







Languages
Pages
Legal


HERBIVORY OF THE COMMON BRUSHTAIL POSSUM
(TRICHOSURUS VULPECULA, MARSUPIALIA: PHALANGERIDAE)
AT DIFFERENT SCALES OF RESOURCE HETEROGENEITY
KAROLINA PETROVIĆ
Master of Science (Biology)
A thesis submitted in fulfilment of the requirements for the degree of
Doctor of Philosophy
September 2014
Faculty of Science
School of Environmental Sciences

Table of Contents
Certificate of Authorship…………………………………………………………………1
Acknowledgments………………………………………………………………………...2
Abstract…………………………………………………………………………………...5
Chapter 1 A world on a plate: Heterogeneity in ecology and animal nutrition……………………...7
General introduction and thesis overview………………………………………………...8
References……………………………………………………………………………….22
Chapter 2 Dietary composition of the common brushtail possum (Trichosurus vulpecula) in a
heterogeneous landscape: Choice determinants at an “all-you-can-eat buffet”………………………. .27
Introduction……………………………………………………………………………..28
Aims and hypotheses…………………………………………………………………….32
Study sites……………………………………………………………………………….33
Methods…………………………………………………………………………………39
Data analysis……………………………………………………………………………..48
Results…………………………………………………………………………………...56
Discussion……………………………………………………………………………….66
References……………………………………………………………………………….78
Chapter 3 Home range attributes of the common brushtail possum (Trichosurus vulpecula):
How does individual variability in resource use, food, and shelter influence spatial patterns? ………….87
Introduction……………………………………………………………………………..88
Aims and hypotheses…………………………………………………………………….92
Study sites……………………………………………………………………………….93
Methods…………………………………………………………………………………95
Data analysis…………………………………………………………………………....102
Results………………………………………………………………………………….106
Discussion……………………………………………………………………………...128
References……………………………………………………………………………...133

Chapter 4 Dietary choices of the common brushtail possum (Trichosurus vulpecula) in a chemically
complex environment: When not to take a green smoothie? ….………………………………….138
Introduction……………………………………………………………………………139
Aims and hypotheses…………………………………………………………………...144
Methods………………………………………………………………………………..145
Data analysis…………………………………………………………………………....159
Results………………………………………………………………………………….160
Discussion……………………………………………………………………………...180
References……………………………………………………………………………...187
Chapter 5 Herbivory in the heterogeneous world……………………………………………..194
Synthesis of results and conclusions……………………………………………………195
References……………………………………………………………………………...202

1
Certificate of Authorship
I hereby declare that this submission is my own work and that, to the best of my
knowledge and belief, it contains no material previously published or written by another
person nor material which to a substantial extent has been accepted for the award of any
other degree or diploma at Charles Sturt University or any other educational institution,
except where due acknowledgment is made in the thesis. Any contribution made to the
research by colleagues with whom I have worked at Charles Sturt University or elsewhere
during my candidature is fully acknowledged. I agree that this thesis be accessible for the
purpose of study and research in accordance with the normal conditions established by the
Executive Director, Library Services or nominee, for the care, loan and reproduction of
theses.
……………………………..
September 2014

2
Acknowledgments
In 2009, Gardner in her seminal work, “The Devolvement of Doctoral Students: Phases of
Challenge and Support,” proposed a conceptual framework for doctoral student identity
development, which is grounded in the idea of growth through challenges and the
provision of support. While my growth through challenges still awaits some empirical
testing, the endless support of my supervisors, colleagues, family, doctors, and friends is an
unquestionable fact.
I am deeply grateful to my supervisors, Assoc. Prof. David M. Watson and Assoc. Prof. Ian
Lunt, for assisting me on my rocky road to academic excellence and professional
fulfilment. You have oriented and supported me with promptness and care and have
always been patient and encouraging in times of new ideas and personal difficulties.
Dave, thank you for giving me this amazing opportunity of living and studying in Australia,
teaching me along the way how to be an independent researcher, critical thinker, and
writer. Ian, thank you for teaching me how to be a logical and organized scientist,
supporting me in all my moments of self-doubt and despair. Without you guys I would not
be here and that is an unquestionable fact!
I am very grateful to my dear colleagues, Simon McDonald and Deanna Duffy, from
Charles Sturt University’s Spatial Analysis Unit, who assisted me in the gruelling task of
data analysis allowing me to see ecological patterns in numbers and maps that my artsy
brain and a short-sighted naturalist eye could not see. I am also sincerely grateful to
Dr Min An from Charles Sturt University’s Environmental and Analytical Laboratories for
helping me get a Charles Sturt University research grant and bringing me all these sweet,
little gifts from China. Also, I am thankful to a talented, young Polish statistician Grzegorz
Golonka, who came up with a novel way of measuring mistletoe biomass.
I have been very privileged to get to know and collaborate with many great scientists
around Australia and learn from their research and experience. I am immensely grateful to
Dr Katherine Handasyde from the University of Melbourne for generously sharing her
practical knowledge of Aussie wildlife and letting me stay in her research station, where
I had a chance to hang around a bunch of awesome possum researchers.
Moreover, it has been a great honour to work with marsupial–eucalypt chemistry pioneers,
Prof. William Foley and Dr Ian Wallis, from the Australian National University.

3
Their technical excellence and tremendous grasp of experimental and theoretical problems
had a great impact on my thinking of herbivory.
Also, I have to acknowledge a great job done by Hugh McGregor, who analysed my
possum faecal samples, and the incredible tree climbing Kenyon brothers from Tree
Tactics, who collected leaf samples for me. Also, I am very grateful to all Charles Sturt
University undergrad and postgrad volunteers, who made my research easier and kept me
company during those long night hours spent chasing possums in the bush. I would also
like to thank landholders in the Strathbogie Ranges that kindly provided access to their
properties, shared their possum stories, and invited me into their homes. You guys really
made me feel a part of the community once when we got to the subject of local gossip.
Finally, I would like to thank my dear family and friends for their never ending faith in me,
constant encouragement, and support. I am incredibly grateful to my beautiful and smart
Polish mother, Katarzyna Piórkowska, for all her acts of kindness, generosity, and love for
me, standing beside me in my darkest hours, and for letting me hear true words of wisdom
from her partner, Salomon Cymerman. I am also immensely grateful to my loving Serbian
father, Ivan Petrovic, who came all way from Europe to help me survive in the Aussie bush
and drove me crazy with his fear of snakes and my driving.
Naming here all my beautiful Aussie, Kiwi, Indian, Indonesian, Polish, and Serbian friends
that supported and entertained me through all these years would take at least a page, but
you guys already know you are in my heart forever. I would especially like to thank here my
dear friends, Anna and Tom, who generously shared their home during the last stage of my
PhD, cooked me amazing food, and hooked me on Taiwanese pineapple cakes.
Last but not the least, I would like to thank all the staff from Karinya Clinic in Albury,
especially Dr. Paresh Ramjee, for giving me personal wisdom, strength, and hope that there
is always light at the end of the tunnel.
During my candidature I was supported by the Australian Government Endeavour Europe
Award and the International Postgraduate Research Scholarship, as well as the Charles
Sturt University Postgraduate Research Scholarship. I wish to express here my immense
gratitude to my supervisor, Assoc. Prof. David M. Watson, and Charles Sturt University’s
Research Office for their ongoing technical and material support during periods when I
experienced health problems.

4
Operational funding and resources for this research were generously provided by the
following organizations, to which I express my gratitude: Charles Sturt University’s
Postgraduate Operating Funds, Charles Sturt University Competitive Grant, and
Holsworth Wildlife Research Endowment. I am also very thankful to my generous Polish
grandmother, Helena Piórkowska, for leaving me an inheritance so I could support myself
during my candidature and concentrate fully on my PhD. The list could go on and on . . .
I love you all and, once again, thank you for your help and support through my phases of
challenge and personal growth.

5
Abstract
Resource heterogeneity has been a broadly discussed concept in ecology used to explain
adaptive responses of herbivores to the distribution, abundance, and nutritional quality of
forage, as well as the patchy distribution of other key resources (i.e., shelter) and
interactions with competitors and predators. However, few studies have considered that
generalist herbivores exhibit resource selectivity at different scales of environmental
heterogeneity. Australian arboreal marsupials represent a unique opportunity for exploring
herbivore nutrition as they represent a continuum from specialist to generalist herbivores,
live in habitats dominated by a single tree genus (Eucalyptus) of varying nutritional quality,
and rely on trees as a shelter and platform for facilitating interactions with conspecifics,
competitors, and predators.
In this study, resource selectivity of a generalist arboreal marsupial, the common brushtail
possum (Trichosurus vulpecula, Marsupialia: Phalangeridae), was examined in the context of
forage availability and nutritional quality at three different scales, landscape, home range,
and individual tree, with tree-dependant parasitic plants representing a special case of small-
scale resource heterogeneity. Furthermore, other factors associated with tree use by
common brushtail possums were explored, including availability of shelter (tree hollows)
and presence of competitors, the mountain brushtail possum (Trichosurus cunninghami).
Integrative field and laboratory techniques were employed, including spotlighting, radio-
tracking, and faecal content analysis. The chemical variability of trees and parasitic plants
was studied using a recently developed method for estimating the nutritional quality of
forage for herbivores or foliar concentrations of available nitrogen.
The present study has demonstrated that common brushtail possums had a broad diet,
including the most common, but chemically defended and hard-to-digest Eucalyptus species;
nitrogen-rich Acacia species; succulent parasitic plants, drooping mistletoe (Amyema pendula)
and cherry ballart (Exocarpos cupressiformis); and supplemented their diet with other plant
groups. However, animals were selective over multiple scales of resource heterogeneity,
ranging from across a landscape to individual home ranges to specific species of trees,
individual trees, and discrete patches of mistletoes within tree canopies. They preferred
parasitic plants over their tree hosts and selectively foraged on trees parasitized by
mistletoes. They used Eucalyptus species less than expected from their availability in the
habitat, and were found foraging on both Monocalyptus and Symphyomyrtus species of
differing nutritional quality.

6
Moreover, tree choices and space-use patterns of common brushtail possum were shaped
by the availability of hollows and the presence of mountain brushtail possums.
The current research has shown that studying herbivory over different spatial scales,
landscape, home range, and individual plants, allows general conclusions to be made about
the nature of herbivore nutrition, focusing not only on food availability and nutritional
quality, but also on a broader context of ecological interactions with conspecifics and
competitors. Moreover, this study has shown that focusing on a single measure of the
nutritional quality of forage for herbivores, available nitrogen, simplifies complex
interactions between multiple nutrients and herbivore-deterrents. Therefore, future studies
should encourage a more holistic approach when analysing availability and nutritional
quality of forage for herbivores.

7
CH
AP
TE
R
1
A world on a plate: Heterogeneity in ecology and animal nutrition

8
CHAPTER 1
A world on a plate: Heterogeneity in ecology and animal nutrition
Introduction
Plant–animal interactions
Antagonistic relationships between plants and animal consumers (herbivores) are one of
the most important ecological interactions in nature (Crawley, 1983; Huntly, 1991; Karban
& Baldwin, 1997). Plants and herbivores represent three-quarters of earth’s biodiversity and
biomass, ranging from microscopic phytoplankton and zooplankton to some of the largest
organisms on earth, the giant sequoia (Sequoiadendron giganteum) and bush elephant
(Loxodonta africana). Herbivores consume around 15% of the plant biomass produced
annually in temperate and tropical ecosystems, making herbivory the most common mode
of animal foraging and the major agent by which energy enters food webs (Agrawal, 2011;
Cyr & Face, 1993).
During approximately 410 million years of shared evolutionary history, plants have
developed an arsenal of physical and chemical defences against herbivores. Consequently,
plant defences have evolved due to long-term selection pressure by herbivores (Agrawal &
Fishbein, 2006; Johnson, 2011). Furthermore, plants respond to short-term threats
imposed by herbivores and can change their metabolic pathways by investing in the
production of chemical defences at the time of attack (Karban & Myers, 1989; Ohgushi,
2005). These induced defences can increase the resistance of the plant to herbivore attack
by reducing the preference for, or effect of, herbivores on the damaged plant. Some plants
harbor many natural enemies, whereas others just a few. Hence, the intensity of selection—
pairwise or diffuse—will depend upon the reliability of attack from a diverse array of
potential herbivores (Ehrlich & Raven, 1964; Janzen, 1980).
For herbivores consumption of plants is a great ecological advantage as plants are the most
abundant food in terrestrial and aquatic ecosystems (Cyr & Face, 1993; Wiegert & Owen,
1971). However, plants are a challenging food source for herbivores as reviewed by Moles
and colleagues (2013). Specifically, plants are considered to be of poor nutritional quality
(low in protein, carbohydrates, and other essential nutrients).

9
Moreover, plants possess physical defences that limit herbivore access and foliage
processing (e.g., hard cuticles, spines, hairs, and latex). Finally, plants have developed a
range of chemical defences that reduce the nutritional quality of the foliage, are hard to
digest (e.g., tannins and fibre), and can be potentially toxic to herbivores (e.g., different
groups of plant secondary metabolites).
Plant secondary metabolites, commonly known as toxins, impose high metabolic costs on
herbivores (Freeland & Janzen, 1974). Upon ingestion of food, plant toxins enter the
animal via absorption from gastrointestinal tract and exert their effects via lipophilicity.
Toxins can cause disruption of cell membrane functions, which lead to the induction of
cysts and tissue damage and can lead ultimately to animal death (Iason, 2005; Scheline,
1991). As a result, herbivore’s ingestion of a particular food type is limited by its ability to
detoxify and excrete specific plant secondary metabolites (Freeland & Janzen, 1974).
Furthermore, plant toxicity is closely linked to the potency of plant secondary metabolites
or the amount and type of toxins that are required to produce a given intoxicating effect on
herbivores. According to Iason and Villalba (2006), all plant secondary metabolites act on
herbivores in a dose-dependent fashion, although their functions, thresholds, and
maximum tolerable doses vary greatly depending on the species of animal.
Some plant secondary metabolites can act as both toxins and digestibility reducers and, in
some instances, as pharmaceuticals exerting beneficial effects on herbivore digestion
(Barbehenn & Constabel, 2011). For instance, tannins—the most ubiquitous plant
secondary metabolites in nature—are known to be toxic if present in high concentrations
for both invertebrate and vertebrate species (Aerts, Barry, & McNabb, 1999). Furthermore,
tannins can reduce the digestibility of forage for herbivores through their protein-binding
capacity or inhibition of enzymatic degradation (Iason, 2005). Nevertheless, the same
process that inhibits digestive protein degradation in ruminants can help ruminants obtain
more essential amino acids through the formation of protein complexes limiting
degradation of protein by rumen bacteria (Min et al., 2003). A new discipline of
Pharm-ecology recognizes the importance of both nutrients and plant secondary
metabolites (toxins and pharmaceuticals) in plant–herbivore interactions (Raubenheimer &
Simpson, 2009; Forbey et al., 2013).

10
To overcome chemical defences imposed by plants, herbivores have developed a series of
anatomical, physiological, and behavioural counter-adaptations in an evolutionary “arms
race” (Brodie III & Brodie Jr, 1999; Mello & Silva-Filho, 2002; Rhoades, 1985). For
instance, to sustain themselves in nutritionally poor environments, herbivores have
increased their body size and gut capacity, lowered their nutrient requirements in
comparison to predatory animals, and developed specialized dentition and detoxification
mechanisms to deal with tough and toxic foliage (Freeland & Janzen, 1974; Karasov &
Diamond, 1988; Van Soest, 1994). Furthermore, herbivores have developed broad diets to
avoid over-ingestion of specific toxins and to prevent overloading of individual
detoxification pathways (Dearing, Mangione, & Karasov, 2000; Marsh et al., 2006). Finally,
to maximize their nutrient acquisition herbivores have developed food preferences by
selecting foliage of particular groups (monocots vs. dicots), favouring certain species or
individuals of the same species, and even choosing individual plant parts that are of higher
nutritional quality (Crawley, 1983; Marquis, 1992).
Herbivory at different scales of resource heterogeneity
Herbivores are known to be selective and alter their movements and foraging behaviour
across different scales of resource heterogeneity, ranging from geographic regions and
landscapes to plant communities, species and individuals (Kotliar & Wiens, 1990; Senft et
al., 1987; Fig. 1). According to Senft et al. (1987), herbivores at a regional scale select
different landscapes based on their geophysical properties (i.e., geomorphology, proximity
to waterways, regional climate), as well as availability and distribution of resources (i.e.,
food and shelter), and adjust their life strategies accordingly from sedentary (home ranging)
to migratory. At a landscape scale, herbivores select foraging areas or plant communities
based on their availability, habitat properties (substrate, topography, and microclimate), and
presence of predators, competitors, and conspecifics. Finally, at a community scale,
herbivores select species, individuals, and plant parts based on their availability or
nutritional quality or both.
The concept of resource selectivity was first introduced by Johnson (1980), who focused
on comparing resource availability with resource usage by animal consumers and
introduced a classification system for defining resource selectivity. He described resource
abundance as the resource quantity in the environment measured independently of the
consumer and distinguished resource availability as the accessibility of the resource to the
consumer.

11
Resource availability is compared with resource usage, or the amount consumed by an
animal in a fixed period of time, to generate an overall understanding of the degree of
resource selectivity, by which the consumer selects a resource disproportionately to its
availability. Based on the concept of resource selectivity, the foraging preferences of
herbivores and other groups of consumers can be established on the premise that all foods
are offered on equal basis within a spatially defined area. The concept of resource selectivity
was further extended to incorporate foraging on different spatial scales (i.e., within
different habitats and animals’ home ranges) and across landscapes and regions (Searle et
al., 2005; Senft et al., 1987).
Furthermore, to recognize the hierarchical nature of resource selectivity, Johnson (1980)
introduced the concept of selection order. First-order selection is defined as the selection
of the physical and geographical range of a species. Within that distributional range,
second-order selection determines the home ranges of individuals and the location of social
groups. Third-order selection refers to the usage of different habitat components by
animals within their home ranges, of which fourth-order selection incorporates differential
usage of food items from those available. Hence, resource selectivity is a hierarchically
structured process (see O'Neill, Johnson, & King, 1989), where processes at different
spatial scales can influence the selection of a resource at one spatial scale. Therefore, it is
critical to test for selection at the appropriate scale(s) at which organisms select resources
and to identify how selection processes vary across scales.

12
Fig. 1 Different scales of resource selectivity by herbivores while foraging

13
Herbivory theories: Past, present and future
The effect of food availability on resource selection by herbivores (known as frequency
dependence sensu Johnson, 1980) incorporates at least two inconsistent theoretical
assumptions as recognized by Di Stefano and Newell (2008). The first assumption is
quantitative and implies the existence of a positive relationship between selection and
relative availability of resources in the habitat (positive frequency dependence). This
assumption is consistent with diet specialization as predicted by the optimal foraging theory
(Pyke, 1984; Pyke, Pulliam, & Charnov, 1977; Table 1). The second assumption is
qualitative and proposes that mixed diets are to be expected if consumption is constrained
by nutrient availability (the partial preference theory; Pulliam, 1975; Westoby, 1978) or if
eating a range of foods reduces the negative effects of plant toxins and other hard-to-digest
compounds (the detoxification limitation theory; Freeland & Janzen, 1974; Marsh et al.,
2006). In this case, a negative relationship between selection and relative availability is to be
expected (negative frequency dependence).The observed negative frequency dependence
can be linked to dietary constraints of herbivores.
Namely, to maximize nutrient intake, dietary preferences of herbivores are non-linearly
related to food availability and nutritional quality and rely also on animal sensory cues
(Forbes, 1998). Forage palatability can be defined as the outcome of herbivores’ specific
foraging behavior and involves complex interactions between the current nutritional state
of herbivores, their past experience with forage, and their perception of the trade-offs
between detrimental aspects of forage (i.e., toughness and toxicity of plant secondary
metabolites) against its nutritional quality (macro- and micro-nutrients). According to Iason
and Villalba (2006), the extent to which these processes determine foraging choices of
herbivores is limited by the ability of animals to experience consequences of their behaviors
and associate particular cues in foods with their specific effects in the body.
For example, Provenza (1996) argues that ruminants possess a degree of nutritional
wisdom in the sense that they generally select foods that meet their nutritional needs and
avoid foods that cause toxicosis. The author postulates that neurally mediated interactions
between the senses (i.e., taste and smell) and the viscera enable ruminants to sense the
consequences of food ingestion. Satiety occurs when animals ingest adequate kinds and
amounts of nutritious foods and acquire preferences (mild to strong) for foods that cause
satiety.

14
Unpleasant feelings of physical discomfort (i.e., malaise) are caused by excesses of nutrients
and toxins and by nutrient deficits in food; animals can acquire aversions to those types of
foods that cause this discomfort (the satiety hypothesis; Provenza et al., 2003).
Recently, Simpson and Raubenheimer (2001) have developed the Geometric Framework
for nutrition to deal with the complexity of nutrient–toxin interactions and their impacts
on herbivore foraging behavior and post-ingestive responses. In this framework both
nutrients and toxins can be accommodated as dimensions in a simple geometrical model
with differing animal intake targets and metabolic costs enabling a relatively simple
comparison between observed and predicted patterns of food consumption. The focus of
this framework is on observing animal free food choices when faced with diverse dietary
options scattered throughout a multi-dimensional space and assessing their rule of
compromise when inhibited from reaching the preferred intake target.
Finally, along with food availability and nutritional or toxic constraints, the intraspecific
and interspecific competition over limited food resources and the risk of predation have
been recognized as important factors shaping foraging patterns of herbivores (Lima & Dill,
1990; Chase et al., 2002). Generally, herbivores with overlapping diets exhibit greater
competitive interactions than species that partition resources due to competitive
displacement in the past and niche separation (Stewart et al., 2002). Predation can reduce
the intensity of competitive interactions among herbivores (Gurevitch, Morrison, &
Hedges, 2000), but can also represent an additional cost for foraging animals (Nersesian,
Banks, & McArthur, 2011). Namely, the predation risk alters how long herbivores are
prepared to forage in certain areas and how long animals spend in risky areas if forage is of
better nutritional quality. Hence, it has been recognized that herbivores are able to quantify,
compare, and balance these different, but proximate costs, altering their foraging behaviour
in the process.

15
Table 1 Key theories explaining foraging behaviour of herbivores
Theory description
Foraging consequences
Evolutionary consequences
Key references
Optimal foraging theory
The optimal diet maximizes the net
rate of energy or mass intake while
minimising food searching and
handling time.
Behavioural and physiological adaptations
to forage of low nutritional quality, ability
to utilize nutrients from a single species
more efficiently.
Increasing availability of a preferred
food in a habitat should lead to diet
specialisation as less preferred foods are
dropped out of the diet.
(Pyke, 1984; Pyke, Pulliam,
& Charnov, 1977)
Partial preference theory
The optimal diet is the outcome of
nutritional balancing of foods with
different nutritional content.
Behavioural and physiological adaptations
to forage of variable nutritional quality, less
efficient in utilizing nutrients from a single
species.
Meeting nutritional needs on low quality
food should lead to a generalist foraging
strategy irrespective of availability of
specific foods.
(Pulliam, 1975; Westoby,
1978)
Detoxification limitation theory
The optimal diet is the outcome of
detoxification processes of different
plant secondary metabolites or
toxins.
Behavioural and physiological adaptations
to forage containing different toxins, less
efficient in detoxifying plant secondary
metabolites from a single species.
Higher detoxification efficiency of
generalist herbivores is realized through
different detoxification pathways for
different plant secondary metabolites.
(Freeland & Janzen, 1974;
Marsh et al., 2006)

16
Herbivory in the Australian context
The concepts generally associated with herbivory, such as resource heterogeneity and
selectivity, have been extensively studied in the Australian context. Australian ecosystems
are dominated by nutritionally poor, highly toxic, and hard to digest Eucalyptus species with
associated arboreal marsupial fauna, including both specialist and generalist species
(DeGabriel, Moore, Marsh, & Foley, 2010; Moore, Wallis, Marsh, & Foley, 2004). Past
research has investigated how variability of abiotic factors (i.e., geological parent material,
climate, elevation, and topography) and biotic factors (i.e., tree species composition,
structure, and abundance) influenced populations of folivorous marsupials over large
spatial scales (i.e., first- and second-order selection; Braithwaite, Binns, & Nowlan, 1988;
Braithwaite, Dudzinski, & Turner, 1983; Braithwaite, Turner, & Kelly, 1984; Pausas,
Braithwaite, & Austin, 1995). Moreover, many studies have examined relationships
between the spatio-temporal variability of biotic factors (including tree availability and
nutritional quality) and marsupial habitat preferences and tree use within their home ranges
(i.e., third- and fourth-order selection DeGabriel, 2008; Moore & Foley, 2005; Moore,
Lawler, Wallis, Beale, & Foley, 2010).
Nutritional studies have found that the extent and structure of resource heterogeneity
determine how and when arboreal marsupials encounter different types of food as they
move through superficially “uniform” Eucalyptus stands (summarized by DeGabriel et al.,
2010). For instance, the strongest influence of resource heterogeneity on arboreal
marsupials is expressed through the chemical variability of Eucalyptus tree assemblages,
because closely related tree species can differ in their palatability (Moore & DeGabriel,
2012). Also, individual trees of the same species can vary greatly in nutrients and plant
secondary metabolites as can different tree parts (i.e., mature vs. young foliage; Edwards et
al., 1993; Lawler et al., 1998). The observed chemical variability may limit or enhance the
opportunity of arboreal marsupials to avoid intoxication by plant secondary metabolites
and their ability to assemble a balanced and nutritious diet (Moore & Foley, 2005; Scrivener
et al., 2004; Youngentob et al., 2011).
Nevertheless, many studies have identified the dietary choices of arboreal marsupials only
in controlled feeding trials in which captive animals were offered Eucalyptus foliage of
varying palatability (e.g., Cork, Hume, & Dawson, 1983; Marsh, Wallis, & Foley, 2003;
Wiggins, McArthur, McLean, & Boyle, 2003).

17
However, the factors that govern the foraging decisions of wild populations have received
less attention, due to difficulties in applying laboratory findings to the complex
environments of wild populations. Almost all studies known to the author, performed both
in laboratory and field conditions, have taken a “eucalypt-centric” approach by looking
only at the eucalypt component of diets of arboreal marsupials (summarized by DeGabriel
et al., 2010; Moore et al., 2004). However, Eucalyptus stands are usually intermixed with
Acacia species, forming a dense understorey and mid-stratum, and have well developed a
grassy and herbaceous ground layer. Both Acacia and understorey species have been
identified as a valuable food resource for arboreal marsupials (McArthur & Turner, 1997;
Shepherd, 1997; Irlbeck & Hume, 2003).
Moreover, what has been largely overlooked in previous studies of the foraging decisions
of arboreal marsupials is the occurrence of parasitic plants within their reach at a tree level.
Aerial parasites, such as native mistletoes (from Loranthaceae and Viscaceae families) and
root parasites (native cherries, Exocarpos spp.; Santalaceae), introduce another level of
resource heterogeneity for arboreal marsupials as they provide additional nutrients without
incurring costs, such as extra travelling or risk of predation. Furthermore, individual
variability and plasticity in foraging choices of Australian arboreal marsupials observed in
other marsupial species, such as the gracile mouse opossum from Brazil (Gracilinanus
microtarsus, Araújo et al., 2009), have been largely overlooked in previous studies.
Arboreal marsupials are dependent on trees not only for food, but also for shelter, with
tree limbs, bark, and hollows providing concealment from predators and protection from
adverse weather conditions (Gibbons & Lindenmayer, 2002). Furthermore, trees facilitate
social interactions and provide a platform on which competition and predation processes
operate (Lindenmayer, 1997; Nersesian, Banks, & McArthur, 2011). Rather than simple
pairwise interactions, individual trees can support multiple individuals of different arboreal
marsupials and their natural enemies, allowing entire food webs to play out within their
canopies. Therefore, the present study takes an integrative approach to explore a basis of
nutrition by looking at the foraging behaviour and tree use of a generalist herbivore, the
common brushtail possum Trichosurus vulpecula (Marsupialia: Phalangeridae; Fig. 2), in
heterogeneous environments of Australian eucalypt forests and woodlands.

18
Fig. 2 The common brushtail possum (Trichosurus vulpecula), photo by H. McGregor

19
The common brushtail possum is a model organism for studying foraging decisions of
herbivores as it represents the epitome of a generalist herbivore in its native range in
Australia and as an introduced species in New Zealand (summarized by Kerle, 2001).
Regardless of a generalist diet, common brushtail possums show clear foraging preferences
at multiple scales of resource heterogeneity and selectively use specific forest types, trees
and understorey species, individual plants and parts, and discrete patches of mistletoes
within tree canopies depending on their availability and nutritional quality (Loney et al.,
2006; McArthur, Goodwin, & Turner, 2000; Scrivener et al., 2004; Sessions & Kelly, 2002).
Furthermore, the current study moves away from nutritional studies done in the past that
attempted to explain foraging decisions of common brushtail possums by deterring effects
of different groups of plant secondary metabolites found in Eucalyptus foliage (e.g. terpenes,
cyanogenic glycosides, formylated phloroglucinol compounds (FPCs); summarized by
Moore et al., 2004). Instead this study focuses on the amount of nitrogen in foliage that is
readily available to animals and not bound with tannins (DeGabriel et al., 2008). Tannins
have been found previously to affect foraging decisions, physiological processes, and
reproductive success in common brushtail possums (Marsh, Wallis, & Foley, 2003;
DeGabriel et al., 2009). Studying the effects of tannins on food nutritional quality instead
of a variety of plant chemical defences against herbivores represents a somewhat a
restrictive approach; but, this approach allows easy cross-taxa comparisons (Eucalyptus vs.
Acacia vs. parasitic plants). This approach also reduces chemical noise resulting from
differences in the amount and type of plant secondary metabolites found in different
species and allows for a clear perception of food as a source of protein for herbivores.

20
Therefore, the overall research objectives of this study are as follows:
1) To determine food preferences, resource selection, and space use patterns of
common brushtail possums at three different scales of resource heterogeneity,
landscape, home range and individual tree-scale, with a special consideration of the
contribution of parasitic plants to small-scale resource heterogeneity.
2) To investigate between and within plant chemical heterogeneity by using a
common currency, available nitrogen, to measure the nutritional value of different
tree species and co-occurring parasitic plants for common brushtail possums.
3) To explore and compare underlying causes of tree use by common brushtail
possums across different spatial scales, taking into account food availability and
nutritional quality, as well as the availability of parasitic plants, tree hollows, and the
presence of competitors.
In the present study, the specific research aims are addressed in Chapters 2–4 while
Chapter 5 summarises and integrates the findings and suggests future research directions.
Chapters 2–4 outline the results of field investigations and laboratory analyses and have
been written as self-contained manuscripts to facilitate subsequent publication.
Consequently, each chapter contains a separate introduction, methods, results, discussion,
and reference list.
In Chapter 2, a landscape-scale study looked at resource selectivity by common brushtail
possums in the context of food availability (species composition of trees and parasitic
plants), hollow availability (used as shelter), as well as potential competition with a
sympatric species, the mountain brushtail possum (Trichosurus cunninghami). This study
focused on the overall coarse-grain patterns of resource selection by establishing the diet
breadth, food preferences, and tree use of the entire population of common brushtail
possums occurring in the study area. In this chapter, standard dietary techniques were used,
direct observations of possum foraging activity and faecal content analysis. This integrative
approach provided a broader nutritional context for the following chapters and enabled
comparisons of resource selectivity on finer scales, home range, and individual tree.

21
In Chapter 3, resource selection (i.e., food and shelter) and space use patterns of common
brushtail possums were explored to establish the home range size and foraging and
denning behavior of individual animals. Both nightly and daily tree sightings of nine
randomly selected individuals (3 males and 6 females) were recorded in a radio-tracking
survey to establish the total home range size for common brushtail possums. This
approach allowed identification of both trees used for foraging and trees used as denning
sites. This information was then depicted in a spatially explicit context to estimate foraging
and denning ranges for each individual possum. Possum food preferences were established
in a more direct way by observing animals foraging within their home ranges. This study
also provided background data for the following chapter by identifying trees used and
avoided by possums while foraging.
In Chapter 4, inter- and intra-specific chemical variability of foliage of the most common
native trees and parasitic plants available to possums within their home ranges was
measured. This approach allowed exploration of the finest scale of resource heterogeneity
affecting foraging decisions of arboreal herbivores. In this study, a recently developed
method by DeGabriel and colleagues (2007, 2009) was used for measuring nutritional value
of browse. It combines the effects of total nitrogen (a proxy of plant protein content),
tannins (protein-binding agents) and fibre (digestibility reducing component of cell walls)
into a single measure of available nitrogen to herbivores. This approach allowed for the use
of a single currency to measure quality of browse for herbivores across different species
and taxa with otherwise unique chemical signatures.
In summary, to understand dietary choices of herbivores in chemically complex
environments, it is important to explore the variability in quantity and quality of forage
available to animals. By knowing what represents a “good” and “easily accessible” food for
herbivores and coupling this knowledge with information gathered from faecal content
analysis and direct observations of animal foraging behavior, we are able to establish the
dietary breadth and foraging preferences of herbivores. Furthermore, studying herbivory
over different spatial scales, landscape, home range, and individual plant, allows for general
conclusions to be made about the nature of herbivory. Finally, it is possible to understand a
broader context of animal nutrition by focusing not only on direct measures of food
availability and nutritional quality, but also on indirect measures of ecological interactions
of herbivores with their conspecifics, predators, and competitors.

22
References
Aerts, R. J., Barry, T. N., & McNabb, W. C. (1999). Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agriculture, Ecosystems & Environment, 75(1), 1–12.
Agrawal, A. A. (2011). Current trends in the evolutionary ecology of plant defence. Functional Ecology, 25(2), 420–432.
Agrawal, A. A., & Fishbein, M. (2006). Plant defense syndromes. Ecology, 87(7), 132–149. Araújo, M. S., Martins, E. G., Cruz, L. D., Fernandes, F. R., Linhares, A. X., Dos Reis, S.
F., & Guimarães, P. R. (2009). Nested diets: A novel pattern of individual‐level resource use. Oikos, 119(1), 81–88.
Armati, P. J., Dickman, C. R., & Hume, I. D. (Eds.) (2006). Marsupials. Cambridge: Cambridge University Press.
Bailey, D. W., Gross, J. E., Laca, E. A., Rittenhouse, L. R., Coughenour, M. B., Swift, D. M., & Sims, P. L. (2006). Mechanisms that result in large herbivore grazing distribution patterns. Journal of Range Management Archives, 49(5), 386–400.
Barbehenn, R. V., & Peter Constabel, C. (2011). Tannins in plant–herbivore interactions. Phytochemistry, 72(13), 1551–1565.
Belovsky, G. E. (1986). Generalist herbivore foraging and its role in competitive interactions. American Zoologist, 26(1), 51–69.
Braithwaite, L. W., Binns, D. L., & Nowlan, R. D. (1988). The distribution of arboreal marsupials in relation to eucalypt forest types in the Eden (NSW) woodchip concession area. Australian Wildlife Research, 15(4), 363–373.
Braithwaite, L. W., Dudzinski, M. L., & Turner, J. (1983). Studies on the arboreal marsupial fauna of eucalypt forests being harvested for woodpulp at Eden, N.S.W. II. Relationship between the fauna density, richness and diversity, and measured variables of the habitat. Australian Wildlife Research, 10(2), 231–247.
Braithwaite, L. W., Turner, J., & Kelly, J. (1984). Studies on the Arboreal Marsupial Fauna of Eucalypt Forests Being Harvested for Wood Pulp at Eden, N.s.w. Iii. Relationships Between Faunal Densities, Eucalypt Occurrence and Foliage Nutrients, and Soil Parent Materials. Australian Wildlife Research, 11(1), 41–48.
Brodie III, E. D., & Brodie Jr, E. D. (1999). Predator-prey arms races. BioScience, 49(7), 557–568.
Brophy, J. J., & Southwell, I. A. (2002). Eucalyptus chemistry. In J. J. W. Coppen (Ed.), Eucalyptus: The Genus Eucalyptus (pp. 102-161). London: Taylor & Francis.
Brown, K., Innes, J., & Shorten, R. (1993). Evidence that possums prey on and scavenge birds’ eggs, birds and mammals. Notornis, 40(3), 169–177.
Chase, J. M., Abrams, P. A., Grover, J. P., Diehl, S., Chesson, P., Holt, R. D., Richards, S. A., Nisbet, R. M., Case, T. J. (2002). The interaction between predation and competition: A review and synthesis. Ecology Letters, 5(2), 302–315.
Chilcott, M., & Hume, I. (1985). Coprophagy and selective retention of fluid digesta: Their role in the nutrition of the common ringtail possum, Pseudocheirus peregrinus. Australian Journal of Zoology, 33(1), 1–15.
Cork, S., Hume, I., & Dawson, T. (1983). Digestion and metabolism of a natural foliar diet (Eucalyptus punctata) by an arboreal marsupial, the koala (Phascolarctos cinereus). Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 153(2), 181–190.
Cowan, P. (1990). Fruits, seeds, and flowers in the diet of brushtail possums, Trichosurus vulpecula, in lowland podocarp/mixed hardwood forest, Orongorongo Valley, New Zealand. New Zealand Journal of Zoology, 17(4), 549–566.
Cowan, P. E. (1989). A vesicular-arbuscular fungus in the diet of brushtail possums, Trichosurus vulpecula. New Zealand Journal of Botany, 27(1), 129–131.

23
Cowan, P. E., & Moeed, A. (1987). Invertebrates in the diet of brushtail possums, Trichosurus vulpecula, in lowland podocarp/broadleaf forest, Orongorongo Valley, Wellington, New Zealand. New Zealand Journal of Zoology, 14(2), 163–177.
Crawley, M. J. (1983). Herbivory. The dynamics of animal–plant interactions. Oxford: Blackwell Scientific Publications.
Cyr, H., & Face, M. L. (1993). Magnitude and patterns of herbivory in aquatic and terrestrial ecosystems. Nature, 361(6408), 148–150.
Dearing, M. D., Mangione, A. M., & Karasov, W. H. (2000). Diet breadth of mammalian herbivores: Nutrient versus detoxification constraints. Oecologia, 123(3), 397-405.
DeGabriel, J. L. (2008). Demographic constraints imposed by plant nutrients and secondary chemistry on the common brushtail possum (Trichosurus vulpecula Kerr.). (Ph. D. thesis), the Australian National University, Canberra.
DeGabriel, J. L., Wallis, I. R., Moore, B. D., & Foley, W. J. (2008). A simple, integrative assay to quantify nutritional quality of browses for herbivores. Oecologia, 156(1), 107–116.
DeGabriel, J. L., Moore, B. D., Foley, W. J., & Johnson, C. N. (2009). The effects of plant defensive chemistry on nutrient availability predict reproductive success in a mammal. Ecology, 90(3), 711–719.
DeGabriel, J., Moore, B. D., Marsh, K. J., & Foley, W. J. (2010). The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology, 20(2), 97–108.
Di Stefano, J., & Newell, G. R. (2008). Diet selection by the swamp wallaby (Wallabia bicolor): Feeding strategies under conditions of changed food availability. Journal of Mammalogy, 89(6), 1540–1549.
Edwards, P. B., Wanjura, W., & Brown, W. (1993). Selective herbivory by Christmas beetles in response to intraspecific variation in Eucalyptus terpenoids. Oecologia, 95(4), 551–557.
Ehrlich, P. R., & Raven, P. H. (1969). Differentiation of populations. Science, 165(3899), 1228–1232.
Eschler, B. M., Pass, D. M., Willis, R., & Foley, W. J. (2000). Distribution of foliar formylated phloroglucinol derivatives amongst Eucalyptus species. Biochemical Systematics and Ecology, 28(9), 813–824.
Evans, M. C. (1992). Diet of the brushtail possum Trichosurus vulpecula (Marsupialia: Phalangeridae) in central Australia. Australian Mammalogy, 15, 25–30.
Foley, W., Iason, G., & McArthur, C. (1999). Role of plant secondary metabolites in the nutritional ecology of mammalian herbivores: How far have we come in 25 years? In H.-J. Jung & G. C. Fahey (Eds.), Nutritional Ecology of Herbivores (pp. 130–209). Savoy, IL: American Society of Animal Science.
Forbes, J. M. (1998). Dietary awareness. Applied Animal Behavior Science, 57(3), 287–297. Forbey, J. S., Dearing, M. D., Gross, E. M., Orians, C. M., Sotka, E. E., & Foley, W. J.
(2013). A Pharm-Ecological Perspective of Terrestrial and Aquatic Plant-Herbivore Interactions. Journal of Chemical Ecology, 39(4), 1–16.
Fox, L. R., & Macauley, B. J. (1977). Insect grazing on Eucalyptus in response to variation in leaf tannins and nitrogen. Oecologia, 29(2), 145–162.
Freeland, W. J., & Janzen, D. H. (1974). Strategies in herbivory by mammals: The role of plant secondary compounds. The American Naturalist, 108(961), 269–287.
Freeland, W. J., & Winter, J. W. (1975). Evolutionary consequences of eating:Trichosurus vulpecula (Marsupialia) and the genus Eucalyptus. Journal of Chemical Ecology, 1(4), 439–455.
Gibbons, P., & Lindenmayer, D. (2002). Tree hollows and wildlife conservation in Australia. Collingwood: CSIRO.

24
Gurevitch, J., Morrison, J. A., & Hedges, L. V. (2000). The interaction between
competition and predation: A meta‐analysis of field experiments. The American Naturalist, 155(4), 435–453.
Hume, I. D. (1999). Marsupial nutrition. Cambridge: Cambridge University Press. Huntly, N. (1991). Herbivores and the dynamics of communities and ecosystems. Annual
Review of Ecology and Systematics, 22(1), 477–503. Iason, G. (2005). The role of plant secondary metabolites in mammalian herbivory:
Ecological perspectives. Proceedings of the Nutrition Society, 64(01), 123–131. Iason, G., & Villalba, J. (2006). Behavioral strategies of mammal herbivores against plant
secondary metabolites: The avoidance-tolerance continuum. Journal of Chemical Ecology, 32(6), 1115–1132.
Irlbeck, N. A., & Hume, I. D. (2003). The role of Acacia in the diets of Australian marsupials? A review. Australian Mammalogy, 25(2), 121–134.
Janzen, D. H. (1980). When is it coevolution. Evolution, 34(3), 611–612. Johnson, D. H. (1980). The comparison of usage and availability measurements for
evaluating resource preference. Ecology, 61(1), 65–71. Johnson, M. T. (2011). Evolutionary ecology of plant defences against herbivores.
Functional Ecology, 25(2), 305–311. Karasov, W. H., & Diamond, J. M. (1988). Interplay between physiology and ecology in
digestion. BioScience, 38(9), 602–611. Karban, R., & Myers, J. H. (1989). Induced plant responses to herbivory. Annual Review of
Ecology and Systematics, 20(1), 331–348. Karban, R., & Baldwin, I. T. (1997). Induced responses to herbivory. Chicago: University of
Chicago Press. Kavanagh, R. P., & Lambert, M. J. (1990). Food selection by the greater glider, Petauroides
volans: Is foliar nitrogen a determinant of habitat quality? Wildlife Research, 17(3), 285–299.
Kerle, A. (2001). Possums: The brushtails, ringtails and greater glider. Sydney: UNSW Press. Kotliar, N. B., & Wiens, J. A. (1990). Multiple scales of patchiness and patch structure:
A hierarchical framework for the study of heterogeneity. Oikos, 59(2), 253–260. Lawler, I. R., Foley, W. J., Eschler, B. M., Pass, D. M., & Handasyde, K. (1998).
Intraspecific variation in Eucalyptus secondary metabolites determines food intake by folivorous marsupials. Oecologia, 116(1), 160–169.
Lima, S. L., & Dill, L. M. (1990). Behavioral decisions made under the risk of predation: A review and prospectus. Canadian Journal of Zoology, 68(4), 619–640.
Lindenmayer, D., Cunningham, R., Tanton, M., Smith, A., & Nix, H. (1990). Habitat requirements of the mountain brushtail possum and the greater glider in the montane ash-type eucalypt forests of the central highlands of Victoria. Wildlife Research, 17(5), 467–478.
Lindenmayer, D. B. (1997). Differences in the biology and ecology of arboreal marsupials in forests of southeastern Australia. Journal of Mammalogy, 78(4), 1117–1127.
Logan, M. (2001). Evidence for the occurrence of rumination‐like behaviour, or merycism, in the koala (Phascolarctos cinereus, Goldfuss). Journal of Zoology, 255(1), 83–87.
Marquis, R. J. (1992). The selective impact of herbivores. In R. S. Fritz & E. L. Simms (Eds.), Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and Genetics (pp. 301–325). Chicago: University of Chicago Press.
Marsh, K. J., Wallis, I. R., & Foley, W. J. (2003). The effect of inactivating tannins on the intake of Eucalyptus foliage by a specialist Eucalyptus folivore (Pseudocheirus peregrinus) and a generalist herbivore (Trichosurus vulpecula). Australian Journal of Zoology, 51(1), 31–42.
Marsh, K. J., Wallis, I. R., Andrew, R. L., & Foley, W. J. (2006). The detoxification limitation hypothesis: Where did it come from and where is it going? Journal of Chemical Ecology, 32(6), 1247–1266.

25
McArthur, C., & Turner, S. (1997). Feeding preferences of captive brushtail possums for eucalypt and acacia foliage. Tasmanian Forests, 9, 155–162.
Mello, M. O., & Silva-Filho, M. C. (2002). Plant-insect interactions: An evolutionary arms race between two distinct defense mechanisms. Brazilian Journal of Plant Physiology, 14(2), 71–81.
Min, B., Barry, T., Attwood, G., & McNabb, W. (2003). The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Animal Feed Science and Technology, 106(1), 3–19.
Moles, A. T., Peco, B., Wallis, I. R., Foley, W. J., Poore, A. G., Seabloom, E. W., Vesk, P. A., Bisigato, A. J., Cella-Pizarro, L., Clark, C. J., Cohen, P. S., Cornwell, W. K., Edwards, W., Ejrnæs, R., Gonzales-Ojeda, T., Graae, B. J., Hay, G., Lumbwe, F. C., Magana-Rodrıguez, B., Moore, B. D., Peri, P. L., Poulsen, J. R., Stegen, J. C., Veldtman, R., Zeipel, H., Andrew, N. R., Boulter, S. L., Borer, E. T., Cornelissen, J. H. C., Farji-Brener, A. G., DeGabriel, J. L., Jurado, E., Kyhn, L. A., Low, B., Mulder, C. P. H., Reardon-Smith K., Rodrıguez-Velazquez, J., De Fortier, A., Zheng, Z., Blendinger, P. G., Enquist, B. J., Facelli, J. M., Knight, T., Majer, J. D., Martınez-Ramos, M., McQuillan, P., & Hui, F. K. C. (2013). Correlations between physical and chemical defences in plants: Tradeoffs, syndromes, or just many different ways to skin a herbivorous cat? New Phytologist, 198(1), 252–263.
Moore, B. D., & DeGabriel, J. L. (2012). Integrating the effects of PSMs on vertebrate herbivores across spatial and temporal scales. In G. R. Iason, M. Dicke & S. E.
Hartley (Eds.), The Ecology of Plant Secondary Metabolites: From Genes to Global Processes (pp. 226–246). Cambridge: Cambridge University Press.
Moore, B. D., & Foley, W. J. (2000). A review of feeding and diet selection in koalas (Phascolarctos cinereus). Australian Journal of Zoology, 48(3), 317–333.
Moore, B. D., & Foley, W. J. (2005). Tree use by koalas in a chemically complex landscape. Nature, 435(7041), 488–490.
Moore, B. D., Lawler, I. R., Wallis, I. R., Beale, C. M., & Foley, W. J. (2010). Palatability mapping: A koala's eye view of spatial variation in habitat quality. Ecology, 91(11), 3165–3176.
Moore, B. D., Wallis, I. R., Marsh, K. J., & Foley, W. J. (2004). The role of nutrition in the conservation of the marsupial folivores of eucalypt forests. In D. Lunney (Ed.), Conservation of Australia’s forest fauna (2 ed., pp. 549–575). Mosman: Royal Zoological Society of New South Wales.
Nersesian, C. L., Banks, P. B., & McArthur, C. (2011). Titrating the cost of plant toxins against predators: Determining the tipping point for foraging herbivores. Journal of Animal Ecology, 80(4), 753–760.
Ohgushi, T. (2005). Indirect interaction webs: Herbivore-induced effects through trait change in plants. Annual Review of Ecology, Evolution, and Systematics, 36, 81–105.
O'Neill, R. V., Johnson, A., & King, A. (1989). A hierarchical framework for the analysis of scale. Landscape Ecology, 3(3), 193–205.
Pahl, L. (1987). Feeding-Behavior and Diet of the Common Ringtail Possum, Pseudocheirus-Peregrinus, in Eucalyptus Woodlands and Leptospermum Thickets in Southern Victoria. Australian Journal of Zoology, 35(5), 487–506.
Pausas, J. G., Braithwaite, L. W., & Austin, M. P. (1995). Modelling habitat quality for arboreal marsupials in the south coastal forests of New South Wales, Australia. Forest Ecology and Management, 78(1–3), 39–49.
Provenza, F. D. (1996). Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. Journal of Animal Science, 74(8), 2010–2020.
Provenza, F. D., Villalba, J. J., Dziba, L. E., Atwood, S. B., & Banner, R. E. (2003). Linking herbivore experience, varied diets, and plant biochemical diversity. Small Ruminant Research, 49(3), 257–274.

26
Pulliam, H. R. (1975). Diet optimization with nutrient constraints. The American Naturalist, 109(970), 765–768.
Pyke, G. H. (1984). Optimal foraging theory: A critical review. Annual Review of Ecology and Systematics, 15(1), 523–575.
Pyke, G. H., Pulliam, H. R., & Charnov, E. (1977). Optimal foraging: A selective review of theory and tests. Quarterly Review of Biology, 52(2), 137–154.
Raubenheimer, D., & Simpson, S. J. (2009). Nutritional PharmEcology: Doses, nutrients, toxins, and medicines. Integrative and Comparative Biology, 49(3), 329–337.
Rhoades, D. F. (1985). Offensive-defensive interactions between herbivores and plants: Their relevance in herbivore population dynamics and ecological theory. The American Naturalist, 125(2), 205–238.
Rothman, J. M., Raubenheimer, D., & Chapman, C. A. (2011). Nutritional geometry: Gorillas prioritize non-protein energy while consuming surplus protein. Biology Letters, 7(6), 847–849.
Scheline, R. P. (1991). Handbook of Mammalian metabolism of Plant Compounds. Boca Raton: CRC Press.
Scrivener, N. J., Johnson, C. N., Wallis, I. R., Takasaki, M., Foley, W. J., & Krockenberger, A. K. (2004). Which trees do wild common brushtail possums (Trichosurus vulpecula) prefer? Problems and solutions in scaling laboratory findings to diet selection in the field. Evolutionary Ecology Research, 6(1), 77–87.
Searle, K. R., Thompson Hobbs, N., & Shipley, L. A. (2005). Should I stay or should I go? Patch departure decisions by herbivores at multiple scales. Oikos, 111(3), 417–424.
Senft, R. L., Coughenour, M. B., Bailey, D. W., Rittenhouse, L. R., Sala, O. E., & Swift, D. M. (1987). Large herbivore foraging and ecological hierarchies. BioScience, 37(11), 789–799.
Shepherd, K. A., Wardell-Johnson, G. W., Loneragan, W. A., & Bell, D. T. (1997). Diet of herbivorous marsupials in a Eucalyptus marginata forest and their impact on the understorey vegetation. Journal of the Royal Society of Western Australia, 80, 47–54.
Simpson, S. J., Raubenheimer, D. (2001). The geometric analysis of nutrient-allelochemical interactions: A case study using locusts. Ecology, 82(2), 422–39.
Stewart, K. M., Bowyer, R. T., Kie, J. G., Cimon, N. J., & Johnson, B. K. (2002). Temporospatial distributions of elk, mule deer, and cattle: Resource partitioning and competitive displacement. Journal of Mammalogy, 83(1), 229–244.
Van Soest, P. J. (1994). Nutritional ecology of the ruminant. Ithaca: Cornell University Press. Westoby, M. (1978). What are the biological bases of varied diets? American Naturalist,
112(985), 627–631. Wiegert, R., & Owen, D. (1971). Trophic structure, available resources and population
density in terrestrial vs. aquatic ecosystems. Journal of Theoretical Biology, 30(1), 69–81. Wiggins, N. L., McArthur, C., McLean, S., & Boyle, R. (2003). Effects of two plant
secondary metabolites, cineole and gallic acid, on nightly feeding patterns of the common brushtail possum. Journal of Chemical Ecology, 29(6), 1447–1464.
Youngentob, K., Wallis, I. R., Lindenmayer, D. B., Wood, J., Pope, M., & Foley, W. J. (2011). Foliage chemistry influences tree choice and landscape use of a gliding marsupial folivore. Journal of Chemical Ecology, 37(1), 71–84.

27
CH
AP
TE
R
2
Dietary composition of the common brushtail possum (Trichosurus vulpecula)
in a heterogeneous landscape: Choice determinants at an “all-you-can-eat buffet”

28
CHAPTER 2
Dietary composition of the common brushtail possum (Trichosurus vulpecula)
in a heterogeneous landscape: Choice determinants at an “all-you-can-eat buffet”
Introduction
Knowledge of what drives foraging choices of herbivores in the wild is essential for our
understanding of the multifaceted dynamics of animal–plant systems. Nutrition is
recognized as a major mechanism regulating the fitness and reproduction of individual
animals, as well as the growth and maintenance of entire populations of herbivores
(Crawley, 1983). The majority of herbivores are generalist foragers satisfying their
nutritional needs from a range of plant species available in the habitat and include such
diverse species as grasshoppers (Unsicker et al., 2008), North American pika (Ochotona
princeps; Dearing, 1996), moose (Alces alces; Belovsky, 1978), and mountain gorilla (Gorilla
gorilla beringei; Watts, 1984). However, plants vary spatially and temporally in concentrations
of nutrients and toxins forcing herbivores to make complex foraging decisions and limiting
their foraging choices to a subset of species available.
Recent studies with captive and domestic animals have revealed the strong effects of plant
nutrients and toxins on diet selection, digestive physiology, and reproductive success of
herbivores (summarized by DeGabriel et al., 2013; Provenza et al., 2009). Nevertheless,
extending our knowledge from the laboratory to the field has proved challenging, and the
existing field studies suffer from several inconsistent theoretical expectations. On one side,
the optimal foraging theory predicts a positive relationship between food selection and
availability in the habitat leading to diet specialization (Pyke, 1984; Pyke et al., 1977).
However, the partial preference theory postulates that a negative relationship and mixed
diets are to be expected if foraging decisions of herbivores are constrained by food nutrient
content (Pulliam, 1975; Westoby, 1974) or, as the detoxification limitation theory
concludes, if eating a range of foods reduces the negative effects of toxins (Freeland &
Janzen, 1974; Marsh et al., 2006a).

29
Ultimately, the foraging theory provides a unifying framework for all the existing theories
and predicts that wild animals will balance the costs and benefits of different foraging
behaviours and their effects on fitness depending upon the animals’ nutritional needs, the
nutritional value and spatial distribution of forage, as well as competition and the risk of
predation (Stephens & Krebs, 1987). Herbivores perceive the risk of predation and plant
toxins as costs to foraging and are able to quantify, compare, and balance these costs,
therefore, alter their foraging behaviour (Nersesian, Banks, & McArthur, 2011).
Furthermore, the availability and proximity of shelter acting as a refuge from predators may
affect herbivore foraging decisions (Duffy & Hay, 1991; Tuft et al., 2011). Herbivores may
manage a risk of predation by selectively foraging in areas close to safety, such as protective
covers, hollows, or borrows.
Moreover, foraging decisions of herbivores in the wild are not simply a trade-off between
nutrient acquisition and the regulation or avoidance of toxins; animal behaviour can be
affected by other ecological factors, such as competition over limited food resources
(Shipley, Forbey, & Moore, 2009). Competitive interactions between animals can lead to
niche partitioning and resource specialization with some herbivores having a limited
realized diet incorporating just a few plant species considered to be “difficult” or low in
nutrients and high in toxins (Robinson & Wilson, 1998). Generalist herbivores, on the
contrary, consume plant species that are wildly available and avoid those that are difficult to
digest or require specialized behavioural, morphological, or physiological adaptations. They
compete well with other herbivores if less difficult types of food are relatively abundant,
but represent poor competitors if plant availability is restricted to difficult types of food
(Shipley, Forbey, & Moore, 2009).
Studying herbivory in Australian ecosystems, dominated by a single genus Eucalyptus,
provides a unique opportunity to investigate foraging decisions of wild animals in a
“field laboratory” where animals are faced with a limited number of food choices. Namely,
despite being widely available, eucalypts are considered to be a difficult food for leaf-eating
insects and arboreal marsupials since they are rich in toxins and poor in nutrients (Fox &
Morrow, 1983; Noble, 1989; Moore et al., 2004a; Moore et al., 2004b; DeGabriel et al.,
2010). Furthermore, the chemical composition of eucalypt foliage changes depending on
the season, habitat, species, individual tree and leaf age (Moore et al., 2004c; Loney et al.,
2006; Andrew, et al., 2007).

30
The feeding experiments with captive arboreal marsupials have shown that animals are
sensitive to types and concentrations of eucalypt toxins and forage selectively on less
difficult species and individuals of the same species (Lawler et al., 1998; Wallis, Watson, &
Foley, 2000; Wiggins et al., 2006b). Nevertheless, these nutritional studies often fail to
establish food preferences of arboreal marsupials in the wild and to look beyond their
eucalypt-dominated diet.
Circumpassing limitations of the previous studies is especially relevant to this study; as
foraging strategies of arboreal marsupials represent a continuum from obligate specialists,
the koala (Phascolarctos cinereus) and greater glider (Petauroides volans), which feed almost
exclusively on foliage of Eucalyptus spp., to a facultative specialist, the common ringtail
possum (Pseudocheirus peregrinus), which supplements its eucalypt diet with foliage of other
tree species, to a facultative generalist, the common brushtail possum (Trichosurus vulpecula),
which forages on a wide variety of woody and seral species. Nonetheless, both specialist
and generalist arboreal marsupials develop food preferences and select some species,
usually high in nutrients and low in toxins, more often than would be expected from their
availability in the habitat. For instance, although koalas have been found to forage on some
120 eucalypt species, they usually use one or two species at a site (reviewed by Moore &
Foley, 2000).
By contrast, common brushtail possums due to their lower detoxification capacity
compared to specialist herbivores are compelled to feed on a variety of species (Marsh et
al., 2006b). To maximize nutrient intake and minimize the negative effects of toxins,
common brushtail possums are known to diet mix (incorporate different plant species and
individuals into their diet, (Wiggins et al., 2003) and diet switch (modify time intervals
between eating different types of food, (Wiggins et al., 2006a). Common brushtail possums
have been found to forage on Eucalyptus and Acacia trees and shrubs, parasitic plants,
different species of herbs and grasses (Evans, 1992; Fitzgerald, 1984; Freeland & Winter,
1975; How & Hillcox, 2000; Irlbeck & Hume, 2003), as well as exotic, ornamental and
orchard trees (Eymann, Herbert, & Cooper, 2006; Sweetapple, Fraser, & Knightbridge,
2004), flowers, pollen, seeds, and fruit (Cowan, 1990) and even some fungi and animal
material (Cowan, 1989).
A special case of selective foraging by arboreal marsupials is expressed at tree level with
animals choosing to feed on mistletoes, parasitic plants dependent on their tree hosts for
water and nutrients.

31
Acting as mineral sinks, mistletoes have nutrient-rich foliage that are considered to be less
chemically and physically defendant, hence, readily eaten by many species of folivores
(Watson, 2001; Watson, 2011). Anecdotal evidence and cafeteria trails with captive koalas
and common brushtail possums demonstrated that animals prefer feeding on mistletoes
over their eucalypt hosts (reviewed by Reid, 1997; Reid & Yan, 2000). Furthermore,
nutritional studies conducted in New Zealand noted serious declines in populations of
native mistletoes in areas where common brushtail possums were found extensively
foraging (Sweetapple et al., 2002; Sweetapple, 2008).
Apart from food availability and nutritional quality, other factors may affect foraging
decisions of arboreal marsupials, including inter- and intra-specific competition over food
and shelter resources and predator avoidance behaviours. For instance, two closely related
species, the common brushtail possum (Trichosurus vulpecula) and mountain brushtail
possum (T. cunninghami), are known to forage on both Eucalyptus and Acacia foliage; yet, the
former species feeds more often on Eucalyptus while the latter on Acacia foliage (Hume,
1999; Burchfield, Agar, & Hume, 2005). It remains to be tested whether this difference is a
result of competitive exclusion, resource partitioning between two sympatric species, or the
existence of inter-specific differences in morphological and physiological adaptations.
Likewise, arboreal marsupials can compete over trees with hollows used as shelter and
concealment from predators forcing animals to defend their trees from intruders or travel
farther from safety in a search for new foraging and denning sites (Smith & Lindenmayer,
1988).
In the current study, foraging behaviour of a wild population of common brushtail
possums (Trichosurus vulpecula) is investigated in relation to food and shelter availability and
competition with a sympatric species, the mountain brushtail possum (T. cunninghami).
In doing so, this study moves away from explaining foraging decisions of arboreal
marsupials using solely nutritional terms, the foliar concentrations of nutrients and toxins,
and explores complex ecological interactions to highlight the existing foraging patterns.
The questions associated with the scales of food availability, measures of diet breadth and
food preferences, as well as competition over food and shelter and a risk of predation are
given due attention. In subsequent chapters foraging decisions of common brushtail
possums are examined in the context of spatial organization of food and shelter resources
and nutritional value of forage.

32
Aims and hypotheses
1. To document diet breadth and food preferences of common brushtail possums
Hypothesis 1: Common brushtail possums are expected to have a mixed diet,
incorporating in addition to eucalypt foliage other species and their parts.
Hypothesis 2: Common brushtail possums show foraging preferences for certain plant
species irrespective of their availability in the habitat.
2. To investigate factors influencing tree use by common brushtail possums
Hypothesis 1: Common brushtail possums are expected to use certain tree species, as well
as trees parasitized by mistletoes more often than expected from their availability in the
habitat.
Hypothesis 2: Trees with hollows are expected to be used more often by common
brushtail possums than expected from their availability in the habitat.
Hypothesis 3: Tree use of common brushtail possums is expected to be affected by
occurrence patterns of a sympatric species, the mountain brushtail possum (T. cunninghami).

33
Study sites
The current landscape-scale study comprised a 70 km network of linear, roadside
vegetation corridors in the Strathbogie Ranges (36° 48´ S, 145° 45´ E) in north-eastern
Victoria, Australia (Fig. 1 and 2, Table 1). The Strathbogie Ranges plateau is a part of the
Victorian Highlands–Northern Fall Bioregion, which covers an area of 1500 km2 about 150
km north-northeast of Melbourne (Phillips et al., 2002). The plateau is part of a batholith
formed from the Middle to Upper Devonian granitic intrusion (370–390 mya) and consists
of a mildly dissected plateau of rolling to hilly tableland, 320–700 m above sea level
(Fig. 3). The plateau consists of coarse-grained granite (Hergt, Phillips, & Ely, 2002).
Reddish duplex or gradational soils have developed from the weathered granite, with gleyed
sandy gradational soils and gleyed loams along drainage lines. The plateau is a part of the
Broken and Goulburn River water catchments (Rundle & Rowe, 1974).
The closest weather-station to the study area is at the township of Strathbogie (35° 51’ 50”
S, 145° 44’ 51’’ E). Mean maximum annual temperature is 18.5°C (1974–2012), and mean
minimum annual temperature is 6.1°C. Mean annual precipitation is 967.4 mm
(1902–2013). February is the hottest and driest month with a mean maximum temperature
of 27.3°C, mean minimum of 11.7°C, and mean monthly precipitation of 44.8 mm.
Winters are cool and wet; July is the coldest month with a mean maximum temperature of
10.2°C and minimum of 1.6°C; mean maximum precipitation of 122.9 mm. Rain (> 1 mm)
falls for 81.4 days per year on average; and snow falls occasionally (Bureau of Meteorology,
Strathbogie 2013).
The pre-European native vegetation on the Strathbogie plateau consists of Herb-rich
Foothill Forest, with Shrubby Dry Forest on the lower slopes. Mountain Dry Woodland
occupies the upper slopes of the plateau, and grassy dry forest occurs along the major river
valleys. Riparian and swamp woodlands or scrubs (i.e., Perched Boggy Shrubland, Swampy
Woodland, and Swampy Riparian Woodland) occur in seasonally or permanently inundated
areas and along watercourses (Carr et al., 2006). Since the 1850s, the relatively high rainfall
and fertile soils have resulted in extensive clearing of the original vegetation for agriculture
and softwood production (Pinus radiata). This clearing produced a highly heterogeneous
landscape (Fig. 4) consisting of native vegetation (29%), pine plantations (5%), and cleared
pastoral land (66%, Martin & Martin, 2004).

34
The most intact remnants of the original vegetation are preserved in linear roadside
corridors (10–40 m wide) and discrete patches of forest on both private and public land.
Roadside vegetation remnants are believed not to have been logged for the last 100 years or
burnt for at least half a century.
Vegetation species composition and associated arboreal marsupial fauna
The surveyed network of roadside vegetation corridors consisted of a mosaic of Herb-rich
Foothill Forest in drier areas (Fig. 5) and Swampy Riparian Woodland in seasonally or
permanently inundated areas and along watercourses (Fig. 6). Herb-rich Foothill Forest
was dominated by Eucalyptus radiata (narrow-leaved peppermint), E. dives (broad-leaved
peppermint), and E. viminalis (manna gum) with occasional E. globulus subsp. bicostata
(Victorian blue gum) and E. obliqua (messmate). Key mid-stratum and understorey tree
species included Acacia dealbata (silver wattle), A. melanoxylon (blackwood) and Exocarpos
cupressiformis (cherry ballart; a root parasitic shrub or small tree in the Santalaceae family
often associated with Eucalyptus hosts).
Similar mid-stratum and understorey tree species occurred in Swampy Riparian Woodland,
with Eucalyptus camphora (mountain swamp gum) dominating in the upper-storey. In the
study area, two mistletoe species were found parasitizing branches of different Eucalyptus
and Acacia species, the dominant Amyema pendula (drooping mistletoe) and the less
abundant Muellerina eucalyptoides (creeping mistletoe, both Loranthaceae family).
The understorey of Herb-rich Foothill Forest was dominated by shrubby, herbaceous, or
grassy species from the families Fabaceae, Asteraceae, Poaceae, and austral bracken
(Pteridium esculentum) while Swampy Riparian Woodland was dominated by rushes and
sedges. The most common exotic species in the understorey was Rubus fruticosus (common
blackberry).
In the study area, Downes, Handasyde, and Elgar (1997) carried out a fauna composition
survey. Two closely related, generalist herbivores, the common brushtail possum
(Trichosurus vulpecula) and the mountain brushtail possum (T. cunninghami), were found to be
common in the study area. The eucalypt specialists, the common ringtail possum
(Pseudocheirus peregrinus), greater glider (Petauroides volans), and koala (Phascolarctos cinereus),
were widespread in the study area as was the acacia-dependant species, the sugar glider
(Petaurus breviceps). Potential predators in the study area included the European red fox
(Vulpes vulpes), feral cat (Felis catus), and a native raptor, the powerful owl (Ninox strenua).

35
Fig. 1 Location of study sites within the Strathbogie Ranges, north-eastern Victoria, Australia

36
Fig. 2 Landscape view of roadside vegetation corridors across the Strathbogie Ranges plateau (photo by K. Petrović)
Table 1 Length of surveyed roadside vegetation corridors in the study area
Road name
Road type
Total length (km)
Surveyed length (km)
Creek Junction Rd sealed road 13.2 12.0
Bonnie Doon Rd sealed road 9.3 7.0
Boundary Hill Rd sealed road 5.9 5.6
Harrys Creek Rd sealed road 8.8 6.8
Euroa-Strathbogie Rd sealed road 5.3 3.5
Spring Creek Rd sealed road 4.6 4.5
Brookleigh Rd unsealed road 4.5 4.1
McGearys Rd unsealed road 2.9 2.9
Mackrells Rd unsealed road 4.5 4.4
Ankers Rd sealed road 14.7 12.3
Tames Rd
sealed road
0.3
0.3
Total n. a. 74.0 63.4

37
Fig. 3 Digital elevation model of the study sites within the Strathbogie Ranges Fig. 4 Land use across the study sites (Primary classification, BRS 2010)

38
Fig. 5 Herb-rich Foothill Forest in roadside vegetation in the Strathbogie Ranges (photo by K. Petrović)
Fig. 6 Swampy Riparian Woodland in roadside vegetation in the Strathbogie Ranges (photo by K. Petrović)

39
Methods
Description and justification of methods used to establish diet breadth, food preferences, and tree use by
common brushtail possums
To investigate foraging behaviour and resource selection of arboreal marsupials and other
groups of herbivores it is important to employ reliable and replicable research methods.
For the purpose of robust dietary and habitat inferences, Manly and colleagues (2002)
categorized resource units as “available,” “used,” and “unused” by animals with resource
selection measured by comparing any two of the three possible combinations. However,
the current techniques that are used for measuring availability of forage to herbivores
impose some logistical challenges, such as the researcher’s a priori knowledge of the type of
resources that animals actually use. Moreover, identification of the resources used by
herbivores is especially difficult in the case of arboreal marsupials as they are
predominantly nocturnal species foraging high in tree canopies (Wayne et al., 2006).
Direct methods, such as catching animals regularly for assessment of mouth or fistula
content, might be difficult to carry out or stressful to animals. Laboratory analytical
methods for establishing the dietary composition of arboreal marsupials can be considered
unethical or prohibitively destructive and involve the killing of animals to examine their
stomach content (Sweetapple & Nugent, 1998). Contrarily, using non-lethal methods, such
as the faecal content analysis, can introduce a certain level of bias to results by
underestimating or overestimating proportions of different plant species in a diet (Dunnet,
Harvie, & Smit, 1973).
Vegetation survey
In this study, a systematic vegetation survey was carried out to determine the composition,
dominance, and structure of tree species, as well as to estimate the relative availability of
different tree species and mistletoes to common brushtail possums. This approach was
deemed to be more appropriate than random sampling as it allowed a large set of data to be
collected within a limited time frame across a large study area, including 70 km of roadside
vegetation corridors. Vegetation corridors were divided into 200 m long strips, 10–40 m
wide, depending on the width of a surveyed corridor (Fig. 7). These strips were located on
both sides of the road as home ranges of common brushtail possums are known to include
both roads and roadside vegetation corridors (Del Borrello, 2009).

40
A single tree at the start of each 200 m vegetation strip was identified as a “systematic tree”
for which the following information was collected: exact location, tree species, height,
diameter at breast height (DBH), and number of hollows and mistletoes. In total 577 trees
were described across the entire study area.
Fig. 7 Sampling design for establishing tree species availability, possum and mistletoe tree occurrence in the study area
To establish the relative availability of tree species and mistletoes to possums, the canopy
volume of 50 randomly selected trees parasitized by mistletoe was measured, as well as the
volume of 334 mistletoes. The exact location of all studied trees was measured with a hand-
held positioning device Garmin GPS 12 Personal Navigator (Garmin International, Inc.,
Olathe, KS, USA). Tree diameter and height were recorded using a hand-held laser
hypsometer (LaserAce®, MDL Laser Systems, Ltd., York, UK). To collect and store data
in the field, a Trimble® Nomad hand-held computer (Sunnyvale, CA, USA) installed with
ArcPad 8.0 Software integrated with ArcGIS 9.3 Software (2009 ESRI, Redlands, CA) was
used.

41
Techniques used for establishing diet breadth and tree use by common brushtail possums
The selection of resources by common brushtail possums was investigated using two
complementary techniques: the faecal content analysis of remnant cuticle fragments and
nightly spotlighting observations of animal foraging activity. These techniques are standard
methods recommended by Jones and Krockenberger (2007) for studying the foraging of
arboreal, nocturnal, and cryptic herbivores. The authors highlighted advantages and
disadvantages of each method. Namely, direct observations of foraging animals produce
the highest diversity of diet; the faecal content analysis requires the least time in the field;
and tree selection allows intraspecific measures of preference to be determined. On the
other hand, direct observations of foraging animals are time-consuming and difficult within
a dense forest; the faecal content analysis underestimates the importance of species with
fragile cuticles; and tree selection is not directly related to food intake and can be ascribed
to availability of hollows and mistletoes, presence of conspecifics, competitors, and
predators.
Faecal content analysis
The faecal content analysis is a standard technique for quantifying the dietary composition
of herbivores. The analysis relies on the premise that a plant cuticle, a waxy non-cellular
layer protecting a leaf from desiccation, is species specific. Cuticles carry the unique imprint
of leaf surface characteristics, such as a size, shape, and alignment of cell patterns, stomata,
and trachoma. This technique is commonly used in rangeland studies of grazing and
browsing behaviour of large herbivores and is considered to be a relatively reliable
technique for establishing the diet composition of arboreal folivorous marsupials (Ellis,
Carrick, Lundgren, Veary, & Cohen, 1999).
The main disadvantage of cuticular analysis is the differential digestion of various plant
species by herbivores, with species with fragile cuticles being totally or partially broken in
the digestion process; therefore, they are missed or under-represented in analysed samples
(Fitzgerald & Waddington, 1979). Different digestibility of plant cuticles means that
feeding trials with captive animals are required to derive correction factors to adjust the raw
data (Dunnet et al., 1973; Fitzgerald & Waddington, 1979; Vavra & Holechek, 1980).
However, Sweetapple and Nugent (2007) point out that the inevitable random sampling
error in the correction factors reduces statistical precision and suggests the stomach
content analysis as a sound alternative.

42
In the current study, it was technically unfeasible and prohibitively destructive to capture or
kill animals to establish their diet. Hence, proportional representation of some plant groups
(i.e., herbs, grasses and fruits) could have been underestimated while others, such as
sclerophyll foliage Eucalyptus and Acacia species, could have been overestimated in the diet
of possums. This fact was treated with caution when discussing results.
Collection of common brushtail possum faeces
To establish the diet breadth, food selection, and preferences of common brushtail
possums in the study area, animal faeces were collected opportunistically from live traps
located at sites where possums were observed foraging (Fig. 8). During two trapping
periods November–December 2009 and January–February 2010 (see Chapter 3 for details
of the used trapping method), 56 clumps of scats were collected, each of which was
deposited by a different animal. To ascertain independence of samples, cage traps were set
up only once in a given location. Moreover, where possible, each animal was identified
from ear tattoos made by previous researchers working on the same population
(Del Borrello, 2009). Only fresh faeces were collected for the analysis. Faeces were
considered fresh if they were moist, maintained a black coating, and fragments of plants
remained green. They were collected each morning during trap clearing, transported to a
field station, and were permanently frozen to avoid degradation.

43
Fig. 8 Faeces collection sites (photo of possum scats by P. Canty)

44
Microhistological analysis of plant fragments in faeces
To identify plant fragments remaining in possum faeces, a reference slide collection of leaf
cuticles was prepared for all native tree species and parasitic plants, as well as grasses and
herbs that were available to animals in the study area (Fig. 9). Reference slides were
prepared by cutting leaf edges to expose the mesophyll tissue within the cuticular layers.
An area approximately 10 mm x 10 mm was sectioned with a sharp scalpel. The section
was placed in 42 g/L sodium hypochlorite (domestic bleach) until the mesophyll was
sufficiently digested. The cuticles were then peeled apart and washed with distilled water to
remove all the residual sodium hypochlorite. Once rinsed, the cuticles were observed under
a dissecting microscope to check if all the mesophyll was removed. Samples were then
stained with 0.2 g/L gentian violet for 3 minutes. The cuticles were rinsed of excess stain in
distilled water and were mounted on a microscope slide with corn syrup. Prepared slides
were photographed with a Nikon digital camera at x10, x20 and x40 magnifications to
create a cuticle reference library.
To analyse the content of faecal samples, scats were prepared by loosely breaking two or
three randomly chosen scats from each clump of scats. The scats were placed in a small
bottle and covered with 42 g/L sodium hypochlorite (domestic bleach), then left to digest
for 12 to 24 hours until most plant fragments in scats were translucent (modified from Ellis
et al., 1999). The plant fragments were rinsed with distilled water and were then washed
through a 1000 μm sieve, followed by a 500 μm sieve. All particles that were too big for the
1000 μm sieve or too small for the 500 μm sieve were discarded. The remaining plant
fragments were considered large enough to be identified since other similar studies found
that the proportion of fragments compromising each species was consistent between sieve
fractions (Ellis et al., 1999; Tuft, Crowther, & McArthur, 2011).
Next, all remaining plant fragments were placed in a Petri dish containing gentian violet
(0.15 g/L) and stained for 3 minutes. The plant fragments were then tipped onto Whatman
filter paper (11 μm pore diameter) atop a Bruckner Funnel, with slight air suction, and were
rinsed with a stream of distilled water until excess stain had washed away. The fragments
were dried on a filter paper and mounted onto a microscope slide using corn syrup. To
analyse the cuticle content of each scat sample, starting from the edge of a slide, the first
100 clear cellular cuticles were identified under a compound light microscope. Species
accumulation curves indicated that 100 cuticle fragments per scat were adequate to detect
the majority of present species (Fig. 10).

45
Fig. 9 Microhistological leaf tissue structure (x20 magnification) of Acacia dealbata (silver wattle) and A. melonoxylon (blackwood), Eucalyptus radiata (narrow-leaved peppermint) and E. dives (broad-leaved peppermint), E. camphora (swamp gum) and E. viminalis (manna gum), E. globulus (Victorian blue gum) and E. obliqua (messmate), Amyema pendula (drooping mistletoe) and Exocarpos cupressiformis (cherry ballart; photo by H. McGregor)

46
Fig. 10 Example of species accumulation curves representing the number of plant cuticle fragments considered sufficient for detecting majority of plant species present within a faecal pellet of the common brushtail possum
Spotlighting survey
The spotlighting survey or direct observations of nocturnal activity of arboreal herbivores
involve locating animals within tree canopies using a hand-held lamp and making
behavioural observations through binoculars. Canopy spotlighting or scanning searches are
performed at night while walking along established transects. Animals are located through
reflected eye-shine, sudden movement, or vocalization. Known biases of this technique
include low detectability estimates and presence of false absences due to differences in
animal individual behaviour (i.e., light or observer shyness), dark colouration of animals,
the location of animal home ranges in relation to forest edges, and differences in tree
allometry or canopy structure (Bennett et al., 1991). To overcome some of these biases, the
spotlighting survey was conducted twice and a second independent observer was
employed.
To locate common brushtail possums in roadside vegetation, a spotlighting survey was
conducted in the spring and summer of 2009. To avoid possible differences in foraging
behaviour, sections of vegetation corridors connected to intact patches of forest or
neighbouring household gardens were omitted from the sampling. Two observers spent a
total of 140 hours of spotlighting, each using an 80-W spotlight and 8 x 30 binoculars,
starting at least 1 hour after sunset and walking at a steady pace of 1 km/h.

47
All individuals were recorded in each 200 m vegetation strip, with the locations of multiple
individuals cross-checked to avoid multiple records of the same individual. The following
data were collected for trees occupied by common brushtail possums: exact location, tree
species, height, and diameter at breast height (DBH), number of mistletoes and hollows. A
hollow was considered a suitable refuge for common brushtail possums if the entrance was
greater than 20 cm diameter since animals cannot enter smaller openings (Gibbons &
Lindenmayer, 2002). Moreover, to investigate the incidence of resource competition with a
sympatric species, the mountain brushtail possum (Trichosurus cunninghami), tree occupancy
and tree species characteristics were also recorded. The exact tree location was measured
with a hand-held positioning device Garmin GPS 12 Personal Navigator (Garmin
International, Inc., Olathe, KS, USA). Tree diameter and height were recorded using a
hand-held laser hypsometer (LaserAce®, MDL Laser Systems, UK). All sightings of
possums were plotted on a digitized base map of a study area with ArcGIS 9.3 Software
(2009 ESRI, Redlands, CA).

48
Data analysis
Estimation of available biomass to common brushtail possums
To establish dietary preferences of common brushtail possums in the study area it was
important to determine the amount of available leaf biomass in an ecologically meaningful
way. In the current study, it was technically unfeasible and prohibitively destructive to
directly measure the leaf biomass of trees and mistletoes since cutting a number of large
trees with high mistletoe loads and separately drying and weighing leaves of both trees and
mistletoes would be required. Instead, the leaf biomass of trees and mistletoes was
estimated using the calculated total aboveground biomass of trees and measured in the field
volumes of tree canopies and mistletoes. Consequently, the leaf biomass of trees and
mistletoes was expressed as a non-parametric measure of proportional availability of trees
and mistletoes to common brushtail possums in the study area.
For the purpose of this study a number of assumptions have been adopted. First, it was
assumed that the leaf biomass of trees was directly proportional to the total aboveground
biomass (Enquist & Niklas, 2002). Secondly, as it was technically unfeasible to estimate
directly mistletoe leaf biomass, the total volume of mistletoes was used instead and
compared with the total canopy volume of a subset of parasitized trees of the known
aboveground biomass. It was assumed that the total volume of tree canopies is directly
proportional to the total aboveground biomass (Dai et al., 2009).
Moreover, it is important to stress here that different species of trees with different
morphology and canopy structure may affect mistletoe traits, such as growth form and leaf
shape (Barlow & Wiens, 1977). Hence, for the purpose of this study it was adopted that
mistletoe leaf biomass is proportional to tree leaf biomass irrespective of tree species (after
Reid, Yan & Fittler, 1994).
To calculate the total aboveground biomass of trees in the study area, a generalized
allometric equation was employed that used tree diameter (DBH) as a predictor variable of
the total aboveground biomass:
lny = -2.3267 + 2.4855 lnx, where y is the aboveground biomass (kg) and x is tree DBH (cm)

49
This generalized allometric equation was developed by Keith, Barrett, and Keenan (2000)
for a suite of species growing in native sclerophyllous forests. The equation was derived by
combining predicted biomass values from specific allometric equations developed for
individual species or sites. The generalized allometric equation accounted for most of the
variation in biomass predicted for individual species or sites (R2 = 0.96).
To estimate proportional availability of mistletoes and different species of trees in the study
area, coefficient was introduced and expressed as the ratio of availability of mistletoes
( ) to availability of trees ( ):
Assuming that availability is directly proportional to volume, we can further express
coefficient as:
In the above equation is the total volume of mistletoes found parasitizing n
number of trees of the total canopy volume while is the average
volume of mistletoes per tree. is the average canopy volume of n number of trees.
In the present analysis, 531 (N) trees were selected systematically to represent the entire
population of trees in the study area, out of which 66 trees (n1) were found to be
parasitized by mistletoes. However, in this case volumes of tree canopies and mistletoes
were not known. Therefore, an independent sample of 50 trees parasitized by mistletoes
(n2) was selected randomly out of all parasitized trees in the study area, and volumes of tree
canopies and mistletoes were measured.
Next, coefficient was calculated for the 50 trees of the known canopy and mistletoe
volumes:

50
To calculate coefficient with 95% confidence intervals, the bootstrap procedure was
used. Namely, 50 trees corresponding to the number of trees of the known canopy and
mistletoe volumes were randomly drawn from the pool of 50 trees (with replacement).
This procedure was repeated 10,000 times.
To calculate the total volume of mistletoes parasitizing 66 trees (n1) in the entire population
of trees (N = 531), it was assumed that the average volume of mistletoes per tree was equal
in both studied subpopulations ( and ):
Next, coefficient or the ratio of the total volume of mistletoes to the total volume of
tree canopies in the study area was calculated. It was assumed that the total volume of
mistletoes was equal to the total volume of mistletoes parasitizing 66 trees
:
The bootstrap procedure (10,000 repetitions) was used to calculate coefficient with
95% confidence intervals (CI).
Knowing the value of coefficient, it is possible to calculate availability of mistletoes
( ), total availability of trees ( ), and availability of each of the tree species
( ) present in the study area. Assuming that availability ( ) is proportional to biomass
( ), we can express availability as coefficient :

51
Knowing that mistletoe availability is we can express
mistletoe availability using the following formula:
In the above equation, is the total aboveground biomass of all trees present in the
study area that can be expressed using the following formula:
symbolizes the total aboveground biomass of a specific tree species: SW (Acacia
dealbata), BG (Eucalyptus globulus), SG (Eucalyptus camphora), MG (Eucalyptus viminalis), MM
(Eucalyptus obliqua), NLP (Eucalyptus radiata), WC (Exocarpos cupressiformis), and EX (exotic
tree).
Finally, assuming that all availabilities added together equal one, we can derive the
following equations for calculating mistletoe availability ( ) and the availability of
each of the tree species ( ) present in the study area:
Diet and food preferences of common brushtail possums
To verify the assumption that a diet of common brushtail possums consists of at least one
Eucalyptus species and might include other plant groups (i.e., acacias, parasitic plants, herbs,
and grasses), the percent share of different forage types in the overall diet was established
and presented as a mean with standard error (s.e), unless otherwise indicated. The collected
data on the faecal composition of 56 common brushtail possums were pooled together for
further analyses while the individual variation was tested by calculating the coefficient of
variation.

52
To explore food preferences of common brushtail possums in the study area the
standardized selection index (B) was employed (after Manly et al., 2002). The standardized
selection index (B) compares resource use (or consumption of different plant species
reconstructed from the faecal content analysis) with resource availability (or estimated
biomass). In this study, the term “food preferences” refers to the positive selection of
different plant species. Namely, a plant species is preferred when its proportion in the diet
is significantly higher than the proportion of available biomass in the habitat; and,
conversely, a plant species is avoided when its proportion in the diet is significantly lower
than the proportion of available biomass in the habitat. Hence, to be selective refers to
both preference and avoidance of different plant species. When a plant species is
consumed in proportion to its availability, no selection is observed.
Resource selection (B) is defined by the following formula:
Bil = (ui/mi)/∑i (ui/mi), where ui is a number of units in category i in the sample of used
units and mi is the number of available units in category i in the sample (i = 1, 2,…, I)
while l is the number of animals.
The values of this index range from 0 to 1 and are interpreted as the probability that a food
type would be the next selected if all food types were equally available. When food types
were available but not used, the 0 use value was replaced by 0.01 (after Di Stefano &
Newell, 2008).
In this study, resource use (U) was restricted only to species of trees and parasitic plants
that were expressed as the proportion of all species consumed by common brushtail
possums. Tree species, Acacia melanoxylon and Eucalyptus obliqua, were excluded from the
analysis because the sample size of A. melanoxylon was too small to produce meaningful
results while E. obliqua was scarcely distributed within a study area and, hence, available
only to a small number of animals. Also, the understorey species were excluded from the
analysis as it was hard to establish available biomass in a meaningful way. Namely, the
studied common brushtail possums were not restricted to roadside vegetation corridors
and often visited adjacent pastured areas (see Chapter 3). Moreover, as this study focused
on overall selection patterns and not individual variation in consumption, the relative
resource availability (M) was the same for all animals across the landscape.

53
This approach is acceptable only in a highly homogenous habitat or when habitat
heterogeneity is expressed at a micro-scale and associated herbivore assemblages are highly
mobile; the last two premises were fulfilled in this study (Zweifel‐Schielly et al., 2011).
The resource use (U), availability (M), and selection (B) of different plant species were
calculated for each individual in the study area (n = 56) and expressed as medians with 95%
confidence intervals (CI). In this study, median appeared to be a better measure of central
tendency than mean since values were proportions ranging from 0 to 1 (after Zar, 2009).
Confidence intervals and significance of selection (B) were calculated using a bootstrap
procedure (10,000 repetitions) in the statistical program R 2.11.1 (2010 The R Foundation
for Statistical Computing). To determine common brushtail possum preferences for
different genera, the logarithmic differences (median ± 95% confidence intervals) were
calculated and their significance tested. Here, the logarithmic difference between two plant
genera was independent from the other genera for which the selection index was also
calculated. Hence, the effects were expressed as the first named food type minus the
second food type; therefore, positive values meant the first named food type was selected
to a greater degree.
Determinants of tree use by common brushtail possums
Tree species preferences of common brushtail and mountain brushtail possums were
determined using a bootstrap or randomization procedure after Jones, Maclagan, and
Krockenberger (2006). A number of trees corresponding to a number of independent
sightings of possums (38 trees used by common brushtail possums and 49 trees used by
mountain brushtail possums) was randomly selected (with replacement) from the total of
577 trees recorded in the study area, and the tree species was recorded. This procedure was
repeated 10,000 times to produce an expected distribution of randomly selected tree
species. The number of sightings of possums in a specific tree species was compared with
the randomly generated distribution, and the probability of sighting was determined directly
from the randomized distribution. To avoid pseudo replication, all trees that were used by
more than one individual were excluded from the analysis.

54
To test the hypotheses that trees parasitized by mistletoes, trees bearing hollows, as well as
medium and large trees (50–100 cm and 100–200 cm DBH) were used more often by
common brushtail and mountain brushtail possums than expected from their availability in
the habitat, the chi-square goodness of fit test was used. To test the relationship between
tree size and the number of mistletoes and hollows, the Spearman's rank test was used. To
compare differences between all measured variables for trees used by common brushtail
possums and mountain brushtail possums, first the Lilliefors test for normality was
employed and next the Wilcoxon signed ranks test was used to compare medians.
All calculations were performed using the statistical program R 2.11.1 (2010 The R
Foundation for Statistical Computing) with a significance level α = 0.05.
Finally, to explain the relationship between common brushtail possum tree presence or
absence and all measured variables (tree species, height and diameter, biomass, number of
mistletoes and hollows, and the occurrence of the mountain brushtail possum) the
binomial logistic regression, a type of the binomial generalized linear model was used.
The binomial logistic regression analysis examines the influence of various factors on a
dichotomous outcome (1—presence; 0—absence) by estimating the probability of the
event’s occurrence. It examines the relationship between one or more independent
variables (continuous or categorical) and the log odds of the dichotomous outcome by
calculating changes in the log odds of the dependent as opposed to the independent
variable itself.
Next, the step Akaike’s Information Criterion (hereafter AIC) function from the MASS
library was employed to identify the best model with the combination of variables
explaining the possum tree presence (Venables & Ripley, 2002). This function is a stepwise
algorithm that sequentially searches through all possible models for the one that minimizes
the AIC. Namely, it starts from the simplest model (the intercept) and adds one step at a
time to a new variable, considered the best by the AIC, to each of the consecutive models.
On each step, the AIC from the new model is compared to the previous model, and if the
AIC from the new model is smaller, the step is repeated. If the AIC from the new model is
not smaller, the algorithm stops and the best model is considered to be the one from the
previous step. If there are no more variables to be added to the new model, the algorithm
stops and the full model is considered the best. The significance of variables from the final
selected model is estimated using the z-test.

55
The AIC was used in this study because the information-theoretic approach is a
recommended alternative to traditional hypothesis testing since it is more tolerant to
violations of parametric statistics that are commonly encountered in ecological data.
Moreover, the AIC method allows for the comparison of multiple working hypotheses and
does not rely solely on the use of ecologically arbitrary P values for determining
significance (Burnham & Anderson, 2002).
Distribution and density of sympatric species, the common brushtail possum and mountain brushtail
possum
All sightings of common brushtail possums and mountain brushtail possums within
roadside vegetation corridors were plotted on a digitised base map of a study area with
ArcGIS 9.3 Software (2009 ESRI, Redlands, CA). Next, the probability density of these two
sympatric species was established using the ArcGIS Kernel Home Range function that
calculates a fixed kernel home range utilization distribution as a grid coverage using the
least squares cross validation for the smoothing parameter (Worton, 1989).

56
Results
Diet breadth and food preferences of common brushtail possums
Based on a total of 5,617 epidermal fragments from 56 analysed faecal samples, the overall
diet composition of common brushtail possums in the study area was established. In total,
nine upper-storey and mid-strata native tree species were identified from the genera
Eucalyptus and Acacia (5 eucalypt and 4 acacia species), along with root and aerial parasites,
Exocarpos cupressiformis (cherry ballart) and Amyema pendula (drooping mistletoe). Exotic trees
included Pinus radiata (radiata pine), Fraxinus angustifolia (narrow-leaved ash), Quercus robur
(English oak), and Prunus cerasifera (cherry plum); due to a low sample size they were pooled
together for further analyses. Understorey species were ascribed to broad functional groups
of herbs and grasses and were excluded from all statistical analyses (see Methods for
justification).
Trees and parasitic plants contributed to the largest proportion of epidermal fragments
identified in faeces of common brushtail possums (60%, Fig. 11). Eucalyptus species from
Monocalyptus and Symphyomyrtus subgenera were found to represent 21.6% of all species
consumed (dominated by E. radiata 14.1%), followed by exotic trees (17.1%) and Acacia
species (9.7%) with silver wattle (A. dealbata) representing 6.3% of Acacia spp. (Fig. 12).
Parasitic plants Amyema pendula and Exocarpos cupressiformis represented 6.8% and 4.3% of all
consumed species (Fig. 12). Understorey species, including forbs and grasses, represented
15.3% of the possum diet while fruit (i.e., common blackberry, Rubus fruticosus) and pollen
(i.e., Acacia and Eucalyptus) constituted 11% of the total diet (Fig. 12). However, 802
epidermal fragments (14.2%) could not be assigned to any plant species, group, or part
(Fig. 12).

57
Fig. 11 The overall diet composition of common brushtail possums occurring in the roadside vegetation corridors of the Strathbogie Ranges, Victoria
Fig. 12 Mean consumption (%) with standard error (s.e) of different plant species, groups, and parts by common brushtail possums occurring in the roadside vegetation corridors of the Strathbogie Ranges, Victoria

58
On average, six plant species, including from one to three Eucalyptus and Acacia species,
were found in 75% of all faeces analysed, followed by exotic trees that compromised 71%
of all faecal samples. Surprisingly, more than 90% of faeces contained key understorey
groups, forbs and grasses. Finally, more than half of all faeces contained fragments of one
of the two parasitic species, Exocarpos cupressiformis or Amyema pendula, fruit and pollen.
In the present study, differences in use of various plant species/genera by common
brushtail possums and their availability in the habitat produced different selection indices
(B, Table 2). Parasitic species, Exocarpos cupressiformis and Amyema pendula, were consumed
more frequently than expected from their availability in the habitat while Eucalyptus species
were consumed less frequently than expected. (P < 0.05, Table 2, Fig. 13). The most
dominant tree species in the habitat, Eucalyptus radiata and E. viminalis, were selectively
avoided regardless of the magnitude of available biomass in the habitat (P < 0.001,
Table 2, Fig. 13). Moreover, when all genera were compared, parasitic plants, Acacia, and
exotic trees were preferred over genus Eucalyptus (P < 0.001, Fig. 14).

59
Table 2 Relative availability, use, and selection of different tree species and parasitic plants by common brushtail possums occurring in the roadside vegetation corridors of the Strathbogie Ranges, Victoria. Values are presented as medians with 95% confidence intervals (CI). Plus (+) symbolizes positive selection or preference for the species, while minus (–) negative selection or avoidance of the species by common brushtail possums. An asterix indicates significance of selection at P < 0.05
Plant species % Available (A) % Used (U) Selected (B)
Acacia dealbata 0.014 (0.008, 0.022) 0.129 (0.091, 0.171) 0.143 (0.078, 0.229) (+)
Eucalyptus globulus 0.034 (0.007, 0.076) 0.001 (0.000, 0.002) 0.006 (0.002, 0.032) (–)*
Eucalyptus camphora 0.088 (0.050, 0.136) 0.078 (0.037, 0.131) 0.021 (0.008, 0.045) (–)*
Eucalyptus viminalis 0.195 (0.124, 0.278) 0.013 (0.002, 0.031) 0.003 (0.001, 0.008) (–)*
Eucalyptus radiata 0.623 (0.530, 0.710) 0.261 (0.184, 0.344) 0.014 (0.006, 0.027) (–)*
Exotic trees 0.035 (0.005, 0.080) 0.316 (0.229, 0.408) 0.196 (0.097, 0.414) (–)
Exocarpos cupressiformis 0.002 (0.001, 0.003) 0.073 (0.040, 0.113) 0.295 (0.169, 0.443) (+)*
Amyema pendula 0.003 (0.001, 0.006) 0.123 (0.073, 0.183) 0.291 (0.167, 0.448) (+)*

60
Fig. 13 Standardized index of plant species selection (B) by common brushtail possums in the study area. Errors are 95% confidence intervals (CI) of the median. The dotted line corresponds to a critical value of 0.125 (1/n where n is the number of plant species) or no selection (i.e., species selected in proportion to their availability). Any bar above the line signify positive selection or preference for the plant species and any bar below the line signify negative selection or avoidance of the species by common brushtail possums. An asterix indicates significance at P < 0.05
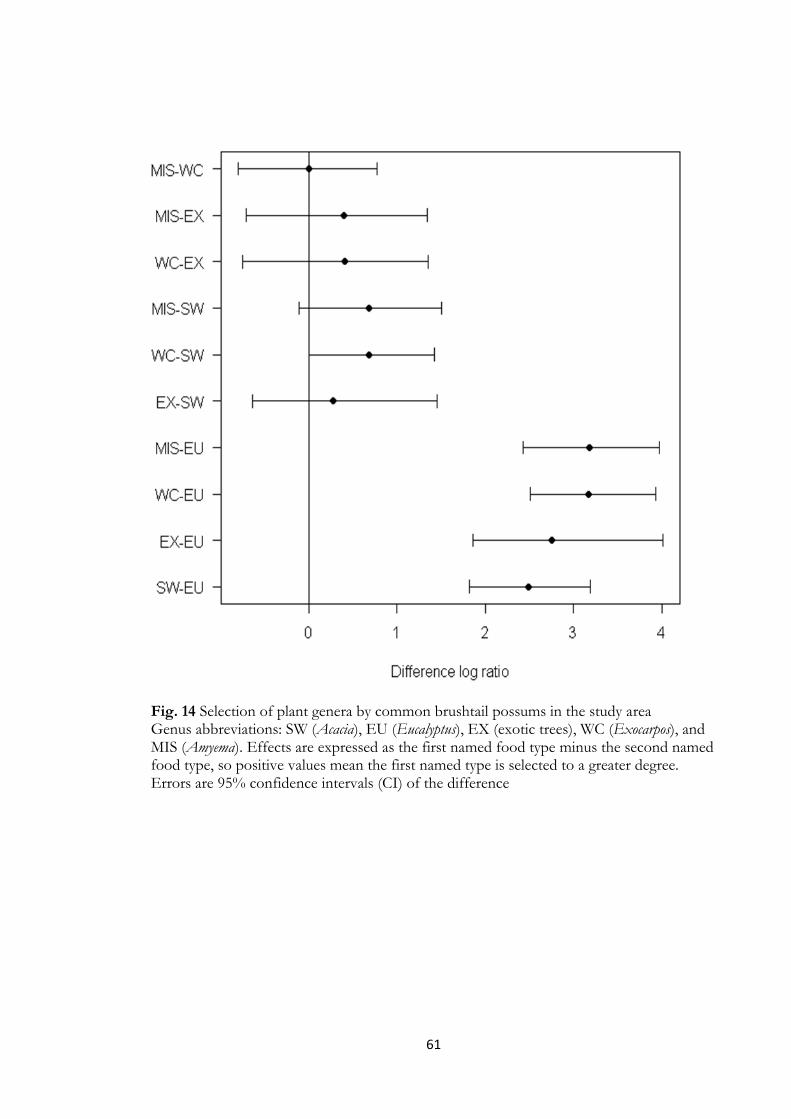
61
Fig. 14 Selection of plant genera by common brushtail possums in the study area Genus abbreviations: SW (Acacia), EU (Eucalyptus), EX (exotic trees), WC (Exocarpos), and MIS (Amyema). Effects are expressed as the first named food type minus the second named food type, so positive values mean the first named type is selected to a greater degree. Errors are 95% confidence intervals (CI) of the difference

62
Determinants of tree use by common brushtail possums
In total, 53 common brushtail possums and 60 mountain brushtail possums were recorded
in the study area. Both species displayed a highly scattered distribution and occupied the
same areas within roadside vegetation corridors (Fig. 15). From ten dominant mid- and
upper-storey species of trees present in the study area, two thirds of sightings of common
brushtail possums were in Eucalyptus camphora (23.7%), E. radiata (21.1%), and E. viminalis
(21.1%). Exotic species of trees were used more often than expected from their availability
in the habitat (15.8%, P < 0.001, Fig. 16). Similarly, mountain brushtail possums were
most often found on E. radiata (34.7%), followed by E. camphora (20.4%), and E. viminalis
(18.4%), which were all used in proportion to their availability (Fig. 16). Surprisingly, Acacia
dealbata (12.2%) was used by mountain brushtail possums less often than expected from its
availability in the habitat (P < 0.05, Fig. 16).
Out of 577 trees recorded in the study area, 11.6% were parasitized by drooping mistletoe
(Amyema pendula) with 4.0 ± 0.5 parasites per tree on average. Mistletoe infection rates were
not uniform across different tree species. Most often mistletoes were found on Eucalyptus
radiata, representing 59.7% of parasitized trees, with on average 3.8 ± 0.7 parasites per tree.
A weak positive correlation was noted between tree diameter and number of mistletoes
(r = 0.24, df = 78, P < 0.05).
Furthermore, almost 12.3% of surveyed trees had hollows large enough to be used as
denning sites by common brushtail possums, with on average 2.4 ± 0.2 hollows per tree.
Tree species with the greatest number of hollows were Eucalyptus radiata (45% of all trees
with hollows, 2.6 ± 0.4 hollows per tree) and E. viminalis (26.7%, 2.5 ± 0.3 hollows per
tree). The number of hollows was positively correlated with tree diameter (r = 0.59, df =
78, P < 0.001).
In the study area, most of the trees were in the small diameter class (5–50 cm; 63.5%),
followed by medium sized trees (50–100 cm, 22.6%) and large trees (> 100 cm, 13.9%).
The species with the largest diameter were E. radiata (69.5 ± 3.8 cm) and E. viminalis (68.3
± 6.2 cm). The medium height of trees in the study area was 11.3 ± 0.2 m and, since height
was highly correlated with tree diameter (r = 0.81, df = 78, P < 0.001), it was omitted from
the further analysis.

63
Fig. 15 Probability density of the common brushtail possums (n = 53) and mountain brushtail possums (n = 60) in the Strathbogie Ranges, Victoria
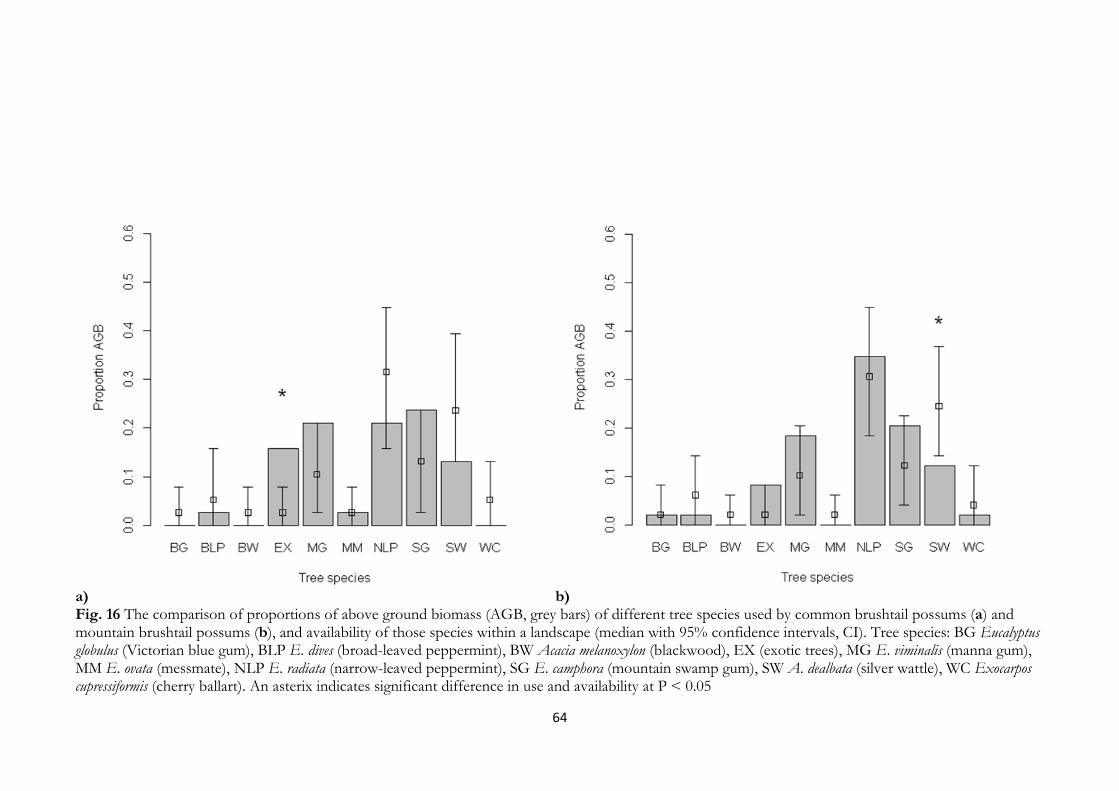
64
a) b) Fig. 16 The comparison of proportions of above ground biomass (AGB, grey bars) of different tree species used by common brushtail possums (a) and mountain brushtail possums (b), and availability of those species within a landscape (median with 95% confidence intervals, CI). Tree species: BG Eucalyptus globulus (Victorian blue gum), BLP E. dives (broad-leaved peppermint), BW Acacia melanoxylon (blackwood), EX (exotic trees), MG E. viminalis (manna gum), MM E. ovata (messmate), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (mountain swamp gum), SW A. dealbata (silver wattle), WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant difference in use and availability at P < 0.05

65
Despite the expectation, there were no significant differences observed in the use of trees
with or without mistletoes by common brushtail possums (χ2 = 0.528, df = 1, P = 0.608).
A similar pattern was observed for mountain brushtail possums (χ2 = 0.803, df = 1,
P = 0.773). Trees with hollows were used by both common and mountain brushtail
possums more often than expected by chance alone (χ2 = 13.778, df = 1, P < 0.001;
χ2 = 3.741, df = 1, P <0.05 respectively). Common brushtail possums more often used
trees larger in diameter (median 71.5 cm; 95% confidence intervals 64.5–79 cm; χ2 =13.778;
df = 1, P < 0.001) than the median diameter of available trees in the study area (28 cm;
95% confidence intervals 23–30 cm). The similar pattern was observed for mountain
brushtail possums (median 54 cm; 95% confidence intervals 35–77 cm; χ2 = 13.067, df = 1,
P < 0.001) Furthermore, no statistically significant differences were detected between
common and mountain brushtail possums in terms of tree use and specific tree traits:
species (χ2 = 6.140, df = 9, P = 0.828), number of mistletoes (W = 875, P = 0.391),
hollows (W = 1049, P = 0.201), and diameter (W = 1063, P = 0.260).
Finally, when taking into account all measured variables, the tree species, height and
diameter, biomass, number of mistletoes and hollows, and the presence of mountain
brushtail possums (T. cunninghami), the binomial generalized linear model indicated that tree
diameter and presence of T. cunninghami were the best variables for predicting tree
occupancy by common brushtail possums (r2 = 0.22, P < 0.001, Tab. 3). The model
explained 76.3% of common brushtail possum tree presences and 76.3% of tree absences.
Table 3 The binomial generalized linear model predicting tree occupancy by common brushtail possums (AIC = 87.73). An asterix indicates significance at P < 0 ‘***’ and P < 0.001 ‘**’ Coefficients Estimate Std. Error z-value P-value Intercept -1.498e+00 4.855e-01 -3.085 0.002** Diameter 2.283e-02 7.343e-03 3.109 0.002** T. cunninghami 1.868e+01 1.974e+03 0.009 0.992 Binomial generalized linear model
df Dev. Resid. df Resid. Dev. P-value Null 75 105.358 Diameter 1 12.117 74 93.242 0.0005*** T. cunninghami 1 11.512 73 81.730 0.0007***

66
Discussion
Summary of key results
In the present study, the common brushtail possum (Trichosurus vulpecula) demonstrated a
generalist feeding strategy consuming a wide variety of plant species and parts. In addition
to different Eucalyptus species, studied animals consumed substantial amounts of foliage
from Acacia species, exotic species of trees, parasitic plants, understorey species, as well as
some fruit and pollen. It is worth noting here, that the root parasite, Exocarpos cupressiformis,
was recorded for the first time in a diet of common brushtail possums. Also, common
brushtail possums were found consuming fruits of an invasive species, Rubus fruticosus.
Nevertheless, despite their generalist feeding strategy common brushtail possums exhibited
clear foraging preferences at multiple scales of resource heterogeneity ranging from forest
types to tree species and parasitic plants within tree canopies. Parasitic plants, Amyema
pendula and Exocarpos cupressiformis, were consumed more frequently than expected from
their availability in the habitat while the dominant Eucalyptus species were consumed less
frequently. Moreover, Acacia species and exotic trees were consumed more often than
expected, but results were not significant.
Common brushtail possums were most often found in Eucalyptus camphora, E. radiata, and
E. viminalis trees during their nightly foraging activity. They used only exotic species of trees
more often than expected from their availability in the habitat. Contrary to expectation,
trees parasitized by mistletoes were not used selectively by common brushtail possums.
A similar pattern of tree use was observed for a sympatric species, the mountain brushtail
possum (T. cunninghami) with animals found most often in canopies of dominant eucalypt
species and hardly ever foraging in trees with mistletoes. Contrary to expectation, mountain
brushtail possums used Acacia dealbata trees less often than expected from their availability
in the habitat. Both common and mountain brushtail possums preferred trees with hollows
and trees larger in diameter. As a result, no significant differences were observed between
trees used by common brushtail and mountain brushtail possums. Tree diameter and
occurrence of mountain brushtail possums were the best variables to explain the observed
patterns of tree use by common brushtail possums.

67
The diet breadth and food preferences of common brushtail possums
Foraging decisions of arboreal marsupials inhabiting eucalypt forests and woodlands are
complex and depend not only on food availability and nutritional quality, but also on
animal ability to locate different types of food and to learn to eat or avoid difficult forage.
In the current study, common brushtail possums had a broad diet and consumed 18
different types of food, including foliage of woody and seral species, as well as fruit and
pollen. Yet, foliage of habitat dominant Eucalyptus species represented less than one quarter
of all consumed forage. Likewise, previous studies demonstrated a great dietary breadth of
common brushtail possums incorporating different groups of plants and plant parts into
their diet (summarized by Kerle, 2001). For instance, a proportion of Eucalyptus foliage in a
diet of common brushtail possums has been found to vary from 90% in eucalypt
woodlands in dry tropics of North Queensland to being completely absent or comprising
less than 10% in wet sclerophyll forests of Tasmania (Fitzgerald, 1984; DeGabriel et al.,
2009b).
Why common brushtail possums consume different amounts of Eucalyptus foliage in
different parts of their range remains unclear, but some general patterns emerge from
recent studies with captive and wild animals. Eucalyptus foliage is known to be low in
nutrients, especially nitrogen (a proxy of protein content), and has high concentrations of
components that interfere with animal digestion and absorption of protein, such as tannins
and fibre (DeGabriel et al., 2008; DeGabriel et al., 2009a). Common brushtail possums are
less effective in digesting fibre than eucalypt specialists, such as the koala and greater glider,
since they lack a mechanism for separating large food particles that allow for subsequent
microbial fermentation (Sakaguchi & Hume, 1990). Unlike common ringtail possums,
common brushtail possums do not practice caecotrophy to maximize nutrient acquisition
and are more sensitive to tannins and associate their astringent taste with post-ingestive
negative effects (Marsh et al., 2003a). Furthermore, eucalypt leaves contain a range of plant
secondary metabolites that are potentially toxic to common brushtail possums, such as
terpenes, cyanogenic glycosides, and formylated phloroglucinol compounds (FPCs),
powerful antifeedants found only in eucalypts from the subgenus Symphyomyrtus (Brophy &
Southwell, 2002; Eschler et al., 2000; Gleadow et al., 2008; Moore et al., 2004).
The inability of common brushtail possums to satisfy their nutritional requirements from
a single species or for that matter to sustain entire populations on a diet composed
exclusively of Eucalyptus foliage was first recognized by Freeland and Winter (1975).

68
The authors observed that while animals spent 66% of their time foraging on eucalypts,
common brushtail possums usually consumed three different foods per night, two species
of Eucalyptus and one non-eucalypt species, in order to reduce the negative effects of any
single toxin. In the current study, common brushtail possums were found consuming an
average of six plant species, including one to three Eucalyptus species with Eucalyptus radiata
from a subgenus Monocalyptus eaten most often (14.1%) and complemented with negligible
amounts of E. camphora (3.8%) and E. viminalis (0.8%) from a subgenus Symphyomyrtus.
The observed high intake of E. radiata by common brushtail possums was unexpected
since this tree species is noted for its high level of phenolics and terpenes approaching up
to 8% in some chemotypes (Foley, 1992). Furthermore, high concentrations of formylated
phloroglucinol compounds (FPCs) have been found to deter possums from foraging on E.
camphora and E. viminalis (Marsh et al., 2003a; Moore et al., 2004a). In a feeding trial
performed by Dearing and Cork (1999), common brushtail possums lost 6.5 ± 1.1 % of
their body weight when offered solely foliage of E. radiata. Yet, animals simultaneously
offered foliage of E. radiata (Monocalyptus) and E. melliodora (Symphyomyrtus) increased their
food intake and consumed 33% more food than on either diet offered singly. The authors
concluded that a diet breadth of common brushtail possums is a product of their
detoxification abilities as these animals are physiologically constrained by their capacity to
absorb, metabolize, and excrete similar types of toxins found in eucalypt foliage.
Recently, DeGabriel and colleagues (2009b) demonstrated that common brushtail possums
occurring in different habitats have developed local physiological and behavioral
adaptations to deal with difficult forage. Contrary to the authors’ expectations, southern
populations inhabiting forests and woodlands dominated by Eucalyptus species proved to be
more sensitive to concentrations of tannins, terpenes, phenolic glycosides, and formylated
phloroglucinol compounds (FPCs) than northern populations. The authors explained these
findings by noting the higher diversity of plant species in southern forests and woodlands
that allowed possums to have broader diets and switch to less difficult forage when
necessary. A positive relationship between availability of forage and a breadth of diet of
common brushtail possums proved to be the case in the current study, where Herb-rich
Foothill Forest and Swampy Riparian Woodland had a well-developed mid-stratum
dominated by Acacia spp. and species-rich understorey composed of native and exotic
species of herbs and grasses.

69
Parasitic plants, Amyema pendula and Exocarpos cupressiformis, were frequently found
parasitizing stems and roots of Eucalyptus and Acacia trees. As a consequence, the studied
common brushtail possum had a broad diet incorporating foliage of Acacia species (9.7%)
with silver wattle (A. dealbata) representing 6.3%, Amyema pendula 6.8%, and Exocarpos
cupressiformis 4.3% while herbs and grasses represented 15.3% of a possum diet.
It is likely that the studied common brushtail possums learned to avoid Eucalyptus foliage to
minimize the negative effects of toxins and selectively foraged on nitrogen-fixing Acacia
species to maximize their nitrogen intake. Yet, Acacia leaves due to their phyllode structure
tend to be fibrous and have high tannin content, and may contain cyanogenic glycosides
and fluroacetate interfering with possum digestion (Irlbeck & Hume, 2003). Fitzgerald
(1984) studied a population of common brushtail possums inhabiting dry sclerophyll
forests in Tasmania where A. dealbata and A. melanoxylon were the most common
understorey species and recorded Acacia foliage (especially A. dealbata) to be present in the
possum diet throughout the year. Similarly, Evans (1992) studied the diet of common
brushtail possums in central arid Australia and noticed that foliage of A. coriacea was a
preferred dietary constituent along with foliage from other Acacia species.
However, in a feeding trial carried out by McArthur and Turner (1997) when common
brushtail possums were offered foliage of acacia species (A. dealbata and A. melanoxylon) and
eucalypt foliage from Monocalyptus and Symphyomyrtus subgenera, animals preferred feeding
on foliage of Symphyomyrtus species and almost completely avoided A. melanoxylon. The
observed diet mixing behaviour could have been a result of nutrient balancing in foraging
decisions of common brushtail possums governed by the nitrogen content of forage.
Raubenheimer and Simpson (2003) demonstrated that a generalist herbivore, the desert
locust (Schistocerca gregaria), selected a diet with a higher nitrogen content. The authors
concluded that the capability of desert locusts to deal with excess ingested protein might be
related to the greater nitrogen content of selected foods rather than their broad dietary
range.
Furthermore, Wiggins and colleagues (2006a, 2008) argued that frequent switching between
chemically diverse foliage reduces physiological constraints imposed by toxin-rich diet and
enables more efficient nutrient acquisition by common brushtail possums. This reduction
in physiological constraints might explain why in the current study common brushtail
possums preferred feeding on parasitic plants over their Eucalyptus and Acacia tree hosts.

70
Similarly, other experimental and field studies have shown that mistletoes from
Loranthaceae family are preferentially eaten by common brushtail possums, common
ringtail possums, koalas, and greater gliders (summarized by Reid 1997).
Mistletoes are considered to be of greater nutritional quality for herbivores as they are rich
in nutrients, have higher foliar moisture content, and are presumably less toxic than their
tree hosts (Watson, 2001, 2011). Foliage of box mistletoe Amyema miquelii, closely related to
drooping mistletoe A. pendula, has been found to have a higher content of potassium and
sodium that can be limiting to some herbivore populations (March & Watson, 2009). Yet,
plant secondary metabolites of genus Amyema remain largely unknown. Both species of
mistletoes were found to contain herbivory-reducing terpenes (Preston & Watson, 2010)
and flavonoid afzelin (Hensens, Lewis & Mulquiney, 1971).
In the present study, diet switching between parasitic plant and their tree hosts could have
helped common brushtail possums to overcome short-term intake constraints resulting
from toxic effects of each species. The foraging decisions of the studied animals might
have been shaped by small-scale heterogeneity of resources with mistletoes representing
alternative food patches within otherwise uniform tree canopies. Namely, Wiggins et al.
(2008) predicted that heterogeneity of plant patches, in relation to distribution of plant
secondary metabolites, and the scale at which this occurs across the landscape, are critical
factors that influence foraging efficiency and fitness of mammalian herbivores.
Furthermore, preferential foraging of common brushtail possums on a root parasite,
Exocarpos cupressiformis (cherry ballart, Santalaceae), deserves special attention as the current
study is the first one to recognize the importance of this species for arboreal marsupials.
Foliage of E. cupressiformis has been previously detected in a diet of the brushtail-tailed rock-
wallaby (Petrogale penicillata, Van Eeden, Di Stefano, & Coulson 2011). The authors argued
that although E. cupressiformis did not appear to form a large portion of wallabies’ diet it
could represent an important highly nutritious component. Yet, no foliar chemical analysis
has been carried out until the present (see Chapter 4 for details). Furthermore, root
parasites from Santalaceae family have been found to sequester some of the plant
secondary metabolites from their hosts, which in turn can negatively affect the foraging of
herbivores (Woldemichael, & Wink, 2002). In the current study, E. cupressiformis parasitized
both nitrogen-fixing acacias and eucalypts containing different groups of plant secondary
metabolites, yet how these host trees affected palatability of E. cupressiformis remains
unknown.

71
In the present study, it was also impossible to distinguish whether common brushtail
possums were “smoothing out their diet” (eating proportionally less of common and more
of rare species), which increases their dietary breadth, or if the foliar chemistry made
Eucalyptus foliage lower quality food for this generalist arboreal marsupial, or if both
reasons together increased their dietary breadth. The experience from sampling different
types of food, such as parasitic plants and transient food aversion to Eucalyptus foliage,
could not be ruled out as a possible explanation for common brushtail possum food
preferences.
Food aversions often called sensory-specific satiety become pronounced when foods
contain high concentrations of toxins or nutrients or both or are nutritionally imbalanced
or deficient (the satiety hypothesis, Provenza et al., 2003). Namely, when animals or
humans are presented with a variety of foods, they prefer an alternative to the one eaten
most recently because interaction among flavours, nutrients, and toxins can cause transient
aversions. Sampling a variety of foods allows animals to learn about the consequences of
ingesting each food or combination of foods enabling them to activate or maintain
different detoxification pathways (Marsh et al., 2006a, Marsh et al., 2006b).
Consequently, in the wild common brushtail possums consume in addition to tree and
mistletoe foliage many different types of foods, such as grasses and forbs, fruits, seeds, and
pollen. In the present study, grasses and forbs represented 15.3% of the possum diet while
fruit and pollen comprised 11% of the total diet. Grasses and forbs were found to
represent 40–60% of the diet of common brushtail possums in dry sclerophyll forests
adjacent to pastures in Tasmania (Fitzgerald, 1984). On this diet digestibility is high
(i.e., cell walls of grasses are low in fibre and plant secondary metabolites) while forbs are
rich in protein, sugars, and lipids (Provenza et al., 2003). Interestingly, although Downey et
al. (1997) found Rubus fruticosus to be relatively rare in the study area, fruits of this invasive
species were readily eaten by common brushtail possums in the present study.
Likewise, in New Zealand seasonally available fruits and seeds of native and exotic species
were found to compromise 70% of a common brushtail possum diet (Dungan et al., 2002).
Williams (1982) compared nutritional properties of some fruits eaten by common brushtail
possums in New Zealand podocarp-forests and concluded that fruits appeared to be a
source of readily digestible energy (rich in carbohydrates and low in fibre), however, they
did not differ from leaves in levels of crude protein, lipids, and minerals.

72
While no such study exists in Australia, the less folivorous and more frugivorous diet of
common brushtail possums compared to other arboreal marsupials’ diets has been
explained by their less specialized dentition and their small body size.
Furthermore, common brushtail possums extended their diet to incorporate several
introduced species, including plantation, ornamental, and orchard trees (Kerle, 2001).
In the current study, exotic species of trees from Europe (Fraxinus angustifolia, Quercus robur
and Prunus cerasifera) and North America (Pinus radiata) represented 17.1% of a possum diet
despite being a minor component in the habitat. Recently, a comprehensive study of
foraging preferences of generalist herbivores from multiple continents demonstrated their
affinity for exotic plants that are evolutionary naïve or lack specific plant secondary
metabolites (Morrison & Hay, 2011). For example, in a feeding trial South American apple
snails (Pomacea sp.) preferred North American macrophytes while North American crayfish
(Procambarus spiculifer) preferred South American, Asian, and Australian macrophytes that
were less chemically defendant than native species.
In the case of introduced herbivores, such as common brushtail possums in New Zealand,
it has been postulated that the degree of diet generalism and access to novel plant species
with a greater nutritional value and lower plant defences may contribute to the successful
invasion and establishment of common brushtail possums (O’Reilly-Wapstra & Cowan,
2010). Dietary plasticity of common brushtail possums probably allowed this generalist
forager to successfully invade broadleaf-podocarp forests and to a lesser degree southern
beech Nothofagus forests in New Zealand. In these forest communities common brushtail
possums are known to eat approximately 100 plant species, yet show strong preferences for
foliage of relatively rare species, such as red mistletoe Peraxilla tetrapetala and yellow
mistletoe Alepis flavida, contributing to mistletoe declines in some areas (Montague, 2000;
Sweetapple et al., 2002; Sweetapple, 2008).
In Australia and even more in New Zealand, common brushtail possums have been found
to supplement their diet with animal sourced protein from arthropods, lizards, bird eggs,
and chicks, and opportunistically scavenge on animal carcasses (Brown, Innes, & Shorten,
1993; Cowan & Moeed, 1987). In the present study, it was hard to distinguish animal
material in possum faeces since this would require a different methodological approach,
such as establishing differences between nitrogen and carbon isotope ratios among
consumed foods and possum body tissues.

73
Nonetheless, it can be speculated that out of 14% of unidentified forage material some part
was of animal origin while the rest was finely digested plant material with no recognizable
cell structures. Different digestibility of plant material could have underestimated the
proportion of seral species, such as herbs and grasses in a diet of the studied common
brushtail possums. The underestimated proportion of seral species could have been
partially accounted for if smaller sieves than the ones used in this study were utilized to
retain finer leaf fragments for further identification.
Also, the relative importance of fruits, flowers, and pollen was difficult to quantify using
the faecal content analysis method. Difficulties in identifying the cell structure of fruits can
partially explain why mistletoe fruits, favoured by possums in some areas (Reid, 1997), were
not detected in faeces of the studied animals. By contrast, it was relatively easy to identify
the well preserved cuticles of different Eucalyptus species and establish their share in the
overall diet of common brushtail possums. Differential digestion may be accounted for
using correction factors developed from regression equations that compensate over- or
underestimation of cuticle types (Dearden, Pegau, & Hansen, 1975). Correction factors are
produced by feeding captive animals known quantities of forage and then assessing the
relative quantity of each species present in their faeces. This method proved to be
unfeasible in the current study due to ethical and logistic constraints associated with
capture and husbandry of common brushtail possums.
Determinants of tree use by common brushtail possums
For arboreal marsupials spending most of their lives in tree canopies, trees represent food,
shelter, and a stage upon which interactions with conspecifics, competitors, and predators
take place. This recognized polygonal use of trees by arboreal marsupials represents a
challenge for researchers trying to single out reasons for the selection of some trees and the
avoidance of others. In the present study, food choices of common brushtail possum were
expected to be reflected in their tree use measured against tree species availability in the
habitat. From ten dominant mid- and upper-storey tree species, two-thirds of the sightings
of common brushtail possums were in Eucalyptus camphora (23.7%), E. radiata (21.1%), and
E. viminalis (21.1%), yet eucalypt species represented just 21.6% of a possum diet and were
avoided by foraging animals.

74
Only exotic species of trees were used more often by common brushtail possums than
expected from their availability in the habitat (15.8%) and this was also reflected in their
diet with exotic trees representing 17.1% of all consumed species. These findings suggest
that common brushtail possums could have used exotic trees exclusively as foraging sites.
Conversely, Eucalyptus trees could have been used by common brushtail possums for other
reasons, including presence of alternative food patches of mistletoes, occurrence of
hollows used as shelter and concealment from predators, and a tree size or branches big
enough to support animal body weight.
However, against expectations common brushtail possums did not selectively use trees
parasitized by mistletoes. This finding challenges the results from the faecal content
analysis showing that common brushtail possums preferentially foraged on Amyema pendula
over its Eucalyptus and Acacia tree hosts. There are many possible and often competing
explanations for these seemingly contradictory results. First, nocturnal marsupials use
visual cues to locate food, at least from a distance, and have monochromatic vision;
therefore, they are unlikely to be able to detect differences in foliage colour between
mistletoes and their tree hosts (Reid & Yan, 2000). Moreover, some species of mistletoes
tend to resemble their tree hosts in appearance presumably to avoid being eaten by
herbivores (Barlow & Wiens, 1997). It has been argued that mistletoe host-resemblance is a
type of cryptic-mimicry where mistletoes increase their fitness by blending into its
vegetative background under the selective pressure exerted by arboreal marsupials.
The question therefore arises as to how common brushtail possums find mistletoes in tree
canopies. Can they similar to primates remember locations of a wide variety of plant foods
and move directly to such foods when and where available without wasting time and
energy searching randomly? There are no studies known to the author measuring the
cognitive abilities of arboreal marsupials. From the nutritional perspective, it can be argued
that a special “taste” of common brushtail possums for mistletoes is the outcome of their
generalist feeding strategy. Animals might consume mistletoes opportunistically when they
forage on eucalypts. For instance, in the current study Eucalyptus radiata represented a major
component of possum diet. Also, E. radiata was parasitized by A. pendula most often and
had the highest number of mistletoes per tree. High mistletoe availability within E. radiata
canopies could have created a specific preference for A. pendula and promoted diet
switching behaviour in possums.

75
Furthermore, tree selection by common brushtail could have been affected by the
occurrence of a sympatric species, the mountain brushtail possum, as both species did not
vary in their use of trees. Furthermore, these sympatric species were never found sharing
the same tree. These findings were confirmed by another study looking at foraging patterns
of common brushtail possums and mountain brushtail possums in the same study area
(Del Borrello 2009). The author suggested that studying these species both in sympatry and
allopatry could reveal differences in tree use and explore underlying mechanisms. Namely,
if in allopatry common brushtail possums would show preferences for a certain tree, as has
been previously demonstrated, than the presence of a competitor might have altered their
foraging behaviour.
However, a resource overlap does not necessary indicate competition as revealed in a study
by Lindenmayer and Cunningham (1997) that looked at patterns of co-occurrence of
arboreal marsupials in the montane ash forests of Victoria. In these forests mountain
brushtail possums were found to co-occur with greater gliders, another strictly folivorous
species. In the current study, koalas were found often foraging on Eucalyptus viminalis, a
species that represented just 0.8% of the studied common brushtail possums’ diet. While
koalas do not appear to be limited by interspecific competition (Norton & Neave, 1990), it
remains to be tested whether they exclude other smaller species from foraging on E.
viminalis.
The other important tree characteristic that might have affected possum tree selection was
the presence of suitable size hollows used as shelter and concealment from predators.
Namely, trees with hollows were preferred by common brushtail possums, with E. radiata
and E. viminalis having the highest hollow number per tree. Johnson et al. (2001)
determined that at high population densities, hollow availability was a limiting factor for
populations of common brushtail possum in mainland Queensland and New South Wales.
Also, a certain level of competition over hollow-bearing trees could have been exhibited
through the sympatric mountain brushtail possums. However, no pattern was observed
when comparing differences in hollow bearing trees used by two species of possums
indicating low population numbers, increased dispersal rates, or high availability of hollows
in the habitat.
Del Borrello (2009) working in the same study area found no evidence for competition
over hollow bearing trees and exclusion of common brushtail possums from suitable
denning sites by mountain brushtail possums.

76
The studied roadside vegetation corridors have been recognized as sustaining an
exceptionally high number of trees with hollows due to the fact that they have not been
logged for more than 100 years (Martin & Martin 2004; Martin & Martin, 2007).
Differences in body size with mountain brushtail possums being larger than common
brushtail possums, hence requiring larger size hollows, could have alleviated competition
between the sympatric species and facilitated their co-existence.
In the current study, both common brushtail possums and mountain brushtail possums
were more often found on larger trees exceeding 50 cm in diameter. A similar pattern was
observed for the specialist herbivore, the koala (Moore & Foley, 2005). Koalas visited trees
with a wider diameter significantly more often than expected from their availability in the
habitat as these trees provided larger patches of food and accounted for a greater
proportion of available biomass (Moore & Foley, 2005). Larger trees are also known to
sustain higher mistletoe populations (Reid & Lange, 1988) and have a higher number of
hollows than trees smaller in diameter (< 50 cm; Gibbons & Lindenmayer, 2002).
Moreover, larger trees are suitable for bird nesting and common brushtail possums might
opportunistically feed on eggs and chicks to obtain extra protein as shown in New Zealand
studies (Brown, Innes, & Shorten 1993; Brown, Moller, & Innes, 1996). Furthermore, by
foraging in larger trees and moving along branches of neighbouring trees, common
brushtail possums might have reduced the risk of predation by avoiding travelling on the
ground between patchily distributed resources. In the study area, the high activity of
ground predators, such as foxes and feral cats, was frequently observed, which might have
influenced common brushtail possums to seek refuge high in tree canopies.
Finally, it is possible that the presence of observers while spotlighting might have altered
the foraging behaviour of the studied animals driving them off the ground and into trees.
Also, the observed discrepancies in tree species selection and consumption could reflect
limitations of spotlighting as a method for establishing food preferences of arboreal
herbivores. Recorded high possum visitations to E. camphora trees could be an artefact of
their open canopy structure enabling easy detection in comparison with the more dense
canopies of E. radiata and E. viminalis. Some of the observations could also reflect animals
moving between feeding locations.

77
Hence, to disentangle the real reasons for selecting some trees and avoiding others, it is
recommended to carry out focal observations of individuals while foraging and establish
their denning sites using radio-telemetry (see Chapter 3). The present study has
demonstrated that foraging decisions of common brushtail possums are complex and
depend not only on food availability and nutritional quality, but also interplay between
many ecological factors: availability of shelter, competition, and predation. An integrative
approach is needed to better understand why the world is green and full of herbivores and
how animals balance the trade-offs between eating and being eaten.

78
References
Alldredge, M. W., Peek, J. M., & Wall, W. A. (2002). Nutritional quality of forages used by elk in northern Idaho. Journal of Range Management, 55(3), 253–259.
Allen, R. B., Fitzgerald, A. E., & Efford, M. G. (1997). Long-term changes and seasonal patterns in possum (Trichosurus vulpecula) leaf diet, Orongorongo Valley, Wellington, New Zealand. New Zealand Journal of Ecology, 21(2), 181–186.
Andrew, R. L., Peakall, R., Wallis, I. R., & Foley, W. J. Spatial distribution of defense chemicals and markers and the maintenance of chemical variation. Ecology, 88(3), 716–728.
Ball, J. P., Danell, K., & Sunesson, P. (2001). Response of a herbivore community to increased food quality and quantity: An experiment with nitrogen fertilizer in a boreal forest. Journal of Applied Ecology, 37(2), 247–255.
Barlow, B. A., & Wiens, D. (1977). Host-parasite resemblance in Australian mistletoes: The case for cryptic mimicry. Evolution, 31(1), 69–84.
Belovsky, G. E. (1978). Diet optimization in a generalist herbivore: The moose. Theoretical Population Biology, 14(1), 105–134.
Bennett, A. F., Lumsden, L. F., Alexander, J. S. A., Duncan, P. E., Johnson, P. G., Robertson, P., & Silveira, C. E. (1991). Habitat use by arboreal mammals along an environment gradient in north-eastern Victoria. Wildlife Research, 18(2), 125–146.
Bjorndal, K. A. (1989). Flexibility of digestive responses in two generalist herbivores, the tortoises Geochelone carbonaria and Geochelone denticulata. Oecologia, 78(3), 317–321.
Braithwaite, L. W., Binns, D. L., & Nowlan, R. D. (1988). The distribution of arboreal marsupials in relation to eucalypt forest types in the Eden (NSW) woodchip concession area. Australian Wildlife Research, 15(4), 363–373.
Braithwaite, L. W., Dudzinski, M. L., & Turner, J. (1983). Studies on the arboreal marsupial fauna of eucalypt forests being harvested for woodpulp at Eden, N.S.W. II. Relationship between the fauna density, richness and diversity, and measured variables of the habitat. Australian Wildlife Research, 10(2), 231–247.
Braithwaite, L. W., Turner, J., & Kelly, J. (1984). Studies on the Arboreal Marsupial Fauna of Eucalypt Forests Being Harvested for Wood Pulp at Eden, N.S.W. Iii. Relationships Between Faunal Densities, Eucalypt Occurrence and Foliage Nutrients, and Soil Parent Materials. Australian Wildlife Research, 11(1), 41–48.
Brophy, J. J., & Southwell, I. A. (2002). Eucalyptus chemistry. In J. J. W. Coppen (Ed.), Eucalyptus: The Genus Eucalyptus (pp. 102–161). London: Taylor & Francis.
Brown, K., Innes, J., & Shorten, R. (1993). Evidence that possums prey on and scavenge birds’ eggs, birds and mammals. Notornis, 40(3), 169–177.
Brown, K., Moller, H., & Innes, J. (1996). Sign left by brushtail possums after feeding on bird eggs and chicks. New Zealand Journal of Ecology, 20(2), 277–284.
Burchfield, E., Agar, N. S., Hume, I. D. (2005). Effects of terpenes and tannins on some physiological and biochemical parameters in two species of phalangerid possums (Marsupialia: Phalangeriadae). Australian Journal of Zoology, 53(6), 395–402.
Bureau of Meteorology, Strathbogie. (2013). Summary climate statistics. Retrieved 13th February, 2013, from http://www.bom.gov.au/climate/averages/tables/cw_082042.shtml
Burnham, K. P., & Anderson, D. R. (2002). Model Selection and Multimodel Inference: A practical Information-Theoretic Approach (2 ed.). New York: Springer.
Carr, G. W., Moysey, E. D., Mathews, S., Frood, D., White, M., Griffioen, P., Rosengren, N. (2006). The location of Spring Soak and Peatland wetlands in the Goulburn Broken Catchment—Wetland Implementation Plan Stage. A report to the Goulburn Broken Catchment Management Authority.

79
Catchment Scale Land Use Map. (2010). Retrieved from http://adl.brs.gov.au/landuse/index.cfm?fa=main.downloadData
Charnov, E. L. (1976). Optimal foraging, the marginal value theorem. Theoretical Population Biology, 9(2), 129-136.
Cork, S. J. (1990). Digestive physiology of koalas. Proceedings of the Australian Physiological and Pharmacological Society, 21(2), 95–104.
Cork, S. J., & Foley, W. J. (1991). Digestive and metabolic strategies of arboreal mammalian folivores in relation to chemical defenses in temperate and tropical forests. In R. T. Palo & C. T. Robbins (Eds.), Plant defenses against mammalian herbivory (pp. 133–166). Boca Raton: CRC Press.
Cork, S. J., & Foley, W. J. (1997). Digestive and metabolic adaptations of arboreal marsupials for dealing with plant antinutrients and toxins. In N. R. Saunders & L. A. Hinds (Eds.), Marsupial Biology—Recent Research, New Perspectives (pp. 204-226). Sydney: UNSW Press.
Cowan, P. (1990). Fruits, seeds, and flowers in the diet of brushtail possums, Trichosurus vulpecula, in lowland podocarp/mixed hardwood forest, Orongorongo Valley, New Zealand. New Zealand Journal of Zoology, 17(4), 549-566.
Cowan, P. E. (1989). A vesicular-arbuscular fungus in the diet of brushtail possums, Trichosurus vulpecula. New Zealand Journal of Botany, 27(1), 129–131.
Cowan, P. E., & Moeed, A. (1987). Invertebrates in the diet of brushtail possums, Trichosurus vulpecula, in lowland podocarp/broadleaf forest, Orongorongo Valley, Wellington, New Zealand. New Zealand Journal of Zoology, 14(2), 163–177.
Crawley, M. J. (1983). Herbivory: The dynamics of animal—plant interactions. Berkeley: University of California Press.
Dai, X., Jia, X., Zhang, W., Bai, Y., Zhang, J., Wang, Y., & Wang, G. (2009). Plant height–crown radius and canopy coverage–density relationships determine above-ground biomass—density relationship in stressful environments. Biology Letters, 5(4), 571–573.
Dearden, B. L., Pegau, R. E., & Hansen, R. M. (1975). Precision of microhistological estimates of ruminant food habits. The Journal of Wildlife Management, 39: 402–407.
Dearing, M. D. (1996). Disparate determinants of summer and winter diet selection of a generalist herbivore, Ochotona princeps. Oecologia, 108(3), 467–478.
DeGabriel, J. L., Wallis, I. R., Moore, B. D., & Foley, W. J. (2008). A simple, integrative assay to quantify nutritional quality of browses for herbivores. Oecologia, 156(1), 107–116.
DeGabriel, J. L., Moore, B. D., Foley, W. J., & Johnson, C. N. (2009a). The effects of plant defensive chemistry on nutrient availability predict reproductive success in a mammal. Ecology, 90(3), 711–719.
DeGabriel, J. L., Moore, B. D., Shipley, L. A., Krockenberger, A. K., Wallis, I. R., Johnson, C. N., & Foley, W. J. (2009b). Inter-population differences in the tolerance of a marsupial folivore to plant secondary metabolites. Oecologia, 161(3), 539–548.
DeGabriel, J. L., Moore, B. D., Marsh, K. J., & Foley, W. J. (2010). The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology, 20(2), 97–108.
DeGabriel, J. L., Moore, B. D., Felton, A. M., Ganzhorn, J. U., Stolter, C., Wallis, I. R., Johnson, C. N., & Foley, W. J (2013). Translating nutritional ecology from the laboratory to the field: Milestones in linking plant chemistry to population regulation in mammalian browsers. Oikos, 123(3), 298–308.
Del Borrello, C. (2009). Home-range and habitat use in sympatric brushtail possums (Trichosurus spp.). (Honours thesis), Melbourne University, Melbourne.
Di Stefano, J., & Newell, G. R. (2008). Diet selection by the swamp wallaby (Wallabia bicolor): Feeding strategies under conditions of changed food availability. Journal of Mammalogy, 89(6), 1540–1549.

80
Digital Elevation Model (DEM-S) Version 1.0. (2002). Retrieved from http://glcf.umd.edu/data/srtm
Downes, S. J., Handasyde, K. A., & Elgar, M. A. (1997). The use of corridors by mammals in fragmented Australian eucalypt forests. Conservation Biology, 11(3), 718–726.
Duffy, J. E., & Hay, M. E. (1991). Food and shelter as determinants of food choice by an herbivorous marine amphipod. Ecology, 72(4), 1286–1298.
Dungan, R. J., O’Cain, M. J., Lopez, M. L., & Norton, D. A. (2002). Contribution by possums to seed rain and subsequent seed germination in successional vegetation, Canterbury, New Zealand. New Zealand Journal of Ecology, 26(2), 121–128.
Dunnet, G. M., Harvie, A. E., & Smit, T. J. (1973). Estimating the proportions of various leaves in the diet of the opossum, Trichosurus vulpecula Kerr, by faecal analysis. Journal of Applied Ecology, 10(3), 737–745.
Ellis, W., Carrick, F., Lundgren, P., Veary, A., & Cohen, B. (1999). The use of faecal cuticle examination to determine the dietary composition of koalas. Australian Zoologist, 31, 127–133.
Enquist, B. J., & Niklas, K. J. (2002). Global allocation rules for patterns of biomass partitioning in seed plants. Science, 295(5559), 1517–1520.
Eschler, B. M., Pass, D. M., Willis, R., & Foley, W. J. (2000). Distribution of foliar formylated phloroglucinol derivatives amongst Eucalyptus species. Biochemical Systematics and Ecology, 28(9), 813–824.
ESRI, ArcGIS Desktop 9.3. (2009). Redlands, CA: Environmental Systems Resource Institute.
ESRI, ArcPad 8.0. (2009). Redlands, CA: Environmental Systems Resource Institute. Evans, M. C. (1992). Diet of the brushtail possum Trichosurus vulpecula (Marsupialia:
Phalangeridae) in central Australia. Australian Mammalogy, 15, 25–30. Eymann, J., Herbert, C., & Cooper, D. (2006). Management issues of urban common
brushtail possums Trichosurus vulpecula: A loved or hated neighbour. Australian Mammalogy, 28(2), 153–171.
Felton, A. M., Felton, A., Raubenheimer, D., Simpson, S. J., Foley, W. J., Wood, J. T., Lindenmayer, D. B. (2009). Protein content of diets dictates the daily energy intake of a free-ranging primate. Behavioral Ecology, 20(4), 685–690.
Fitzgerald, A. E. (1984). Diet of the possum (Trichosurus vulpecula) in three Tasmanian forest types and its relevance to the diet of possums in New Zealand forests. In A. P. Smith & I. D. Hume (Eds.), Possums and gliders (pp. 137–143). Sydney: Surrey Beatty & Sons.
Fitzgerald, A. E., & Waddington, D. C. (1979). Comparison of two methods of fecal analysis of herbivore diet. The Journal of Wildlife Management, 43(2), 468–473.
Fitzgerald, B. M., & Gibb, J. (2001). Introduced mammals in a New Zealand forest: Long-term research in the Orongorongo Valley. Biological Conservation, 99(1), 97–108.
Fox, L. R., & Morrow, P. A. (1983). Estimates of damage by herbivorous insects on Eucalyptus trees. Australian Journal of Ecology, 8(2), 139–147.
Freeland, W. J., & Janzen, D. H. (1974). Strategies in herbivory by mammals: The role of plant secondary compounds. The American Naturalist, 108(961), 269–287.
Freeland, W. J., & Winter, J. W. (1975). Evolutionary consequences of eating:Trichosurus vulpecula (Marsupialia) and the genus Eucalyptus. Journal of Chemical Ecology, 1(4), 439–455.
Gibbons, P., & Lindenmayer, D. (2002). Tree hollows and wildlife conservation in Australia. Collingwood: CSIRO Publishing.
Gleadow, R. M., Haburjak, J., Dunn, J. E., Conn, M. E., & Conn, E. E. (2008). Frequency and distribution of cyanogenic glycosides in Eucalyptus L'Hérit. Phytochemistry, 69(9), 1870–1874.

81
Grant, C. C., & Scholes, M. C. (2006). The importance of nutrient hot-spots in the conservation and management of large wild mammalian herbivores in semi-arid savannas. Biological Conservation, 130(3), 426–437.
Green, W. (1985). A review of ecological studies relevant to management of the common brushtail possum: New Zealand Forest Service.
Green, W. Q., & Coleman, J. D. (1986). Movement of possums(Trichosurus vulpecula) between forest and pasture in Westland, New Zealand: Implications for bovine tuberculosis transmission. New Zealand Journal of Ecology, 9, 57–70.
Hardy, A. C. (1967). Great waters: A voyage of natural history to study whales, plankton and the waters of the Southern Ocean in the old Royal Research Ship "Discovery". London: Collins.
Hergt, J. M., Phillips, G. N., & Ely, K. S. (2002). Strathbogie Igneous Complex, central Victoria. In G. N. Phillips & K. S. Ely (Eds.), Victoria Undercover: Benalla 2002 Conference Proceedings and Field guide: Collaborative Geoscience in Northern Victoria (pp. 43–50). Collingwood: CSIRO Publishing.
Hindell, M. A., Handasyde, K. A., & Lee, A. K. (1985). Tree species selection by free-ranging koala populations in Victoria. Wildlife Research, 12(2), 137–144.
How, R., & Hillcox, S. (2000). Brushtail possum, Trichosurus vulpecula, populations in south-western Australia: Demography, diet and conservation status. Wildlife Research, 27(1), 81–89.
Hume, I. D. (1999). Marsupial nutrition. Cambridge: Cambridge University Press. Hunter, M. D., & Price, P. W. (1992). Playing chutes and ladders: Heterogeneity and the
relative roles of bottom-up and top-down forces in natural communities. Ecology, 73(3), 723–732.
Irlbeck, N. A., & Hume, I. D. (2003). The role of Acacia in the diets of Australian marsupials ? A review. Australian Mammalogy, 25(2), 121–134.
Johnson, A. R., Wiens, J. A., Milne, B. T., & Crist, T. O. (1992). Animal movements and population dynamics in heterogeneous landscapes. Landscape Ecology, 7(1), 63–75.
Johnson, C. N., Clinchy, M., Taylor, A. C., Krebs, C. J., Jarman, P. J., Payne, A., & Ritchie, E. G. (2001). Adjustment of offspring sex ratios in relation to the availability of resources for philopatric offspring in the common brushtail possum. Proceedings of the Royal Society of London. Series B: Biological Sciences, 268(1480), 2001–2005.
Jones, K. M. W., & Krockenberger, A. K. (2007). Determining the diet of cryptic folivores: An assessment of diet analysis techniques using the green ringtail possum (Pseudochirops archeri) as a case study. Wildlife Research, 34(5), 352–358.
Jones, K. M. W., MacLagan, S. J., & Krockenberger, A. K. (2006). Diet selection in the green ringtail possum (Pseudochirops archeri): A specialist folivore in a diverse forest. Australian Ecology, 31(7), 799–807.
Kauffman, M. J., Varley, N., Smith, D. W., Stahler, D. R., MacNulty, D. R., & Boyce, M. S. (2007). Landscape heterogeneity shapes predation in a newly restored predator–prey system. Ecology Letters, 10(8), 690–700.
Keith, H., Barrett, D., & Keenan, R. (2000). Review of allometric relationships for estimating woody biomass for New South Wales, the Australian Capital Territory, Victoria, Tasmania, and South Australia National Carbon Accounting System Technical Report 5B. Canberra: Australian Greenhouse Office.
Kerle, J. A. (2001). Possums: The brushtails, ringtails and greater glider. Sydney: UNSW Press. Kerle, J. A. (1984). Variation in ecology of Trichosurus: Its adaptive significance. In A. P.
Smith & I. D. Hume (Eds.), Possums and gliders (pp. 115–128). Sydney: Surrey Beatty & Sons.
Ketterings, Q. M., Coe, R., van Noordwijk, M., Ambagau, Y., & Palm, C. A. (2001). Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology and Management, 146(1), 199–209.

82
Kirmani, S. N., Banks, P. B., & McArthur, C. (2010). Integrating the costs of plant toxins and predation risk in foraging decisions of a mammalian herbivore. Oecologia, 164(2), 349–356.
Lawler, I. R., Foley, W. J., Eschler, B. M., Pass, D. M., & Handasyde, K. (1998). Intraspecific variation in Eucalyptus secondary metabolites determines food intake by folivorous marsupials. Oecologia, 116(1), 160–169.
Li, H., Madden, J. L., & Potts, B. M. (1995). Variation in volatile leaf oils of the Tasmanian Eucalyptus species—1. Subgenus Monocalyptus. Biochemical Systematics and Ecology, 23(3), 299–318.
Lindenmayer, D. B., & Cunningham, R. B. (1997). Patterns of co-occurrence among arboreal marsupials in the forests of central Victoria, southeastern Australia. Australian Journal of Ecology, 22(3), 340–346.
Loney, P. E., McArthur, C., Potts, B. M., & Jordan, G. J. (2006). How does ontogeny in a Eucalyptus species affect patterns of herbivory by Brushtail Possums? Functional Ecology, 20(6), 982–988.
MacLennan, D. G. (1984). The feeding behaviour and activity patterns of the brushtail possum, Trichosurus vulpecula, in an open eucalypt woodland in Southeast Queensland. In A. P. Smith & I. D. Hume (Eds.), Possums and gliders (pp. 155–161). Sydney: Surrey Beatty & Sons.
Mangione, A. M., Dearing, M. D., & Karasov, W. H. (2000). Interpopulation differences in tolerance to creosote bush resin in desert woodrats (Neotoma lepida). Ecology, 81(8), 2067–2076.
Manly, B. F. J., McDonald, L. L., Thomas, D. L., McDonald, T. L., & Erickson, W. P. (2002). Resource selection by animals: Statistical design and analysis for field studies. London: Kluwer Academic Pub.
March, W. A., & Watson, D. M. (2007). Parasites boost productivity: Effects of mistletoe on litterfall dynamics in a temperate Australian forest. Oecologia, 154(2), 339–347.
March, W. A., & Watson, D. M. (2010). The contribution of mistletoes to nutrient returns: Evidence for a critical role in nutrient cycling. Austral Ecology, 35(7), 713–721.
Maron, J. L., & Crone, E. (2006). Herbivory: Effects on plant abundance, distribution and population growth. Proceedings of the Royal Society B: Biological Sciences, 273(1601), 2575–2584.
Marsh, K. J., Foley, W. J., Cowling, A., & Wallis, I. R. (2003a). Differential susceptibility to Eucalyptus secondary compounds explains feeding by the common ringtail (Pseudocheirus peregrinus) and common brushtail possum (Trichosurus vulpecula). Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 173(1), 69–78.
Marsh, K. J., Wallis, I. R., & Foley, W. J. (2003b). The effect of inactivating tannins on the intake of Eucalyptus foliage by a specialist Eucalyptus folivore (Pseudocheirus peregrinus) and a generalist herbivore (Trichosurus vulpecula). Australian Journal of Zoology, 51(1), 31–42.
Marsh, K. J., Wallis, I. R., Andrew, R. L., & Foley, W. J. (2006a). The detoxification limitation hypothesis: Where did it come from and where is it going? Journal of Chemical Ecology, 32(6), 1247–1266.
Marsh, K. J., Wallis, I. R., McLean, S., Sorensen, J. S., & Foley, W. J. (2006b). Conflicting demands on detoxification pathways influence how common brushtail possums choose their diets. Ecology, 87(8), 2103–2112.
Martin, J. K., & Martin, A. A. (2004). Wildlife conservation beyond the parks: The Strathbogie Ranges. In D. Lunney (Ed.), Conservation of Australia's Forest Fauna (2nd ed., pp. 508–512). Mosman: Royal Zoological Society of New South Wales.
Martin, J. K., & Martin, A. A. (2007). Resource distribution influences mating system in the bobuck (Trichosurus cunninghami: Marsupialia). Oecologia, 154(1), 227–236.

83
McArthur, C., & Turner, S. (1997). Feeding preferences of captive brushtail possums for eucalypt and acacia foliage. Tasforests Hobart, 9, 155–162.
McArthur, C., Goodwin, A., & Turner, S. (2000). Preferences, selection and damage to seedlings under changing availability by two marsupial herbivores. Forest Ecology and Management, 139(1-3), 157–173.
McArthur, C., Hagerman, A., & Robbins, C. T. (1991). Physiological strategies of mammalian herbivores against plant defenses. In R. T. Palo & C. T. Robbins (Eds.), Plant Defenses against Mammalian Herbivores (pp. 103–114). Boca Raton: CRC Press.
McLean, S., & Foley, W. J. (1997). Metabolism of Eucalyptus terpenes by herbivorous marsupials. Drug Metabolism Reviews, 29(1–2), 213–218.
Montague, T. L. (2000). The brushtail possum: Biology, impact and management of an introduced marsupial. Lincoln: Manaaki Whenua Press.
Moore, B. D., & Foley, W. J. (2000). A review of feeding and diet selection in koalas (Phascolarctos cinereus). Australian Journal of Zoology, 48(3), 317–333.
Moore, B. D., Wallis, I. R., Marsh, K. J., & Foley, W. J. (2004a). The role of nutrition in the conservation of the marsupial folivores of eucalypt forests. In D. Lunney (Ed.), Conservation of Australia’s forest fauna (2 ed., pp. 549–575). Mosman: Royal Zoological Society of New South Wales.
Moore, B. D., Wallis, I. R., Palá-Paúl, J., Brophy, J. J., Willis, R. H., & Foley, W. J. (2004b). Antiherbivore chemistry of Eucalyptus—cues and deterrents for marsupial folivores. Journal of Chemical Ecology, 30(9), 1743–1769.
Moore, B. D., Wallis, I. R., Wood, J. T., & Foley, W. J. (2004c). Foliar nutrition, site quality, and temperature influence foliar chemistry of tallowwood (Eucalyptus microcorys). Ecological Monographs, 74(4), 553–568.
Moore, B. D., & Foley, W. J. (2005). Tree use by koalas in a chemically complex landscape. Nature, 435(7041), 488–490.
Moore, B. D., Lawler, I. R., Wallis, I. R., Beale, C. M., & Foley, W. J. (2010). Palatability mapping: A koala's eye view of spatial variation in habitat quality. Ecology, 91(11), 3165–3176.
Morrison, W. E., & Hay, M. E. (2011). Herbivore preference for native vs. exotic plants: Generalist herbivores from multiple continents prefer exotic plants that are evolutionary naïve. PLoS One, 6(3), e17227.
Nersesian, C. L., Banks, P. B., & McArthur, C. (2011). Titrating the cost of plant toxins against predators: Determining the tipping point for foraging herbivores. Journal of Animal Ecology, 80(4), 753–760.
Nichols, W. F., Killingbeck, K. T., & August, P. V. (1998). The Influence of Geomorphological Heterogeneity on Biodiversity II. A Landscape Perspective. Conservation Biology, 12(2), 371–379.
Noble, I. R. (1989). Ecological traits of the Eucalyptus L’Hérit. Subgenera Monocalyptus and Symphyomyrtus. Australian Journal of Botany, 37(3), 207–224.
Norton, T. W, & Neave, H. M. (1990). Koala habitat: A conceptual functional model. In G. Gordon (Ed.), Koalas: Research for Management (pp. 93-101). Brisbaine: World Koala Research Inc.
O’Reagain, P. J., & Schwartz, J. (1995). Dietary selection and foraging strategies of animals on rangeland. Coping with spatial and temporal variability. Paper presented at the International Symposium on the Nutrition of Herbivores.
O’Reilly-Wapstra, J. M., & Cowan, P. (2010). Native plant/herbivore interactions as determinants of the ecological and evolutionary effects of invasive mammalian herbivores: The case of the common brushtail possum. Biological Invasions, 12(2), 373–387.
Pacala, S. W., & Roughgarden, J. (1982). Spatial heterogeneity and interspecific competition. Theoretical Population Biology, 21(1), 92–113.

84
Pass, D. M., Foley, W. J., & Bowden, B. (1998). Vertebrate herbivory on Eucalyptus—identification of specific feeding deterrents for common ringtail possums (Pseudocheirus peregrinus) by bioassay-guided fractionation of Eucalyptus ovata foliage. Journal of Chemical Ecology, 24(9), 1513–1527.
Pausas, J. G., Braithwaite, L. W., & Austin, M. P. (1995). Modelling habitat quality for arboreal marsupials in the south coastal forests of New South Wales, Australia. Forest Ecology and Management, 78(1-3), 39–49.
Phillips, G. N., Ely, K. S., Hergt, J. M., Cownley, D. G., Paul, B., & Dickins, C. (2002). 3-dimensional geometry of the Strathbogie batholith. In G. N. Phillips & K. S. Ely (Eds.), Victoria Undercover: Benalla 2002 Conference Proceedings and Field guide: Collaborative Geoscience in Northern Victoria (pp. 55–60). Collingwood: CSIRO Publishing.
Provenza, F. D., Villalba, J. J., Dziba, L. E., Atwood, S. B., & Banner, R. E. (2003). Linking herbivore experience, varied diets, and plant biochemical diversity. Small Ruminant Research, 49(3), 257–274.
Provenza, F. D., Villalba, J. J., Wiedmeier, R., Lyman, T., Owens, J., Lisonbee, L.,… & Lee, S (2009). Value of plant diversity for diet mixing and sequencing in herbivores. Rangelands, 31(1), 45–49.
Pulliam, H. R. (1975). Diet optimization with nutrient constraints. The American Naturalist, 109(970), 765–768.
Pyke, G. H. (1984). Optimal foraging theory: A critical review. Annual Review of Ecology and Systematics, 15(1), 523–575.
Pyke, G. H., Pulliam, H. R., & Charnov, E. L. (1977). Optimal foraging: A selective review of theory and tests. Quarterly Review of Biology, 52(2), 137–154.
R: A language and environment for statistical computing. R 2.11.1. (2010). Vienna, Austria: R Foundation for Statistical Computing. Retrieved from http://www.R-project.org
Raubenheimer, D., & Simpson, S. J. (2003). Nutrient balancing in grasshoppers: Behavioural and physiological correlates of dietary breadth. Journal of Experimental Biology, 206(10), 1669–1681.
Reid, N., & Lange, R. T. (1988). Host specificity, dispersion and persistence through drought of two arid zone mistletoes. Australian Journal of Botany, 36(3), 299–313.
Reid, N. (1997). Control of mistletoes by possums and fire: A review of the evidence. The Victorian Naturalist, 114(3), 149–158.
Reid, N., & Yan, Z. (2000). In P. J. Keane, G. A. Kile, F. D. Podger & B. N. Brown (Eds.), Diseases and Pathogens of Eucalypts (pp. 353–375). Collingwood: CSIRO Publishing.
Ritchie, M. E. (2009). Scale, heterogeneity, and the structure and diversity of ecological communities (Vol. 45). Princeton: Princeton University Press.
Robinson, B. W., Wilson, D. S. (1998) Optimal foraging, specialization, and a solution to Liem’s Paradox. The American Naturalist, 151(3), 223–235.
Rundle, A. S., & Rowe, R. K. (1974). A Study of the Land in the Catchment of the Broken River. Kew: Soil Conservation Authority of Victoria.
Sakaguchi, E., & Hume, I. D. (1990). Digesta retention and fibre digestion in brushtail possums, ringtail possums and rabbits. Comparative Biochemistry and Physiology Part A: Physiology, 96(2), 351–354.
Scrivener, N. J., Johnson, C. N., Wallis, I. R., Takasaki, M., Foley, W. J., & Krockenberger, A. K. (2004). Which trees do wild common brushtail possums (Trichosurus vulpecula) prefer? Problems and solutions in scaling laboratory findings to diet selection in the field. Evolutionary Ecology Research, 6(1), 77–87.
Searle, K. R., Hobbs, N. T., & Gordon, I. J. (2007). It's the" foodscape", not the landscape: Using foraging behavior to make functional assessments of landscape condition. Israel Journal of Ecology and Evolution, 53(3), 297–316.
Sedinger, J. S., & Raveling, D. G. (1984). Dietary selectivity in relation to availability and quality of food for goslings of cackling geese. The Auk, 101(2), 295–306.

85
Senft, R. L., Coughenour, M. B., Bailey, D. W., Rittenhouse, L. R., Sala, O. E., & Swift, D. M. (1987). Large herbivore foraging and ecological hierarchies. BioScience, 37(11), 789–799.
Sessions, L. A., & Kelly, D. (2002). Heterogeneity in vertebrate and invertebrate herbivory and its consequences for New Zealand mistletoes. Austral Ecology, 26(6), 571–581.
Shipley, L. A., Forbey, J. S., & Moore, B. D. (2009). Revisiting the dietary niche: When is a mammalian herbivore a specialist? Integrative and Comparative Biology, 49(3), 274–290.
Speiser, B., & Rowell-Rahier, M. (1991). Effects of food availability, nutritional value, and alkaloids on food choice in the generalist herbivore Arianta arbustorum (Gastropoda: Helicidae). Oikos, 62(3), 306–318.
Spotfire S+® Version 8.1 (2010). TIBCO Software Inc., Somerville. Sweetapple, P. J., & Nugent, G. (1998). Comparison of two techniques for assessing
possum (Trichosurus vulpecula) diet from stomach contents. New Zealand Journal of Ecology, 22(2), 181–188.
Sweetapple, P. J., Nugent, G., Whitford, J., & Knightbridge, P. I. (2002). Mistletoe (Tupeia antarctica) recovery and decline following possum control in a New Zealand forest. New Zealand Journal of Ecology, 26(1), 61–72.
Sweetapple, P. J., Fraser, K. W., & Knightbridge, P. I. (2004). Diet and impacts of brushtail possum populations across an invasion front in South Westland, New Zealand. New Zealand Journal of Ecology, 28(1), 19–33.
Sweetapple, P. J. (2008). Spatial variation in impacts of brushtail possums on two Loranthaceous mistletoe species. New Zealand Journal of Zoology, 32(2), 177–185.
Tuft, K. D., Crowther, M. S., Connell, K., Müller, S., & McArthur, C. (2011). Predation risk and competitive interactions affect foraging of an endangered refuge-dependent herbivore. Animal Conservation, 14(4), 447–457.
Tuft, K. D., Crowther, M. S., & McArthur, C. (2011). Multiple scales of diet selection by brush-tailed rock-wallabies (Petrogale penicillata). Australian Mammalogy, 33(2), 169–180.
Turner, M. G. (1989). Landscape ecology: The effect of pattern on process. Annual Review of Ecology and Systematics, 20, 171–197.
Unsicker, S. B., Oswald, A., Köhler, G., & Weisser, W. W. (2008). Complementary effects through dietary mixing enhance the performance of a generalist insect herbivore. Oecologia, 156(2), 313–324.
Van Der Ree, R. (2012). Ecology of arboreal marsupials in a network of remnant linear habitats. (Ph. D. thesis), Deakin University, Geelong.
Van Eeden, L., Di Stefano, J., & Coulson, G. (2011). Diet selection by the brush-tailed rock-wallaby (Petrogale penicillata) in East Gippsland, Victoria. Australian Mammalogy, 32(2), 162–168.
Van Tienderen, P. H. (1991). Evolution of generalists and specialist in spatially heterogeneous environments. Evolution, 45(6), 1317–1331.
Venables, W. N., & Ripley, B. D. (2002). Modern Applied Statistics with S (4 ed.). New York: Springer.
Wallis, I. R., Watson, M. L., & Foley, W. J. (2002). Secondary metabolites in Eucalyptus melliodora: Field distribution and laboratory feeding choices by a generalist herbivore, the common brushtail possum. Australian Journal of Zoology, 50(5), 507–519.
Watson, D. M. (2001). Mistletoe—A Keystone Resource in Forests and Woodlands Worldwide. Annual Review of Ecology and Systematics, 32(1), 219–249.
Watson, D. M. (2011). Mistletoes of southern Australia. Collingwood: CSIRO Publishing. Watts, D. P. (1984). Composition and variability of mountain gorilla diets in the central
Virungas. American Journal of Primatology, 7(4), 323–356.

86
Wayne, A. F., Cowling, A., Rooney, J. F., Ward, C. G., Wheeler, I. B., Lindenmayer, D. B., & Donnelly, C. F. (2006). Factors affecting the detection of possums by spotlighting in Western Australia. Wildlife Research, 32(8), 689–700.
Westoby, M. (1974). An analysis of diet selection by large generalist herbivores. The American Naturalist, 108(961), 290–304.
Westoby, M. (1978). What are the biological bases of varied diets? The American Naturalist, 112(985), 627–631.
Wiggins, N. L., McArthur, C., McLean, S., & Boyle, R. (2003). Effects of two plant secondary metabolites, cineole and gallic acid, on nightly feeding patterns of the common brushtail possum. Journal of Chemical Ecology, 29(6), 1447–1464.
Wiggins, N., McArthur, C., & Davies, N. W. (2006a). Diet switching in a generalist mammalian folivore: Fundamental to maximising intake. Oecologia, 147(4), 650–657.
Wiggins, N., McArthur, C., & Davies, N. W., & McLean, S. (2006b). Behavioral responses of a generalist mammalian folivore to the physiological constraints of a chemically defended diet. Journal of Chemical Ecology, 32(6), 1133–1147.
Wiggins, N. L., McArthur, C., Davies, N. W., & McLean, S. (2008). Spatial scale of the patchiness of plant poisons: A critical influence on foraging efficiency. Ecology, 87(9), 2236–2243.
Williams, C. K. (1982). Nutritional properties of some fruits eaten by the possum Trichosurus vulpecula in a New Zealand broadleaf-podocarp forest. New Zealand Journal of Ecology, 5, 16–20.
Woldemichael, G. M., & Wink, M. (2002). Concomitant occurrence of pyrrolizidine and quinolizidine alkaloids in the hemiparasite Osyris alba L. (Santalacea). Biochemical Systematics and Ecology, 30(2), 139–149.
Worton, B. J. (1989). Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology, 7(1), 164–168.
Young, H. E. (1937). The mistletoe problem. Brisbaine: Queensland Forestry Department. Zar, J. H. (2009). Biostatistical Analysis (5 ed.). New Jersey: Pearson.
Zweifel‐Schielly, B., Leuenberger, Y., Kreuzer, M., & Suter, W. (2011). A herbivore's food landscape: Seasonal dynamics and nutritional implications of diet selection by a red deer population in contrasting Alpine habitats. Journal of Zoology, 286(1), 68–80.

87
CH
AP
TE
R
3
Home range attributes of the common brushtail possum (Trichosurus vulpecula):
How does individual variability in resource use, food, and shelter influence spatial patterns?

88
CHAPTER 3
Home range attributes of the common brushtail possum (Trichosurus vulpecula):
How does individual variability in resource use, food, and shelter influence spatial patterns?
Introduction
Understanding the factors that shape home range size, resource use, and the spatial
organization of animals within a landscape is one of the fundamental challenges in ecology.
An animal’s home range is defined as the area traversed by an individual performing daily
life activities, such as foraging, resting, shelter-seeking, mating, and raising young (Burt,
1943). Long ago, Seton (1909, p. 23) observed, “No wild animal roams at random over the
country; each has a home region, even if it has not an actual home.” This area restricted use
of space has profound consequences for many ecological processes including animal
distribution and abundance (Gautestad & Mysterud, 2005), habitat use and resource
selection (Rhodes, McAlpine, Lunney, & Possingham, 2005), community structure (Fagan,
Lutscher, & Schneider, 2007), and predator-prey interactions (Lewis & Murray, 1993).
Home range is a spatial expression of individual behaviours that an animal performs to
survive, reproduce, and avoid predators and competitors; therefore, its movement
pathways are influenced by many different biotic and abiotic factors (summarized by
Börger et al., 2006; DeGabriel, Moore, Marsh, & Foley, 2010, Fig. 1). According to
Moorcroft and Lewis (2006) these macroscopic spatial patterns are determined by a large
number of single movement steps, each of which results from the interactions among
individual animal life traits, physiological state, and the external environment. Furthermore,
herbivores living in nutritionally poor environments modify their movement pathways in
response to the availability and nutritional quality of food and adjust accordingly their
foraging behaviour and physiological state (Dearing, Foley, & McLean, 2005).

89
Fig. 1 Graphical model of animal space-use processes (modified from Börger et al., 2006)

90
However, a home range size is not a universal measure of animal space use; it varies
between species, populations, and individuals and depends on the interactions between
individual energetic requirements and the spatial distribution of limiting resources across a
landscape. This individual variability is associated with differences in body size, sex and age,
reproductive status, habitat type, and availability of food and shelter to individuals
(Cederlund & Sand, 1994; Harestad & Bunnel, 1979; Milton & May, 1976; Perry & Garland
Jr, 2002). While shelter can be a limiting resource, the availability, distribution, and quality
of food usually strongly influence the home range size of individuals (DeGabriel, Moore,
Foley, & Johnson, 2009; Mares et al., 1982; Vedder, 2005). Individuals in food-rich
environments are expected to meet their energy requirements over a reduced area,
compared to animals living in areas with sparse or patchily distributed resources (McIntyre
& Wiens, 1999; Said et al., 2009). Social structure and mating patterns can also affect the
home range size of different members of a population with males usually having larger
home ranges than females to maximize mating opportunities and reproductive success
(Martin & Martin, 2007; Sandell, 1989).
Despite this recognition, variability in resource selection and space-use among individuals
within the same population has long been dismissed as ecological noise around the mean
(Ringler, 1983). As summarized by Bourke, Magnan, and Rodriguez (2005), although
individual variability in foraging patterns of fishes was well described by Ivlev (1961), its
ecological significance for the home ranging behaviour and social structure of different
animal species was only recognized much later (Lomnicki, 1988). Roughgarden (1972,
1979) identified two components of individual variability in resource use: the within-
individual component represents the variation in resource use by a single individual
through time, and the between-individual component represents resource diversity used by
different individuals in a population. Between-individual variability may develop from
physiological, behavioural, and morphological differences among animals while within-
individual variability may be caused by specific physiological states, such as hunger and
fear, and behavioural contexts, such as animal experience, food type or distribution, and
motivation. Individual variability in turn can create two types of spatial organisation of
animals: The first type affects an individual’s home range size, while the second type affects
the spacing of animals within a landscape.

91
Arboreal marsupials, with their versatility in diet, dependence on large hollows for shelter
and reproduction, and broad habitat tolerances, are an ideal group to clarify determinants
of individual variability in space use and resource selection. In particular, the study of
arboreal marsupial home ranging behaviour can answer questions regarding whether
individual variability in a home range size is the outcome of differences in energy needs
(i.e., females rearing young, males competing over females or dispersing subadult males),
and availability and nutritional quality of forage.
Previous nutritional studies have shown that variability in the chemical composition of
foliage of closely related Eucalyptus species and individuals of the same species can explain
some of the observed spatial patterns at scales ranging from a couple of meters to
thousands of kilometres (Andrew, Peakall, Wallis, & Foley, 2007; DeGabriel, Moore,
Shipley, et al., 2009; Lawler, Foley, & Eschler, 2000; Moore, Lawler, Wallis, Beale, & Foley,
2010). In addition, the patchy distribution of hollow-bearing trees is considered a limiting
resource in Australia’s fragmented and intensively managed forests and woodlands
affecting the spatial organization of entire populations of arboreal marsupials (Gibbons &
Lindenmayer, 2002).
The present study examines the effects of within- and between-individual variability in
resource use, food and shelter, on the home ranging behaviour of a model generalist
herbivore, the common brushtail possum (Trichosurus vulpecula: Marsupialia, Phalangeridae).
Following the findings presented in Chapter 2 that highlighted the importance of parasitic
plants and different species of trees in a possum diet, this chapter will explore their
combined effects on space use and resource selection by individual animals. This study also
provides baseline data on tree use and the foraging of possums on specific species of trees
and individual trees which will be used in Chapter 4 to measure the nutritional quality of
preferred and avoided trees. Studying individual responses to variation in food resources
and other potentially limiting factors will allow for a better understanding of the common
brushtail possum ecology, its behavioural adaptations to nutritionally poor environments,
spatial organization, and evolutionary success.

92
Aims and hypotheses
The main aim of this study was to determine how availability of resources (i.e., food and shelter) shapes
spatial patterns, foraging, denning behaviour, and tree use of the common brushtail possum (Trichosurus
vulpecula).
In particular, the following hypotheses were tested:
Hypothesis 1: Total home range size and core areas differ among common brushtail
possum individuals and sexes and are influenced by food and shelter availability.
Hypothesis 2: Common brushtail possums show preferences for specific trees and
differentiate trees used as denning sites from those used as a foraging sites.
Hypothesis 3: Trees parasitized by drooping mistletoe (Amyema pendula) are used more
often by common brushtail possums while foraging than would be expected from their
availability within home ranges.
Hypothesis 4: Trees with suitably sized hollows are used as denning sites by common
brushtail possums more often than would be expected from their availability within home
ranges.

93
Study sites
This study was carried out in the Strathbogie Ranges (36° 48´ S, 145° 45´ E) in north-
eastern Victoria, Australia (see Chapter 2 for details; Fig. 2). It compromised a network of
vegetation corridors along Bonnie Doon Rd., Boundary Hill Rd., Creek Junction Rd., and
Ankers Rd (Fig. 3). The surveyed roadsides supported a fine-scale mosaic of Herb-rich
Foothill Forest in drier areas and Swampy Riparian Woodland in seasonally or permanently
inundated areas and along watercourses. The dominant tree species in Herb-rich Foothill
Forest were Eucalyptus radiata (narrow-leaved peppermint), E. viminalis (manna gum), and a
root parasitic shrub/small tree, Exocarpos cupressiformis (cherry ballart). In Swampy Riparian
Woodland, the dominant tree species were Eucalyptus camphora (mountain swamp gum) and
Acacia dealbata (silver wattle). Two mistletoe species were found parasitizing Eucalyptus and
Acacia trees, the abundant Amyema pendula (drooping mistletoe) and the relatively scarce
Muellerina eucalyptoides (creeping mistletoe).
Fig. 2 Herb-rich Foothill Forest and Swampy Riparian Woodland within roadside vegetation corridors in the Strathbogie Ranges (photo by K. Petrović)

94
Fig. 3 Surveyed roadside vegetation corridors in the Strathbogie Ranges, north-eastern Victoria

95
Methods
Description and justification of methods used for establishing a home range size, foraging, and denning
patterns of common brushtail possums
To determine a home range size, foraging, and denning patterns of individual common
brushtail possums, very high frequency (VHF) radio-telemetry was used to track individual
animals from a distance. The presence of a common brushtail possum in a tree at night was
considered a useful proxy of its foraging activity. This method has been widely used in
studies of foraging behaviour of nocturnal arboreal herbivores (Jones & Krockenberger,
2007; Martin & Handasyde, 1999; Moore & Foley, 2005; Scrivener et al., 2004) which are
cryptic or easily disturbed and are, therefore, rarely observed foraging. However, this
method can produce biased results as animals use trees for different reasons. For instance,
animals can use trees as movement routes between foraging patches, as den sites and
escape from predators, or to facilitate social interactions. This method allowed for trees
used as den sites to be identified with full certainty because animals were found resting in a
single tree during the day.
Animal capture
Initial information on the occurrence of common brushtail possums in the study area was
obtained from the spotlighting survey (Chapter 2). From this information, a trapping
procedure was designed that reduced capture of non-target species and minimized stress to
animals. All trapping, handling, and radio-tracking of possums was conducted under the
auspices of the CSU Animal Care and Ethics Committee, approval No 08/090 and
Department of Sustainability and Environment Flora and Fauna permit, no 10004604.
Animal trapping was conducted over 60 nights spread over two trapping periods that
varied in length and location of the capture sites:
1) 15 November–15 December 2008
2) 3 January–2 February 2009
At each site 10–20 large wire traps (Professional Trapping Supplies 600 mm x 260 mm x
240 mm) were set one hour before dusk for three consecutive nights until suitable animals
had been captured (Fig. 4). Traps were baited with apple and peanut butter and placed on
the ground at the base of a tree with hollows or where animals were previously seen
foraging.

96
Traps were partly covered with a durable plastic cover and leaf litter to protect captured
animals from inclement weather. Traps were checked before dawn, and animals not needed
for the study were immediately released at the site of capture. Suitable animals were
transferred from a trap to a hessian bag and transported to the field station for further
processing.
Animal handling
All captured common brushtail possums were weighed to the nearest 0.01 kg and sedated
with an intramuscular injection of tiletamine/zolazepam (Zoletil©, Virbac Australia Pty.
Ltd, Peakhurst, NSW, Australia). A single dose of Zoletil© 10 mg/kg animal body weight
was administrated to facilitate handling and minimise distress for the animals (after Jackson
2003). Ear pinnae of all individuals were tattooed using a tattoo punch and given a unique
number for subsequent identification and future studies. Animal sex was recorded and
standard morphometric measurements were taken to the nearest 0.1 mm (i.e., head length,
head width, hind foot or pes length, and ear length). The pouch condition and reproductive
status of females were recorded; if a pouch young was present, its sex was checked and
head length measured (an index of age). Male scrotal width and testis size were measured
(i.e., length, width, and depth). In addition, a tooth wear class, an index of age based on
wear patterns of the first molar in the upper jaw (1–7; Winter, 1980), and a rump wear
class, an index of body condition (0–3), were established. Based on the criteria set up by
Martin and colleagues (2007) that included body weight, female pouch condition, male
testis size, and tooth wear class, each individual was classified as juvenile, sub-adult, or
adult.
Next, nine randomly selected individuals (3 males and 6 females) out of 16 trapped were
fitted with 30 g VHF radiocollars (Sirtrack Pty. Ltd., Havelock North, New Zealand); each
collar was marked with a reflective tape to facilitate identification at night (Fig. 4). Each
animal was given an apple and left to recover from sedation and handling in a large
wooden box with fresh air for at least 6 hours before being released. In all instances,
possums recovered well without showing any signs of distress and consumed the food that
was provided. Animals were released one hour after dusk at the point of capture or at the
base of their den tree if known. At the end of the study period all animals wearing radio-
collars were retrapped to retrieve the collars and were released at the point of capture. No
animal health or welfare issues arose during the capture, handling, and radio-tracking of
animals.

97
Radio-telemetry survey
To determine tree use (i.e., foraging and denning behaviour) and establish home range size,
animals were fitted with radio-collars operating at discrete frequencies (151–152 MHz);
their movements were radio-tracked using a R100 telemetry receiver (Communications
Specialist, Inc., Orange, CA, USA) and a hand-held three-element yagi antenna (A. F.
Antronics, Inc., Urbana IL, USA). Animals were radio-tracked for 30 days and 30 nights
over four months (November 2008 to February 2009) to collect 60 fixes for each animal.
The previous radio-telemetry studies have found that 60 fixes per animal is the
recommended minimum number of fixes to correctly establish the home range size of
sedentary animals (after Kenward, 2001; Millspaugh & Marzluff, 2001). To minimize the
confounding effects of mating and territorial behaviour and to avoid any inadvertent
impacts on animal welfare, animals were not radio-tracked during a breeding season in
March and April.

98
Fig. 4 Common brushtail possum captured in a wire trap, fitted with a VHF radio-collar and released to its den tree (photo by K. Petrović)

99
Radio-tracking was carried out during the spring and summer, which was considered the
best time to explore how animals responded to the chemical variability of trees containing
the highest concentrations of plant secondary metabolites (see Chapter 4). Moreover,
spreading radio-tracking over a longer period minimized the bias introduced by spatio-
temporal autocorrelation in the animal movement analysis (Hansteen, Andreassen, & Ims,
1997). The same principle was applied when radio-tracking animals over a single night.
Namely, the time at which tracking commenced and the order in which individuals were
tracked was altered each time. All individuals were located between 1 hour after dusk and
approximately 2 hours before dawn or after they emerged from their dens and started
foraging. Radio-tracking was continued during their activity through the night and before
they returned to their dens (Fig. 5). To eliminate the possibility of spatio-temporal
autocorrelation between night fixes, the location of each individual was recorded only once
per night. In all cases, animals were identified with the use of an 80-W spotlight with a red
filter and 8 x 30 binoculars before recording their exact location using a hand-held
positioning device, a Garmin GPS 12 Personal Navigator (Garmin International, Inc.,
Olathe, KS, USA).
Common brushtail possums rested in one location during the day; hence, only one fix per
day was required to determine the location of den trees (Fig. 5). Day radio-fixes or passive
fixes represented animal den sites while night radio-fixes or active fixes represented
foraging activity. Both day and night fixes were considered in the home range analysis as
den sites have been found to influence ranging behaviour of brushtail possums in roadside
vegetation corridors (Del Borrello, 2009; Martin, Handasyde, & Taylor, 2007). Finally,
to estimate a home range size, all day and night fixes were plotted on a digitised base map
with ArcGIS 10.0 Software (ESRI, Redlands, CA, USA) using the animal movement
program, an ESRI ArcView extension (Hooge, Eichenlaub, & Solomon, 1999).
Vegetation survey
Within the established home range boundaries of the nine studied common brushtail
possums, the following information was collected regarding used trees: exact location, tree
species, height and diameter at breast height (DBH), and the number of hollows and
mistletoes. A hollow was considered suitable for possums if its entrance was greater than
10 cm in diameter (Gibbons et al., 2002). Trees used as foraging and denning sites were
described, and each tree was marked with a numbered tag for subsequent identification and
collection of leaf samples for the chemical analyses as described in Chapter 4.

100
To determine the availability of different tree species and the availability of trees with
hollows and mistletoes within each possum’s home range, additional information about
tree species composition and structure was collected. Trees were surveyed systematically at
20 m intervals along transect that was established across the entire home range area on
both sides of the road (Fig. 6). The exact location and identity of all studied trees was
recorded with a hand-held positioning device (Garmin GPS 12 Personal Navigator,
Garmin International, Inc., Olathe, KS, USA). Tree diameter and height were measured
using a hand-held laser hypsometer (LaserAce®, MDL Laser Systems, Ltd., York, UK).
Fig. 5 Night and day radio-tracking of common brushtail possums within roadside vegetation corridors in the Strathbogie Ranges (photo by A. Eberhard)

101
Fig. 6 Schematic diagram depicting vegetation sampling design used within the estimated home range of each common brushtail possum

102
Data analysis
Home range attributes
A home range size was estimated for each of nine studied common brushtail possums
using two different methods: the minimum convex polygon (MCP; Mohr, 1947) and fixed
kernel (FK; Hooge et al., 1999; Worton, 1989). The minimum convex polygon method is
useful due to its simplicity and comparability between studies; it calculates the area
bounded by the outermost fixes. However, there are several problems with this method,
particularly the strong influence of outliers which can inflate estimates of home range size
(Harris et al., 1990; Powell, 2000). The fixed kernel method is, therefore, used in many
studies. This non-parametric statistical method calculates a kernel home range estimate
using Hooge’s algorithm (Hooge et al., 1999). It is considered less sensitive to outliers
because it identifies the areas of greatest use for each individual (Pope, Lindenmayer, &
Cunningham, 2004; Powell, 2000).
In this study, the fixed kernel method was used with a smoothing factor determined by the
least-square cross validation. This method calculated the core home range area (50%), the
area with the highest animal activity, and the total home range area (90%) using both day
(i.e., representing den trees) and night fixes (i.e., representing food trees). The same home
range areas were calculated by the minimum convex polygon method. Following the
findings of Martin and colleagues (2007), the entire width of the habitat, including the road,
was considered when calculating the total home range size as common brushtail possums
were observed crossing the road on numerous occasions. To identify whether the total
home range estimates were calculated correctly, minimum convex polygon areas were
plotted against the number of position fixes for each individual. Home range asymptotes
approached a plateau in all nine cases (Fig. 7).

103
Fig. 7 The minimum convex polygon area (MCP) accumulation curves for the nine radio-tracked common brushtail possums (3 males and 6 females)
Using two methods, the differences between sexes of the calculated total and core home-
range sizes were studied. First, the Shapiro-Wilk test was completed to test for the
normality of data. Next, the F-test was performed to test if variances of the two
populations were equal. If the obtained P-value was > 0.05, then the t-test was used to
compare two populations under the assumption of equal variances. However, if the P-value
of the F-test was < 0.05 then the non-parametric, the Wilcoxon signed ranks test, was used.
To determine the entire area of common brushtail possum arboreal activity, including all
trees that were used as den or foraging sites by individual animals, 100% minimum convex
polygon estimates, as well as foraging and denning areas, were calculated (after
Lindenmayer, Welsh, & Donnelly, 1997; Martin, 2006). In this study, repeated use of the
same den or food tree by common brushtail possums was treated as a single data point to
avoid biasing range estimates by multiple records. The observed differences in denning and
foraging ranges between sexes were compared using the t-test and the Wilcoxon signed
ranks test. Finally, the Pearson product-moment correlation was used to investigate
whether foraging and denning areas influenced the total home range size.

104
Determinants of tree use by common brushtail possums
To identify individual variability in tree use, each animal was treated as a separate case and
results were pooled only when it was possible to make general conclusions about the
studied animals. All calculations were performed in R version 2.11.1 (Copyright (C) 2010
The R Foundation for Statistical Computing) with a significance level α = 0.05 unless
otherwise indicated.
Foraging preferences of each common brushtail possum were determined using a
bootstrap or randomization procedure (after Jones, Maclagan and Krockenberger 2006).
First, the above ground biomass of each tree species in each home range was calculated
using a general allometric equation (after Keith et al., 2000, see Chapter 2 for details):
lny = -2.3267 + 2.4855 lnx, where y is the aboveground biomass (kg) and x is tree DBH (cm)
Next, 30 records (corresponding with the number of nightly tree sightings for each
common brushtail possum) were randomly selected (with replacement) from the total
number of trees sampled within each home range, and the tree species was recorded. This
procedure was repeated 10,000 times to produce an expected distribution of randomly
selected tree species. The number of sightings of common brushtail possums in a specific
tree species was compared with the randomly generated distribution, and the probability of
sighting was determined directly from the randomized distribution.
To establish whether trees with hollows of a suitable size for common brushtail possums
or trees parasitized by mistletoes were used more often than would be expected from their
availability within animal home ranges, contingency tables were produced. The significance
of the difference between the two proportions was assessed using the Pearson's chi-square
test with Yates' continuity correction to avoid overestimation of P values. When
proportions were close or equal to zero, the bootstrap procedure (100,000 repetitions) was
used instead. The Wilcoxon rank-sum test was used to identify possible differences in the
distance of single and multiple used trees from the closest den tree. The same analysis was
performed to ascertain differences in the distance between trees parasitized by drooping
mistletoe or free from mistletoe and the closest den tree.

105
To investigate the relationship between used and unused trees by common brushtail
possums and predictor variables (i.e., tree species, height and diameter, biomass, and
number of mistletoes and hollows) the binomial generalized linear mixed model (GLMM)
was fitted with an animal considered a random factor. The same model was used to
investigate the relationship between food or den trees and predictor variables. Firstly, any
highly intercorrelated explanatory variables (Spearman’s Rho > 0.7) were excluded from
the analysis to avoid multicollinearity in the modelling process (Tabachnick & Fidell, 1996).
Next, GLMM was fitted with Laplace approximation used to generate parameter estimates.
Finally, the step Akaike’s Information Criterion (hereafter AIC) function from the MASS
library was employed to identify the best model with a combination of variables explaining
the possum tree presence (Venables & Ripley, 2002). This function is a stepwise algorithm
that sequentially searches through all possible models for the one that minimizes the AIC.
Namely, it starts from the simplest model (the intercept) and adds one step at a time to a
new variable, considered the best by the AIC, to each of the consecutive models. On each
step, the AIC from the new model is compared to the previous model, and if the AIC from
the new model is smaller, the step is repeated. If the AIC from the new model is not
smaller, the algorithm stops and the best model is considered to be the one from the
previous step. If there are no more variables to be added to the new model, the algorithm
stops and the full model is considered the best. The significance of variables from the final
selected model is estimated using the z-test.
It is worth noting, that the AIC (Akaike’s Information Criterion) was used because the
information-theoretic approach is a recommended alternative to traditional hypothesis
testing since it is more tolerant to violations of parametric statistics that are commonly
encountered in ecological data. Moreover, the AIC method allows for the comparison of
multiple working hypotheses and does not rely solely on the use of ecologically arbitrary
P values for determining significance (Burnham & Anderson, 2002). Analysis was carried
out using the statistical program Spotfire S+® Version 8.1 (2010, TIBCO Software Inc.
Somerville, MA).

106
Results
Home range attributes
In this study, the nine common brushtail possums (3 males and 6 females, Table 1) were
radio-tracked over a four-month period (Table 2) to establish their total home range size
(90%), core area (50%), as well as their denning and foraging ranges (100%). The location
of home ranges of the nine radio-tracked individuals within a landscape of the Strathbogie
Ranges is shown in Fig. 8. The average total home range size (90%), calculated using the
minimum convex polygon method for both sexes combined, was 2.2 ± 0.6 ha (mean ± s.e.)
while the average core area (50%) was 0.4 ± 0.2 ha. The average total home range of males
was 4.1 ± 0.6 ha and of females 1.2 ± 0.4 ha. The total home range of males was
significantly larger than those of females (t = - 3.98, df = 7, P < 0.05; Table 3).
The average core area of males was more than twice the size of females, but the result was
not significant (0.8 ± 0.4 ha males; 0.3 ± 0.2 ha; W = 3, P = 0.167; Table 3). The total and
core home range areas of the nine common brushtail possums (3 males and 6 females)
calculated using the minimum convex polygon method are shown in Fig. 9.
Similarly, the average total home range (90%) of the nine common brushtail possums
calculated using the fixed kernel method was 2.4 ± 1.0 ha, and a core area 0.5 ± 0.2 ha,
with males having larger total and core home ranges than females (t = -2.25, df = 7, P =
0.06; t = - 2.52, df = 7, P < 0.05 respectively; Table 3). The average total home range of
males was 4.9 ± 2.2 ha and of females 1.2 ± 0.5 ha while the average core area of males
was 1.0 ± 0.4 ha and of females 0.3 ± 0.1 ha (Table 3). The total and core home range
areas of the nine common brushtail possums (3 males and 6 females), which were
calculated using the fixed kernel method, are shown in Fig. 10.
The total home range (100%) and foraging and denning ranges of the nine common
brushtail possums (3 males and 6 females) were established using the minimum convex
polygon method. The total home range and foraging ranges of the nine radio-tracked
common brushtail possums were strongly correlated (r = 0.91, P < 0.001) whereas the total
home range was less strongly correlated with denning ranges of all studied animals (r =
0.64, P = 0.07). The percent of overlap between the foraging area and total-home range
was 72.3% for males and 98% for females. The average foraging area of males was 4.8 ±
0.9 ha and of females’ 2.0 ± 0.8 ha while the average denning area of males was 1.5 ± 0.8
ha and of females’ 0.2 ± 0.1 ha (Table 4).

107
Males had larger foraging and denning ranges than females, but the result was not
significant (t = -2.18, df = 7, P = 0.07; W = 2, P = 0.10 respectively). Both sexes had
significantly larger foraging than denning home ranges (W = 0, P < 0.05). The total home
range (100%) and denning and foraging ranges of the nine common brushtail possums
(3 males and 6 females), which were calculated using the minimum convex polygon
method, are shown in Fig. 11.
Determinants of tree use by common brushtail possums
Approximately two-thirds of nightly sightings of the nine common brushtail possums
(3 males and 6 females) were in exotic species of trees (30.3%) and Eucalyptus radiata
(28.7%), followed by less frequently used E. camphora (13.4%) and E. viminalis (12.2%), and
occasionally used Exocarpos cupressiformis (3.9%) and Acacia dealbata (3.5%). In 5% of cases,
common brushtail possums were recorded foraging on the ground. Of the six tree species
most commonly used by common brushtail possums while foraging, Eucalyptus species
were used in accordance with their availability expressed as above ground biomass. Exotic
species of trees and a root parasite Exocarpos cupressiformis were used proportionally to their
availability and only Acacia dealbata was used significantly less often than expected (P <
0.001, Fig. 12).
Out of 520 recorded trees available to common brushtail possums within their home
ranges, 9.6% were parasitized by drooping mistletoe (Amyema pendula) with on average 3.7
± 0.7 mistletoe clumps per tree. Mistletoe infection rates were not uniform across tree
species. Most often mistletoes were found on E. radiata, representing almost half of all
parasitized trees (46%) with on average 5.0 ± 1.2 mistletoe clumps per tree, followed by E.
viminalis (24%) with 1.6 ± 0.2 mistletoe clumps per tree. By contrast, out of 210 trees used
by common brushtail possums while foraging, 20.5% were parasitized by drooping
mistletoe with on average 4.7 ± 0.9 mistletoe clumps per tree. Eucalyptus radiata represented
67.4% parasitized trees with on average of 5.6 ± 1.3 mistletoe clumps per tree. The chi-
square test showed that common brushtail possums used mistletoe parasitized trees more
often while foraging than expected by chance alone (χ2 = 14.911, df = 1, P < 0.001).

108
Table 1 Morphometric features and a life stage of the nine radio-tracked common brushtail possums (3 males and 6 females)
Animal Weight Head Ear Pes Tooth Rump Life ID
(kg)
Length (mm)
Width (mm)
Length (mm)
Length (mm)
Wear class
Wear class
Stage
Male 1 3.0 91.4 49.1 59.3 53.3 4 0 Adult
Male 2 3.3 94.6 51.1 57.5 43.4 5 2 Adult
Male 3 2.4 98.3 57.7 73.9 60.1 2 2 Subadult
Female 1 1.6 83.6 44.9 61.6 59.2 2 1 Subadult
Female 2 2.9 93.7 50.9 79.6 61.9 5 0 Lactating
Female 3 2.0 92.2 52.1 67.7 63.4 3 0 Adult
Female 4 3.0 100.2 60.4 63.4 57.8 4 0 Adult
Female 5 2.7 100.8 56.7 77.0 60.2 6 0 Lactating
Female 6 2.6 93.6 52.2 56.3 63.3 2 3 Adult

109
Table 2 Details of a radio-telemetry survey of the nine common brushtail possums (3 males and 6 females)
Animal ID
Tracking period
Days tracked
Total No. passive fixes (Den trees)
Total No. active fixes (Food trees)
Male 1
16.11.2008.–07.04.2009.
32
32 (15)
30 (27)
Male 2 25.11.2008.–12.04.2009. 30 30 (4) 29 (27)
Male 3 28.1120.08.–19.04.2009. 32 29 (12) 32 (28)
Female 1 25.11.2008.–18.04.2009. 30 30(13) 30 (25)
Female 2 12.01.2009.–19.04.2009. 30 30 (2) 30 (28)
Female 3 12.01.2009.–23.04.2009. 30 30 (3) 29 (25)
Female 4 11.01.2009.–21.04.2009. 30 30 (5) 30 (27)
Female 5 03.12.2008.–08.04.2009. 31 31 (3) 30 (25)
Female 6 14.12.2008.–18.04.2009. 30 30 (1) 30 (26)

110
Fig. 8 Location of home ranges of the nine common brushtail possums (3 males and 6 females) within a landscape of the Strathbogie Ranges, Victoria

111
Table 3 The total home range (90%) and core area (50%) of the nine common brushtail possums (3 males and 6 females) calculated using the minimum convex polygon (MCP) and fixed kernel (FK) method. A zero value represents repeated occupancy of the same tree by the animal
Table 4 The total home range (100%), foraging and denning ranges of the nine common brushtail possums (3 males and 6 females) calculated using the minimum convex polygon (MCP) method. A zero value represents repeated occupancy of the same tree by the animal
Animal ID MCP 90% (ha) MCP 50% (ha) FK 90% (ha) FK 50% (ha)
male01 3.07 1.29 5.05 1.26 male02 4.99 0.02 0.93 0.24 male03 4.39 1.02 8.71 1.56 female01 1.66 1.00 3.54 0.67 female02 0.30 0.09 0.70 0.19 female03 2.02 0.43 1.45 0.37 female04 2.68 0.00 1.13 0.33 female05 0.17 0.01 0.05 0.01 female06 0.23 0.00 0.09 0.03
Animal ID
Total home-range (ha)
Foraging range(ha)
Denning range (ha)
male01 6.36 3.47 1.55 male02 6.59 6.59 0.06 male03 7.14 4.46 2.87 female01 3.96 3.89 0.58 female02 0.34 0.30 0.01 female03 2.04 1.94 0.50 female04 4.81 4.79 0.01 female05 0.26 0.26 0.00 female06
0.61
0.61
0.00

112
Fig. 9 The total home range (90%) and core area (50%) of the nine common brushtail possums (3 males and 6 females) calculated using the minimum convex polygon method. For the size of individual home range refer to the scale

113
Fig. 10 The total home range (90%) and core area (50%) of the nine common brushtail possums (3 males and 6 females) calculated using the fixed kernel method. For the size of individual home range refer to the scale

114
Fig. 11 The total home range (100%) and foraging and denning ranges of the nine common brushtail possums (3 males and 6 females) calculated using the minimum convex polygon method. For the size of individual home range refer to the scale
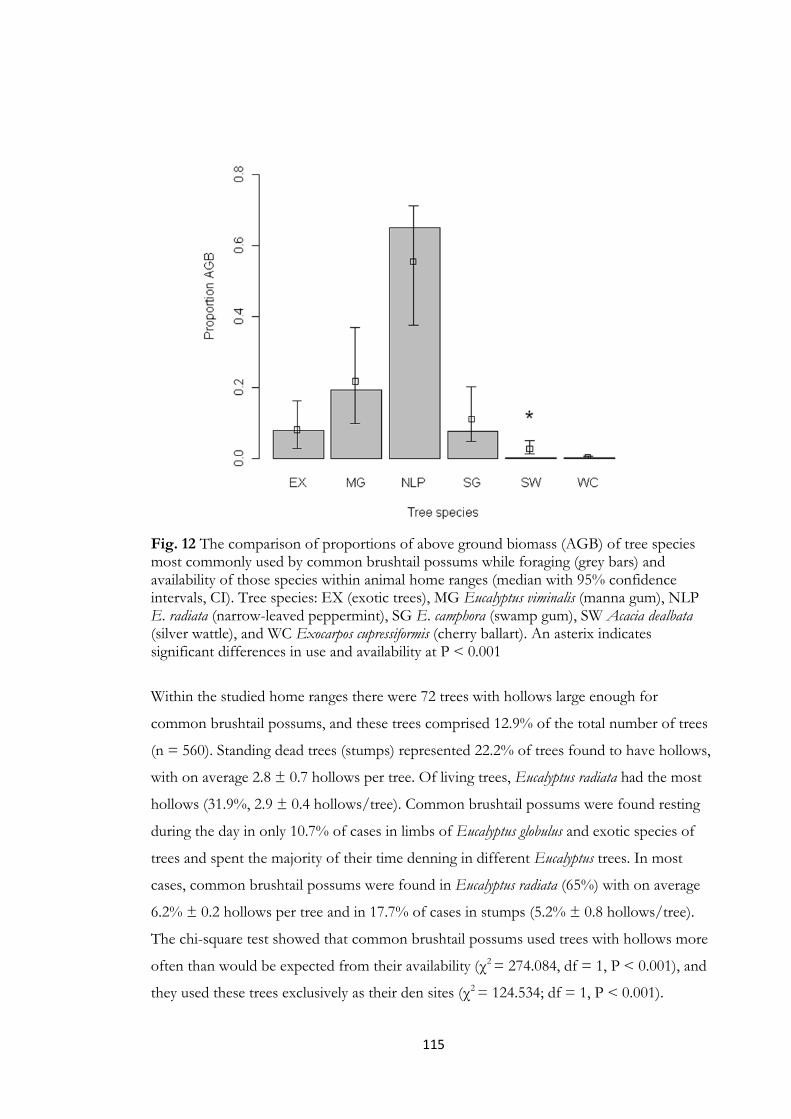
115
Fig. 12 The comparison of proportions of above ground biomass (AGB) of tree species most commonly used by common brushtail possums while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001
Within the studied home ranges there were 72 trees with hollows large enough for
common brushtail possums, and these trees comprised 12.9% of the total number of trees
(n = 560). Standing dead trees (stumps) represented 22.2% of trees found to have hollows,
with on average 2.8 ± 0.7 hollows per tree. Of living trees, Eucalyptus radiata had the most
hollows (31.9%, 2.9 ± 0.4 hollows/tree). Common brushtail possums were found resting
during the day in only 10.7% of cases in limbs of Eucalyptus globulus and exotic species of
trees and spent the majority of their time denning in different Eucalyptus trees. In most
cases, common brushtail possums were found in Eucalyptus radiata (65%) with on average
6.2% ± 0.2 hollows per tree and in 17.7% of cases in stumps (5.2% ± 0.8 hollows/tree).
The chi-square test showed that common brushtail possums used trees with hollows more
often than would be expected from their availability (χ2 = 274.084, df = 1, P < 0.001), and
they used these trees exclusively as their den sites (χ2 = 124.534; df = 1, P < 0.001).

116
Common brushtail possums used on average 6.7 ± 1.9 different trees as den sites, with
males having a significantly higher number of repeatedly used den trees than females (11.3
± 3.7; 4.3 ± 1.8 respectively, P < 0.001).
Moreover, common brushtail possums used medium-sized trees (50–100 cm in diameter
and 10–20 m high) and large-sized trees (100–150 cm and > 150 cm in diameter and > 20
m high) significantly more often than small-sized trees (5–50 cm in diameter, <10 m high;
χ2 = 352.668, df = 3, P < 0.001; χ2 = 207.956, df = 2, P < 0.001 respectively). The number
of mistletoes was weakly correlated with tree diameter and height (r = 0.18, r = 0.26
respectively, P < 0.001) while the number of hollows was strongly correlated with tree
diameter (r = 0.76, P < 0.001) and less strongly with tree height (r = 0.48, P < 0.001).
The binomial generalized linear mixed model showed that the key parameters that
separated trees used by common brushtail possums from unused ones were tree species,
diameter, and the number of hollows (r2 = 0.37, P < 0.001). Trees with a larger diameter
and a higher number of hollows were used more often by animals (Table 5).
The model explained 75.7% of common brushtail possum tree presences and 86.1% of
absences. Furthermore, tree species, diameter, number of hollows, and mistletoes were the
most important factors separating common brushtail possum food trees from den trees (r2
= 0.60, P < 0.001). Eucalyptus camphora and exotic trees were used as food trees while
stumps and E. radiata were used as den trees; E. viminalis was used both as a food and den
tree by common brushtail possums. Food trees had a smaller diameter, less hollows, and
more mistletoes than den trees (Table 6). The model explained 90.0% of common
brushtail possum foraging records and 88.2% of denning records.

117
Table 5 The binomial generalized linear mixed model predicting tree occupancy by common brushtail possums (AIC = 921.27, BIC = 990.55). Tree species: BLP Eucalyptus dives (broad-leaved peppermint), BW Acacia melanoxylon (blackwood), EX (exotic trees), MG E. viminalis (manna gum), MM E. obliqua (messmate), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), ST (stump), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significance at P < 0 ‘***’, P < 0.001 ‘**’, P < 0.01 ‘*’
Random effects:
Var. Std. Dev. Animal (Intercept) 0.257 0.507 Fixed effects:
Estimate Std. Error z-value P-value Intercept -0.964 0.449 -2.148 0.032* Species BLP -4.306 1.026 -4.196 2.72e-05*** Species BW -0.285 0.852 -0.335 0.738 Species EX -0.409 0.442 -0.927 0.354 Species MG -1.521 0.471 -3.226 0.001** Species MM -0.626 0.768 -0.816 0.414 Species NLP -1.504 0.411 -3.661 0.0003*** Species SG -1.545 0.477 -3.242 0.001** Species ST -1.855 0.526 -3.527 0.0004*** Species SW -2.170 0.528 -4.106 4.03e-05*** Species WC -0.805 0.582 -1.384 0.166346 Diameter 0.031 0.004 8.441 < 2e-16*** Hollow 0.354 0.065 5.437 5.41e-08*** Binomial generalized linear mixed model: Null fit: Visitation ~ 1 + Animal Fit: Visitation ~ Species + Diameter + Hollow + Animal df AIC BIC logLik Dev. Chi sq. Chi df P-value Null fit 2 1432.59 1442.48 -714.29 1428.59 Fit 14 921.27 990.55 -446.64 893.27 535.32 12 < 2.2e-16***

118
Table 6 The binomial generalized linear mixed model predicting tree use (food and den) by common brushtail possums (AIC = 295.88, BIC = 358.55). Tree species: BLP Eucalyptus dives (broad-leaved peppermint), BW Acacia melanoxylon (blackwood), EX (exotic trees), MG E. viminalis (manna gum), MM E. obliqua (messmate), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), ST (stump), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significance at P < 0 ‘***’, P < 0.001 ‘**’, P < 0.01 ‘*’
Random effects:
Var. Std. Dev. Animal (Intercept) 0.843 0.918 Fixed effects:
Estimate Std. Error z-value P-value Intercept 1.253e+00 7.208e-01 1.738 0.082 Species BLP 2.183e+01 5.619e+02 0.039 0.969 Species BW 1.809e+01 2.906e+03 0.006 0.995 Species Stump -1.677e+01 4.261e+03 -0.004 0.997 Species EX 2.040e+00 7.034e-01 2.900 0.004** Species MG 2.469e+00 7.250e-01 3.406 0.001*** Species MM 2.351e+01 6.050e+04 0.000 0.999 Species NLP 2.969e+00 6.019e-01 4.932 8.12e-07*** Species SG 4.672e+00 9.880e-01 4.729 2.25e-06*** Species SW 1.436e+01 7.583e+02 0.019 0.985 Species WC 1.850e+01 6.084e+03 0.003 0.998 Diameter -2.959e-02 6.886e-03 -4.297 1.73e-05*** Mistletoe 2.128e-01 9.910e-02 2.147 0.032* Hollow -6.276e-01 1.067e-01 -5.882 4.06e-09*** Binomial generalized linear mixed model: Null fit: Visitation ~ 1 + Animal Fit: Visitation ~ Species + Diameter + Mistletoe + Hollow + Animal
Df AIC BIC logLik Dev. Chi sq. Chi df P-value Null fit 2 664.18 672.54 330.09 660.18 Fit 15 295.88 358.55 132.94 265.88 394.31 13 < 2.2e-16***

119
Individual variability in tree use
Tree use varied among the studied common brushtail possum individuals (3 males and 6
females) with animals found foraging in 6.7% to 86.7% of cases in exotic species of trees.
In addition, tree use of Eucalyptus radiata was highly uneven ranging from 6.7% to 60.7%,
and a similar pattern was observed for E. viminalis and E. camphora trees (Table 7). When
the biomass of trees used as foraging sites by common brushtail possums was compared to
available tree biomass that was measured within each individual animal home range, strong
individual differences were observed among the studied animals. For instance, male
possum 1 and females 1 and 5 used E. radiata proportionally to its availability within the
home ranges while male 2 and female 3 used it less often than expected from its availability
(P < 0.05, Fig. 13). Male 2 and female 3 used E. viminalis more often than expected by
chance alone (P < 0.001) while female 5 avoided this species (P < 0.05). E. camphora was
used less often by females 1, 3, 4 and 6 (P < 0.05). Similarly, in all home ranges where
Acacia dealbata was found, common brushtail possums avoided using this species during
their nightly foraging activity (P < 0.05). However, when foraged trees where plotted
against the number of night radio fixes taken for each animal, the accumulation curves did
not reach a plateau in all cases (Fig. 14). This suggests that further radio-tracking could
produce different visitation rates to analysed trees and could change possum species
preferences.
Common brushtail possums were much less versatile in their use of den trees; most
animals (2 males and 4 females) were found significantly more often denning in Eucalyptus
radiata trees (P < 0.001), with only female 3 using E. camphora (P = 0.19) and female 6 E.
viminalis (P < 0.001) as their primary den sites. Here, accumulation curves plateaued for six
of the studied animals (1 male and 6 females) and increased for male 1 and 3 and female 1
(Fig. 15). Almost 90% of the nightly sightings of common brushtail possums were in trees
that were used only once during the sampling period (Fig. 16). There was a significant
difference in distances traversed by common brushtail possums from den trees to single
and multiple used trees (P < 0.001). Multiple used trees were on average 56.5 ± 17.5 m
from den trees while single used trees were on average 130.9 ± 8.8 m from den trees. No
significant difference was observed in distances travelled by common brushtail possums
between their den trees and trees parasitized by mistletoes (109.3 ± 21.5 m) or free from
mistletoes (123.0 ± 8.9 m, P = 0.788).

120
Table 7 Percentage of available and used trees by the nine common brushtail possums (3 males and 6 females) during their nightly foraging activity within each home range. Maximum and minimum visits (%) to each of the tree species by different individuals are marked in red
Animal ID Male 1 Male 2 Male 3 Female 1 Female 2 Female 3 Female 4 Female 5 Female 6
Species Avail Use Avail Use Avail Use Avail Use Avail Use Avail Use Avail Use Avail Use Avail Use Acacia dealbata 27.6 3.6 50.7 17.9 16.7 0.0 25.0 0.0 11.6 0.0 4.0 3.6 4.0 0.0 33.3 6.7 12.2 0.0
Acacia melanoxylon 0.0 3.6 0.0 0.0 0.0 0.0 5.0 10.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 4.1 0.0
Eucalyptus camphora 1.7 0.0 1.3 3.6 0.0 0.0 35.0 56.7 0.0 0.0 46.0 7.1 46.0 50.0 2.2 0.0 12.2 0.0
Eucalyptus globulus 10.3 3.6 0.0 0.0 4.8 4.0 0.0 0.0 1.4 0.0 0.0 0.0 0.0 0.0 6.7 23.3 0.0 0.0
Eucalyptus viminalis 0.0 0.0 5.3 0.0 6.0 8.0 0.0 3.3 20.3 6.7 6.0 10.7 6.0 0.0 17.8 20.0 32.7 65.4
Eucalyptus dives 3.4 0.0 0.0 0.0 26.2 8.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Eucalyptus radiata 48.3 60.7 29.3 35.7 28.6 48.0 35.0 30.0 23.2 6.7 24.0 32.1 24.0 21.4 13.3 26.7 2.0 0.0
Eucalyptus obliqua 0.0 3.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.3 13.3 0.0 0.0
Exocarpos cupressiformis 0.0 0.0 0.0 0.0 7.1 4.0 0.0 0.0 1.4 0.0 4.0 0.0 4.0 3.6 8.9 3.3 36.7 26.9
Exotic trees 8.6 25.0 13.3 42.9 10.7 28.0 0.0 0.0 42.0 86.7 16.0 46.4 16.0 25.0 4.4 6.7 0.0 7.7

121
Fig. 13 The comparison of proportions of above ground biomass (AGB) of tree species used by individual common brushtail possums (3 males and 6 females) while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001

122
Fig. 13 cont. The comparison of proportions of above ground biomass (AGB) of tree species used by individual common brushtail possums (3 males and 6 females) while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001

123
Fig. 13 cont. The comparison of proportions of above ground biomass (AGB) of tree species used by individual common brushtail possums (3 males and 6 females) while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001
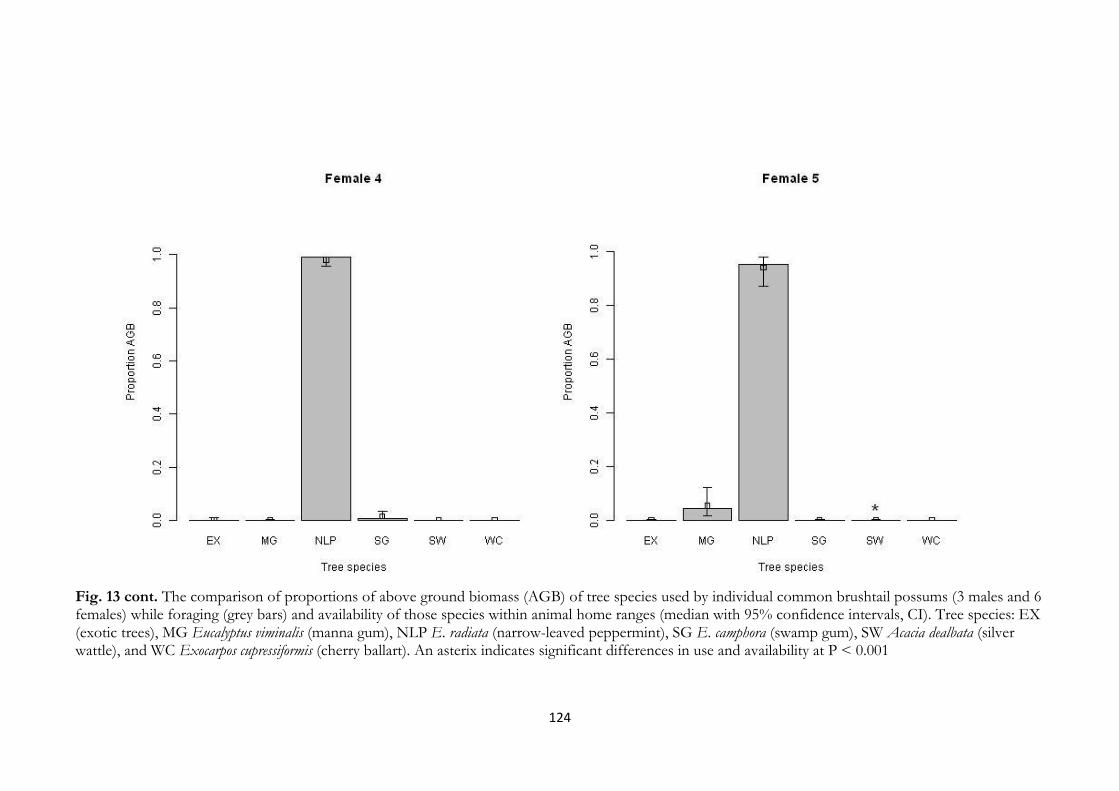
124
Fig. 13 cont. The comparison of proportions of above ground biomass (AGB) of tree species used by individual common brushtail possums (3 males and 6 females) while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001
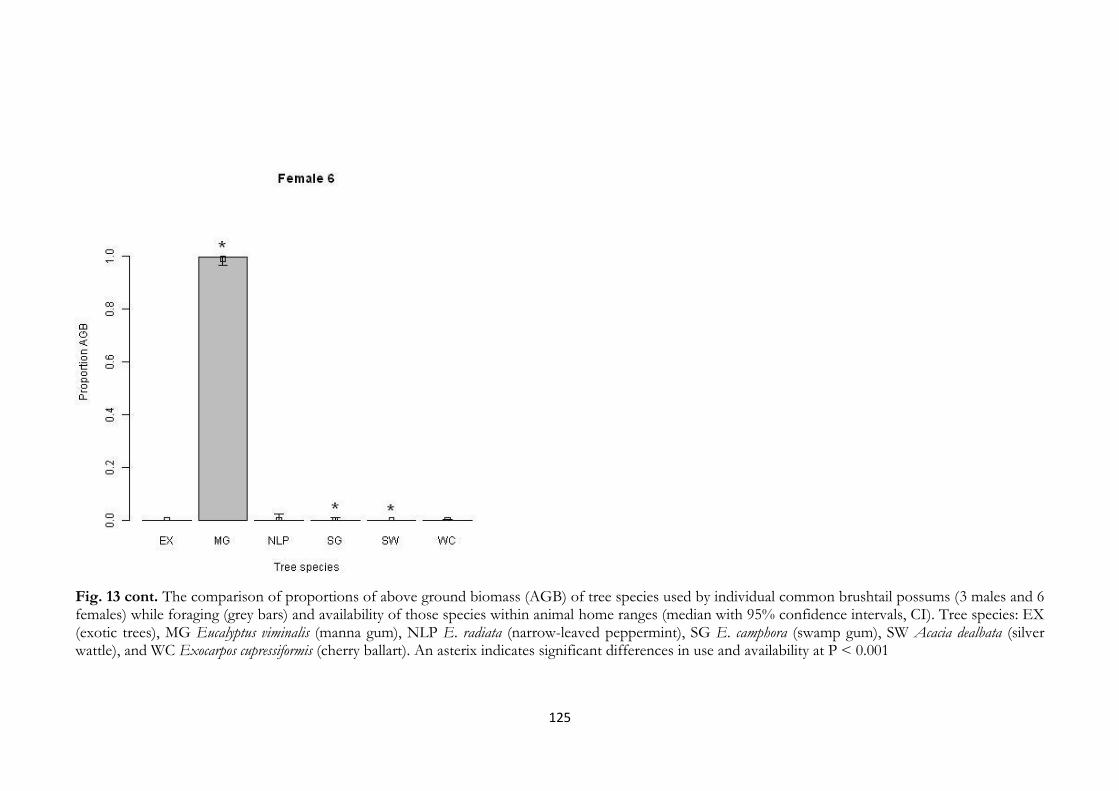
125
Fig. 13 cont. The comparison of proportions of above ground biomass (AGB) of tree species used by individual common brushtail possums (3 males and 6 females) while foraging (grey bars) and availability of those species within animal home ranges (median with 95% confidence intervals, CI). Tree species: EX (exotic trees), MG Eucalyptus viminalis (manna gum), NLP E. radiata (narrow-leaved peppermint), SG E. camphora (swamp gum), SW Acacia dealbata (silver wattle), and WC Exocarpos cupressiformis (cherry ballart). An asterix indicates significant differences in use and availability at P < 0.001

126
Fig. 14 Accumulation curves of the number of food trees recorded during night radio-tracking of the nine common brushtail possums (3 males and 6 females)
Fig. 15 Accumulation curves of the number of den trees recorded during day radio-tracking of the nine common brushtail possums (3 males and 6 females)

127
Fig. 16 Percentage (%) of trees used by common brushtail possums once or more during their nightly foraging activity

128
Discussion
Summary of key results
In the present study, male common brushtail possums had larger home ranges than
females. The total home range size of both males and females was strongly correlated with
their foraging ranges and only weakly correlated with their denning ranges, suggesting that
food availability was a more limiting factor for common brushtail possums than the
availability of hollow-bearing trees. Moreover, the studied common brushtail possums
preferred different species of trees when foraging and repeatedly used individuals of the
same species, as well as trees parasitized by mistletoes. Possums differentiated between
trees used as foraging and denning sites based on species, diameter, and number of hollows
and mistletoes, with trees having a higher number of hollows used more often as denning
sites.
Home range attributes
In general, a home range size of arboreal marsupials inhabiting eucalypt forests and
woodlands is considered to be the outcome of animal foraging, denning, and mating
behaviours. In this study, areas traversed by common brushtail possums during their
nightly foraging activity, along with areas occupied during the day by animals resting in a
single hollow-bearing tree, produced different estimates of the total home range size. Male
common brushtail possums had significantly larger home ranges (4.1 ± 0.6 ha) than
females (1.2 ± 0.4 ha). This finding is consistent with other studies looking at differences in
home range size between sexes, with male common brushtail possums having home ranges
from 1 to 11 ha and females from 0.7 to 7.4 ha in size (summarized by Kerle, 2001).
Sex differences in the home range size of males and females (5.1 ± 0.8 ha; 2.1 ± 0.3 ha
respectively) have been found previously for a closely related species, the mountain
brushtail possum (Trichosurus cunninghami), occurring in the same study area (Martin et al.,
2007). The authors suggested that males of this polygynous species had to range over larger
areas than females to access sufficient food resources and maximize reproductive success
by overlapping their home ranges with home ranges of multiple females.

129
Similarly, common brushtail possums have a predominantly polygynous mating system
(Isaac & Johnson, 2003), and males often have overlapping home ranges and compete for
access to multiple estrous females (Winter, 1976). Hence, larger male home ranges could be
attributed to different reproductive imperatives of males and females.
Moreover, the sexual dimorphism observed in the studied common brushtail possums,
with males being larger than females, could have affected the differences in home range
size between the sexes. McNab (1963) suggested that the home range size of herbivores is
related to animal body weight and energetic requirements. Namely, herbivores’ energetic
requirements increase with body weight forcing animals to forage over larger areas,
especially in areas with patchily distributed resources. Yet, in habitats of greater
environmental productivity and with aggregated resources, herbivores are expected to
range over smaller areas according to the habitat productivity hypothesis (Harestad &
Bunnel, 1979). This could partially explain the relatively small home ranges of both male
and female common brushtail possums occurring in the study area. The roadside
vegetation corridors in the Strathbogie Ranges are considered highly productive habitats
since they have not been logged for the last 100 years or burnt for the last 50 years (Martin
& Martin, 2007). As a result, these habitats are characterised by a high density of large trees
used as foraging and denning sites by arboreal marsupials.
Furthermore, other studies carried out in New Zealand have demonstrated that patches of
high quality habitat can maintain higher densities of common brushtail possums, and in
consequence, individuals can have relatively small home ranges (summarized by Cowan,
2001). For instance, where common brushtail possums live on farmland with scattered
patches of remnant forest or scrub, they show two types of ranging behaviour (Cowan &
Clout, 2000): Some have small home ranges centred on preferred habitats, such as stream-
side willows or swamps and never venture far out onto farmland; others range up to 1600
m over open pasture and have annual home ranges of up to 60 ha (Brockie et al., 1997).
Hence, the mobility of a species can affect the individual home range size, especially in the
habitats where resources are rare and patchily distributed. In these habitats, more mobile
animals have larger home ranges in order to include a sufficient number of suitable food
trees (DeGabriel et al., 2010).

130
Finally, abundance and quality of food have been previously recognized as the major
factors shaping home range size of herbivores (Säid et al., 2009). DeGabriel and colleagues
(2009a) found that the size of common brushtail possum home ranges was affected by the
nutritional quality of trees, with larger home ranges occurring in areas of lower nutritional
quality. The significantly larger foraging ranges than denning home ranges observed in the
studied common brushtail possums could have been the outcome of their generalist
feeding strategy that forced animals to range over larger areas in search of different types of
food. In the current study, the total home range size of both males and females was
strongly correlated with foraging area and only weakly correlated with denning area,
suggesting that food availability might have been a more limiting factor for common
brushtail possums than availability of hollow-bearing trees.
In the current study, male common brushtail possums had a significantly higher number of
dens than females (11.3 ± 3.7, 4.3 ± 1.8 respectively). By contrast, How (1981) found that
in north-eastern New South Wales male common brushtail possums used on average 1.5
dens and females 3.5 dens. In Queensland Winter (1976) found that only one to three dens
were used on a regular basis with females often using the same den for months. The small
number of dens recorded in earlier studies could indicate that the number of trees with
suitable size hollows for possums is limited in many parts of Australia subjected to heavy
deforestation and fragmentation. However, in the Strathbogie Ranges the roadside
vegetation corridors have been identified as sustaining a large number of trees bearing
hollows allowing mountain brushtail possums to occupy multiple den trees (males
16.5 ± 1.5 and females 7.4 ± 0.6; Martin et al., 2007).
In summary, resources available to animals within their home ranges do not affect animals
separately. Recently Di Stefano and colleagues (2011) proposed a conceptual framework
for quantifying the influence of multiple resources on the home range size of herbivores.
According to the authors, separate resources (i.e., food and shelter) interact to form a
continuous, multi-dimensional surface exhibiting an emergent environmental pattern with a
home range size more closely correlated with resource heterogeneity than with simple
additive or interactive effects of separate resources. They demonstrated that the availability
of different types of food (i.e., forbs, shrubs, ferns, monocots, and trees) and shelter
(i.e., lateral cover) affected the home range size of a terrestrial herbivore, the swamp
wallaby (Wallabia bicolor).

131
Individual variability in tree use
It has been recognized previously that individuals of the same species can exploit resources
differently to fulfil their requirements for growth, survival, and reproduction, and maximise
in this way their fitness contribution to future generations (Dall, McNamara, & Houston,
2004). For herbivores, several regulating mechanisms have been identified, including
variation in structure, abundance, and spatial distribution of forage within their home
ranges (Spalinger & Hobbs, 1992; Hobbs et al., 2003). In the current study, individual
possums used different tree species during their nightly foraging activity, and individual
home ranges differed in terms of tree species composition and abundance. For instance,
some home ranges spanned across both Herb-rich Foothill Forest and Swampy Riparian
Woodland (Female 1, 3 and 4) while others were dominated by a single species, Eucalyptus
radiata (Male 1 and 3). Interestingly, one of the older females carrying a second young in
season (Female 2) had a home range dominated by exotic species of trees. Hence, selection
against particular tree species by one common brushtail possum could have been due to its
high abundance within animal home range while selection for the same species by a
different possum could have been due to its rarity within its home range.
Alternatively, resource partitioning among the nine studied common brushtail possums
could have occurred with individuals using different species of trees as their foraging sites
within otherwise relatively uniform habitats. Individual differences in dietary preferences
have been identified as “individual specialization” or the expression of animal personalities
in a situation where generalist populations are composed of specialized foragers with
dietary preferences representing subsets of the broader species-specific generalist diet
(Bolnick et al., 2003). For example, the gracile mouse opossum (Gracilinanus microtarsus), an
omnivorous arboreal marsupial that inhabits the Atlantic rainforest and cerrado in Brazil,
has been found to exhibit significant individual specialization with diets of specialists being
nested within the diets of generalists (Araújo et al., 2009). Alternatively, the diet differences
may be exaggerated by direct interference between individuals, with dominant individuals
securing preferred foods by forcing subordinate individuals to feed on suboptimal
resources.
Apart from observed differences in tree species preferences, the studied common brushtail
possums repeatedly used individual trees of the same species that were usually closer to
their den trees.

132
Similar patterns were observed for opossum (Didelphis virginiana), with its den sites located
within close proximity to food (Allen, Marchinton, & Lentz, 1985). Opposite to
expectation, common brushtail possums were more likely to visit trees closer to dens
irrespective of mistletoe presence (i.e., they were not prepared to travel further to visit trees
with mistletoe although they preferred them). The observed inconsistencies in the findings
of this study could have been caused by a low sample size and if further radio-tracking was
possible it would allow for the separation of different factors.
Likewise, common brushtail possums do not always choose trees based solely on their
availability and proximity to shelter; other factors, such as competition with co-occurring
arboreal marsupials, can shape their tree choices. In the current study, possums showed a
negative selection for Acacia dealbata which is the preferred food of mountain brushtail
possums known to exhibit a certain level of territoriality in protecting their key resources
from competitors (Martin & Martin, 2007). Del Borrello (2009) looked at differences in
tree use by common and mountain brushtail possums in the same study area and suggested
a certain level of dietary partitioning between the sympatric species. Furthermore, two
species of possums are known to have different tolerances to plant secondary metabolites,
such as tannins and terpenes, with the latter being found only in Eucalyptus species eaten by
common brushtail possums (Burchfield, Agar, & Hume, 2006). Similarly, in tropical
rainforests in north-east Queensland, sympatric folivores (i.e., Lumholtz’s tree kangaroo,
Dendrolagus lumholtzi; lemuroid ringtail possum, Hemibelideus lemuroides; Herbert River ringtail
possum, Pseudochirulus herbertensis; and coppery brushtail possum, Trichosurus johnstonii)
partitioned resources based on their tolerance of different plant secondary metabolites
(Kanowski, Irvine, & Winter, 2003).
The current study has demonstrated that the distances travelled between foraging and
denning sites and the intensity of tree use by common brushtail possums shape their home
range size. Animals that exhibit the observed behaviour are altering the rate and bias of
their movement in response to heterogeneous features of the landscape (i.e. the availability
of different tree species, mistletoes and hollows, and the chemical composition of forage).
Moreover, intra-specific differences were observed with individual animals differing in their
responsiveness to environmental variability across space and time. Further investigation of
the mechanisms underlying individual variability in foraging and denning behaviour and
space use patterns can yield interesting insights into individual animal personalities, fitness
consequences, and the evolutionary success of this generalist herbivore.

133
References
Allen, C. H., Marchinton, R. L., & Lentz, W. M. (1985). Movement, habitat use and denning of opossums in the Georgia Piedmont, 113(2). American Midland Naturalist, 408–412.
Andrew, R. L., Peakall, R., Wallis, I. R., & Foley, W. J. (2007). Spatial distribution of defense chemicals and markers and the maintenance of chemical variation. Ecology, 88(3), 716–728.
Araújo, M. S., Martins, E. G., Cruz, L. D., Fernandes, F. R., Linhares, A. X., Dos Reis, S.
F., & Guimarães, P. R. (2009). Nested diets: A novel pattern of individual‐level resource use. Oikos, 119(1), 81–88.
Bolnick, D. I., Svanbäck, R., Fordyce, J. A., Yang, L. H., Davis, J. M., Hulsey, C. D., & Forister, M. L. (2003). The ecology of individuals: Incidence and implications of individual specialization. The American Naturalist, 161(1), 1–28.
Börger, L., Franconi, N., Ferretti, F., Meschi, F., De Michele, G., Gantz, A., & Coulson, T. (2006). An Integrated Approach to Identify Spatiotemporal and Individual-Level Determinants of Animal Home Range Size. The American Naturalist, 168(4), 471–485.
Bourke, P., Magnan, P., & Rodriguez, M. (2005). Individual variations in habitat use and morphology in brook charr. Journal of Fish Biology, 51(4), 783–794.
Brockie, R. E., Ward, G. D., & Cowan, P. E. (1997). Possums (Trichosurus vulpecula) on Hawke's Bay farmland: Spatial distribution and population structure before and after a control operation. Journal of the Royal Society of New Zealand, 27(2), 181–191.
Burchfield, E., Agar, N., & Hume, I. (2006). Effects of terpenes and tannins on some physiological and biochemical parameters in two species of phalangerid possums (Marsupialia: Phalangeridae). Australian Journal of Zoology, 53(6), 395–402.
Burdette, W. J., & Gehan, E. A. (1970). Planning and analysis of clinical studies. Springfield: Thomas.
Burt, W. H. (1943). Territoriality and home range concepts as applied to mammals. Journal of Mammalogy, 24(3), 346–352.
Cederlund, G., & Sand, H. (1994). Home-range size in relation to age and sex in moose. Journal of Mammalogy, 75(4), 1005–1012.
Chilcott, M., & Hume, I. (1985). Coprophagy and selective retention of fluid digesta: Their role in the nutrition of the common ringtail possum, Pseudocheirus peregrinus. Australian Journal of Zoology, 33(1), 1–15.
Comport, S., Ward, S., & Foley, W. (1996). Home ranges, time budgets and food-tree use in a high-density tropical population of greater gliders, Petauroides volans minor (Pseudocheiridae : Marsupialia). Wildlife Research, 23(4), 401–419.
Cowan, P. E. (2000). Factors Affecting Possum Re-infestation—Implications for Management: Department of Conservation Wellington.
Cowan, P. E. (2001). Advances in New Zealand mammalogy 1990–2000: Brushtail possum. Journal of the Royal Society of New Zealand, 31(1), 15–29.
Cowan, P. E., & Clout, M. (2000). Possums on the move: Activity patterns, home ranges, and dispersal. In T. Montague, T. (Ed.) The brushtail possum: Biology, impact and management of an introduced marsupial (pp. 24–34). Lincoln: Manaaki Whenua Press.
Dall, S. R., Houston, A. I., & McNamara, J. M. (2004). The behavioural ecology of personality: Consistent individual differences from an adaptive perspective. Ecology Letters, 7(8), 734–739.
Dearing, M. D., Foley, W. J., & McLean, S. (2005). The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates. Annual Review of Ecology, Evolution, and Systematics, 36, 169–189.

134
DeGabriel, J. L. (2008). Demographic constraints imposed by plant nutrients and secondary chemistry on the common brushtail possum (Trichosurus vulpecula Kerr.). (Ph. D. Thesis), the Australian National University, Canberra.
DeGabriel, J. L., Moore, B. D., Foley, W. J., & Johnson, C. N. (2009a). The effects of plant defensive chemistry on nutrient availability predict reproductive success in a mammal. Ecology, 90(3), 711–719.
DeGabriel, J., Moore, B. D., Shipley, L., Krockenberger, A. K., Wallis, I. R., Johnson, C. N., & Foley, W. J. (2009b). Inter-population differences in the tolerance of a marsupial folivore to plant secondary metabolites. Oecologia, 161(3), 539–548.
DeGabriel, J., Moore, B. D., Marsh, K. J., & Foley, W. J. (2010). The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology, 20(2), 97–108.
Del Borrello, C. (2009). Home-range and habitat use in sympatric brushtail possums (Trichosurus spp.). (Honours thesis), the University of Melbourne, Melbourne.
Di Stefano, J., Coulson, G., Greenfield, A., & Swan, M. (2011). Resource heterogeneity influences home range area in the swamp wallaby Wallabia bicolor. Ecography, 34(3), 469–479.
Dussault, C., Ouellet, J.-P., Courtois, R., Huot, J., Breton, L., & Jolicoeur, H. (2005). Linking moose habitat selection to limiting factors. Ecography, 28(5), 619–628.
Efford, M., Warburton, B., & Spencer, N. (2000). Home-range changes by brushtail possums in response to control. Wildlife Research, 27(2), 117–127.
Ellis, W., Melzer, A., Carrick, F., & Hasegawa, M. (2002). Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland. Wildlife Research, 29(3), 303–311.
Enoksson, B., & Nilsson, S. G. (1983). Territory size and population density in relation to food supply in the nuthatch Sitta europaea (Aves). The Journal of Animal Ecology, 52(3), 927–935.
ESRI, ArcGIS Desktop 10.0. (2012). Redlands, CA: Environmental Systems Resource Institute.
Fagan, W. F., Lutscher, F., & Schneider, K. (2007). Population and Community Consequences of Spatial Subsidies Derived from Central Place Foraging. The American Naturalist, 170(6), 902–915.
Fisher, D. O., & Lara, M. C. (1999). Effects of body size and home range on access to mates and paternity in male bridled nailtail wallabies. Animal Behaviour, 58(1), 121–130.
Fisher, D. O., & Owens, I. P. (2000). Female home range size and the evolution of social organization in macropod marsupials. Journal of Animal Ecology, 69(6), 1083–1098.
Ganzhorn, J. U. (1988). Food partitioning among Malagasy primates. Oecologia, 75(3), 436–450.
Gautestad, A. O., & Mysterud, I. (2005). Intrinsic scaling complexity in animal dispersion and abundance. The American Naturalist, 165(1), 44–55.
Gibbons, P., & Lindenmayer, D. (2002). Tree hollows and wildlife conservation in Australia. Collingwood: CSIRO.
Hansteen, T. L., Andreassen, H. P., & Ims, R. A. (1997). Effects of spatiotemporal scale on autocorrelation and home range estimators. The Journal of Wildlife Management, 61(2), 280–290.
Harestad, A. S., & Bunnel, F. L. (1979). Home range and body weight—a reevaluation. Ecology, 60(2), 389–402.
Harris, S., Cresswell, W. J., Forde, P. G., Trewhella, W. J., Woollard, T., & Wray, S. (1990). Home-range analysis using radio-tracking data—a review of problems and techniques particularly as applied to the study of mammals. Mammal Review, 20(2–3), 97–123.

135
Harvey, P. H., & Clutton-Brock, T. H. (1981). Primate home-range size and metabolic needs. Behavioral Ecology and Sociobiology, 8(2), 151–155.
Hews, D. K. (1993). Food resources affect female distribution and male mating opportunities in the iguanian lizard Uta palmeri. Animal Behaviour, 46(2), 279–291.
Hobbs, T. N., Gross, J. E., Shipley, L. A., Spalinger, D. E., & Wunder, B. A. (2003). Herbivore functional response in heterogeneous environments: A contest among models. Ecology, 84(3), 666–681.
Hooge, P. N., Eichenlaub, W., & Solomon, E. (1999). The animal movement program. USGS, Alaska Biological Science Center.
How, R. A. (1972). The ecology and management of Trichosurus species (Marsupialia) in N.S.W. (Ph.D. thesis), the University of New England, Armidale.
How, R. A. (1981). Population Parameters of Two Congeneric Possums, Trichosurus spp., in North Eastern New South Wales. Australian Journal of Zoology, 29(2), 205–215.
Isaac, J. L., & Johnson, C. N. (2003). Sexual dimorphism and synchrony of breeding: Variation in polygyny potential among populations in the common brushtail possum, Trichosurus vulpecula. Behavioral Ecology, 14(6), 818–822.
Ivlev, V. S. (1961). Experimental ecology of the feeding of fishes. New Haven: Yale University Press.
Johnson, D. H. (1980). The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61(1), 65–71.
Johnson, C. N., Clinchy, M., Taylor, A. C., Krebs, C. J., Jarman, P. J., Payne, A., & Ritchie, E. G. (2001). Adjustment of offspring sex ratios in relation to the availability of resources for philopatric offspring in the common brushtail possum. Proceedings of the Royal Society of London. Series B: Biological Sciences, 268(1480), 2001–2005.
Jones, K. M. W., & Krockenberger, A. K. (2007). Determining the diet of cryptic folivores: An assessment of diet analysis techniques using the green ringtail possum (Pseudochirops archeri) as a case study. Wildlife Research, 34(5), 352–358.
Kanowski, J., Irvine, A. K., & Winter, J. W. (2003). The relationship between the floristic composition of rain forests and the abundance of folivorous marsupials in north-east Queensland. Journal of Animal Ecology, 72(4), 627–632.
Kenward, R. (2001). A manual for wildlife radio tagging. London: Academic Press. Kerle, A. (2001). Possums: The brushtails, ringtails and greater glider. Sydney: UNSW Press. Lawler, I. R., Foley, W. J., & Eschler, B. M. (2000). Foliar concentration of a single toxin
creates habitat patchiness for a marsupial folivore. Ecology, 81(5), 1327–1338. Lewis, M. A., & Murray, J. D. (1993). Modelling territoriality and wolf-deer interactions.
Nature, 366(6457), 738–740. Lindenmayer, D. B. (1997). Differences in the biology and ecology of arboreal marsupials
in forests of southeastern Australia. Journal of Mammalogy, 1117–1127. Lindenmayer, D. B., Cunningham, R. B., Tanton, M. T., Smith, A. P., & Nix, H. A. (1991).
Characteristics of hollow-bearing trees occupied by arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, south-east Australia. Forest Ecology and Management, 40(3–4), 289–308.
Lomnicki, A. (1988). Population Ecology of Individuals. Princeton: Princeton University Press. Main, M. B. (2008). Reconciling competing ecological explanations for sexual segregation
in ungulates. Ecology, 89(3), 693–704. Mares, M. A., Lacher Jr, T. E., Willig, M. R., Bitar, N. A., Adams, R., Klinger, A., & Tazik,
D. (1982). An experimental analysis of social spacing in Tamias striatus. Ecology, 63(2), 267–273.
Martin, J. K. (2005). Behavioural ecology of bobuck (Trichosurus cunninghami). (Ph. D. thesis), the University of Melbourne, Melbourne.
Martin, J. K., Handasyde, K. A., & Taylor, A. C. (2007). Linear roadside remnants: Their
influence on den‐use, home range and mating system in bobucks (Trichosurus cunninghami). Austral Ecology, 32(6), 686–696.

136
Martin, J. K., Handasyde, K. A., Taylor, A. C., & Coulson, G. (2007). Long-term pair-bonds without mating fidelity in a mammal. Behaviour, 144(11), 1419–1445.
Martin, J. K., & Martin, A. A. (2007). Resource distribution influences mating system in the bobuck (Trichosurus cunninghami: Marsupialia). Oecologia, 154(1), 227–236.
Martin, R. W., & Handasyde, K. A. (1999). The Koala: Natural history, Conservation and Management. Sydney: University of New South Wales.
McGlynn, T. P., Shotell, M. D., & Kelly, M. S. (2003). Responding to a variable environment: home range, foraging behavior, and nest relocation in the Costa Rican rainforest ant Aphaenogaster araneoides. Journal of Insect Behavior, 16(5), 687–701.
McIntyre, N. E., & Wiens, J. A. (1999). Interactions between landscape structure and animal behavior: The roles of heterogeneously distributed resources and food deprivation on movement patterns. Landscape Ecology, 14(5), 437–447.
McNab, B. K. (1963). Bioenergetics and the determination of home range size. American Naturalist, 97(894), 133–140.
Millspaugh, J. J., & Marzluff, J. M. (2001). Radio tracking and animal populations. San Diego: Academic Press.
Milton, K., & May, M. L. (1976). Body weight, diet and home range area in primates. Nature, 259(5543), 459–462.
Mohr, C. O. (1947). Table of equivalent populations of North American small mammals. American Midland Naturalist, 37(1), 223–249.
Montague, T. L. (2000). The brushtail possum: Biology, impact and management of an introduced marsupial. Lincoln: Manaaki Whenua Press.
Moorcroft, P., & Lewis, M. A. (2006). Mechanistic home range analysis. Princeton: Princeton University Press.
Moore, B. D., & Foley, W. J. (2005). Tree use by koalas in a chemically complex landscape. Nature, 435(7041), 488–490.
Moore, B. D., Lawler, I. R., Wallis, I. R., Beale, C. M., & Foley, W. J. (2010). Palatability mapping: A koala's eye view of spatial variation in habitat quality. Ecology, 91(11), 3165–3176.
Nersesian, C., Banks, P., & McArthur, C. (2012). Influences of plant toxins and their spatial distribution on foraging by the common brushtail possum, a generalist mammalian herbivore. Journal of Chemical Ecology, 38(12), 1544–1551.
Ostfeld, R. S. (1985). Limiting resources and territoriality in microtine rodents. American Naturalist, 126(1), 1–15.
Paterson, B., Morris, R., Weston, J., & Cowan, P. (1995). Foraging and denning patterns of brushtail possums, and their possible relationship to contact with cattle and the transmission of bovine tuberculosis. New Zealand Veterinary Journal, 43(7), 281–288.
Perry, G., & Garland Jr, T. (2002). Lizard home ranges revisited: Effects of sex, body size, diet, habitat, and phylogeny. Ecology, 83(7), 1870–1885.
Pope, M. L., Lindenmayer, D. B., & Cunningham, R. B. (2004). Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. I. Home range size and movements. Wildlife Research, 31(6), 559–568.
Powell, R. A. (2000). Animal home ranges and territories and home range estimators. In L. Boitani & T. K. Fuller (Eds.), Research techniques in animal ecology: Controversies and consequences (pp. 65–110). New York: Columbia University Press.
Rhodes, J. R., McAlpine, C. A., Lunney, D., & Possingham, H. P. (2005). A spatially explicit habitat selection model incorporating home range behavior. Ecology, 86(5), 1199–1205.
Ringler, N. H. (1983). Variation in foraging tactics of fishes. In D. L. G. Noakes, D. G. Lindquist, G. S. Helfman & J. A. Ward (Eds.), Predation and prey in fishes (pp. 159–171). Hague: Junk.
Roughgarden, J. (1972). Evolution of niche width. The American Naturalist, 106(952), 683–718.

137
Roughgarden, J. (1979). Theory of population genetics and evolutionary ecology. New York: Macmillan.
Säid, S., Gaillard, J., Widmer, O., Débias, F., Bourgoin, G., Delorme, D., & Roux, C.
(2009). What shapes intra‐specific variation in home range size? A case study of female roe deer. Oikos, 118(9), 1299–1306.
Sandell, M. (1989). The mating tactics and spacing patterns of solitary carnivores. In J. L. Gittleman (Ed.), Carnivore behavior, ecology, and evolution (pp. 164–182). New York: Springer.
Scrivener, N. J., Johnson, C. N., Wallis, I. R., Takasaki, M., Foley, W. J., & Krockenberger, A. K. (2004). Which trees do wild common brushtail possums (Trichosurus vulpecula) prefer? Problems and solutions in scaling laboratory findings to diet selection in the field. Evolutionary Ecology Research, 6(1), 77–87.
Seton, E. T. (1909). Life-histories of northern animals: An account of the mammals of Manitoba (Vol. 2). New York: Charles Scribner's Sons.
Smith, A. C., & Schaefer, J. A. (2002). Home-range size and habitat selection by American marten (Martes americana) in Labrador. Canadian Journal of Zoology, 80(9), 1602–1609.
Smith, G. C., Mathieson, M., & Hogan, L. (2007). Home range and habitat use of a low-density population of greater gliders, Petauroides volans (Pseudocheiridae: Marsupialia), in a hollow-limiting environment. Wildlife Research, 34(6), 472–483.
Spalinger, D. E., & Hobbs, N. T. (1992). Mechanisms of foraging in mammalian herbivores: New models of functional response. American Naturalist, 140(2), 325–348.
Sullivan, T. P., Sullivan, D. S., & Krebs, C. J. (1983). Demographic responses of a chipmunk (Eutamias townsendii) population with supplemental food. The Journal of Animal Ecology, 52(3), 743–755.
Tufto, J., Andersen, R., & Linnell, J. (1996). Habitat use and ecological correlates of home range size in a small cervid: The roe deer. Journal of Animal Ecology, 65(6), 715–724.
Van Beest, F. M., Rivrud, I. M., Loe, L. E., Milner, J. M., & Mysterud, A. (2011). What determines variation in home range size across spatiotemporal scales in a large browsing herbivore? Journal of Animal Ecology, 80(4), 771–785.
Vedder, A. L. (2005). Movement patterns of a group of free‐ranging mountain gorillas (Gorilla gorilla beringei) and their relation to food availability. American Journal of Primatology, 7(2), 73–88.
Winter, J . W. (1976). The behaviour and social organisation of the brush-tail possum (Trichosurus vulpecula Kerr). (Ph.D. thesis), University of Queensland, St Lucia.
Winter, J. W. (1980). Tooth wear as an age index in a population of the brush-tailed possum, Trichosurus vulpecula (Kerr). Australian Wildlife Research, 7(3), 359–363.
Worton, B. J. (1989). Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology, 7(1), 164–168.
Youngentob, K., Wallis, I. R., Lindenmayer, D. B., Wood, J., Pope, M., & Foley, W. J. (2011). Foliage chemistry influences tree choice and landscape use of a gliding marsupial folivore. Journal of Chemical Ecology, 37(1), 71–84.
Youngentob, K. N., Yoon, H.-J., Coggan, N., & Lindenmayer, D. B. (2012). Edge effects influence competition dynamics: A case study of four sympatric arboreal marsupials. Biological Conservation, 155, 68–76.

138
CH
AP
TE
R
4
Dietary choices of the common brushtail possum (Trichosurus vulpecula) in a chemically
complex environment: When not to take a green smoothie?

139
CHAPTER 4
Dietary choices of the common brushtail possum (Trichosurus vulpecula) in a chemically complex
environment: When not to take a green smoothie?
Introduction
In 1977 Janzen wrote,
Plants are not just food for animals, and animals are not just decorations on the
vegetation. The world is not green. It is coloured lectin, tannin, cyanide, caffeine,
aflatoxin, and canavanine. And there is a lot of cellulose thrown in to make the mix
even more inedible. Animals are not ambulatory bomb calorimeters. They starve,
they ache, they abort, they vomit, they remember, they die, and they evolve (Janzen,
2007: 706).
Throughout the last 50 years of herbivore nutritional ecology, numerous studies have
focused on the influence of various plant chemical and physical properties on animal
dietary choices (reviewed by Agrawal & Fishbein, 2006; Carmona, Lajeunesse, & Johnson,
2011). This work has primarily focused on three dominant plant properties: nutritional
quality, including minerals, nitrogen and water content (Scriber, 1977; Mattson, 1980;
McNaughton, 1988); anti-herbivore activity of plant secondary metabolites (Bennett &
Wallsgrove, 1994; Dearing, Foley, & McLean, 2005); and mechanical properties of fibre
and leaf toughness (Lucas, Turner, Dominy, & Yamashita, 2000).
In the past, researchers have acknowledged the ecological importance of both nutrients and
plant secondary metabolites for herbivores (Coley & Barone, 1996; Molyneux & Ralphs,
1992). Yet, they have rarely managed to reconcile the complex interactions between these
opposite, but mutually complementary, aspects of plant chemistry. Many researchers
considered the availability of plant protein, expressed as foliar total nitrogen content, as a
single limiting factor for wild populations of herbivores, directly affecting their
maintenance, growth and reproduction (Cockfield, 1988; Joern & Behmer, 1997; White,
1984). Other studies analysed the toxic effects of different groups of plant secondary
metabolites (i.e. phenolics, terpenes, cyanogenic glycosides) on the metabolism and
foraging behaviour of herbivores (Rosenthal & Berenbaum, 1992).

140
Conversely, one of the most ubiquitous groups of plant secondary metabolites, tannins,
do not exhibit toxic effects on herbivores and instead act as forage digestibility reducers
and herbivory deterrents (Mole & Waterman, 1987; Schultz, 1989).Tannins are known to
represent 5–10% of plant biomass and are water-soluble phenolic compounds that bind
and precipitate proteins and other macronutrients within aqueous solutions, severely
decreasing the nutritive value of browse by reducing its digestibility (Barbehenn & Peter
Constabel, 2011; Mueller-Harvey, 2006). Herbivores not only consume tannins that render
nitrogen unavailable, but also tannins that may cause oxidative stress on an animal’s
digestive tract via auto-oxidation at high pH or enzymatic oxidation at lower pH
(Barbehenn & Peter Constabel, 2011; Salminen & Karonen, 2011). Nutritional
requirements of herbivores are further constrained as tannins decrease fibre digestibility by
binding bacterial enzymes or forming indigestible complexes with cell wall constituents or
both (Reed, 1995).
Differential activity of tannins has encouraged the evolution of various strategies by
herbivores, such as avoiding tannins or tolerating the negative effects of tannins (Iason &
Villalba, 2006). Some animals remain highly sensitive to even the smallest amounts of
tannins and cease feeding after experiencing the distinctively astringent taste, e.g., rodents
(Shimada & Saitoh, 2003), lagomorphs (Clausen, Provenza, Burritt, Reichardt, & Bryant,
1990) and primates (Takemoto, 2003). Other groups, like ruminants, have developed post-
ingestive physiological adaptations to counteract tannins, including the secretion of tannin-
binding proteins in the saliva, the increased gastrointestinal mucus production, the
degradation of tannins by gut microorganisms, and the activation of detoxifying enzymes
(reviewed by McArthur, Hagerman, & Robbins, 1991). Insects for instance, maintain highly
alkaline gut conditions to dissolve tannin-protein complexes (i.e., lepidopteran larvae,
Berenbaum, 1980) or use enzymes β-glucosidase to hydrolyse tannic acid or adsorb it on
the peritrophic membrane (i.e., locust, Bernays, 1981).
On the other side, arboreal marsupials inhabiting Australian forests and woodlands
dominated by a single genus Eucalyptus subsist dominantly on nitrogen-poor, but tannin-
rich and highly fibrous foliage. Tannins can represent up to 27% and nitrogen 0.8–2% of
leaf dry matter, and arboreal marsupials have developed a suite of morphological,
physiological, and behavioural adaptations to digest low quality forage (Macauley & Fox,
1980; Moore et al., 2004a). For instance, a eucalypt specialist, the koala (Phascolarctos
cinereus), possesses gut microflora actively metabolizing tannin-protein complexes.

141
Namely, Streptococcus bovis biotope I and tannin-protein complex-enterobacterium, occurring
frequently in the digestive tract of koala, have a unique property of degrading proteins
complexed with hydrolysable tannins (Osawa, 1991).
Another eucalypt specialist, the common ringtail possum (Pseudocheirus peregrinus), is less
sensitive to tannins and has a well-developed caecum that selectively retains bacteria and
other nitrogen-rich small particles and a well-developed dentition that reduces highly
fibrous foliage to small particles (Hume, 1999). This species also practices caecotrophy to
return nitrogen-rich caecal contents to the digestive tract (Marsh et al., 2003; McArthur &
Sanson 1991, 1993). Conversely, a dietary generalist, the common brushtail possum
(Trichosurus vulpecula), is more sensitive to tannins because of the immediate oral sensation
of the astringent taste of foliage. In addition, the common brushtail possum has less
specialized dentition and a small caecum relative to its body size with no evidence of a
separation mechanism (Hume, 1999).
High sensitivity of common brushtail possums to tannins has been demonstrated by Foley
and Hume (1987), who added polyethylene glycol 4000 (PEG), a tannin-binding agent, to
the drinking water of captive animals maintained on the sole diet of Eucalyptus melliodora
foliage. E. melliodora foliage contains 20–29% total phenolics and supplementation with
PEG increased the common brushtail possums dry matter intake by 32%, their digestibility
of neutral-detergent fibre by 77%, and consequently increased their intake of metabolisable
energy by 67%. The effects were attributed to the reversal of the tannin-microbial enzyme
complexes in the hindgut. Furthermore, common brushtail possums are known to possess
tannin-binding proteins in saliva, but their efficacy against detrimental effects of tannins is
limited (McArthur et al., 1995).
Apart from tannins Eucalyptus foliage typically contains a plethora of toxins, including
terpenes, cyanogenic glycosides and potent antifeedants, which are formylated
phloroglucinol compounds (FPCs) found only in eucalypts from subgenus Symphyomyrtus
(Eschler et al., 2000; Moore et al., 2004b; Gleadow et al., 2008). Nevertheless, common
brushtail possums can overcome the negative effects of toxins by having a broad diet and
activating different detoxification pathways (Marsh et al., 2006b; Wiggins, McArthur, &
Davies, 2006; Nersesian et al., 2012).

142
For instance, in contrast to more specialized mountain brushtail possums, common
brushtail possums are tolerant of sesquiterpenes, one of the most widespread terpene
groups among eucalypt species, due to their high glucuronidation capacity involved in the
detoxification of terpenes (Trichosurus cunninghami; Burchfield, Agar & Hume 2005).
Furthermore, common brushtail possums can recognise cyanogenic glycosides, such as
cyanide, and avoid these highly toxic compounds while foraging (Clapperton et al., 1996).
Also, formylated phloroglucinol compounds known to deter koalas and common ringtail
possums from foraging on many eucalypt species and individuals of the same species have
been found to affect common brushtail possums to a lesser extent (Marsh et al., 2003).
Consequently, this recognized tolerance of plant secondary metabolites by common
brushtail possums has shifted once again nutritional ecologists’ attention from plant
toxicity to plant nutritional quality.
Recently, a new foraging theory, the nitrogen availability hypothesis, has been proposed by
DeGabriel et al. (2009a, 2010) to explain herbivore foraging behaviour, habitat suitability,
and life strategies. According to this hypothesis, foliar concentration of available nitrogen,
rather than total nitrogen, tannins, and fibre individually, is a more ecologically meaningful
measure of plant nutritional quality for herbivores. A method developed by DeGabriel and
colleagues (2008) uses a simple in vitro assay to integrate foliar total nitrogen, fibre, tannins,
and dry matter digestibility into a single measure of available nitrogen to herbivores (see
Methods for detailed description).
The nitrogen availability hypothesis has been tested empirically on arboreal marsupials
(DeGabriel et al., 2009a; Stalenberg, 2010; Youngentob et al., 2011), moose (McArt et al.,
2009), and spider monkeys (Felton et al., 2009b). These nutritional studies ranged from
laboratory based feeding trials to foraging observations of wild populations and uniformly
found that both specialist and generalist herbivores preferred leaves from species and
individuals with lower levels of tannins and higher available nitrogen. Furthermore,
DeGabriel et al. (2009) demonstrated that the natural variation of available nitrogen rather
than total nitrogen among eucalypts growing in otherwise homogenous habitats has
affected the reproductive success of individual common brushtail possums. Hence, tannins
were identified as the main dietary constituent that limits the availability of nitrogen and
best explains reproductive output, explicitly linking herbivore nutrition and population
dynamics.

143
Wallis et al. (2012) argued that the main advantage of measuring foliar concentrations of
available nitrogen is in its broad, simple approach that allows the comparison of diverse
vegetation groups using the same currency. This new approach is especially relevant in the
Australian context, where the previous studies rarely acknowledged the dietary breadth of
arboreal marsupials and focused only on measuring the nutritional quality of eucalypts. A
great deal of work has been dedicated to explain how the Eucalyptus chemical composition
and variability affect arboreal marsupial foraging behaviour; however, herbivory on native
Acacia species has received much less attention (Dynes & Schlink, 2002; Irlbeck & Hume,
2003).
Additionally, hardly any data exist on the chemical composition of plants parasitising roots
and branches of different Eucalyptus and Acacia species and their overall effects on
herbivores (Close, Davidson, & Davies, 2006; Hensens, Lewis, & Mulquiney, 1971;
Preston, An, & Watson, 2010). Hence, the present study assessed for the first time the
nutritional quality of co-occurring Eucalyptus and Acacia species and parasitic plants,
Amyema pendula and Exocarpos cupressiformis, for a generalist arboreal marsupial, the common
brushtail possum (Trichosurus vulpecula).

144
Aims and hypotheses
The specific aims of this work focused on two research areas with underlying hypotheses
drawn from the existing research:
1. To investigate the chemical variability of trees and parasitic plants available to common brushtail
possums within their home ranges.
Hypothesis 1: Nitrogen-fixing Acacia species will have the highest foliar total nitrogen
concentrations, followed by nitrogen-accumulating parasitic plants and nitrogen-poor
Eucalyptus species (Irlbeck & Hume, 2003; Watson, 2009).
Hypothesis 2: Species from the same genera will have similar concentrations of foliar total
nitrogen, but will vary in concentrations of available nitrogen, tannins, and fibre
(DeGabriel, 2008; Wallis, Nicolle, & Foley, 2010).
Hypothesis 3: Parasitic plants should have the highest dry matter digestibility and lowest
dry matter content, followed by sclerophyllous Eucalyptus species and Acacia species
(McLeod, 1973; Umucalilar, Gulsen, Coskun, Hayirli, & Dural, 2007).
Hypothesis 4: Intra-specific chemical variability will be observed among different tree
species and parasitic plants, with the latter being more variable depending on the species of
the tree host (Lawler, Foley, Eschler, Pass, & Handasyde, 1998; Snyder, Fineschi, Linhart,
& Smith, 1996).
2. To establish the influence of chemical variability on the common brushtail possum’s dietary choices
and their home range size.
Hypothesis 1: Common brushtail possums are expected to prefer genera, species, and
individuals with higher foliar concentrations of available nitrogen, dry matter digestibility,
and lower dry matter content (after DeGabriel et al., 2009a).
Hypothesis 2: The size of common brushtail possum home ranges will be affected by the
nutritional quality of browse, with larger home ranges occurring in areas of lower
nutritional quality (after DeGabriel et al., 2009a).

145
Methods
Description and justification of methods used for measuring the nutritional quality of forage
The key factors explaining herbivore dietary choices are foliar concentrations of total
nitrogen, a proxy of protein content; tannins, one of the most ubiquitous plant secondary
metabolites; and fibre as a cell-wall constituent. In the past, many studies of the nutritional
quality of forage for herbivores overlooked the complex interactions between these
mutually complementary factors. Only recently, DeGabriel and colleagues (2008) proposed
a new integrative approach for measuring the additive and interactive effects of tannins and
fibre on nitrogen availability to herbivores. This method begins with two-stage in vitro
enzymatic digestion to establish the amount of foliar nitrogen and fibre digested by
herbivores; it then uses polyethylene glycol (PEG), a tannin binding agent, to determine the
amount of nitrogen inhibited by tannins. Application of PEG provides a direct method for
measuring the biological effects of tannins on herbivores without a need to classify them
into hydrolysable or condensed tannins (Silanikove et al., 1996). Additionally, this method
provides a uniform comparative approach for studying cross-taxa chemical variability of
plants, overcoming the problem of otherwise incomparable genera and species (W. J. Foley,
pers. comm., February 2009). Hence, this simple integrative approach was considered to be
the most suitable method for quantifying and comparing the nutritional quality of different
trees and parasitic plants available to common brushtail possums within their home ranges.
Another problem encountered in this study was finding a robust method for chemically
analysing a large number of leaf samples from the common brushtail possums’ relatively
large home ranges (see Chapter 3). Near Infrared Spectroscopy (NIRS) proved to be a
simple, time- and cost-effective alternative to conventional analytical methods (Foley et al.,
1998). This method employs multivariate statistical models to relate spectral characteristics
of a plant material to its chemical properties. In particular, when samples are irradiated with
near-infrared light their reflectance spectra represent a mixture of chemical bonds (C—H,
N—H, and O—H) characteristic for a specific chemical compound. These spectra are then
statistically calibrated against the reference values for a specific chemical compound that is
analysed using traditional laboratory methods. Next, multivariate regression equations are
developed using the partial least square regression method to estimate a chemical
composition of multiple samples. Thus, NIRS represents a practical and robust alternative
to the complicated, slow, and expensive chemical methods used for estimating foliar
chemical composition of multiple species over large geographic areas (Ebbers et al., 2002).

146
Sampling design and collection of leaf material
To describe the foraging environment of common brushtail possums and quantify and
compare nutritional quality of different trees and parasitic plants, leaf material was collected
from six randomly chosen home ranges (2 male and 4 female, as described in Chapter 3).
The sampling design included trees used (n1 = 98) and unused by common brushtail
possums (n2 = 98) while foraging at night, as well as trees parasitized by drooping mistletoe
(Amyema pendula; n3 = 75) and unparasitized trees (n4 = 75; Fig. 1). Additionally, 75 samples
of mistletoe foliage were collected from both Eucalyptus and Acacia tree hosts. Used trees
were considered to represent an accurate proxy of possum foraging; however, the
possibility of false absences could not be ruled out (Martin & Handasyde, 1999).
Figure 2 illustrates locations of all sampled trees, including possum used and mistletoe
parasitized trees. Tree diameter and height were recorded using a hand-held laser
hypsometer (LaserAce®, MDL Laser Systems, UK). Since diameter and height were highly
correlated (Spearman’s Rho = 0.79, P < 0.001), only tree diameter was reported as a proxy
of tree age in this study. Moreover, biomass of all sampled trees was calculated using a
general allometric equation developed by Keith, Barrett, and Keenan (2000; see Chapter 2
for details).
The collection of leaf material was carried out during a two week sampling period in April
2009 following the end of a radio-tracking survey of common brushtail possum performed
to establish space use patterns and resource selection (see Chapter 3). A single sampling
effort was considered to be an accurate representation of the nutritional quality of browse
available to possums at a given point in time; therefore, overcoming the problem of
seasonal fluctuations in foliar nitrogen and tannin levels (Cooper, Owen-Smith, & Bryant,
1988; Fox & Macauley, 1977). In total, 418 leaf samples were collected, with on average
69.7 ± 4.2 samples per home range. These samples included nine species from Eucalyptus,
two from Acacia, and the parasitic species, Amyema pendula and Exocarpos cupressiformis
(Table 1).
For the purpose of this study, the genus Eucalyptus was divided into subgenera Monocalyptus
and Symphyomyrtus, since the two subgenera are known to differ in plant secondary
metabolites and dependant groups of herbivores (Eschler et al., 2000; Gleadow et al., 2008;
Noble, 1989). Despite a small sample size (n = 5), Eucalyptus obliqua from the subgenus
Monocalyptus was included in the analysis since its foliar chemical composition has not been
previously studied.

147
To establish chemical differences between parasitic plants and their tree hosts, only
samples of Amyema pendula were analysed since it was impossible to identify tree hosts of
the root parasite, Exocarpos cupressiformis, without extensive root excavation (Table 2). A
single terminal branch containing both mature and juvenile leaves was collected from the
centre of a tree canopy using handheld secateurs mounted on a telescopic aluminium pole
or with the help of a professional arborist (Tree Tactics, Pty Ltd). Since common brushtail
possums are known to forage on both mature and juvenile leaves, they (50–100 g) were
analysed together to establish average browse nutritional quality (Loney, McArthur, Potts,
& Jordan, 2006). Collected samples were placed on ice in the field and later transferred to a
-20°C freezer to prevent chemical degradation of leaf material. Leaves were then freeze
dried (CSIRO, Canberra, ACT) and passed through a 1 mm sieve in a Cyclotec 1093 mill
(Tecator, Sweden).
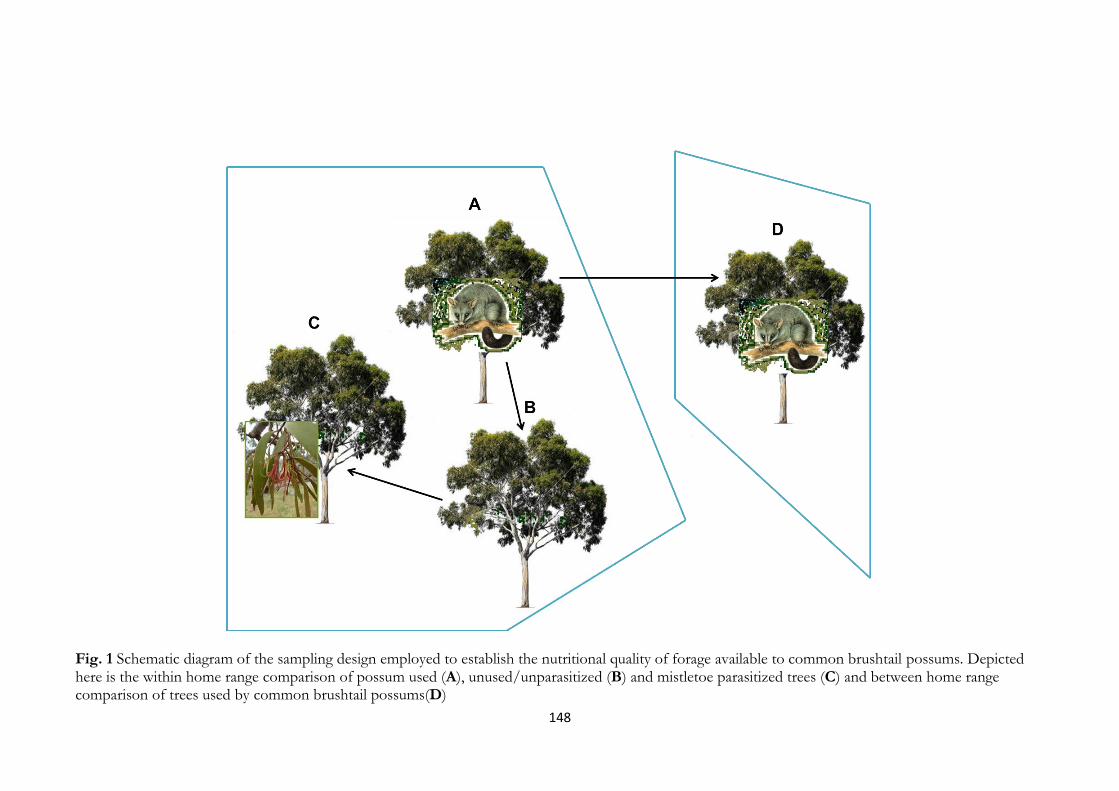
148
Fig. 1 Schematic diagram of the sampling design employed to establish the nutritional quality of forage available to common brushtail possums. Depicted here is the within home range comparison of possum used (A), unused/unparasitized (B) and mistletoe parasitized trees (C) and between home range comparison of trees used by common brushtail possums(D)

149
Fig. 2 Location of the sampled trees within home ranges of the six common brushtail possums (CBP; 2 male and 4 female) in the Strathbogie Ranges, Victoria

150
Table 1 Summary of foliage samples collected within the six common brushtail possum home ranges
Table 2 Summary of Amyema pendula foliage samples collected from different species of tree hosts within the six common brushtail possum home ranges
Genus
Species
No samples
Acacia
Acacia dealbata 41 Acacia melanoxylon
21
Amyema
Amyema pendula
75
Exocarpos
Exocarpos cupressiformis
22
Monocalyptus
Eucalyptus radiata 95 Eucalyptus dives Eucalyptus obliqua
17 5
Symphyomyrtus
Eucalyptus camphora 75 Eucalyptus viminalis 39 Eucalyptus globulus
21
Genus
Species
No mistletoe samples
Acacia
Acacia dealbata
12
Monocalyptus
Eucalyptus radiata 31 Eucalyptus dives
7
Symphyomyrtus
Eucalyptus camphora 12 Eucalyptus viminalis 8 Eucalyptus globulus
5

151
Near-Infrared Spectroscopy (NIRS)
Collection of near-infrared spectra of leaf material
To minimize residual moisture and particle size affecting readings of near-infrared spectra,
all freeze dried and ground leaf samples (uniform sample particle size 1 mm) were placed in
an oven at 40⁰C for at least 1 h to remove residual moisture. Samples were then spread
evenly in a scanning quarter cup and were scanned with a near-infrared reflectance
spectrometer with a spinning cup attachment (Model 6500, Foss NIRSystems, Silver
Spring, MD) at the Australian National University, Canberra, ACT (after Foley et al., 1998).
Reflectance spectra of all samples were collected between 408–1093 nm and 1108–2493
nm. Each sample was scanned twice or until the root mean square of the two scans (scored
as log (1/reflectance)) was less than 3.0 x 10-4. The spectra were then averaged.
For an independent validation set, 20 samples including all studied species were selected.
Calibration set and NIRS equations development
The purpose of the calibration process in this study was to develop Near-Infrared
Spectroscopy equations used to predict the concentrations of chemical compounds for all
samples based on their near-infrared spectra (n = 418; see Laboratory Analyses for
detailed description of selected chemical compounds). To identify samples most suitable
for NIRS equations development, the algorithm SELECT was used in the NIRS 3, version
4.00 Software (WinISI Infrasoft International, Port Matilda, PA). The algorithm SELECT
employs principal component analysis (PCA) and Mahalanobis distances (H statistics) to
identify a subset of samples with spectra representing the full spectral variability for all
collected samples; these samples formed the calibration set (Shenk & Westerhaus, 1991).
The calibration set was then subjected to chemical analyses to obtain reference values for
developing the best NIRS equations. In this study, 158 samples were selected as the most
representative for a calibration set and their chemical composition was analysed in the
laboratory.

152
The next step in developing NIRS equations was to choose by a trial-and-error approach
the equations with lowest standard error of calibration (SEC) and highest coefficient of
determination (R2) for each of the studied chemical compounds using different regression
models and mathematical treatments to reduce background effects (Foley et al., 1998;
Woolnough & Foley, 2002). After trying a number of regression models, the modified
partial-least squares regression (MPLS) proved to be the most suitable for developing
NIRS equations apart from one case where partial-least squares regression (PLS) was used
(Table 3). The MPLS and PLS techniques combine the principal component analysis
(PCA) and multiple linear regression analyses. PCA is used to reduce the number of
measured near infrared spectra to just a few combinations suitable for describing most
spectral variability while multiple linear regression relates spectra to the reference values of
the calibration set. In addition to MPLS and PLS, a variety of mathematical treatments
were used to reduce the potential variation in spectra due to differences in sample particle
size and residual moisture, as well as to reduce multicollinearity caused by developing
equations based on more than one wavelength (Batten, 1998; Table 3). For example, a
mathematical treatment of “2, 4, 4, 1” refers to using the second derivative, leaving the gap
of four wavebands between calculated values, performing a first smoothing over four
wavebands and then a second smoothing over one waveband.

153
Table 3 Calibration statistics and mathematical treatments of the Near-Infrared Spectroscopy equations†.† Abbreviations: constituent = foliar chemical
attribute measured in the presence and absence of polyethylene glycol (PEG); n = number of samples used to develop the NIRS equations; = mean value of samples; SD = standard deviation from the mean; SECV = standard error of cross validation; R² and 1-VR = degree of correlation between predicted and actual measures; Scatter correction = standard normal variate (SNV) and detrend of mathematical transformations; Math treatment = Savitzy-Golay spectral-smoothing function; Regression type = modified partial least square (MPLS) and partial least square regression (PLS)
Constituent
N
SD
R²
SECV
1-VR
Wavelength used
Scatter correction
Math treatment
Regression type
Total N 154 1.59 0.48 0.99 0.07 0.98 All SNV Detrend 2, 4, 4, 1 MPLS
DMD + PEG 148 46.76 7.76 0.85 3.42 0.81 All SNV 1, 8, 8, 1 MPLS
DMD 153 48.29 8.38 0.86 4.09 0.76 All SNV 2, 8, 8, 1 MPLS
N digestibility + PEG 151 0.70 0.07 0.55 0.05 0.45 1108–2492.8 None 2, 4, 4, 1 PLS
N digestibility 150 0.44 0.16 0.86 0.08 0.79 All SNV Detrend 2, 6, 4, 1 MPLS
Available N + PEG 153 1.11 0.37 0.94 0.11 0.91 1108–2492.8 SNV 2, 8, 8, 1 MPLS
Available N
150
0.74
0.41
0.93
0.12
0.91
All
SNV Detrend
2, 4, 4, 1
MPLS

154
Validation procedures
The validity of the NIRS equations was tested twice. The first test was used when
performing the cross-validation procedure during the development of equations for all
analysed samples (Shenk & Westerhaus, 1991). This cross-validation procedure involves
separating all samples into groups and performing calibrations on all but one of the groups,
which is then used as an independent validation set. The procedure is repeated multiple
times until all groups of samples have been cross-validated. In this way, SECV (standard
error of cross-validation) and the coefficient of the determination of cross-validation
(1-VR) are generated, which explained the error associated with predictions (Table 3).
In the second test, the validity of the NIRS equations was evaluated only for a calibration
set (n = 158) by fitting simple linear regression models of predicted concentrations of
selected chemical compounds against concentrations of the same compounds measured in
the laboratory. Predicted concentrations of the selected foliar compounds strongly
correlated with the concentrations measured in the laboratory (P < 0.001, Fig. 3).
Moreover, to further investigate the accuracy and robustness of NIRS equations, an
independent set of 20 samples was analysed using conventional laboratory techniques. The
validity of the NIRS equations was again assessed using simple linear regression. The
predicted concentrations of the selected foliar compounds fitted well with the measured
ones (P < 0.001, Fig. 4).

155
a)
b)
c) Fig. 3 Linear regression of Near-Infrared Spectroscopy predicted and measured in the laboratory concentrations of the selected foliar compounds for a calibration set (n = 158). Figures represent a) total nitrogen (% DM); b) available nitrogen (% DM); c) dry matter digestibility (% DMD). Dotted lines at y = x are for reference only

156
a)
b)
c) Fig. 4 Linear regression of Near-Infrared Spectroscopy predicted and measured in the laboratory concentrations of the selected foliar compounds for a validation set (n = 20). Figures represent a) total nitrogen (%DM); b) available nitrogen (%DM); c) dry matter digestibility (%DMD). Dotted lines at y = x are for reference only

157
Laboratory chemical analyses
In vitro determinations of foliar digestibility and nutrient availability
Considering that only limited amounts of foliar dry matter and total nitrogen are actually
digestible by herbivores, a multiple-step chemical analysis was performed to establish the
amount of nitrogen available to herbivores (Fig. 5). First, to quantify foliar dry matter
digestibility and nitrogen digestibility, the in vitro two-stage enzymatic digestion was
performed using pepsin/cellulase following the method of DeGabriel et al. (2008).
The in vitro pepsin/cellulase digestion emulated digestion of foliar material in hindgut
fermenters, with pepsin simulating nitrogen digestion and cellulase simulating fibre
digestion. In detail, samples from a calibration set (n = 158) and an independent validation
set (n = 20) were weighed into pre-weighed filter bags (ANKOM F57, ANKOM
Technology, Macedon, NY) in four 0.80 ± 0.01 g replicates. Next, duplicate samples were
treated with polyethylene glycol (PEG 4000 analytical grade, Sigma, St. Louis, MO) in
0.05 M Tris-BASE buffer. The remaining control samples were treated only with buffer.
The samples were placed in an incubator at 33°C and gently stirred for 24 h. All samples
were washed, dried, and weighed to a constant mass. Samples were then incubated in
pepsin (2.00 g 1:10,000 pepsin in 1 L 0.1 N HCl) for 24 h, washed, and placed in cellulase
(6.25 g cellulase in 1 L of 100 mmol acetic acid buffer) for 48 h and stirred gently at 33°C.
Following enzymatic digestion and washing, samples were oven-dried at 60°C to a constant
mass and remaining indigested residue weighed to calculate dry matter digestibility (%).
To estimate nitrogen digestibility (Equation 1, Fig. 5), the concentrations of total nitrogen
(% DM) and nitrogen left after digestion (% DM) were measured using the Dumas
technique with a LECO TrueSpec combustion N analyser calibrated against EDTA
(LECO Corporation, St. Joseph, MI). Next, available nitrogen (% DM) concentration was
obtained by multiplying nitrogen digestibility with total nitrogen (% DM) in the presence
and absence of PEG (Equation 2). Finally, by calculating the amount of available nitrogen
(% DM) bound with PEG, it was possible to establish amounts of tannins and fibre
bounded with nitrogen (Equations 3 and 4). The concentration of available nitrogen,
tannin- and fibre-bound nitrogen summed up to total N (% DM; Equation 5, Fig. 5).
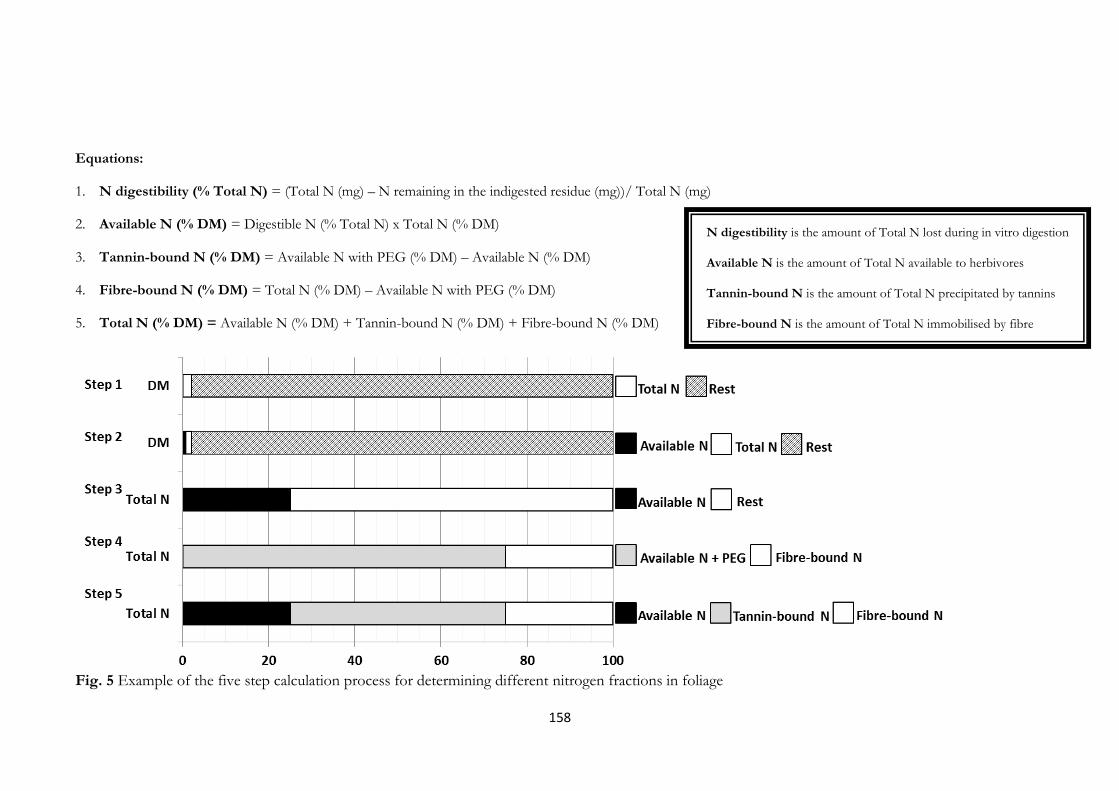
158
Equations: 1. N digestibility (% Total N) = (Total N (mg) – N remaining in the indigested residue (mg))/ Total N (mg) 2. Available N (% DM) = Digestible N (% Total N) x Total N (% DM) 3. Tannin-bound N (% DM) = Available N with PEG (% DM) – Available N (% DM) 4. Fibre-bound N (% DM) = Total N (% DM) – Available N with PEG (% DM) 5. Total N (% DM) = Available N (% DM) + Tannin-bound N (% DM) + Fibre-bound N (% DM)
Fig. 5 Example of the five step calculation process for determining different nitrogen fractions in foliage
N digestibility is the amount of Total N lost during in vitro digestion
Available N is the amount of Total N available to herbivores
Tannin-bound N is the amount of Total N precipitated by tannins
Fibre-bound N is the amount of Total N immobilised by fibre

159
Data analysis
To investigate relationships between the studied chemical compounds, the Spearman’s
rank-order correlation and the simple linear regression were performed. To compare inter-
and intra-specific chemical variability of trees and parasitic plants available to common
brushtail possums within their home ranges, the one-way analysis of variance (ANOVA)
analysis with the post-hoc Tukey’s multiple comparisons test was carried out. Data were
tested for normality using the Shapiro-Wilk test and were log transformed if they departed
from normality. To investigate differences in proportions of available nitrogen, tannin-
bound nitrogen, and fibre-bound nitrogen across different species, the test for equality of
proportions was used, followed by a pair-wise comparison of species with the Bonferroni
correction for alpha.
To establish differences between trees used and unused by common brushtail possums, as
well as trees parasitized and unparasitized by mistletoes, the Welch’s t-test was used. The
Kruskal-Wallis test was used to compare species differences across common brushtail
possum home ranges. To identify which chemical compounds best explained possum tree
use and mistletoe tree parasitism, the binomial generalized linear model was fitted to all
species and next to each species individually (Nelder & Wedderburn, 1972). Firstly, any
highly inter-correlated explanatory variables (Spearman’s Rho > 0.7) were excluded from
the analysis to avoid multicollinearity in the modelling process (Tabachnick & Fidell, 1996).
Secondly, the stepAIC function from the MASS library was used to find the best
combination of explanatory variables. The stepAIC function used AIC as the step criterion
in both directions. All results were reported as observed means ± s.e. and statistical
significance followed the convention of P < 0.05. All statistical analyses were performed in
Spotfire S+® Version 8.1 (2010, TIBCO Software Inc. Somerville, MA).

160
Results
Foliar chemical attributes
Foliar concentrations of total and available nitrogen (% DM) were strongly positively
correlated in all studied plant genera (Spearman’s Rho = 0.86, P < 0.001, Fig. 6) with
R2 ranging from 0.86 for Acacia to 0.96 for Amyema (Fig. 8). An exception was the
subgenus Monocalyptus, in which foliar concentrations of available nitrogen were more
variable than those observed for total nitrogen, leading to a much lower coefficient of
correlation (R2 = 0.37) compared to subgenus Symphyomyrtus (R2 = 0.88). Further
examination of variation in the subgenus Monocalyptus showed strong differences between
species with R2 values ranging from 0.70 for Eucalyptus radiata to 0.30 for E. obliqua. Both
total and available nitrogen were negatively correlated with tree diameter across all genera
(Spearman’s Rho = - 0.53, - 0.58 respectively, P < 0.001).
No correlation was observed between foliar concentrations of total nitrogen and tannin-
bound nitrogen (% DM) when considering all species (Spearman’s Rho = 0.06, P < 0.001,
Fig. 6). However, when analysed separately, tannin-bound nitrogen of Acacia foliage
proved to be moderately correlated with total nitrogen (% DM; R2 = 0.39, Fig. 9).
Conversely, tannin-bound nitrogen was negatively correlated with available nitrogen (%
DM) while both total and available nitrogen were positively correlated with fibre-bound
nitrogen (% DM; Fig. 6). Foliar concentrations of fibre-bound nitrogen (% DM) were
highly variable for genera Amyema and Symphyomyrtus producing lower R2 values compared
to the other genera (Fig. 10). Tree diameter was negatively correlated with fibre-bound
nitrogen (% DM; Spearman’s Rho = - 0.46, P < 0.001) and positively correlated with
tannin-bound nitrogen (% DM; Spearman’s Rho = 0.16, P < 0.001).
Nitrogen digestibility (% total N) and dry matter digestibility (% DMD) were positively
correlated (Spearman’s Rho = 0.68, P < 0.001, Fig. 7). Furthermore, both nitrogen
digestibility and dry matter digestibility were positively correlated with available nitrogen
(% DM; Spearman’s Rho = 0.78, 0.36 respectively), negatively correlated with tannin-
bound nitrogen (% DM; Spearman’s Rho = - 0.63, - 0.73 respectively), and negatively
correlated with foliar dry matter content (Spearman’s Rho = - 0.36, - 0.30 respectively, P <
0.001, Fig. 7).

161
Fig. 6 Spearman’s Rho correlations of selected chemical variables for all species (n = 413). An asterix (*) indicates a significant correlation at P < 0.001. Arrows show direction and strength of correlation
Figure 7 Spearman’s Rho correlations of foliar nitrogen and dry matter digestibility for all species (n = 413). An asterix (*) indicates a significant correlation at P < 0.001. Arrows show direction and strength of correlation

162
Fig. 8 Linear regression of total nitrogen (% DM) against available nitrogen (% DM) of different genera and Eucalyptus subgenera. The equation and R2 value refer to linear regression performed on all samples (n = 413)
Fig. 9 Linear regression of total nitrogen (% DM) against tannin-bound nitrogen (% DM) of different genera and Eucalyptus subgenera. The equation and R2 value refer to linear regression performed on all samples (n = 413)

163
Fig. 10 Linear regression of total nitrogen (% DM) against fibre-bound nitrogen (% DM) of different genera and Eucalyptus subgenera. The equation and R2 value refer to linear regression performed on all samples (n = 413)

164
Foliar nitrogen composition of trees and parasitic plants
The foliar total nitrogen concentration was highest in nitrogen-fixing Acacia, followed by
the root parasite, Exocarpos, and eucalypts from subgenus Symphyomyrtus (% DM; P < 0.001,
Fig. 11). Against expectations, an aerial parasite Amyema and eucalypts from the subgenus
Monocalyptus had the lowest concentrations of total nitrogen (% DM; P < 0.001, Fig. 11).
Plant genera varied in foliar concentrations of available nitrogen, tannin- and fibre-bound
nitrogen (% DM; P < 0.001, Fig. 12). Genus Monocalyptus had the lowest concentration of
available nitrogen and was followed by genera Amyema and Symphyomyrtus, which had similar
concentrations of available nitrogen. The genus with the highest concentration of available
nitrogen was Acacia, which had also the highest concentration of fibre-bound nitrogen
while Monocalyptus had the highest concentration of tannin-bound nitrogen (% DM;
P < 0.001, Fig. 12).

165
Fig. 11 Mean foliar concentration of total nitrogen (% DM) with standard error (s.e.) of different plant genera and Eucalyptus subgenera. Letters above the bars represent significant differences between the groups at P < 0.001
Fig. 12 Mean foliar concentrations of available nitrogen (black bar), tannin-bound nitrogen (grey), and fibre-bound nitrogen (white) with standard error (s.e.) of different plant genera and Eucalyptus subgenera. The foliar concentrations of available nitrogen, tannin-bound nitrogen and fibre-bound nitrogen sum to total nitrogen (% DM) concentration

166
From all species Acacia dealbata had the highest concentration of foliar total nitrogen
(%DM) while Eucalyptus camphora had the highest total nitrogen (% DM) of all Eucalyptus
species (P < 0.001, Fig. 13, Table 4). Of the two parasitic species, Exocarpos cupressiformis
had higher concentration of foliar total nitrogen (% DM) than Amyema pendula (P < 0.001,
Fig. 13, Table 4). Acacia dealbata had the highest available nitrogen (% DM), followed by
Exocarpos cupressiformis and Eucalyptus camphora while species from Monocalyptus subgenera
had the lowest (P < 0.001, Fig. 13, Table 4). Moreover, Monocalyptus species had the
significantly highest foliar concentrations of tannin-bound nitrogen (% DM), especially
Eucalyptus radiata (P < 0.001, Fig. 13, Table 4). Foliar concentrations of tannin-bound
nitrogen (% DM) were the lowest in E. viminalis and Amyema pendula while fibre-bound
nitrogen (% DM) was the highest in the two Acacia species, A. dealbata and A. melanoxylon
(P < 0.001, Fig. 13, Table 4).
All studied species had similar proportions of fibre-bound nitrogen ranging from 26 to
33% (χ2 = 1.65, df = 9, P = 0.996 Fig. 14). Also, most of the species had similar
proportions of tannin-bound nitrogen (13–17%), apart from the species from the subgenus
Monocalyptus (χ2 = 34.49, df = 9, P < 0.001) with Eucalyptus radiata having 44 % of total
nitrogen bound with tannins. Consequently, due to extremely high proportions of tannins
in Monocalyptus species, only small amounts of total nitrogen (5–27 %) were actually
available to herbivores in comparison to other species where the proportions of available
nitrogen ranged from 41 % for E. globulus to 61 % for Exocarpos cupressiformis (χ2 = 24.19,
df = 9, P < 0.05).

167
Fig. 13 Mean foliar concentrations of available nitrogen (black bar), tannin-bound nitrogen (grey), and fibre-bound nitrogen (white) with standard error (s.e.) of different plant genera, Eucalyptus subgenera and species. The foliar concentrations of available nitrogen, tannin-bound nitrogen, and fibre-bound nitrogen sum to total nitrogen (% DM) concentration

168
Fig. 14 Percentage (%) of available nitrogen (black bar), tannin-bound nitrogen (grey), and fibre-bound nitrogen (white) in foliage of different plant species. The foliar concentrations of available nitrogen, tannin-bound nitrogen, and fibre-bound nitrogen sum to total nitrogen (% DM) concentration

169
Dry matter digestibility and dry matter content of trees and parasitic plants
The species with the highest dry matter digestibility (% DMD) was drooping mistletoe
Amyema pendula (P < 0.001, Fig. 15, Table 4), followed by Exocarpos cupressiformis and
Eucalyptus viminalis. The species with the lowest dry matter digestibility (% DMD) were
Eucalyptus dives and E. radiata, both from genus Monocalyptus (P < 0.001, Fig. 15, Table 4).
The genera with the significantly lowest dry matter content (%) were parasitic Amyema and
Exocarpos while Monocalyptus had the highest (P < 0.001, Fig. 16).
Fig. 15 Mean foliar dry matter digestibility (% DMD) with standard error (s.e.) of different plant genera and Eucalyptus subgenera. Letters above the bars represent significant differences among the groups at P < 0.001
Fig. 16 Mean foliar dry matter content (%) with standard error (s.e.) of different plant genera and Eucalyptus subgenera. Letters above the bars represent significant differences among the groups at P < 0.001

170
Table 4 Summary statistics of foliar chemical attributes of different tree species and parasitic plants available to common brushtail possums within their home ranges. Letters represent significant differences among the groups at P < 0.001
Species Statistics Acacia dealbata
Acacia melanoxylon
Amyema pendula
Exocarpos cupressiformis
Eucalyptus dives
Eucalyptus obliqua
Eucalyptus radiata
Eucalyptus camphora
Eucalyptus globulus
Eucalyptus viminalis
No. samples
41 21 75 22 17 5 95 75 21 41
Total N Mean 2.60e 2.27d 1.44b 1.89c 1.20a 1.08a 1.43b 1.76c 1.11a 1.49b
(% DM) s.e. 0.04 0.04 0.03 0.05 0.02 0.03 0.01 0.02 0.02 0.02
CV (%) 9.39 8.46 20.21 11.29 8.41 5.86 8.40 10.88 8.39 8.30
Digestible N Mean 0.54c 0.52c 0.55c 0.59d 0.28a 0.31a 0.26a 0.52c 0.45b 0.56c
(% DM) s.e. 0.01 0.01 0.01 0.01 0.01 0.02 0.01 0.00 0.01 0.01
CV (%) 7.19 7.54 9.37 8.65 19.57 17.98 22.90 7.39 8.14 8.06
Available N Mean 1.35d 1.24d 0.80b 1.15d 0.43a 0.31a 0.39a 0.95c 0.45a 0.82b
(% DM) s.e. 0.03 0.03 0.03 0.04 0.03 0.03 0.01 0.02 0.02 0.02
CV (%) 13.99 11.89 28.15 15.53 24.39 23.56 28.16 14.54 15.99 13.55
Tannin-bound N Mean 0.44b 0.31a 0.22a 0.24a 0.45b 0.46b 0.63c 0.33a 0.22a 0.18a
(% DM) s.e. 0.01 0.01 0.01 0.02 0.01 0.02 0.01 0.01 0.02 0.02
CV (%) 16.19 20.30 36.68 42.05 13.26 9.20 9.46 33.90 34.32 55.51
Fibre-bound N Mean 0.82d 0.73c 0.41b 0.50b 0.33a 0.31a 0.42b 0.48b 0.44b 0.49b
(% DM) s.e. 0.01 0.02 0.01 0.02 0.01 0.02 0.01 0.01 0.02 0.02
CV (%) 7.98 13.72 17.35 14.66 9.55 16.75 13.95 21.43 17.00 19.72
DM Mean 50.10b 48.22a 44.59a 45.64a 53.24b 47.48a 54.55b 48.65a 55.13b 56.24b
(%) s.e. 1.12 1.34 0.60 1.10 0.54 1.32 0.62 0.60 1.39 0.88
CV (%) 14.34 12.77 11.61 11.28 4.21 6.23 11.06 10.76 11.59 10.03
DMD Mean 47.68b 43.21a 59.81e 45.87b 37.46a 40.30a 39.71a 51.46c 50.12c 53.20d
(%) s.e. 0.28 0.41 0.24 0.35 0.51 1.05 0.29 0.22 0.54 0.24
CV (%) 3.71 4.38 3.48 3.61 5.62 5.82 7.04 3.70 4.93 2.89
DBH Mean 17.22a 24.05a n. a. 16.09a 74.47d 62.60c 79.36 44.20b 41.57b 65.41c
(%) s.e. 1.52 3.40 n. a. 1.66 5.83 13.65 3.30 2.20 5.98 3.59
CV (%) 56.67 64.79 n. a. 48.29 32.29 48.75 40.47 44.15 65.87 35.18

171
Intra-specific variability in the foliar attributes
Chemical variability within species was expressed as the coefficient of variation, which
describes the variation as a proportion of the mean (Table 4). There was little within-
species variation observed for all studied tree species in terms of total nitrogen. Tree
species with the highest coefficient of variation of available nitrogen were from the genus
Monocalyptus exceeding 20% while Symphyomyrtus species were the most variable in terms of
tannins and fibre. From all species, Eucalyptus viminalis was the most variable in foliar tannin
levels (> 50%), followed by Exocarpos cupressiformis (42%) while Eucalyptus camphora was
most variable in terms of fibre (Table 4).
The most variable in terms of foliar concentrations of total and available nitrogen was
drooping mistletoe (Amyema pendula, Table 4). Drooping mistletoe’s total nitrogen (% DM)
was lower than the total nitrogen of its tree hosts, Acacia dealbata (t = 9.19, df = 21.79,
P < 0.001, Fig. 17) and Eucalyptus camphora (t = 5.13, df = 21.83, P < 0.001, Fig. 17), but
higher than E. dives (t = -2.36, df = 4.81, P = 0.0667). No significant differences in total
nitrogen (% DM) were found between mistletoes and their tree hosts, E. radiata or E.
viminalis (t = 1.66, df = 47.61, P = 0.1026; t = 2.067, df = 12.78, P = 0.06 respectively,
Fig. 17).
Foliar concentrations of total and available nitrogen (% DM) of Amyema pendula depended
on the species of host and were highest in mistletoes parasitizing Acacia dealbata (P < 0.001,
Fig. 18). No significant differences in concentrations of total and available nitrogen
(% DM) were detected in mistletoes growing on eucalypt species from the Monocalyptus and
Symphyomyrtus subgenera (% DM, Fig. 18). Mistletoe tannin-bound nitrogen was not
significantly different among different species of hosts while fibre-bound nitrogen was
highest in mistletoes parasitizing Amyema pendula. No significant relationship was found
between foliar concentrations of total nitrogen (% DM) between mistletoes and their tree
hosts (P < 0.05, Fig. 19).
Mistletoes parasitizing different species of tree hosts had significantly lower dry matter
content (% DM) and higher dry matter digestibility (% DMD) than their hosts (P < 0.001).
Mistletoe foliar dry matter content (% DM) varied depending on the species of tree host
and was highest in mistletoes parasitizing E. radiata (46.11 ± 0.95) and lowest in mistletoes
parasitizing E. dives (39.67 ± 1.65; P < 0.001).

172
Dry matter digestibility (% DMD) was highest in mistletoes parasitizing E. camphora (60.98
± 0.32) while lowest in mistletoes parasitizing E. dives (58.20 ± 0.50; P < 0.001).
The Welch’s t-test revealed no significant differences in foliar concentrations of total
nitrogen in among trees parasitized and free from Amyema pendula. Only the parasitized
trees of E. dives and Acacia dealbata had lower foliar concentrations of total nitrogen
(% DM) than unparasitized ones (t = - 2.49, df = 6.42, P < 0.05; t = - 2.62, df = 20.64,
P < 0.05 respectively). Also, parasitized trees Eucalyptus dives and Acacia dealbata had lower
foliar concentrations of available nitrogen (% DM) than unparasitized ones (t = - 3.34,
df = 11.99, P < 0.05; t = - 2.35, df = 20.38, P < 0.05 respectively). No significant
differences were observed among parasitized and unparasitized trees in terms of foliar dry
matter content (% DM) and dry matter digestibility (% DMD). Mistletoe parasitized E.
radiata and E. camphora had a significantly higher diameter than trees free from Amyema
pendula (t = 2.11, df = 59.14, P < 0.05; t = 3.09, df = 18.23, P < 0.001 respectively).
Fig. 17 Mean foliar total nitrogen content (% DM) with standard error (s.e.) of different tree hosts and drooping mistletoe (Amyema pendula). An asterix (*) indicates significant differences between the groups at P < 0.001
*

173
Fig. 18 Mean foliar concentrations of available nitrogen (black bar), tannin-bound nitrogen (grey), and fibre-bound nitrogen (white) with standard error (s.e.) of dropping mistletoe (Amyema pendula) parasitizing different species of tree hosts. The foliar concentrations of available nitrogen, tannin-bound nitrogen, and fibre-bound nitrogen sum to total nitrogen (% DM) concentration
Figure 19 Linear regression of total nitrogen (% DM) of different tree hosts against total nitrogen (% DM) of drooping mistletoe (Amyema pendula)

174
The binomial generalized linear model showed that a key parameter separating trees
parasitized by Amyema pendula from those free from mistletoes was tree diameter (r2 = 0.03,
P < 0.05). Trees larger in diameter were more often parasitized by Amyema pendula
(Table 5). The model explained 56% of mistletoe tree presences and 64% of absences.
Table 5 The binomial generalized linear model predicting tree occupancy by drooping mistletoe Amyema pendula (AIC = 205.45). An asterix indicates significance at P < 0 ‘***’, P < 0.001 ‘**’ and P < 0.05 ‘*’
Coefficients Estimate Std. Error z-value P-value Intercept 0.762 0.350 2.178 0.029 * Diameter -0.012 0.005 -2.452 0.014 * Binomial generalized linear model
df Dev. Resid. df Resid. Dev. P-value
Null 149 207.94 Diameter 1 6.4938 148 201.45 0.011*
Inter-specific differences of trees used by common brushtail possums
Among all trees used by common brushtail possums, rarely-used Acacia dealbata (see
Chapter 2 and 3) had the highest foliar concentrations of total and available nitrogen (%
DM) and the highest concentration of fibre-bound nitrogen (P < 0.001, Fig. 20). From
five Eucalyptus species present in the study area, E. camphora, the second most used tree
species by common brushtail possums, had the highest concentrations of total and
available nitrogen (% DM, P < 0.001, Fig. 20). Conversely, the most often used tree
species, Eucalyptus radiata, had the lowest foliar concentration of available nitrogen (% DM)
and the highest tannin-bound nitrogen concentration of all tree species (P < 0.001, Fig.
20). The rarely used species, Eucalyptus obliqua and Eucalyptus globulus, had the lowest
concentration of total nitrogen (% DM; P < 0.001, Fig. 20). Furthermore, Eucalyptus
viminalis had the highest concentration of fibre-bound nitrogen of all Eucalyptus species (%
DM; P < 0.001, Fig. 20) and the highest dry matter content (58. 36 ± 1.21%), but also the
highest dry matter digestibility of all studied species (52.84 ± 0.44%). Tree species with the
lowest dry matter content was E. camphora (47.60 ± 1.21%) while E. radiata had the lowest
dry matter digestibility of all studied species (40.27 ± 0.41%).

175
Fig. 20 Mean foliar concentrations of available nitrogen (black bar), tannin-bound nitrogen (grey), and fibre-bound nitrogen (white) with standard error (s.e.) of different tree species used by common brushtail possums. The foliar concentrations of available nitrogen, tannin-bound nitrogen, and fibre-bound nitrogen sum to total nitrogen (% DM) concentration

176
Intra-specific differences of trees used by common brushtail possums
Hardly any significant differences were observed among trees used by common brushtail
possums and the closest unused trees of the same species in terms of all measured chemical
attributes (P > 0.05, Fig. 21 and 22, Table 6). A significant difference in diameter was
revealed for Eucalyptus radiata trees (P < 0.05). However, across common brushtail possum
home ranges, few significant differences between trees of the same species were discerned;
for example, Eucalyptus viminalis and E. camphora varied in foliar concentrations of total and
available nitrogen, dry matter content, and dry matter digestibility (P < 0.05).
a)
b)
Fig. 21 Intra-specific differences in foliar chemistry of used and unused trees by common brushtail possums within their home ranges. Figures show (a) total nitrogen (% DM) and (b) available nitrogen (% DM) with standard error (s.e.) of different tree species

177
a)
b) Fig. 22 Intra-specific differences in foliar chemistry of used and unused trees by common brushtail possums within their home ranges. Figures show (a) dry matter digestibility (% DMD) and (b) dry matter content (%) with standard error (s.e.) of different tree species

178
Table 6 Summary statistics of foliar chemical attributes of different tree species used and unused by common brushtail possums within their home ranges. An asterix (*) indicates significant differences between the groups at P < 0.05
Species Stats Eucalyptus camphora
Eucalyptus viminalis
Eucalyptus globulus
Eucalyptus radiata
Eucalyptus obliqua
Acacia dealbata
Acacia melanoxylon
Exocarpos cupressiformis
No. samples 42 26 12 80 6 6 8 16
Total N (% DM) Mean ± s.e.
Used 1.76 ± 0.04 1.55 ± 0.04 1.14 ± 0.03 1.44 ± 0.02 1.05 ± 0.03 2.67 ± 0.14 2.19 ± 0.07 1.83 ± 0.07
Unused 1.83 ± 0.04 1.46 ± 0.04 1.06 ± 0.06 1.44 ± 0.02 1.10 ± 0.04 2.37 ± 0.02 2.33 ± 0.02 1.80 ± 0.08
Digest. N (% total N) Mean ± s.e.
Used 0.52 ± 0.01 0.57 ± 0.01 0.44 ± 0.01 0.27 ± 0.01 0.35 ± 0.01* 0.55 ± 0.03 0.51 ± 0.02 0.58 ± 0.02
Unused 0.53 ± 0.01 0.57 ± 0.01 0.44 ± 0.02 0.26 ± 0.01 0.26 ± 0.02 0.52 ± 0.01 0.53 ± 0.01 0.56 ± 0.02
Avail. N (% DM) Mean ± s.e.
Used 0.95 ± 0.03 0.87 ± 0.03 0.45 ± 0.02 0.41 ± 0.01 0.33 ± 0.05 1.39 ± 0.14 1.17 ± 0.07 1.10 ± 0.06
Unused 1.01 ± 0.03 0.80 ± 0.03 0.44 ± 0.04 0.39 ± 0.02 0.27 ± 0.03 1.20 ± 0.02 1.28 ± 0.01 1.09 ± 0.06
Tannins (% DM) Mean ± s.e.
Used 0.33 ± 0.03 0.17 ± 0.02 0.25 ± 0.03 0.62 ± 0.01 0.43 ± 0.02* 0.41 ± 0.01 0.32 ± 0.02 0.19 ± 0.01
Unused 0.35 ± 0.02 0.14 ± 0.02 0.17 ± 0.04 0.64 ± 0.01 0.50 ± 0.01 0.32 ± 0.04 0.32 ± 0.02 0.15 ± 0.01
Fibre (% DM) Mean ± s.e.
Used 0.48 ± 0.03 0.50 ± 0.03 0.43 ± 0.02 0.41 ± 0.01 0.29 ± 0.04 0.88 ± 0.02 0.70 ± 0.05 0.54 ± 0.02
Unused 0.46 ± 0.02 0.52 ± 0.02 0.46 ± 0.02 0.41 ± 0.01 0.33 ± 0.01 0.85 ± 0.03 0.72 ± 0.03 0.57 ± 0.02
DM (%) Mean ± s.e.
Used 47.60 ± 1.21 58.36 ± 1.21 54.73 ± 2.36 56.00 ± 1.09 48.38 ± 1.28 56.27 ± 6.26 51.04 ± 1.94 48.46 ± 2.73
Unused 46.66 ± 0.75 55.66 ± 1.52 58.05 ± 3.48 54.85 ± 0.91 45.11 ± 2.07 47.33 ± 4.43 52.90 ± 4.86 44.50 ± 0.49
DMD (%) Mean ± s.e.
Used 51.11 ± 0.36 52.84 ± 0.44 50.65 ± 1.18 40.27 ± 0.41* 41.50 ± 1.21 47.09 ± 0.94 43.29 ± 0.70 45.85 ± 0.77
Unused 50.71 ±0.30 53.18 ± 0.48 49.07 ± 1.19 39.84 ± 0.45 38.13 ± 0.74 47.02 ± 0.54 44.28 ± 0.70 45.66 ± 0.18
DBH (cm) Mean ± s.e.
Used 46.76 ±3.67 68.69 ± 6.05 60.00 ± 15.55 91.30 ± 6.13 58.67 ± 21.15 18.33 ± 5.84 29.00 ± 4.93 15.75 ± 3.76
Unused 45.05 ± 4.34 64.31 ± 5.57 38.50 ± 7.23 67.41 ± 3.46 75.67 ± 14.33 16.00 ± 1.00 26.75 ± 4.26 17.25 ± 1.89

179
Determinants of tree use by common brushtail possums
To discriminate which of the measured tree attributes best explained overall tree use by
common brushtail possums the binomial generalized linear model was fitted to all tree
species and each species individually. The final model indicated that tree diameter (P <
0.05) and foliar concentration of available nitrogen (P = 0.09) were the best variables for
predicting the presence of common brushtail possums across all analysed trees (Table 7).
The model explained 60.2% of common brushtail possum tree presences and 61.2% of
absences. For the most visited species, Eucalyptus radiata, the model predicted that diameter
(P < 0.001) was the best variable for predicting 60% of common brushtail possum
presences and 72.5% of absences while for E. camphora, foliar concentration of available
nitrogen (P = 0.14) explained 66.7% of common brushtail possum tree presences and
71.4% of absences.
Table 7 The binomial generalized linear model predicting tree use by common brushtail possums (AIC = 269.08). An asterix indicates significance at P < 0 ‘***’, P < 0.001 ‘**’ and P < 0.05 ’*’ Coefficients Estimate Std. Error z-value P-value Intercept -1.501 0.613 -2.448 0.014* Diameter 0.015 0.006 2.791 0.005* Available N 0.852 0.510 1.668 0.095 Binomial generalized linear model
df Dev. Resid. df Resid. Dev. P-value
Null 195 271.71 Diameter 1 5.798 194 265.92 0.016 * Available N 1 2.837 193 263.08 0.092
Relationships between tree attributes and common brushtail possum home range size
Finally, when comparing the nutritional quality (mean foliar concentration of available
nitrogen) of individual possum home ranges, significant differences (P < 0.001) were
observed between individual animals (female 1 and 5, female 4 and female 5, female 4 and
male 3, and female 1 and male 3). However, the Spearman’s rank-order correlation showed
that none of the measured foliar attributes influenced possum home range size (P > 0.05).

180
Discussion
Summary of key results
In the present study, the phytochemical environment of common brushtail possums
included species of different nutritional quality. The nitrogen-fixing Acacia dealbata had the
highest concentrations of foliar total and available nitrogen, as well as the highest
concentration of fibre-bound nitrogen, of all species. The co-occurring root parasite,
Exocarpos cupressiformis, and eucalypts from subgenus Symphyomyrtus had relatively high
concentrations of available nitrogen with Eucalyptus viminalis found to have the lowest
concentration of tannin-bound nitrogen of all species. The aerial parasite, Amyema pendula,
had moderate concentrations of available nitrogen, which varied among individuals
depending on a species of host, with the most foliar nitrogen found in mistletoes
parasitizing Acacia dealbata trees. Mistletoes also had the lowest dry matter content and
highest dry matter digestibility. Eucalypts from the subgenus Monocalyptus were found to be
of the poorest nutritional quality for common brushtail possums, due to their low
concentrations of available nitrogen, high concentrations of tannin-bound nitrogen, and
low dry matter digestibility.
Contrary to expectation, common brushtail possums were found most often foraging in
Eucalyptus radiata (Monocalyptus), which contains the lowest foliar concentration of available
nitrogen and highest concentration of tannin-bound nitrogen. The common brushtail
possums also foraged in E. camphora and E. viminalis (Symphyomyrtus), which contain the
highest available nitrogen concentrations of all Eucalyptus species. However, possums were
found only occasionally on Acacia dealbata and A. melanoxylon that had the highest available
nitrogen concentrations of all studied species. Despite the fact that parasitic plants were of
high nutritional quality for common brushtail possums, the results of this study were
inconclusive. These inconclusive results could be attributed to a small sample size of
Exocarpos cupressiformis and a lack of certainty about tree use in the case of trees parasitized
by mistletoes. Surprisingly, there were few significant intra-specific differences stated
between trees used by common brushtail possums and the closest unused ones. In this
study, none of the measured foliar chemical attributes influenced a home range size while
tree diameter and available nitrogen explained 60.2% of common brushtail possum tree
presences and 61.2% of absences.

181
The nutritional value of different tree species and parasitic plants available to common brushtail possums
To understand the nature of chemically complex “foodscapes” and their ecological effects
on common brushtail possums, it is important to ascertain the nutritional quality of forage
available to herbivores within their home ranges and to describe any between- and within-
species variability affecting animal foraging choices. In this study, the chemical analysis of
the nutritional quality of forage for common brushtail possums revealed that, despite being
subjected to similar environmental conditions, co-occurring genera and species exhibited
broad differences in foliar concentrations of total and available nitrogen, reflecting their
diverse life-history strategies (modes of nitrogen acquisition and partitioning), plant
genotype differences, and environmental variation affecting chemotypes.
For instance, acacias are known to reach the highest concentrations of foliar total nitrogen
through the symbiotic relations with root prokaryotic bacteria (Burdon et al., 1999). Yet,
acacias are considered to be of low nutritional quality for herbivores due to presumed high
concentrations of tannins (Dynes & Schlink, 2002). Nevertheless, in the present study
Acacia species had similar concentrations of tannin-bound nitrogen (c.a. 20%) as parasitic
plants and eucalypts from the subgenus Symphyomyrtus. It is important to stress here that
crude measurements of polyethylene glycol (PEG) tannin binding capacity in this study
ignore the structural complexity of different groups of tannins and their diverse effects on
herbivores.
In a more detailed study, Cork and Pahl (1984) investigated nutritional quality, including
total nitrogen, total tannins, and condensed tannins, of Acacia melanoxylon and different
Monocalyptus and Symphyomyrtus species for common ringtail possums. This study showed
that A. melanoxylon had the highest total nitrogen concentration, but similar concentration
of total tannins as Monocalyptus species, and a lower concentration of condensed tannins
than both Monocalyptus and Symphyomyrtus species. On the other side, the rarity of Australian
herbivores found feeding on Acacia foliage might be attributed to the presence of plant
secondary metabolites, such as alkaloids, cyanogenic glycosides, flavonoids and small
amounts of terpenoids (Dynes & Schlink, 2002). By contrast, Eucalyptus species that
represent a large portion of the diet of arboreal marsupials are heavily defended by several
groups of plant secondary metabolites, including terpenes, cyanogenic glycosides, and
formylated phloroglucinol compounds (FPCs; Moore et al., 2004b).

182
Between species differences in foliar concentrations of plant secondary metabolites further
complicate the situation and require a holistic metabolomic approach to identify chemical
traits that consistently vary between Acacia and Eucalyptus subgenera as suggested by Tucker
and colleagues (2010).
In the present study, foliar total nitrogen concentrations varied among Eucalyptus subgenera
from moderate to low, with Symphyomyrtus species having slightly higher concentrations of
total nitrogen than Monocalyptus species, with a single Symphyomyrtus species Eucalyptus
camphora having the highest concentration of total nitrogen of all eucalypts. Conversely,
distinct differences were observed in the foliar concentrations of available nitrogen with
Symphyomyrtus species containing more than half the available nitrogen of that of subgenus
Monocalyptus. Wallis, Nicolle and Foley (2010) found that 31 species of Monocalyptus
contained slightly less total nitrogen than 82 Symphyomyrtus species (1.00 vs. 1.12% DM) and
less than half of the available nitrogen (0.27 vs. 0.59% DM).
Ecological differences between subgenera Monocalyptus and Symphyomyrtus have been known
since the 1980s, with the tannin-rich Monocalyptus subgenera recognised to carry a lower
diversity of herbivores and suffer less leaf loss and damage than Symphyomyrtus (Noble,
1989). For instance, Youngentob et al. (2010) demonstrated that for a eucalypt specialist,
the greater glider (Petauroides volans), lower concentrations of available nitrogen in E. radiata
compared to E. viminalis explained the tree use of Monocalyptus species. By contrast, the
concentration of formylated phloroglucinol compounds, syderoxylonals, was more
important than available nitrogen in the case Symphyomyrtus species.
In the present study, the parasitic plants, Amyema pendula and Exocarpos cupressiformis, were
expected to have moderate to slightly higher foliar concentrations of total nitrogen than
their tree hosts due to mineral absorption and accumulation, hence they would experience
higher levels of herbivory (Bannister, 1989; Canyon & Hill, 1997). However, this study,
contrary to other studies (Bannister, Strong, & Andrew, 2002; March & Watson, 2010),
showed that A. pendula contained lower concentrations of total nitrogen than most of its
tree hosts. Additionally, mistletoe foliar concentrations of total nitrogen varied across
different species of hosts, being highest in mistletoes parasitizing acacias with hardly any
differences observed in those parasitizing eucalypt species. By contrast, the root parasite
Exocarpos cupressiformis had significantly higher concentration of total nitrogen than A.
pendula possibly due to its tendency to selectively parasitize nitrogen-fixing hosts or acquire
nitrogen independently from the soil (Sinclair, 2006).

183
Furthermore, this study has shown that foliar concentrations of tannin-bound nitrogen of
parasitic plants were similar to those observed for both Acacia and Eucalyptus host species,
suggesting that some other factors may affect the dietary choices of herbivores. Mistletoes
are known to accumulate large amounts of potassium and phosphorus, macronutrients
important in herbivore diets, but at the same time sequester plant secondary metabolites
from their tree hosts, such as alkaloids and cyanogenic glycosides (Martin Cordero et al.,
1993; Schulze, Turner, & Glatzel, 1984). Conversely, the secondary chemistry of the root
parasite, Exocarpos cupressiformis, is poorly researched. Only recently, the presence of
exocarpic acid was discovered in a tropical rainforest species, Exocarpos latifolius. This acid is
known to exhibit antibacterial properties but has unknown effects on herbivores (Koch et
al., 2009).
Differences in dry matter digestibility (% DMD) revealed one more level of chemical
variability between studied species and genera, with Amyema pendula being the most
digestible, and fibrous species from Monocalyptus being the least. Higher dry matter
digestibility observed in Symphyomyrtus than Monocalyptus species was also confirmed by
other studies (Wallis et al., 2010; Youngentob et al., 2011). In general, high fibre content
was suggested as the main cause of the low digestibility of Eucalyptus species as microscopic
observations of plant fragments in the caecum and faeces of common brushtail possums
revealed few bacteria attached to fibrous tissues (Foley & Hume, 1986). Furthermore,
Acacia species were found to be moderately digestible with phyllodinous Acacia melanoxylon
being higher in fibre and less digestible than the bipinnate leaves of Acacia dealbata.
Common brushtail possums are thought to select Acacia leaves based on their nutrient and
fibre content since they have a relatively small gut capacity (Crowe & Hume, 1997). Thus,
enzymatic digestion in the small intestine is relatively more important, and microbial
fermentation in the hindgut is less so, hence the possum’s strong selection against dietary
fibre.
In conclusion, there is no currently accepted uniform cross-taxa definition of the
nutritional quality of forage for herbivores, making it hard to generalise which genera and
species are more suitable and readily consumed by herbivores. The measured effects of
tannins and fibre on nitrogen availability can explain nutritional differences between
Eucalyptus genera. However, total nitrogen is a better nutritional measure for Acacia and
parasitic plant species.

184
Similarly, if we identify genera and species with high foliar concentrations of total and
available nitrogen that herbivores avoid eating, we might wish to explore other reasons for
this behaviour, such as the presence and high concentrations of different plant secondary
metabolites. Hence, the next section will focus on animal foraging choices in a chemically
complex environment, hopefully overcoming our inherently biased perception of what
represents “good food” for herbivores.
Common brushtail possum foraging choices in a chemically complex environment
Unlike the findings of other similar studies, the present study demonstrated that common
brushtail possums used trees from both Eucalyptus subgenera, Monocalyptus and
Symphyomyrtus (see Chapters 2 and 3 for details). A currently accepted view is that common
brushtail possums rarely feed on Monocalyptus species in order to avoid the high costs of
tannins on their nitrogen metabolism and instead prefer Symphyomyrtus species in mixed
eucalypt stands (Marsh et al., 2003; Moore et al., 2004). However, in this study the most
often used eucalypts, Eucalyptus radiata (Monocalyptus) and E. camphora (Symphyomyrtus), were
diametrically different in their nutritional quality. Eucalyptus radiata had the highest foliar
concentration of tannin-bound nitrogen and consequently the lowest available nitrogen
from all used species while Eucalyptus camphora had the highest total and available nitrogen
from all eucalypts.
The fact that common brushtail possums were most often found foraging on tannin-rich
Eucalyptus radiata foliage was unexpected as possums are believed to be sensitive to tannins
due to their distinctively astringent taste and nauseating effect that cause animals to cease
feeding (Marsh et al., 2003). To counteract the negative effects of tannins, animals secrete
proline-rich salivary proteins that bind tannins; yet, McArthur and colleagues (1995) noted
low rates of protein secretion in common brushtail possums. Other detoxification
strategies can be employed by animals, such as associations with gut microbes that
dissociate tannin-protein complex or secrete tannase (DeGabriel et al., 2009a, 2009b).
However, in this study the Symphyomyrtus species, despite having low tannin concentrations
were found to be eaten less often by possums, likely due to presence of other potent
herbivore deterrents, formylated phloroglucinol compounds (FPCs, Eschler et al., 2000).

185
Formylated phloroglucinol compounds are known to damage enterochromaffin cells in the
gut of herbivores, releasing serotonin that is subsequently detected by the emetic system
producing the sensation of nausea (Lawler et al., 1998). Research with captive koalas,
common brushtail possums, and ringtail possums has shown that herbivores regulated their
intake of formylated phloroglucinol compounds when they were isolated from eucalypts
and added to artificial diets (Lawler et al., 1998; Wallis, Watson, & Foley, 2002). Scrivener
and colleagues (2004) demonstrated that wild common brushtail possums largely avoided
trees with high concentrations of formylated phloroglucinol compounds while their use of
trees with low concentrations of these compounds was highly variable suggesting the
importance of tannins and other plant secondary metabolites. Recently a new group of
chemicals has been discovered in Monocalyptus, free flavanones with no substitution in Ring
B; yet, their effects on herbivores remain to be verified (Tucker et al., 2010).
Regardless of the nature of the chemical differences between Eucalyptus subgenera, this
study has demonstrated that common brushtail possums have a much broader diet than
previously assumed and consume both Symphyomyrtus and Monocalyptus species depending
on their availability (see Chapters 2 and 3 for details). Similar patterns were observed in
the study of foraging choices of co-occurring greater gliders in a habitat dominated by
Eucalyptus radiata (Monocalyptus) and E. viminalis (Symphyomyrtus) by Youngentob et al. (2011).
The authors explained this phenomenon as the result of animals using different metabolic
detoxification pathways for different groups of plant secondary metabolites. For instance,
gliders will eat foliage from Symphyomyrtus species until the physiological effects of
formylated phloroglucinol compounds force them to stop; then they switch to Monocalyptus
while detoxification processes remove formylated phloroglucinol compounds. Next, they
would revert to eating Symphyomyrtus foliage before ingesting debilitating amounts of
tannins from Monocalyptus species.
Furthermore, intra-specific differences in the concentrations of nutrients and chemical
defences have been recognised for a long time as drivers of dietary choices of arboreal
marsupials (Lawler, Foley, & Eschler, 2000; Lawler et al., 1998). Genetically and
environmentally driven intra-specific chemical differences in foliar concentrations of
available nitrogen, tannins, and formylated phloroglucinol compounds among Eucalyptus
species have been postulated to influence tree selectivity and foliage intake of common
brushtail possums and koalas (Andrew et al., 2010; O'Reilly-Wapstra, McArthur, & Potts,
2002; O'Reilly-Wapstra et al., 2005).

186
However, in this study no significant differences were observed between used trees and the
closest unused trees of the same species in terms of all measured tree variables, apart from
a few exceptions. The lack of difference between the trees may be explained by physical
barriers in pollen dispersal or by habitat uniformity or autocorrelation processes causing
genetically similar trees to occur in close proximity to each other (Andrew et al., 2007). It is
important to stress here that a low sample size could have affected the results, creating a
large number of false negatives or trees used by possums that were not recorded in this
study. Nevertheless, across possum home ranges, few significant intra-specific differences
were discerned. This suggests that intra-specific differences might occur on a broader scale
across landscapes and regions, exerting adverse effects on different herbivore populations.
Recently, the Geometric Framework for nutrition has been proposed by Raubenheimer,
Simpson and Mayntz (2009) that enables a better understanding of animal nutritional
strategies and interpretation of food choices that are otherwise difficult to explain. The
focus of this framework is on observing free food choices of individual animals when faced
with diverse dietary options and relating these choices to several nutritionally relevant
measures within simple geometrical models. The nutritional measures can include the
following: the optimal balance and amount of nutrients required to be ingested and
allocated to growth over a given time period (the intake and growth targets), the animal’s
current state in relation to these requirements, available foods and the consequences for the
animal upon ingesting them, and the amount of nutrients and plant secondary metabolites
that are retained and detoxified by the animal. Future studies on nutritional requirements of
arboreal marsupials should consider incorporating the Geometric Framework into their
research design.

187
References
Agrawal, A. A. (2001). Phenotypic plasticity in the interactions and evolution of species. Science, 294(5541), 321–326.
Agrawal, A. A. (2011). Current trends in the evolutionary ecology of plant defence. Functional Ecology, 25(2), 420–432.
Agrawal, A. A., & Fishbein, M. (2006). Plant defense syndromes. Ecology, 87(7), 132–149. Andrew, R. L., Peakall, R., Wallis, I. R., & Foley, W. J. (2007). Spatial distribution of
defense chemicals and markers and the maintenance of chemical variation. Ecology, 88(3), 716–728.
Andrew, R. L., Wallis, I. R., Harwood, C. E., & Foley, W. J. (2010). Genetic and environmental contributions to variation and population divergence in a broad-spectrum foliar defence of Eucalyptus tricarpa. Annals of Botany, 105(5), 707–717.
Bannister, P. (1989). Nitrogen concentration and mimicry in some New Zealand mistletoes. Oecologia, 79(1), 128–132.
Bannister, P., Strong, G. L., & Andrew, I. (2002). Differential accumulation of nutrient elements in some New Zealand mistletoes and their hosts. Functional Plant Biology, 29(11), 1309–1318.
Barbehenn, R. V., & Peter Constabel, C. (2011). Tannins in plant-herbivore interactions. Phytochemistry, 72(13), 1551–1565.
Batten, G. D. (1998). Plant analysis using near infrared reflectance spectroscopy: The potential and the limitations. Australian Journal of Experimental Agriculture, 38(7), 697–706.
Bell, T. L., & Adams, M. A. (2011). Attack on all fronts: Functional relationships between aerial and root parasitic plants and their woody hosts and consequences for ecosystems. Tree Physiology, 31(1), 3–15.
Bennett, R. N., & Wallsgrove, R. M. (1994). Tansley Review No. 72. Secondary metabolites in plant defence mechanisms. New Phytologist, 127(4), 617–633.
Berenbaum, M. (1980). Adaptive significance of midgut pH in larval Lepidoptera. The American Naturalist, 115(1), 138–146.
Bernays, E. A. (1981). Plant tannins and insect herbivores: An appraisal. Ecological Entomology, 6(4), 353–360.
Boland, D. J., Brophy, J. J., & House, A. P. N. (1991). Eucalyptus leaf oils: Use, chemistry, distillation and marketing. Melbourne: Inkata Press.
Braithwaite, L. W., Turner, J., & Kelly, J. (1984). Studies on the Arboreal Marsupial Fauna of Eucalypt Forests Being Harvested for Wood Pulp at Eden, N.s.w. Iii. Relationships Between Faunal Densities, Eucalypt Occurrence and Foliage Nutrients, and Soil Parent Materials. Australian Wildlife Research, 11(1), 41–48.
Burchfield, E., Agar, N. S., & Hume, I. D. (2006). Effects of terpenes and tannins on some physiological and biochemical parameters in two species of phalangerid possums (Marsupialia: Phalangeridae). Australian Journal of Zoology, 53(6), 395–402.
Burdon, J. J., Gibons, A. H., Searle, S. D., Woods, M. J., & Brockwell, J. (1999). Variation in the effectiveness of symbiotic associations between native rhizobia and temperate Australian Acacia: Within-species interactions. Journal of Applied Ecology, 36(3), 398–408.
Canyon, D. V., & Hill, C. J. (1997). Mistletoe host-resemblance: A study of herbivory, nitrogen and moisture in two Australian mistletoes and their host trees. Australian Journal of Ecology, 22(4), 395–403.
Carmona, D., Lajeunesse, M. J., & Johnson, M. T. J. (2011). Plant traits that predict resistance to herbivores. Functional Ecology, 25(2), 358–367.

188
Choo, G. M., Waterman, P. G., McKey, D. B., & Gartlan, J. S. (1981). A simple enzyme assay for dry matter digestibility and its value in studying food selection by generalist herbivores. Oecologia, 49(2), 170–178.
Clapperton, B. K., Matthews, L. R., Fawkes, M. S., & Pearson, A. J. (1996). Lithium and cyanide-induced conditioned food aversions in brushtail possums. The Journal of Wildlife Management, 60(1), 195–201.
Clausen, T. P., Provenza, F. D., Burritt, E. A., Reichardt, P. B., & Bryant, J. P. (1990). Ecological implications of condensed tannin structure: A case study. Journal of Chemical Ecology, 16(8), 2381–2392.
Close, D. C., Davidson, N. J., & Davies, N. W. (2006). Seasonal fluctuations in pigment chemistry of co-occurring plant hemi-parasites of distinct form and function. Environmental and Experimental Botany, 58(1-3), 41–46.
Cockfield, S. D. (1988). Relative availability of nitrogen in host plants of invertebrate herbivores: Three possible nutritional and physiological definitions. Oecologia, 77(1), 91–94.
Coley, P. D., & Barone, J. A. (1996). Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systematics, 27, 305–335.
Cooper, S. M., Owen-Smith, N., & Bryant, J. P. (1988). Foliage acceptability to browsing ruminants in relation to seasonal changes in the leaf chemistry of woody plants in a South African savanna. Oecologia, 75(3), 336–342.
Crowe, O., & Hume, I. D. (1997). Morphology and function of the gastrointestinal tract of Australian folivorous possums. Australian Journal of Zoology, 45(4), 357–368.
Cyr, H., & Face, M. L. (1993). Magnitude and patterns of herbivory in aquatic and terrestrial ecosystems. Nature, 361(6408), 148–150.
Dearing, M. D., Foley, W. J., & McLean, S. (2005). The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates. Annual Review of Ecology, Evolution, and Systematics, 36, 169–189.
DeGabriel, J. L. (2008). Demographic constraints imposed by plant nutrients and secondary chemistry on the common brushtail possum (Trichosurus vulpecula Kerr.). (Ph. D. thesis), The Australian National University, Canberra.
DeGabriel, J., Wallis, I., Moore, B., & Foley, W. (2008). A simple, integrative assay to quantify nutritional quality of browses for herbivores. Oecologia, 156(1), 107–116.
DeGabriel, J. L., Moore, B. D., Foley, W. J., & Johnson, C. N. (2009a). The effects of plant defensive chemistry on nutrient availability predict reproductive success in a mammal. Ecology, 90(3), 711–719.
DeGabriel, J., Moore, B. D., Shipley, L., Krockenberger, A. K., Wallis, I. R., Johnson, C. N., & Foley, W. J. (2009b). Inter-population differences in the tolerance of a marsupial folivore to plant secondary metabolites. Oecologia, 161(3), 539–548.
DeGabriel, J., Moore, B. D., Marsh, K. J., & Foley, W. J. (2010). The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology, 20(2), 97–108.
Di Stefano, J., & Newell, G. R. (2008). Diet selection by the swamp wallaby (Wallabia bicolor): Feeding strategies under conditions of changed food availability. Journal of Mammalogy, 89(6), 1540–1549.
Dynes, R. A., & Schlink, A. C. (2002). Livestock potential of Australian species of Acacia. Conservation Science Western Australia, 4(3), 117–124.
Ebbers, M. J. H., Wallis, I. R., Dury, S., Floyd, R., & Foley, W. J. (2002). Spectrometric prediction of secondary metabolites and nitrogen in fresh Eucalyptus foliage: Towards remote sensing of the nutritional quality of foliage for leaf-eating marsupials. Australian Journal of Botany, 50(6), 761–768.
Eschler, B. M., Pass, D. M., Willis, R., & Foley, W. J. (2000). Distribution of foliar formylated phloroglucinol derivatives amongst Eucalyptus species. Biochemical Systematics and Ecology, 28(9), 813–824.

189
Felton, A. M., Felton, A., Lindenmayer, D. B., & Foley, W. J. (2009a). Nutritional goals of wild primates. Functional Ecology, 23(1), 70–78.
Felton, A. M., Felton, A., Raubenheimer, D., Simpson, S. J., Foley, W. J., Wood, J. T., ... & Lindenmayer, D. B. (2009b). Protein content of diets dictates the daily energy intake of a free-ranging primate. Behavioral Ecology, 20(4), 685–690.
Foley, W. J., & Hume, I. D. (1987). Digestion and metabolism of high-tannin Eucalyptus foliage by the brushtail possum (Trichosurus vulpecula)(Marsupialia: Phalangeridae). Journal of Comparative Physiology B, 157(1), 67–76.
Foley, W. J., McIlwee, A., Lawler, I. R., Aragones, L., Woolnough, A. P., & Berding, N. (1998). Ecological applications of near infrared reflectance spectroscopy—a tool for rapid, cost-effective prediction of the composition of plant and animal tissues and aspects of animal performance. Oecologia, 116(3), 293–305.
Forbey, J. S., Dearing, M. D., Gross, E. M., Orians, C. M., Sotka, E. E., & Foley, W. J. (2013). A pharm-ecological perspective of terrestrial and aquatic plant–herbivore interactions. Journal of Chemical Ecology, 39(4), 465–480.
Fox, L. R., & Macauley, B. J. (1977). Insect grazing on Eucalyptus in response to variation in leaf tannins and nitrogen. Oecologia, 29(2), 145–162.
Freeland, W. J., & Janzen, D. H. (1974). Strategies in herbivory by mammals: The role of plant secondary compounds. The American Naturalist, 108(961), 269–287.
Glatzel, G., & Geils, B. W. (2009). Mistletoe ecophysiology: Host–parasite interactions. Botany, 87(1), 10–15.
Gleadow, R. M., Haburjak, J., Dunn, J. E., Conn, M. E., & Conn, E. E. (2008). Frequency and distribution of cyanogenic glycosides in Eucalyptus L'Hérit. Phytochemistry, 69(9), 1870–1874.
Hansen, A. P., Pate, J. S., & Atkins, C. A. (1987). Relationships between acetylene reduction activity, hydrogen evolution and nitrogen fixation in nodules of Acacia spp.: Experimental background to assaying fixation by acetylene reduction under field conditions. Journal of Experimental Botany, 38(1), 1–12.
Hensens, O. D., Lewis, K. G., & Mulquiney, C. E. (1971). Some extractives from mistletoe. Australian Journal of Chemistry, 24(2), 431–434.
Iason, G., & Villalba, J. (2006). Behavioral strategies of mammal herbivores against plant secondary metabolites: The avoidance-tolerance continuum. Journal of Chemical Ecology, 32(6), 1115–1132.
Irlbeck, N. A., & Hume, I. D. (2003). The role of Acacia in the diets of Australian marsupials ? A review. Australian Mammalogy, 25(2), 121–134.
Janzen, D. H., (1977). Promising Directions of Study in Tropical Animal–Plant Interactions. Annals of the Missouri Botanical Garden, 64(4), 706-736.
Joern, A., & Behmer, S. T. (1997). Importance of dietary nitrogen and carbohydrates to survival, growth, and reproduction in adults of the grasshopper Ageneotettix deorum (Orthoptera: Acrididae). Oecologia, 112(2), 201–208.
Kavanagh, R. P., & Lambert, M. J. (1990). Food selection by the greater glider, Petauroides volans: Is foliar nitrogen a determinant of habitat quality? Wildlife Research, 17(3), 285–299.
Keith, H., Barrett, D., & Keenan, R. (2000). Review of allometric relationships for estimating woody biomass for New South Wales, the Australian Capital Territory, Victoria, Tasmania, and South Australia National Carbon Accounting System Technical Report 5B. Canberra: Australian Greenhouse Office.
Koch, M., Bugni, T. S., Pond, C. D., Sondossi, M., Dindi, M., Piskaut, P., Barrows, L. R. (2009). Antimycobacterial activity of Exocarpos latifolius is due to exocarpic acid. Planta Med, 75(12), 1326–1330.
Lawler, I. R., Foley, W. J., & Eschler, B. M. (2000). Foliar concentration of a single toxin creates habitat patchiness for a marsupial folivore. Ecology, 81(5), 1327-1338.

190
Lawler, I. R., Foley, W. J., Eschler, B. M., Pass, D. M., & Handasyde, K. (1998). Intraspecific variation in Eucalyptus secondary metabolites determines food intake by folivorous marsupials. Oecologia, 116(1), 160–169.
Lawler, I. R., Foley, W. J., Pass, G. J., & Eschler, B. M. (1998). Administration of a 5HT3 receptor antagonist increases the intake of diets containing Eucalyptus secondary metabolites by marsupials. Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 168(8), 611–618.
Lawler, I. R., Stapley, J., Foley, W. J., & Eschler, B. M. (1999). Ecological example of conditioned flavor aversion in plant-herbivore interactions: Effect of terpenes of Eucalyptus leaves on feeding by common ringtail and brushtail possums. Journal of Chemical Ecology, 25(2), 401–415.
Loney, P. E., McArthur, C., Potts, B. M., & Jordan, G. J. (2006). How does ontogeny in a Eucalyptus species affect patterns of herbivory by Brushtail Possums? Functional Ecology, 20(6), 982–988.
Lucas, P. W., Turner, I. M., Dominy, N. J., & Yamashita, N. (2000). Mechanical defences to herbivory. Annals of Botany, 86(5), 913–920.
March, W. A., & Watson, D. M. (2010). The contribution of mistletoes to nutrient returns: Evidence for a critical role in nutrient cycling. Austral Ecology, 35(7), 713–721.
Marsh, K. J., Foley, W. J., Cowling, A., & Wallis, I. R. (2003). Differential susceptibility to Eucalyptus secondary compounds explains feeding by the common ringtail (Pseudocheirus peregrinus) and common brushtail possum (Trichosurus vulpecula). Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 173(1), 69–78.
Marsh, K. J., Wallis, I. R., & Foley, W. J. (2003). The effect of inactivating tannins on the intake of Eucalyptus foliage by a specialist Eucalyptus folivore (Pseudocheirus peregrinus) and a generalist herbivore (Trichosurus vulpecula). Australian Journal of Zoology, 51(1), 31–42.
Marsh, K. J., Wallis, I. R., Andrew, R. L., & Foley, W. J. (2006a). The detoxification limitation hypothesis: Where did it come from and where is it going? Journal of Chemical Ecology, 32(6), 1247–1266.
Marsh, K. J., Wallis, I. R., McLean, S., Sorensen, J. S., & Foley, W. J. (2006b). Conflicting demands on detoxification pathways influence how common brushtail possums choose their diets. Ecology, 87(8), 2103–2112.
Martin Cordero, C., Gil Serrano, A. M., & Ayuso Gonzalez, M. J. (1993). Transfer of bipiperidyl and quinolizidine alkaloids to Viscum cruciatum Sieber (Loranthaceae) hemiparasitic on Retama sphaerocarpa boissier (Leguminosae). Journal of Chemical Ecology, 19(10), 2389–2393.
Martin, R. W., & Handasyde, K. A. (1999). The Koala: Natural history, Conservation and Management. Sydney: University of New South Wales.
Mattson, W. J. (1980). Herbivory in relation to plant nitrogen content. Annual Review of Ecology and Systematics, 11(1), 119–161.
McArt, S. H., Spalinger, D. E., Collins, W. B., Schoen, E. R., Stevenson, T., & Bucho, M. (2009). Summer dietary nitrogen availability as a potential bottom-up constraint on moose in south-central Alaska. Ecology, 90(5), 1400–1411.
McArthur, C., Hagerman, A., & Robbins, C. T. (1991). Physiological strategies of mammalian herbivores against plant defenses. In R. T. Palo & C. T. Robbins (Eds.), Plant Defenses against Mammalian Herbivores (pp. 103–114). Boca Raton: CRC Press.
McArthur, C., Sanson, G. D., & Beal, A. M. (1995). Salivary proline-rich proteins in mammals: Roles in oral homeostasis and counteracting dietary tannin. Journal of Chemical Ecology, 21(6), 663–691.
McLean, S., & Foley, W. J. (1997). Metabolism of Eucalyptus terpenes by herbivorous marsupials. Drug Metabolism Reviews, 29(1–2), 213–218.

191
McLean, S., Brandon, S., Boyle, R. R., & Wiggins, N. L. (2008). Development of tolerance to the dietary plant secondary metabolite 1, 8-cineole by the brushtail possum (Trichosurus vulpecula). Journal of Chemical Ecology, 34(5), 672–680.
McLeod, M. N. (1973). The digestibility and the nitrogen, phosphorus and ash contents of the leaves of some Australian trees and shrubs. Australian Journal of Experimental Agriculture, 13(62), 245–250.
McNaughton, S. J. (1988). Mineral nutrition and spatial concentrations of African ungulates. Nature, 334, 343–345.
Mole, S., & Waterman, P. G. (1987). Tannins as Antifeedants to Mammalian Herbivores—Still an Open Question? In G. R. Waller (Ed.), Allelochemicals: Role in Agriculture and Forestry (Vol. 330, pp. 572–587): American Chemical Society.
Molyneux, R. J., & Ralphs, M. H. (1992). Plant toxins and palatability to herbivores. Journal of Range Management, 45(1), 13–18.
Moore, B. D., Wallis, I. R., Marsh, K. J., & Foley, W. J. (2004a). The role of nutrition in the conservation of the marsupial folivores of eucalypt forests. In D. Lunney (Ed.), Conservation of Australia’s Forest Fauna (2 ed., pp. 549–575). Mosman: Royal Zoological Society of New South Wales.
Moore, B. D., Wallis, I. R., Palá-Paúl, J., Brophy, J. J., Willis, R. H., & Foley, W. J. (2004b). Antiherbivore chemistry of Eucalyptus—cues and deterrents for marsupial folivores. Journal of Chemical Ecology, 30(9), 1743–1769.
Moore, B. D., & Foley, W. J. (2005). Tree use by koalas in a chemically complex landscape. Nature, 435(7041), 488–490.
Mueller-Harvey, I. (2006). Unravelling the conundrum of tannins in animal nutrition and health. Journal of the Science of Food and Agriculture, 86(13), 2010–2037.
Nelder, J. A., & Wedderburn, R. W. M. (1972). Generalized linear models. Journal of the Royal Statistical Society. Series A (General), 135(3), 370–384.
Nersesian, C. L., Banks, P. B., Simpson, S. J., & McArthur, C. (2012). Mixing nutrients mitigates the intake constraints of a plant toxin in a generalist herbivore. Behavioral Ecology, 23(4), 879–888.
Noble, I. R. (1989). Ecological traits of the Eucalyptus subgenera Monocalyptus and Symphyomyrtus. Australian Journal of Botany, 37(3), 207–224.
O'Reilly-Wapstra, J., McArthur, C., & Potts, B. M. (2002). Genetic variation in resistance of Eucalyptus globulus to marsupial browsers. Oecologia, 130(2), 289–296.
O'Reilly-Wapstra, J. M., Potts, B. M., McArthur, C., & Davies, N. W. (2005). Effects of nutrient variability on the genetic-based resistance of Eucalyptus globulus to a mammalian herbivore and on plant defensive chemistry. Oecologia, 142(4), 597–605.
Press, M. C., & Phoenix, G. K. (2005). Impacts of parasitic plants on natural communities. New Phytologist, 166(3), 737–751.
Preston, A. L., An, M., & Watson, D. M. (2010). Chemical profile differences in endemic parasitic weeds: A study of host-parasite chemical profiles in select mistletoe and eucalypt species. Paper presented at the Seventeenth Australasian Weeds Conference, Christchurch, New Zealand.
Raubenheimer, D., Simpson, S. J., & Mayntz, D. (2009). Nutrition, ecology and nutritional ecology: Toward an integrated framework. Functional Ecology, 23(1), 4–16.
Reed, J. D. (1995). Nutritional toxicology of tannins and related polyphenols in forage legumes. Journal of Animal Science, 73(5), 1516–1528.
Rosenthal, G. A., & Berenbaum, M. R. (1992). Herbivores: Their interactions with secondary plant metabolites: Ecological and Evolutionary Processes (Vol. 2). San Diego: Academic Press.
Salminen, J. P., & Karonen, M. (2011). Chemical ecology of tannins and other phenolics: We need a change in approach. Functional Ecology, 25(2), 325–338.
Schultz, J. C. (1989). Tannin—insect interactions. In R. W. Hemingway & J. J. Karchesy (Eds.), Chemistry and Significance of Condensed Tannins (pp. 417–433). New York: Plenum Press.

192
Schulze, E. D., Turner, N. C., & Glatzel, G. (1984). Carbon, water and nutrient relations of two mistletoes and their hosts: A hypothesis. Plant, Cell & Environment, 7(5), 293–299.
Scriber, J. M. (1977). Limiting effects of low leaf-water content on the nitrogen utilization, energy budget, and larval growth of Hyalophora cecropia (Lepidoptera: Saturniidae). Oecologia, 28(3), 269–287.
Scrivener, N. J., Johnson, C. N., Wallis, I. R., Takasaki, M., Foley, W. J., & Krockenberger, A. K. (2004). Which trees do wild common brushtail possums (Trichosurus vulpecula) prefer? Problems and solutions in scaling laboratory findings to diet selection in the field. Evolutionary Ecology Research, 6(1), 77–87.
Shenk, J. S., & Westerhaus, M. O. (1991). Population definition, sample selection, and calibration procedures for near infrared reflectance spectroscopy. Crop Science, 31(2), 469–474.
Shimada, T., & Saitoh, T. (2003). Negative effects of acorns on the wood mouse Apodemus speciosus. Population Ecology, 45(1), 7–17.
Silanikove, N., Gilboa, N., Nir, I., Perevolotsky, A., & Nitsan, Z. (1996). Effect of a daily supplementation of polyethylene glycol on intake and digestion of tannin-containing leaves (Quercus calliprinos, Pistacia lentiscus, and Ceratonia siliqua) by goats. Journal of Agricultural and Food Chemistry, 44(1), 199–205.
Simpson, S. J., & Raubenheimer, D. (2012). The nature of nutrition: A unifying framework. Australian Journal of Zoology, 59(6), 350–368.
Sinclair, S. J. (2006). The influence of dwarf cherry (Exocarpos strictus) on the health of river red gum (Eucalyptus camaldulensis). Australian Forestry, 69(2), 137–141.
Snyder, M. A., Fineschi, B., Linhart, Y. B., & Smith, R. H. (1996). Multivariate discrimination of host use by dwarf mistletoe Arceuthobium vaginatum subsp. cryptopodum: Inter- and intra-specific comparisons. Journal of Chemical Ecology, 22(2), 295–305.
Stalenberg, E. (2010). Spatial variation in habitat quality effects fine-scale resource use by a low density koala population. (Honours thesis), the Australian National University, Canberra.
Spotfire S+® Version 8.1. (2010). TIBCO Software Inc., Somerville. Tabachnick, B. G., & Fidell, L. S. (1996). Multivariate statistics. New York: Harper Collins
College Publishing. Takemoto, H. (2003). Phytochemical determination for leaf food choice by wild
chimpanzees in Guinea, Bossou. Journal of Chemical Ecology, 29(11), 2551–2573. Tixier, H., Duncan, P., Scehovic, J., Yant, A., Gleizes, M., & Lila, M. (1997). Food selection
by European roe deer (Capreolus capreolus): Effects of plant chemistry, and consequences for the nutritional value of their diets. Journal of Zoology, 242(2), 229–245.
Tucker, D., Wallis, I. R., Bolton, J., Marsh, K. J., Rosser, A., Brereton, I.,Foley, W. J. (2010). A metabolomic approach to identifying chemical mediators of mammal–plant interactions. Journal of Chemical Ecology, 36(7), 727–735.
Umucalilar, H., Gulsen, N., Coskun, B., Hayirli, A., & Dural, H. (2007). Nutrient composition of mistletoe (Viscum album) and its nutritive value for ruminant animals. Agroforestry Systems, 71(2), 77–87.
Venables, W. N., & Ripley, B. D. (2002). Modern Applied Statistics with S (4 ed.). New York: Springer.
Wallis, I. R., Watson, M. L., & Foley, W. J. (2002). Secondary metabolites in Eucalyptus melliodora: Field distribution and laboratory feeding choices by a generalist herbivore, the common brushtail possum. Australian Journal of Zoology, 50(5), 507–519.
Wallis, I. R., Nicolle, D., & Foley, W. J. (2010). Available and not total nitrogen in leaves explains key chemical differences between the eucalypt subgenera. Forest Ecology and Management, 260(5), 814–821.

193
Wallis, I. R., Edwards, M. J., Windley, H., Krockenberger, A. K., Felton, A., Quenzer, M., ... & Foley, W. J. (2012). Food for folivores: Nutritional explanations linking diets to population density. Oecologia, 169(2), 281–291.
Watson, D. M. (2001). Mistletoe—a keystone resource in forests and woodlands worldwide. Annual Review of Ecology and Systematics, 32(1), 219–249.
Watson, D. M. (2009). Determinants of parasitic plant distribution: The role of host quality. Botany, 87(1), 16–21.
White, T. C. R. (1984). The abundance of invertebrate herbivores in relation to the availability of nitrogen in stressed food plants. Oecologia, 63(1), 90–105.
Wiggins, N., McArthur, C., & Davies, N. W. (2006). Diet switching in a generalist mammalian folivore: Fundamental to maximising intake. Oecologia, 147(4), 650–657.
Woolnough, A. P., & Foley, W. J. (2002). Rapid evaluation of pasture quality for a critically endangered mammal, the northern hairy-nosed wombat (Lasiorhinus krefftii). Wildlife Research, 29(1), 91–100.
Youngentob, K., Wallis, I. R., Lindenmayer, D. B., Wood, J., Pope, M., & Foley, W. J. (2011). Foliage chemistry influences tree choice and landscape use of a gliding marsupial folivore. Journal of Chemical Ecology, 37(1), 71–84.
Zweifel‐Schielly, B., Leuenberger, Y., Kreuzer, M., & Suter, W. (2011). A herbivore's food landscape: Seasonal dynamics and nutritional implications of diet selection by a red deer population in contrasting Alpine habitats. Journal of Zoology, 286(1), 68–80.

194
CH
AP
TE
R
5
Herbivory in the heterogeneous world

195
CHAPTER 5
Herbivory in the heterogeneous world
Synthesis of results and conclusions
Herbivores are known to make decisions and alter their movements and foraging
behaviour at different scales of resource heterogeneity, ranging from geographic regions
and landscapes to plant communities, species, individuals of the same species, and even
plant parts. To understand the ecology of herbivores in heterogeneous environments it is
important to ascertain the critical scales at which the processes involved in resource
selection occur. Johnson (1980) identified four orders of resource selection by animals:
the geographic range of a species, the home range of an individual, the various habitats
used by an individual within its home range, and the individual resources selected within
each habitat. Hence, resource selection of herbivores is influenced by broad scale
processes, such as regional climate and soil types; landscape-scale processes, such as the
amount and configuration of habitat; and fine-scale processes, such as the chemistry of
individual plants or plant parts (Moore & DeGabriel, 2012; Senft et al., 1987).
The present work has focused on exploring underlying patterns of resource selection
(i.e., food and shelter) and space use-patterns of the common brushtail possum (Trichosurus
vulpecula, Marsupialia: Phalangeridae). This generalist arboreal herbivore has been observed
to respond to multiple scales of resource heterogeneity (i.e., regional, landscape, and
habitat), and plant internal heterogeneity or chemical variability of different tree species,
individuals, and tree parts (DeGabriel, Moore, Marsh, & Foley, 2010). Hence, this system
provides a unique opportunity to study complex plant–herbivore interactions in a species
rich and chemically heterogeneous environment and to relate these interactions to other
spatial ecological processes, such as competition over resources (i.e., food and shelter) and
a potential risk of predation. In the present study, additive and interacting effects of
environmental heterogeneity on resource selection by common brushtail possums were
studied at three spatial scales: the landscape, home range, and tree scale.

196
In Chapter 2, resource selection by the entire population of common brushtail
possums (Trichosurus vulpecula) and indirectly measured competition with a sympatric
species, the mountain brushtail possum (T. cunninghami), were studied at a
landscape-scale.
Next, in Chapter 3, individual variability in resource use (i.e., food and shelter) and
space-use patterns of selected common brushtail possums were studied at a home
range scale.
Finally, in Chapter 4, food selection by common brushtail possums in response to
the nutritional quality of forage was explored at a tree-scale.
By studying resource selection and space-use patterns of common brushtail possums at
different scales, landscape, home range, and individual tree, the opportunity arose to
compare and contrast findings across different ecological metrics and answer more general
questions about herbivory: What is the realized diet breadth of generalist herbivores and
are they facultative specialists in a heterogeneous world of many choices? Do individual
differences in resource use, such as selective foraging on different species of plants and
individuals of the same species, lead to the expression of different animal personalities?
Is using a single currency to measure the nutritional value of forage for herbivores
obscuring the complex effects of multiple nutrients and toxins?
In Chapter 2 resource selectivity by common brushtail possums was examined in the
context of food availability (species composition of trees and parasitic plants), hollow
availability (used as shelter), as well as potential competition with a sympatric species, the
mountain brushtail possum. The main aim of this study was to document the diet breadth
and food preferences of common brushtail possums and to investigate underlying factors
influencing their tree use at a broad landscape-scale. As demonstrated in other similar
studies (Freeland & Winter, 1975; Marsh, Wallis, McLean, Sorensen, & Foley, 2006;
Nersesian, Banks, Simpson, & McArthur, 2012), common brushtail possums behaved as
true generalist herbivores consuming a wide variety of foods apart from Eucalyptus leaves,
including leaves of different species of Acacia; exotic trees; parasitic plants, Amyema pendula
and Exocarpos cupressiformis; as well as understorey species and fruits of Rubus fruticosus.

197
However, despite their generalist feeding strategy, common brushtail possums exhibited
clear foraging preferences, ranging across forest types and species of trees to discrete
patches of mistletoes within tree canopies. Acacia dealbata, exotic trees, and parasitic plants
were eaten more frequently than expected from their availability in the habitat while habitat
dominant Eucalyptus species were consumed less frequently than expected. Additional
factors such as the availability of hollows and competition with mountain brushtail
possums might have shaped tree use by common brushtail possums.
In Chapter 3, space-use patterns and resource selection (i.e., food and shelter) of common
brushtail possums were explored within their home ranges. As expected from previous
studies (DeGabriel, Moore, Foley, & Johnson, 2009; Martin & Martin, 2007), males had
larger home ranges than females. The total home range size of both males and females was
strongly correlated with foraging area and only weakly correlated with denning area,
suggesting that food availability might have been a more limiting factor for common
brushtail possums than availability of hollow-bearing trees. Moreover, individual animals
preferred different species of trees for foraging and denning and repeatedly used individual
trees of the same species, as well as trees parasitized by mistletoes. Common brushtail
possums differentiated between trees used as foraging and denning sites based on the
species and the number of hollows and used trees with a greater number of hollows more
frequently as denning sites. Individual animals showed preferences for different species of
trees used as foraging sites, reflecting different tree species composition or the structural
heterogeneity of home ranges leading to dietary specialization.
In Chapter 4, a single measure of forage nutritional quality, available nitrogen, was used
for cross-taxa comparisons of forage available to common brushtail possums within their
home ranges. This measure combined the foliar concentrations of total nitrogen and
digestibility-reducing tannins and fibre on the amount of nitrogen that was readily available
to herbivores. The chemical analysis of the nutritional quality of forage revealed that,
despite being subjected to similar environmental conditions, co-occurring tree genera and
species reflected their diverse life strategies by exhibiting broad differences in foliar
concentrations of available nitrogen, tannins, and fibre. Unlike previous studies (Marsh,
Foley, Cowling, & Wallis, 2003; Moore et al., 2004), the present research demonstrated that
common brushtail possums used both Monocalyptus and Symphyomyrtus species while
foraging at night.

198
The most often used species, Eucalyptus radiata (Monocalyptus) and E. camphora
(Symphyomyrtus), differed in their nutritional quality. The former species had the lowest
available nitrogen and highest tannin-bound nitrogen concentrations of all used species
while the latter had the highest available nitrogen of all analysed eucalypts. Occasionally
consumed Acacia dealbata had the highest concentration of available nitrogen while parasitic
plants which were preferred by common brushtail possums, Exocarpos cupressiformis and
Amyema pendula, had moderate to high concentrations of available nitrogen, with foliar
nitrogen concentrations of mistletoe varying depending on the species of tree host.
Contrary to expectations, hardly any intraspecific differences were observed among trees
repeatedly used by common brushtail possums, suggesting that some other unmeasured
plant chemical and physical factors combined with a potential risk of predation might have
been involved in animal foraging decisions.
In summary, the findings of this study have highlighted the previous studies’ over-
simplification of the complex food-web interaction in Australian forests and woodlands.
The previous studies focused exclusively on the availability and nutritional quality for
arboreal marsupials of one genus, Eucalyptus, and at the same time excluded other tree
genera and parasitic plants. This study highlighted the need to move on from a traditional
division of arboreal marsupials into either specialist or generalist herbivores, by showing
that generalist species, such as the common brushtail possum, can be selective at multiple
scales of resource heterogeneity, ranging across landscapes, home ranges, and individual
trees to discrete patches of mistletoes within tree canopies (Fig. 1).

199
Fig. 1 Drivers of resource selectivity by common brushtail possums at different spatial scales

200
Furthermore, based on the recognized dietary preferences of entire populations of
generalist herbivores and the specific “food tastes” of individual members of a population,
we can conceptualise generalist herbivores as a specialized group of consumers when
offered a smörgåsbord. Moreover, this study demonstrated that applying a single currency,
available nitrogen, to measure the nutritional quality of different plant species and taxa for
generalist herbivores simplifies complex interactions between multiple nutrients and toxins
that shape animal dietary choices. Only by acknowledging the complexities of generalist
herbivores’ diet balancing processes and by taking into account other confounding
processes, such as competition and predation, it is possible to understand how animals
select their diets in heterogeneous environments.
Future studies need to reconcile the five nutritional goals of herbivores: nitrogen (protein)
maximization (Mattson, 1980), energy maximization (Belovsky, 1986), avoidance and
regulation of plant secondary metabolites (Freeland & Janzen, 1974), limitation of fibre
(Demment & Soest, 1985), and nutrient balancing (Westoby, 1978). To achieve this
synthesis, I suggest structuring research around six levels of inquiry: Level (1) What food is
available within the animal’s home range? Level (2) What is the animal dietary breadth?
Level (3) Which properties of food determine inclusion in the overall diet? Level (4) Which
properties determine the preference ranking of foods that are a part of the overall diet?
Level (5) What are the nutritional goals that underlie the choice and ranking of foods?
Level (6) What are other confounding factors affecting herbivore nutrition?
The current study provides a baseline for future nutritional studies by highlighting the
specialized nature of generalized diets of herbivores expressed over multiple scales of
resource heterogeneity and encourages a more holistic approach in studies of nutritional
quality of forage for herbivores. Recently, DeGabriel and colleagues (2013) identified four
steps that need to be achieved in order to understand nutritional complexity of herbivores:
(1) knowing what foods and how much of these foods wild browsers eat, as well as what
they avoid eating; (2) knowing the relevant aspects of plant nutritional and defensive
chemistry to measure in a given system and how to measure them; (3) understanding the
spatial distribution of nutrients and plant secondary metabolites in plant communities, the
costs they impose on foraging, and the effects on animals’ distributions; and (4) having
appropriate statistical tools to analyze the data.

201
Yet, as the current study has demonstrated, identifying and quantifying what herbivores eat
and even more what they avoid eating is difficult and subjected to many biases and
technical difficulties. Namely, direct foraging observations are difficult in the case of
arboreal marsupials as discussed in Chapter 2 and 3; yet, some technological advances,
such as audio-telemetry and video-tracking are promising (Logan & Sanson, 2002).
Similarly, the fecal content analysis that underestimates participation of species with highly
digestible cuticles can be replaced by methods that use plant cuticular markers (i.e., n-
alkanes, alcohols and long-chain fatty acids coupled with carbon isotope ratios or genetic
markers to determine herbivore diets from faeces (Dove & Mayes, 2006; Bezabih et al.,
2011; Valentini et al., 2009).
Also, measuring relevant nutrients and toxins for herbivore dietary choices represents a
challenge for nutritional ecologists as discussed in Chapter 4. So far, we know more about
type and concentrations of toxins that make herbivores avoid certain foods than
macronutrients that make them consume others. The Geometric Framework (Simpson &
Raubenheimer, 2001) provides a unifying approach to explore effects of both toxins and
nutrients on animal food intake and digestion. Moreover, types and concentrations of
nutrients and toxins vary within and between species, regions, and seasons, and in order to
measure environmental chemical variability, rapid and cheap methods for analysing a large
number of samples are needed. One well-established solution is near infrared reflectance
spectroscopy (NIRS; see Chapter 4 for details), and recent advances include the
development of portable devices to record spectra in the field and the use of airborne
hyper-spectral sensing techniques to measure the spectra from whole tracts of forests
(HYMAP; Dury, Turner, & Foley, 2001). Furthermore, new statistical tools are needed to
deal with the complexity of nutritional data and the non-linear relationship between
nutrient and toxin concentrations and the food intake of herbivores as discussed by
DeGabriel and colleagues (2013). Finally, the effects of increased atmospheric CO2 on
foliar concentrations of carbon-based secondary or structural compounds may have far-
reaching consequences for herbivory and should be incorporated in future studies
exploring the foraging patterns of herbivores.

202
References
Belovsky, G. E. (1986). Optimal foraging and community structure: Implications for a guild of generalist grassland herbivores. Oecologia, 70(1), 35–52.
Bezabih, M., Pellikaan, W. F., Tolera, A., & Hendriks, W. H. (2011). Evaluation of n-alkanes and their carbon isotope enrichments (δ13C) as diet composition markers. Animal, 5(01), 57–66.
DeGabriel, J., Moore, B. D., Marsh, K. J., & Foley, W. J. (2010). The effect of plant secondary metabolites on the interplay between the internal and external environments of marsupial folivores. Chemoecology, 20(2), 97–108.
DeGabriel, J. L., Moore, B. D., Foley, W. J., & Johnson, C. N. (2009). The effects of plant defensive chemistry on nutrient availability predict reproductive success in a mammal. Ecology, 90(3), 711–719.
DeGabriel, J. L., Moore, B. D., Felton, A. M., Ganzhorn, J. U., Stolter, C., Wallis, I. R., ... & Foley, W. J. (2013). Translating nutritional ecology from the laboratory to the field: Milestones in linking plant chemistry to population regulation in mammalian browsers. Oikos, 123(3), 298–308.
Demment, M. W., & Soest, P. J. V. (1985). A Nutritional Explanation for Body-Size Patterns of Ruminant and Nonruminant Herbivores. The American Naturalist, 125(5), 641–672.
Dove, H., & Mayes, R. W. (2006). Protocol for the analysis of n-alkanes and other plant-wax compounds and for their use as markers for quantifying the nutrient supply of large mammalian herbivores. Nature Protocols, 1(4), 1680–1697.
Dury, S. J., Turner, B. J., & Foley, W. J. (2001). Can hyperspectral data be used to map koala and possum habitat?. In Geoscience and Remote Sensing Symposium, 2001. IGARSS'01. IEEE 2001 International (Vol. 4, pp. 1648–1650). IEEE.
Freeland, W. J., & Janzen, D. H. (1974). Strategies in herbivory by mammals: The role of plant secondary compounds. The American Naturalist, 108(961), 269–287.
Freeland, W. J., & Winter, J. W. (1975). Evolutionary consequences of eating:Trichosurus vulpecula (Marsupialia) and the genus Eucalyptus. Journal of Chemical Ecology, 1(4), 439–455.
Johnson, D. H. (1980). The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61(1), 65–71.
Logan, M., & Sanson, G. D. (2002). The effects of tooth wear on the activity patterns of free-ranging koalas (Phascolarctos cinereus Goldfuss). Australian Journal of Zoology, 50(3), 281–292.
Marsh, K. J., Foley, W. J., Cowling, A., & Wallis, I. R. (2003). Differential susceptibility to Eucalyptus secondary compounds explains feeding by the common ringtail (Pseudocheirus peregrinus) and common brushtail possum (Trichosurus vulpecula). Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology, 173(1), 69–78.
Marsh, K. J., Wallis, I. R., McLean, S., Sorensen, J. S., & Foley, W. J. (2006). Conflicting demands on detoxification pathways influence how common brushtail possums choose their diets. Ecology, 87(8), 2103–2112.
Martin, J. K., & Martin, A. A. (2007). Resource distribution influences mating system in the bobuck (Trichosurus cunninghami: Marsupialia). Oecologia, 154(1), 227–236.
Mattson, W. J. (1980). Herbivory in relation to plant nitrogen content. Annual Review of Ecology and Systematics, 11, 119–161.
Moore, B. D., & DeGabriel, J. L. (2012). Integrating the effects of PSMs on vertebrate herbivores across spatial and temporal scales. In G. R. Iason, M. Dicke & S. E. Hartley (Eds.), The Ecology of Plant Secondary Metabolites: From Genes to Global Processes (pp. 226–246). Cambridge: Cambridge University Press.

203
Moore, B. D., Wallis, I. R., Marsh, K. J., & Foley, W. J. (2004). The role of nutrition in the conservation of the marsupial folivores of eucalypt forests. In D. Lunney (Ed.), Conservation of Australia’s forest fauna (2 ed., pp. 549–575). Mosman: Royal Zoological Society of New South Wales.
Nersesian, C. L., Banks, P. B., Simpson, S. J., & McArthur, C. (2012). Mixing nutrients mitigates the intake constraints of a plant toxin in a generalist herbivore. Behavioral Ecology, 23(4), 879–888.
Senft, R. L., Coughenour, M. B., Bailey, D. W., Rittenhouse, L. R., Sala, O. E., & Swift, D. M. (1987). Large herbivore foraging and ecological hierarchies. BioScience, 37(11), 789–799.
Simpson, S. J., & Raubenheimer, D. (2001). The geometric analysis of nutrient—allelochemical interactions: A case study using locusts. Ecology, 82(2), 422–439.
Valentini, A., Miquel, C., Nawaz, M. A., Bellemain, E. V. A., Coissac, E., Pompanon, F., ... & Taberlet, P. (2009). New perspectives in diet analysis based on DNA barcoding and parallel pyrosequencing: the trnL approach. Molecular Ecology Resources, 9(1), 51–60.
Westoby, M. (1978). What are the biological bases of varied diets? The American Naturalist, 112(985), 627–631.
Top Related