







Languages
Pages
Legal


MOLECULARPHYLOGENETICSAND
Molecular Phylogenetics and Evolution 32 (2004) 951–977
EVOLUTION
www.elsevier.com/locate/ympev
Generic delimitation and phylogenetic relationshipswithin the subtribe Chironiinae (Chironieae: Gentianaceae),with special reference to Centaurium: evidence from nrDNA
and cpDNA sequences
Guilhem Mansiona,* and Lena Struweb
a Laboratoire de Botanique Evolutive, Universit�e de Neuchatel, CH-2007 Neuchatel, Switzerlandb Department of Ecology, Evolution, and Natural Resources, Rutgers University, New Brunswick, NJ 08901, USA
Received 21 October 2003; revised 7 March 2004
Available online 4 June 2004
Abstract
To better understand the evolutionary history of the genus Centaurium and its relationship to other genera of the subtribe
Chironiinae (Gentianaceae: Chironieae), molecular analyses were performed using 80 nuclear ribosomal ITS and 76 chloroplast
trnLF (both the trnL UAA intron and the trnL–F spacer) sequences. In addition, morphological, palynological, and phytochemical
characters were included to a combined data matrix to detect possible non-molecular synapomorphies. Phylogenetic reconstructions
support the monophyly of the Chironiinae and an age estimate of ca. 22 million years for the subtribe. Conversely, both molecular
data sets reveal a polyphyletic Centaurium, with four well-supported main clades hereafter treated as separate genera. The primarily
Mediterranean Centaurium s.s. is closely related to southern African endemics Chironia and Orphium, and to the Chilean species
Centaurium cachanlahuen. The resurrected Mexican and Central American genus Gyrandra is closely related to Sabatia (from eastern
North America). Lastly, the monospecific genus Exaculum (Mediterranean) forms a monophyletic group together with the two new
genera: Schenkia (Mediterranean and Australian species) and Zeltnera (all other indigenous American centauries). Several bio-
geographical patterns can be inferred for this group, supporting a Mediterranean origin followed by dispersals to (1) North
America, Central America, and South America, (2) southern Africa (including the Cape region), and (3) Australia and Pacific
Islands.
� 2004 Elsevier Inc. All rights reserved.
Keywords: Biogeography; Character evolution; Centaurium; Gentianaceae; Gyrandra; Molecular clock; Phylogeny; Schenkia; Zeltnera
1. Introduction
Early classifications of the Gentianaceae were estab-lished by Endlicher (1838), Grisebach (1839), Bentham
and Hooker (1876), Knoblauch (1894), and Gilg (1895).
Despite important systematic progress achieved either
with traditional data (e.g., Broome, 1973; M�esz�aros,1994, 1996; Struwe et al., 1994; Zeltner, 1970) or mo-
lecular approaches (e.g., Chassot et al., 2001; Yuan
and K€upfer, 1995; Yuan et al., 1996, 2003), only the
* Corresponding author. Fax: +41-32-718-3001.
E-mail address: [email protected] (G. Mansion).
1055-7903/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.ympev.2004.03.016
infrafamilial classification of Gilg (1895), based on
pollen characters and gross morphology, was used as a
reference during the 20th century. With the increase inmolecular tools and results, a new classification was
proposed, which combined phylogenetic approaches
and traditional data (Struwe et al., 2002). In this treat-
ment, the gentian family (87 genera and ca. 1615–1688
species) has been organized into six monophyletic tribes
(Chironieae, Exaceae, Gentianeae, Helieae, Potalieae,
and Saccifolieae) based on cladistic analyses of trnL
(UAA) intron and matK sequence data. The tribeChironieae is an important assemblage of 23 genera and
ca. 160 species (Struwe et al., 2002). This tribe was
further subdivided in three monophyletic subtribes, the

952 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
Canscorinae, Chironiinae, and Coutoubeinae, on thebasis of pollen type (monads vs. tetrads), corolla tube
length, and anther spiralization.
Within the subtribe Chironiinae, the genus Centau-
rium (commonly called centaury) comprises mostly
short-lived (annual or biennial, rarely perennial) her-
baceous species and is characterized by the coiling of the
anthers after pollen release. The taxa are widely dis-
tributed in wet and disturbed habitats worldwide, wherecompetition is reduced. They can be encountered along
roadsides, on stream banks, in fields and pastures or in
open forests in most parts of the northern hemisphere.
Two main regions of diversification are recognized: one
in the Old World and another in the New World. The
Old World species are centered in the Mediterranean
basin and radiate in a northeasterly manner. About 25
diploid and polyploid species occur in this region(Zeltner, 1970). All New World species (�30 spp.) are
polyploids and most occur in western North America
from Mexico to Canada (British Columbia), with cen-
ters of high diversity in California and Texas. Some
species extend southward to Central and South
America.
Centaurium appears to be the largest genus of the
Chironieae, even though the exact number of species isnot yet clearly established. The species circumscription
within Centaurium is difficult because traditional mor-
phological characters discriminate species poorly.
Moreover, a large amount of morphological plasticity
occurs due to varying environmental conditions and
natural hybridization that may obscure species bound-
aries (Melderis, 1931; Zeltner, 1970). The abundance of
polymorphisms found within different taxa has led tothe naming of many species in Centaurium and different
nomenclatural opinions among authors (e.g., Gilg, 1895;
Grisebach, 1839; Hegi, 1966; Melderis, 1931; Robyns,
1954; Zeltner, 1970). Based on field experience and ex-
amination of different herbaria collections, a good esti-
mate of the actual species number appears to be 55–60
(Mansion, unpublished).
The phylogenetic position of Centaurium withinChironieae is not clear. Molecular trees inferred from
combined sequences of the matK and trnL intron re-
gions support its inclusion in the subtribe Chironiinae,
close to the genus Sabatia (Struwe et al., 2002). ITS
results placed Centaurium in the same subtribe, but as a
sister clade to Chironia plus Orphium (Thiv et al.,
1999a). Only a few species of Centaurium have been
investigated so far with molecular tools, and somegenera such as Exaculum were not sampled. The previ-
ous molecular-based hypotheses of Chironieae phylog-
eny do not reflect the whole range of diversity within the
subtribe. Consequently, the intergeneric relationship of
Centaurium has remained obscure. No cladistic studies
have previously been performed on the entire genus, and
the monophyly of Centaurium has not been established.
In addition, only a few morphological studies have beenperformed on the Gentianaceae, and little is known
concerning the evolution of vegetative and floral char-
acters within the subtribe Chironiinae and particularly
Centaurium.
The main goals of this study were (1) to resolve the
phylogenetic position of the subtribe Chironiinae
within the tribe Chironieae, (2) to investigate the in-
tergeneric relationships within the subtribe Chironiinae,(3) to test the monophyly of Centaurium, and (4) to use
the phylogenetic reconstruction of the Chironiinae as a
framework to infer biogeographic scenarios and char-
acter evolution in this group. For these purposes se-
quences of the internal transcribed spacer (ITS) of
nrDNA and a combination of the trnL UAA intron
with the trnL–F spacer (trnLF region) of cpDNA were
used for cladistic analyses. These regions have beenwidely used to infer phylogenetic relationships within
the Gentianaceae at both tribal, generic and species
levels (Chassot et al., 2001; Gielly and Taberlet, 1996;
Struwe et al., 2002; Thiv et al., 1999a,b; von Hagen and
Kadereit, 2001; Yuan and K€upfer, 1995, 1997; Yuan
et al., 1996, 2003).
2. Materials and methods
2.1. Sampling
In total, 80 taxa were included in the ITS data set and
76 in the trnL intron+ trnL–F spacer matrix (hereafter
called trnLF). To investigate the phylogenetic position
of the Chironiinae within the Chironieae, several ac-cessions representing neighboring tribes and subtribes
recognized by Struwe et al. (2002) were included in the
analyses (Table 1). One of the more basal tribes in the
family, Exaceae, represented by the genera Exacum and
Sebaea, was chosen as a functional outgroup (Struwe
et al., 2002; Yuan et al., 2003). Within the Chironieae,
subtribe Canscorinae was represented by Canscora,
Hoppea, Microrphium, Schinziella, and subtribe Cou-toubeinae by Coutoubea, Deianira, Schultesia, and
Symphyllophyton. Most of the genera comprising sub-
tribe Chironiinae, i.e., Blackstonia, Centaurium, Cicen-
dia, Chironia, Eustoma, Exaculum, Geniostemon,
Ixanthus, Orphium, and Sabatia, were sequenced for
several species for the ITS (58 accessions) and trnLF
regions (48 accessions). Only the rare Brazilian genus
Zygostigma is missing due to lack of suitable materialfor molecular studies. The Caribbean endemic Bis-
goeppertia was excluded since recent results indicate that
Bisgoeppertia, tentatively included by Struwe et al.
(2002) in the subtribe Chironiinae, is more closely re-
lated to Lisianthius and thus fits better in the Potalieae
(M. Thiv, pers. comm.). Most of the Centaurium species
included in this study were collected and determined in

Table 1
Origin of plant material, voucher information, and GenBank accessions for DNA sequences used in this paper
Taxa Voucher information Location GenBank Accession Nos. References
ITS1 ITS2 trnL (UAA)
intron
trn L-F spacer
Anthocleista amplexicaulis Baker S. Wohlhauser SWPBZT (NEU) Madagascar n/a n/a AJ490189* n/a Yuan et al.
(2003)
Anthocleista grandiflora Gilg M. Callmander s.n. (NEU) Madagascar AJ489864* AJ489864* AJ490190* AY251777 Yuan et al.
(2003)/This study
Anthocleista vogelii Planch. S. A. Thompson & J.E. Rawlins 1399
(NY)
n/a AY251688 AY251718 AF102377 n/a Struwe et al.
(1998)
Blackstonia acuminata (W.D.J. Koch)
Domin
Licht (MJG) Italy AJ011468* AJ011477* n/a n/a Thiv et al.
(1999a,b)
Blackstonia grandiflora Maire G. Mansion 010832 (NEU) Spain AY251684 AY251714 AY251742 AY251768 This study
Blackstonia imperfoliata (L.F.) Samp. G. Mansion 010833 (NEU) Spain AY251685 AY251715 AY251743 AY251769 This study
Blackstonia imperfoliata (L.F.) Samp. M. Thiv (HBM) Italy AJ011470* AJ011480* n/a n/a Thiv et al.
(1999a,b)
Blackstonia perfoliata (L.) Huds. G. Mansion 98712 (NEU) France AY047793 AY047878 AF402198 AF402254 This study
Blackstonia perfoliata (L.) Huds. L. & N. Zeltner 2050 (NEU) Spain AY251686 AY251716 n/a n/a This study
Canscora alata (Roth) Wallich J. C. Piso, S. Wohlhauser & L. Zeltner
MO24 (NEU)
Madagascar AJ489865* AJ489865* AJ490191* n/a Yuan et al.
(2003)
Canscora andrographioides Griff. P. Chassot 99-234 (NEU) Thailand AJ489866* AJ489866* AJ490192* n/a Yuan et al.
(2003)
Canscora diffusa (Vahl.) Roem.
& Schult.
Kokou s.n. (TOGO) n/a AY256386 AY256391 AF102389* AY251779 Struwe et al.
(1998)/This study
Canscora diffusa (Vahl.) Roem.
& Schult.
P. Chassot 99-231 (NEU) Thailand AJ489867* AJ489867* AJ490193* AY251780 Yuan et al.
(2003)/This study
Centaurium cachanlahuen (Molina)
B. L. Robinson
L. Zeltner 020501 (NEU) Chile AY251694 AY251724 AY251749 AY251786 This study
Centaurium cachanlahuen (Molina)
B. L. Robinson
L. Zeltner 020502 (NEU) Chile AY251695 AY251725 AY251750 AY251787 This study
Centaurium erythraea Rafn Anonymous 717367 (MEL) Australia AY251669 AY251699 AY251729 AY251753 This study
Centaurium favargeri Zeltner L. & N. Zeltner 2044 (NEU) Spain AY251670 AY251700 AY251730 AY251754 This study
Centaurium gypsicola (Boiss. et Reut.)
Ronniger
L. & N. Zeltner 2081 (NEU) Spain AY251671 AY251701 AY251731 AY251755 This study
Centaurium littorale (D. Turner) Gilmour L. & N. Zeltner 1816 (NEU) England AY251672 AY251702 AY251732 AY251756 This study
Centaurium maritimum (L.) Fritch G. Mansion 98904 (NEU) France AY251673 AY251703 AY251733 AY251757 This study
Centaurium pulchellum (Sw.) Druce G. Mansion 98505 (NEU) France AY047787 AY047872 AY251734 AY251758 This study
Centaurium scilloides (L. fil.) Samp. L. & N. Zeltner 981111 (NEU) France AY251675 AY251705 AY251737 AY251761 This study
Centaurium tenuiflorum (Hoffmgg.
& Link) Fritsch
L. & N. Zeltner 1767 (NEU) Morocco AY047773 AY047858 AY251735 AY251759 This study
Centaurium tenuiflorum (Hoffmgg.
& Link) Fritsch
L. & N. Zeltner 1766 (NEU) Morocco AY251674 AY251704 AY251736 AY251760 This study
Chelonanthus alatus (Aubl.) Pulle F. Bretagnolle & J. Piguet C3 (NEU) Ecuador AJ489868* AJ489868* AJ490194* AY251775 Yuan et al.
(2003)/This study
Chelonanthus angustifolius Gilg F. Bretagnolle & J. Piguet T11 (NEU) Ecuador AJ489869* AJ489869* AJ490195* AY251776 Yuan et al.
(2003)/This study
Chelonanthus purpurascens (Aubl.)
Struwe, S. Nilsson, & V. Albert
F. Bretagnolle & J. Piguet C92 (NEU) Colombia AJ489870* AJ489870* AJ490196* n/a Yuan et al.
(2003)
G.Mansio
n,L.Stru
we/Molecu
larPhylogenetics
andEvolutio
n32(2004)951–977
953

Table 1 (continued)
Taxa Voucher information Location GenBank Accession Nos. References
ITS1 ITS2 trnL (UAA)
intron
trn L-F spacer
Chironia baccifera L. M. Callmander & S. Wohlhauser A005
(NEU)
South Africa AY251690 AY251720 AY251746 AY251783 This study
Chironia laxa Gilg M. Callmander & S. Wohlhauser A003
(NEU)
South Africa AY251691 AY251721 n/a n/a This study
Chironia linoides L. M. Callmander & S. Wohlhauser A004
(NEU)
South Africa AY251692 AY251722 AY251747 AY251784 This study
Cicendia filiformis (L.) Delarbre M. Thiv 2156 (MJG) France AJ011463* AJ011473* AF102403* n/a Thiv et al.
(1999a,b)
Cicendia quadrangularis (Lam.) Griseb. P. Maas 8154 (U) n/a AY251682 AY251712 AF102404* AY251765 Struwe et al.
(1998)/This study
Coutoubea minor H.B.K. G. A. Romero 1684 (NY) n/a n/a n/a AF102407* n/a Struwe et al.
(1998)
Coutoubea ramosa Aubl. B. Hoffman & C. Capellaro 984 (NY) n/a n/a n/a AF102408* n/a Struwe et al.
(1998)
Coutoubea spicata Aubl. S. Mori 24349 n/a SM24075 SM24349 AY251745 AY251778 This study
Coutoubea spicata Aubl. S. Mori 24075 (NY) French Guiana n/a n/a AF102409* n/a Struwe et al.
(1998)
Deianira pallescens Cham. & Schlecht. W. A. Anderson 9385 (NY) n/a n/a n/a AF102410* AY251782 Struwe et al.
(1998)/This study
Eustoma exaltatum (L.) Salisb. L. & N. Zeltner 980610-1 (NEU) Mexico AY251697 AY251727 n/a n/a This study
Eustoma exaltatum (L.) Salisb. L. & N. Zeltner 980528-1 (NEU) Mexico AY251698 AY251728 AY251752 AY251789 This study
Eustoma russelianum G. Don G. Mansion s. n. (NEU) Switzerland
(cultivated)
AY251696 AY251726 AY251751 AY251788 This study
Exaculum pusillum Caruel P. K€upfer s. n. (NEU) Italy AY251681 AY251711 AY251740 AY251764 This study
Exacum affine Balf. Miller et al. 8238 Yemen, Socotra AJ489877* AJ489877* AJ490202* AY251770 Yuan et al.
(2003)/This study
Exacum caeruleum Balf. Miller et al. 11356 Yemen, Socotra AJ489882* AJ489882* AJ490207* AY251771 Yuan et al.
(2003)/This study
Exacum tetragonum Roxb. Ludin & Klackenberg 332 India AJ489907* AJ489907* n/a n/a Yuan et al.
(2003)
Fagraea fragrans Roxb. C.-H. Tsou 207 (NY) n/a AY251689* AY251719* AF102421* n/a Struwe et al.
(1998)
Geniostemon gypsophilum B.L. Turner G. Nesom et al. 7621 (LL) Mexico n/a n/a AF102429* AY251766 Struwe et al.
(1998)/This study
Gyrandra brachycalyx (Standley & L.O.
Willams) Mansion
G. Mansion, L. & N. Zeltner 990239
(NEU)
Mexico AY047770 AY047855 n/a n/a This study
Gyrandra brachycalyx (Standley & L.O.
Willams) Mansion
G. Mansion, L. & N. Zeltner 990205
(NEU)
Mexico AY047771 AY047856 AF402184 AF402240 This study
Gyrandra tenuifolia (Martens & Galeotti)
Mansion
G. Mansion, L. & N. Zeltner 990228
(NEU)
Mexico AY047772 AY047857 AF402186 AF402242 This study
Hoppea dichotoma Willd. C.D.K. Cook RHT307 (MJG) n/a n/a n/a AF102440* n/a Struwe et al.
(1998)
Ixanthus viscosus (Aiton) Griseb. M. Thiv s.n. Spain, Tenerife AJ011471* AJ011481* n/a n/a Thiv et al.
(1999a,b)
Ixanthus viscosus (Aiton) Griseb. P. K€upfer s.n. (NEU) Spain, Tenerife AY251683 AY251713 AY251741 AY251767 This study
954
G.Mansio
n,L.Stru
we/Molecu
larPhylogenetics
andEvolutio
n32(2004)951–977

Macrocarpaea macrophylla (Kunth) Gilg R. Callejas & H. Balslev 1030 (NY) Colombia AY256385 AY256390 AF102455 * n/a Struwe et al.
(1998)/This study
Macrocarpaea angelliae J.R. Grant
& Struwe
J.R. Grant 02-4289 (NEU) Ecuador AY397760 AY397761 n/a n/a Yuan et al.
(2003)
Megacodon stylophorus (C.B. Clarke)
H. Smith
Y.-M. Yuan 93-142 (NEU) China Z48109* Z48137* AJ315200* AY251773 Yuan et al.
(2003)/This study
Microphium pubescens C.B. Clarke P. Chassot 99-243 (NEU) Thailand AJ489916* AJ489916* AJ490241* AY251781 Yuan et al.
(2003)/This study
Orphium frutescens E. Meyer M. Callmander & S. Wohlhauser A001
(NEU)
South Africa AY251693 AY251723 AY251748 AY251785 This study
Sabatia angularis (L.) Pursh. T. Ghammers 4860 (NY) USA AJ011467* AJ011476* n/a n/a Thiv et al., 2000
Sabatia campestris Nutt. G. Mansion, L. & N. Zeltner 97706
(NEU)
USA AY256382 AY256387 AY255692 AY255696 This study
Sabatia dodecandra (L.) Britton,
Sterns & Poggenb.
J. Grant 97-2858 (NEU) USA AY256383 AY256388 AY255693 AY255697 This study
Sabatia stellaris Pursh. J. Grant 97-2871 (NEU) USA AY256384 AY256389 AY255694 AY255698 This study
Schenkia australis (R. Br.) Mansion L. & N. Zeltner 001203 (NEU) Australia AY251676 AY251706 n/a n/a This study
Schenkia australis (R. Br.) Mansion L. & N. Zeltner 001206 (NEU) Australia AY251677 AY251707 n/a n/a This study
Schenkia australis (R. Br.) Mansion L. & N. Zeltner 001209 (NEU) Australia AY251678 AY251708 n/a n/a This study
Schenkia australis (R. Br.) Mansion L. & N. Zeltner 001216 (NEU) Australia AY251679 AY251709 n/a n/a This study
Schenkia clementii (Domin.) Mansion L. & N. Zeltner 001201 (NEU) Australia AY251680 AY251710 AY251739 AY251763 This study
Schenkia spicata (L.) Mansion G. Mansion 981005 (NEU) France AY047791 AY047876 AF402196 AF402252 This study
Schenkia spicata (L.) Mansion L. & N. Zeltner 1756 (NEU) Morocco AY047792 AY047877 AF402197 AF402253 This study
Schinziella tetragona (Schinz) Gilg Malaisse 13852 (BR) n/a n/a n/a AF102479* n/a Struwe et al.
(1998
Schultesia guianensis (Aubl.) Malme C.C. Berg & A.J. Henderson BG 661
(NY)
n/a n/a n/a AF102480* n/a Struwe et al.
(1998)
Sebaea brachyphylla Griseb. J. Raynal 19414 Madagascar AJ489920* AJ489920* AY251744 AY251772 Yuan et al.
(2003)/This study
Sebaea madagascariensis Klack. J. C. Piso, S. Wohlhauser & L. Zeltner
MO18 (NEU)
Madagascar AJ489921* AJ489921* n/a n/a Yuan et al.
(2003)
Swertia perennis L. P. K€upfer s.n. (NEU) Switzerland AY251687 AY251717 AY255695 AY251774 This study
Symphyllophyton caprifolioides Gilg Ratter 6742 (E) Brasil AJ011462* AJ011472* AF102490* n/a Thiv et al.
(1999a,b)
Zeltnera abramsii (Munz) Mansion F. Bretagnolle & G. Mansion 990903
(NEU)
California AY047712 AY047797 AF402145 AF402201 This study
Zeltnera arizonica (A. Gray) Mansion G. Mansion, L. & N. Zeltner 97726
(NEU)
Texas AY047725 AY047810 AF402155 AF402211 This study
Zeltnera beyrichii (Torr. & A. Gray)
Mansion
G. Mansion, L. & N. Zeltner 96526
(NEU)
Texas AY047732 AY047817 AF402160 AF402216 This study
Zeltnera calycosa (Buckl.) Mansion G. Mansion, L. & N. Zeltner 96519
(NEU)
Texas AY047740 AY047825 AF402164 AF402220 This study
Zeltnera glandulifera (Correll) Mansion G. Mansion, L. & N. Zeltner 96518
(NEU)
Texas AY047731 AY047816 AF402159 AF402215 This study
Zeltnera madrensis (Hemsl.) Mansion G. Mansion, L. & N. Zeltner 990230
(NEU)
Mexico AY047742 AY047827 AF402166 AF402222 This study
Zeltnera martinii (Broome) Mansion G. Mansion, L. & N. Zeltner 990215
(NEU)
Mexico AY047749 AY047834 AF400268 AF402224 This study
Zeltnera multicaulis (B.L. Robinson)
Mansion
G. Mansion, L. & N. Zeltner 96515
(NEU)
Arizona AY047726 AY047811 AF402156 AF402212 This study
G.Mansio
n,L.Stru
we/Molecu
larPhylogenetics
andEvolutio
n32(2004)951–977
955
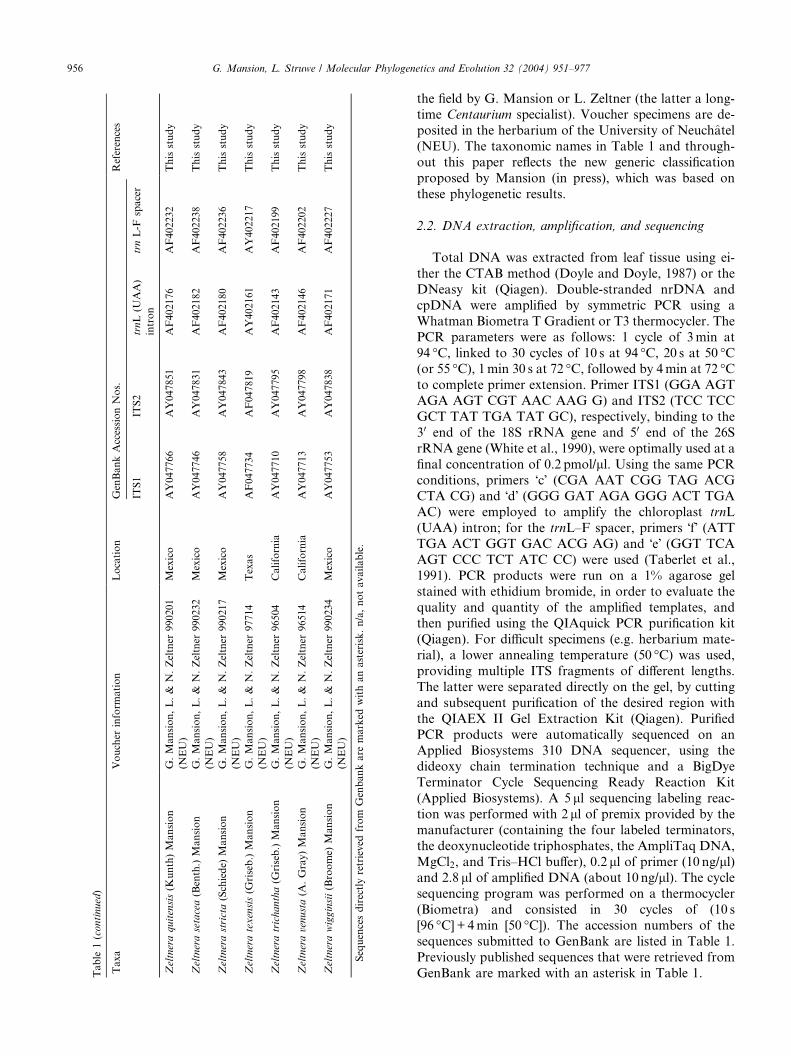
Table
1(continued)
Taxa
Voucher
inform
ation
Location
GenBankAccessionNos.
References
ITS1
ITS2
trnL(U
AA)
intron
trnL-F
spacer
Zeltneraquitensis(K
unth)Mansion
G.Mansion,L.&
N.Zeltner
990201
(NEU)
Mexico
AY047766
AY047851
AF402176
AF402232
Thisstudy
Zeltnerasetacea(Benth.)Mansion
G.Mansion,L.&
N.Zeltner
990232
(NEU)
Mexico
AY047746
AY047831
AF402182
AF402238
Thisstudy
Zeltnerastricta(Schiede)
Mansion
G.Mansion,L.&
N.Zeltner
990217
(NEU)
Mexico
AY047758
AY047843
AF402180
AF402236
Thisstudy
Zeltneratexensis(G
riseb.)Mansion
G.Mansion,L.&
N.Zeltner
97714
(NEU)
Texas
AF047734
AF047819
AY402161
AY402217
Thisstudy
Zeltneratrichantha(G
riseb.)Mansion
G.Mansion,L.&
N.Zeltner
96504
(NEU)
California
AY047710
AY047795
AF402143
AF402199
Thisstudy
Zeltneravenusta(A
.Gray)Mansion
G.Mansion,L.&
N.Zeltner
96514
(NEU)
California
AY047713
AY047798
AF402146
AF402202
Thisstudy
Zeltnerawigginsii(Broome)
Mansion
G.Mansion,L.&
N.Zeltner
990234
(NEU)
Mexico
AY047753
AY047838
AF402171
AF402227
Thisstudy
Sequencesdirectlyretrieved
from
Genbankare
marked
withanasterisk.n/a,notavailable.
956 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
the field by G. Mansion or L. Zeltner (the latter a long-time Centaurium specialist). Voucher specimens are de-
posited in the herbarium of the University of Neuchatel
(NEU). The taxonomic names in Table 1 and through-
out this paper reflects the new generic classification
proposed by Mansion (in press), which was based on
these phylogenetic results.
2.2. DNA extraction, amplification, and sequencing
Total DNA was extracted from leaf tissue using ei-
ther the CTAB method (Doyle and Doyle, 1987) or the
DNeasy kit (Qiagen). Double-stranded nrDNA and
cpDNA were amplified by symmetric PCR using a
Whatman Biometra T Gradient or T3 thermocycler. The
PCR parameters were as follows: 1 cycle of 3min at
94 �C, linked to 30 cycles of 10 s at 94 �C, 20 s at 50 �C(or 55 �C), 1min 30 s at 72 �C, followed by 4min at 72 �Cto complete primer extension. Primer ITS1 (GGA AGT
AGA AGT CGT AAC AAG G) and ITS2 (TCC TCC
GCT TAT TGA TAT GC), respectively, binding to the
30 end of the 18S rRNA gene and 50 end of the 26S
rRNA gene (White et al., 1990), were optimally used at a
final concentration of 0.2 pmol/ll. Using the same PCR
conditions, primers �c� (CGA AAT CGG TAG ACGCTA CG) and �d� (GGG GAT AGA GGG ACT TGA
AC) were employed to amplify the chloroplast trnL
(UAA) intron; for the trnL–F spacer, primers �f� (ATT
TGA ACT GGT GAC ACG AG) and �e� (GGT TCA
AGT CCC TCT ATC CC) were used (Taberlet et al.,
1991). PCR products were run on a 1% agarose gel
stained with ethidium bromide, in order to evaluate the
quality and quantity of the amplified templates, andthen purified using the QIAquick PCR purification kit
(Qiagen). For difficult specimens (e.g. herbarium mate-
rial), a lower annealing temperature (50 �C) was used,
providing multiple ITS fragments of different lengths.
The latter were separated directly on the gel, by cutting
and subsequent purification of the desired region with
the QIAEX II Gel Extraction Kit (Qiagen). Purified
PCR products were automatically sequenced on anApplied Biosystems 310 DNA sequencer, using the
dideoxy chain termination technique and a BigDye
Terminator Cycle Sequencing Ready Reaction Kit
(Applied Biosystems). A 5 ll sequencing labeling reac-
tion was performed with 2 ll of premix provided by the
manufacturer (containing the four labeled terminators,
the deoxynucleotide triphosphates, the AmpliTaq DNA,
MgCl2, and Tris–HCl buffer), 0.2 ll of primer (10 ng/ll)and 2.8 ll of amplified DNA (about 10 ng/ll). The cyclesequencing program was performed on a thermocycler
(Biometra) and consisted in 30 cycles of (10 s
[96 �C]+ 4min [50 �C]). The accession numbers of the
sequences submitted to GenBank are listed in Table 1.
Previously published sequences that were retrieved from
GenBank are marked with an asterisk in Table 1.

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 957
2.3. Maximum parsimony analyses
DNA sequences were initially aligned using the pro-
gram Clustal W (Higgins et al., 1996). The obtained
multiple alignment was manually adjusted by sequential
pairwise comparisons. Several parameters such as se-
quence length, transition/transversion ratio, GC con-
tent, and number of informative characters and pairwise
distance divergence values were computed using version4.0b10 of PAUP (Swofford, 2002).
Maximum parsimony (MP) analyses were first con-
ducted for each set of ITS and trnLF sequences sepa-
rately. Heuristic searches were performed under the
Fitch parsimony criterion using the following options in
Winclada version 1.00.08 (Nixon, 2002) and version 2.0
of NONA (Goloboff, 1999): 100 random repetitions
with five starting trees (seed¼ 0), saving at most 10,000most parsimonious (MP) trees, and using the mult*
max* option for branch swapping. Branch support was
performed using the jackknifing method (Farris et al.,
1996) as implemented in Winclada/NONA, with the
following settings: 100 repetitions with five replicates
each search, five starting trees (seed¼ 0). Homoplasy
measures were computed using the consistency index
(CI), excluding all uninformative characters, and theretention index (RI) as implemented in Winclada/
NONA.
2.4. Assessment of congruence
Three alternative strategies have been proposed for
handling multiple data sets, and this topic has given rise
to considerable debate regarding the advantages ordisadvantages of the so-called combined, the condi-
tional, or consensus approaches, respectively (De Que-
iroz et al., 1995; Huelsenbeck et al., 1996). The real
issues are how to determine the source of conflict be-
tween data sets, and more important, if there are any
evolutionary explanations for any differences between
data sets. Several statistical approaches have recently
been proposed to assess whether or not data are a merepartition of a global data set, thus sharing the same
natural history and evolutionary processes (Johnson
and Soltis, 1998).
The incongruence length difference test (ILD; Farris
et al., 1995) was used first to examine the phylogenetic
congruence between the respective nrDNA and cpDNA
data sets. This test, implemented in PAUP as the par-
tition homogeneity test (PHT), was performed with 100replicates of heuristic searches (invariant and ambigu-
ous characters excluded, Maxtree¼ 500 and TBR
branch swapping). It has been suggested that PHT p
values greater than p ¼ 0:01 reflect congruent data sets
that, if combined, will either improve or will not neg-
atively affect phylogenetic accuracy (Cunningham et al.,
1998).
Second, the strict consensus trees, obtained from theseparate analyses of each data set, were compared for
topology and branch support, allowing a possible
identification of potential localized sources of conflict.
2.5. Maximum likelihood analyses
Once the placement of the tribe Chironieae within the
Gentianaceae was clarified, a smaller 53-taxon data set,including only the Chironiinae and its neighboring sub-
tribes (Canscorinae and Coutoubeinae), was used to
perform full maximum likelihood (ML) analyses. In the
53-taxon data set chosen, the respective ITS and trnLF
partitions were combined following the total evidence
approach (Kluge, 1989; Section 3). Heuristic searches
were performed under theML criterion, using the general
time reversible (GTR) model of sequence substitutionwith the following settings: assumed nucleotide frequency
A¼ 0.2559, C¼ 0.2672, G¼ 0.2364, T¼ 0.240; substitu-
tion rates between A–C¼ 0.9152, A–G¼ 1.7496,
A–T¼ 0.8489, C–G¼ 0.4627, C–T¼ 2.5705, shape pa-
rameter gamma¼ 4.8303, assumed number of invariable
positions¼ 0. The best model was evaluated through the
Hierarchical Likelihood Ratio Test procedure as imple-
mented in Modeltest version 3.06 (Posada and Crandall,1998). The resulting ML tree was aimed to be used as a
framework for subsequent molecular clock estimations,
phenotypic characters mapping, and biogeographic
reconstructions.
2.6. Molecular clock and divergence time calculation
The estimation of divergence time using a molecularclock is highly controversial and should be treated with
caution (Sanderson, 1998). Nevertheless, it is the only
way of inferring diversification rates and lineage age
when fossil data are lacking, which is usually the case for
annual plants such as Centaurium and relatives.
To test the assumption of a clockwise molecular
evolution of DNA sequences for the subtribe Chiro-
niinae, a likelihood ratio test (LRT) was performed onthe 53-taxon data set, under the general time revers-
ible (GTR) model of sequence substitution. The LRT
was calculated as 2 lnðLclock=Lno�clockÞ, and assumed to
follow a v2 distribution with (n� 2) degree of free-
dom, where n is the number of taxa (Sanderson,
1998).
If the LRT failed to support a clock-like evolution of
the data set, relative branching time was estimated fromnon-parametric rate smoothing analyses (NPRS, clock-
independent method), using the default settings in
TreeEdit version 1.0a8 (Rambaut and Charleston, 2001;
Sanderson, 1997). The NPRS method does not assume
an underlying molecular clock and will score better than
a clock-based method if the sequences are not evolving
in a clock-like fashion (Sanderson, 1997).

958 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
To estimate absolute diversification rates, it is neces-sary to fix at least one node on the cladogram to a known
date, which can be done by using either paleobotanical
records or geological events (Sanderson, 1998). Only two
fossil pollen records are known from the Gentianaceae,
and neither of these are from Chironieae (Graham, 1984;
Scott, 1995), so we used geological dating based on the
age estimate of Gran Canaria of the Macaronesian Is-
lands, ca. 15 million years (MY) (Juan et al., 2000). Thisgeological estimation may be seen as the maximum age
of the divergence between the disjunct sister genera
Ixanthus (an endemic of Gran Canaria and more recent
islands of the Macaronesian Archipelago) and Blacks-
tonia (of Mediterranean origin) (Thiv et al., 1999a).
2.7. Phenotypic character optimization
To detect synapomorphic features supporting the
clades obtained with molecular data, and to better un-
derstand the state transformation of various important
characters within the Chironiinae, 27 phenotypic char-
acters (mainly morphological and phytochemical ones,
Table 2) were added to the 53-taxon molecular matrix.
These characters were mapped on the combined ML
cladogram, using the trace Character function andACCTRAN optimization as implemented in MacClade
(Maddison and Maddison, 1997). The character coding
for Centaurium species was largely based on personal
observations on living and herbarium material; for other
Table 2
Morphological, palynological, and phytochemical characters and their chara
1. Life form: 0¼ tree, shrub, or suffrutescent herb; 1¼perennial, non-suffr
2. Stem-x-section: 0¼ terete; 1¼ quadrangular; 2¼winged
3. Inflorescence: 0¼dichasial; 1¼monochasial; 2¼ cluster of sessile flowe
4. Calyx: 0¼ polymerous [8–12-merous]; 1¼ 5-merous; 2¼ 4-merous; 3¼ 2
5. Fusion of sepals: 0¼ scarcely; 1¼half; 2¼ almost completely
6. Abaxial side of calyx lobes: 0¼ smooth; 1¼keeled; 2¼winged
7. Calycine colleters: 0¼ absent; 1¼present
8. Corolla color: 0¼ green, yellow, or white; 1¼ blue, lilac, pink, or red
9. Corolla merosity: 0¼polymerous [8-12–merous]; 1¼ 5-merous; 2¼ 4-m
10. Corolla shape: 0¼ rotate (saucer-shaped); 1¼ funnel-shaped (infundibu
11. Petal fusion: 0¼ scarcely; 1¼half; 2¼ almost completely
12. Floral nectaries: 0¼ none (or rudimentary); 1¼on the corolla; 2¼ gyno
13. Anther shape: 0¼ non-sagittate; 1¼ sagittate
14. Anther shape at anthesis: 0¼not twisted; 1¼ recurved; 2¼helically twi
15. Filament bases: 0¼ not united; 1¼united by a membrane
16. Stamen insertion: 0¼near the base of the corolla tube; 1¼ between the ba
tube
17. Ovary shape: 0¼ globular; 1¼oval; 2¼ elliptic, long
18. Stigma: 0¼ simple, capitate, or subcapitate (to slightly bilobed); 1¼bilo
19. Fruit type: 0¼ capsular; 1¼ baccate
20. Pollen when released: 0¼monad; 1¼ tetrad; 2¼polyad
21. Pollen aperture: 0¼ colpi; 1¼ colpori; 2¼ pori
22. Secoiridoid biosynthesis end-product: 0¼ sweroside; 1¼ swertiamarine;
23. Xanthone O-glycosides: 0¼ absent; 1¼ present
24. Xanthone 3-oxygenated: 0¼ absent; 1¼ present
25. Xanthone 4-oxygenated: 0¼ absent; 1¼ present
26. Xanthone 5-oxygenated: 0¼ absent; 1¼ present
27. Xanthone 6-oxygenated: 0¼ absent; 1¼ present
cases literature resources were used (M�esz�aros et al.,2002, with further additions and modifications).
2.8. Biogeographic reconstructions
During the past decade, several analytical biogeo-
graphic methods have become available to test alterna-
tive biogeographic hypotheses (Morrone and Crisci,
1995). Dispersal biogeography represents an approachthat emphasizes dispersal from source areas (ancestral
area or center of origin), by coding areas as a multistate
character and optimizing states on a taxa cladogram,
using either the Camin–Sokal or Fitch parsimony cri-
terion (Bremer, 1992; Maddison et al., 1992; Morrone
and Crisci, 1995). In contrast to dispersalism, vicariance
biogeography assumes a correspondence between taxo-
nomic phylogeny and area relationships and allows theconstruction of area cladograms, showing hypotheses of
historical relationships between areas (van Veller et al.,
2001). Lastly, dispersal–vicariance analysis (Ronquist,
1997) is a compromise method that permits both dis-
persal and vicariance hypotheses. This approach, im-
plemented in DIVA (Ronquist, 1996), reconstructs
ancestral distribution in a given phylogeny without any
a priori assumptions about area relationships, minimizesdispersal and extinction and allows reticulate relation-
ships among areas (Ronquist, 1997).
The historical biogeography of the Chironieae was
inferred using character optimization methods based
cter states used in this study
utescent herb; 2¼ annual or biennial herb
rs; 3¼ solitary
-merous
erous
lar) to campanulate; 2¼ salver-shaped (hypocrateriform) or tubular
ecial (disk?)
sted
se and the mouth of the corolla tube; 2¼near the mouth of the corolla
bed, the lobes well-separated; 2¼decurrent
2¼ gentiopicroside

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 959
either on dispersalism (ancestral areas method; Bremer,1992; Fitch optimization; Maddison et al., 1992) or on
dispersal vicariance approach (DIVA; Ronquist, 1997).
Unlike vicariance-based methods, character optimiza-
tion methods allow the reconstruction of ancestral dis-
tributions without the constraint of an area-cladogram
(Bremer, 1992; Maddison et al., 1992).
Bremer�s ancestral area method was performed with
PAUP, by considering each area as a binary character,and optimizing the number of gains and losses for each
character on the taxon cladogram, using the Camin–
Sokal parsimony criterion.
Fitch optimization was achieved with MACCLADE,
on a data matrix with geographic areas coded as a single
multistate character. In this method, polymorphism is
restricted to terminal taxa (i.e., widespread species)
whereas ancestors are reconstructed as monomorphic(occurring in a single area).
For DIVA, the data matrix was constructed by
scoring the taxa for presence or absence in each area.
DIVA method is similar to Fitch optimization but al-
lows the treatment of widespread ancestors (Ronquist,
1997). Since the DIVA 1.1 computer program requires a
limited number of taxa and a fully resolved cladogram,
the reference 53-taxon matrix was reduced to a 25-taxonmatrix, joining the sister species occurring in the same
area into one terminal component. The polytomy en-
countered in Zeltnera (Section 3) was arbitrary resolved
by grouping the Z. multicaulis clade with the Z. martinii
clade. Alternative solutions gave the same results (not
shown). DIVA optimizations were then conducted with
either an unrestricted number of areas assignated to
each node, or with a number restricted to two (for most
Table 3
Alignment and sequence characteristics of the different nrDNA and cpDNA
Characteristics ITS data set (ITS1/ITS2) trn
Sequence length range (bp) 398–469 682
(222–238/194–247) (37
Sequence length mean (bp) 457 778
(230/227) (40
G+C content (range%) 55.5–70.3 33.
(54–71/56–71) (34
G+C content (mean%) 62.2 34.
(61/63) (36
Aligned sequence length (bp) 507 114
(254–257) (60
Parsimony informative characters 311 224
(158/153) (85
Constant characters 136 787
(68/68) (44
Uninformative characters 64 134
(28/36) (75
Most-parsimonious trees (N) 48 30
Tree length 1428 548
CI 0.46 0.7
RI 0.78 0.8
RC 0.36 0.7
of the taxa investigated do not occur in more than twoareas).
The geographic areas defined for these analyses were:
the Mediterranean (A), southern Africa (including the
Cape region (B), Australia (C), Western North America
(D), Eastern North America (E), Central America (F),
South America (G), and Southeastern Asia (H).
3. Results
3.1. DNA sequence variation
The length of the unaligned ITS1 and ITS2 sequences
ranged between 222–238 and 194–247 base pairs (bp),
respectively. The GC content varied from 54–71%
(ITS1) to 56–71% (ITS2), with respective mean values of61% (ITS1) and 63% (ITS2). The alignment of 80 ITS
sequences produced a matrix of 507 characters with 311
(61.3%) of these being parsimony informative. The
boundaries of ITS regions were identified based on a
comparison with reference sequences of Gentiana frigida
ITS1 and ITS2 (GenBank Accessions Nos. Z48053 and
Z48084). One region covering 12 ambiguous positions
was poorly alignable and was experimentally excludedfrom all analyses. For this exclusion did not affect the
results (not shown), all the characters were kept for
parsimony statistics calculation (Table 3).
In the trnLF region, sequence length ranged from 682
to 873 bp (378–490 bp for trnL intron and 290–451 bp
for trnL–F spacer), with minimum and maximum GC
values of 33.5 and 36.9%, respectively (Table 3). The
limits of the trnL intron and trnL–F spacer was defined
regions investigated in the present study
LF data set (trnL intron/trnL–F spacer) Combined data set
–873 831–1341
8–490/290–451)
1189
4/374)
5–36.9 43.9–51.3
–38/31–37)
3 45.4
.2/33.2)
5 1656
7–538)
507
/139)
962
7/340)
187
/59)
20
1890
9 0.56
8 0.77
1 0.43

960 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
by comparison with Gentiana frigida (GenBank Acces-sion Nos. X77883 and AJ315277, respectively). This
matrix included 75 sequences and 1145 aligned charac-
ters, of which 224 (20%) were phylogenetically infor-
mative. Most of the indels necessary for the matrix
alignment do not contain any phylogenetic information.
Nevertheless, some indels appear to be informative for
the tribe Exaceae (position 337–462 in Exacum and 363–
462 in Sebaea) or the subtribe Coutoubeinae (position567–583).
A summary of the main characteristics for the re-
spective regions is presented in Table 3.
3.2. Phylogenetic analyses
3.2.1. MP analysis of the ITS data set
Heuristic searches performed on the ITS matrix re-sulted in 48 most parsimonious (MP) trees of 1428 steps,
with a CI of 0.46, and a RI of 0.78. One arbitrary chosen
cladogram is shown in Fig. 1, where the sign ‘‘Ø’’ in-
dicates the nodes that collapse on the strict consensus
tree. Jackknife values (J) for node supports are indicated
below the branches (when equal to or greater than 50%).
The ITS strict consensus tree is well resolved, with good
branch support for most of the tribes and subtribesdefined by Struwe et al. (2002). Nevertheless, the
monophyly of the tribe Chironieae is not supported
(branch collapsing on the ITS strict consensus tree). Yet,
the respective subtribes are well delimited, with high
jackknife values for the Canscorinae (J ¼ 100) and
Coutoubeinae (J ¼ 100), but weak support for Chiro-
niinae (J < 50). Within the subtribe Chironiinae, all the
species of Centaurium (except C. cachanlahuen) are dis-tributed into four independent, well-separated clades,
each well-supported and here named C1 (J ¼ 87), C2
(J ¼ 100), C3 (J ¼ 99), and C4 (J ¼ 90; see Fig. 1).
Some clades are well supported, such as (Blacksto-
nia+ Ixanthus; J ¼ 100), (Orphium+Chironia+Centau-
rium cachanlahuen; J ¼ 78), (Sabatia+Centaurium clade
C2; J ¼ 71), and (Exaculum+Centaurium clade
C3+Centaurium clade C4; J ¼ 100). Finally, themonophyly of several additional genera in the Chiro-
niinae is established by having maximum node support
of J ¼ 100 (e.g., Blackstonia, Cicendia, Eustoma, and
Sabatia).
3.2.2. MP analysis of the trnLF data set
Phylogenetic analyses performed on the trnLF data
set, under the MP criterion, gave 30 MP trees of 548steps (CI¼ 0.80, RI¼ 0.89). One MP phylogram is
shown in Fig. 2 (with jackknife branch support). In-
ferred trees from the trnLF sequences analyses are
generally well resolved at the intergeneric level, and
usually received good branch support. The groups
identified on the ITS cladogram such as subtribes
Canscorinae (J ¼ 99), Coutoubeinae (J ¼ 91), or the
(Blackstonia+ Ixanthus) clade (J ¼ 99), are also stronglysupported by the cpDNA analysis. Furthermore, tribe
Chironieae and subtribe Chironiinae received significant
branch support (J ¼ 100 and J ¼ 97, respectively). The
polyphyly of Centaurium is partially confirmed, with
high jackknife values for the clades C1 (J ¼ 71, but
excluding C. maritimum), C3 (J ¼ 96), and C4 (J ¼ 96),
respectively, but not for the unresolved clade C2. The
intergeneric relationships within the Chironiinae arerather poorly solved in the trnLF result (Fig. 2). Some
clades depicted by the ITS analysis such as (Chiro-
nia+Orphium+Centaurium cachanlahuen), (Gyran-
dra+Sabatia), or (Exaculum+Schenkia+clade C4) are
not resolved in the trnLF strict consensus tree.
3.2.3. Assessment of congruence
The partition homogeneity test, performed on acombined data set of 65 sequences (with constant and
ambiguous characters excluded), indicate incongruence
between the ITS and trnLF data sets (p ¼ 0:01). Nev-
ertheless, a comparison of the cladograms obtained for
each separate partition suggests a general congruence in
topologies and branch supports. In both the ITS and
trnLF analyses: (1) subtribes Canscorinae, Chironiinae,
and Coutoubeinae are monophyletic; (2) most of thegenera, within the Chironiinae, received quite good
branch support; and (3) Centaurium s.l. is polyphyletic.
Thus, the rejection of the ILD test may be due to
minor conflicts in species placement. Some sister rela-
tionships observed on the ITS cladogram such as (C.
pulchellum+C. maritimum) and (Gyrandra tenuifo-
lia+G. brachycalyx) are not supported by the trnLF
topology (Figs. 1 and 2). The latter argues for two al-ternative clades, (C. pulchellum+C. tenuiflorum) and (G.
tenuifolia+Sabatia dodecandra), respectively. The ex-
clusion of these species in the ILD test increases the
support for congruence between the data set
(P ¼ 0:949).
3.2.4. Combined MP and ML analyses
Once the conflicting species have been detected, wecombined the two data sets in a ‘‘total evidence’’ ap-
proach (Kluge, 1989; Fig. 3). Sequences of the combined
region range from 831 to 1341 bp, with a mean GC
content of 45.4%. The combined matrix of 65 taxa
comprised 1656 aligned characters with 507 (ca. 31%)
being parsimony informative.
Cladistic analyses performed on the combined matrix
resulted in 20 MP trees of 1890 steps, with CI¼ 0.56 andRI¼ 0.77 (Fig. 3). The strict consensus tree is similar to
the ITS one, with generally a better resolution and
branch support, except for some unresolved clades such
as (Centaurium+Chironia) or (Exaculum+Schen-
kia+Zeltnera). This lack of resolution may be the
consequence of weak conflict between the nrDNA and
plastid data sets, in the relationships of these taxa.
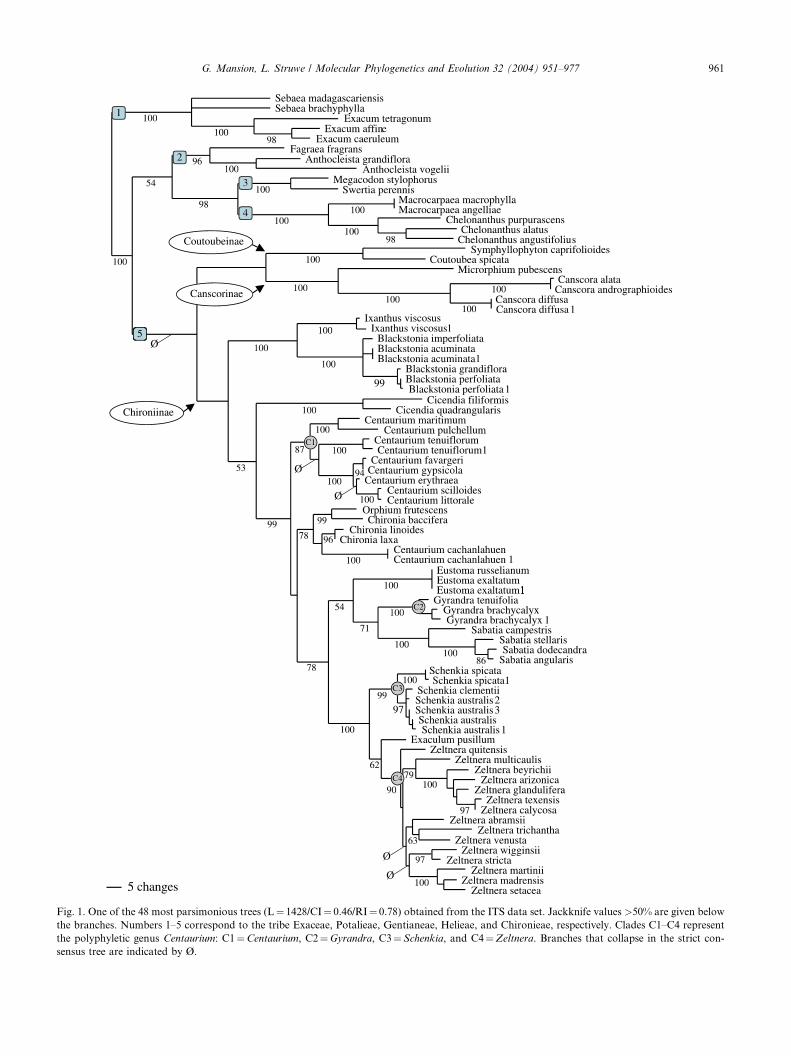
Fig. 1. One of the 48 most parsimonious trees (L¼ 1428/CI¼ 0.46/RI¼ 0.78) obtained from the ITS data set. Jackknife values >50% are given below
the branches. Numbers 1–5 correspond to the tribe Exaceae, Potalieae, Gentianeae, Helieae, and Chironieae, respectively. Clades C1–C4 represent
the polyphyletic genus Centaurium: C1¼Centaurium, C2¼Gyrandra, C3¼Schenkia, and C4¼Zeltnera. Branches that collapse in the strict con-
sensus tree are indicated by Ø.
G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 961

Fig. 2. One of the 30 most parsimonious trees (L¼ 548/CI¼ 0.79/RI¼ 0.88) obtained from the trnLF data set. Jackknife values >50% are given
below the branches. Numbers 1–5 correspond to the tribe Exaceae, Potalieae, Gentianeae, Helieae, and Chironieae, respectively. Clades C1–C4
represent the polyphyletic genus Centaurium: C1¼Centaurium, C3¼Schenkia, and C4¼Zeltnera. Clade C2 is not supported. Branches that collapse
in the strict consensus tree are indicated by Ø.
962 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977

Fig. 3. One of the 20 most parsimonious trees (L¼ 1890/CI¼ 0.56/RI¼ 0.77) obtained from the combined (ITS+ trnLF) data set. Jackknife values
>50% are given below the branches. Numbers 1–5 correspond to the tribe Exaceae, Potalieae, Gentianeae, Helieae, and Chironieae, respectively.
Clades C1–C4 represent the polyphyletic genus Centaurium: C1¼Centaurium, C2¼Gyrandra, C3¼Schenkia, and C4¼Zeltnera. Branches that
collapse in the strict consensus tree are indicated by Ø.
G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 963
964 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
Finally, hard conflicting species, responsible of the ILDfailure, are resolved as in the ITS topology.
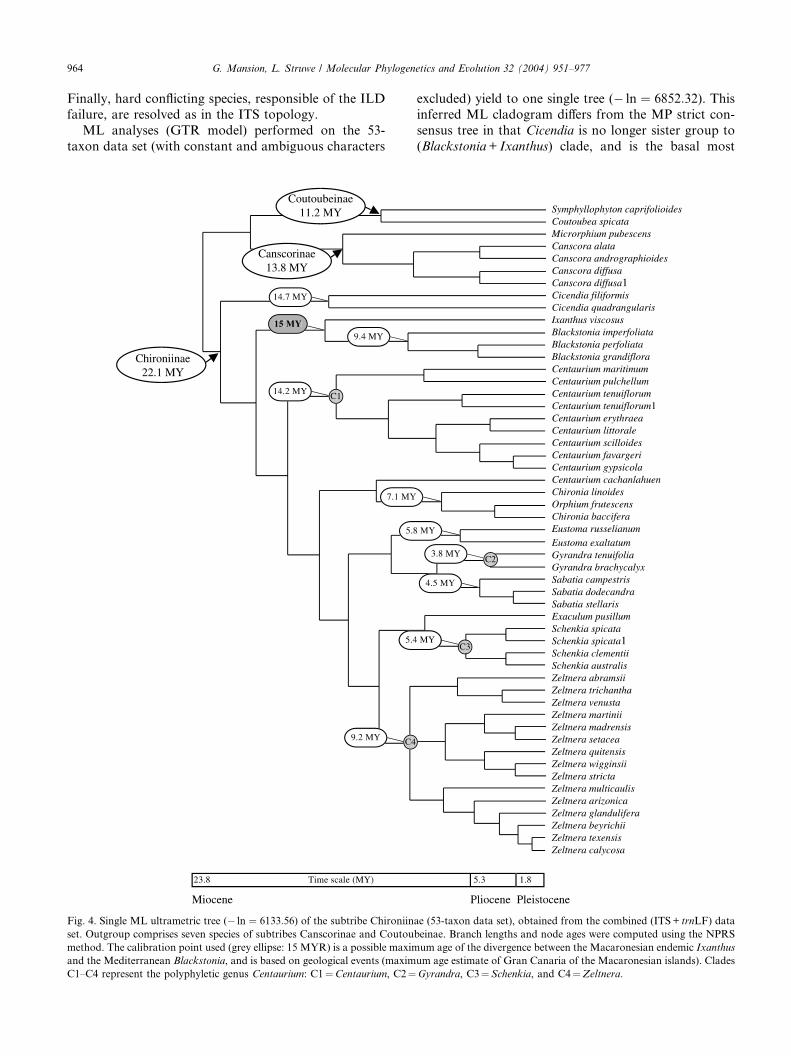
ML analyses (GTR model) performed on the 53-
taxon data set (with constant and ambiguous characters
Fig. 4. Single ML ultrametric tree (� ln ¼ 6133:56) of the subtribe Chironiin
set. Outgroup comprises seven species of subtribes Canscorinae and Coutou
method. The calibration point used (grey ellipse: 15 MYR) is a possible maxim
and the Mediterranean Blackstonia, and is based on geological events (maxim
C1–C4 represent the polyphyletic genus Centaurium: C1¼Centaurium, C2¼
excluded) yield to one single tree (� ln ¼ 6852:32). Thisinferred ML cladogram differs from the MP strict con-
sensus tree in that Cicendia is no longer sister group to
(Blackstonia+ Ixanthus) clade, and is the basal most
ae (53-taxon data set), obtained from the combined (ITS+ trnLF) data
beinae. Branch lengths and node ages were computed using the NPRS
um age of the divergence between the Macaronesian endemic Ixanthus
um age estimate of Gran Canaria of the Macaronesian islands). Clades
Gyrandra, C3¼Schenkia, and C4¼Zeltnera.

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 965
genus in the subtribe Chironiinae (Fig. 4). Only onepolytomy have been encountered in Zeltnera (Node C4).
3.3. Molecular dating
By enforcing a molecular clock on the 53-taxon data
set, we obtained a ML tree with a log-likelihood score of
6908.83. Further comparison with the non-clock model
(phylogram with a score of 6852.32) suggested therejection of the MC assumption (LRT¼ 113,
df ¼ 51; P < 0:01). The respective branch lengths were
thus calibrated using the clock-independent NPRS
method, with the Blackstonia+ Ixanthus node fixed to
Fig. 5. Character mapping of morphological characters on the fully resol
polytomy in Zeltnera has been resolved arbitrary: see text). Character states a
text. Illustrated species examples are noted in brackets after each character st
1¼Perennial herb [Centaurium scilloides, scale bar¼ 1 cm]; 2¼Annual herb
(0¼Terete, not winged stem [Blackstonia perfoliata]; 1¼Quadrangular, not
Scale bar¼ 5mm). 3. Corolla color (0¼Yellow corolla [Cicendia filiformis
merosity (0¼Polymerous corolla [Blackstonia perfoliata]; 1¼Pentamerous c
lum]. Scale bar¼ 1 cm). 5. Corolla shape (0¼Corolla rotate [Sabatia camp
shaped [Zeltnera trichantha]. Scale bar¼ 1 cm). Arrows represent the length
twisted (not illustrated); 1¼Recurved anther (a. Eustoma, b. Sabatia); 2¼H
Stamen insertion (0¼Stamens inserted between the base and the mouth
brachycalyx]. Scale bar¼ 5mm). Arrows show the respective stamen insertio
ovary [Gyrandra pauciflora]; 2¼Elliptic and long ovary [Zeltnera maryanna
shape (0¼ Stigma simple, capitate or subcapitate [e.g., Zeltnera]; 1¼ Stigma b
The arbitrarily chosen character corolla color (3) is optimized on the cladog
15 million years ago (MYA). By using this calibration,the maximum age of the tribe Chironieae and subtribe
Chironiinae was estimated to the early Miocene (23.3
and 22.1 MY, respectively, Fig. 4).
3.4. Phenotypic characters variation
Among the 27 phenotypic characters investigated
(Table 2), only a few appeared to contain some phylo-genetic information when optimized on the 53-taxon
cladogram (Fig. 5). The phylogenetic importance and
evolution of these nine selected characters will be
discussed.
ved ML cladogram obtained from combined 53-taxon data set (the
nd illustrations of the main morphological features are discussed in the
ate. 1. Life form (0¼Tree or shrub [e.g., Fagraea sp., scale bar¼ 2m];
[Zeltnera martinii, scale bar¼ 1 cm]). 2. Stem shape in cross-section
winged stem [Zeltnera glandulifera]; 2¼Winged stem [Canscora alata].
]; 1¼Pink corolla [Exaculum pusillum]. Scale bar¼ 1 cm). 4. Corolla
orolla [Eustoma exaltatum]; 2¼Tetramerous corolla [Exaculum pusil-
estris]; 1¼Corolla funnel-shaped [Exacum affine]; 2¼Corolla salver-
of the corolla tube. 6. Anther shape at anthesis (0¼Not recurved or
elically twisted anther (c. Chironia, d. Zeltnera). Scale bar¼ 1mm). 7.
[Zeltnera venusta]; 1¼Stamens inserted near the mouth [Gyrandra
ns. 8. Ovary shape (0¼Globular ovary [Chironia baccifera]; 1¼Oval
and Centaurium rhodense, respectively]. Scale bar¼ 5mm). 9. Stigma
ilobed, the lobes well-separated [e.g., Centaurium]. Scale bar¼ 20mm).
ram (Fitch and ACCTRAN optimization).

Table 4
Results of the ancestral area analysis of the Chironiinae
Area Gain Loss G/L (%) Coefficient
Mediterranean basin 5 6 83.4 1.00
Western North America 4 7 57.1 0.68
Central America 5 10 50 0.60
South America 3 7 42.9 0.51
Southern Africa 1 5 20.0 0.24
Eastern North America 1 7 14.3 0.17
Australia 1 8 12.5 0.15
966 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
Most of the vegetative and floral characters (e.g., type
of inflorescence, occurrence of calycine colleters or floral
nectaries, petal fusion) are highly homoplastic. On the
other hand, palynological (pollen release and type of
aperture) and phytochemical (secoiridoid biosynthesis)
features were poorly investigated at the tribal level, andgenerally remained uninformative.
3.5. Biogeographic reconstruction
Both the ancestral areas method (Bremer, 1992) and
Fitch optimization (Maddison et al., 1992), performed
on 53-taxon data set, support a Mediterranean origin
for the subtribe Chironiinae (Table 4, Fig. 6).DIVA analyses resulted in multiple combination of
optimal reconstructions. Constraining the number of
maximal areas to two resulted in a unique Mediterra-
nean ancestral distribution for the subtribe Chironiinae
(instead of a widespread ancestral distribution in the
unconstrained analysis; see Fig. 4). Overall, DIVA
analyses require either 11 (unconstrained analysis) or 13
(constrained analysis) dispersal events, and suggest arather complicated biogeographical history for the
subtribe. One possible scenario involved several dis-
persal from the Mediterranean to South Africa, Aus-
tralia, or the Americas (Fig. 6).
4. Discussion
4.1. Utility, limits, and congruence of the molecular data
sets
Sequences of the ITS and trnLF regions have pro-
vided good results in previous cladistic analyses per-
formed on the Gentianaceae at both generic and specific
levels (Chassot et al., 2001; Gielly and Taberlet, 1996;
Thiv et al., 1999a,b; von Hagen and Kadereit, 2001;Yuan and K€upfer, 1995; Yuan et al., 1996, 2003). The
current study confirms the utility of the ITS sequences to
resolve interspecific relationships within the Gentiana-
ceae, and particularly in the Chironiinae. In general,
strict consensus trees show well-resolved groups, and
branch support is high. On the other hand, trnLF
sequences evolve slower than the ITS region, but theyprovide good resolution at the generic level. Yet, most of
the intergeneric relationships within the Chironiinae,
inferred with ITS sequences are supported by the trnLF
data set.
Nevertheless, interspecific relationships are poorly
resolved on the trnLF cladograms, and may result in
soft incongruence between the respective data sets
(Wendel and Doyle, 1998). Pruning taxa with low res-olution on the trnLF cladogram (e.g., C. maritimum or
C. cachanlahuen, Fig. 2) do not change the results of the
ILD test (results not shown). Alternatively, two con-
flicting nodes (with J > 70) have been identified in the
respective ITS and trnLF cladograms as source of hard
incongruence. Their exclusion of the combined data set
resulted in the acceptation of the ILD test.
It is likely that some of the taxa present in the con-flicting clades have separate histories for the nuclear
(ITS) and the plastid (trnLF) genomes. Thus, evolu-
tionary (reticulate) processes known to perturb cladistic
reconstruction in plants, such as hybridization, intro-
gression, lineage sorting, gene duplication or interlocus
concerted evolution may be considered to explain the
present phylogenetic incongruence (Doyle, 1992; Wen-
del and Doyle, 1998).In Centaurium, topological incongruence between
phylogenetic cladograms inferred from ITS and trnLF
data sets have proven to be good tools for estimation of
reticulate evolution and detection of allopolyploidy
(Mansion et al., unpublished). Several examples of in-
terspecific hybridization involving, e.g., C. maritimum,
C. tenuiflorum, or C. pulchellum have been documented,
based on karyological grounds (Zeltner, 1970), andrecently confirmed with molecular tools (Mansion
et al., unpublished). Regarding the importance of in-
terspecific gene flow within Centaurium, hypothesis of
nuclear introgression could be evoked for the mor-
phologically distinct species C. maritimum and C. pul-
chellum that have a sister relationship with the ITS tree,
but divergent positions in the trnLF cladogram (Figs. 1
and 2).Introgressive hybridization is an important trend in
plant evolution known to contribute in intraspecific
variation (Anderson, 1948). Introgression have been
reported in Sabatia (Bell and Lester, 1978), and artificial
hybridization also revealed a high intersectional cross-
ability between species (Perry, 1971). Since cytoplasmic
introgression is not uncommon in taxa capable of in-
terspecific hybridization (Rieseberg, 1995), the sisterrelationships depicted between Gyrandra tenuifolia and
Sabatia dodecandra (Fig. 2) suggest possible episodes of
chloroplast capture. In this case, one explanation for its
occurrence may be long-distance dispersal since Gyran-
dra tenuifolia occurs in Mexico (Broome, 1973), and
Sabatia dodecandra is encountered from Connecticut to
Louisiana (Wilbur, 1955).

Fig. 6. Biogeographic patterns in the Chironiinae. (A) DIVA reconstruction mapped on the fully resolved ML cladogram obtained from combined
53-taxon data set (the polytomy in Zeltnera has been resolved arbitrary: see text). The outgroup includes seven species of subtribes Canscorinae and
Coutoubeinae. Letters A to H are the defined geographic areas for the analysis: A¼Mediterranean basin and surrounding areas, B¼ Southern
Africa, C¼Australia, D¼Western North America, E¼Eastern North America, F¼Central America, G¼ South America, H¼Southeastern Asia
Letters in parentheses are DIVA optimization with number of areas constrained to two. Branch patterns are Fitch optimization (ACCTRAN) of the
character ‘‘repartition’’ on the cladogram. (B) A possible scenario for the biogeographic history of the Chironiinae (dashed lines¼ dispersal; full
lines¼ vicariance). Dispersalism methods and DIVA (maxareas¼ 2) argue for a Mediterranean origin (A) for the Chironiinae, with a primary
dispersal into South Africa (B: Chironia, Orphium) and South America (G: Centaurium cachanlahuen, Cicendia quadrangularis), followed by several
later episodes of colonization into North America (D,E,F) or Australia (C).
G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 967

968 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
4.2. Generic delimitation of Centaurium s.l. and system-
atic implications
One of the most unexpected results of this study is the
polyphyly of Centaurium. This genus has always been
regarded as a morphologically well-delimited assem-
blage by most authors of systematic treatments (e.g.,
Broome, 1973; Gilg, 1895; Grisebach, 1839; Zeltner,
1970). Our molecular analyses depict four major clades:clade C1 which mainly clusters the Eurasian species of
Centaurium; clades C2 and C3, comprising some Mexi-
can species, Mediterranean and Australian ones, re-
spectively; and clade C4 which includes the remaining
native American centauries.
In the early classifications of Centaurium, established
by Schmidt (1828), Grisebach (1839, 1845), and Gilg
(1895), no segregation between Old World and NewWorld species of centauries was undertaken. Only
Grisebach (1845, 1853) placed one species from Mexico
in his new genus Gyrandra (G. chironioides), and a spe-
cies from Hawaii in another new genus Schenkia (S.
sebaeoides). In Gilg�s (1895) taxonomic treatment, he
presented a classification based mainly on pollen char-
acters, and this classification was used for over a cen-
tury. He recognized the genus Erythraea (withCentaurium and Gyrandra in synonymy) and placed it in
subtribe Erythraeinae, close to Sabatia. The classifica-
tion of Struwe et al. (2002) combined phylogenetic ap-
proaches and traditional data, and close affinities were
proposed between Centaurium, Sabatia, and Chironia.
Even if such relationships had been previously advo-
cated (Broome, 1973; Weaver and R€udenberg, 1975;
Wilbur, 1955), the apparent polyphyly of Centaurium
has never been proposed earlier. Similar cases, where
molecular analyses do not support an apparent mor-
phological homogeneity, have been reported in the
Polemoniaceae within the genus Linanthus (Bell and
Patterson, 2000). In that genus, which has mainly di-
verged in response to the spreading drought that oc-
curred in California during the Pliocene, no clear
morphological characters were available to segregatebetween the different molecular-based clades. Several
other examples may be cited as well, particularly Lotus
(Fabaceae), which is a group showing a strikingly sim-
ilar geographical distribution to Centaurium, and in
which two very distinct North American and Eurasian
clades have been depicted by ITS analyses (Allan and
Porter, 2000).
This polyphyly not only disrupts the infragenericclassification of the group, but intergeneric relationships
as well. A new classification has recently been proposed
to resolve the problem of the polyphyly of Centaurium
(Mansion, in press). In this classification, several taxo-
nomic changes are made as a result of the findings in this
paper. Clade C1 represents the Eurasian species of the
genus Centaurium Hill, and the �true� Centaurium. The
genera Gyrandra and Schenkia, both described byGrisebach (1845, 1853), on the basis of G. chironioides
(¼Centaurium chironioides) and S. sebaeoides (¼Cen-
taurium sebaeoides), are resurrected and extended to
include additional species. Finally, a new genus Zeltnera
was proposed for the remaining American species cor-
responding to clade C4 (Mansion, in press). This no-
menclatural treatment, dividing Centaurium into four
genera, is followed in the present discussion.
4.3. Phylogenetic relationships within the Chironiinae
This study, based on ITS and trnLF data sets, sup-
ports the monophyly of the tribe Chironieae, and con-
firms the subtribes Canscorinae and Coutoubeinae as
sister clades with the Chironiinae, as suggested by
Struwe et al. (2002). The relationships described below,unless otherwise stated, are based on the ITS cladogram
that allows a better resolution at the specific level
(Fig. 1).
4.3.1. The (Blackstonia + Ixanthus) clade
A close relationship between Blackstonia and Ixan-
thus was suggested by Anderson (1948), Grisebach
(1839), and more recently by Thiv et al. (1999a).Blackstonia comprises four annual, diploid or tetraploid
species of mainly Mediterranean distribution (Zeltner,
1970). The monotypic Ixanthus, one of 28 endemic
genera of the Canary Islands (Kunkel, 1993), is a pe-
rennial, basally woody, and sparsely branched herb,
which reaches a height of up to 2m. These genera share
some morphological features such as the presence of
perfoliate bracts, calycine colleters, and a yellow corolla.A more detailed comparison, including anatomy–his-
tology, morphology, karyology, and ecological charac-
ters, was presented by Thiv et al. (1999a). In our study,
the relatively basal position and sister group relationship
between Blackstonia and Ixanthus are reinforced.
4.3.2. Large phylogenetic distance between Cicendia and
Exaculum
The phylogenetic position of Cicendia within the
Chironiinae remains problematic. MP analyses suggest
an intermediate placement of Cicendia between the basal
(Blackstonia+ Ixanthus) clade and the remaining genera
(Figs. 1 and 3) whereas the inferred ML cladogram
supports a basal most position of Cicendia within the
subtribe (Fig. 4). Earlier hypotheses of close affinities
between the Mediterranean genera Cicendia and Exac-
ulum (Grisebach, 1839), and to a lesser extent with the
Mexican genus Geniostemon (Thiv and Kadereit, 2002a),
are not supported by our molecular analyses (Fig. 2). All
the inferred cladograms (Figs. 1–3) reveal a large phy-
logenetic distance between Cicendia and Exaculum.
These genera share a similar morphological appearance
(slender herbs, sparsely branched, with minute flowers

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 969
and untwisted anthers after pollen release) and an affinityfor the same type of open habitats (Favarger, 1960). In
contrast toCicendia, the flowers ofExaculum are pink and
possess a short calyx tube with the calyx lobes exceeding
the corolla tube in length, and the corolla tube is longer
than the corolla lobes (vs. equal length in Cicendia). In
Cicendia, the stigma is subcapitate whereas Exaculum
possesses a bilobed stigma (Vald�es et al., 1987). Lastly, thechromosome number differs between these taxa, withn ¼ 13 for Cicendia filiformis and n ¼ 10 for Exaculum
pusillum (Favarger, 1960).
The present cladistic analyses support phylogenetic
affinities of Exaculum with either Zeltnera (ITS, Fig. 1)
or Schenkia (trnLF, Fig. 2), or an unresolved position
(combined analysis, Fig. 3). Schenkia comprises plants
with spicate, cymose inflorescences, centered in the
Mediterranean basin, but is also found in Russia, Japan,Australia, and on the Hawaiian Islands. Molecular,
morphological (spike-like cyme inflorescence, subsessile
flowers, and subcapitate stigma) and karyological
(n ¼ 11) evidence support the generic identity of
Schenkia and argue against the inclusion of Exaculum in
it. The genus Zeltnera also differs from Exaculum in
several aspects such as the chromosome number
(n ¼ 17; n ¼ 20; n ¼ 21; and n ¼ 22; Mansion andZeltner, in press), stamen insertion (in the upper half of
the corolla tube in Zeltnera vs. in the throat of the co-
rolla in Exaculum), coiling of the anthers (absent in
Exaculum), and geographic repartition (North and
Central America for Zeltnera). Thus, molecular, kary-
ological and morphological data support the recognition
of a monospecific genus Exaculum.
4.3.3. The South African (Chironia +Orphium) clade and
Centaurium
Most species of Chironia (ca. 30 species) are found in
grassy places of southern Africa, mainly in the Fynbos
vegetation of the Cape region, where the endemic
monotypic genus Orphium also occurs. Orphium is a
suffrutescent herb, which closely resembles the pubes-
cent species of Chironia, such as C. baccifera. Fewmorphological differences (mainly the keeled calyx of
Orphium) discriminate between the two genera; hence,
all the generic characters described for Orphium are also
encountered in Chironia. Both ITS and trnLF support a
close relationship between Orphium and Chironia bac-
cifera (Figs. 1–3), suggesting either a broader circum-
scription of Chironia (including Orphium) or a
paraphyletic genus Chironia.Phylogenetic analyses suggest a derived position of
Chironia and Orphium compared to the Mediterranean
genus Centaurium (Figs. 1 and 4), or failed to resolve the
position of the respective clades (Fig. 3). Chironia and
Orphium share morphological features with Centaurium
such as pink to purple corollas (rarely white or yellow)
and anthers coiling after dehiscence (Marais and Verd-
oon, 1963). The main morphological difference betweenthese genera is the corolla shape, i.e., rotate to funnel-
shaped with the corolla tube shorter than the corolla
lobes in Chironia vs. salver-shaped, with the corolla tube
equaling or exceeding the corolla lobes in Centaurium.
Furthermore, perennial species with a basally woody
stem are missing in Centaurium, but frequently en-
countered in Chironia. It is noteworthy that the Cape
flora has a surprisingly low proportion of annuals (ca.7% of the species) compared to other regions with
Mediterranean climates (Goldblatt and Manning, 2000).
Indeed, the Gentianaceae, with about 18 annual species
(mainly Chironia and Sebaea), is one of the families with
the highest number of annuals in the Cape flora
(Goldblatt and Manning, 2000, p. 15).
4.3.4. Position of the South American Centaurium cachan-
lahuen
The unique phylogenetic position ofC. cachanlahuen is
of interest because it groups with Chironia and not with
any other Centaurium species (Figs. 1 and 3), which im-
plies a striking South American–South African disjunc-
tion (see below). This important medicinal herb (Molina,
1787; Schneider, 1974), endemic to the Andean areas of
Chile and Argentina, is frequent from the Pacific littoralto the Andean pre-Cordillera and from the Atacama
Province in the North to the Chilo�e Province in the South(Gunckel, 1924). The gross morphology of C. cachan-
lahuen evokes Zeltnera quitensis. However, the style dif-
fers in being very short (i.e., as long as the stigma lobes)
and the divided stigma much resembles that of Chironia
krebsii (Mansion, personal observations), whereas Zelt-
nera species generally share undivided stigma (Mansion,in press).
Further comparative morphological studies are
planned to resolve the taxonomic status of C. cachan-
lahuen and the relationships with the South American
Chironiinae taxa (Zygostigma australe and Centaurium
ameghinoi) that were not included in the present study
(Mansion and Zeltner, in progress).
4.3.5. Relationships among the (Gyrandra+Sabatia)
clade and Eustoma
The (Gyrandra + Sabatia) clade received strong
branch support in the ITS result (J ¼ 71). Sabatia is a
rather species-rich genus within the Chironiinae (ca. 20
taxa), comprising distinctive taxa classified in several
sections on the basis of morphological and karyological
data and incompatibility among hybrids (Perry, 1971;Wilbur, 1955). The present study includes one species
each from four out of seven sections, and gives pre-
liminary support for the monophyly of Sabatia.
The genus Gyrandra was first described by Grisebach
(1839), and contains five species (Mansion, in press).
The ITS and combined cladograms (Figs. 1, 3, and 4)
support the sister position of Gyrandra to Sabatia,

970 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
whereas trees based on cpDNA characters (Fig. 2) arguefor the inclusion of Gyrandra speciosa in Sabatia.
Eustoma is a small genus occurring in the southern
United States, Mexico, and Greater Antilles with two
species (NatureServe, 2003), E. russellianum (Hook.) G.
Don (including E. grandiflora (Raf.) Shinners) and E.
exaltatum (L.) Salisb. (cf. Shinners, 1957). This genus
was first placed in the tribe Tachiinae (Gilg, 1895), close
to Lisianthius, due to similar pollen features (reticulatepollen).
It is noteworthy that the taxa belonging to the (Eus-
toma+Gyrandra+Sabatia) clade share the chromosome
number of 2n ¼ 72 (with a high dysploid variation in
Sabatia), a larger-sized corolla (except Sabatia arenicola
and Gyrandra brachycalyx) than other Chironiinae, and
stamen insertion near the sinus of the corolla lobes
(Mansion, in press). Moreover, their respective geo-graphic distribution (Caribbean, Mexico and eastern
North America) may support a common ancestral origin
and vicariant patterns. One morphological difference is
the degree of anther coiling after anthesis, being helically
twisted in Gyrandra, slightly coiled in Eustoma and re-
curved to circinnately coiled or half-twisted laterally in
Sabatia. Another difference is stigma shape, which is
subcapitate to slightly bilobed in Gyrandra, bilobed andwell separated in Eustoma, and deeply bilobed and
twisted around each other at anthesis in Sabatia.
4.4. Character evolution in the Chironiinae
The Chironieae have been divided in three subtribes
based on molecular data (Struwe et al., 2002). None-
theless, no morphological cladistic analysis has beenperformed on the whole tribe, except some more general
works covering the Gentianaceae (M�esz�aros et al., 1996,2002). The latter study failed to detect synapomorphic
characters supporting current tribe or subtribe delimi-
tations. This lack of synapomorphies for above-generic
ranks is also noted in this analysis combining molecular
data and structural characters, with some exceptions
where a few distinct characteristics support supragenericgroupings or genera.
4.4.1. Morphological characters
Vegetative characters appear to contain poor, if any,
phylogenetic information at the supergeneric level. Most
of the Chironiinae investigated are annual herbs, and
the suffrutescent habit found in Chironia, Coutoubea,
Ixanthus, Orphium, and Symphyllophyton has developedindependently several times (Fig. 5, character 1). It
seems that the basal woodiness in Ixanthus is best in-
terpreted as secondary, and has either evolved as a
consequence of the perennial life cycle or as an adap-
tation to the laurel forest habitat (Thiv et al., 1999a).
The suffrutescent habit encountered in Chironia and
Orphium could be a derived character, when compared
to the herbaceous condition found in Centaurium. Thus,the cause for woodiness in insular Ixanthus and conti-
nental genera (Chironia and Orphium) may differ. The
stems are mainly quadrangular in cross-section within
the Chironiinae (Fig. 5, character 2), except for the basal
genera (Blackstonia, Cicendia, Ixanthus) and Eustoma,
which are all terete.
Corolla color varies significantly within Chironieae.
Yellow, white or green corollas appear to be a plesio-morphic state (Fig. 5, character 3). The pink to lilac
corollas seem to be synapomorphic for most of the
genera of the Chironiinae, except for the basal Blacks-
tonia, Cicendia, and Ixanthus, and some species of
Centaurium (C. maritimum: yellow corollas) or Sabatia
(white corollas). Most of the pink-flowered genera often
include certain populations with white-flowered indi-
viduals (Mansion, pers. obs.). Pentamerous corollas aremost common within the tribe Chironieae and have been
regarded as the ancestral merosity state for the family
(M�esz�aros et al., 2002). Yet, tetramerous flowers are
also frequent in several genera of Chironieae, either
consistently (e.g., Canscora, Cicendia, Coutoubea,
Cracosna, Exaculum, Hoppea, or Schinziella) or occa-
sionally (e.g., Centaurium, Ixanthus, Schenkia or Zelt-
nera), as well as polymerous ones (Blackstonia, Sabatiasection Dodecandrae, and outside of the Chironieae:
Anthocleista, Potalia, and Urogentias). Our combined
analysis supports the pentamery of the corolla as a
plesiomorphic state for the Chironieae. On the other
hand, the tetramerous flowers appear to be synapo-
morphic for a part of the subtribe Canscorinae (Fig. 5,
character 4), as recently found by Thiv and Kadereit
(2002b), and based on a cladistic analysis of morpho-logical characters. Most of the genera in the tribe have a
salver-shaped corolla. Among the exceptions are
Blackstonia, Gyrandra, and Sabatia, all three with a
rotate corolla, and (Chironia+Orphium) with a funnel-
shaped corolla (Fig. 5, character 5).
An unusual specialization of the androecium is the
coiling of the anther after pollen release, hitherto con-
sidered as a typical characteristic of Centaurium s.l. Withthe generic break up of Centaurium into four separate
genera, this character is no longer a possible synapo-
morphy. Such spiralization or recurvation of the thecae
is also present to various degrees in other genera of the
Chironiinae, such as Chironia, Eustoma, Orphium, and
Sabatia, but is absent within the basal genera (Blacks-
tonia, Cicendia, Geniostemon or Ixanthus) and in Exac-
ulum (Fig. 5, character 6). Within the subtribe, thestamens are basifixed and generally inserted in the upper
half of the corolla tube, except for the (Sabatia+Gyr-
andra+Eustoma) clade where the insertion occurs
higher up in the corolla throat, near the sinuses of the
rotate or funnelform corolla (Fig. 5, character 7).
The gynoecium of the Chironieae is bicarpellate, with
a superior ovary variable in shape (Fig. 5, character 8),

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 971
and surmounted by a style of variable length. The latteris either slightly divided (apically divided in Blackstonia,
twisted in Sabatia, or slightly branched in Centaurium or
Chironia spp.) or oblong-capitate (terminally hooked in
Chironia spp. and Orphium, or subcapitate or slightly
bilobed to funnelform in Gyrandra, Schenkia or Zelt-
nera). The degree of style division is generally a good
discriminating character at the generic level (Fig. 5,
character 9). Another character of importance in thesystematics of the Chironiinae is the stigma morphol-
ogy. This feature is particularly useful to discriminate
between Chironia species (Marais and Verdoon, 1963).
Within the centauries, stigmas with fan forms generally
appear in Zeltnera, whereas Centaurium species mainly
have elliptic or reniform ones (Mansion, in press).
4.4.2. Palynology
Within subtribe Chironiinae, 3-colporate and finely
reticulate pollen grains have previously been reported in
14 species of Centaurium s.l. (K€ohler, 1905). Further-more, similar pollen morphology (defined as ‘‘Centau-
rium pulchellum’’ pollen type) has been encountered in
six species of Centaurium and in Schenkia spicata (Punt
and Nienhuis, 1976). This preliminary pollen study does
not support the distinctive phylogenetic distance be-tween Centaurium and Schenkia as found by us.
Recently, Nilsson (2002) described striate to striato-re-
ticulate pollen grains in both Chironia and Orphium,
thus supporting the possible inclusion of Orphium into
Chironia.
The palynology of Zeltnera has been poorly studied.
Elias and Robyns (1975) reported pollen characters for
Zeltnera quitensis that do not differ much from themorphology found in the ‘‘Centaurium pulchellum’’
pollen type. Nonetheless, variation has been observed
regarding the pollen shape (subprolate in Zeltnera vs.
sphaeroidal in Centaurium) and exine features (lirae
irregularly interlaced vs. lirae parallel to colpi and
sometimes abruptly changing direction; Rao and
Chinnappa, 1983).
Overall, palynological data remain fragmental withinthe subtribe Chironiinae, but some features may con-
stitute possible generic synapomorphies. A palynologi-
cal study that includes all representative taxa is greatly
needed.
4.4.3. Phytochemistry
Phytochemical characters have been used extensively
for taxonomic purposes in the Gentianaceae, and aresometimes helpful to discriminate between tribes, genera
or even sections (Gottlieb, 1982; Hostettmann-Kaldas
et al., 1981; Jensen and Schripsema, 2002). Particular
compounds that are known to occur in aerial parts and
roots of many gentians are xanthones (often yellow) and
secoiridoids (very bitter). The xanthones are biosyn-
thetically of mixed acetate and shikimate origin, and
two main hydroxylation patterns (or ‘‘parent patterns’’)can be found (Carpenter et al., 1969; Hostettmann and
Wagner, 1977; Jensen and Schripsema, 2002). Xant-
hones can be encountered in most of the genera of the
Chironieae, except in some species of Cicendia (C. fili-
formis), Centaurium (C. maritimum) and Sabatia (S.
angularis, S. elliotii; Schaufelberger, 1986). These com-
pounds also occur in some Texan species of Zeltnera
(Mansion and Rodriguez, unpublished data).Diverse oxidation patterns of the parent xanthones
have been recorded in the Gentianaceae, and are not
distributed equally within the family. The variation in
oxidation patterns has been used for cladistic recon-
structions by Gottlieb (1982), van der Sluis (1985), and
M�esz�aros (1994), in which the polarity of this character
has been expounded in different ways. A more satisfac-
tory proposition has been recently suggested by Jensenand Schripsema (2002), who arranged the xanthones of
the Gentianaceae in four biosynthetic groups based on
how �primitive� they are in the biosynthetic chain. In our
analysis, xanthones of group 1, the most primitive state
(1-3-7 and/or 1-3-7-8 substituted xanthones), constitute
a homoplastic state since they are randomly distributed
in five genera within the respective tribes. They appear,
for example, in Blackstonia and Orphium, but not intheir respective sister genera Ixanthus and Chironia, re-
spectively. On the other hand, particular hexa-substi-
tuted xanthones, mainly corresponding to group 4 of
Jensen and Schripsema (2002), are frequent in the Chi-
ronieae and may be a synapomorphy for this group (not
shown).
4.5. Biogeography and evolutionary history of the Chiro-
niinae
Extant species of the Chironieae are distributed in
three main geographic regions. The Canscorinae and
Coutoubeinae are of Paleotropical and Neotropical dis-
tributions, respectively, whereas theChironiinae aremore
frequent in the subtropical to temperate northern hemi-
sphere. The Chironiinae particularly occurs in the WestPalearctic (Blackstonia, Centaurium, Chironia, Cicendia
p.p., Exaculum, Ixanthus, and Schenkia p.p.) or in the
southern part of the Nearctic (Cicendia p.p., Eustoma,
Geniostemon, Gyrandra, Sabatia, and Zeltnera). Some
genera are encountered in the southern hemisphere as
well, mainly in Argentina, Brazil, and Chile (Centaurium
cachanlahuen, Zygostigma australe), South Africa (Chi-
ronia, Orphium), and western Australia (Schenkia p.p.).Our ancestral area analysis strongly supports a
Mediterranean origin for the Chironiinae with sub-
sequent dispersal into the Americas, South Africa, and
Australia (Table 4). Constrained DIVA analyses and
Fitch optimization also argue for a Mediterranean ori-
gin for the subtribe, but with a primary dispersal into
South Africa (Chironia, Orphium) and South America

972 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
(C. cachanlahuen), followed by several episodes of col-onization into North America or Australia (Fig. 6).
Furthermore, DIVA results suggest that the most recent
common ancestor (MRCA) of the (Eustoma+Gyran-
dra+Sabatia) clade first colonized Central America
(giving rise to Eustoma and Gyrandra) and then eastern
North America (becoming Sabatia). Later, a possible
dispersal to Australia would explain the distributional
patterns in the extant genus Schenkia. Coinciding withthis was a large species diversification in Zeltnera in
western North America (Fig. 6). Alternatively, a Medi-
terranean taxon may have evolved locally providing the
MRCA of Schenkia and Exaculum, and the MRCA of
Zeltnera, which further dispersed to western North
America. Based on this, several alternative dispersal
hypotheses can be inferred, suggesting dispersal from
the Mediterranean area to either (1) North America,Central America, and South America, (2) South Africa
and Cape region, or (3) Australia and the Pacific Is-
lands. Particularly disjunct patterns such as the amphi-
tropical repartition of Cicendia quadrangularis, or the
unusual transatlantic connection between Chironia and
C. cachanlahuen will be discussed below.
4.5.1. Connections between regions with Mediterranean
climate
Most of the genera in the Chironiinae are restricted to
either the Mediterranean basin and surrounding areas
(Blackstonia, Cicendia, Centaurium, Exaculum, Ixanthus,
and Schenkia) or to diverse regions with mainly Medi-
terranean climates, such as southwestern North America
(Cicendia, Eustoma, Geniostemon, Zeltnera), South Af-
rica (Chironia, Orphium), Australia (Schenkia) and Chile(C. cachanlahuen). In these latter regions, introduced
Chironieae species from the Mediterranean are also
frequent (e.g., Blackstonia, Centaurium, or Cicendia). A
few genera are restricted to smaller climatic regions,
such as tropical climates with summer rain Gyrandra or
Zeltnera in Mexico, or hot and humid climates (Sabatia
in eastern North America).
Biogeographic patterns inferred from the DIVAanalysis indicate that several migrations to the Americas
have occurred in the Chironiinae. Two major routes are
known to have provided connections between the Old
and New World, namely, the Bering Land Bridge (BLB)
and the North Atlantic Land Bridge (NALB). The first
bridge (BLB) between northwestern North American
and northeastern Asia was present throughout the
Tertiary with repeated formation over the past 20 mil-lion years during the Miocene, Pliocene, and Pleistocene
(Graham, 1999; Tiffney, 1985a; Wen, 1999), and has
permitted many possibilities for exchange of plants and
animals between Eurasia and North America. During
the Miocene, the BLB appears to have been the main
connection for many temperate, deciduous plants cur-
rently present in North America and Asia. However, the
temperatures at these high latitudes were too cool formost herbaceous groups (Tiffney, 1985a). Movements
via the BLB still remained possible by successive colo-
nization of disturbed sites (or alternatively by a southern
Aleutian bridge), or spread by random long-distance
dispersal (Tiffney, 1985a). Considering that many of the
Chironiinae taxa are frequently found in disturbed sites
(e.g., Blackstonia, Centaurium, Schenkia or Zeltnera)
where competition is weak, the first solution appears tobe more likely. Nevertheless, the current lack of Chiro-
niinae in eastern Asia (except Centaurium pulchellum in
China and Schenkia japonica in Japan) provides a weak
support for the BLB hypothesis. The second alternative
bridge (NALB) involved at least four geographical links:
two between North America and Greenland, one be-
tween Fennoscandia and Greenland, and one between
Greenland and the British Isles (McKenna, 1983). Thelatter link broke by the early Eocene (ca. 50 MYA),
whereas some other connections between Eurasia and
North America remained available until the end of the
Eocene (ca. 38 MYA) allowing plant migrations (Tiff-
ney, 1985b; Tiffney and Manchester, 2001). These ranges
are not congruent with current age estimations for the
subtribe Chironiinae (22.1 MY; Fig. 4). Even with a
larger estimation of the divergence time to 30 MY, be-tween Ixanthus and Orphium (based on a matK–trnL
data set; Thiv, unpublished), the NALB does not ap-
pear to have played a major role in the Chironieae
diversification.
The relationship found between the Mediterranean
native Centaurium and the South African genera Chi-
ronia and Orphium (Fig. 3), exemplifies a distributional
pattern known in several other groups of plants (e.g.,genera in the Dipsacaceae, Papaveraceae, and Thym-
eleaceae). Only 4% of the genera encountered in South
Africa are thought to have disjunct affinities with Eur-
asian taxa (Goldblatt, 1978). The direction of these ex-
changes is largely unknown, with some South African
genera being either basal or derived compared to their
Mediterranean counterparts (e.g., Crassulaceae [Mort
et al., 2001] or Colchicaceae [Caujape-Castells et al.,2002]). Our study weakly support a derived position of
Chironia relative to Centaurium (Figs. 1 and 4).
One significant feature of the Tertiary history of Af-
rica is the union of the African-Arabian block with
Eurasia, thereby closing the Tethys Sea, by the mid-
Miocene (ca. 15 MYA; Axelrod and Raven, 1978). Di-
rect migration of the Eurasian flora into Africa became
then possible, and two main routes have been proposed(Quezel, 1978); (1) incursion into central Sahara via the
Ahaggar and Tibesti mountains, and (2) incursion of
Mediterranean woodland vegetation via the Red Sea
hills of Egypt and Somalia (Wickens, 1976). The pres-
ence of Centaurium pulchellum, along with the enigmatic
Monodiella flexuosa Maire (¼Centaurium flexuosum
(Maire) Lebrun & Marais), in the Tibesti Mountains

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 973
(between southern Libya and northern Chad) supportsthis view. A new uplift in East Africa by the Miocene
caused the formation of cooler and drier areas, and
provided additional routes for migration and exchange
between Mediterranean and South African floras. By
the Pliocene and Pleistocene, exchanges with South
Africa seem to be impossible due to a tropical–equato-
rial barrier (Quezel, 1978). Therefore, a probable Mio-
cene origin (5–25 MY) may be proposed for a (Chironia+ Orphium) divergence from a Mediterranean ancestor,
which is congruent with the node age estimation of this
clade using our geological calibration point (7.1 MY;
Fig. 4). By the Miocene, the development of a cold
circum-Antarctic current (around 11–14 MY) favored
the aridification of the Cape region (Richardson et al.,
2001). This was an important factor in initiating the
transformation of the Miocene subtropical forest to theFynbos vegetation of today (Axelrod and Raven, 1978).
The Canarian fauna and flora is related to that of
other Atlantic islands, such as Madeira and, to some
lesser extent, that of the Azores and the Cape Verde
Islands. This flora has affinities mainly with the Medi-
terranean region, although some elements are related to
more remote regions such as South America or Aus-
tralia (Juan et al., 2000). The genus Ixanthus occurs inthe most recent islands of the Canary archipelago such
as (from east to west) Gran Canaria (14–16 MY), Ten-
erife (11.6 MY), La Gomera (10 MY), La Palma (2
MY), and El Hierro (1 MY), where it grows in the un-
derstory of laurel forests (Bramwell and Bramwell,
1990). The genus is absent from the two eastern islands,
Fuerteventura (20 MY) and Lanzarote (15.5 MY).
Relative ages of the respective islands are based on po-tassium argon dating (Juan et al., 2000). As the Canary
Islands are of oceanic origin (Juan et al., 2000), long-
distance dispersal is required for plant colonization.
Thus, one can expect a single introduction of either the
MRCA of the (Blackstonia+ Ixanthus) clade or Ixanthus
on Gran Canaria, followed by a stepwise colonization
sequence from this relatively older to the younger is-
lands in the chain (i.e., from Gran Canaria, to Tenerife,and the more western islands). This type of single col-
onization pattern has been demonstrated for many en-
demic Macaronesian genera, including Argyranthemum
(Francisco-Ortega et al., 1997), Pericallalis (Panero
et al., 1999), or Sonchus (Kim et al., 1996) for the As-
teraceae, Echium (Boraginaceae, Bohle et al., 1996),
Crambe (Brassicaceae, Francisco-Ortega et al., 1999),
Aenium (Crassulaceae, Mes et al., 1996), Bencomia
(Rosaceae, Helfgott et al., 2000), and Sideritis (Lamia-
ceae, Barber et al., 2002). Alternatively, Ixanthus may
have colonized older arid eastern islands such as Fu-
erteventura (20 MY), or arose directly in continental
laurel forests, and became extinct locally due to the
aridification of the habitat or volcanic activity (Juan
et al., 2000).
The presence of some Blackstonia, Centaurium orCicendia species in Australia can be interpreted as an-
thropozoogenic introduction, but an alternative sce-
nario may be proposed for Schenkia. Schenkia is
widespread in damp and sandy places and occurs in all
the Australian states (Adams, 1996). The current dis-
tribution of Schenkia shows that diploid taxa are com-
mon around the Mediterranean basin and radiate
throughout Asia, and tetraploid species are distributedin western Australia, Hawaii, and the Easter Islands
(Zeltner, pers. comm.). A long-distance dispersal of an
ancestral diploid of Mediterranean origin to Australia
and Pacific islands (Hawaii or Easter Islands) remains
possible. The rare endemic Hawaiian species S. sebaeo-
ides is tetraploid (Carr, 1978; Zeltner, pers. comm.), and
morphologically closely related to S. australis. Relative
maximum age estimation within Schenkia is 5.4 MY(Fig. 4), i.e., by late Miocene. The migration of diploid
or tetraploid populations accompanying the invasion of
Asian plants in northeastern Australia, could have been
possible during the late Miocene, by the New Guinea
bridge permitting the junction of the Australian conti-
nent with the Philippine Sea plate (Hill, 1994; Renner
et al., 2001). The occurrence of S. japonica, a morpho-
logically close relative to S. australis (not included in ouranalyses), in Taiwan and Japan supports this view. Such
recent colonization (the end of Miocene to Pliocene) of
the Australasian region has been documented for the
genus Myosotis (Winkworth et al., 2002).
4.5.2. Genera with disjunct repartition
Among 18 North American–European disjunct an-
giosperm genera (Qian, 1999), four are bispecific genera(Cicendia, Corema, Hottonia, and Mespilus). All of them
are disjunct between the western Old World and the
western New World, especially the Mediterranean and
the Madrean regions (Takhtajan, 1986). Likewise, some
species of Centaurium (C. erythraea, C. pulchellum or C.
tenuiflorum) or Schenkia (S. spicata) from Europe, are
naturalized in North America. Node age estimations of
the Cicendia split (ca. 14.7 MY, Fig. 4) indicate a pos-sible introduction to North America by the middle
Miocene, more likely via the BLB. Recent discoveries
suggest that the Neotropical climates were unstable over
the past 2 million years during the Pleistocene (Whit-
more and Prance, 1987). Cyclic glacial events led to
periods of cooler and/or drier climate in which
(rain)forest species may have withdrawn to small refugia
pockets (Haffer, 1982). Other recent geological phe-nomena that have been suggested for driving Neotrop-
ical speciation are the uplift of the northwestern Andes
(ca. 5 MY) and the bridging of Isthmus of Panama,
achieved ca. 3.5 MY (Gentry, 1982). Thus, one can
hypothesize for Cicendia quadrangularis a migration to
South America via the existing Panama land bridge,
between 3.5 and 2 MY. Alternatively, a dispersal event

974 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
from North to South America by long-distance migra-tion, via an intermediate series of appropriate ‘‘way
stations’’ such as Peru or Bolivia (Raven, 1963), remains
probable. Such a pattern has been recently documented
in the amphitropical disjunct genus Larrea (Lia et al.,
2001).
The transatlantic disjunct pattern described between
South African Chironia and New World C. cachanlah-
uen, has been encountered in only four other genera ofthe Gentianaceae, namely Enicostema, Neurotheca
[Potalieae], Schultesia [Chironieae], and Voyria. Several
hypotheses have been proposed, according to the rela-
tive age of the groups investigated, and sometimes
evocating possible Gondwanic vicariance processes or
boreotropic taxon interchange (Struwe et al., 2002). The
last connection between Africa and South America is
estimated to 105 MY, but many of the angiosperm lin-eages have radiated more recently in the Tertiary, and
do not extend back to the Cretaceous. Therefore, vi-
cariant species resulting from the break-up of the wes-
tern Gondwana cannot satisfactory explain these
disjunct angiosperm lineages. Moreover, both conti-
nents continued to be connected via a series of volcanic
islands until the end of the Eocene (ca. 38 MY), when
they were separated by 1400 km and fewer islands re-mained (Raven and Axelrod, 1974). Nevertheless, dis-
persal directly across the Atlantic Ocean (although
perhaps feasible in some cases) seems unlikely to be a
general explanation. An alternative migratory road has
been proposed for some groups, such as the Malpighi-
aceae (Davis et al., 2002). Members of this family have
migrated from South America into North America via
scattered continental or volcanic islands that connectedboth the continent at various times in the Tertiary.
From North America these lineages may have migrated
eastward to Laurasia via a series of connections across
the North Atlantic Land Bridge, subsequently diversi-
fying in Africa (Madagascar), and in some cases in
Eurasia. In addition, the Bering Land Bridge facilitated
the spread of northern hemisphere biota during periods
of climatic warming in the late Paleocene-early Eocene.This pattern is also encountered in Melastomataceae,
and in the opposite direction, in Annonaceae and
Lauraceae. Nonetheless, such explanations may not be
available for more recently disjunct groups such as
(Chironia+C. cachanlahuen).
To summarize, our work based on molecular and
morphological data sets strongly supports the mono-
phyly of the subtribe Chironiinae, with all genera exceptone (Zygostigma) investigated. These results also argue
for a polyphyletic genus Centaurium, split up into four
well-supported clades. From a systematic and biologist�spoint of view, species of Centaurium are best recognized
in four distinct genera, namely Centaurium (ca. 20 spp.),
Gyrandra (5 spp.), Schenkia (c. 5 spp.), and Zeltnera (c.
25 spp.). Intergeneric relationships revealed by molecu-
lar data significantly change the established classifica-tion of the subtribe and may have further important
evolutionary consequences. From a biogeographic point
of view, the Mediterranean origin of the Chironiinae is
clearly established in the light of the current investiga-
tion, but some unresolved patterns remain to be inves-
tigated. For instance, the transatlantic connection
between the southern African (Chironia+Orphium), and
the Chilean C. cachanlahuen, cannot be determinedwithout including the remaining South American species
belonging to the Chironiinae (e.g. Zygostigma australe
or Centaurium ameghinoi). Such incomplete sampling of
the subtribe may also explain the difficulty in finding
good morphological synapomorphies or other distinc-
tive traits such as chromosomal, palynological, or sec-
ondary chemistry features. A better comprehension of
the evolutionary trends within the subtribe Chironiinae,and the Gentianaceae as a whole, definitively requires
the phylogenetic resolutions of the four newly proposed
genera, along with the hitherto poorly investigated ones
(Chironia or Sabatia).
Acknowledgments
The authors gratefully thank Jason R. Grant for help
with grammar in the early stages of the manuscript,
Mike Thiv for providing DNA samples, Louis Zeltner
for sharing unpublished results, Kristin Mylecraine and
Cynthia Frasier for comments on an earlier version of
the manuscript, and Philippe K€upfer for general super-vision of G.M.�s PhD thesis. This study was financially
supported by the Fonds National Suisse de RechercheScientifique (Grant No. 31-52885.97). L.S. thanks Rut-
gers University for funding during the final stages of
manuscript preparation.
References
Adams, L.G., 1996. Gentianaceae. In: Wilson, A. (Ed.), Flora of
Australia, vol. 28. CSIRO, Melbourne, pp. 72–103.
Allan, G.J., Porter, J.M., 2000. Tribal delimitation and phylogenetic
relationships of Loteae and Coronilleae (Faboideae: Fabaceae)
with special reference to Lotus: evidence from nuclear ribosomal
ITS sequences. Am. J. Bot. 87, 1871–1881.
Anderson, E., 1948. Introgressive Hybridization. Wiley, New York.
Axelrod, D.I., Raven, P.H., 1978. Late Cretaceous and Tertiary
vegetation history of Africa. In: Wergers, M.J.A. (Ed.), Biogeog-
raphy and Ecology of Southern Africa. W. Junk, The Hague.
Barber, J.C., Francisco-Ortega, J., Santos-Guerra, A., Turner, K.G.,
Jansen, R.K., 2002. Origin of Macaronesian Sideritis L. (Lamioi-
deae: Lamiaceae) inferred from nuclear and chloroplast sequence
data sets. Mol. Phylogenet. Evol. 23, 293–306.
Bell, N.B., Lester, L.J., 1978. Genetic and morphological detection of
introgression in a clinal population of Sabatia section Campestrina
(Gentianaceae). Syst. Bot. 3, 87–103.
Bell, C.D., Patterson, R.W., 2000. Molecular phylogeny and biogeog-
raphy of Linanthus (Polemoniaceae). Am. J. Bot. 87, 1857–1870.

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 975
Bentham, G., Hooker, J., 1876. Gentianeae. In: Williams, Norgates,
(Eds.), Genera Plantarum. Lovell & Reeve, London, pp. 799–820.
Bohle, U.-R., Hilger, H.H., Martin, W.F., 1996. Island colonisation
and evolution of the insular woody habit in Echium L. (Boragin-
aceae). Proc. Natl. Acad. Sci. USA 93, 11740–11745.
Bramwell, D., Bramwell, Z., 1990. Flores silvestres de las Islas
Canarias. Rueda, Madrid.
Bremer, K., 1992. Ancestral areas: a cladistic reinterpretation of the
center of origin concept. Syst. Biol. 4, 436–445.
Broome, C.R., 1973. Systematics of Centaurium (Gentianaceae) of
Mexico and Central America. Thesis, Duke University, Duke.
Carpenter, I., Locksley, H.D., Scheinmann, F., 1969. Xanthones in
higher plants: biogenetic proposals and a chemotaxonomical
survey. Phytochemistry 8, 2013–2026.
Carr, G.D., 1978. Chromosome number of Hawaiian flowering plants
and the significance of cytology in selective taxa. Am. J. Bot. 65,
236–242.
Caujape-Castells, J., Jansen, R.K., Pedrola-Monfort, J., Membrives,
M., 2002. Space-time diversification of Androcymbium Willd.
(Colchicaceae) in western South Africa. Plant Syst. Evol. 232,
73–88.
Chassot, P., Nemomissa, S., Yuan, Y.-M., K€upfer, P., 2001. High
paraphyly of Swertia L. (Gentianaceae) in the Gentianella-lineage
as revealed by nuclear and chloroplast DNA sequence variation.
Plant Syst. Evol. 229, 1–21.
Cunningham, C.W., Omland, K.E., Oakley, T.H., 1998. Reconstruct-
ing ancestral character states: a critical reappraisal. Trends Ecol.
Evol. 13, 361–366.
Davis, C.C., Bell, C.D., Mathews, S., Donoghue, M.J., 2002. Laur-
asian migration explains Gondwanan disjunctions: evidences from
Malpighiaceae. Proc. Natl. Acad. Sci. USA 99, 6833–6837.
De Queiroz, A., Donoghue, M.J., Kim, J., 1995. Separate versus
combined analysis of phylogenetic evidence. Annu. Rev. Ecol. Syst.
26, 657–681.
Doyle, J.J., 1992. Gene trees and species trees: molecular systematics as
one character taxonomy. Syst. Bot. 17, 144–163.
Doyle, J.J., Doyle, J.L., 1987. A rapid DNA isolation procedure for
small quantities of fresh leaf tissue. Phytochem. Bull. 19, 11–15.
Elias, T.S., Robyns, A., 1975. Family 160. Gentianaceae. In: Wood-
son, R.E., Schery, R.W. (Eds.), Flora of Panama, Part 8. Ann.
Missouri Bot. Gard., vol. 62, pp. 61–101.
Endlicher, S.L., 1838. Genera plantarum secundum ordines naturales
disposita. Vienna.
Farris, J.S., K€allersj€o, M., Kluge, A.G., Bult, C., 1995. Testing
significance of incongruence. Cladistics 10, 315–319.
Farris, J.S., Albert, V.A., K€allersj€o, M., Lipscomb, D., Kluge, A.G.,
1996. Parsimony jackknifing outperforms neighbor-joining. Cla-
distics 12, 99–124.
Favarger, C., 1960. Etude cytologique du Cicendia filiformis et du
Microcala pusilla (Gentianac�ees). Bull. Soc. Bot. France 107, 94–
98.
Francisco-Ortega, J., Santos-Guerra, A., Hines, H., Jansen, R.K.,
1997. Molecular evidence for a Mediterranean origin of the
macaronesian endemic genus Argyranthemum (Asteraceae). Am.
J. Bot. 84, 1595–1613.
Francisco-Ortega, J., Fuertes-Aguilar, J., Gomez-Campo, C., Santos-
Guerra, A., Jansen, R.K., 1999. Internal transcribed spacer
sequence phylogeny of Crambe L. (Brassicaceae): molecular data
reveal two Old World disjunction. Mol. Phylogenet. Evol. 11, 361–
380.
Gentry, A.H., 1982. Neotropical floristic diversity: phytogeographical
connections between Central and South America, Pleistocene
climatic fluctuations, or an accident of the Andean orogeny?
Ann. Missouri Bot. Gard. 69, 557–593.
Gielly, L., Taberlet, P., 1996. A phylogeny of the European gentians
inferred from chloroplast trnL (UAA) intron sequences. Bot. J.
Linn. Soc. 120, 57–75.
Gilg, E., 1895. Gentianaceae. In: Engler, A., Prantls, K. (Eds.), Die
Nat€urlichen Pflanzenfamilien. Verlag von Wilhelm Engelmann,
Leipzig, pp. 50–108.
Goldblatt, P., 1978. An analysis of the flora of Southern Africa: its
characteristics, relationships and origins. Ann. Missouri Bot. Gard.
65, 369–386.
Goldblatt, P., Manning, J., 2000. Cape plants: a conspectus of the
Cape Flora of South Africa. Strelitzia 9, 7–36.
Goloboff, P., 1999. NONA (NO NAME) ver. 2. Published by the
author, Tucum�an, Argentina.
Gottlieb, O.R., 1982. Evolution of xanthones in Gentianaceae and
Guttiferae. In: Micromolecular Evolution, Systematics and Ecol-
ogy. Springer, Berlin, pp. 89–95.
Graham, A., 1984. Lisianthius pollen from the Eocene of Panama.
Ann. Missouri Bot. Gard. 71, 987–993.
Graham, A., 1999. The Tertiary history of the northern temperate
element in the northern Latin America biota. Am. J. Bot. 86, 32–38.
Grisebach, A.H.R., 1839. Genera et species Gentianearum adjectis
observationibus quibusdam phytogeographycis. J.G. Cotta, Stutt-
gart and Tubingen.
Grisebach, A.H.R., 1845. Gentianaceae. In: de Candolles, A. (Ed.),
Prodromus systematis naturalis regni vegetabilis. Fortin Masson
Sociorum, Paris, pp. 39–141.
Grisebach, A.H.R., 1853. Schenkia, novum genus Gentianaerum.
Bonplandia 1, 226.
Gunckel, L.H., 1924. Contribuci�on al estudio de la Erythreae chilensis
Pers. (Cachanlagua). Thesis, Universidad de Concepci�on.Haffer, J., 1982. General aspects of the refuge theory. In: Prance, G.T.
(Ed.), Biological Diversification in the Tropics. Columbia Univer-
sity Press, New York, pp. 6–24.
Hegi, G., 1966. Gentianaceae. In: Illustrierte Flora von Mittel-Europa.
Carl Hanser Verlag, Munich, pp. 1953–2047.
Helfgott, D.M., Francisco-Ortega, J., Santos-Guerra, A., Jansen,
R.K., Simpson, B.B., 2000. Biogeography and breeding system
evolution of the woody Bencomia alliance (Rosaceae) in Macaro-
nesia based on ITS sequence data. Syst. Bot. 25, 82–97.
Higgins, D.G., Thompson, J.D., Gibson, T.J., 1996. Using CLUSTAL
for multiple sequence alignments. Methods Enzymol. 266, 383–402.
Hill, R.S., 1994. History of the Australian Vegetation: Cretaceous to
Recent. Cambridge University Press, Cambridge.
Hostettmann, K., Wagner, H., 1977. Xanthone glycosides. Phyto-
chemistry 16, 821–829.
Hostettmann-Kaldas, M., Hostettmann, K., Sticher, O., 1981. Xant-
hones, flavones and secoiridoids of American Gentiana species.
Phytochemistry 20, 443–446.
Huelsenbeck, J.P., Bull, J.J., Cunningham, C.W., 1996. Combining
data in phylogenetic analysis. Trends Ecol. Evol. 11, 152–157.
Jensen, S.R., Schripsema, J., 2002. Chemotaxonomy and pharmacol-
ogy of the Gentianaceae. In: Struwe, L., Albert, V.A. (Eds.),
Gentianaceae—Systematics and Natural History. Cambridge Uni-
versity Press, Cambridge, pp. 573–631.
Johnson, L.A., Soltis, D.E., 1998. Assessing incongruence: empirical
examples from molecular data. In: Soltis, D.E., Soltis, P.S., Doyle,
J.J. (Eds.), Molecular Systematics of Plants 2: DNA Sequencing.
Kluwer, Boston, pp. 297–348.
Juan, C., Emerson, B.T.C., Oromi, P., Hewitt, G.M., 2000. Coloni-
zation and diversification: toward a phylogeographic synthesis for
the Canary Island. Trends Ecol. Evol. 15, 104–109.
Kim, S.-C., Crawford, D.J., Francisco-Ortega, J., Santos-Guerra, A.,
1996. A common origin for woody Sonchus and five related genera
in the Macaronesian islands: molecular evidence for extensive
radiation. Proc. Natl. Acad. Sci. USA 93, 7743–7748.
Kluge, A.G., 1989. A concern for evidence and the phylogenetic
hypothesis of relationships among Epicrates (Boidae, Serpentes).
Syst. Zool. 38, 7–25.
Knoblauch, E., 1894. Beitr€age zur Kenntniss der Gentianaceae. Bot.
Centralbl. 60, 321–401.

976 G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977
K€ohler, A., 1905. Der Systematische Wert der Pollenbeschaffenkeit bei
den Gentianaceen. Mitteil. Bot. Mus. Z€urich, 25.Kunkel, G., 1993. Die Kanarischen Inseln und ihre Pflanzenwelt.
Gustav Fischer, Stuttgart Jena, New York.
Lia, V.V., Confalioneri, V.A., Comas, C.I., Hunziker, J.H., 2001.
Molecular phylogeny of Larrea and its allies (Zygophyllaceae):
reticulate evolution and the probable time of creosote bush arrival
to North America. Mol. Phylogenet. Evol. 21, 309–320.
Maddison, W.P.R., Maddison, D.R., 1997. MacClade 3.07. Sinauer
Associates, Sunderland, MA.
Maddison, D.R., Ruvolo, M., Swofford, D.L., 1992. Geographic
origins of human mitochondrial DNA: phylogenetic evidence from
control region sequences. Syst. Biol. 41, 111–124.
Mansion, G., in press. A new classification of the polyphyletic genus
Centaurium Hill (Chironiinae-Gentianaceae): description of the
New-World endemic Zeltnera, and reinstatement of Gyrandra
Griseb. and Schenkia Griseb. Taxon.
Mansion, G., Zeltner, L., in press. Phylogenetic relationships within
the New World endemic Zeltnera, (Gentianaceae-Chironiinae)
inferred from molecular and karyological data. Am. J. Bot.
Marais, W., Verdoon, I.C., 1963. Gentianaceae. In: Dyer, R.A., Codd,
L.E., Rycrofts, H.B. (Eds.), Flora of Southern Africa. Department
of Agricultural Technical Services, Republic of South Africa,
Pretoria, pp. 171–243.
McKenna, M.C., 1983. Cenozoic paleogeography of North Atlantic
land bridges. In: Bott, M.H.P., Saxov, S., Talwani, M., Thiedes, J.
(Eds.), Structure and Development of the Greenland–Scotland
Ridge. Plenum Press, New York.
Melderis, A., 1931. Genetical and taxonomical studies in the genus
Erythraea Rich. Acta Horti. Bot. Univ. Latv. 6, 123–256.
Mes, T.H.M., van Brederode, J., Hart, H.T., 1996. Origin of the
woody Macaronesian Sempervivioideae and the phylogenetic
position of the East African of Aeonium. Bot. Acta 109, 47–491.
M�esz�aros, S., 1994. Evolutionary significance of xanthones in Gen-
tianaceae: a reappraisal. Biochem. Syst. Ecol. 22, 85–94.
M�esz�aros, S., De Laet, J., Smets, E., 1996. Phylogeny of temperate
Gentianaceae: a morphological approach. Syst. Bot. 21,
153–168.
M�esz�aros, S., de Laet, J., Goethals, V., Smets, E., Nilsson, S., 2002.
Cladistics of the Gentianaceae. In: Struwe, L., Albert, V.A. (Eds.),
Gentianaceae—Systematics and Natural History. Cambridge Uni-
versity Press, Cambridge, pp. 310–376.
Molina, J.I., 1787. Saggio Sulla Storia Civile del Chili. Bologna, Nella
Stamper�ia de S. Tommaso d�Aquino.
Morrone, J.J., Crisci, J.V., 1995. Historical biogeography: introduc-
tion to methods. Annu. Rev. Ecol. Syst. 26, 373–401.
Mort, M.E., Soltis, D.E., Soltis, P.S., Francisco-Ortega, J., Santos-
Guerra, A., 2001. Phylogenetic relationships and evolution of
Crassulaceae inferred from matK sequence data. Am. J. Bot. 88,
76–91.
NatureServe, 2003. NatureServe Explorer: an online encyclopedia of
life [web application]. Version 1.8. NatureServe, Arlington, Vir-
ginia. Available from <http://www.natureserve.org/explorer>.
Nilsson, S., 2002. Gentianaceae: a review of palynology. In: Struwe,
L., Albert, V.A. (Eds.), Gentianaceae—Systematics and Natural
History. Cambridge University Press, Cambridge, pp. 377–497.
Nixon, K.C., 2002. WinClada ver. 1.00.08. Published by the author,
Ithaca, NY.
Panero, J.L., Francisco-Ortega, J., Jansen, R.K., Santos-Guerra, A.,
1999. Molecular evidences for multiple origins of woodiness and a
New-World biogeographic connection of the Macaronesian Island
endemic Pericallis (Asteraceae: Senecioneae). Proc. Natl. Acad. Sci.
USA 96, 13886–13891.
Perry, J.D., 1971. Biosystematic studies in the North American genus
Sabatia (Gentianaceae). Rhodora 73, 309–369.
Posada, D., Crandall, K.A., 1998. ModelTest: testing the model of
DNA substitution. Bioinformatics 14, 817–818.
Punt, W., Nienhuis, W., 1976. The northwest European pollen flora. 6.
Gentianaceae. Rev. Palaeobot. Palynol. 21, 89–123.
Qian, H., 1999. Floristic analysis of vascular plant genera of North
America north of Mexico: characteristics of phytogeography. J.
Biogeogr. 26, 1307–1327.
Quezel, P., 1978. Analysis of the flora of Mediterranean and Saharan
Africa. Ann. Missouri Bot. Gard. 65, 479–534.
Rambaut, A., Charleston, M., 2001. TreeEdit: Phylogenetic Tree
Editor v1.0 alpha 10. Available from <http://evolve.zoo.ox.ac.uk/
software>.
Rao, S.K., Chinnappa, C.C., 1983. Pericolporate pollen in the
Gentianaceae. Can. J. Bot. 61, 174–178.
Raven, P.H., 1963. Amphitropical relationships in the floras of North
and South America. Q. Rev. Biol. 38, 151–177.
Raven, P.H., Axelrod, D.I., 1974. Angiosperm biogeography and past
continental movements. Ann. Missouri Bot. Gard. 61, 539–673.
Renner, S.S., Clausing, G., Meyer, K., 2001. Historical biogeography
of the Melastomataceae: the role of Tertiary migration and long-
distance dispersal. Am. J. Bot. 88, 1290–1300.
Richardson, J.E., Pennington, R.T., Pennington, T.D., Hollingsworth,
P.M., 2001. Rapid diversification of a species-rich genus of
Neotropical rain forest trees. Science 293, 2242–2245.
Rieseberg, L.H., 1995. The role of hybridization in evolution: old wine
in new skins. Am. J. Bot. 82, 944–944.
Robyns, A., 1954. Essai d��etude syst�ematique et �ecologique des
Centaurium de Belgique. Bull. J. Bot. Bruxelle 24, 349–398.
Ronquist, F., 1996. DIVA version 1.1. Computer program and manual
available by anonymous FTP at (ftp.uu.se or ftp.systbot.uu.se).
Ronquist, F., 1997.Dispersal-vicariance analysis: a new approach to the
quantification of historical biogeography. Syst. Biol. 46, 196–203.
Sanderson, M.J., 1997. A nonparametric approach to estimating
divergence times in the absence of rate constancy. Mol. Biol. Evol.
14, 1218–1231.
Sanderson, M.J., 1998. Estimating rate and time in molecular
phylogenies: beyond the molecular clock? In: Soltis, D.E., Soltis,
P.M., Doyle, J.J. (Eds.), Molecular Systematics of Plants II. DNA
Sequencing. Kluwer Academic Publishers, Boston, pp. 242–264.
Schaufelberger, D., 1986. Contribution �a l��etude phytochimique des
Gentianac�ees. Thesis, Lausanne University, Lausanne.
Schmidt, G.L.E., 1828. De Erythraea. Berolini, Berlin.
Schneider, W., 1974. Pflanzliche drogen. Govi Verlag, Frankfurt.
Shinners, L.H., 1957. Synopsis of the genus Eustoma (Gentianaceae).
Southwest Nat. 2, 38–43.
Scott, L., 1995. Pollen evidence for vegetational and climatic change in
Southern Africa during the Neogene and Quaternary. In: Vrba, E.,
Denton, G.H., Partridge, T.C., Burckle, L.H. (Eds.), Paleoclimate
and Human Evolution with Emphasis on Human Origins. Yal
University Press, pp. 65–76.
Struwe, L., Albert, V.A., Bremer, B., 1994. Cladistics and family level
classification of the Gentianales. Cladistics 10, 175–206.
Struwe, L., Kadereit, J.W., Klackenberg, J., Nilsson, S., Thiv, M., von
Hagen, K.B., Albert, V.A., 2002. Systematics, character evolution,
and biogeography of Gentianaceae, including a new tribal and
subtribal classification. In: Struwe, L., Albert, V.A. (Eds.), Gen-
tianaceae—Systematics and Natural History. Cambridge Univer-
sity Press, Cambridge, pp. 21–309.
Swofford, D.L., 2002. PAUP*. Phylogenetic Analysis Using Parsi-
mony (*and Other Methods). Version 4. Sinauer Associates,
Sunderland, MA.
Taberlet, P., Gielly, L., Pautou, G., Bouvet, J., 1991. Universal primers
for amplification of three non-coding regions of chloroplast DNA.
Plant Mol. Biol. 17, 1105–1109.
Takhtajan, A., 1986. Floristic Regions of the World. University of
California Press, Berkeley, CA.
Thiv, M., Kadereit, J.W., 2002a. Tribe Chironieae. In: Struwe, L.,
Albert, V.A. (Eds.), Gentianaceae—Systematics and Natural His-
tory. Cambridge University Press, Cambridge, pp. 108–137.

G. Mansion, L. Struwe / Molecular Phylogenetics and Evolution 32 (2004) 951–977 977
Thiv, M., Kadereit, J.W., 2002b. A morphological analysis of
Gentianaceae–Canscorinae and the evolution of anisomorphic
androecia in the subtribe. Syst. Bot. 27, 708–788.
Thiv, M., Struwe, L., Kadereit, J.W., 1999a. The phylogenetic
relationships and evolution on the Canarian laurel forest endemic
Ixanthus viscosus (Aiton) Griseb. (Gentianaceae): evidence from
matK and ITS sequences, and floral morphology and anatomy.
Plant Syst. Evol. 218, 299–317.
Thiv, M., Struwe, L., Albert, V.A., Kadereit, J.W., 1999b. The
phylogenetic relationships of Saccifolium bandeirae (Gentianaceae)
reconsidered. Harvard Pap. Bot. 4, 519–526.
Tiffney, B.H., 1985a. Perspectives on the origin of the floristic
similarity between eastern Asia and eastern North America. J.
Arnold Arbor. 66, 73–94.
Tiffney, B.H., 1985b. The Eocene north Atlantic land bridge: its
importance in Tertiary and modern phytogeography of the
northern hemisphere. J. Arnold Arbor. 66, 243–273.
Tiffney, B.H., Manchester, S.R., 2001. The use of geological and
paleontological evidence in evaluating plant phylogeographic
hypotheses in the northern hemisphere tertiary. Int. J. Plant Sci.
162, 3–17.
Vald�es, B., Talavera, S., Fern�andez-Galiano, E., 1987. Gentianaceae.
In: Flora vascular de Andaluc�ia Occidental, vol. 2. S.A. Ketres
Editora, Barcelona, pp. 339–345.
van der Sluis, W.G., 1985. Chemotaxonomical investigations of the
genera Blackstonia and Centaurium (Gentianaceae). Plant Syst.
Evol. 149, 253–286.
van Veller, M.G.P., Zandee, M., Kornet, D.J., 2001. Measures for
obtaining inclusive sets of area cladograms under assumptions
zero, 1 and 2 with different methods for vicariance biogeography.
Cladistics 17, 248–259.
von Hagen, K.B., Kadereit, J.W., 2001. The phylogeny of Gentianella
(Gentianaceae) and its colonization of the Southern hemisphere as
revealed by nuclear and chloroplast DNA sequence variation. Org.
Divers. Evol. 1, 61–79.
Weaver, R.E., R€udenberg, L., 1975. Cytotaxonomic notes on some
Gentianaceae. J. Arnold Arbor. 56, 211–222.
Wen, J., 1999. Evolution of eastern Asian and eastern North American
disjunct distributions in flowering plants. Annu. Rev. Ecol. Syst.
30, 421–455.
Wendel, J.F., Doyle, J.J., 1998. Phylogenetic incongruence: window
into genome history and molecular evolution. In: Soltis, D.E.,
Soltis, P.M., Doyle, J.J. (Eds.), Molecular Systematics of Plants II.
DNA Sequencing. Kluwer Academic Publishers, Boston, pp.
265–296.
White, T.J., Bruns, T., Lee, S., Taylor, J., 1990. Amplification and
direct sequencing of fungal ribosomal DNA for phylogenetics. In:
Innis, M.A., Gelfund, D.H., Sninsky, J.J., Whites, T.J. (Eds.), PCR
Protocols: A Guide to Methods and Applications. Academic Press,
San Diego, pp. 315–322.
Whitmore, T.C., Prance, G.T., 1987. Biogeography and Quaternary
History in Tropical America. Clarendon Press, Oxford.
Wickens, G.E., 1976. The Flora of the Jebel. Kew Bull add Marra
(Sudan Republic) and its geographical affinities. Kew Bull Add.
Series 5, 1–368.
Wilbur, R.L., 1955. A revision of the North American genus Sabatia
(Gentianaceae). Rhodora 57, 1–104.
Winkworth, R.C., Grau, J., Robertson, A.W., Lockhart, P.J., 2002.
The origins and evolution of the genusMyosotis L. (Boraginaceae).
Mol. Phylogenet. Evol. 24, 180–193.
Yuan, Y.-M., K€upfer, P., 1995. Molecular phylogenetics of the
subtribe Gentianinae (Gentianaceae) inferred from the sequences
of the internal transcribed spacer (ITS) of nuclear ribosomal DNA.
Plant Syst. Evol. 196, 206–226.
Yuan, Y.-M., K€upfer, P., 1997. The monophyly and rapid evolution of
Gentiana sect. Chondrophyllae Bunge s.l. (Gentianaceae): evidence
from the nucleotide sequences of the internal transcribed spacer of
nuclear ribosomal DNA. Bot. J. Linn. Soc. 123, 25–43.
Yuan, Y.-M., K€upfer, P., Doyle, J.J., 1996. Infrageneric phylogeny of
the genus Gentiana (Gentianaceae) inferred from nucleotide
sequences of the internal transcribed spacer (ITS) of nuclear
ribosomal DNA. Am. J. Bot. 83, 641–652.
Yuan, Y.-M., Wolhauser, S., M€oller, M., Chassot, P., Mansion, G.,
Grant, J., K€upfer, P., Klackenberg, J., 2003. Monophyly and
relationships of the tribe Exaceae (Gentianaceae) inferred from
nuclear ribosomal and chloroplast DNA sequences. Mol. Phylo-
genet. Evol. 28, 500–517.
Zeltner, L., 1970. Recherches de biosyst�ematique sur les genres
Blackstonia Huds. et Centaurium Hill (Gentianaceae). Bull. Soc.
Neuchatel Sci. Nat. 93, 1–164.
Top Related