







Languages
Pages
Legal


Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 256549 9 pageshttpdxdoiorg1011552013256549
Research ArticlePhotosystem II Photochemistry and Phycobiliprotein ofthe Red Algae Kappaphycus alvarezii and Their Implications forLight Adaptation
Xiangyu Guan1 Jinfeng Wang2 Jianyi Zhu3 Chunyan Yao3 Jianguo Liu2
Song Qin4 and Peng Jiang2
1 School of Ocean Sciences China University of Geosciences Beijing 100083 China2 Key Laboratory of Experimental Marine Biology Institute of Oceanology Chinese Academy of Sciences Qingdao 266071 China3Department of Biology Changshu Institute of Technology Changshu 215500 China4Yantai Institute of Coastal Zone Research Chinese Academy of Sciences Yantai 264003 China
Correspondence should be addressed to Song Qin sqinyicaccn and Peng Jiang jiangpengqdio163com
Received 1 September 2013 Accepted 19 October 2013
Academic Editor Hanzhi Lin
Copyright copy 2013 Xiangyu Guan et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Photosystem II photochemistry and phycobiliprotein (PBP) genes of red algaeKappaphycus alvarezii rawmaterial of 120581-carrageenanused in food and pharmaceutical industries were analyzed in this study Minimum saturating irradiance (119868119896) of this algal specieswas less than 115120583molmminus2 sminus1 Its actual PSII efficiency (yield II) increased when light intensity enhanced and decreased when lightintensity reached 200 120583molmminus2 sminus1 Under dim light yield II declined at first and then increased on the fourth day Under highlight yield II retained a stable value These results indicate that K alvarezii is a low-light-adapted species but possesses regulativemechanisms in response to both excessive and deficient light Based on the PBP gene sequencesK alvarezii together with other redalgae assembled faster and showed a closer relationship with LL-Prochlorococcus compared to HL-Prochlorococcus Many aminoacid loci in PBP sequences of K alvarezii were conserved with those of LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were also found The diversities of PE and PC are proposed to have playedsome roles during the algal evolution and divergence of light adaption
1 Introduction
Kappaphycus alvarezii (Doty) Doty (Rhodophyta Solieri-aceae) is a commercially important marine alga for itshigh production of polysaccharide 120581-carrageenan which iscommonly used in food medicine and cosmetic industries[1 2] Extensive application and increasing demand makeK alvarezii widely cultivated around Southeast Asia EastAfrica and South America and promote approximately 8growth rates annually in the carrageenan industry [3 4]Photosynthetic responses of K alvarezii have frequentlybeen detected to clarify the optimal conditions that wouldmaximize its photosynthesis and growth For example theeffects of temperature salinity and UVB radiation on dif-ferent morphotypes of K alvarezii were evaluated undercontrolled conditions [5ndash7] However little is known about
their physiological changes associated with light intensitiesand the optimal photosynthetic available radiation levels ofmany locally cultivated strains remain to be determinedPhotosystem (PS) II photochemical parameters measured bypulse amplitude modulation (PAM) fluorometry have longbeen used for evaluating the photosynthetic physiology ofland plants [8 9] Moreover its application development caneasily be found in algal research [10ndash12] It should be a feasibletool for in vivo testing of the photosynthetic behaviors of Kalvarezii to different radiations as well as for ascertaining thebest light conditions
Photosynthetic behaviors and light response of plantsare largely determined and regulated by their photosyntheticunits [13 14] Phycobilisomes (PBSs) are major photosyn-thetic units that endue K alvarezii and other red algaecyanobacteria several cryptomonads and brown algae with
2 BioMed Research International
Table 1 Degenerate PCR primers of PBP genes
Primers Sequence Direction TargetKPE-1 51015840-ATGCTTGAC(T)GCA(G)TTTTCT(C)AG-31015840 Forward CPEKPE-2 51015840-TTAGCA(GT)TAA(G)A(T)GA(C)GTTGATG(T)ACG(A)-31015840 Reverse CPEKPC-1 51015840-ATGT(C)TAGAT(C)GCATTTGCC(T)AA-31015840 Forward CPCKPC-2 51015840-TTAG(A)CTTAGCGT(C)ATTAATAGC-31015840 Reverse CPCKAPC-1 51015840-ATGAGTATTG(A)TTACA(TG)AAG(A)TC-31015840 Forward APCKAPC-2 51015840-TTAACT(C)TAA(G)A(G)CCAGAAC-31015840 Reverse APC
the capability of light-harvesting and energy migration [15]For their biomedical value combinational biosynthesis andpurification of recombinant PBSs have been carried out withthe help of modern biotechnology [16 17] PBSs are com-posed of linker polypeptides and phycobiliproteins (PBPs)mainly allophycocyanin (APC) phycoerythrin (PE) andphycocyanin (PC) PBSs are one of the ways by which algaesurvive in varying and sometimes extreme habitats [18] someexamples would be Prochlorococcus and Synechococcus twogenera of abundant unicellular organisms Prochlorococcusconsist of two ecotypes specifically adapted to either low-light(LL) or high-light (HL) conditions Their pigmentation andgene complement are quite different [19] By altering theirPBPs structure and PBSs composition such as the PE diver-gence in LL-Prochlorococcus andHL-Prochlorococcus [20 21]algae evolve to accommodate different light conditions Priorstudies demonstrated that PE genes in LL-Prochlorococcussuffer frompositive selection and the selected sites are relatedto light-harvesting or energy-transferring which is consis-tent with the findings in other PBPs [22] Similarly basedon their PBS structure and composition Synechococcus weredivided into three types type 1 rods are composed of C-PConly type 2 rods are composed of eitherC-PCorR-PCIII anda PEI-like PBP type 3 rods are composed of R-PC and twoPEtypes (PEI and PEII) [23] Accordingly nucleic or amino acidsequences of PBPs and PBSs linker family may have hiddensome helpful information related to algal photosynthesis andlight adaptation [24 25] Unfortunately so far only PBPgenes of very few red algae such as Aglaothamnion neglectumand Gracilaria tenuistipitata were sequenced [26 27] Theinformation of K alvarezii is still unknown
The present study measured PSII photochemistry toevaluate photosynthetic efficiency and light response of Kalvarezii In addition PBP genes were firstly sequenced andaligned with those of other algae The results of this studywould improve our understanding of the impact of changinglight conditions on K alvarezii and the potential mechanismof light adaption of this red alga
2 Materials and Methods
21 Algal Material K alvarezii were collected from a farmpopulation in Lingshui Hainan Province China Healthyfronds were cultivated in NP-enriched seawater (N043mM P 0019mM) at 23∘C under 12 h 12 h (LD)photoperiod and 40 120583molmminus2 sminus1 light intensity The culturemedium was renewed once a week
22 Chlorophyll Fluorescent Measurement Chlorophyll flu-orescence of K alvarezii thalli was cultivated under alight intensity of 40120583molmminus2 sminus1 and then cultivated underfive different light intensities of 10 25 50 100 and200120583molmminus2 sminus1 in four days Healthy samples were dark-adapted for 15min after which actual PSII efficiency (yield II)and rapid light curves (RLC) were measured using a Water-PAM chlorophyll fluorometer with Water-EDF Fiberoptics-Emitter-Detector Unit (Walz Effeltrich Germany) Thesteady-state fluorescence level during exposure to cultivatedlight intensity (Ft) was detected bymeasuring the value undera 03 120583molmminus2 sminus1 modulated light and a 40 120583molmminus2 sminus1actinic light Maximum fluorescence level during illumi-nation (Fm1015840) was measured by a 08 s saturating pulse at4000 120583molmminus2 sminus1 Yield II was calculated according to theformula yield II = (Fm1015840minusFt)Fm1015840 Ft and Fm1015840 weremeasuredunder nine different and increasingly actinic lights (PAR = 043 64 92 138 209 309 479 and 714 120583molmminus2 sminus1) of a 10 sduration and a 08 s saturating pulse at 4000 120583molmminus2 sminus1Relative electron transport rate (rETR) was calculated asrETR = yield times PAR times 119860 times 05 in which PAR symbolizesresponses to photosynthetic active radiation (119860 absorbancyindex = 085 in this study) From the RLC relative maximumelectron transport rate (rETRmax) and initial slope (120572) wereobtainedMinimum saturating irradiance (119868119896) = rETRmax120572Results were expressed as mean plusmn SD in which data fromthree independent experiments were analyzed with the sta-tistical software STATISTICA70 (119905-test 119875 lt 005)
23 DNA Extraction and Sequencing of PBP Genes Algalsamples were ground in liquid nitrogen and the genomicDNA was extracted with a Plant Genome DNA Kit (TiangenBeijing China) according to the manufacturerrsquos instruc-tions
Based on the conserved sequences of red algae Aneglectum (Z11906 Z11905) Ceramium boydenii (AF526383)Gracilaria lemaneiformis (AF275685) G tenuistipitata varliui (AY673996) Griffithsia monilis (Z98528) Porphyra hai-tanensis (DQ449071 AY372218) and Porphyra yezoensis(DQ666487) degenerate oligonucleotide primers of PBPgenes were designed (Table 1) The PCR program utilized aninitial denaturation at 94∘C for 5min followed by 35 cycles at94∘C for 1min 47∘C for 1min and 72∘C for 1min 30 s with afinal elongation at 72∘C for 10min Amplification reactionswere performed in a Biometra Thermal Cycler (BiometraGottingen Germany)
BioMed Research International 3
PCR products were purified with a TIANgel Midi Purifi-cation Kit (Tiangen Beijing China) and then constructedinto a pMD18-T vector system (Takara Dalian China) Theligation products were transferred into Escherichia coli TOP10 strains Five positive clones of each amplification productwere checked by their electrophoretic mobility and PCRanalysis after the recombinant plasmids were selected undera 100 120583gmLminus1 concentration of ampicillin Double strandsof these target segments were sequenced by Shanghai SunnyBiotechnology Co Ltd (Sunny Shanghai China)
24 Multiple Sequence Alignment and Phylogenetic AnalysisA twenty-four-cyanobacterial-genome databasewas accessedfrom JGI (httpwwwjgidoegov) in FASTA format andthe species Gloeobacter violaceus (GV) Trichodesmiumerythraeum (TE) Synechococcus (S) and Prochlorococ-cus (P) were examined PBP sequences of Rhodophyta(including species of Bangiophyceae and Florideophycease)Cyanophora paradoxa (M11159 CP) and Guillardia theta(AM183803) were searched at NCBI [28 29] PBP nucleotidesequences of K alvarezii (KA) were cloned in this paper andtranslated to amino acid sequences by Omiga [30] Proteinsequences of the PBSs previously described were used asdatabase queries Each protein in this query dataset was usedto search for potential novel sequences in above sequencedcyanobacterial species genomes using the BLASTP andTBLASTN programs [31] Sequences giving better reciprocalBLAST hits were assumed to be capable of identifyinghomologous counterparts in these species if they could bealigned with at least the BLAST-Score gt90 and the 119864-valuelt1eminus10 The search was iterated until convergence and thenexamined individually
Multiple protein sequences alignment was performedusing ClustalX [32] Neighbor joining (NJ) and maximumparsimony (MP) methods in MEGA5 were used to constructthe phylogenetic tree [33] in which the confidence level ofeach branch was determined by analyzing 1000 bootstrapreplicates Bootstrap values gt50 were generated
25 Tertiary Structure Prediction Tertiary structure of PE 120572-and 120573-subunits of K alvarezii was analyzed using homol-ogy modeling Next 120572- and 120573-PE amino acid sequencesof K alvarezii were submitted to the protein-modelingserver SWISSMODEL (httpswissmodelexpasyorg) andpredicted with PDB-1b8dK using PDB-1liaB as the modeltemplate All the manipulations were performed using Pdb-Viewer
3 Results
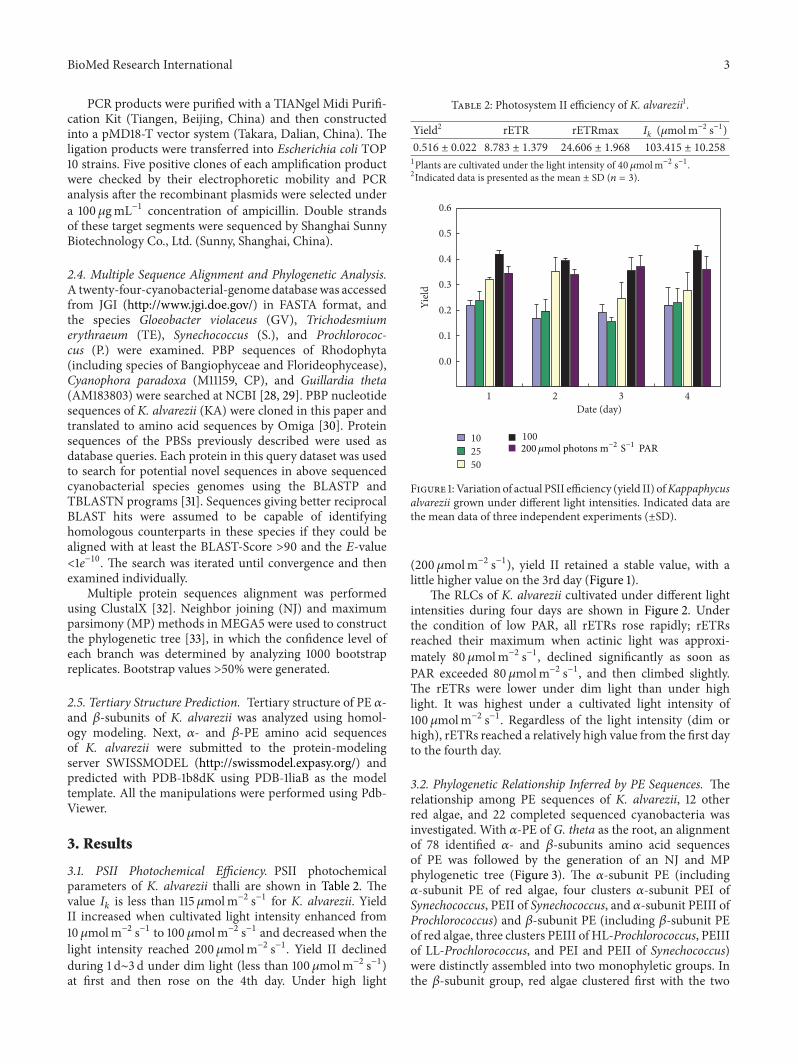
31 PSII Photochemical Efficiency PSII photochemicalparameters of K alvarezii thalli are shown in Table 2 Thevalue 119868119896 is less than 115 120583molmminus2 sminus1 for K alvarezii YieldII increased when cultivated light intensity enhanced from10 120583molmminus2 sminus1 to 100 120583molmminus2 sminus1 and decreased when thelight intensity reached 200 120583molmminus2 sminus1 Yield II declinedduring 1 dsim3 d under dim light (less than 100120583molmminus2 sminus1)at first and then rose on the 4th day Under high light
Table 2 Photosystem II efficiency of K alvarezii1
Yield2 rETR rETRmax 119868119896 (120583molmminus2 sminus1)0516 plusmn 0022 8783 plusmn 1379 24606 plusmn 1968 103415 plusmn 10258
1Plants are cultivated under the light intensity of 40120583molmminus2 sminus12Indicated data is presented as the mean plusmn SD (119899 = 3)
00
01
02
03
04
05
06
1 2 3 4
Yield
Date (day)
102550
100200120583mol photons mminus2 Sminus1 PAR
Figure 1Variation of actual PSII efficiency (yield II) ofKappaphycusalvarezii grown under different light intensities Indicated data arethe mean data of three independent experiments (plusmnSD)
(200120583molmminus2 sminus1) yield II retained a stable value with alittle higher value on the 3rd day (Figure 1)
The RLCs of K alvarezii cultivated under different lightintensities during four days are shown in Figure 2 Underthe condition of low PAR all rETRs rose rapidly rETRsreached their maximum when actinic light was approxi-mately 80 120583molmminus2 sminus1 declined significantly as soon asPAR exceeded 80 120583molmminus2 sminus1 and then climbed slightlyThe rETRs were lower under dim light than under highlight It was highest under a cultivated light intensity of100 120583molmminus2 sminus1 Regardless of the light intensity (dim orhigh) rETRs reached a relatively high value from the first dayto the fourth day
32 Phylogenetic Relationship Inferred by PE Sequences Therelationship among PE sequences of K alvarezii 12 otherred algae and 22 completed sequenced cyanobacteria wasinvestigated With 120572-PE of G theta as the root an alignmentof 78 identified 120572- and 120573-subunits amino acid sequencesof PE was followed by the generation of an NJ and MPphylogenetic tree (Figure 3) The 120572-subunit PE (including120572-subunit PE of red algae four clusters 120572-subunit PEI ofSynechococcus PEII of Synechococcus and 120572-subunit PEIII ofProchlorococcus) and 120573-subunit PE (including 120573-subunit PEof red algae three clusters PEIII of HL-Prochlorococcus PEIIIof LL-Prochlorococcus and PEI and PEII of Synechococcus)were distinctly assembled into two monophyletic groups Inthe 120573-subunit group red algae clustered first with the two
4 BioMed Research International
0
2
4
6
8
10
12
14
20010050
2510
rETR
0 200 400 600 800Light intensities (120583mol photons mminus2 sminus1)
1 d
(a)
0
2
4
6
8
10
12
14
0 200 400 600 800
rETR
20010050
2510
Light intensities (120583mol photons mminus2 sminus1)
2 d
(b)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
3 d
(c)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
4 d
(d)
Figure 2 Rapid light curves (RLCs) of Kappaphycus alvarezii cultivated under different light intensities Indicated data are the mean data ofthree independent experiments (plusmnSD)
cyanobacteria G violaceus and T erythraeum and thenassembled with the PEI and PEII clades of SynechococcusCompared with HL-Prochlorococcus they have a closer rela-tionship with LL-Prochlorococcus Red algal PE sequences aremore similar to LL-Prochlorococcus PEs than to HL-PEs Apossible explanation of this result is that HL-Prochlorococcusevolves faster than LL and also exhibits much higher GCcontent Hence the similarities among red algal- HL-and LL-PEs do not necessarily reflect their photosyntheticproperties and relationships In the 120572-subunit group redalgae successively assembled with PEI of Synechococcus LL-Prochlorococcus and T erythraeum PEII of Synechococcusformed a cluster by itself
33 Tertiary Structure of PE Forty-eight amino acid lociconserved with LL-Prochlorococcus but diverged with HL-Prochlorococcuswere mapped onto 120573-PE tertiary structure ofK alvarezii in blue color (Figure 4(a)) These loci distributeto the following six domains (1) N-terminal 2-10aa domainis obviously conserved including the loci 3D function ofinteraction of N-terminus of 120572- and 120573-subunits (2) 65-81aadomain (except 67I 68A 72N 75T 76N and 79M) is adjacentto themotif of chromophore interaction (3) 92-100aa domain(including 92Y 94S 96A 98L 99A and 100G) in which91R 95Y are functions of 120572- and 120573-interaction and 100G isat the end of helix E (4) 107-115aa domain (including 107D108R 111N 112G and 115E) includes the functional loci 108R
BioMed Research International 5
PCCMP1986-120573-HPAS9601-120573-HPMIT9312-120573-HPMIT9303-120573-HPMIT9313-120573-H
PCCMP1357-III120573-LPMIT9211-120573-L
PNATL2A-III120573-LPNATL2A-120573-L
82728584
9367
9999
7267
9055
66
50
54
65
76
53
50
5853
69
65
9472
9887
9895
9280
6357
98979786
9999
7753
6851
63556956
7876
9788
9794
9892
97999897
9996
7254
9589
8776
9187
10099
10099
10099
10099
KA-120573
120572KA-
GL-120573CB-120573PB-120573
PP-120573PT-120573PH-120573PY-120573
CO-120573GT-120573
RV-120573AN-120573GM-120573
GVPCC7421-120573TEIMS101-120573
SCC9311-I120573SCC9902-I120573SBL107-I120573SRCC307-I120573SRS9916-I120573
SCC9605-I120573SWH8102-I120573
SWH7803-I120573SWH7805-I120573
SCC9311-II120573
SCC9902-II120573SBL107-II120573
SRCC307-II120573SRS9916-II120573
SCC9605-II120573SWH8102-II120573
SWH7803-II120573
PCCMP1357-III120572-LPMIT9211-120572-LPNATL2A-III120572-LPNATL1A-120572-L
PMIT9303-120572-LPMIT9313-120572-L
GL-120572
CB-120572PB-120572
PP-120572PT-120572PH-120572
PY-120572
CO-120572
GT-120572RV-120572
AN-120572GM-120572
SCC9311-I120572SRCC307-I120572SCC9902-I120572
SRS9916-I120572SBL107-I120572SCC9605-I120572SWH8102-I120572
SWH7803-I120572SWH7805-I120572
TEIMS101-120572SCC9311-II120572
SRCC307-II120572SRS9916-II120572
SCC9605-II120572SWH8102-II120572
SBL107-II120572SCC9902-II120572SWH7803-II120572
20
7268
74609498
9998
ProchlorococcusSynechococcusKappaphycus alvareziiGracilaria lemaneiformisCeramium boydeniiPolysiphonia boldiiPorphyra purpureaPorphyra teneraPorphyra haitanensisPorphyra yezoensisCorallina officinalisGracilaria tenuistipitataRhodella violaceaAglaothamnion neglectumGriffithsia monilisGloeobacter violaceus
PS
KAGLCBPBPPPTPHPYCOGTRVANGMGV
Guillardia theta-120572
Figure 3 Phylogenetic tree ofKappaphycus alvarezii 12 other red algae and sequenced cyanobacteria based onPENumbers at nodes indicatebranch support given as bootstrap values from neighbor joining (NJ)maximum parsimony (MP) analysis Numbers are only shown if theyexceed 50 respectively
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
Table 1 Degenerate PCR primers of PBP genes
Primers Sequence Direction TargetKPE-1 51015840-ATGCTTGAC(T)GCA(G)TTTTCT(C)AG-31015840 Forward CPEKPE-2 51015840-TTAGCA(GT)TAA(G)A(T)GA(C)GTTGATG(T)ACG(A)-31015840 Reverse CPEKPC-1 51015840-ATGT(C)TAGAT(C)GCATTTGCC(T)AA-31015840 Forward CPCKPC-2 51015840-TTAG(A)CTTAGCGT(C)ATTAATAGC-31015840 Reverse CPCKAPC-1 51015840-ATGAGTATTG(A)TTACA(TG)AAG(A)TC-31015840 Forward APCKAPC-2 51015840-TTAACT(C)TAA(G)A(G)CCAGAAC-31015840 Reverse APC
the capability of light-harvesting and energy migration [15]For their biomedical value combinational biosynthesis andpurification of recombinant PBSs have been carried out withthe help of modern biotechnology [16 17] PBSs are com-posed of linker polypeptides and phycobiliproteins (PBPs)mainly allophycocyanin (APC) phycoerythrin (PE) andphycocyanin (PC) PBSs are one of the ways by which algaesurvive in varying and sometimes extreme habitats [18] someexamples would be Prochlorococcus and Synechococcus twogenera of abundant unicellular organisms Prochlorococcusconsist of two ecotypes specifically adapted to either low-light(LL) or high-light (HL) conditions Their pigmentation andgene complement are quite different [19] By altering theirPBPs structure and PBSs composition such as the PE diver-gence in LL-Prochlorococcus andHL-Prochlorococcus [20 21]algae evolve to accommodate different light conditions Priorstudies demonstrated that PE genes in LL-Prochlorococcussuffer frompositive selection and the selected sites are relatedto light-harvesting or energy-transferring which is consis-tent with the findings in other PBPs [22] Similarly basedon their PBS structure and composition Synechococcus weredivided into three types type 1 rods are composed of C-PConly type 2 rods are composed of eitherC-PCorR-PCIII anda PEI-like PBP type 3 rods are composed of R-PC and twoPEtypes (PEI and PEII) [23] Accordingly nucleic or amino acidsequences of PBPs and PBSs linker family may have hiddensome helpful information related to algal photosynthesis andlight adaptation [24 25] Unfortunately so far only PBPgenes of very few red algae such as Aglaothamnion neglectumand Gracilaria tenuistipitata were sequenced [26 27] Theinformation of K alvarezii is still unknown
The present study measured PSII photochemistry toevaluate photosynthetic efficiency and light response of Kalvarezii In addition PBP genes were firstly sequenced andaligned with those of other algae The results of this studywould improve our understanding of the impact of changinglight conditions on K alvarezii and the potential mechanismof light adaption of this red alga
2 Materials and Methods
21 Algal Material K alvarezii were collected from a farmpopulation in Lingshui Hainan Province China Healthyfronds were cultivated in NP-enriched seawater (N043mM P 0019mM) at 23∘C under 12 h 12 h (LD)photoperiod and 40 120583molmminus2 sminus1 light intensity The culturemedium was renewed once a week
22 Chlorophyll Fluorescent Measurement Chlorophyll flu-orescence of K alvarezii thalli was cultivated under alight intensity of 40120583molmminus2 sminus1 and then cultivated underfive different light intensities of 10 25 50 100 and200120583molmminus2 sminus1 in four days Healthy samples were dark-adapted for 15min after which actual PSII efficiency (yield II)and rapid light curves (RLC) were measured using a Water-PAM chlorophyll fluorometer with Water-EDF Fiberoptics-Emitter-Detector Unit (Walz Effeltrich Germany) Thesteady-state fluorescence level during exposure to cultivatedlight intensity (Ft) was detected bymeasuring the value undera 03 120583molmminus2 sminus1 modulated light and a 40 120583molmminus2 sminus1actinic light Maximum fluorescence level during illumi-nation (Fm1015840) was measured by a 08 s saturating pulse at4000 120583molmminus2 sminus1 Yield II was calculated according to theformula yield II = (Fm1015840minusFt)Fm1015840 Ft and Fm1015840 weremeasuredunder nine different and increasingly actinic lights (PAR = 043 64 92 138 209 309 479 and 714 120583molmminus2 sminus1) of a 10 sduration and a 08 s saturating pulse at 4000 120583molmminus2 sminus1Relative electron transport rate (rETR) was calculated asrETR = yield times PAR times 119860 times 05 in which PAR symbolizesresponses to photosynthetic active radiation (119860 absorbancyindex = 085 in this study) From the RLC relative maximumelectron transport rate (rETRmax) and initial slope (120572) wereobtainedMinimum saturating irradiance (119868119896) = rETRmax120572Results were expressed as mean plusmn SD in which data fromthree independent experiments were analyzed with the sta-tistical software STATISTICA70 (119905-test 119875 lt 005)
23 DNA Extraction and Sequencing of PBP Genes Algalsamples were ground in liquid nitrogen and the genomicDNA was extracted with a Plant Genome DNA Kit (TiangenBeijing China) according to the manufacturerrsquos instruc-tions
Based on the conserved sequences of red algae Aneglectum (Z11906 Z11905) Ceramium boydenii (AF526383)Gracilaria lemaneiformis (AF275685) G tenuistipitata varliui (AY673996) Griffithsia monilis (Z98528) Porphyra hai-tanensis (DQ449071 AY372218) and Porphyra yezoensis(DQ666487) degenerate oligonucleotide primers of PBPgenes were designed (Table 1) The PCR program utilized aninitial denaturation at 94∘C for 5min followed by 35 cycles at94∘C for 1min 47∘C for 1min and 72∘C for 1min 30 s with afinal elongation at 72∘C for 10min Amplification reactionswere performed in a Biometra Thermal Cycler (BiometraGottingen Germany)
BioMed Research International 3
PCR products were purified with a TIANgel Midi Purifi-cation Kit (Tiangen Beijing China) and then constructedinto a pMD18-T vector system (Takara Dalian China) Theligation products were transferred into Escherichia coli TOP10 strains Five positive clones of each amplification productwere checked by their electrophoretic mobility and PCRanalysis after the recombinant plasmids were selected undera 100 120583gmLminus1 concentration of ampicillin Double strandsof these target segments were sequenced by Shanghai SunnyBiotechnology Co Ltd (Sunny Shanghai China)
24 Multiple Sequence Alignment and Phylogenetic AnalysisA twenty-four-cyanobacterial-genome databasewas accessedfrom JGI (httpwwwjgidoegov) in FASTA format andthe species Gloeobacter violaceus (GV) Trichodesmiumerythraeum (TE) Synechococcus (S) and Prochlorococ-cus (P) were examined PBP sequences of Rhodophyta(including species of Bangiophyceae and Florideophycease)Cyanophora paradoxa (M11159 CP) and Guillardia theta(AM183803) were searched at NCBI [28 29] PBP nucleotidesequences of K alvarezii (KA) were cloned in this paper andtranslated to amino acid sequences by Omiga [30] Proteinsequences of the PBSs previously described were used asdatabase queries Each protein in this query dataset was usedto search for potential novel sequences in above sequencedcyanobacterial species genomes using the BLASTP andTBLASTN programs [31] Sequences giving better reciprocalBLAST hits were assumed to be capable of identifyinghomologous counterparts in these species if they could bealigned with at least the BLAST-Score gt90 and the 119864-valuelt1eminus10 The search was iterated until convergence and thenexamined individually
Multiple protein sequences alignment was performedusing ClustalX [32] Neighbor joining (NJ) and maximumparsimony (MP) methods in MEGA5 were used to constructthe phylogenetic tree [33] in which the confidence level ofeach branch was determined by analyzing 1000 bootstrapreplicates Bootstrap values gt50 were generated
25 Tertiary Structure Prediction Tertiary structure of PE 120572-and 120573-subunits of K alvarezii was analyzed using homol-ogy modeling Next 120572- and 120573-PE amino acid sequencesof K alvarezii were submitted to the protein-modelingserver SWISSMODEL (httpswissmodelexpasyorg) andpredicted with PDB-1b8dK using PDB-1liaB as the modeltemplate All the manipulations were performed using Pdb-Viewer
3 Results
31 PSII Photochemical Efficiency PSII photochemicalparameters of K alvarezii thalli are shown in Table 2 Thevalue 119868119896 is less than 115 120583molmminus2 sminus1 for K alvarezii YieldII increased when cultivated light intensity enhanced from10 120583molmminus2 sminus1 to 100 120583molmminus2 sminus1 and decreased when thelight intensity reached 200 120583molmminus2 sminus1 Yield II declinedduring 1 dsim3 d under dim light (less than 100120583molmminus2 sminus1)at first and then rose on the 4th day Under high light
Table 2 Photosystem II efficiency of K alvarezii1
Yield2 rETR rETRmax 119868119896 (120583molmminus2 sminus1)0516 plusmn 0022 8783 plusmn 1379 24606 plusmn 1968 103415 plusmn 10258
1Plants are cultivated under the light intensity of 40120583molmminus2 sminus12Indicated data is presented as the mean plusmn SD (119899 = 3)
00
01
02
03
04
05
06
1 2 3 4
Yield
Date (day)
102550
100200120583mol photons mminus2 Sminus1 PAR
Figure 1Variation of actual PSII efficiency (yield II) ofKappaphycusalvarezii grown under different light intensities Indicated data arethe mean data of three independent experiments (plusmnSD)
(200120583molmminus2 sminus1) yield II retained a stable value with alittle higher value on the 3rd day (Figure 1)
The RLCs of K alvarezii cultivated under different lightintensities during four days are shown in Figure 2 Underthe condition of low PAR all rETRs rose rapidly rETRsreached their maximum when actinic light was approxi-mately 80 120583molmminus2 sminus1 declined significantly as soon asPAR exceeded 80 120583molmminus2 sminus1 and then climbed slightlyThe rETRs were lower under dim light than under highlight It was highest under a cultivated light intensity of100 120583molmminus2 sminus1 Regardless of the light intensity (dim orhigh) rETRs reached a relatively high value from the first dayto the fourth day
32 Phylogenetic Relationship Inferred by PE Sequences Therelationship among PE sequences of K alvarezii 12 otherred algae and 22 completed sequenced cyanobacteria wasinvestigated With 120572-PE of G theta as the root an alignmentof 78 identified 120572- and 120573-subunits amino acid sequencesof PE was followed by the generation of an NJ and MPphylogenetic tree (Figure 3) The 120572-subunit PE (including120572-subunit PE of red algae four clusters 120572-subunit PEI ofSynechococcus PEII of Synechococcus and 120572-subunit PEIII ofProchlorococcus) and 120573-subunit PE (including 120573-subunit PEof red algae three clusters PEIII of HL-Prochlorococcus PEIIIof LL-Prochlorococcus and PEI and PEII of Synechococcus)were distinctly assembled into two monophyletic groups Inthe 120573-subunit group red algae clustered first with the two
4 BioMed Research International
0
2
4
6
8
10
12
14
20010050
2510
rETR
0 200 400 600 800Light intensities (120583mol photons mminus2 sminus1)
1 d
(a)
0
2
4
6
8
10
12
14
0 200 400 600 800
rETR
20010050
2510
Light intensities (120583mol photons mminus2 sminus1)
2 d
(b)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
3 d
(c)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
4 d
(d)
Figure 2 Rapid light curves (RLCs) of Kappaphycus alvarezii cultivated under different light intensities Indicated data are the mean data ofthree independent experiments (plusmnSD)
cyanobacteria G violaceus and T erythraeum and thenassembled with the PEI and PEII clades of SynechococcusCompared with HL-Prochlorococcus they have a closer rela-tionship with LL-Prochlorococcus Red algal PE sequences aremore similar to LL-Prochlorococcus PEs than to HL-PEs Apossible explanation of this result is that HL-Prochlorococcusevolves faster than LL and also exhibits much higher GCcontent Hence the similarities among red algal- HL-and LL-PEs do not necessarily reflect their photosyntheticproperties and relationships In the 120572-subunit group redalgae successively assembled with PEI of Synechococcus LL-Prochlorococcus and T erythraeum PEII of Synechococcusformed a cluster by itself
33 Tertiary Structure of PE Forty-eight amino acid lociconserved with LL-Prochlorococcus but diverged with HL-Prochlorococcuswere mapped onto 120573-PE tertiary structure ofK alvarezii in blue color (Figure 4(a)) These loci distributeto the following six domains (1) N-terminal 2-10aa domainis obviously conserved including the loci 3D function ofinteraction of N-terminus of 120572- and 120573-subunits (2) 65-81aadomain (except 67I 68A 72N 75T 76N and 79M) is adjacentto themotif of chromophore interaction (3) 92-100aa domain(including 92Y 94S 96A 98L 99A and 100G) in which91R 95Y are functions of 120572- and 120573-interaction and 100G isat the end of helix E (4) 107-115aa domain (including 107D108R 111N 112G and 115E) includes the functional loci 108R
BioMed Research International 5
PCCMP1986-120573-HPAS9601-120573-HPMIT9312-120573-HPMIT9303-120573-HPMIT9313-120573-H
PCCMP1357-III120573-LPMIT9211-120573-L
PNATL2A-III120573-LPNATL2A-120573-L
82728584
9367
9999
7267
9055
66
50
54
65
76
53
50
5853
69
65
9472
9887
9895
9280
6357
98979786
9999
7753
6851
63556956
7876
9788
9794
9892
97999897
9996
7254
9589
8776
9187
10099
10099
10099
10099
KA-120573
120572KA-
GL-120573CB-120573PB-120573
PP-120573PT-120573PH-120573PY-120573
CO-120573GT-120573
RV-120573AN-120573GM-120573
GVPCC7421-120573TEIMS101-120573
SCC9311-I120573SCC9902-I120573SBL107-I120573SRCC307-I120573SRS9916-I120573
SCC9605-I120573SWH8102-I120573
SWH7803-I120573SWH7805-I120573
SCC9311-II120573
SCC9902-II120573SBL107-II120573
SRCC307-II120573SRS9916-II120573
SCC9605-II120573SWH8102-II120573
SWH7803-II120573
PCCMP1357-III120572-LPMIT9211-120572-LPNATL2A-III120572-LPNATL1A-120572-L
PMIT9303-120572-LPMIT9313-120572-L
GL-120572
CB-120572PB-120572
PP-120572PT-120572PH-120572
PY-120572
CO-120572
GT-120572RV-120572
AN-120572GM-120572
SCC9311-I120572SRCC307-I120572SCC9902-I120572
SRS9916-I120572SBL107-I120572SCC9605-I120572SWH8102-I120572
SWH7803-I120572SWH7805-I120572
TEIMS101-120572SCC9311-II120572
SRCC307-II120572SRS9916-II120572
SCC9605-II120572SWH8102-II120572
SBL107-II120572SCC9902-II120572SWH7803-II120572
20
7268
74609498
9998
ProchlorococcusSynechococcusKappaphycus alvareziiGracilaria lemaneiformisCeramium boydeniiPolysiphonia boldiiPorphyra purpureaPorphyra teneraPorphyra haitanensisPorphyra yezoensisCorallina officinalisGracilaria tenuistipitataRhodella violaceaAglaothamnion neglectumGriffithsia monilisGloeobacter violaceus
PS
KAGLCBPBPPPTPHPYCOGTRVANGMGV
Guillardia theta-120572
Figure 3 Phylogenetic tree ofKappaphycus alvarezii 12 other red algae and sequenced cyanobacteria based onPENumbers at nodes indicatebranch support given as bootstrap values from neighbor joining (NJ)maximum parsimony (MP) analysis Numbers are only shown if theyexceed 50 respectively
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
BioMed Research International 3
PCR products were purified with a TIANgel Midi Purifi-cation Kit (Tiangen Beijing China) and then constructedinto a pMD18-T vector system (Takara Dalian China) Theligation products were transferred into Escherichia coli TOP10 strains Five positive clones of each amplification productwere checked by their electrophoretic mobility and PCRanalysis after the recombinant plasmids were selected undera 100 120583gmLminus1 concentration of ampicillin Double strandsof these target segments were sequenced by Shanghai SunnyBiotechnology Co Ltd (Sunny Shanghai China)
24 Multiple Sequence Alignment and Phylogenetic AnalysisA twenty-four-cyanobacterial-genome databasewas accessedfrom JGI (httpwwwjgidoegov) in FASTA format andthe species Gloeobacter violaceus (GV) Trichodesmiumerythraeum (TE) Synechococcus (S) and Prochlorococ-cus (P) were examined PBP sequences of Rhodophyta(including species of Bangiophyceae and Florideophycease)Cyanophora paradoxa (M11159 CP) and Guillardia theta(AM183803) were searched at NCBI [28 29] PBP nucleotidesequences of K alvarezii (KA) were cloned in this paper andtranslated to amino acid sequences by Omiga [30] Proteinsequences of the PBSs previously described were used asdatabase queries Each protein in this query dataset was usedto search for potential novel sequences in above sequencedcyanobacterial species genomes using the BLASTP andTBLASTN programs [31] Sequences giving better reciprocalBLAST hits were assumed to be capable of identifyinghomologous counterparts in these species if they could bealigned with at least the BLAST-Score gt90 and the 119864-valuelt1eminus10 The search was iterated until convergence and thenexamined individually
Multiple protein sequences alignment was performedusing ClustalX [32] Neighbor joining (NJ) and maximumparsimony (MP) methods in MEGA5 were used to constructthe phylogenetic tree [33] in which the confidence level ofeach branch was determined by analyzing 1000 bootstrapreplicates Bootstrap values gt50 were generated
25 Tertiary Structure Prediction Tertiary structure of PE 120572-and 120573-subunits of K alvarezii was analyzed using homol-ogy modeling Next 120572- and 120573-PE amino acid sequencesof K alvarezii were submitted to the protein-modelingserver SWISSMODEL (httpswissmodelexpasyorg) andpredicted with PDB-1b8dK using PDB-1liaB as the modeltemplate All the manipulations were performed using Pdb-Viewer
3 Results
31 PSII Photochemical Efficiency PSII photochemicalparameters of K alvarezii thalli are shown in Table 2 Thevalue 119868119896 is less than 115 120583molmminus2 sminus1 for K alvarezii YieldII increased when cultivated light intensity enhanced from10 120583molmminus2 sminus1 to 100 120583molmminus2 sminus1 and decreased when thelight intensity reached 200 120583molmminus2 sminus1 Yield II declinedduring 1 dsim3 d under dim light (less than 100120583molmminus2 sminus1)at first and then rose on the 4th day Under high light
Table 2 Photosystem II efficiency of K alvarezii1
Yield2 rETR rETRmax 119868119896 (120583molmminus2 sminus1)0516 plusmn 0022 8783 plusmn 1379 24606 plusmn 1968 103415 plusmn 10258
1Plants are cultivated under the light intensity of 40120583molmminus2 sminus12Indicated data is presented as the mean plusmn SD (119899 = 3)
00
01
02
03
04
05
06
1 2 3 4
Yield
Date (day)
102550
100200120583mol photons mminus2 Sminus1 PAR
Figure 1Variation of actual PSII efficiency (yield II) ofKappaphycusalvarezii grown under different light intensities Indicated data arethe mean data of three independent experiments (plusmnSD)
(200120583molmminus2 sminus1) yield II retained a stable value with alittle higher value on the 3rd day (Figure 1)
The RLCs of K alvarezii cultivated under different lightintensities during four days are shown in Figure 2 Underthe condition of low PAR all rETRs rose rapidly rETRsreached their maximum when actinic light was approxi-mately 80 120583molmminus2 sminus1 declined significantly as soon asPAR exceeded 80 120583molmminus2 sminus1 and then climbed slightlyThe rETRs were lower under dim light than under highlight It was highest under a cultivated light intensity of100 120583molmminus2 sminus1 Regardless of the light intensity (dim orhigh) rETRs reached a relatively high value from the first dayto the fourth day
32 Phylogenetic Relationship Inferred by PE Sequences Therelationship among PE sequences of K alvarezii 12 otherred algae and 22 completed sequenced cyanobacteria wasinvestigated With 120572-PE of G theta as the root an alignmentof 78 identified 120572- and 120573-subunits amino acid sequencesof PE was followed by the generation of an NJ and MPphylogenetic tree (Figure 3) The 120572-subunit PE (including120572-subunit PE of red algae four clusters 120572-subunit PEI ofSynechococcus PEII of Synechococcus and 120572-subunit PEIII ofProchlorococcus) and 120573-subunit PE (including 120573-subunit PEof red algae three clusters PEIII of HL-Prochlorococcus PEIIIof LL-Prochlorococcus and PEI and PEII of Synechococcus)were distinctly assembled into two monophyletic groups Inthe 120573-subunit group red algae clustered first with the two
4 BioMed Research International
0
2
4
6
8
10
12
14
20010050
2510
rETR
0 200 400 600 800Light intensities (120583mol photons mminus2 sminus1)
1 d
(a)
0
2
4
6
8
10
12
14
0 200 400 600 800
rETR
20010050
2510
Light intensities (120583mol photons mminus2 sminus1)
2 d
(b)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
3 d
(c)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
4 d
(d)
Figure 2 Rapid light curves (RLCs) of Kappaphycus alvarezii cultivated under different light intensities Indicated data are the mean data ofthree independent experiments (plusmnSD)
cyanobacteria G violaceus and T erythraeum and thenassembled with the PEI and PEII clades of SynechococcusCompared with HL-Prochlorococcus they have a closer rela-tionship with LL-Prochlorococcus Red algal PE sequences aremore similar to LL-Prochlorococcus PEs than to HL-PEs Apossible explanation of this result is that HL-Prochlorococcusevolves faster than LL and also exhibits much higher GCcontent Hence the similarities among red algal- HL-and LL-PEs do not necessarily reflect their photosyntheticproperties and relationships In the 120572-subunit group redalgae successively assembled with PEI of Synechococcus LL-Prochlorococcus and T erythraeum PEII of Synechococcusformed a cluster by itself
33 Tertiary Structure of PE Forty-eight amino acid lociconserved with LL-Prochlorococcus but diverged with HL-Prochlorococcuswere mapped onto 120573-PE tertiary structure ofK alvarezii in blue color (Figure 4(a)) These loci distributeto the following six domains (1) N-terminal 2-10aa domainis obviously conserved including the loci 3D function ofinteraction of N-terminus of 120572- and 120573-subunits (2) 65-81aadomain (except 67I 68A 72N 75T 76N and 79M) is adjacentto themotif of chromophore interaction (3) 92-100aa domain(including 92Y 94S 96A 98L 99A and 100G) in which91R 95Y are functions of 120572- and 120573-interaction and 100G isat the end of helix E (4) 107-115aa domain (including 107D108R 111N 112G and 115E) includes the functional loci 108R
BioMed Research International 5
PCCMP1986-120573-HPAS9601-120573-HPMIT9312-120573-HPMIT9303-120573-HPMIT9313-120573-H
PCCMP1357-III120573-LPMIT9211-120573-L
PNATL2A-III120573-LPNATL2A-120573-L
82728584
9367
9999
7267
9055
66
50
54
65
76
53
50
5853
69
65
9472
9887
9895
9280
6357
98979786
9999
7753
6851
63556956
7876
9788
9794
9892
97999897
9996
7254
9589
8776
9187
10099
10099
10099
10099
KA-120573
120572KA-
GL-120573CB-120573PB-120573
PP-120573PT-120573PH-120573PY-120573
CO-120573GT-120573
RV-120573AN-120573GM-120573
GVPCC7421-120573TEIMS101-120573
SCC9311-I120573SCC9902-I120573SBL107-I120573SRCC307-I120573SRS9916-I120573
SCC9605-I120573SWH8102-I120573
SWH7803-I120573SWH7805-I120573
SCC9311-II120573
SCC9902-II120573SBL107-II120573
SRCC307-II120573SRS9916-II120573
SCC9605-II120573SWH8102-II120573
SWH7803-II120573
PCCMP1357-III120572-LPMIT9211-120572-LPNATL2A-III120572-LPNATL1A-120572-L
PMIT9303-120572-LPMIT9313-120572-L
GL-120572
CB-120572PB-120572
PP-120572PT-120572PH-120572
PY-120572
CO-120572
GT-120572RV-120572
AN-120572GM-120572
SCC9311-I120572SRCC307-I120572SCC9902-I120572
SRS9916-I120572SBL107-I120572SCC9605-I120572SWH8102-I120572
SWH7803-I120572SWH7805-I120572
TEIMS101-120572SCC9311-II120572
SRCC307-II120572SRS9916-II120572
SCC9605-II120572SWH8102-II120572
SBL107-II120572SCC9902-II120572SWH7803-II120572
20
7268
74609498
9998
ProchlorococcusSynechococcusKappaphycus alvareziiGracilaria lemaneiformisCeramium boydeniiPolysiphonia boldiiPorphyra purpureaPorphyra teneraPorphyra haitanensisPorphyra yezoensisCorallina officinalisGracilaria tenuistipitataRhodella violaceaAglaothamnion neglectumGriffithsia monilisGloeobacter violaceus
PS
KAGLCBPBPPPTPHPYCOGTRVANGMGV
Guillardia theta-120572
Figure 3 Phylogenetic tree ofKappaphycus alvarezii 12 other red algae and sequenced cyanobacteria based onPENumbers at nodes indicatebranch support given as bootstrap values from neighbor joining (NJ)maximum parsimony (MP) analysis Numbers are only shown if theyexceed 50 respectively
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
0
2
4
6
8
10
12
14
20010050
2510
rETR
0 200 400 600 800Light intensities (120583mol photons mminus2 sminus1)
1 d
(a)
0
2
4
6
8
10
12
14
0 200 400 600 800
rETR
20010050
2510
Light intensities (120583mol photons mminus2 sminus1)
2 d
(b)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
3 d
(c)
0
2
4
6
8
10
12
14
rETR
0 200 400 600 800PAR (120583mol photons mminus2 sminus1)
20010050
2510
4 d
(d)
Figure 2 Rapid light curves (RLCs) of Kappaphycus alvarezii cultivated under different light intensities Indicated data are the mean data ofthree independent experiments (plusmnSD)
cyanobacteria G violaceus and T erythraeum and thenassembled with the PEI and PEII clades of SynechococcusCompared with HL-Prochlorococcus they have a closer rela-tionship with LL-Prochlorococcus Red algal PE sequences aremore similar to LL-Prochlorococcus PEs than to HL-PEs Apossible explanation of this result is that HL-Prochlorococcusevolves faster than LL and also exhibits much higher GCcontent Hence the similarities among red algal- HL-and LL-PEs do not necessarily reflect their photosyntheticproperties and relationships In the 120572-subunit group redalgae successively assembled with PEI of Synechococcus LL-Prochlorococcus and T erythraeum PEII of Synechococcusformed a cluster by itself
33 Tertiary Structure of PE Forty-eight amino acid lociconserved with LL-Prochlorococcus but diverged with HL-Prochlorococcuswere mapped onto 120573-PE tertiary structure ofK alvarezii in blue color (Figure 4(a)) These loci distributeto the following six domains (1) N-terminal 2-10aa domainis obviously conserved including the loci 3D function ofinteraction of N-terminus of 120572- and 120573-subunits (2) 65-81aadomain (except 67I 68A 72N 75T 76N and 79M) is adjacentto themotif of chromophore interaction (3) 92-100aa domain(including 92Y 94S 96A 98L 99A and 100G) in which91R 95Y are functions of 120572- and 120573-interaction and 100G isat the end of helix E (4) 107-115aa domain (including 107D108R 111N 112G and 115E) includes the functional loci 108R
BioMed Research International 5
PCCMP1986-120573-HPAS9601-120573-HPMIT9312-120573-HPMIT9303-120573-HPMIT9313-120573-H
PCCMP1357-III120573-LPMIT9211-120573-L
PNATL2A-III120573-LPNATL2A-120573-L
82728584
9367
9999
7267
9055
66
50
54
65
76
53
50
5853
69
65
9472
9887
9895
9280
6357
98979786
9999
7753
6851
63556956
7876
9788
9794
9892
97999897
9996
7254
9589
8776
9187
10099
10099
10099
10099
KA-120573
120572KA-
GL-120573CB-120573PB-120573
PP-120573PT-120573PH-120573PY-120573
CO-120573GT-120573
RV-120573AN-120573GM-120573
GVPCC7421-120573TEIMS101-120573
SCC9311-I120573SCC9902-I120573SBL107-I120573SRCC307-I120573SRS9916-I120573
SCC9605-I120573SWH8102-I120573
SWH7803-I120573SWH7805-I120573
SCC9311-II120573
SCC9902-II120573SBL107-II120573
SRCC307-II120573SRS9916-II120573
SCC9605-II120573SWH8102-II120573
SWH7803-II120573
PCCMP1357-III120572-LPMIT9211-120572-LPNATL2A-III120572-LPNATL1A-120572-L
PMIT9303-120572-LPMIT9313-120572-L
GL-120572
CB-120572PB-120572
PP-120572PT-120572PH-120572
PY-120572
CO-120572
GT-120572RV-120572
AN-120572GM-120572
SCC9311-I120572SRCC307-I120572SCC9902-I120572
SRS9916-I120572SBL107-I120572SCC9605-I120572SWH8102-I120572
SWH7803-I120572SWH7805-I120572
TEIMS101-120572SCC9311-II120572
SRCC307-II120572SRS9916-II120572
SCC9605-II120572SWH8102-II120572
SBL107-II120572SCC9902-II120572SWH7803-II120572
20
7268
74609498
9998
ProchlorococcusSynechococcusKappaphycus alvareziiGracilaria lemaneiformisCeramium boydeniiPolysiphonia boldiiPorphyra purpureaPorphyra teneraPorphyra haitanensisPorphyra yezoensisCorallina officinalisGracilaria tenuistipitataRhodella violaceaAglaothamnion neglectumGriffithsia monilisGloeobacter violaceus
PS
KAGLCBPBPPPTPHPYCOGTRVANGMGV
Guillardia theta-120572
Figure 3 Phylogenetic tree ofKappaphycus alvarezii 12 other red algae and sequenced cyanobacteria based onPENumbers at nodes indicatebranch support given as bootstrap values from neighbor joining (NJ)maximum parsimony (MP) analysis Numbers are only shown if theyexceed 50 respectively
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
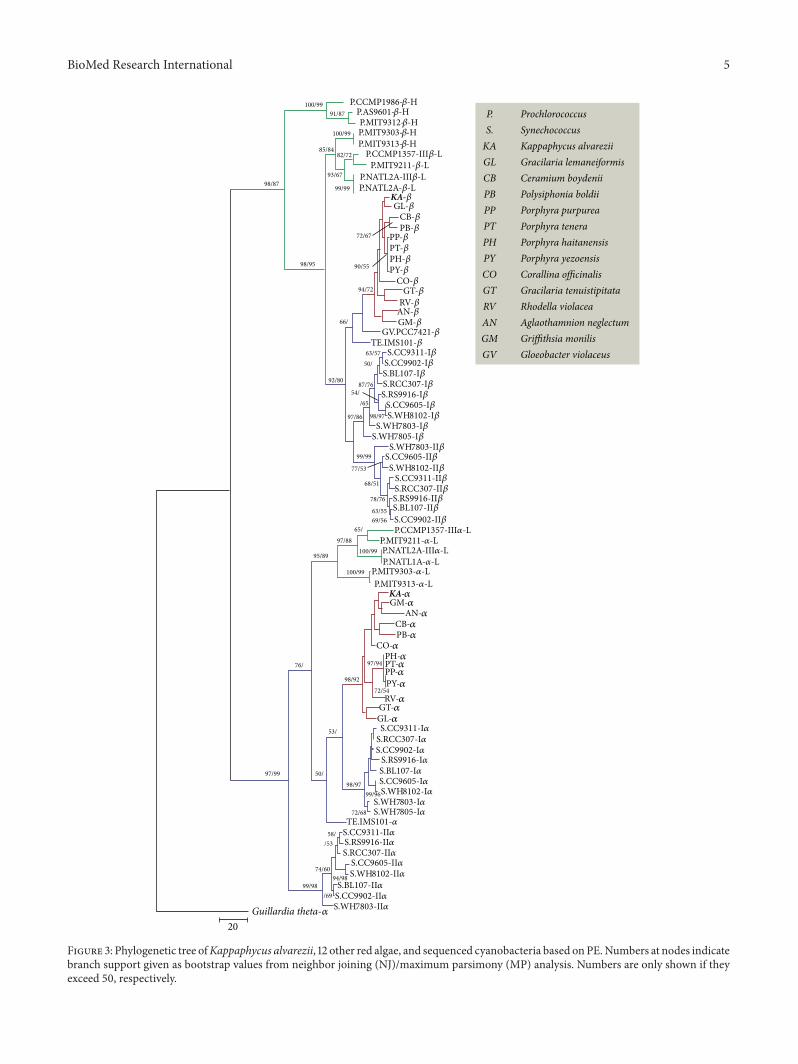
BioMed Research International 5
PCCMP1986-120573-HPAS9601-120573-HPMIT9312-120573-HPMIT9303-120573-HPMIT9313-120573-H
PCCMP1357-III120573-LPMIT9211-120573-L
PNATL2A-III120573-LPNATL2A-120573-L
82728584
9367
9999
7267
9055
66
50
54
65
76
53
50
5853
69
65
9472
9887
9895
9280
6357
98979786
9999
7753
6851
63556956
7876
9788
9794
9892
97999897
9996
7254
9589
8776
9187
10099
10099
10099
10099
KA-120573
120572KA-
GL-120573CB-120573PB-120573
PP-120573PT-120573PH-120573PY-120573
CO-120573GT-120573
RV-120573AN-120573GM-120573
GVPCC7421-120573TEIMS101-120573
SCC9311-I120573SCC9902-I120573SBL107-I120573SRCC307-I120573SRS9916-I120573
SCC9605-I120573SWH8102-I120573
SWH7803-I120573SWH7805-I120573
SCC9311-II120573
SCC9902-II120573SBL107-II120573
SRCC307-II120573SRS9916-II120573
SCC9605-II120573SWH8102-II120573
SWH7803-II120573
PCCMP1357-III120572-LPMIT9211-120572-LPNATL2A-III120572-LPNATL1A-120572-L
PMIT9303-120572-LPMIT9313-120572-L
GL-120572
CB-120572PB-120572
PP-120572PT-120572PH-120572
PY-120572
CO-120572
GT-120572RV-120572
AN-120572GM-120572
SCC9311-I120572SRCC307-I120572SCC9902-I120572
SRS9916-I120572SBL107-I120572SCC9605-I120572SWH8102-I120572
SWH7803-I120572SWH7805-I120572
TEIMS101-120572SCC9311-II120572
SRCC307-II120572SRS9916-II120572
SCC9605-II120572SWH8102-II120572
SBL107-II120572SCC9902-II120572SWH7803-II120572
20
7268
74609498
9998
ProchlorococcusSynechococcusKappaphycus alvareziiGracilaria lemaneiformisCeramium boydeniiPolysiphonia boldiiPorphyra purpureaPorphyra teneraPorphyra haitanensisPorphyra yezoensisCorallina officinalisGracilaria tenuistipitataRhodella violaceaAglaothamnion neglectumGriffithsia monilisGloeobacter violaceus
PS
KAGLCBPBPPPTPHPYCOGTRVANGMGV
Guillardia theta-120572
Figure 3 Phylogenetic tree ofKappaphycus alvarezii 12 other red algae and sequenced cyanobacteria based onPENumbers at nodes indicatebranch support given as bootstrap values from neighbor joining (NJ)maximum parsimony (MP) analysis Numbers are only shown if theyexceed 50 respectively
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
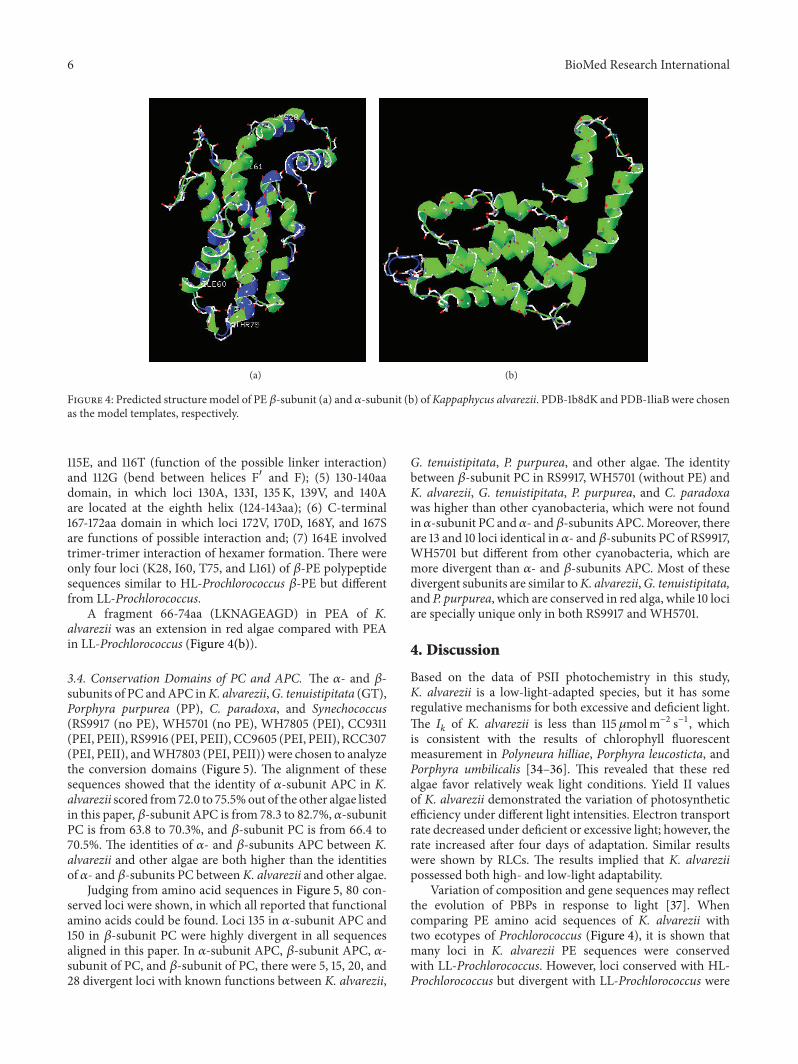
6 BioMed Research International
(a) (b)
Figure 4 Predicted structure model of PE 120573-subunit (a) and 120572-subunit (b) ofKappaphycus alvarezii PDB-1b8dK and PDB-1liaB were chosenas the model templates respectively
115E and 116T (function of the possible linker interaction)and 112G (bend between helices F1015840 and F) (5) 130-140aadomain in which loci 130A 133I 135 K 139V and 140Aare located at the eighth helix (124-143aa) (6) C-terminal167-172aa domain in which loci 172V 170D 168Y and 167Sare functions of possible interaction and (7) 164E involvedtrimer-trimer interaction of hexamer formation There wereonly four loci (K28 I60 T75 and L161) of 120573-PE polypeptidesequences similar to HL-Prochlorococcus 120573-PE but differentfrom LL-Prochlorococcus
A fragment 66-74aa (LKNAGEAGD) in PEA of Kalvarezii was an extension in red algae compared with PEAin LL-Prochlorococcus (Figure 4(b))
34 Conservation Domains of PC and APC The 120572- and 120573-subunits of PC andAPC inK alvareziiG tenuistipitata (GT)Porphyra purpurea (PP) C paradoxa and Synechococcus(RS9917 (no PE) WH5701 (no PE) WH7805 (PEI) CC9311(PEI PEII) RS9916 (PEI PEII) CC9605 (PEI PEII) RCC307(PEI PEII) andWH7803 (PEI PEII)) were chosen to analyzethe conversion domains (Figure 5) The alignment of thesesequences showed that the identity of 120572-subunit APC in Kalvarezii scored from720 to 755out of the other algae listedin this paper 120573-subunit APC is from 783 to 827 120572-subunitPC is from 638 to 703 and 120573-subunit PC is from 664 to705 The identities of 120572- and 120573-subunits APC between Kalvarezii and other algae are both higher than the identitiesof 120572- and 120573-subunits PC betweenK alvarezii and other algae
Judging from amino acid sequences in Figure 5 80 con-served loci were shown in which all reported that functionalamino acids could be found Loci 135 in 120572-subunit APC and150 in 120573-subunit PC were highly divergent in all sequencesaligned in this paper In 120572-subunit APC 120573-subunit APC 120572-subunit of PC and 120573-subunit of PC there were 5 15 20 and28 divergent loci with known functions between K alvarezii
G tenuistipitata P purpurea and other algae The identitybetween 120573-subunit PC in RS9917 WH5701 (without PE) andK alvarezii G tenuistipitata P purpurea and C paradoxawas higher than other cyanobacteria which were not foundin 120572-subunit PC and 120572- and 120573-subunits APCMoreover thereare 13 and 10 loci identical in 120572- and 120573-subunits PC of RS9917WH5701 but different from other cyanobacteria which aremore divergent than 120572- and 120573-subunits APC Most of thesedivergent subunits are similar toK alvareziiG tenuistipitataand P purpurea which are conserved in red alga while 10 lociare specially unique only in both RS9917 and WH5701
4 Discussion
Based on the data of PSII photochemistry in this studyK alvarezii is a low-light-adapted species but it has someregulative mechanisms for both excessive and deficient lightThe 119868119896 of K alvarezii is less than 115 120583molmminus2 sminus1 whichis consistent with the results of chlorophyll fluorescentmeasurement in Polyneura hilliae Porphyra leucosticta andPorphyra umbilicalis [34ndash36] This revealed that these redalgae favor relatively weak light conditions Yield II valuesof K alvarezii demonstrated the variation of photosyntheticefficiency under different light intensities Electron transportrate decreased under deficient or excessive light however therate increased after four days of adaptation Similar resultswere shown by RLCs The results implied that K alvareziipossessed both high- and low-light adaptability
Variation of composition and gene sequences may reflectthe evolution of PBPs in response to light [37] Whencomparing PE amino acid sequences of K alvarezii withtwo ecotypes of Prochlorococcus (Figure 4) it is shown thatmany loci in K alvarezii PE sequences were conservedwith LL-Prochlorococcus However loci conserved with HL-Prochlorococcus but divergent with LL-Prochlorococcus were
BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
APC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
APC-120573
PC-120573
PC-120572KAGTPPCPSRS9917SWH5701
SCC9311SRS9916SCC9605SRCC307SWH7803
KAGTPPCPSRS9917SWH5701SWH7805
SWH7805
SWH7805
SWH7805
SCC9311SRS9916SCC9605SRCC307SWH7803 Highly conserved amino acid positions of which the function has been speculated
Highly divergent amino acid positionsDivergent amino acid in highly conserved positionsDivergent amino acid positions in red algaeIdentical amino acid positions in Synechococcus RS9917 and WH5701
Figure 5 Highly conserved or divergent amino acid loci in APC and PC sequences of Kappaphycus alvarezii
also found The divergent loci were mainly located in 120572-helix and had several reported functional domains (egsubunitsrsquo interaction linkersrsquo interaction and chromophoresrsquointeraction) According to the previous studies [38] PEgenes in LL-Prochlorococcus suffered from positive selectionand the selected loci were related to light-harvesting orenergy-transferring We speculate the mutative domains orloci were related to light responses of K alvarezii Theextension domain near the chromophoresrsquo interaction areain PE-120572 of red algae lacking in LL-Prochlorococcus is alsohypothesized to have an effect on low-light response Furtherexperiments are still needed to prove these assumptionsMoreover by comparing APC and PC (Figure 5) we alsofound that APC sequences are more conserved than PC and120573-subunits are more divergent than 120572-subunits inK alvareziiand other algae Although PBP sequences are divergent inall algae we found that the loci with basic function aremore conserved such as subunitsrsquo interaction linkersrsquo inter-action and chromophoresrsquo interaction The conservationmay ensure PBPsrsquo function in absorption and transfer oflight and energy The diversities of PE and PC may takepart in algal surviving and reviving under different lightconditions
Our study evaluated photosynthetic characteristics ofK alvarezii and attempted to explain light adaptation byanalyzing secondary and tertiary structure of photosyntheticprotein The results of this study would shed some light onunderstanding the potential mechanism of light response ofK alvarezii and other red algae
Acknowledgments
This work was supported by the National Science amp Tech-nology Pillar Program of China (no 2011BAD13B0902)and National Natural Science Foundation of China (no41006094) The authors thank Baosheng Ge Shaofang Liuand Yingjie Chen for providing assistance with the pigmentcomposition measurement
References
[1] P Matanjun S Mohamed K Muhammad and N MMustapha ldquoComparison of cardiovascular protective effects oftropical seaweeds Kappaphycus alvarezii Caulerpa lentilliferaand Sargassum polycystum on high-cholesterolhigh-fat diet inratsrdquo Journal of Medicinal Food vol 13 no 4 pp 792ndash800 2010
8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
[2] S W Chan H Mirhosseini F S Taip T C Ling and C PTan ldquoComparative study on the physicochemical propertiesof kappa-carrageenan extracted from Kappaphycus alvarezii(doty) doty ex Silva in Tawau Sabah Malaysia and commercialkappa-carrageenansrdquo Food Hydrocolloids vol 30 no 2 pp 581ndash588 2013
[3] D JMcHugh ldquoProspects for seaweed production in developingcountriesrdquo FAO Fisheries Circular 986 FAO Rome Italy 2001
[4] M S Bindu and IA Levine ldquoThe commercial red seaweedKap-paphycus alvarezii-an overview on farming and environmentrdquoJournal of Applied Phycology vol 23 no 4 pp 789ndash796 2011
[5] E Aguirre-Von-Wobeser F L Figueroa and A Cabello-PasinildquoPhotosynthesis and growth of red and green morphotypesof Kappaphycus alvarezii (Rhodophyta) from the PhilippinesrdquoMarine Biology vol 138 no 4 pp 679ndash686 2001
[6] E C Schmidt M Maraschin and Z L Bouzon ldquoEffects ofUVB radiation on the carragenophyte Kappaphycus alvarezii(Rhodophyta Gigartinales) changes in ultrastructure growthand photosynthetic pigmentsrdquoHydrobiologia vol 649 no 1 pp171ndash182 2010
[7] L Hayashi G S M Faria B G Nunes et al ldquoEffects of salinityon the growth rate carrageenan yield and cellular structure ofKappaphycus alvarezii (Rhodophyta Gigartinales) cultured invitrordquo Journal of Applied Phycology vol 23 no 3 pp 439ndash4472011
[8] B Loggini A Scartazza E Brugnoli and F Navari-IzzoldquoAntioxidative defense system pigment composition and pho-tosynthetic efficiency in two wheat cultivars subjected todroughtrdquo Plant Physiology vol 119 no 3 pp 1091ndash1099 1999
[9] H-H Kunz M Scharnewski K Feussner et al ldquoThe ABCtransporter PXA1 and peroxisomal 120573-oxidation are vital formetabolism in mature leaves of Arabidopsis during extendeddarknessrdquo Plant Cell vol 21 no 9 pp 2733ndash2749 2009
[10] M T Meyer T Genkov J N Skepper et al ldquoRubisco small-subunit 120572-helices control pyrenoid formation in Chlamy-domonasrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 109 no 47 pp 19474ndash19479 2012
[11] A E Allen C L Dupont M Obornık et al ldquoEvolutionand metabolic significance of the urea cycle in photosyntheticdiatomsrdquo Nature vol 473 no 7346 pp 203ndash207 2011
[12] M L Yallop A M Anesio R G Perkins et al ldquoPhotophysiol-ogy and albedo-changing potential of the ice algal communityon the surface of the Greenland ice sheetrdquo The ISME Journalvol 6 no 12 pp 2302ndash2313 2012
[13] A Lepisto S Kangasjarvi E-M Luomala et al ldquoChloro-plast nadph-thioredoxin reductase interacts with photoperiodicdevelopment inArabidopsisrdquoPlant Physiology vol 149 no 3 pp1261ndash1276 2009
[14] N Nelson and A Ben-Shem ldquoThe complex architecture of oxy-genic photosynthesisrdquo Nature Reviews Molecular Cell Biologyvol 5 no 12 pp 971ndash982 2004
[15] K E Apt J L Collier and A R Grossman ldquoEvolution of thephycobiliproteinsrdquo Journal of Molecular Biology vol 248 no 1pp 79ndash96 1995
[16] X Guan S Qin Z Su et al ldquoCombinational biosynthesis of afluorescent cyanobacterial holo-120572-phycocyanin in Escherichiacoli by using one expression vectorrdquo Applied Biochemistry andBiotechnology vol 142 no 1 pp 52ndash59 2007
[17] B Ge Z Tang F Zhao Y Ren Y Yang and S Qin ldquoScale-up offermentation and purification of recombinant allophycocyaninover-expressed in Escherichia colirdquo Process Biochemistry vol40 no 10 pp 3190ndash3195 2005
[18] C S Ting G Rocap J King and S W Chisholm ldquoCyanobac-terial photosynthesis in the oceans the origins and significanceof divergent light-harvesting strategiesrdquo Trends inMicrobiologyvol 10 no 3 pp 134ndash142 2002
[19] L R Moore G Rocap and S W Chisholm ldquoPhysiology andmolecular phytogeny of coexisting Prochlorococcus ecotypesrdquoNature vol 393 no 6684 pp 464ndash467 1998
[20] C Steglich A F Post and W R Hess ldquoAnalysis of naturalpopulations of Prochlorococcus spp in the northern Red Seausing phycoerythrin gene sequencesrdquo Environmental Microbi-ology vol 5 no 8 pp 681ndash690 2003
[21] Z I Johnson E R Zinser A Coe N P McNulty E M SWoodward and S W Chisholm ldquoNiche partitioning amongProchlorococcus ecotypes along ocean-scale environmental gra-dientsrdquo Science vol 311 no 5768 pp 1737ndash1740 2006
[22] F Zhao and S Qin ldquoComparative molecular population genet-ics of phycoerythrin locus in Prochlorococcusrdquo Genetica vol129 no 3 pp 291ndash299 2007
[23] C Six J-C Thomas L Garczarek et al ldquoDiversity andevolution of phycobilisomes in marine Synechococcus spp acomparative genomics studyrdquo Genome Biology vol 8 no 12article R259 2007
[24] F Zhao X ZhangC Liang JWuQ Bao and SQin ldquoGenome-wide analysis of restriction-modification system in unicellularand filamentous cyanobacteriardquo Physiological Genomics vol 24no 3 pp 181ndash190 2006
[25] X Guan S Qin F Zhao X Zhang and X Tang ldquoPhycobili-somes linker family in cyanobacterial genomes divergence andevolutionrdquo International Journal of Biological Sciences vol 3 no7 pp 434ndash445 2007
[26] J C Hagopian M Reis J P Kitajima D Bhattacharya and MC De Oliveira ldquoComparative analysis of the complete plastidgenome sequence of the red alga Gracilaria tenuistipitata varliui provides insights into the evolution of rhodoplasts and theirrelationship to other plastidsrdquo Journal of Molecular Evolutionvol 59 no 4 pp 464ndash477 2004
[27] K E Apt and A R Grossman ldquoCharacterization and tran-script analysis of the major phycobiliprotein subunit genesfromAglaothamnion neglectum (Rhodophyta)rdquo PlantMolecularBiology vol 21 no 1 pp 27ndash38 1993
[28] K D Pruitt T Tatusova G R Brown andD RMaglott ldquoNCBIReference Sequences (RefSeq) current status new features andgenome annotation policyrdquo Nucleic Acids Research vol 40 noD1 pp D130ndashD135 2012
[29] DA BensonMCavanaugh K Clark et al ldquoGenBankrdquoNucleicAcids Research vol 41 no D1 pp D36ndashD42 2013
[30] J A Kramer ldquoOmigaŮ a PC-based sequence analysis toolrdquoApplied Biochemistry and Biotechnology B vol 19 no 1 pp 97ndash106 2001
[31] C Camacho G Coulouris V Avagyan et al ldquoBLAST+ archi-tecture and applicationsrdquo BMC Bioinformatics vol 10 article421 2009
[32] M A Larkin G Blackshields N P Brown et al ldquoClustalW andClustal X version 20rdquo Bioinformatics vol 23 no 21 pp 2947ndash2948 2007
[33] K Tamura D Peterson N Peterson G Stecher M Nei andS Kumar ldquoMEGA5 molecular evolutionary genetics analysisusing maximum likelihood evolutionary distance and max-imum parsimony methodsrdquo Molecular Biology and Evolutionvol 28 no 10 pp 2731ndash2739 2011
BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
[34] W Nultsch J Pfau and K Huppertz ldquoPhotoinhibition ofphotosynthetic oxygen production and its recovery in thesubtidal red alga Polyneura hilliaerdquo Botanica Acta vol 103 no1 pp 62ndash67 1990
[35] F L Figueroa S Salles J Aguillera et al ldquoEffects of solarradiation on photoinhibition and pigmentation in the red algaPorphyra leucostictardquoMarine Ecology Progress Series vol 151 pp81ndash90 1997
[36] D-P Hader A Groniger C Hallier M Lebert F L Figueroaand C Jimenez ldquoPhotoinhibition by visible and ultravioletradiation in the red macroalga Porphyra umbilicalis grown inthe laboratoryrdquo Plant Ecology vol 145 no 2 pp 351ndash358 1999
[37] R C Everroad and A M Wood ldquoPhycoerythrin evolution anddiversification of spectral phenotype in marine Synechococcusand related picocyanobacteriardquo Molecular Phylogenetics andEvolution vol 64 no 3 pp 381ndash392 2012
[38] F Zhao and S Qin ldquoEvolutionary analysis of phycobiliproteinsimplications for their structural and functional relationshipsrdquoJournal of Molecular Evolution vol 63 no 3 pp 330ndash340 2006
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
Top Related