mL PBS without touching the fat layer at the side of the bottles, divided between
two 50-mL Falcon tubes and washed three times with 10 mL cold PBS (centrifugation at 200
x g for 10 min at 4 °C). The final milk somatic cell pellets were suspended in 1 mL of RPMI
1640 (Gibco Brl., Scotland, UK) supplemented with 1% bovine serum albumin (Merck KGaA,
Darmstadt, Germany). Twenty μL of the suspension was diluted with 380 μl healthy milk
(SCC < 50 cells/mL) and measured with a DeLaval cell counter to estimate the cell
PBS
S. chromogenesIM
S. chromogenesTA
S. fleurettii
PBS
S. chromogenesIM
S. chromogenesTA
S. fleurettii
Intramammaryinoculation
12 h after inoculation1 h before inoculation
4 CXCR1c.980AG heifers
4 CXCR1c.980GG heifers

Chapter 6 Gene expression in milk cells
183
concentration using following formula; SCCsample (in cells/μl) = .
Approximately 5*106 cells were pipetted in a 2 mL test tube, pelleted by centrifugation at
16,100 x g for 1 min at 4 °C and resuspended in 1 mL TRI Reagent Solution (Ambion, Austin,
TX). If less cells were available, all of the cell suspension was used. Samples were frozen and
stored at -20 °C for 8-10 months.
2.4 RNA extraction and cDNA synthesis
RNA was isolated following the manufacturer’s instructions of TRI Reagent Solution
(Ambion, Austin, TX). Genomic DNA was removed by adding 4 μL RQ1 DNase (0.5 U/μL,
Promega, Leiden, Netherlands) and 2.7 μL RQ1 DNase 10X Reaction Buffer (Promega)
followed by incubation for 30 min at 37 °C. The reaction was terminated by adding 3 μL RQ1
DNase Stop Solution (Promega) followed by incubation for 10 min at 65 °C. The RNA was
purified by spin-column centrifugation (Amicon Ultra-0.5 centrifugal filter device, Merck
Millipore, Billerica, MA) to approximately 15 μL. Its concentration and purity was estimated
using a ND-1000 spectrophotometer (NanoDrop, Wilmington, NC).
DNA contamination was assessed by performing a minus RT control. A PCR mix, in a total
volume of 10 μL, containing 0.4 μL sample (double the equivalent of RNA input in the qPCR
reaction), 1 μL 10×FastStart Taq DNA Polymerase Buffer (Roche Applied Science), 0.2 μl
dNTP Mix (10 mM each; BIOLINE, London, UK), 0.5 μL forward primer [YWHAZ+1, 5 μM, 5’-
GCATCCCACAGACTATTTCC-3', IDT (Integrated DNA Technologies), Leuven, Belgium], 0.5 μL
reverse primer (YWHAZ-1, 5 μM, 5’-GCAAAGACAATGACAGACCA-3', IDT) and 0.1 μL Taq DNA
Polymerase (5 U/μl, Roche Applied Science) was made. The PCR program consisted of an
initiation step of 4 min at 95 °C followed by 40 amplification cycles (denaturation for 10 s at
95 °C, annealing for 10 s at 58 °C and extension for 20 s at 72 °C) and a final 2-min elongation
step at 72 °C. DNA amplification (120 bp) was examined by electrophoresis on ethidium
bromide-stained agarose (2%) gel (150 V, 20 min).
Improm-II reverse transcriptase (Promega) was used to convert DNA-free RNA into cDNA.
First, 10 μL sample containing a maximum of 1 μg RNA was mixed with 0.8 μL random
hexamer primers (10 μM, IDT) and 0.8 μL oligo(dT)15 primer (10 μM, IDT). If less than 1 μg
RNA was extracted, all available RNA was used. The primer/template mix was thermally
denatured by 5 min incubation at 70 °C followed by 5 min incubation on ice. Secondly, 4 μL

Chapter 6 Gene expression in milk cells
184
Improm-II 5x reaction buffer, 2.4 μL MgCl2 (25 mM), 1 μL dNTP Mix (10 mM each; BIOLINE)
and 1 μL Improm-II reverse transcriptase (20 U/μL) was added. The final volume of 20 μL was
incubated 5 min at 25 °C (primer annealing), 60 min at 42 °C (first-strand synthesis reaction),
and 15 min at 70 °C (inactivation of reverse transcriptase). Samples were 5 times diluted and
stored at -20 °C.
2.5 PCR assay to assess cDNA integrity
Complementary DNA integrity was assessed using two 4-primer PCR assays multiplying
fragments of approximately 100, 500 and 900 bps of YWHAZ and CXCR1, respectively. For
both assays, a forward and 3 reverse PCR primers were designed using Primer3Plus
(Untergasser et al., 2007) and synthesized by IDT. Sequences are shown in Table 6.1. Regions
forming potential secondary structures were identified with Mfold (Zuker, 2003) and
avoided. Specificity of binding of the primers was analyzed using NCBI BLAST (Altschul et al.,
1990). For both assays, a PCR mix, in a total volume of 10 μL, containing 2 μL 5x diluted
cDNA, 1.0 μL 10×FastStart Taq DNA Polymerase Buffer (Roche Applied Science), 0.3 μL dNTP
Mix (10 mM each; BIOLINE), 1 μL forward primer (5 μM), 0.3 μL reverse primer 1 (5 μM), 0.3
μL reverse primer 2 (5 μM), 0.6 μL reverse primer 3 (5 μM) and 0.1 μL Taq DNA Polymerase
(5 U/μl, Roche Applied Science) was made. The PCR program consisted of an initiation step
of 4 min at 95°C followed by 40 amplification cycles (denaturation for 45 s at 95 °C,
annealing for 45 s at the optimal annealing temperature and extension for 1 min 30 s at 72
°C) and a final 4-min elongation step at 72 °C. The optimal annealing temperature for the
YWHAZ and CXCR1 assay (60 °C and 65 °C, respectively) were determined experimentally.
DNA amplification was examined by electrophoresis on ethidium bromide-stained agarose
(0.8%) gel (150 V, 25 min). The cDNA integrity was considered excellent, sufficient or
insufficient if, respectively, 3, 2 or 1 bands were visible in both assays (Figure 6.2).
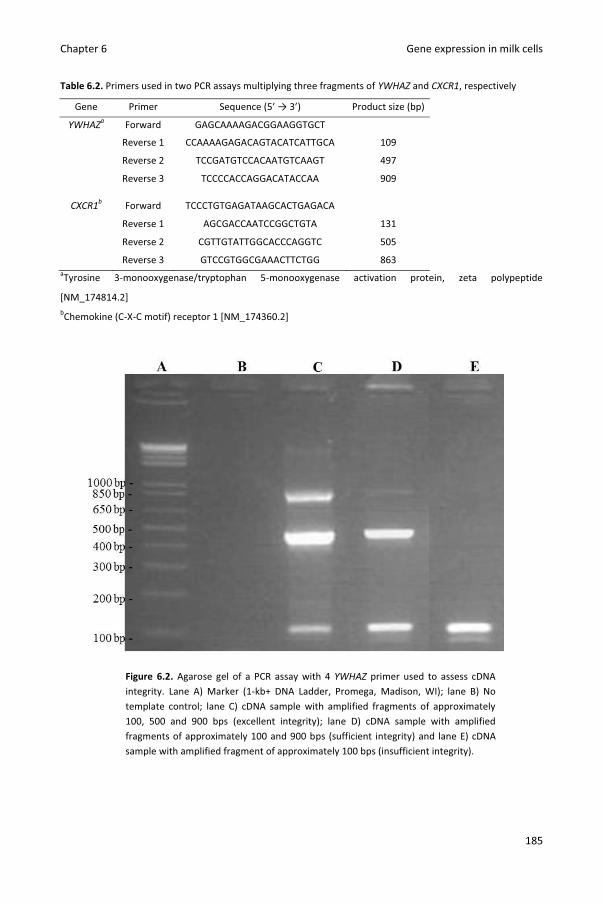
Chapter 6 Gene expression in milk cells
185
Table 6.2. Primers used in two PCR assays multiplying three fragments of YWHAZ and CXCR1, respectively
Gene Primer Sequence (5’ → 3’) Product size (bp)
YWHAZa Forward GAGCAAAAGACGGAAGGTGCT
Reverse 1 CCAAAAGAGACAGTACATCATTGCA 109
Reverse 2 TCCGATGTCCACAATGTCAAGT 497
Reverse 3 TCCCCACCAGGACATACCAA 909
CXCR1b Forward TCCCTGTGAGATAAGCACTGAGACA
Reverse 1 AGCGACCAATCCGGCTGTA 131
Reverse 2 CGTTGTATTGGCACCCAGGTC 505
Reverse 3 GTCCGTGGCGAAACTTCTGG 863 aTyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide
[NM_174814.2] bChemokine (C-X-C motif) receptor 1 [NM_174360.2]
Figure 6.2. Agarose gel of a PCR assay with 4 YWHAZ primer used to assess cDNA integrity. Lane A) Marker (1-kb+ DNA Ladder, Promega, Madison, WI); lane B) No template control; lane C) cDNA sample with amplified fragments of approximately 100, 500 and 900 bps (excellent integrity); lane D) cDNA sample with amplified fragments of approximately 100 and 900 bps (sufficient integrity) and lane E) cDNA sample with amplified fragment of approximately 100 bps (insufficient integrity).

Chapter 6 Gene expression in milk cells
186
2.6 RT-qPCR
Ten candidate reference genes were selected based upon previous research (Goossens et
al., 2005; Stevens et al., 2011; Varshney et al., 2012): ACTB, B2M, H2A, HPRT1, PPP1R11,
RPS15A, SDHA, TBP, UBC, and YWHAZ. Primers were ordered from IDT. Gene, primer and
amplicon information is listed in Table 6.2. A PCR mix of 10 μL containing 5 μL 2x SYBR Green
I Master Mix (Roche Diagnostics, Basel, Switzerland), 1 μL forward primer (5 μM), 1 μL
reverse primer (5 μM) and 2 μL cDNA sample was made. The PCR program consisted of an
initiation step of 3 min at 95°C, followed by 40 amplification cycles (denaturation for 30 s at
95 °C, annealing-elongation for 40 s at the optimal annealing temperature and detection of
fluorescent signals generated by SYBR Green I binding to dsDNA). Samples were heated from
75 °C to 95 °C in 0.5 °C increments per 5 s while continuously measuring fluorescence. The
generated melt curve was used to confirm a single gene-specific peak and to detect
primer/dimer formation. Optimal annealing temperatures were determined experimentally
by gradient qPCR on a 4-fold serial dilution until 1/1024 of pooled cDNA of all samples. All
reactions were performed in duplicate. In each run, the serial dilution and a no template
control were included to analyze calibration curves, PCR efficiency (E) and squared
correlation coefficient (r²) and check for PCR contamination. All qPCRs were performed in
PCR strips (Bio-Rad, Hercules, CA) using a CFX96 Touch™ Real-Time PCR Detection System
(Bio-Rad). Quantification cycles (Cqs) were analyzed with CFX Manager™ Software v3.1 (Bio-
Rad). The raw Cq values were converted to quantiles (Q) using following formula; Q = (1 +
E)(CqS – CqL) with E = PCR efficiency, CqS = Cq value of the sample and CqL = lowest Cq value of
all samples.

Chapter 6 Gene expression in milk cells
187
Table 6.2. Gene, primer and amplicon information
Gene full name NCBI Gene ID Genbank Accession
number
Sequence (5’ → 3’) Genomic location
Product size (bp) Ta (°C)
ACTB 280979 NM_173979.3 F1 CCTCACGGAACGTGGTTACA CDS2 87 58 Actin, beta R1 TCCTTGATGTCACGCACAATTT CDS B2M 280729 NM_173893.3 F AGACACCCACCAGAAGATGG CDS 206 59 Beta-2-microglobulin R CGGCAGCTGTACTGATCCTT CDS CXCR1 281863 NM_174360.2 F TCCCTGTGAGATAAGCACTGAGACAC CDS 118 64 Chemokine (C-X-C motif) receptor 1 R GCTGTATAAGATGACCAGCATCACCA CDS H2A 506900 NM_001205596.1 F GTCGTGGCAAGCAAGGAG CDS 182 60 Histone 2 alpha R GATCTCGGCCGTTAGGTACTC CDS HPRT1 281229 NM_001034035.2 F TGCTGAGGATTTGGAGAAGG CDS 154 58 Hypoxanthine phosphoribosyl-transferase I R CAACAGGTCGGCAAAGAACT CDS PPP1R114 504846 NM_001100295.1 F ACCATCAAACTTCGGAAACG CDS 166 57 Protein phosphatase 1, regulatory (inhibitor) subunit 11 R CCTCCTCTTCCTCGTCATCA CDS RPS15A 337888 NM_001037443.2 F AATGTCCTGGCTGATGCTCT CDS 218 59 Ribosomal protein S15a R GGGCTGATCACTCCACACTT CDS SDHA 281480 NM_174178.2 F GCAGAACCTGATGCTTTGTG CDS 185 60 Succinate dehydrogenase flavoprotein subunit A R CGTAGGAGAGCGTGTGCTT CDS TBP5 516578 NM_001075742.1 F AATGGCTGCTGTGTTCTCCT 3’UTR3 214 60 TATA box binding protein R TGACGCTCTGGTGTTCTCTGT 3’UTR UBC 444874 NM_001206307.1 F AGTTCAGTCTTCGTTCTTCTGTG 5’UTR4 88 58 Ubiquitin C R GGTTTTACCAGTGAGGGTCTT CDS YWHAZ 287022 NM_174814.2 F GCATCCCACAGACTATTTCC CDS 120 60 Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, zeta polypeptide
R GCAAAGACAATGACAGACCA CDS 1Forward and reverse primer. 2Coding sequence. 33’ untranslated region. 45’ untranslated region.
2.7 Analysis of gene expression stability
Stability of the different candidate reference genes were analyzed using Normfinder
(version 0.953; Andersen et al., 2004) Excel Add-In. Samples were grouped as (1) all samples
1 h before inoculation, (2) samples from quarters inoculated with PBS 12 h after inoculation,
and (3) samples from quarters inoculated with S. chromogenes IM 12 h after inoculation.
Normfinder estimates an expression stability measure (ρ) per candidate reference gene
based on the overall variation of the expression and the variation of the expression between
the subgroups (Andersen et al., 2004).
2.7 Data analysis
Differences in gene expression between CXCR1 genotype and sample subgroups were
further studied using SAS 9.4 (SAS Institute Inc., NC, USA). First, expression of B2M, CXCR1,
H2A, HPRT1, SDHA and YWHAZ were normalized by the geometric mean of the three most

Chapter 6 Gene expression in milk cells
188
stable genes (ACTB, RPS15A and UBC; see further). Expression of ACTB, RPS15A and UBC
were normalized to the geometric mean of the other two most stable genes. Data were log
transformed to obtain a normalized distribution. Secondly, a linear mixed regression model
was fit with relative expression as outcome variable and heifer and quarter as random effect
to correct for clustering of quarters within cows and two observations per quarter,
respectively (PROC MIXED, SAS 9.4). Sample subgroup (1, 2 and 3) was added as fixed effect.
In the model for relative CXCR1 expression, genotype (c.980AG and c.980GG) and the
interaction between sample subgroup and genotype were also tested. Statistical significance
was assessed at P ≤ 0.05.
3. Results
3.1 Experimental infection
All milk samples taken 24, 12 and 1 h before experimental inoculation were culture
negative. Quarters (n = 16) had a geometric mean SCC of 48,000 cells/ml [Interquartile range
(IQR) 26,000 – 84,000] 1 h before inoculation. Neighboring quarters remained culture
negative after inoculation. Quarters inoculated with the S. chromogenes strain originating
from a chronic infection developed subclinical mastitis as no local or systemic signs were
observed and the challenge isolate was recovered from all quarters 6 h after inoculation.
Twelve hours after inoculation, neighboring and infected quarters had a geometric mean
SCC of 77,000 cells/ml (IQR 44,000 to 126,000) and 1,692,000 cells/ml (IQR 861,000 to
3,371,000), respectively.
3.2 Quality control of nucleic acids
Total amount of isolated RNA ranged from 27.6 ng to 36.5 μg per sample. An average
A260/A280 ratio of 2.04 was measured (range: 1.81-2.23). The DNase treatment was repeated
for one sample because of a positive minus RT test. The cDNA integrity was assessed using
two four-primer PCR assays amplifying three different YWHAZ fragments and three different
CXCR1 fragments. In two samples, gel electrophoresis of both assays showed no or only one
band indicating cDNA integrity to be too low to amplify the medium and large fragments.
Latter samples were not further analyzed. The cDNA integrity of the remaining samples was

Chapter 6 Gene expression in milk cells
189
considered sufficient (2 bands, n = 8) or excellent (3 bands, n = 22). Results of the YWHAZ
and CXCR1 assay were concordant.
3.3 RT-qPCR and gene expression stability
The gradient qPCR of PPP1R11 and TBP on the 1/1024 dilution of the pooled cDNA
showed weak fluorescent signals and melt peaks indicating low expression of these genes in
the samples. They were not further tested as reference genes. The calibration curves of the
remaining candidate reference genes and CXCR1 demonstrated PCR efficiencies close to
100% and correlation coefficients close to 1 indicating good assay performance. The SD of
the Cq values of replicate samples were limited demonstrating good repeatability (Table
6.3). Normfinder identified UBC, RPS15A and ACTB as the most stable genes based on their
low inter- and intragroup variation in expression (Figure 6.3).
Table 6.3. qPCR information
Gene Median
Cq SD Cq1 Calibration curve2
slope y intercept E (%) r2 ACTB 22.5 0.125 -3.541 18.607 91.6 0.997
B2M 17.7 0.137 -3.4261 13.756 95.8 0.998
CXCR1 24.3 0.077 -3.195 17.240 105.6 0.999
H2A 23.1 0.186 -3.370 19.463 98.0 0.996
HPRT1 25.4 0.140 -3.463 20.475 94.4 0.997
RPS15A 19.3 0.151 -3.433 15.213 95.6 0.999
SDHA 23.4 0.148 -3.534 19.385 91.8 0.999
UBC 22.5 0.122 -3.248 18.150 103.2 0.998
YWHAZ 22.8 0.097 -3.368 19.139 98.1 0.999
1Average standard deviation of the Cq values of replicate samples. 2Slope, y intercept, PCR efficiency and correlation coefficient estimated by qPCR on a 4-fold serial dilution of
pooled cDNA of all samples.

Chapter 6 Gene expression in milk cells
190
Figure 6.3. Gene expression stability of candidate reference genes in milk somatic cells analysed using
Normfinder software. Intergroup variation (+ intragroup variation) of expression of 8 candidate reference
genes in milk somatic cells isolated from quarters inoculated with PBS (n = 8) or Staphylococcus chromogenes
(n = 8) of 8 dairy heifers. Candidate reference genes are ranked on gene expression stability (ρ) calculated using
Normfinder (Andersen et al., 2004) with most stable genes on the right side (smallest inter- and intragroup
variation).
3.4 Effect of genotype and experimental infection on gene expression
Genotype and the interaction between genotype and sample subgroup were non-
significant (P = 0.55 and 0.26, respectively) and removed from the model for relative
expression of CXCR1. Relative expression of CXCR1 was significantly higher in milk somatic
cells from the infected and neighboring quarters 12 h after inoculation compared to milk
somatic cells isolated 1 h before inoculation (both P < 0.01). Additionally, relative expression
of B2M, H2A, HPRT1, SDHA and YWHAZ differed significantly between milk somatic cells
from the infected quarters 12 h after inoculation and milk somatic cells isolated 1 h before
inoculation (P < 0.05). Furthermore, relative expression of B2M and YWHAZ differed
significantly between milk somatic cells from the neighboring quarters 12 h after inoculation
and milk somatic cells isolated 1 h before inoculation. Relative expression of the three most
stable genes (ACTB, RPS15A and UBC) did not differ significantly between the subgroups
(Table 6.4).
ρ=0.51 ρ=0.44
ρ=0.42 ρ=0.40 ρ=0.36 ρ=0.29 ρ=0.28
ρ=0.17
-0.5
-0.3
-0.1
0.1
0.3
0.5
0.7
0.9
Inte
rgro
up v
aria
tion
All quarters 1h before inoculation
Neighboring quarters 12h after inoculation
Infected quarters 12h after inoculation

Chapter 6 Gene expression in milk cells
191
Table 6.4 Linear mixed models describing the difference in gene expression in milk somatic cells before and
after experimental intramammary infection
Gene All quarters Neighboring quarters Infected quarters 1 h before inoculation1 12 h after inoculation 12 h after inoculation (n = 152) (n = 8) (n = 7 2)
β03 SE4 β3 SE4 P- β3 SE4 P-value
ACTB 0.27 0.07 -0.06 0.08 0.46 0.10 0.0 0.27 B2M 0.04 0.06 0.15 0.07 <0.05 0.41 0.0 <0.01
CXCR1 -1.30 0.15 0.94 0.26 <0.01 1.71 0.2 <0.01 H2A -0.04 0.06 0.17 0.07 <0.05 -0.21 0.0 <0.01
HPRT1 -0.08 0.05 -0.03 0.07 0.66 -0.31 0.0 <0.01 RPS15A -0.10 0.10 -0.07 0.12 0.59 -0.06 0.1 0.62 SDHA -0.13 0.05 0.04 0.08 0.67 -0.22 0.0 <0.05 UBC -0.17 0.05 0.13 0.08 0.10 -0.03 0.0 0.67
YWHAZ -0.13 0.08 0.05 0.09 0.61 0.28 0.0 <0.01 1Two quarters of 8 dairy heifers were inoculated with PBS or Staphylococcus chromogenes. 2Two samples showed insufficient cDNA integrity and were therefore removed from the dataset. 3Regression coefficient. 4Standard error.
Heifer and quarter were added as random effect to correct for clustering of quarters within cows and two
observations per quarter, respectively.
4. Discussion
Gene expression analysis in experimentally infected and healthy quarters allows for
identification of differentially expressed genes and pathways. Quantitative real-time PCR in
milk somatic cells isolated prior to inoculation and at different stages of experimental
infection enables a detailed follow-up of the host response. In this study, we analyzed
associations between SNP CXCR1c.980A>G and CXCR1 expression in milk somatic cells and
studied the influence of an experimental intramammary infection with S. chromogenes on
expression of CXCR1 and eight commonly used reference genes.
Compared to biopsies, milk somatic cells allow for easy resampling but yield less RNA,
especially if the SCC is low (Jacobs et al., 2013). Because of the low yield in some samples,
we opted to isolate RNA from all milk somatic cells rather than a subpopulation (e.g.
neutrophils). RNA integrity can influence RT-qPCR results but is not easy to assess (Bustin et
al., 2009). Instead of analyzing rRNA integrity or performing a 3’:5’ integrity assay, we
designed two four-primer PCR assays amplifying three fragments of YWHAZ and CXCR1 to
test cDNA integrity. The assays are based on the fact that if integrity is too low, amplification
of large fragments of approximately 500 and 900 bp will be affected.

Chapter 6 Gene expression in milk cells
192
Polymorphism c.980A>G was not associated with CXCR1 expression in milk somatic cells.
Yet, relative CXCR1 expression increased in milk somatic cells isolated from infected quarters
which corresponds well with in vitro research showing increased CXCR1 expression in blood
neutrophils after in vitro lipopolysaccharide challenge (Leyva-Baca et al., 2008). Transcription
also increased in milk somatic cells from neighboring quarters indicating that both local and
systemic responses following the intramammary challenge can influence gene expression
(Mitterhuemer et al., 2010) and demonstrating that neighboring quarters cannot be
considered as fully independent control quarters.
The experimental IMI had a significant effect on the relative expression of 5 out of 8
candidate reference genes. Important to mention is that candidate reference genes were
selected based on a stable expression in other studies (Goossens et al., 2005; Stevens et al.,
2011; Varshney et al., 2012). Reference genes in experimental infection studies should be
stably expressed and unaffected by IMI. Validation of the reference gene to normalize gene
expression data is not always published (Bustin et al., 2009). Although normalization to a
single reference gene can cause relative large errors (Vandesompele et al., 2002), it is often
practiced (Beecher et al., 2012; Mitterhuemer et al., 2010). If the expression of this single
reference gene is affected by IMI, certain genes might be falsely identified as up- or
downregulated whereas truly up- or downregulated genes might be missed.
5. Conclusions
In conclusion, CXCR1 expression in milk somatic cells was not associated with SNP
c.980A>G but was upregulated following experimental IMI with S. chromogenes.
Additionally, differential expression was observed for housekeeping genes B2M, H2A,
HPRT1, SDHA and YWHAZ. The effect of IMI on expression of reference genes should be
tested and reported in future studies on gene expression in milk somatic cells.
6. Acknowledgements
This research was financed by a PhD grant (n° 101206) by the Agency for Innovation by
Science and Technology in Flanders (IWT Vlaanderen). Technical assistance by Kristine
Piccart, Sofie Piepers and Lars Hulpio was appreciated.

Chapter 6 Gene expression in milk cells
193
7. References
Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic Local Alignment
Search Tool. J Mol Biol. 215:403-410.
Andersen, C. L., J. L. Jensen, and T. F. Orntoft. 2004. Normalization of real-time quantitative
reverse transcription-PCR data: A model-based variance estimation approach to identify
genes suited for normalization, applied to bladder and colon cancer data sets. Cancer
Res. 64:5245-5250.
Barber, M. R., and T. J. Yang. 1998. Chemotactic activities in nonmastitic and mastitic
mammary secretions: Presence of interleukin-8 in mastitic but not nonmastitic
secretions. Clin. Diagn. Lab Immunol. 5:82-86.
Beecher, C., M. Daly, R. P. Ross, J. Flynn, T. V. McCarthy, and L. Giblin. 2012. Characterization
of the bovine innate immune response in milk somatic cells following intramammary
infection with Streptococcus dysgalactiae subspecies dysgalactiae. J. Dairy Sci. 95:5720-
5729.
Bustin, S. A., V. Benes, J. A. Garson, J. Hellemans, J. Huggett, M. Kubista, R. Mueller, T. Nolan,
M. W. Pfaffl, G. L. Shipley, J. Vandesompele, and C. T. Wittwer. 2009. The MIQE
guidelines: minimum information for publication of quantitative real-time PCR
experiments. Clin. Chem. 55:611-622.
De Vliegher, S., G. Opsomer, A. Vanrolleghem, L. A. Devriese, O. C. Sampimon, J. Sol, H. W.
Barkema, F. Haesebrouck, and A. de Kruif. 2004. In vitro growth inhibition of major
mastitis pathogens by Staphylococcus chromogenes originating from teat apices of dairy
heifers. Vet. Microbiol. 101:215-221.
Goossens, K., M. Van Poucke, A. Van Soom, J. Vandesompele, A. Van Zeveren, and L. J.
Peelman. 2005. Selection of reference genes for quantitative real-time PCR in bovine
preimplantation embryos. BMC Dev. Biol. 5:27.
Jacobs, A. A. A., J. Dijkstra, W. H. Hendriks, J. van Baal, and A. M. van Vuuren. 2013.
Comparison between stearoyl-CoA desaturase expression in milk somatic cells and in
mammary tissue of lactating dairy cows. J. Anim. Physiol. Anim. Nutr. 97:353-362.
Leyva-Baca, I., G. Pighetti, and N. A. Karrow. 2008. Genotype-specific IL8RA gene expression
in bovine neutrophils in response to Escherichia coli lipopolysaccharide challenge. Anim.
Genet. 39:298-300.

Chapter 6 Gene expression in milk cells
194
Mitterhuemer, S., W. Petzl, S. Krebs, D. Mehne, A. Klanner, E. Wolf, H. Zerbe, and H. Blum.
2010. Escherichia coli infection induces distinct local and systemic transcriptome
responses in the mammary gland. Reprod. Domest. Anim. 45:34-35.
National Mastitis Council. 1999. Laboratory Handbook on Bovine Mastitis. National Mastitis
Counc. Inc., Madison, WI.
Paape, M., J. Mehrzad, X. Zhao, J. Detilleux, and C. Burvenich. 2002. Defense of the bovine
mammary gland by polymorphonuclear neutrophil leukocytes. J. Mammary. Gland Biol.
Neoplasia 7:109-121.
Piessens, V., E. Van Coillie, B. Verbist, K. Supre, G. Braem, A. Van Nuffel, L. De Vuyst, M.
Heyndrickx, and S. De Vliegher. 2011. Distribution of coagulase-negative Staphylococcus
species from milk and environment of dairy cows differs between herds. J. Dairy Sci.
94:2933-2944.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.
Rambeaud, M., R. Clift, and G. M. Pighetti. 2006. Association of a bovine CXCR2 gene
polymorphism with neutrophil survival and killing ability. Vet. Immunol. Immunopathol.
111:231-238.
Stevens, M. G. H., L. J. Peelman, B. De Spiegeleer, A. Pezeshki, G. R. Van De Walle, L.
Duchateau, and C. Burvenich. 2011. Differential gene expression of the toll-like receptor-
4 cascade and neutrophil function in early- and mid-lactating dairy cows. J. Dairy Sci.
94:1277-1288.
Untergasser, A., H. Nijveen, X. Rao, T. Bisseling, R. Geurts, and J. A. M. Leunissen. 2007.
Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 35:W71-W74.
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe, and F.
Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by
geometric averaging of multiple internal control genes. Genome Biol. 3.
Varshney, N., A. K. Mohanty, S. Kumar, J. K. Kaushik, A. K. Dang, M. Mukesh, B. P. Mishra, R.
Kataria, S. P. Kimothi, T. K. Mukhopadhyay, D. Malakar, B. S. Prakash, S. Grover, and V. K.
Batish. 2012. Selection of suitable reference genes for quantitative gene expression
studies in milk somatic cells of lactating cows (Bos indicus). J. Dairy Sci. 95:2935-2945.

Chapter 6 Gene expression in milk cells
195
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-group
specific association between CXCR1 polymorphisms and subclinical mastitis in dairy
heifers. J. Dairy Res. 79:341-351.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2014. Association of
CXCR1 polymorphisms with apoptosis, necrosis and concentration of milk neutrophils in
early lactating dairy heifers. Res. Vet. Sci. 97:55-59.
Zuker, M. 2003. Mfold web server for nucleic acid folding and hybridization prediction.
Nucleic Acids Res. 31:3406-3415.

196

197
Chapter 7
General discussion

Chapter 7 General discussion
198

Chapter 7 General discussion
199
1. Introduction
Mastitis resistance is partly genetically determined (Rupp and Boichard, 2003). The
main aim of this thesis was to study single nucleotide polymorphisms (SNPs) in the candidate
gene CXCR1 (Chemokine (C-X-C motif) receptor 1) in relation to mammary gland immunity
and udder health. Cross-sectional and cohort observational studies as well as in vivo and in
vitro experiments were performed and the results were combined within this thesis.
Each study design has its advantages and disadvantages. Observational studies have a
high external validity but outcome variables are influenced by many other factors besides
the main predictor of interest (in casu SNPs in the CXCR1 gene). Experiments allow for
standardization but can raise ethical questions if living animals are used or lack external
validity, especially if in vitro models are used. We should mention that associations, detected
through observational studies, do not prove causality. However, consistent observation of an
association in studies on different populations using different designs indicates a real effect
(Grimes and Schulz, 2002). Therefore, results of the different studies in this thesis are
compared with each other, put next to results of (different types of) studies performed by
other research groups, and form the basis for some interesting conclusions.
In this chapter, the role of polymorphisms c.735C>G and c.980A>G in bovine
neutrophils and mastitis are discussed in detail. In addition, c.735C>G and c.980A>G
genotype frequencies in different populations are compared. Effects of SNPs are not
necessarily additive. The SNPs might interact with each other and with SNPs in other genes.
Certain combined genotypes or haplotypes could alter pathways whereas the single SNPs
have little effect (Pighetti et al., 2012). In the population of 140 Flemish heifers (Chapter
4.1), many SNPs were in full linkage disequilibrium with each other. Five haplotypes and 12
different CXCR1 genotypes were observed. In this thesis, detailed phenotypes such as
pathogen-group specific IMI status or pathogen-specific incidence rate of clinical mastitits
(IRCM) were studied. Due to the limited number of cases per CXCR1 genotype, we opted to
analyze associations with SNPs. By doing so, we maximized the likelihood of detecting
associations between SNPs and (pathogen-specific) mastitis resistance but might have
missed potential effects of specific haplotypes.
Not all detected associations are easily translated to selection perspectives. For some
traits, the lowest or highest value is beneficial whereas other traits most likely have an

Chapter 7 General discussion
200
optimal value (Table 7.1). For example, apoptosis reduces functionality of neutrophils but
also avoids tissue damage caused by necrosis (Van Oostveldt et al., 2002). Similarly, reactive
oxygen species (ROS) generation eliminates pathogens but also damages the tissue (Paape et
al., 2003). Intramammary infections (IMI) with minor pathogens can cause a intermediate
increase in somatic cell count (SCC) (Supré et al., 2011) yet were found to be associated with
a higher milk yield (Piepers et al., 2010). Additionally, pre-existing IMI or even teat apex
colonization with minor pathogens might have a protective effect against new infection with
the more detrimental major pathogens (De Vliegher et al., 2003, 2004; Lam et al., 1997).
High SCC indicates IMI but very low SCC might be associated with reduced mastitis resistance
(Detilleux et al., 2004; Schukken et al., 2003).
Table 7.1. Different traits studied in this thesis
Chapter Trait What is beneficial?
3.1, 4.2 Milk neutrophil apoptosis Optimal value
3.1, 4.2 Milk neutrophil necrosis Low value
3.1 Milk neutrophil concentration Optimal value
3.2 Blood neutrophil reactive oxygen species generation Optimal value
4.1 Major pathogen intramammary infection Low value
4.1 Minor pathogen intramammary infection Optimal value
4.1, 4.2, 5.2 Somatic cell count Optimal value
5.2 (Pathogen-specific) incidence rate of clinical mastitis Low value
5.2 Milk yield High value
Additional to CXCR1 polymorphisms, this thesis broadens the knowledge on priming of
bovine neutrophils, incidence of clinical mastitis in Flemish dairy herds and gene expression
in milk somatic cells. These results are discussed in the part “other findings”. Finally,
potential future research is suggested, and practical applications are summarized.
2. The role of CXCR1 polymorphisms in bovine neutrophils traits and mastitis
2.1. The role of polymorphism c.735C>G in bovine neutrophil traits and mastitis
Polymorphism c.735C>G has been studied for more than a decade (Table 7.2). It causes
an amino acid change in the third intracellular loop (p.His245Glu) potentially affecting G-
protein binding and signal transduction (Damaj et al., 1996; Pighetti et al., 2012). The higher

Chapter 7 General discussion
201
interleukin 8 (IL-8) induced intacellular calcium release in c.735CC neutrophils compared to
c.735GG neutrophils reinforces this hypothesis (Rambeaud and Pighetti et al., 2007). This
might also be explained by an association between SNP c.735C>G and ligand binding. In
Chapter 4.1, SNP c.735C>G was found to be in full linkage disequilibrium with newly
detected SNPs c.37A>T, c.38T>A and c.68G>A causing amino acid changes p.Ile13Tyr and
p.Gly23Glu in the N-terminus of CXCR1 known to have an important role in the first steps of
binding IL-8 (Liou et al., 2014) (Figure 7.1).
Figure 7.1. Visual representation of bovine CXCR1, updated based on the reseach included in this thesis as
compared with Figure 1.8. The amino acid sequence and positions of the identified single nucleotide
polymorphisms (SNP) in the coding DNA sequence are noted. Positions of newly detected SNPs in this thesis
are indicated as 33. Amino acids with a neutral, cationic and anionic charge are colored blue, green and orange,
respectively. Amino acids encoded by a codon with a non-synonymous and synonymous SNP have a red and
yellow outline, respectively. The structure is based on publications on human CXCR1 (Baggiolini et al., 1994;
Damaj et al., 1996). A putative disulfide bridge is indicated in purple. The SNP c.621G>A described by Pighetti et
al. (2012) was not detected in the populations included in this thesis.

Chapter 7 General discussion
202
Associations between SNP c.735C>G and IL-8 signal transduction or binding could also
be explained by linkage disequilibrium with causal mutations in regulatory regions. Beecher
et al. (2012) described a higher CXCR1 gene expression in c.735GG milk somatic cells
compared to c.735CC milk somatic cells but failed to provide sufficient experimental data
such as assessment of gDNA contamination, validation of reference genes and qPCR
validation (e.g. PCR efficiency) (Chapter 6). Nevertheless, results correspond well with the
previously described higher IL-8 receptor expression in c.735GG neutrophils compared to
c.735CC neutrophils (Rambeaud and Pighetti, 2007).
The higher CXCR1 functionality in c.735GG cows might affect neutrophil migration,
viability and activity. In vitro, c.735GG blood neutrophils showed a higher rhIL-8 induced
upregulation of adhesion molecules and more chemotaxis to rhIL-8 than c.735CC blood
neutrophils (Rambeaud and Pighetti, 2005). In contrast, rhIL-8 incubation decreased
apoptosis more in c.735CC compared to c.735GG neutrophils (Rambeaud et al., 2006). In
Chapter 3.1, IMI status was associated with a higher decrease of % milk neutrophil apoptosis
in c.735GG compared to c.735CC heifers. At first sight, these results seem contradictory.
However, in vitro rhIL-8 incubation differs from in vivo IMI. In the large majority of the
infected quarters, coagulase-negative staphylococci were isolated. Yet, the duration of
infection was unknown. More chronic IMI in c.735CC compared to c.735GG heifers could
explain the higher % milk neutrophil apoptosis in infected compared to uninfected quarters.
Rambeaud et al. (2006) demonstrated a higher phorbol 12-myristate 13-acetate (PMA)
induced ROS generation in c.735GG blood neutrophils compared to c.735CC blood
neutrophils. Yet, this association could not be confirmed in our research (Chapter 3.2). Single
nucleotide polymorphisms c.735C>G was neither associated with PMA or opsonized
zymosan particle (OZP) induced ROS generation nor with the effect of rbIL-8 incubation on
ROS generation. The clear priming effect of IL-8 on ROS generation as described in human
neutrophils was not observed in our study (Daniels et al., 1992; Wozniak et al., 1993) (see
further).
The higher neutrophil migration and activity (Rambeaud and Pighetti, 2005; Rambeaud
et al., 2006) correspond well with the lower incidence of subclinical mastitis in c.735GG cows
observed by Youngerman et al. (2004). As host responses and impact on milk yield heavily
depend on the invading mastitis pathogen, associations between CXCR1 polymorphisms and
udder health (and mammary gland immunity) are ideally studied using pathogen-specific

Chapter 7 General discussion
203
data (Piepers et al., 2010; Schukken et al., 2009; Schukken et al., 2011). We studied
pathogen-specific prevalence of subclinical heifer mastitis but detected no association with
SNP c.735C>G (Chapter 4.1). Yet, in a larger study, c.735GG cows were found to have a lower
S. aureus incidence rate of clinical mastitis (IRCM) than c.735CC cows. Additionally, overall, S.
dysgalactiae and E. coli IRCM were non-significantly lower in c.735GG cows compared to
c.735CC cows (Chapter 5.2) contradicting the previously reported higher overall IRCM in
c.735GG cows compared to c.735CC cows (Galvao et al., 2011).
Beecher et al. (2010) reported a lower SCC in c.735GG cows compared to c.735CC
cows. Yet, the analysis was performed on data originating from only one experimental herd.
Similar to other studies on multiple commercial herds (Beecher et al., 2010; Galvao et al.,
2011; Goertz et al., 2009), we detected no association between c.735C>G and SCC (Chapter
4.1 and 5.2). Additional to a lower subclinical masitis incidence (Youngerman et al., 2004)
and S. aureus IRCM, c.735GG cows out-produced c.735CC cows with 0.8 kg milk/day
(Chapter 5.2). Results correspond well with the higher milk fat yield observed in offspring of
c.735GG sires compared to offspring of c.735CG sires (Beecher et al., 2010) but contradict
the previously described lower milk yield in c.735GG cows compared to c.735CG cows in
smaller U.S. populations (Galvao et al., 2011; Youngerman et al., 2004).
Based on the higher CXCR1 functionality (Rambeaud and Pighetti, 2007), the higher
neutrophil migration and activity (Rambeaud and Pighetti, 2005; Rambeaud et al., 2006), the
lower subclinical mastitis incidence (Youngerman et al., 2004) and the lower S. aureus IRCM
(Chapter 5.2) in c.735GG cows compared to c.735CC cows, we hypothize allele c.735G to be
associated with a higher mastitis resistance (Figure 7.2). Some contradictory findings suggest
that certain genotypes are beneficial in one population but detrimental in others. Not only
mastitis resistance but also mastitis tolerance is partly genetically determined (Detilleux,
2009). Under equal pathogen burden, host responses might be more severe in c.735GG cows
compared to c.735CC. This could result in a higher likelihood of clinical symptoms and more
inflammation-induced tissue damage explaining the higher overall IRCM and lower milk yield
(Galvao et all., 2011), respectively. However, we should mention that the sample size in
most studies is limited (Table 7.2) and that some contradictory findings might be explained
by false positive or negative results.
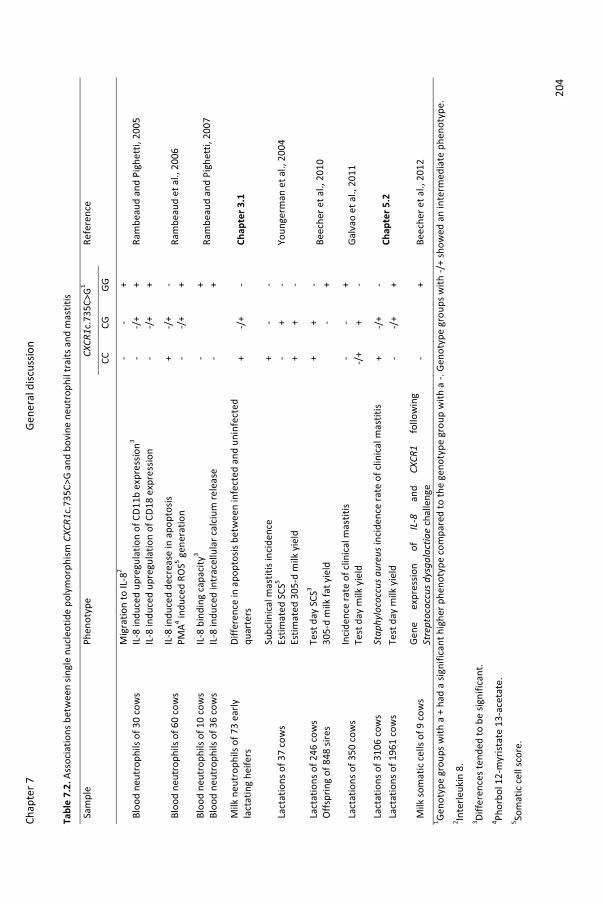
Chap
ter 7
Gene
ral d
iscus
sion
204
Tabl
e 7.
2. A
ssoc
iatio
ns b
etw
een
singl
e nu
cleo
tide
poly
mor
phism
CXC
R1c.
735C
>G a
nd b
ovin
e ne
utro
phil
trai
ts a
nd m
astit
is
Sam
ple
Ph
enot
ype
CX
CR1c
.735
C>G
1
Refe
renc
e
CC
CG
GG
Bloo
d ne
utro
phils
of 3
0 co
ws
M
igra
tion
to IL
-82
-
- +
Ra
mbe
aud
and
Pigh
etti,
200
5
IL-8
indu
ced
upre
gula
tion
of C
D11b
exp
ress
ion3
-
-/+
+
IL
-8 in
duce
d up
regu
latio
n of
CD1
8 ex
pres
sion
-
-/+
+
Bloo
d ne
utro
phils
of 6
0 co
ws
IL
-8 in
duce
d de
crea
se in
apo
ptos
is
+
-/+
-
Ram
beau
d et
al.,
200
6
PMA4 in
duce
d RO
S5 gen
erat
ion
-
-/+
+
Bloo
d ne
utro
phils
of 1
0 co
ws
IL
-8 b
indi
ng c
apac
ity3
-
+
Ra
mbe
aud
and
Pigh
etti,
200
7 Bl
ood
neut
roph
ils o
f 36
cow
s
IL-8
indu
ced
intr
acel
lula
r cal
cium
rele
ase
-
+
Milk
neu
trop
hils
of 7
3 ea
rly
lact
atin
g he
ifers
Diffe
renc
e in
apo
ptos
is be
twee
n in
fect
ed a
nd u
ninf
ecte
d qu
arte
rs
+
-/+
-
Chap
ter 3
.1
Lact
atio
ns o
f 37
cow
s
Subc
linic
al m
astit
is in
cide
nce
+
- -
Yo
unge
rman
et a
l., 2
004
Es
timat
ed S
CS5
-
+ -
Estim
ated
305
-d m
ilk y
ield
+ +
-
Lact
atio
ns o
f 246
cow
s
Test
day
SCS
3
+ +
-
Beec
her e
t al.,
201
0 O
ffspr
ing
of 8
48 si
res
30
5-d
milk
fat y
ield
-
+
Lact
atio
ns o
f 350
cow
s
Inci
denc
e ra
te o
f clin
ical
mas
titis
- -
+
Galv
ao e
t al.,
201
1
Test
day
milk
yie
ld
-/
+ +
-
Lact
atio
ns o
f 310
6 co
ws
St
aphy
loco
ccus
aur
eus i
ncid
ence
rate
of c
linic
al m
astit
is
+
-/+
-
Chap
ter 5
.2
Lact
atio
ns o
f 196
1 co
ws
Te
st d
ay m
ilk y
ield
- -/
+ +
Milk
som
atic
cel
ls of
9 c
ows
Gene
ex
pres
sion
of
IL-8
an
d CX
CR1
follo
win
g St
rept
ococ
cus d
ysga
lact
iae
chal
leng
e
-
+
Beec
her e
t al.,
201
2 1 Ge
noty
pe g
roup
s with
a +
had
a si
gnifi
cant
hig
her p
heno
type
com
pare
d to
the
geno
type
gro
up w
ith a
-. G
enot
ype
grou
ps w
ith -/
+ sh
owed
an
inte
rmed
iate
phe
noty
pe.
2 Inte
rleuk
in 8
. 3 Di
ffere
nces
tend
ed to
be
signi
fican
t. 4 Ph
orbo
l 12-
myr
istat
e 13
-ace
tate
. 5 So
mat
ic c
ell s
core
.

Chapter 7 General discussion
205
Figure 7.2. From genotype to phenotype: effects of CXCR1 polymorphism c.735C>G. 1c.735GG milk somatic cells showed a higher CXCR1 gene expression compared to c.735CC milk somatic cells
(Beecher et al., 2012). 2IL-8 receptor expression, IL-8 affinity and IL-8 induced intracellular calcium release was higher in c.735GG
neutrophils compared to c.735CC neutrophils (Rambeaud and Pighetti, 2007). 3IL-8 induced upregulation of adhesion molecules and chemotaxis to IL-8 was higher in c.735GG neutrophils
compared to c.735CC neutrophils (Rambeaud and Pighetti, 2005). 4IL-8 induced decrease in apoptosis and PMA induced ROS generation was higher in c.735GG neutrophils
compared to c.735CC neutrophils (Rambeaud et al, 2006). 5Subclinical mastitis incidence and milk yield were lower in c.735GG cows compared to c.735CC cows
(Youngerman et al., 2004). 6Overall clinical mastitis incidence was lower in c.735GG cows compared to c.735CC cows (Galvao et al., 2011). 7Staphylococcus aureus clinical mastitis incidence was lower and milk yield was higher in c.735GG cows
compared to c.735CC cows (Chapter 5.2). 8Somatic cell count was lower in c.735GG compared to c.735CC cows and milk fat yield was higher in offspring
from c.735GG sires compared to offspring from c.735CC sires (Beecher et al., 2010).

Chapter 7 General discussion
206
2.2. The role of polymorphism c.980A>G in bovine neutrophil traits and mastitis
Results in this thesis indicate that not only SNP c.735C>G but also SNP c.980A>G plays
a role in bovine neutrophil traits and mastitis. Single nucleotide polymorphism c.980A>G was
found to be in full linkage disequilibrium with SNP c.995A>G (Chapter 4.1). These SNPs cause
amino acid changes p.Lys327Arg and p.His332Arg in the C-terminal region known to be
important for internalization and chemotaxis of human neutrophils (Richardson et al., 2003)
(Figure 7.1). Additionally, SNP c.980A>G might be in linkage disequilibrium with causal
mutations in regulatory regions. We detected no association between c.980A>G genotype
and CXCR1 gene expression in milk somatic cells but the sample size (n = 8 cows) might have
been too small to do so (Chapter 6).
In this thesis, neutrophil migration was not studied in specific. Yet, the larger
difference in milk neutrophil concentration between infected and uninfected quarters
(Chapter 3.1) and the higher SCC in the early fase of experimental infection (Chapter 4.2)
indicate neutrophil migration to be higher in c.980GG heifers compared to c.980AG heifers.
Milk neutrophil viability was studied in healthy, naturally infected and experimentally
infected quarters (Chapter 3.1 and 4.2) Regardless of infection stage, necrosis has
detrimental effects as it decreases milk neutrophil functionality and causes leakage of
cellular content being toxic to the surrounding tissue (Mehrzad et al., 2004; Van Oostveldt et
al., 2001). Apoptosis is unwanted in the early stage of infection as it is also associated with
decreased neutrophil functionality but is vital to elegantly remove neutrophils in later stage
of infection (Rainard and Riollet, 2006; Van Oostveldt et al., 2002). In uninfected quarters, %
milk neutrophil apoptosis was lower in c.980AG compared to c.980GG heifers whereas no
differences in % milk neutrophil necrosis were observed (Chapter 3.1). Yet, during early
infection with S. chromogenes, % milk neutrophil apoptosis and necrosis were higher in
c.980AG compared to c.980GG heifers (Chapter 4.2). The lower neutrophil viability during
early infection could hinder c.980AG heifers to rapidly eliminate invading pathogens
(Mehrzad et al., 2004; Van Oostveldt et al., 2002). Similar to SNPc.735C>G, SNP c.980A>G
neither associated with PMA or opsonized zymosan particle (OZP) induced ROS generation
nor with the effect of rbIL-8 incubation on ROS generation (Chapter 3.2).
In Chapter 4.1, associations between CXCR1 polymorphism and subclinical mastitis in
early lactating heifers were studied. By stratifying between IMI caused by coagulase-
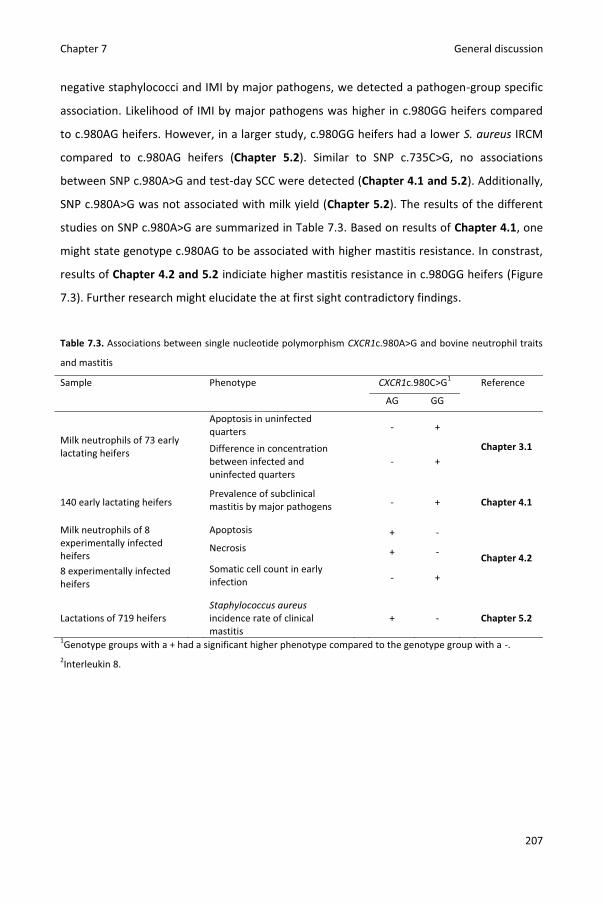
Chapter 7 General discussion
207
negative staphylococci and IMI by major pathogens, we detected a pathogen-group specific
association. Likelihood of IMI by major pathogens was higher in c.980GG heifers compared
to c.980AG heifers. However, in a larger study, c.980GG heifers had a lower S. aureus IRCM
compared to c.980AG heifers (Chapter 5.2). Similar to SNP c.735C>G, no associations
between SNP c.980A>G and test-day SCC were detected (Chapter 4.1 and 5.2). Additionally,
SNP c.980A>G was not associated with milk yield (Chapter 5.2). The results of the different
studies on SNP c.980A>G are summarized in Table 7.3. Based on results of Chapter 4.1, one
might state genotype c.980AG to be associated with higher mastitis resistance. In constrast,
results of Chapter 4.2 and 5.2 indiciate higher mastitis resistance in c.980GG heifers (Figure
7.3). Further research might elucidate the at first sight contradictory findings.
Table 7.3. Associations between single nucleotide polymorphism CXCR1c.980A>G and bovine neutrophil traits
and mastitis
Sample Phenotype CXCR1c.980C>G1 Reference
AG GG
Milk neutrophils of 73 early lactating heifers
Apoptosis in uninfected quarters
- +
Chapter 3.1
Difference in concentration between infected and uninfected quarters
- +
140 early lactating heifers Prevalence of subclinical
mastitis by major pathogens
- +
Chapter 4.1
Milk neutrophils of 8 experimentally infected heifers
Apoptosis + -
Chapter 4.2 Necrosis + -
8 experimentally infected heifers
Somatic cell count in early infection
- +
Lactations of 719 heifers Staphylococcus aureus
incidence rate of clinical mastitis
+ -
Chapter 5.2
1Genotype groups with a + had a significant higher phenotype compared to the genotype group with a -. 2Interleukin 8.
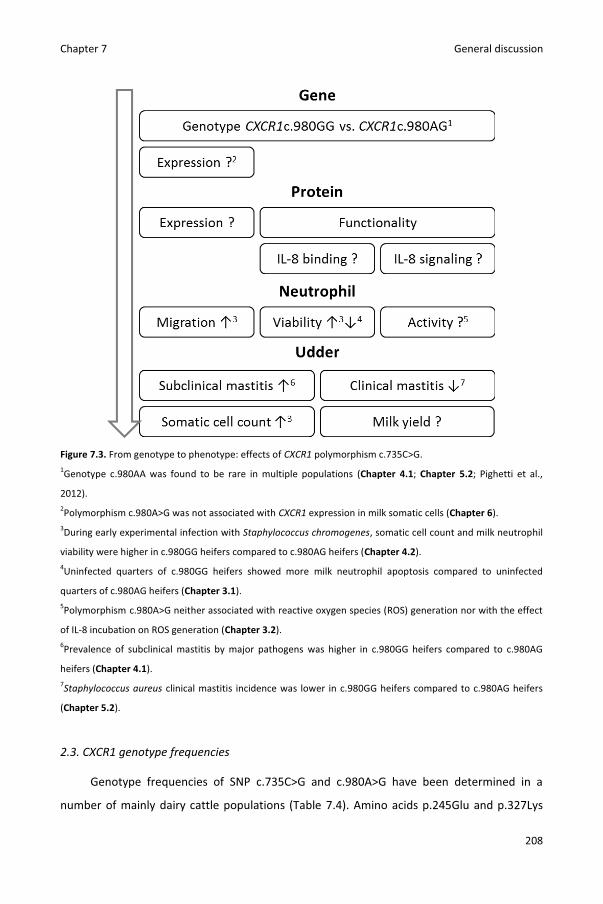
Chapter 7 General discussion
208
Figure 7.3. From genotype to phenotype: effects of CXCR1 polymorphism c.735C>G. 1Genotype c.980AA was found to be rare in multiple populations (Chapter 4.1; Chapter 5.2; Pighetti et al.,
2012). 2Polymorphism c.980A>G was not associated with CXCR1 expression in milk somatic cells (Chapter 6). 3During early experimental infection with Staphylococcus chromogenes, somatic cell count and milk neutrophil
viability were higher in c.980GG heifers compared to c.980AG heifers (Chapter 4.2). 4Uninfected quarters of c.980GG heifers showed more milk neutrophil apoptosis compared to uninfected
quarters of c.980AG heifers (Chapter 3.1). 5Polymorphism c.980A>G neither associated with reactive oxygen species (ROS) generation nor with the effect
of IL-8 incubation on ROS generation (Chapter 3.2). 6Prevalence of subclinical mastitis by major pathogens was higher in c.980GG heifers compared to c.980AG
heifers (Chapter 4.1). 7Staphylococcus aureus clinical mastitis incidence was lower in c.980GG heifers compared to c.980AG heifers
(Chapter 5.2).
2.3. CXCR1 genotype frequencies
Genotype frequencies of SNP c.735C>G and c.980A>G have been determined in a
number of mainly dairy cattle populations (Table 7.4). Amino acids p.245Glu and p.327Lys

Chapter 7 General discussion
209
encoded by c.735C and c.980A, respectively, were found to be conserved across species
suggesting c.735C and c.980A to be “the wild type” (Pighetti et al., 2012). Nevertheless, high
frequencies of c.735CG, c.735GG, c.980AG and c.980GG (“mutant” genotypes) have been
reported as well. In most Holstein populations, allele frequencies of c.735G were close to
50% resulting in approximately 25% c.735CC, 50% c.735CG and 25% c.735GG (Galvao et al.,
2011; Leyva-Baca et al., 2008; Pighetti et al., 2012; Chapter 4.1 and 5.2). Different
frequencies were observed in a population of Irish Holstein sires. Only 1% c.735CC was
observed (Beecher et al., 2010). Genotype frequencies of c.735C>G were also studied in two
small Jersey populations. Allele frequencies of c.735G were even higher compared to the
Holstein populations resulting in more than 60% c.735GG (Beecher et al., 2010; Youngerman
et al., 2004). Both Pighetti et al. (2012) and we observed high allele frequencies of c.980G in
Holstein populations. More than half of the animals had genotype c.980GG. Additionally,
both research groups detected strong linkage disequilibrium between c.735C>G and
c.980A>G with very limited frequencies of haplotype c.735G980A.
In Chapter 5.2, c.735GG cows were found to outproduce c.735CC cows. Intense
selection for milk yield during the past decennia might explain the low frequencies of the
“wild” genotypes c.735CC and c.980AA. To test this hypothesis, we determined c.735C>G
and c.980A>G genotype frequencies in a sample of Belgian Blue animals (n =50, belonging to
5 different herds) selected for muscle mass rather than milk yield (Grobet et al., 1997).
Genotype at SNP c.735C>G and c.980A>G were determined using our fluorescent multiprobe
PCR assay (Chapter 5.2). Chi-square tests demonstrated genotype frequencies of c.735C>G
and c.980A>G to differ between the 50 Belgian Blue animals and the 3,106 Holstein cows
included in Chapter 5.2 [P( ≥ 54) < 0.0001 and P( ≥ 10) < 0.01, respectively].
Unlike Holstein and Jersey animals, a high frequency of c.735CC (70%) was detected in
Belgian Blues. Results further indicate that selection for milk yield might have caused the
high c.735G allele frequency. However, allele frequency of c.980G was even higher than in
Holstein studies (Table 7.4). If selection pressure against c.980A existed, it occurred both in
Holsteins and in Belgian Blue.

Chapter 7 General discussion
210
Table 7.4. CXCR1 genotype frequencies (in %) in different studies Country Breed n Sex CXCR1 genotype Reference
c.735C>G c.980A>G
CC CG GG AA AG GG
U.S. Jersey 42 Cows 2 22 76 Youngerman et al., 2004
U.S. Holstein 37 Cows 24 65 11 Youngerman et al., 2004
Canada Holstein 334 Sires 27 55 19 Leyva-Baca et al., 2008
Ireland Jersey 47 Cows 13 25 62 Beecher et al., 2010
Ireland Holstein 77 Cows 14 50 36 Beecher et al., 2010
Ireland Holstein 848 Sires ..1 68 31 Beecher et al., 2010
U.S. Holstein 350 Cows 22 50 28 Galvao et al., 2011
U.S. Holstein 88 Cows 30 45 25 15 35 50 Pighetti et al., 2012
Belgium Holstein 140 Cows 26 48 26 2 30 68 Chapter 4.1
Belgium Holstein 3106 Cows 25 48 27 5 34 61 Chapter 5.2
Belgium Belgian Blue 50 Both 70 26 4 0 18 82 Verbeke et al., unpublished data
3. Other findings
3.1 Neutrophil priming
The clear priming effect of IL-8 on ROS generation as described in human neutrophils
(Daniels et al., 1992; Wozniak et al., 1993) was not observed in Chapter 4.1. To our surprise,
rbIL-8 incubation for 2 or 6 h even had inhibitory effects on OZP induced ROS generation.
Incubation with rbIL-8 for 18 h primes blood neutrophils for higher Mannheimia
haemolytica-induced ROS generation (Mitchell et al., 2003). However, this effect might be
caused by an IL-8 induced delay of apoptosis. No priming effect on ROS generation was
observed when shorter incubation periods were applied (Galligan and Coomber, 2000;
Mitchell et al., 2003). Human CXCR1 but not hCXCR2 is important for the priming effect on
ROS generation (Jones et al., 1996; Jones et al., 1997). Functional differences between the
receptors were attributed to amino acid sequence differences in the cytoplasmic tail (C-
terminus) causing a faster receptor phosphorylation and internalization of hCXCR2
compared to hCXCR1 (Nasser et al., 2005; Richardson et al., 1998; Richardson et al., 2003).
Interestingly, the amino acid sequences of the cytoplasmic tails of bovine CXCR1 and CXCR2
are highly similar and both share more identity with hCXCR2 than with hCXCR1 (Lahouassa et
al., 2008)(Figure 7.4, Table 7.5).

Chapter 7 General discussion
211
Figure 7.4. Multiple alignment of the amino acid sequences of the cytoplasmic tails of human and bovine IL-8
receptors performed using Clustal W (Larkin et al., 2007). Sequences were retrieved from GenBank
(NP_000625.1, NP_001161770.1, NP_001098508.1, and NP_001094755.1, respectively).
Table 7.5. Amino acid sequence identity (%) between the cytoplasmic tails of human and bovine IL-8 receptors
hCXCR1 hCXCR2 bCXCR1 bCXCR2 hCXCR1 100 hCXCR2 55 100 bCXCR1 54 79 100 bCXCR2 59 85 95 100
Sequences were retrieved from GenBank (NP_000625.1, NP_001161770.1, NP_001098508.1, and
NP_001094755.1, respectively) and compared with each other using protein BLAST (Altschul et al., 1997).
3.2 Clinical mastitis in Flanders
In Chapter 5.1, the pathogen-specific incidence rate of clinical mastitis (IRCM) was
estimated in Flanders. Few Flemish dairy producers record or sample clinical mastitis cases.
Nevertheless, the relatively high response rate in our study (79%) demonstrates high
awareness of the importance of clinical mastitis. With a herd mean and median of 7.4 and
5.3 quarter cases/10,000 cow-days at risk, IRCM was estimated to be moderate compared to
other regions (Barkema et al., 1998; Barnouin et al., 2005; Bradley et al., 2007; Olde
Riekerink et al., 2008; Wolff et al., 2012). The previously described higher IRCM in early
lactation in heifers compared to multiparous cows (Barkema et al., 1998; McDougall et al.,
2007) was confirmed in our study stressing the importance of heifer mastitis (De Vliegher et
al., 2012). Similar to clinical mastitis samples submitted to the central milk quality lab of the
Milk Control Centre, environmental pathogens S. uberis and E. coli were the most frequently
isolated pathogens (Milk Control Centre Flanders, 2012). Most cases showed mild clinical
signs. Although isolation of E. coli was more likely in moderate and severe cases compared
to mild cases, less than half of the severe cases were culture positive for E. coli and isolation
of E. coli was also common in mild cases. Solely based on severity, pathogens cannot be
identified. The IRCM showed a large between-herd variation, suggesting management plays

Chapter 7 General discussion
212
an important role. In some herds, few cases were sampled whereas in other herds the
number of sampled cases equaled the average number of lactating animals. Especially when
bulk milk SCC was low, producers were not aware of the high IRCM on their herd at the start
of the study. Due to lack of clinical mastitis records, premium payments for low bulk milk
SCC, and a penalty system for high bulk milk SCC, udder health on Flemish herds is mainly
quantified using bulk milk SCC records. However, bulk milk SCC does not reflect the IRCM
being an equally important udder health parameter (Schukken et al., 2003). In our study,
some herds with low bulk milk SCC had high IRCM and vice versa (Figure 7.5).
Figure 7.5. Scatterplot of the incidence rate of clinical mastitis (IRCM) by average bulk milk somatic cell count
(in 1,000 cells/mL) in 50 randomly selected Flemish dairy herds. The median IRCM (5.3) and bulk milk somatic
cell count (220) are indicated with lines. White markers represent herds with an automatic milking system.
Due to the relatively limited number of herds (n = 50), an extensive herd-level risk
factor analysis as described for Dutch herds was not feasible (Barkema et al., 1999; Schukken
et al., 1990; Schukken et al., 1991). Because of the frequent isolation of so-called
environmental pathogens, we focused on associations between pathogen-specific IRCM and
herd hygiene. We opted to categorize herds as clean or dirty based on the udder hygiene
score of a number of cows. Udder hygiene scores depend less on the last cleaning and better
reflect teat exposure to dirt and manure compared to other hygiene measurement such as
cubicle cleanliness (Schreiner and Ruegg, 2003; Schukken et al., 1990). More frequent
scoring would have allowed a better estimation of the herd hygiene. Nevertheless, herds
0
5
10
15
20
25
0 50 100 150 200 250 300 350 400 450 500
Qua
rter
cas
es p
er 1
0,00
0 co
w-d
ays a
t risk
Average bulk milk somatic cell count

Chapter 7 General discussion
213
categorized as dirty had higher overall and E. coli IRCM compared to herds categorized as
clean which corresponds well with British research demonstrating a higher risk of clinical
mastitis in general and E. coli clinical mastitis in specific in cows with dirty udders (Breen et
al., 2009). Although hygiene was measured at herd level in our study and at cow level by
Breen et al. (2009), corresponding associations suggest causality.
3.3 Gene expression in milk somatic cells
A number of experimental intramammary infection studies have supplied more insight
in host-pathogen interactions (Bannerman, 2009; Schukken et al., 2011). Yet, our knowledge
on mammary gland immunity and abilities to enhance resistance is far from complete.
Measurement of gene expression allows for identification of up- or down-regulated genes
and pathways during pathogen-specific immune responses. In Chapter 6, gene expression
was measured before and after experimental IMI with S. chromogenes. No associations were
detected between CXCR1 polymorphisms and CXCR1 expression. Yet, the research provided
interesting information for other scientists studying gene expression in milk somatic cells.
Both in our and other research (Jacobs et al., 2013), some samples of milk somatic cells
yielded little RNA. Normalization to reference genes controls for variation of mRNA content,
enzymatic efficiencies and other factors across samples. However, normalization to
reference genes with unstable expression can cause considerable bias., especially when only
one reference gene is used (Vandesompele et al., 2002). Reference genes in experimental
infection studies should be stably expressed and unaffected by IMI. Yet, expression of 5 out
of 8 tested candidate reference genes was affected in our study. We recommend to test and
report the effect of IMI on expression of reference genes in future studies on gene
expression in milk somatic cells.
4. Future research
Despite the relatively high number of scientific articles published on the subject, many
further research opportunities exist. In the following paragraphs, some suggestions are
made.
Additional to CXCR1, CXCR2 should be screened for polymorphisms by DNA
sequencing. Subsequently, associations with similar traits as in the CXCR1 studies could be

Chapter 7 General discussion
214
tested. New studies could be performed but blood or DNA samples of previous studies could
also be genotyped for CXCR2 polymorphisms. Fluorescent multiprobe PCR assays as
described in Chapter 5.2 could be designed to allow rapid CXCR2 genotyping. Using a similar
approach, other genes involved in the mammary gland immunity could be studied in more
detail. Preferably, genes in close proximity of markers associated with mastitis resistance in
genome-wide association studies are selected.
The impact of CXCR1 polymorphisms on neutrophil functionality could be further
unraveled by in vitro studies focusing on new phenotypes such as phagocytosis and
diapedesis (Stevens et al., 2011). Association studies that focused entirely on SNP c.735C>G
(Rambeaud and Pighetti, 2005; Rambeaud et al., 2006; Rambeaud and Pighetti, 2007) could
be repeated for SNP c.980A>G and other CXCR1 or CXCR2 polymorphisms. As effects of SNPs
might depend on the ligand concentration, we suggest testing the association between
genotype and neutrophil functionality under different concentrations of IL-8. This way,
differences in dose-response curves could be analyzed. The unexpected inhibitory effects of
IL-8 on neutrophil ROS generation (Chapter 3.2) indicate the need for more research on the
role of IL-8, CXCR1 and CXCR2 in bovine innate immunity. Research findings on human
neutrophils do not necessarily apply to bovine neutrophils.
The sample size in the heifer mastitis study (Chapter 4.1) allowed a pathogen-group
specific analysis but not a pathogen-specific analysis as performed in the clinical mastitis
study (Chapter 5.2). A longitudinal study in which milk samples of a larger population of
heifers and multiparous cows are cultured on a regular basis would allow analysis of
associations between CXCR1 SNPs and pathogen-specific incidence rate of subclinical
masitis. Because of the absence of association between CXCR1 genotype and SCC in many
studies (Galvao et al., 2011; Goertz et al., 2009)(Chapter 4.1 and 5.2) and the detected
pathogen-specific associations in our studies (Chapter 4.1 and 5.2), we discourage
observational studies solely using SCC traits. The results of the clinical mastitis study
(Chapter 5.2) also should motivate to perform experimental infection studies with S. aureus.
Methods described in Chapter 6 could be used to study differences in host response in
detail.
As previously mentioned, CXCR1 polymorphisms do not only have the potential to
affect mastitis resistance but might also influence mastitis tolerance. This hypothesis could
be further tested in experimental studies or cohort studies comparing milk yield losses

Chapter 7 General discussion
215
following pathogen-specific subclinical or clinical mastitis in animals with different CXCR1
genotypes.
Because of the small samples sizes in most studies and the variable associations within
different populations, validation of the research is required prior to introduction of CXCR1
genotypes in marker assisted selection. With little effort, we could genotype sires currently
available with the fluorescent multiprobe qPCR assay and analyze records of offspring in
detail. Such study could include recordsfrom Nordic countries to analyze the relation with
veterinary treated clinical mastitis.
Last but not least, the study on the IRCM in Flemish herds (Chapter 5.1) indicates
mastitis research as well as prevention and control in Flanders should mainly focus on the
so-called environmental pathogens. Practical research on how we can motivate producers to
record and prevent clinical mastitis cases is warranted. Housing and management practices
rapidly evolve but the influences of many new techniques (e.g. the use of recycled manure
solids) on herd hygiene and udder health are often poorly understood.
5. Conclusions and practical applications
The associations detected in the genome-wide association study (Sodeland et al.,
2011) and the multiple candidate gene association studies (Table 7.2 and 7.3) strongly
indicate CXCR1 to be an important gene for mastitis resistance. The upregulated CXCR1
expression in milk somatic cells of infected and neighboring quarters following IMI further
substantiates this finding (Chapter 6).
Several studies indicate alleles c.735G and c.980G to be associated with higher mastitis
resistance (Beecher et al., 2010; Rambeaud et al., 2006; Rambeaud and Pighetti, 2005;
Rambeaud and Pighetti, 2007; Youngerman et al., 2004, Chapter 4.2 and 5.2). After
validation in a large (international) population (see future research), effects of both SNPs
might be estimated precisely and this information might be used in the estimation of
breeding values. Linkage disequilibrium between markers in DNA chips and SNPs c.735C>G
and c.980A>G could be determined. Markers in high linkage disequilibrium with SNPs
c.735C>G and c.980A>G could be used to select against certain CXCR1 genotypes.
Besides potential applications for selection, this thesis offers more insight in the
mammary gland immunity and stresses the important role of CXCR1. In human medicine,

Chapter 7 General discussion
216
therapeutic agents targeting CXCR1 are being tested in relation to breast cancer (Singh et al.,
2013). Similar agents targeting bovine CXCR1 could enhance mastitis resistance, mastitis
tolerance or both.
The results of the clinical mastitis study (Chapter 5.1) can be used by producers and
veterinarians to compare the IRCM on a herd with the mean and median IRCM in Flanders.
However, to do so, producers need to record cases of clinical mastitis. Regular screening of
the udder hygiene score of a number of cows can be used to monitor herd hygiene.
Measurements to improve herd hygiene could reduce the overall number of clinical mastitis
cases and especially the number of E. coli cases. To improve udder health, innovative tools
such as marker assisted selection for certain CXCR1 genotypes should always be combined
with a more “classic” approach as described in the NMC 10-point mastitis control program
(National Mastitis Council, 2014).
6. References
Altschul, S. F., T. L. Madden, A. A. Schaffer, J. H. Zhang, Z. Zhang, W. Miller, and D. J. Lipman.
1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search
programs. Nucleic Acids Res. 25:3389-3402.
Baggiolini, M., B. Dewald, and B. Moser. 1994. Interleukin-8 and related chemotactic
cytokines - CXC and CC chemokines. Adv. Immunol. 55:97-179.
Bannerman, D. D. 2009. Pathogen-dependent induction of cytokines and other soluble
inflammatory mediators during intramammary infection of dairy cows. J. Anim Sci. 87:10-
25.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, H. Wilmink, G. Benedictus, and A.
Brand. 1998. Incidence of clinical mastitis in dairy herds grouped in three categories by
bulk milk somatic cell counts. J. Dairy Sci. 81:411-419.
Barkema, H. W., Y. H. Schukken, T. J. Lam, M. L. Beiboer, G. Benedictus, and A. Brand. 1999.
Management practices associated with the incidence rate of clinical mastitis. J. Dairy Sci.
82:1643-1654.
Barnouin, J., S. Bord, S. Bazin, and M. Chassagne. 2005. Dairy management practices
associated with incidence rate of clinical mastitis in low somatic cell score herds in
France. J. Dairy Sci. 88:3700-3709.

Chapter 7 General discussion
217
Beecher, C., M. Daly, S. Childs, D. P. Berry, D. A. Magee, T. V. McCarthy, and L. Giblin. 2010.
Polymorphisms in bovine immune genes and their associations with somatic cell count
and milk production in dairy cattle. BMC Genet. 11:99.
Beecher, C., M. Daly, R. P. Ross, J. Flynn, T. V. McCarthy, and L. Giblin. 2012. Characterization
of the bovine innate immune response in milk somatic cells following intramammary
infection with Streptococcus dysgalactiae subspecies dysgalactiae. J. Dairy Sci. 95:5720-
5729.
Bradley, A. J., K. A. Leach, J. E. Breen, L. E. Green, and M. J. Green. 2007. Survey of the
incidence and aetiology of mastitis on dairy farms in England and Wales. Vet. Rec.
160:253-258.
Breen, J. E., M. J. Green, and A. J. Bradley. 2009. Quarter and cow risk factors associated with
the occurrence of clinical mastitis in dairy cows in the United Kingdom. J. Dairy Sci.
92:2551-2561.
Damaj, B. B., S. R. McColl, K. Neote, N. Songqing, K. T. Ogborn, C. A. Hebert, and P. H.
Naccache. 1996. Identification of G-protein binding sites of the human interleukin-8
receptors by functional mapping of the intracellular loops. FASEB J. 10:1426-1434.
Daniels, R. H., M. J. Finnen, M. E. Hill, and J. M. Lackie. 1992. Recombinant human monocyte
Il-8 primes NADPH-oxidase and phospholipase-A2 activation in human neutrophils.
Immunology 75:157-163.
Detilleux, J. C.. 2004. Neutrophils in the war against Staphylococcus aureus: predator-prey
models to the rescue. J. Dairy Sci. 87:3716-3724.
Detilleux, J. C. 2009. Genetic factors affecting susceptibility to udder pathogens. Vet.
Microbiol. 134:157-164.
De Vliegher, S., H. Laevens, L. A. Devriese, G. Opsomer, J. L. Leroy, H. W. Barkema, and A. de
Kruif A. 2003. Prepartum teat apex colonization with Staphylococcus chromogenes in
dairy heifers is associated with low somatic cell count in early lactation. Vet. Microbiol.
92:245-252.
De Vliegher, S., G. Opsomer, A. Vanrolleghem, L. A. Devriese, O. C. Sampimon, J. Sol, H. W.
Barkema, F. Haesebrouck, and A. de Kruif. 2004. In vitro growth inhibition of major
mastitis pathogens by Staphylococcus chromogenes originating from teat apices of dairy
heifers. Vet. Microbiol. 101:215-221.

Chapter 7 General discussion
218
De Vliegher, S., L. K. Fox, S. Piepers, S. McDougall, and H. W. Barkema. 2012. Invited review:
Mastitis in dairy heifers: nature of the disease, potential impact, prevention, and control.
J. Dairy Sci. 95:1025-1040.
Galligan, C. L., and B. L. Coomber. 2000. Effects of human IL-8 isoforms on bovine neutrophil
function in vitro. Vet. Immunol. Immunopathol. 74:71-85.
Galvao, K. N., G. M. Pighetti, S. H. Cheong, D. V. Nydam, and R. O. Gilbert. 2011. Association
between interleukin-8 receptor-alpha (CXCR1) polymorphism and disease incidence,
production, reproduction, and survival in Holstein cows. J. Dairy Sci. 94:2083-2091.
Goertz, I., C. Baes, C. Weimann, N. Reinsch, and G. Erhardt. 2009. Association between single
nucleotide polymorphisms in the CXCR1 gene and somatic cell score in Holstein dairy
cattle. J. Dairy Sci. 92:4018-4022.
Grimes, D. A., and K. F. Schulz. 2002. Bias and causal associations in observational research.
Lancet. 359:248-252.
Grobet, L., L. J. R. Martin, D. Poncelet, D. Pirottin, B. Brouwers, J. Riquet, A. Schoeberlein, S.
Dunner, F. Menissier, J. Massabanda, R. Fries, R. Hanset, and M. Georges. 1997. A
deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle.
Nat. Genet. 17:71-74.
Jones, S. A., M. Wolf, S. X. Qin, C. R. Mackay, and M. Baggiolini. 1996. Different functions for
the interleukin 8 receptors (IL-8R) of human neutrophil leukocytes: NADPH oxidase and
phospholipase D are activated through IL-8R1 but not IL-8R2. Proc. Natl. Acad. Sci. U. S.
A. 93:6682-6686.
Jones, S. A., B. Dewald, I. ClarkLewis, and M. Baggiolini. 1997. Chemokine antagonists that
discriminate between interleukin-8 receptors - Selective blockers of CXCR2. J. Biol Chem.
272:16166-16169.
Lahouassa, H., P. Rainard, A. Caraty, and C. Riollet. 2008. Identification and characterization
of a new interleukin-8 receptor in bovine species. Mol. Immunol. 45:1153-1164.
Lam, T. J., Y. H. Schukken, J. H. van Vliet, F. J. Grommers, M. J. Tielen, and A. Brand. 1997.
Effect of natural infection with minor pathogens on susceptibility to natural infection
with major pathogens in the bovine mammary gland. Am. J. Vet. Res. 58:17-22.
Larkin, M. A., G. Blackshields, N. P. Brown, R. Chenna, P. A. McGettigan, H. McWilliam, F.
Valentin, I. M. Wallace, A. Wilm, R. Lopez, J. D. Thompson, T. J. Gibson, and D. G. Higgins.
2007. Clustal W and clustal X version 2.0. Bioinformatics 23:2947-2948.

Chapter 7 General discussion
219
Leyva-Baca, I., F. Schenkel, J. Martin, and N. A. Karrow. 2008. Polymorphisms in the 5 '
upstream region of the CXCR1 chemokine receptor gene, and their association with
somatic cell score in Holstein cattle in Canada. J. Dairy Sci. 91:407-417.
Liou, J. W., F. T. Chang, Y. Chung, W. Y. Chen, W. B. Fischer, and H. J. Hsu. 2014. In silico
analysis reveals sequential interactions and protein conformational changes during the
binding of chemokine CXCL-8 to its receptor CXCR1. Plos One 9:e94178.
McDougall, S., K. E. Agnew, R. Cursons, X. X. Hou, and C. R. Compton. 2007. Parenteral
treatment of clinical mastitis with tylosin base or penethamate hydriodide in dairy cattle.
J. Dairy Sci. 90:779-789.
Mehrzad, J., L. Duchateau, and C. Burvenich. 2004. Viability of milk neutrophils and severity
of bovine coliform mastitis. J. Dairy Sci. 87:4150-4162.
Milk Control Centre Flanders. 2012. Annual Report. Milk Control Centre Flanders, Lier,
Belgium.
Mitchell, G. B., B. N. Albright, and J. L. Caswell. 2003. Effect of interleukin-8 and granulocyte
colony-stimulating factor on priming and activation of bovine neutrophils. Infect. Immun.
71:1643-1649.
Nasser, M. W., R. J. Marjoram, S. L. Brown, and R. M. Richardson. 2005. Cross-desensitization
among CXCR1, CXCR2, and CCR5: Role of protein kinase C-epsilon. J. Immunol. 174:6927-
6933.
National Mastitis Council. 2014. Recommended mastitis control program. NMC INC. ,
Madison, WI. http://www. nmconline. org/docs/NMCchecklistINT. pdf.
Olde Riekerink, R. G. M., H. W. Barkema, D. F. Kelton, and D. T. Scholl. 2008. Incidence rate
of clinical mastitis on Canadian dairy farms. J. Dairy Sci. 91:1366-1377.
Paape, M. J., D. D. Bannerman, X. Zhao, and J. W. Lee. 2003. The bovine neutrophil:
Structure and function in blood and milk. Vet. Res. 34:597-627.
Piepers, S., G. Opsomer, H. W. Barkema, A. de Kruif, and S. De Vliegher. 2010. Heifers
infected with coagulase-negative staphylococci in early lactation have fewer cases of
clinical mastitis and higher milk production in their first lactation than noninfected
heifers. J. Dairy Sci. 93:2014-2024.
Pighetti, G. M., C. J. Kojima, L. Wojakiewicz, and M. Rambeaud. 2012. The bovine CXCR1
gene is highly polymorphic. Vet. Immunol. Immunopathol. 145:464-470.

Chapter 7 General discussion
220
Rainard, P., and C. Riollet. 2006. Innate immunity of the bovine mammary gland. Vet. Res.
37:369-400.
Rambeaud, M., and G. M. Pighetti. 2005. Impaired neutrophil migration associated with
specific bovine CXCR2 genotypes. Infect. Immun. 73:4955-4959.
Rambeaud, M., R. Clift, and G. M. Pighetti. 2006. Association of a bovine CXCR2 gene
polymorphism with neutrophil survival and killing ability. Vet. Immunol. Immunopathol.
111:231-238.
Rambeaud, M., and G. M. Pighetti. 2007. Differential calcium signaling in dairy cows with
specific CXCR1 genotypes potentially related to interleukin-8 receptor functionality.
Immunogenetics 59:53-58.
Richardson, R. M., B. C. Pridgen, B. Haribabu, H. Ali, and R. Snyderman. 1998. Differential
cross-regulation of the human chemokine receptors CXCR1 and CXCR2 - Evidence for
time-dependent signal generation. J. Biol. Chem. 273:23830-23836.
Richardson, R. M., R. J. Marjoram, L. S. Barak, and R. Snyderman. 2003. Role of the
cytoplasmic tails of CXCR1 and CXCR2 in mediating leukocyte migration, activation, and
regulation. J. Immunol. 170:2904-2911.
Rupp, R., and D. Boichard. 2003. Genetics of resistance to mastitis in dairy cattle. Vet. Res.
34:671-688.
Schreiner, D. A., and P. L. Ruegg. 2003. Relationship between udder and leg hygiene scores
and subclinical mastitis. J. Dairy Sci. 86:3460-3465.
Schukken, Y. H., F. J. Grommers, D. Vandegeer, H. N. Erb, and A. Brand. 1990. Risk factors for
clinical mastitis in herds with a low bulk milk somatic cell count. 1. Data and risk factors
for all cases. J. Dairy Sci. 73:3463-3471.
Schukken, Y. H., F. J. Grommers, D. Vandegeer, H. N. Erb, and A. Brand. 1991. Risk factors for
clinical mastitis in herds with a low bulk milk somatic cell count. 2. Risk-Factors for
Escherichia coli and Staphylococcus aureus. J. Dairy Sci. 74:826-832.
Schukken, Y. H., J. Hertl, D. Bar, G. J. Bennett, R. N. Gonzalez, B. J. Rauch, C. Santisteban, H. F.
Schulte, L. Tauer, F. L. Welcome, and Y. T. Grohn. 2009. Effects of repeated Gram-positive
and Gram-negative clinical mastitis episodes on milk yield loss in Holstein dairy cows. J.
Dairy Sci. 92:3091-3105.
Schukken, Y. H., D. J. Wilson, F. Welcome, L. Garrison-Tikofsky, and R. N. Gonzalez. 2003.
Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 34:579-596.

Chapter 7 General discussion
221
Schukken, Y. H., J. Gunther, J. Fitzpatrick, M. C. Fontaine, L. Goetze, O. Holst, J. Leigh, W.
Petzl, H. J. Schuberth, A. Sipka, D. G. E. Smith, R. Quesnell, J. Watts, R. Yancey, H. Zerbe,
A. Gurjar, R. N. Zadoks, and H. M. Seyfert. 2011. Host-response patterns of
intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 144:270-289.
Singh, J. K., G. Farnie, N. J. Bundred, B. M. Simoes, A. Shergill, G. Landberg, S. J. Howell, and
R. B. Clarke. 2013. Targeting CXCR1/2 significantly reduces breast cancer stem cell
activity and increases the efficacy of inhibiting HER2 via HER2-dependent and -
independent mechanisms. Clin. Cancer Res. 19:643-656.
Sodeland, M., M. P. Kent, H. G. Olsen, M. A. Opsal, M. Svendsen, E. Sehested, B. J. Hayes, and
S. Lien. 2011. Quantitative trait loci for clinical mastitis on chromosomes 2, 6, 14 and 20
in Norwegian Red cattle. Anim. Genet. 42:457-465.
Stevens, M. G. H., L. J. Peelman, B. De Spiegeleer, A. Pezeshki, G. R. Van De Walle, L.
Duchateau, and C. Burvenich. 2011. Differential gene expression of the toll-like receptor-
4 cascade and neutrophil function in early- and mid-lactating dairy cows. J. Dairy Sci.
94:1277-1288.
Supré, K., F. Haesebrouck, R. N. Zadoks, M. Vaneechoutte, S. Piepers, and S. De Vliegher.
2011. Some coagulase-negative Staphylococcus species affect udder health more than
others. J. Dairy Sci. 94:2329-2340.
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe, and F.
Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by
geometric averaging of multiple internal control genes. Genome Biol. 3.
Van Oostveldt, K., F. Vangroenweghe, H. Dosogne, and C. Burvenich. 2001. Apoptosis and
necrosis of blood and milk polymorphonuclear leukocytes in early and midlactating
healthy cows. Vet. Res. 32:617-622.
Van Oostveldt, K., M. J. Paape, H. Dosogne, and C. Burvenich. 2002. Effect of apoptosis on
phagocytosis, respiratory burst and CD18 adhesion receptor expression of bovine
neutrophils. Domest. Anim. Endocrinol. 22:37-50.
Wolff, C., M. Espetvedt, A. K. Lind, S. Rintakoski, A. Egenvall, A. Lindberg, and U. Emanuelson.
2012. Completeness of the disease recording systems for dairy cows in Denmark,
Finland, Norway and Sweden with special reference to clinical mastitis. BMC Vet. Res.
8:131.

Chapter 7 General discussion
222
Wozniak, A., W. H. Betts, G. A. Murphy, and M. Rokicinski. 1993. Interleukin-8 primes human
neutrophils for enhanced superoxide anion production. Immunology 79:608-615.
Youngerman, S. M., A. M. Saxton, S. P. Oliver, and G. M. Pighetti. 2004. Association of CXCR2
polymorphisms with subclinical and clinical mastitis in dairy cattle. J. Dairy Sci. 87:2442-
2448.

223
Summary

Summary
224

Summary
225
Mastitis impairs milk quality and production, and increases the culling probability of
dairy cows substantially. Furthermore, clinical mastitis affects animal welfare. Mastitis
resistance differs between animals and is partly genetically determined. Although many
genes are involved in mammary gland immunity, polymorphisms in genes encoding proteins
with an important role in the immune response might alter mastitis resistance of cows
significantly. Neutrophils play a major role in the mammary gland immunity. Binding of
interleukin-8 (IL-8) on chemokine (C-X-C motif) receptor 1 (CXCR1) has a pleiotropic effect, as
it enhances activity, causes migration from blood to the site of infection and delays
apoptosis of neutrophils. Despite its important function, many single nucleotide
polymorphisms (SNP) were detected in the coding region of the bovine CXCR1 gene
potentially altering receptor functionality.
Chapter 1 provides a review of literature on bovine mastitis, mammary gland immunity
and the genetics of mastitis resistance. Additionally, an overview is given on previous
research on CXCR1 polymorphisms and its association with mastitis resistance. The main aim
of this thesis was to gain more insight in the role of CXCR1 polymorphisms for mammary
gland immunity and udder health (Chapter 2). Associations between CXCR1 polymorphisms
and neutrophil characteristics (Chapter 3), subclinical mastitis (Chapter 4) and clinical
mastitis (Chapter 5) were analyzed.
Before analyzing associations, the entire coding region of the CXCR1 gene was
screened for polymorphism by DNA sequencing of 140 Flemish Holstein heifers originating
from 20 commercial dairy farms (Chapter 4.1). In total, 16 polymorphisms were detected of
which 8 cause a change in amino acid sequence and 6 were not described before.
In Chapter 3.1, associations between the CXCR1 genotype (whole coding region) and
apoptosis, necrosis, and concentration of milk neutrophils of 292 quarters belonging to 73
early lactating dairy heifers were studied. Associations between CXCR1 SNPs and milk
neutrophil viability and concentration depended on the intramammary infection (IMI) status.
In uninfected quarters, % milk neutrophil apoptosis was higher in heifers with genotype
c.980GG compared to heifers with genotype c.980AG, whereas in infected quarters, % milk
neutrophil apoptosis was higher in heifers with genotype c.642GG compared to heifers with
genotype c.642AG. Differences in milk neutrophil concentration between infected and

Summary
226
uninfected quarters were smaller in c.980AG heifers than in c.980GG heifers. An association
between the CXCR1 genotype and necrosis of milk neutrophils could not be demonstrated.
Additional to viability, activity of neutrophils might associate with CXCR1
polymorphisms. To test this hypothesis, blood neutrophils were isolated from twenty early
lactating heifers with a different CXCR1 genotype at position 735 or 980, incubated with
different concentrations of recombinant bovine IL-8 (rbIL-8) for 2 or 6 h and stimulated with
phorbol 12-myristate 13-acetate (PMA) or opsonized zymosan particles (OZP) (Chapter 3.2).
Associations between CXCR1 genotype and the reactive oxygen species (ROS) generation,
measured by luminol chemiluminescence, were analyzed. Single nucleotide polymorphisms
c.735C>G and c.980A>G were neither associated with ROS generation in general nor with the
effect of rbIL-8 incubation on ROS generation. However, neutrophils incubated with rbIL-8
for 2 or 6 h showed higher total PMA induced ROS generation but lower total OZP induced
ROS generation compared to neutrophils incubated without rbIL-8. Against expectations,
inhibitory effects of IL-8 on ROS generation in bovine neutrophils were observed.
In Chapter 4.1, associations between CXCR1 polymorphisms and subclinical heifer
mastitis of 140 heifers belonging to 20 commercial dairy herds were analyzed. Phenotypic
data included quarter bacteriological culture results and somatic cell count (SCC) in early
lactation and composite milk SCC during first lactation. Polymorphism c.980A>G was
associated with pathogen-group specific IMI: heifers with genotype AG were less likely to
have an IMI due to major mastitis pathogens compared to heifers with genotype GG but did
not have less IMI by coagulase-negative staphylococci, so-called minor pathogens. CXCR1
genotype was neither associated with quarter SCC in early lactation nor with composite SCC
during lactation.
To further study the influence of SNP c.980A>G on mammary gland immunity, mammary
glands of 4 c.980AG and 4 c.980GG Holstein heifers were experimentally challenged with a
Staphylococcus chromogenes isolate originating from a chronic IMI (Chapter 4.2). Quarters
from heifers with genotype c.980AG and c.980GG both became subclinically infected but
showed differences in the early stage of infection. Bacterial count at 18 h post challenge
tended to be higher in quarters from c.980AG heifers compared to c.980GG heifers. Somatic
cell count (SCC) was lower at 6 h post challenge and tended to be lower at 9 h post challenge
in c.980AG heifers compared to c.980GG heifers. Yet, milk production decreased similarly.

Summary
227
Milk neutrophils of c.980AG heifers showed more apoptosis at 9 h post challenge and
tended to show more necrosis at 6, 9 and 12 h post challenge than c.980GG heifers.
Differences were less pronounced in the later stage (> 18 h) of infection. Results
demonstrate that CXCR1 polymorphism can influence SCC and milk neutrophil viability
following experimental IMI.
Unlike other regions, little information on clinical mastitis was available in Flanders.
Before testing associations between clinical mastitis and CXCR1 genotype, pathogen-specific
incidence rate of clinical mastitis (IRCM) was estimated on 50 randomly selected commercial
Flemish dairy herds (Chapter 5.1). Additionally, the severity of the cases and the potential
associations with herd hygiene were studied. During a one-year survey, participating
producers sampled 845 cases and 692 dairy cows. The mean and median IRCM was
estimated at 7.4 and 5.3 quarter cases per 10,000 cow-days at risk, respectively. A large
between-herd variation was observed. In general, the IRCM was lower in heifers compared
to multiparous cows. However, the overall IRCM in the first week after calving was higher in
heifers compared to cows. Streptococcus uberis (18.2% of the cases) and Escherichia coli
(15.5%) were the most frequently isolated pathogens and no growth was observed in 19.9%
of the cases. The majority of the cases (63.1%) were mild (only clots in milk). Moderate (hard
quarter without general signs) and severe symptoms (systemic illness) were observed in
29.9%, and 7.0% of the cases, respectively. Isolation of E. coli (vs. any other culture result)
was more likely in moderate and severe cases compared to mild cases. Overall IRCM and E.
coli IRCM were higher in dirty compared to clean herds based on udder hygiene scores.
Results broaden the knowledge on clinical mastitis in Flemish dairy herds and underlines the
high risk of clinical mastitis in early lactating heifers, the role of the so-called environmental
pathogens, and herd hygiene.
In total, 3,106 cows from the abovementioned survey were genotyped (Chapter 5.2). A
fluorescent multiprobe PCR assay was designed specifically and validated to genotype SNPs
c.735C>G and c.980A>G simultaneously on a reliable and cost-effective manner. Associations
between genotype and pathogen-specific IRCM, test-day somatic cell count (SCC) and test-
day milk yield were analyzed. Monthly SCC and milk yield records were available for a subset
of 1,961 cows of 32 herds participating in the DHI (dairy herd improment) program. Cows
with genotype c.735GG had lower S. aureus IRCM compared to cows with genotype c.735CC.

Summary
228
Additionally, a parity-specific association between S. aureus IRCM and SNP c.980A>G was
detected. Heifers with genotype c.980GG had a lower S. aureus IRCM compared to heifers
with genotype c.980AG. Differences were less pronounced in multiparous cows. Associations
between CXCR1 genotype and SCC were not detected. However, milk yield was associated
with SNP c.735C>G. Cows with genotype c.735GG out-produced cows with genotype
c.735CC by 0.8 kg milk/day.
RNA was isolated from milk somatic cells one hour before and 12 hours after the
experimental IMI described in Chapter 4.2. Expression of CXCR1 and eight candidate
reference genes (ACTB, B2M, H2A, HPRT1, RPS15A, SDHA, UBC and YWHAZ) was measured
by reverse transcription quantitative real-time PCR (RT-qPCR). Differences in relative CXCR1
expression between c.980AG heifers and c.980GG heifers were studied and the effect of the
experimental IMI on relative expression of CXCR1 and the candidate reference genes was
analyzed (Chapter 6). Relative expression of CXCR1 was not associated with polymorphism
c.980A>G but was significantly upregulated following the experimental intramammary
infection. Additionally, differential expression was detected for B2M, H2A, HPRT1, SDHA and
YWHAZ. This study reinforces the importance of CXCR1 in mammary gland immunity and
demonstrates the potential effect of experimental IMI on expression of housekeeping genes
in milk somatic cells.
In Chapter 7, results of the abovementioned studies are compared to each other and to
results of studies performed by other research groups. The role of CXCR1 polymorphisms on
mastitis resistance is discussed and suggestions for marker assisted selection and future
research are made. Based on the many associations we concluded CXCR1 to be an important
gene for mastitis resistance (Figure 8.1).

Sum
mar
y
229
Fi
gure
8.1
. Fro
m g
enot
ype
to p
heno
type
: sum
mar
y of
the
resu
lts. 1 sin
gle
nucl
eotid
e po
lym
orph
isms;
2 phor
bol 1
2-m
yrist
ate
13-a
ceta
te a
nd o
pson
ized
zym
osan
par
ticle
in
duce
d re
activ
e ox
ygen
spec
ies g
ener
atio
n.

230

231
Samenvatting

Samenvatting
232

Samenvatting
233
Mastitis heeft een negatieve invloed op de melkkwaliteit en -productie en verhoogt de
kans dat een koe opgeruimd wordt aanzienlijk. Bovendien schaadt klinische mastitis het
dierenwelzijn. De weerstand tegen mastitis verschilt tussen dieren en is deels genetisch
bepaald. Vele genen zijn betrokken bij de immuniteit van de melkklier. Desalnietemin spelen
polymorfismen in genen coderend voor eiwitten met een belangrijke functie in de
immuunrespons mogelijks een grote rol. Neutrofielen zijn cruciaal voor de immuniteit van de
melkklier. De binding van interleukine 8 (IL-8) op de chemokine (C-X-C motief) receptor 1
(CXCR1) heeft een pleiotroop effect op neutrofielen. Het verhoogt hun activiteit, veroorzaakt
migratie van bloed naar de plaats van infectie en vertraagt de apoptose. Ondanks de
belangrijke functie werden veel “single nucleotide polymorphisms” (SNPs) gedetecteerd in
het coderend gedeelte van het boviene CXCR1 gen die mogelijks de receptor functionaliteit
beïnvloeden.
Hoofdstuk 1 vat de literatuur rond boviene mastitis, immuniteit van de melkklier en de
genetica van mastitisresistentie samen. Bijkomend wordt een overzicht gegeven van het
reeds uitgevoerde onderzoek naar CXCR1 polymorfismen en mastitis resistentie. Het
hoofddoel van deze thesis is om meer inzicht verkrijgen in de rol van CXCR1 polymorfismen
op de immuniteit van de melkklier en de uiergezondheid (Hoofdstuk 2). De eventuele
associaties tussen CXCR1 polymorfismen en neutrofiel eigenschappen (Hoofdstuk 3),
subklinische mastitis (Hoofdstuk 4) en KM (Hoofdstuk 5) werden bestudeerd.
Voorafgaand aan de associatiestudies werd het volledig coderend gedeelte van het
CXCR1 gen naar SNPs door middel van DNA sequenering van 140 Vlaamse Holstein vaarzen
afkomstig van 20 commerciële melkveebedrijven gescreend (Hoofdstuk 4.1). Zestien
polymorfismen waarvan er 8 een wijziging in de aminozuur sequentie veroorzaken en 6
voordien niet werden beschreven konden worden gedetecteerd.
In Hoofdstuk 3.1 onderzochten we associaties tussen het CXCR1 genotype (volledig
coderend gedeelte) en apoptose, necrose en de concentratie van melk neutrofielen
geïsoleerd uit 292 kwartieren van 73 vroeg lacterende melkveevaarzen. De associatie tussen
CXCR1 SNPs en melk neutrofiel viabiliteit en concentratie was afhankelijk van de
intramammaire infectie (IMI) status. Niet-geïnfecteerde kwartieren hadden een hoger %
melk neutrofiel apoptose in vaarzen met genotype c.980GG dan in vaarzen met genotype
c.980AG. Geïnfecteerde kwartieren hadden een hoger % melk neutrofiel apoptose in
c.642GG vaarzen dan in c.642AG vaarzen. Het verschil in melk neutrofiel concentratie tussen

Samenvatting
234
geïnfecteerde en niet-geïnfecteerde was kleiner bij c.980AG vaarzen dan bij c.980GG
vaarzen. We konden geen associatie tussen het CXCR1 genotype en melk neutrofiel necrose
aantonen.
Naast viabiliteit beïnvloeden CXCR1 polymorfismen mogelijks ook de activiteit van
neutrofielen. Om deze hypothese te testen werden bloed neutrofielen geïsoleerd uit 20
vroeg lacterende vaarzen met verschillend CXCR1 genotype op positie 735 of 980,
geïncubeerd met verschillende concentraties recombinant bovien IL-8 (rbIL-8) en
gestimuleerd met forbol 12-myristaat 13-acetaat (PMA) of geopsoniseerde zymosan
partikels (OZP) (Hoofdstuk 3.2). Vervolgens gingen we eventuele associates tussen CXCR1 en
“reactive oxygen species” (ROS) generatie gemeten door middel van luminol
chemieluminescentie na. We vonden geen verbanden tussen CXCR1 genotype en ROS
generatie in het algemeen of het effect van rbIL-8 incubatie op ROS generatie. Neutrofielen
geïncubeerd met rbIL-8 gedurende 2 of 6 u vertoonden een hogere PMA geïnduceerde ROS
generatie maar een lagere OZP geïnduceerde ROS generatie dan neutrofielen geïncubeerd
zonder rbIL-8. Tegen onze verwachtingen in observeerden we inhiberende effecten van IL-8
op ROS generatie in boviene neutrofielen.
In Hoofdstuk 4.1 werden mogelijke associaties tussen CXCR1 polymorfismen en
subklinische vaarzenmastitis bij 140 vaarzen van 20 commerciële melkveebedrijven
geanalyseerd. In de vroege lactatie werden kwartiermelkstalen bacteriologisch onderzocht
en werd het kwartiercelgetal bepaald. Gedurende de eerste lactatie waren maandelijkse
koecelgetal gegevens beschikbaar. Polymorfisme c.980A>G kon geassocieerd worden met
pathogeen-groep specifieke IMI status: c.980GG vaarzen hadden een hogere kans op IMI
door major pathogenen dan c.980AG vaarzen maar hadden niet minder IMI door coagulase-
negatieve stafylokokken, zogenaamde minor pathogenen. Er werden geen verbanden
gevonden tussen het CXCR1 genotype en het kwartiercelgetal tijdens de vroege lactatie en
het koecelgetal gedurende lactatie.
Om de invloed van SNP c.980A>G op de immuniteit van de melkklier verder te
onderzoeken infecteerden we de melkklieren van 4 c.980AG en 4 c.980GG Holstein vaarzen
met een Staphylococcus chromogenes stam geïsoleerd uit een chronische IMI (Hoofdstuk
4.2). In de vroege fase van de subklinische infectie werden verschillen tussen de
genotypegroepen opgemerkt. Zestien uur na inoculatie was het kiemgetal hoger bij c.980AG
dan bij c.980GG vaarzen. Zes en negen uur na inoculatie was het celgetal lager bij c.980AG

Samenvatting
235
vaarzen dan bij c.980GG vaarzen. De melkproductie daalde op vergelijkbare wijze. Melk
neutrofielen van c.980AG vaarzen vertoonden daarentegen meer apoptose 9 uur na
inoculatie en meer necrose 6, 9 en 12 uur na inoculatie dan melk neutrofielen van c.980GG
vaarzen. De verschillen waren minder uitgesproken in de late fase (> 18 u) van infectie. Dit
experiment toonde aan dat CXCR1 polymorfisme het celgetal en de melkneutrofielviabiliteit
na experimentele infectie kan beïnvloeden.
In tegenstelling tot andere regio’s zijn er in Vlaanderen weinig gegevens rond klinische
mastitis beschikbaar. Vooraleer associaties tussen klinische mastitis en CXCR1 genotype na
te gaan hebben we de pathogeen-specifieke incidentie ratio van klinische mastitis (IRKM) op
50 willekeurig geselecteerde commerciële Vlaamse melkveebedrijven geschat (Hoofdstuk
5.1). Bijkomend bestudeerden we de ernst van de gevallen en mogelijke verbanden tussen
de bedrijfshygiëne en de IRKM. Gedurende een jaar bemonsterden de deelnemende
melkveehouders 845 gevallen bij 692 melkkoeien. Het gemiddelde en mediaan IRKM werd
geschat op respectievelijk 7.4 en 5.3 kwartiergevallen per 10,000 dagen “at risk”. We
observeerden een grote bedrijfsvariatie. Vaarzen hadden over het algemeen een lagere
IRKM dan koeien maar een hogere IRKM in de eerste week na kalven. Streptococcus uberis
(18.2% van de gevallen) en Escherichia coli (15.5%) waren de vaakst geïsoleerde pathogenen.
19.9% van de gevallen vertoonde geen groei. De meerderheid van de gevallen (63.1%)
vertoonde een mild verloop (enkel vlokken in de melk). Respectievelijk 29.9% en 7% van de
gevallen vertoonden matige (hard kwartier zonder algemene symptomen) en ernstige
symptomen (systemische ziekte). Isolatie van E. coli (t.o.v. andere bacteriologische uitslagen)
was meer waarschijnlijk in matige en ernstige gevallen dan in milde gevallen. Zowel
algemene als E. coli IRKM waren hoger op vuile dan op propere bedrijven gebaseerd op
uierhygiëne scores. De resultaten verruimen onze kennis rond klinische mastitis op Vlaamse
melkveebedrijven en onderstrepen het hoge risico op klinische mastitis bij vroeg lacterende
vaarzen, de rol van zogenaamde omgevingskiemen en het belang van bedrijfshygiëne.
Vervolgens werd het genotype van 3,106 koeien van de bovengenoemde survey
nagegaan (Hoofdstuk 5.2). Om het genotype op SNPs c.735C>G en C.980A>G simultaan te
bepalen op een betrouwbare en kosteneffectieve wijze ontwikkelden en valideerden we een
fluorescerende multiprobe PCR assay. Associaties tussen genotype en pathogeen-specifieke
IRKM, celgetal en melkproductie werden geanalyseerd. Het celgetal en de melkproductie
van 1,961 koeien op 32 van de 50 bedrijven werden maandelijks bepaald als onderdeel van

Samenvatting
236
de melkproductieregistratie (MPR). Koeien met genotype c.735GG hadden een lagere
Staphylococcus aureus IRKM dan koeien met genotype c.735CC. Bijkomend detecteerden we
een pariteit-specifieke associatie tussen S. aureus IRKM en SNP c.980A>G. Vaarzen met
genotype c.980GG hadden een lagere S. aureus IRKM dan vaarzen met genotype c.980AG.
De verschillen waren minder uitgesproken bij multipare koeien. Het CXCR1 genotype kon
geassocieerd worden met de melkproductie maar niet met het celgetal. Koeien met
genotype c.735GG produceerden gemiddeld 0.8 kg melk meer per dag dan koeien met
genotype c.735CC.
RNA werd geïsoleerd uit melk somatische cellen een uur voor en 12 uur na de
experimentele IMI beschreven in Hoofdstuk 4.2. Expressie van CXCR1 en 8 vaak gebruikte
referentiegenen (ACTB, B2M, H2A, HPRT1, RPS15A, SDHA, UBC en YWHAZ) werd opgemeten
aan de hand van “reverse transcription quantitative real-time PCR (RT-qPCR)”. Verschillen in
CXCR1 expressie tussen c.980AG vaarzen en c.980GG vaarzen werden bestudeerd en de
invloed van de experimentele IMI op expressie van CXCR1 en de kandidaat referentiegenen
werd geanalyzeerd. Expressie van CXCR1 was niet geassocieerd met polymorfisme c.980A>G.
De experimentele IMI had een invloed op expressie van ACTB, B2M, CXCR1, HPRT1 en
YWHAZ. Deze studie bevestigt het belang van CXCR1 in de afweer van de melkklier en toont
aan dat experimentele IMI de expressie van kandidaat referentiegenen in melk somatische
cellen kan beïnvloeden.
In Hoofdstuk 7 vergeleken we de resultaten van de bovengenoemde studies met elkaar
en met resultaten van studies uitgevoerd door andere onderzoeksgroepen. De rol van CXCR1
polymorfismen bij mastitisresistentie en suggesties voor merker geassisteerde selectie en
toekomstig onderzoek werden bediscussieerd. Op basis van de vele gevonden associaties
concludeerden we dat CXCR1 een belangrijk gen is voor mastitis resistentie (Figure 8.1).

Sam
enva
ttin
g
237
Fi
guur
1. V
an g
enot
ype
tot
feno
type
: sam
enva
ttin
g va
n de
res
ulta
ten.
1 ”sin
gle
nucl
eotid
e po
lym
orph
isms”
; 2 forb
ol 1
2-m
yrist
aat
13-a
ceta
at a
nd g
eops
onize
erde
zym
osan
part
ikel
s geï
nduc
eerd
e “r
eact
ive
oxyg
en sp
ecie
s” g
ener
atie
.

238

239
Curriculum vitae – Publications

Curriculum vitae - Publications
240

Curriculum vitae - Publications
241
Curriculum vitae
Joren Verbeke werd geboren op 8 december 1986 te Brugge. In 2004 behaalde hij het
diploma hoger secundair onderwijs aan het Sint-Lodewijks college te Brugge (richting
Wetenschappen Wiskunde) en vatte hij de studie Diergeneeskunde te Gent aan. Na
succesvolle onderzoekstages aan de Universiteit van Jimma (Ethiopië) en de Universiteit
Gent behaalde hij in 2010 het diploma van dierenarts met onderscheiding. Zijn masterproef
“Polymorphism screening and association study of CXCR1 (IL8RA) with udder health and milk
production of dairy heifers” (promotor Prof. dr. Sarne De Vliegher, medepromotor Prof. dr.
Luc Peelman) werd bekroond met de prijs “beste masterproef in de optie Onderzoek”.
In september 2010 begon hij een doctoraatstudie getiteld “Verschillen tussen coagulase-
negatieve Staphylococcus soorten en hun interactie met de koe”. Na het behalen van een
specialisatiebeurs gefinancierd door het agentschap voor Innovatie door Wetenschap en
Technologie (IWT) in januari 2011 wijzigde het onderwerp naar “Het boviene CXCR1 gen,
neutrofielen en uiergezondheid” met als promotoren Prof. dr. Sarne De Vliegher (Vakgroep
Voortplanting, Verloskunde en Bedrijfsdiergeneeskunde) en Prof. dr. Luc Peelman (Vakgroep
Voeding, Genetica en Ethologie). In het kader van dit onderzoek publiceerde hij meerdere
internationale wetenschappelijke artikelen en gaf hij presentaties op verschillende
internationale congressen.
Als lid van het t MMM--team voerde hij bacteriologisch onderzoek op melkstalen uit, gaf hij amamma vov
lezingen op studiedagen, schreef hij artikelen in landbouwtijdschriften rond uiergezondheid
en begeleide hij masterproefstudenten. In samenwerking met het Melkcontrolecentrum
Vlaanderen en Dierengezondheidszorg Vlaanderen coördineerde hij het Veepeiler
deelproject “Klinische mastitis op Vlaamse melkveebedrijven”. Bijkomend was Joren actief
betrokken bij de doctoraatstudie van Dr. Tadele Tolosa binnen het samenwerkingsproject
tussen Jimma University en de Vlaamse universiteiten (IUC-JU).

Curriculum vitae - Publications
242
Publications in international journals
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Pathogen-
group specific association between CXCR1 polymorphisms and subclinical mastitis in
dairy heifers. J. Dairy Res. 79:341-351.
Tolosa, T., J. Verbeke, S. Piepers S, K. Supré, and S. De Vliegher. 2013. Risk factors associated
with subclinical mastitis as detected by California Mastitis Test in smallholder dairy farms
in Jimma, Ethiopia using multilevel modelling. Prev. Vet. Med. 112:68-75.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2014. Association of
CXCR1 polymorphisms with apoptosis, necrosis and concentration of milk neutrophils in
early lactating dairy heifers. Res. Vet. Sci. 97:55-59.
Verbeke, J., S. Piepers, K. Supré, and S. De Vliegher. 2014. Pathogen-specific incidence rate
of clinical mastitis in Flemish dairy herds, severity, and association with herd hygiene. J.
Dairy Sci. 97:6926-6934.
Verbeke, J., M. Van Poucke, L. Peelman, S. Piepers, and S. De Vliegher. Associations between
CXCR1 polymorphisms and pathogen-specific incidence rate of clinical mastitis, test-day
somatic cell count, and test-day milk yield. J. Dairy Sci. 97:7927-7939.
Verbeke, J., K. Piccart, S. Piepers, L. Peelman, M. Van Poucke, A. De Visscher and S. De
Vliegher. Somatic cell count and milk neutrophil viability of dairy heifers with specific
CXCR1 genotypes following experimental intramammary infection with Staphylococcus
chromogenes originating from milk. Vet. J.: Revisions.
Verbeke, J., M. Van Poucke, L. Peelman, and S. De Vliegher. Differential expression of CXCR1
and commonly used reference genes in bovine milk somatic cells following experimental
intramammary infection. Submitted.
Verbeke, J., X. Boulougouris, C. Rogiers, C. Burvenich, L. Peelman, B. De Spiegeleer, and S. De
Vliegher. Reactive oxygen species generation of bovine blood neutrophils with different
CXCR1 (IL8RA) genotype following interleukin-8 incubation. Submitted.
Tolosa, T., J. Verbeke, Z. Ayana, S. Piepers, K. Supré, and S. De Vliegher. 2013. Pathogen
group specific risk factors for clinical mastitis, subclinical mastitis and blind quarters at
the herd, cow and quarter level in smallholder dairy farms in Jimma, Ethiopia. Submitted.
Tolosa, T., J. Verbeke, S. Piepers, M. Teferaa, Y. Getachewa, K. Supré, and S. De Vliegher.
Dairy farming and milk quality in Jimma, Ethiopia. In preparation.

Curriculum vitae - Publications
243
Oral presentations and contributions
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2011. Association
between CXCR1 gene polymorphisms and udder health in dairy heifers. 6th European
congress of bovine health management: abstracts, 7-9 September, Liège, Belgium p. 56.
Deberdt, K., S. Piepers, R. March, R. Guix, A. De Visscher, J. Verbeke, and S. De Vliegher.
2011. Immunological response to an experimental intramammary inocculation with a
killed Staphylococcus aureus strain in vaccinated and non-vaccinated lactating dairy
cows. 6th European congress of bovine health management: abstracts, 7-9 September,
Liège, Belgium p. 59.
Piepers, S., K. Deberdt, A. De Visscher, J. Verbeke, and S. De Vliegher. 2011. Immunological
response to an experimental intramammary inocculation with a killed Staphylococcus
aureus strain in vaccinated and non-vaccinated lactating dairy cows. 3rd International
symposium on Mastitis and Milk Quality: abstracts, 22-24 September, St. Louis, Missouri.
Deberdt, K., S. Piepers, A. Prenafeta, R. March, A. Foix, R. Guix, A. De Visscher, J. Verbeke,
and S. De Vliegher. 2011. Immunological response to an experimental intramammary
inocculation with a killed Staphylococcus aureus strain in vaccinated and non-vaccinated
lactating dairy cows. Proceedings of the international conference on Udder Health and
Communication, 25-27 October, Utrecht, The Netherlands p. 353-358.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2011. Polymorphism
screening and association study of CXCR1 with udder health in dairy heifers. Proceedings
of the international conference on Udder Health and Communication, 25-27 October,
Utrecht, The Netherlands p. 367-372.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2011. Association of
CXCR1 gene polymorphism with apoptosis of blood and milk neutrophils in early lactating
dairy heifers. Physiology and Genomics of Mastitis: abstracts, 31 October-2 November,
Tutzing, Germany p. 36.
Verbeke, J., S. Piepers, L. Peelman, M. Van Poucke, and S. De Vliegher. 2012. Association
between CXCR1 gene polymorphisms and udder health in dairy heifers. National Mastitis
Council 51st Annual Meeting: abstracts, 22-24 January, St Pete Beach, Florida, p. 157-158.

Curriculum vitae - Publications
244
Piccart, K., S. Piepers, J. Verbeke, N. Melo De Sousa, J. Beckers, and S. De Vliegher. 2014.
Milk prolactin response after experimental infection with different coagulase-negative
staphylococci in dairy heifers. 2014 Joint Annual Meeting, 20-24 July, Kansas City,
Missouri, Paper 413.
Verbeke J., K. Piccart, S. Piepers, M. Van Poucke, L. Peelman, and S. De Vliegher. 2014.
Experimental intramammary challenge with Staphylococcus chromogenes in dairy heifers
with specific CXCR1 genotypes. 2014 Joint Annual Meeting, 20-24 July, Kansas City,
Missouri, Paper 175.
Verbeke J., M. Van Poucke, L. Peelman, S. Piepers, and S. De Vliegher. 2014. Association of
CXCR1 gene polymorphisms with incidence rate of clinical mastitis, somatic cell count
and milk production in dairy cattle. 2014 Joint Annual Meeting, 20-24 July, Kansas City,
Missouri, Paper 176.
Poster presentation
Verbeke J., M. Van Poucke, L. Peelman, S. Piepers, and S. De Vliegher. 2014. Association of
CXCR1 gene polymorphisms with incidence rate of clinical mastitis, and test-day somatic
cell count and milk production. 2014 National Mastitis Council Regional Meeting, 4-6
August, Ghent, Belgium

245
Dankwoord – Acknowledgements

Dankwoord - Acknowledgements
246
A problem is a chance to do your best. Tijdens mijn doctoraat heb ik heel wat kansen
gekregen om mijn best te kunnen doen. Gelukkig kon ik telkens rekenen op de steun van
mensen rondom mij.
Als eerste wens ik mijn promotor te danken. Beste Sarne, ondanks je drukke agenda
kon ik steeds bij jou terecht met al mijn vragen. Iedere letter en cijfer binnen dit doctoraat
las je na. Dankzij jouw hulp en motivatie leerde ik studies opzetten, data analyseren,
artikelen schrijven en resultaten presenteren. Bedankt om een geweldige promotor voor mij
te zijn! Het M-team zou het M-team niet zijn zonder Sofie Piepers. Beste Sofie, jouw kritische
geest stimuleerde mij om het beste uit mezelf te halen. Jouw passie voor uiergezondheid
kent geen gelijke. Zonder jou waren chapter 3.1 en 4.1 nooit tot stand gekomen. Bedankt
voor al jouw hulp tijdens mijn doctoraat. Ik wens je veel succes bij het oprichten van de spin-
off. Niet enkel mastitis experts maar ook twee rasechte genetici maakten deel uit van mijn
begeleidingscommissie: Luc en Mario. Beste Luc, jouw lessen tijdens de derde bachelor
wekten mijn interesse in de genetica op. Bedankt voor alle vertrouwen en advies die je mij
schonk als copromotor van mijn doctoraat. Beste Mario, jij leerde mij de kneepjes van het
genetische vak. Dankzij jouw hulp kon ik het CXCR1 gen van duizenden koeien bestuderen.
Ook als het niet zo vlot liep in het labo kon ik bij jou terecht. Je bent een geweldige labo-
coördinator.
Geen doctoraat zonder examencommissie. Bedankt professor de Kruif om deze
commissie voor te zitten. Jaren geleden vertelde je mij dat je bij een presentatie maar een
van de twee volgende groepen kan boeien, ofwel de 10% experts ofwel de overige 90% van
het publiek. Ik heb vaak teruggedacht aan deze raad en hoop dat ik vandaag slechts 10% van
het publiek verveeld heb. Bedankt prof. Burvenich, prof. Cox en prof. Van Immerseel voor de
vele waardevolle opmerkingen. Merci prof. Detilleux pour tous vos conseils. Votre article
“Genetic factors affecting susceptibility to udder pathogens” m’a aidé beaucoup pendant
mon doctorat. Dear prof. Kühn, thank you for being a member of the examination
committee. As an editor of JDS, you have read many of my manuscripts. Although not all
were accepted, you have always send me very valuable comments. Binnen de universiteit
raken we soms wat gevangen in theoretische gedachtegangen. Bedankt Erwin om mij na te
doen nadenken over hoe we de resultaten kunnen valoriseren en toepassen in de praktijk.
In de zomer van 2010 was ik niet het enige nieuwe lid van het M-team. Ik had de eer
en het genoegen vergezeld te worden door twee knappe, verstandige blondines: Kathelijne

Dankwoord - Acknowledgements
247
en Anneleen. De eerste dagen waren wat onwennig maar ook de bureaus naast de onze
zullen snel gemerkt hebben dat we het goed met elkaar konden vinden. Ik wens jullie veel
succes in alles wat jullie ondernemen! Het jaar na Kathelijne en Anneleen nam Sarne de
minder blonde, maar daarom niet minder knappe en verstandige Marina aan. Een brok
Limburgse energie raasde door onze vakgroep. Respect voor het vele werk dat je al verzette,
Marina. Veel succes bij het verwerken van alle gegevens! In dezelfde zomer vervoegde ook
Dimitri onze rangen. Zowel in Chicago als in Gent was het plezant om samen met “den Dimi”
een biertje te drinken. Ik wens je veel succes bij het vervolg van je onderzoek en je job bij de
Orde maar bovenal veel plezier samen met Lynn en de kleine Alina. Tot mijn grote vreugde
kon ik mijn Hasseltse vriendin Kristine overhalen om bij ons te werken. Klein is haar gestalte
maar groot haar gevoel voor humor. Ik dank ook Reshat, Pieter, Els, Steven en Lars voor de
vele fijne momenten. Hoewel hun verblijf beperkt was tot enkele maanden heb ik mij goed
geamuseerd met Wannes V. en Bert V. Ik hoop deze lente opnieuw mee te kunnen schapen
scheren Bert! Met de komst van de “moderne dames” (uitleg vraag je best aan Sarne)
Zyncke, Justine en Ameline is de toekomst van het M-team verzekerd.
De DI08 is meer dan enkel het M-team. Bedankt Leïla en Sandra voor alle hulp bij de
administratieve uitdagingen. Bedankt Veronique, Marnik en Ria om materiaal klaar te zetten
of een pittig verhaal te vertellen. Samen koeien in stukken snijden, weefsel verzamelen in
het slachthuis of varkens op de weegschaal zetten klinkt weinig plezant maar was het toch.
Bedankt Jenne, Lotte, Ilse, Annelies, Ruben en Rubèn! Veel succes bij jouw revalidatie Lotte,
aarzel niet om mij te bellen als ik je ergens mee kan helpen. Bedankt Jenne, Miel en Kristine
voor de fijne tijd in Kansas City!
Behalve DI08 heb ik nog heel wat andere plaatsen op de faculteit onveilig gemaakt.
Tijdens mijn thesis en doctoraat bracht ik vele uren door op het labo voor dierlijke genetica.
Bedankt Mario, Domi, Ruben, Caro, Karen, Cindy, Linda, Sara, Jozefien, Typhanie, Tiphanie,
Alice, Wenwen, Wei, Philippe, Sofie en Laetitia voor de hulp en de fijne koffie- en
middagpauzes! Ik hoop dat ik nog eens mee kan badmintonnen. Dankzij de vele bezoekjes
aan de flowcytometer leerde ik Koen, Donna, Jella, Kristel en Evelyne van biochemie kennen.
Bedankt voor de fijne gesprekken en de hulp bij “flowcytometrische uitdagingen”. Samen
met Caro en Xanthippe van fysiologie was ik zot genoeg om een studie uit te voeren waarbij
we koeien binnen de 24u na afkalven bemonsterden. Bedankt voor jullie hulp! Bedankt
Christian om jouw fenomenale kennis rond de fysiologie van de uier met mij te delen.

Dankwoord - Acknowledgements
248
Jammer dat ik pas op het einde van mijn doctoraat met jullie samen werkte. Naast bewoners
van de faculteit ontmoette ik de uiterst aangename ILVOrianen Stephanie, Ingrid, Bert V.,
Liesbet en Veerle. Veel succes in alles wat jullie doen!
“It began in Africa.”. I gained my first research experience in Jimma, Ethiopia. Thank
you Prof. Janssens for sending me there to conduct my master thesis. Thank you prof.
Duchateau for heading the IUC-JU project. I admire your passion for research in Africa.
During my stay in 2008, I received a lot of help from Seleshi and got to know Tadele. I was
very happy to return to Jimma a couple of months ago. Jimma is changing rapidly, but many
challenges remain. I will do my very best to continue helping you with your research Tadele.
I am looking forward to welcome both of you in Belgium soon!
Een studie op 50 melkveebedrijven verspreid over Vlaanderen voer je niet in je eentje
uit. Bedankt Luc, Karlien, Koen L., Linda, Werner (MCC), Koen D., Hans en Jo (DGZ) voor de
vlotte samenwerking! Bedankt Emilie, Jenne, Dimitri en Ilse om mee bloedstalen te nemen
op enkele bedrijven. Bedankt aan alle deelnemende melkveehouders en dierenartsen om
melkstalen te nemen. Zonder jullie was er geen Chapter 5.1 of 5.2. Bedankt Lydia, Karel en
André voor jullie hulp bij de infectiestudie op de proefhoeve! Bedankt Raf, Guus, Erwin en
Stan voor jullie hulp bij de studie bij de pasafgekalfde vaarzen. Ik vond het leuk om ook in
Zeeuws-Vlaanderen onderzoek uit te voeren.
Het leven bestaat uit meer dan werken. Goede vrienden stonden telkens klaar voor mij
om onderzoeksuccessen mee te vieren en tegenslagen te helpen vergeten. Bedankt Thomas,
Leen, Laurens, Floor, Jan, Emilie, Sean, Wannes, Floor, Davy, Jozefien, Ruben, Pieter, Bruno,
Steven D., Ellen, Brecht en alle vrienden die zich na 3/1 inschreven om naar mijn
doctoraatsverdediging te komen. Bedankt ook aan de vrienden die wouden maar niet
konden komen.
Ik heb het geluk gehad in een geweldig gezin op te groeien. Bedankt ma en pa voor alle
mogelijkheden die jullie mij gaven! Ik ben wie ik ben dankzij jullie genen en opvoeding. Met
Tim, Rein en Janne heb ik twee geweldige broers en een fantastische zus. Tel daar nog de
twee super “schone zussen” Saar en Clara bij en je weet dat de pret niet op kan op onze
familiefeesten. Ik kijk er naar uit om nonkel te worden Saar en Tim! Bedankt voor de hulp bij
de cover van dit boekje Clara! Bedankt oma, pepe en meme om naar Merelbeke te komen.
Jammer dat opa er niet bij kan zijn. Ook wat mijn schoonfamilie betreft ben ik met mijn gat
in de boter gevallen. Bedankt Ingrid, Johan, Bram, Gerda en André!

Dankwoord - Acknowledgements
249
Save the best for the last: één iemand in mijn leven verdient een aparte pagina in dit
proefschrift. Liefste Ilse, zoetje, echtgenote, beste vriendin, … Bedankt om samen met mij
alle vreugde en verdriet te delen. Ik houd zielsveel van jou!
Joren
Top Related