







Languages
Pages
Legal


Microbial
Metabolism
Chapter 7
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

Metabolism and the Role of Enzymes
•Metabolism: pertains to all chemical reactions and physical
workings of the cell
•Anabolism:
- any process that results in synthesis of cell
molecules and structures
- a building and bond-making process that forms larger
macromolecules from smaller onesmacromolecules from smaller ones
- requires the input of energy
•Catabolism:
- breaks the bonds of larger molecules into smaller
molecules
- releases energy

Simplified Model of Metabolism
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Rel
ativ
e co
mpl
exity
of m
olec
ules
ANABOLISM
ANABOLISM
ANABOLISM
Peptidoglycan
Proteins
CATABOLISM
Glu
Glucose
Macromolecules
Bacterialcell
Buildingblocks
Rel
ativ
e co
mpl
exity
of m
olec
ules
Nutrientsfrom outsideor frominternalpathways
Glycolysis
Krebs cycle
Respiratorychain
Fermentation
Yields energy Uses energy Uses energy Uses energy
Some assemblyreactions occurspontaneously
Complex lipids
RNA + DNA
Peptidoglycan
Amino acids
Sugars
Nucleotides
Fatty acidsGlyceraldehyde-3-P
Acetyl CoA
Pyruvate
Precursormolecules
blocks

Checklist of Enzyme Characteristics

Enzymes: Catalyzing the Chemical Reactions of
Life
•Enzymes
- chemical reactions of life cannot proceed
without them
- are catalysts that increase the rate of chemical
reactions without becoming part of the products
or being consumed in the reaction

How Do Enzymes Work?
•Reactants are converted into products by bond
formation or bond breakage
- substrates: reactant molecules acted on by an
enzyme
Speed up the rate of reactions without increasing the •Speed up the rate of reactions without increasing the
temperature
•Much larger in size than substrates
•Have unique active site on the enzyme that fits only the
substrate

How Do Enzymes Work? (cont’d)
•Binds substrate
•Participates directly in changes to substrate
•Does not become part of the products
•Not used up by the reaction
•Can be used over and over again
•Enzyme speed
- the number of substrate molecules converted
per enzyme per second
- catalase: several million
- lactate dehydrogenase: a thousand

Conjugated Enzyme Structure
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
CoenzymeCoenzyme
Metalliccofactorcofactor
ApoenzymesMetalliccofactor

Enzyme Structure
•Simple enzymes consist of protein alone
•Conjugated enzymes contain protein and nonprotein
molecules
- sometimes referred to as a holoenzyme
- apoenzyme: protein portion of a conjugated
enzyme
- cofactors: either organic molecules called
coenzymes or inorganic elements (metal ions)

Enzyme-Substrate Interactions
•A temporary enzyme-substrate union must occur at the
active site
- fit is so specific that it is described as a “lock-
and-key” fit
•Bond formed between the substrate and enzyme are
weak and easily reversibleweak and easily reversible
•Once the enzyme-substrate complex has formed, an
appropriate reaction occurs on the substrate, often with
the aid of a cofactor
•Product is formed
•Enzyme is free to interact with another substrate

Enzyme-Substrate Reactions
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Substrates
Products
EEnzyme (E)Doesnot fit
(a) (b)
ES complex
(c)

Cofactors: Supporting the Work of Enzymes
•The need of microorganisms for trace elements arises
from their roles as cofactors for enzymes
- iron, copper, magnesium, manganese, zinc,
cobalt, selenium, etc.
•Participate in precise functions between the enzyme •Participate in precise functions between the enzyme
and substrate
- help bring the active site and substrate close
together
- participate directly in chemical reactions with
the enzyme-substrate complex

Cofactors: Supporting the Work of Enzymes
(cont’d)
•Coenzymes
- organic compounds that work in conjunction
with an apoenzyme
- general function is to remove a chemical -group from one substrate molecule and add
it to another substrate molecule
- carry and transfer hydrogen atoms, electrons,
carbon dioxide, and amino groups
- many derived from vitamins

Classification of Enzyme Functions
•Each enzyme also assigned a common name that
indicates the specific reaction it catalyzes- carbohydrase: digests a carbohydrate substrate
- amylase: acts on starch
- maltase: digests maltose
proteinase, protease, peptidase: hydrolyzes the - proteinase, protease, peptidase: hydrolyzes the
peptide bonds of a protein
- lipase: digests fats
- deoxyribonuclease (DNase): digests DNA
- synthetase or polymerase: bonds many small molecules
together

Regulation of Enzyme Function
•Constitutive enzymes: always present in relatively constant amounts regardless of
the amount of substrate
•Regulated enzymes: production is turned on (induced) or turned off (repressed) in
responses to changes in concentration of the substrate
Regulated EnzymesConstitutive Enzymes
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Add moresubstrate.
Enzyme is induced.
or
Enzyme is repressed.
Removesubstrate.
(b)
(a)
Add moresubstrate.
No change inamount of enzyme.

Regulation of Enzyme Function (cont’d)
•Activity of enzymes influenced by the cell’s
environment
- natural temperature, pH, osmotic pressure
- changes in the normal conditions causes
enzymes to be unstable or labile
•Denaturation
- weak bonds that maintain the native shape of
the apoenzyme are broken
- this causes disruption of the enzyme’s shape
- prevents the substrate from attaching to the
active site

Metabolic PathwaysCopyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
A
B
C
U
O
M
N
P
A
B
X
YV
W
Z
Multienzyme Systems
Branched
Convergent
Linear Cyclic
T input
KrebsCycle
S product
Divergent
D
E
O2
O
O1
P
Q
R
M
C
N
ZW
XY
Example:Glycolysis
Example:Amino acidsynthesis
Cycle

Direct Controls on the Action of Enzymes
•Competitive inhibition
- inhibits enzyme activity by supplying a
molecule that resembles the enzyme’s normal
substrate
- “mimic” occupies the active site, preventing
the actual substrate from binding
•Noncompetitive inhibition
- enzymes have two binding sites: the active site
and a regulatory site
- molecules bind to the regulatory site
- slows down enzymatic activity once a certain
concentration of product is reached

Two Common Control Mechanisms for Enzymes
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Competitive Inhibition Noncompetitive Inhibition
SubstrateCompetitiveinhibitor withsimilar shape
Active site
Regulatory site
Normalsubstrate
Both moleculescompete forthe active site.
Enzyme
Regulatory
Enzyme
Reaction proceeds. Reaction is blockedbecause competitiveinhibitor is incapableof becoming a product.
Product
Reaction proceeds. Reaction is blocked becausebinding of regulatory moleculein regulatory site changesconformation of active site sothat substrate cannot enter.
Regulatorymolecule(product)

Controls on Enzyme Synthesis
•Enzymes do not last indefinitely; some wear out, some
are degraded deliberately, and some are diluted with
each cell division
•Replacement of enzymes can be regulated according to
cell demand
•Enzyme repression: genetic apparatus responsible for
replacing enzymes is repressed
- response time is longer than for feedback
inhibition
•Enzyme induction: enzymes appear (are induced) only
when suitable substrates are present

Enzyme RepressionCopyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
1
2
3
6
RNA translated into protein
DNA transcribed into RNA
Protein
Excess product binds to
4
5
7
= +
Excess product binds toDNA and shuts downfurther enzyme production. DNA can not be transcribed;
the protein cannot be made.
Substrate
Folds to form functionalenzyme structure
EnzymeProducts Substrate

The Pursuit and Utilization of Energy
•Cells require constant input and expenditure of usable
energy
•Energy comes directly from light or is contained in
chemical bonds and released when substances are
catabolized or broken down
•Energy is stored in ATP
•Only chemical energy can routinely drive cell
transactions
•Chemical reactions are the universal basis of cellular
energetics

Energy in Cells
•Energy is managed in the form of chemical reactions
that involve the making and breaking of bonds and the
transfer of electrons
•Exergonic reactions release energy, making it available
for cellular workfor cellular work
•Endergonic reactions are driven forward with the
addition of energy
•Exergonic and endergonic reactions are often coupled
so that released energy is immediately put to work

Energy in Cells (cont’d)
•Cells extract chemical energy already present in
nutrient fuels and apply that energy toward useful work
in the cell
•Cells possess specialized enzyme systems that trap the
energy present in the bonds of nutrients as they are energy present in the bonds of nutrients as they are
progressively broken
•During exergonic reactions, energy released by bonds is
stored in high-energy phosphate bonds such as ATP
•ATP fuels endergonic cell reactions

Oxidation and Reduction
•Oxidation: loss of electrons
- when a compound loses electrons, it is oxidized
•Reduction: gain of electrons
- when a compound gains electrons, it is reduced
•Oxidation-reduction (redox) reactions are common in
the cell and are indispensable to the required energy
transformations

Oxidation and Reduction (cont’d)
•Oxidoreductases: enzymes that remove electrons from
one substrate and add them to another
- their coenzyme carriers are nicotinamide
adenine dinucleotide (NAD) and flavin adenine
dinucleotide (FAD) dinucleotide (FAD)
•Redox pair: an electron donor and an electron acceptor
involved in a redox reaction
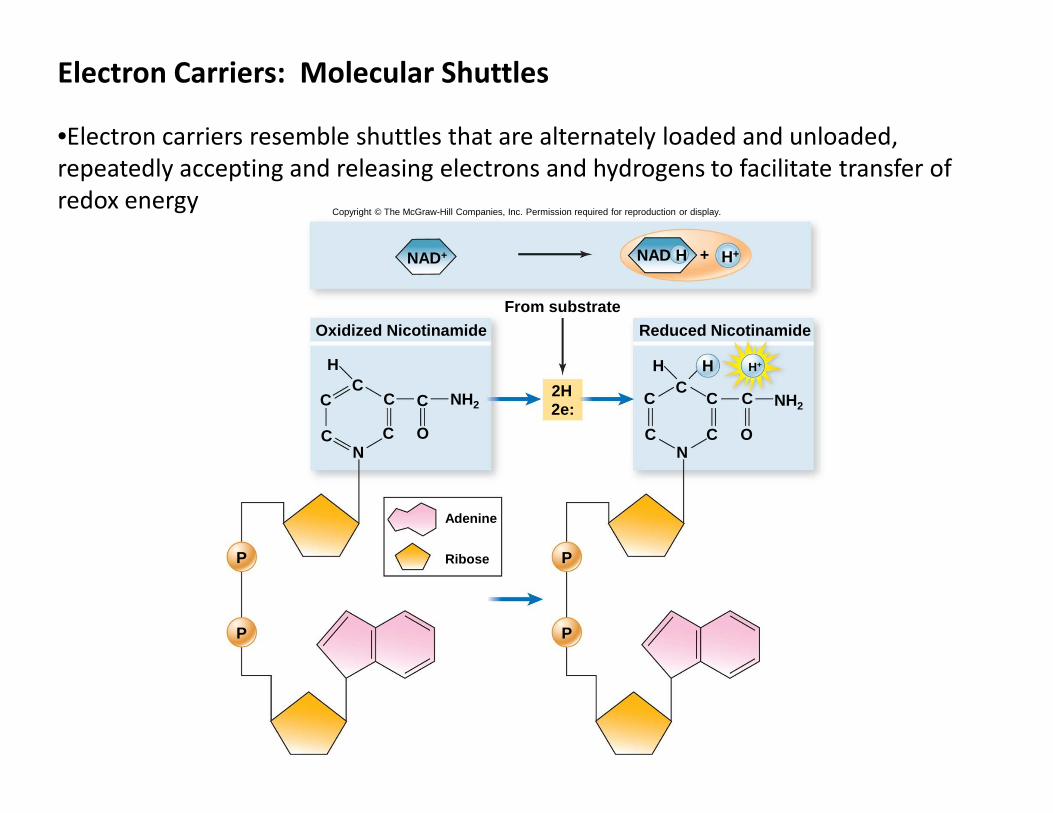
Electron Carriers: Molecular Shuttles
•Electron carriers resemble shuttles that are alternately loaded and unloaded,
repeatedly accepting and releasing electrons and hydrogens to facilitate transfer of
redox energy
H+
H++NAD+ NAD H
Reduced Nicotinamide
From substrate
Oxidized Nicotinamide
NH22H2e:
H
C C CC
H
NH2
H
C C CC
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
P
P
P
P
Adenine
Ribose
NH22e:C
C C
C C
O
NH2C
C C
C C
ON N

ATP: Metabolic Money•Three-part molecule
- nitrogen base (adenine)
- 5-carbon sugar (ribose)
- chain of three phosphate
groups bonded to ribose
- phosphate groups are
N
NN
N N
H H
H
H
Adenine
AdenosineAdenosine
Diphosphate(ADP)
AdenosineTriphosphate
(ATP)
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
- phosphate groups are
bulky and carry negative
charges, causing a strain
between the last two
phosphates
- the removal of the terminal
phosphate releases energy
O
HHH H
O
O
O
O
P O
O
H
HPPHO
OH OH OH
OH
Ribose
OHBond that releasesenergy when broken

The Metabolic Role of ATP
•ATP utilization and replenishment is an ongoing cycle
- energy released during ATP hydrolysis powers
biosynthesis
- activates individual subunits before they are
enzymatically linked together
•Used to prepare molecules for catabolism•Used to prepare molecules for catabolism
•When ATP is utilized, the terminal phosphate is removed
to release energy and ADP is formed
- input of energy is required to replenish ATP
•In heterotrophs, catabolic pathways provide the energy
infusion that generates the high-energy phosphate to form
ATP from ADP

Catabolism
•Metabolism uses enzymes to catabolize organic
molecules to precursor molecules that cells then use to
anabolize larger, more complex molecules
•Reducing power: electrons available in NADH and
FADH2FADH2
•Energy: stored in the bonds of ATP
- both are needed in large quantities for anabolic
metabolism
- both are produced during catabolism

Overview of the Three Main Catabolic Pathways
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
ANAEROBIC RESPIRATION FERMENTATIONAEROBIC RESPIRATION
CO2
NAD H
ATP
CO2
NAD H
ATP
NAD HCO2
Yields 2 ATPs
CO2
NAD H
ATP
NAD HCO2
KrebsCycle
KrebsCycle
Gly
coly
sis
Gly
coly
sis
Gly
coly
sis
Fermentation
ATP ATP
ATPFADH2
Using organiccompounds as
electron acceptor
Electron Transport System Electron Transport System
Alcohols, acids
2 ATPs2–36 ATPs36–38 ATPsMaximum net yield
Yields variableamount ofenergy
Yields 2 GTPsFADH2 ATP
Using O 2 as electron acceptor Using non- O 2 compound as electron acceptor
(So42–, NO3–, CO3
2–)

Glycolysis
•Turns glucose into pyruvate, which yields energy in the pathways that
follow Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Table 7.2
C C C C C C
Fructose-1, 6-diphosphate
C C C C C C
Glycolysis
Energy Lost or Gained
Uses 2 ATPs
Overview Details
Three reactions alter and rearrange the6-C glucose molecule into 6-C fructose-1,6diphosphate.
Glucose
One reaction breaks fructose-1,6-diphosphateinto two 3-carbon molecules.
Five reactions convert each 3 carbon moleculeinto the 3C pyruvate.
Pyruvate is a molecule that is uniquely suited for chemicalreactions that will produce reducing power (which w illeventually produce ATP).
C C CC C C
C C CC C C
Yields 4 ATPs and 2 NADHs
Total Energy Yield: 2 ATPs and2 NADHs
Pyruvate Pyruvate

The Krebs Cycle:
A Carbon and Energy Wheel
•After glycolysis, pyruvic acid is still energy-rich
•The Krebs cycle takes place in the cytoplasm of bacteria and in the
mitochondrial matrix of eukaryotes
- a cyclical metabolic pathway that begins with acetyl CoA,
which joins with oxaloacetic acid, and then participates in
seven other additional transformationsseven other additional transformations
- transfers the energy stored in acetyl CoA to NAD+ and FAD
by reducing them (transferring hydrogen ions to them)
- NADH and FADH2 carry electrons to the electron transport
chain
- 2 ATPs are produced for each molecule of glucose
through phosphorylation

The Krebs Cycle
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Table 7.3
Each acetyl CoA yields 1 GTP, 3 NADHs, In the first reaction, acetyl CoA
C C C
Pyruvate
CC CC CC
Details
The Krebs Cycle
Energy Lost or Gained Overview
Pyruvate
The 3C pyruvate is converted to2C acetyl CoA in one reaction.
Acetyl CoA
Remember: Thishappens twice for
each glucosemolecule that
One CO2 is liberated and one NADH isformed.
Each acetyl CoA yields 1 GTP, 3 NADHs,1 FADH, and 2 CO 2 molecules.
Total Yield per 2 acetyl CoAs:CO2: 4 In the course of seven more
reactions, citrate is manipulatedto yield energy and CO 2 andoxaloacetate is regenerated.
Intermediate molecules on thewheel can be shunted into othermetabolic pathways as well.
In the first reaction, acetyl CoAdonates 2Cs to the 4C moleculeoxaloacetate to form 6C citrate.
Energy: 2 GTPs, 6 NADHs, 2 FADHs
Otherintermediates GTP
CO2
CO2
Yields:3 NADHs1 FADH2
Citrate
Oxaloacetate
Acetyl CoAmolecule that
enters glycolysis.
C C C C
C C C C C C
C CC

The Respiratory Chain:
Electron Transport
•A chain of special redox carriers that receives reduced
carriers (NADH, FADH2) generated by glycolysis and the
Krebs cycle
- passes them in a sequential and orderly fashion
from one to the next
- highly energetic
- allows the transport of hydrogen ions outside
of the membrane
- in the final step of the process, oxygen accepts
electrons and hydrogen, forming water

The Respiratory Chain:
Electron Transport (cont’d)
•Principal compounds in the electron transport chain:
- NADH dehydrogenase
- flavoproteins
- coenzyme Q (ubiquinone)
- cytochromes

The Respiratory (Electron Transport) ChainCopyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Table 7.4
Reduced carriers (NADH, FADH) transfer electrons an d H+ to firstelectron carrier in chain: NADH dehydrogenase.
These are then sequentially transferred to the next four to sixcarriers with progressively more positive reduction potentials.The carriers are called cytochromes. The number of carriers varies,depending on the bacterium.
Simultaneous with the reduction of the electron car riers,protons are moved to the outside of the membrane, c reating aconcentration gradient (more protons outside than i nside thecell). The extracellular space becomes more positiv ely chargedand more acidic than the intracellular space. This conditioncreates the proton motive force, by which protons f low down theconcentration gradient through the ATP synthase emb edded in themembrane. This results in the conversion of ADP to ATP.
The Respiratory (Electron Transport) Chain
H+
H+
H+
H+
ATPsynthase
Once inside the cytoplasm, protons combine with O 2 toform water (in aerobic respirers [left]), and with a variety ofO-containing compounds to produce more reduced comp ounds.
Anaerobic respiration yields less per NADH and FADH .
Aerobic respiration yields a maximum of 3 ATPs peroxidized NADH and 2 ATPs per oxidized FADH.
Anaerobicrespirers
Aerobicrespirers
CytoplasmH2O NO2
– HS–
O2
H+
CellmembraneWith ETS
Cell wallH+
H+
H+
H+
H+
H+
H+H+
H+
H+
Cytochromes
NAD H
ATPADP
synthase
NO3–
SO42–

The Electron Transport Chain (cont’d)
•Electron transport carriers and enzymes are embedded in the cell
membrane in prokaryotes and on the inner mitochondrial membrane in
eukaryotes Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Intermembranespace
CristaeH+ ions

The Electron Chain (cont’d)
•Released energy from electron carriers in the electron
transport chain is channeled through ATP synthase
•Oxidative phosphorylation: the coupling of ATP
synthesis to electron transportsynthesis to electron transport
- each NADH that enters the electron transport
chain can give rise to 3 ATPs
- Electrons from FADH2 enter the electron
transport chain at a later point and have less
energy to release, so only 2 ATPs result

The Terminal Step
•Aerobic respiration
- catalyzed by cytochrome aa3, also known as
cytochrome oxidase
- adapted to receive electrons from cytochrome c,
pick up hydrogens from solution, and react with
oxygen to form water
2H+ + 2e- + ½ O2 � H20

The Terminal Step (cont’d)
•A potential side reaction of the respiratory chain is the
incomplete reduction of oxygen to the superoxide ion
(O2-) and hydrogen peroxide (H2O2)
•Aerobes produce enzymes to deal with these toxic
oxygen products
- superoxide dismutase- superoxide dismutase
- catalase
- Streptococcus lacks these enzymes but still
grows well in oxygen due to the production of
peroxidase

The Terminal Step (cont’d)
•Anaerobic Respiration
- the terminal step utilizes oxygen-containing ions,
rather than free oxygen, as the final electron
acceptor
Nitrate reductase
�
NO3- + NADH �NO2
- + H2O + NAD+
•Nitrate reductase catalyzes the removal of oxygen from
nitrate, leaving nitrite and water as products

Anaerobic Respiration (cont’d)
•Denitrification
- some species of Pseudomonas and Bacillus
possess enzymes that can further reduce
nitrite to nitric oxide (NO), nitrous oxide (N2O),
and even nitrogen gas
- important step in recycling nitrogen in the - important step in recycling nitrogen in the
biosphere
•Other oxygen-containing nutrients reduced
anaerobically by various bacteria are carbonates and
sulfates
•None of the anaerobic pathways produce as much ATP
as aerobic respiration

After Pyruvic Acid II: Fermentation
•Fermentation
- the incomplete oxidation of glucose or other
carbohydrates in the absence of oxygen
- uses organic compounds as the terminal
electron acceptors
- yields a small amount of ATP- yields a small amount of ATP
- used by organisms that do not have an electron
transport chain
- other organisms repress the production of
electron transport chain proteins when oxygen is
lacking in their environment to revert to
fermentation

Fermentation (cont’d)
•Only yields 2 ATPs per molecule of glucose
•Many bacteria grow as fast as they would in the
presence of oxygen due to an increase in the rate of
glycolysis
•Permits independence from molecular oxygen•Permits independence from molecular oxygen
- allows colonization of anaerobic environments
- enables adaptation to variations in oxygen
availability
- provides a means for growth when oxygen
levels are too low for aerobic respiration

Fermentation (cont’d)
•Bacteria and ruminant cattle
- digest cellulose through fermentation
- hydrolyze cellulose to glucose
- ferment glucose to organic acids which are absorbed
as the bovine’s principal energy source
Human muscle cells•Human muscle cells
- undergo a form of fermentation that permits short
periods of activity after the oxygen supply has been
depleted
- convert pyruvic acid to lactic acid, allowing
anaerobic production of ATP
- accumulated lactic acid causes muscle fatigue

Fermentation
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Table 7.5
Pyruvic acid from glycolysis can itself become the electronacceptor.
Pyruvic acid can also be enzymatically altered and then serve asthe electron acceptor.C CH
H
H
C C C
CO2
Pyruvic acid
Remember: Thishappens twice for
each glucosemolecule that
enters glycolysis.
Fermentation
The NADs are recycled to reenter glycolysis.
The organic molecules that became reduced in their role aselectron acceptors are extremely varied, and often yield usefulproducts such as ethyl alcohol, lactic acid, propio nic acid,butanol, and others.
OH
CC C
H
H
H
H
O
C C
H
H
H
H
H
Lactic acid
OH
OH
NAD+
Ethyl alcohol
OH
Acetaldehyde
NAD H NAD H

Products of Fermentation in Microorganisms
•Alcoholic beverages: ethanol and CO2
•Solvents: acetone, butanol
•Organic acids: lactic acid, acetic acid
•Vitamins, antibiotics, and hormones
•Large-scale industrial syntheses by microorganisms
often utilize entirely different fermentation mechanisms
for the production of antibiotics, hormones, vitamins,
and amino acids

Catabolism of Noncarbohydrate Compounds
•Complex polysaccharides broken into component
sugars, which can enter glycolysis
•Lipids broken down by lipases
- glycerol converted to dihydroxyacetone
phosphate, which can enter midway into phosphate, which can enter midway into
glycolysis
- fatty acids undergo beta oxidation, whose
products can enter the Krebs cycle as acetyl CoA

Catabolism of Noncarbohydrate Compounds
(cont’d)
•Proteins are broken down into amino acids by
proteases
- amino groups are removed through - amino groups are removed through
deamination
- remaining carbon compounds are converted
into Krebs cycle intermediates

Amphibolic Pathways of Glucose MetabolismCopyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Table 7.6
Intermediates from glycolysis are fed into the amin oacid synthesis pathway. From there, the compounds a reformed into proteins. Amino acids can then contribu tenitrogenous groups to nucleotides to form nucleic a cids.
Glucose and related simple sugars are made intoadditional sugars and polymerized to form complexcarbohydrates.
The glycolysis product acetyl CoA can be oxidized t o formfatty acids, critical components of lipids.
Catabolic PathwaysIn addition to the respiration and fermentation pat hwaysalready described, bacteria can deaminate amino aci ds,which leads to the formation of a variety of metabo lic
CA
TA
BO
LIS
MA
NA
BO
LIS
M
Amphibolic Pathways of Glucose Metabolism
Anabolic Pathways
Beta oxidationDeamination
GLUCOSE
Building block
Macromolecule
Cellstructure
Membranesstorage
Cell wallstorage
Enzymes/Membranes
Chromosomes
Lipids/Fats
Starch/CelluloseProteinsNucleic
acids
Fatty acidsCarbohydratesAmino acidsNucleotides
intermediates, including pyruvate and acetyl CoA.
Also, fatty acids can be oxidized to form acetyl Co A.C
AT
AB
OLI
SM
Gly
coly
sis
Metabolicpathways
Simplepathways
Pyruvic acid
Acetyl coenzymeA
KrebsCycle
NH3 H2O
CO2
Top Related