







Languages
Pages
Legal


23
Jonathan M. EIBLDepartment of Biology, Virginia Commonwealth University,
Richmond, Virginia 23284, U.S.A.
Gregory M. PLUNKETTDepartment of Biology, Virginia Commonwealth University,
Richmond, Virginia 23284, [email protected]
Porter P. LOWRY IIMissouri Botanical Garden, P.O. Box 299,
St. Louis, Missouri 63166-0299, [email protected]
Laboratoire de Phanérogamie, Muséum national d’Histoire naturelle,16 rue Buffon, 75005 Paris, France.
Evolution of Polyscias sect. Tieghemopanax (Araliaceae) based on nuclear and chloroplast DNA sequence data
ABSTRACTPolyscias sect. Tieghemopanax comprises approximately 26 species almostentirely endemic to New Caledonia. Three independent molecular markerswere used to infer phylogenetic relationships and evolutionary patterns withinthe Tieghemopanax group, and to evaluate the status of a single highly variablespecies, P. dioica. Sequence data from nuclear ribosomal ITS and 5S spacersalong with intron and spacer sequences from the plastid trnL-trnF region wereanalyzed separately and in each possible combination of the three data sets.Combined analyses provided topologies largely congruent with each of theseparate analyses but with increased resolution, higher bootstrap support, anddecreased computational time. The resulting cladograms confirm themonophyly of section Tieghemopanax and suggest an allopatric or adaptive di-vergence model of speciation in response to geography, elevation, and/or sub-strate type for most species. In contrast, P. dioica may represent an assemblageof once distinct but closely related species whose boundaries have recently bro-ken down through hybridization and introgression. The results suggest that theTieghemopanax group originated and diversified on New Caledonia (23 spp.);subsequent long-distance dispersal to other archipelagos resulted in the evolu-tion of three local endemics (1 sp. each in Vanuatu, Fiji, and Australasia) andthe widespread distribution of P. cissodendron in the SW Pacific.
ADANSONIA, sér. 3 • 2001 • 23 (1) : 23-48© Publications Scientifiques du Muséum national d’Histoire naturelle, Paris.
KEY WORDSspeciation, Araliaceae,
biogeography, New Caledonia,
Polyscias, Tieghemopanax.

INTRODUCTION
Araliaceae (the ginseng family) comprise about50 genera and approximately 1,200-1,400 speciesof vascular plants (MABBERLEY 1997; PLUNKETT
et al. 2001). Within this family, Polyscias J.R. Forst.& G. Forst. (c. 130 spp.) and Schefflera J.R. Forst.& G. Forst. (about 650 spp.) are the two largestgenera. In contrast to the pantropical distributionof Schefflera, Polyscias (as currently circumscribed)is restricted to the Old World tropics, from Africaacross the Indian Ocean to Malesia and Australia,and thence across the islands of the south Pacificas far east as Tahiti (BERNARDI 1971; PHILIPSON
1979; LOWRY 1989). BERNARDI (1979) hypothe-sized that Malesia may have been the cradle ofaraliad origin, and noted that New Caledonia hasmore species per unit area than any other regionon earth — over 90 species in eight genera(LOWRY unpubl.). Polyscias is well represented on
this southwest Pacific island, with 23 indigenousspecies (all but one endemic) and four cultivatedspecies of Indo-Malesian origin (LOWRY in prep.).The native species form a morphologically coher-ent group (BERNARDI 1979), including trees orshrubs with odd-pinnate leaves that lack sheath-ing petioles, and leaflets that are opposite, sessileto subsessile, entire or toothed, and pinnatelyveined. Their flowers have (4-)5 petals, (4-)5 sta-mens, (4-)5 anthers, 2-locular ovaries, 2 styles(free or partially connate), and 2 carpels; they areusually arranged in panicles of umbellules,racemules or heads, or in compound dichasia.These species have mostly an andromonoecious(or less commonly dioecious) mating system(SCHLESSMAN et al. 1990, 2001) and their fruitsare fleshy drupes that are strongly compressed lat-erally (HUTCHINSON 1967; LOWRY 1989). Theuniformity of the indigenous New Caledonianspecies of Polyscias led some authors to recognize
Eibl J.M., Plunkett G.M. & Lowry II P.P.
24 ADANSONIA, sér. 3 • 2001 • 23 (1)
MOTS CLÉSspéciation, Araliaceae,
biogéographie, Nouvelle-Calédonie,
Polyscias, Tieghemopanax.
RÉSUMÉEvolution de Polyscias sect. Tieghemopanax (Araliaceae) à partir de l’analyse del’ADN nucléaire et chloroplastique.Polyscias sect. Tieghemopanax renferme environ 26 espèces presque toutesendémiques de Nouvelle-Calédonie. Trois marqueurs moléculaires indépen-dants ont été utilisés pour déduire les affinités et les modèles d’évolution ausein du groupe Tieghemopanax, et pour évaluer le statut d’une espèce parti-culièrement variable, P. dioica. Les séquences ITS et espaceur 5S de l’ADNribosomique nucléaire ainsi que les séquences de l’intron et de l’espaceur de larégion trnL-trnF de l’ADN chloroplastique ont été analysées séparément etdans chacune des combinaisons possibles de ces trois ensembles de données.Les topologies obtenues à partir des analyses combinées s’avèrent largementcongruentes avec celles de chacune des analyses individuelles mais avec unemeilleure résolution, des valeurs de bootstrap plus élevées et un temps decalcul moindre. Les cladogrammes qui en résultent confirment la monophyliede la section Tieghemopanax et suggèrent un modèle de spéciation allopa-trique ou d’adaptation divergente, en liaison avec la géographie, l’altitude,et/ou le type de substrat, pour la plupart des espèces. Au contraire, P. dioicareprésenterait un ensemble d’espèces proches autrefois distinctes entres les-quelles les barrières ont récemment disparu par hybridation et introgression.Les résultats obtenus suggèrent que le groupe Tieghemopanax est originaire deNouvelle-Calédonie où il s’est diversifié (23 spp.) et qu’une dispersion ulté-rieure à longue distance vers d’autres archipels a permis l’évolution de troisendémiques locaux (une espèce au Vanuatu, une à Fiji et une en Australasie)et d’une autre espèce (P. cissodendron) largement répandue dans le SWPacifique.

them as a distinct genus, Tieghemopanax R. Vig.(VIGUIER 1905; GUILLAUMIN 1948), which alsoincludes several members in Fiji (SMITH &STONE 1968; SMITH 1985), as well as Vanuatu(formerly the New Hebrides) and a number ofother south Pacific islands (LOWRY et al. 1986;LOWRY 1989). More recently, however,Tieghemopanax has been treated as an informalgroup or section within a more broadly definedPolyscias (SMITH & STONE 1968; PHILIPSON
1978, 1979; BERNARDI 1979; SMITH 1985;LOWRY 1989).
As originally defined, Polyscias included taxawith pinnately compound leaves, 4-5 styles and4-5 carpels (FORSTER & FORSTER 1776), withP. pinnata J.R. Forst. & G. Forst. [= P. scutellaria(Burm. f.) Fosb.] as the type species (see SMITH &STONE 1968; LOWRY 1989). The original circum-scription of Tieghemopanax and another genus,Nothopanax Miq., included pinnate leavedspecies having 2-3 carpellate ovaries (MIQUEL
1855, 1856; VIGUIER 1905). However, STONE
(1965a,b) placed Nothopanax in synonymy underPolyscias and transferred several species ofTieghemopanax from Vanuatu to Polyscias, sug-gesting that the same might also be warranted forthe New Caledonian members of the genus.More recently, species previously assigned toTieghemopanax have likewise been treated in abroadly defined Polyscias following PHILIPSON’s(1978, 1979) expanded circumscription (e.g.,LOWRY et al. 1986; LOWRY 1989). A recent phy-logenetic study by PLUNKETT et al. (2001) sug-gested that this broadly defined Polyscias isparaphyletic. The monophyly of species tradi-tionally assigned to Tieghemopanax, however, wasaffirmed. In light of this finding, it now seemsopportune to explore in more detail questionsregarding the evolution and diversification ofspecies belonging to the Tieghemopanax group.Given its center of distribution in NewCaledonia, where most species are endemic,Tieghemopanax represents a model for the studyof diversification on an island with a particularlyintriguing geological and botanical history.
The New Caledonian flora, with c. 3,250native species of seed plants, exhibits remarkablyhigh levels of endemism both at the level ofspecies (c. 79%) and genus (~14% of the nearly
770 genera recorded) in an area of only about17,000 km2 (MORAT 1993; LOWRY 1998;MORAT et al. 2001). These include many relictualtaxa representing ancient Australasian lineagespresent at the time New Caledonia separatedfrom Australia c. 74 MY ago (KROENKE 1996),and others derived from more recent coloniza-tions by long-distance dispersal. Evolution andspeciation have been driven in part by thepresence of highly selective ultramafic substratescovering nearly 1/3 of the island (JAFFRÉ 1976;MORAT et al. 1986; JAFFRÉ et al. 1987) and aremarkable diversity of bioclimatic and ecologicalconditions resulting in a wide range of vegetationformations (MORAT 1993; LOWRY 1998), includ-ing low elevation to montane rainforest (MORAT
et al. 1984), dry sclerophyllous forests on cal-careous substrates (JAFFRÉ et al. 1993; MORAT etal. 2001), and maquis vegetation (MORAT et al.1986), a characteristic low, heath-like, sclerophyl-lous formation largely restricted to ultramafics.This unique combination of geological, climato-logical, and ecological features makes NewCaledonia a “hotspot” for both paleo- andneoendemics, and presents an ideal context forstudying evolutionary processes.
Based on evidence from molecular data(nuclear Internal Transcribed Spacer-ITS) in con-junction with present-day geographical distribu-tions, PLUNKETT et al. (2001) speculated that anearly member of section Tieghemopanax may havearrived in New Caledonia by long-distance dis-persal from Australasia, and subsequently under-gone broad radiation across the island. Inaddition to confirming the monophyly of thosespecies of Polyscias traditionally assigned toTieghemopanax, the ITS study suggested that theAustralian endemic P. sambucifolia also belongswithin the Tieghemopanax clade and that a sec-ond Australasian species, P. elegans, the sole mem-ber of sect. Gelibia (Hutch.) Philipson, may besister to Tieghemopanax. These results pose inter-esting questions regarding the origin, dispersal,and radiation of section Tieghemopanax withinNew Caledonia. For example, did the twoAustralian species descend from the presumedAustralasian progenitor species of Tieghemo-panax, or do they represent secondary dispersals
Polyscias sect. Tieghemopanax (Araliaceae)
25ADANSONIA, sér. 3 • 2001 • 23 (1)

back to Australia from New Caledonia? Similarly,are the New Caledonian species descendents of asingle dispersal event from Australasia or multipledispersals? And how might the biophysical fea-tures of New Caledonia (e.g., vegetation, climate,elevation, edaphic conditions) have contributedto speciation within the Tieghemopanax groupafter the initial dispersal(s)? Unfortunately, ITSdata alone were not sufficient to resolve relation-ships within the section. Given these limitations,the present study has employed additional non-coding sequences from the intron and intergenicspacer of the plastid trnL-trnF region, and fromthe non-transcribed spacer of nuclear 5S rDNA.These markers were chosen because they accumu-late mutations at a similar or more rapid rate thanITS. Altogether, three data sets (with comparablesampling) were assembled and analyzed, repre-senting three independent (presumably unlinked)markers, three types of non-coding sequences (anintron, two transcribed spacers, and two non-transcribed spacers), from two inheritance units(nuclear and plastid DNA). The data sets wereanalyzed, both separately and in combination, inan effort to examine phylogenetic relationshipswithin the Tieghemopanax group.
Circumscriptions at and below the genus levelin most plant groups have in general been basedon traditional taxonomic methods, relying largelyon the comparison of morphological characters.These methods have proven useful in delimitingspecies within section Tieghemopanax, wherenumerous straightforward morphological,edaphic and/or geographical features are availableto distinguish the 26 currently recognized taxa(some of which remain to be described; LOWRY
1989 and unpubl.). In applying this traditionalconcept to Tieghemopanax, however, it has beennecessary to recognize one very broadly definedspecies, P. dioica, which exhibits an unusuallyhigh level of morphological variability and is dis-tributed very widely within New Caledonia. Ascurrently circumscribed, P. dioica includes severalpreviously recognized species [e.g., P. pulchella(Baill.) Harms, P. schlechteri Harms, P. simabae-folia (R. Vig.) Harms, P. suborbicularis (Baill.)Harms, Tieghemopanax nigrescens Pancher exR. Vig., and T. viguieri Däniker], but the corre-sponding morphological types often appear to
grade into one another and frequently co-occur ata give locality (LOWRY unpubl.). While the plantsnow placed in P. dioica share several features thatreadily allow them to be distinguished from othermembers of the Tieghemopanax group, it is never-theless unclear whether they represent a singlespecies with significant phenotypic variation orperhaps several distinct taxa. Resolution of thisdilemma may be difficult in the absence of anexplicit phylogenetic framework to assess rela-tionships within the Tieghemopanax clade and thepopulations comprising P. dioica.
Apart from this single, highly variable species,the other members of section Tieghemopanax aremorphologically coherent and apparently com-prise a series of discrete, well-defined species pairs(or trios). Therefore, in using traditional taxo-nomic approaches we must apply two very differ-ent species definitions in section Tieghemopanax.This may belie two distinct modes of evolutionand speciation in a single group of closely relatedtaxa. In such cases, a robust phylogenetic hypoth-esis (especially one based on data independent ofthe morphological treatments, such as moleculardata) can provide the needed framework for eval-uating evolutionary patterns.
The distribution of section Tieghemopanax,centered on New Caledonia, provides an idealopportunity to examine evolution on this intrigu-ing island system, characterized by such extraor-dinary levels of both paleo- and neoendemism.To date, however, no detailed studies of this kindhave been completed. Thus, we hope that thepresent paper will provide not merely a test ofspeciation patterns in Polyscias, but perhaps also afirst step in evaluating evolutionary modes acrossmany of the endemic plant groups of NewCaledonia. The study reported here represents apreliminary analysis to explore further the evolu-tionary patterns found in New CaledonianPolyscias. Specifically, we have used moleculardata to: (1) confirm the monophyly of sectionTieghemopanax; (2) elucidate phylogenetic rela-tionships among the species within the section;(3) infer evolutionary patterns (e.g., patterns ofspeciation, biogeography, and morphologicalcharacter evolution) among these species; (4)assess the phylogenetic status of plants nowassigned to the highly variable species P. dioica;
Eibl J.M., Plunkett G.M. & Lowry II P.P.
26 ADANSONIA, sér. 3 • 2001 • 23 (1)

and (5) consider additional taxa that may providefurther tests of evolutionary mechanisms in theNew Caledonian flora.
MATERIALS AND METHODS
Newly derived data sets were collected fromone nuclear marker (the 5S rDNA spacer) andone chloroplast marker (trnL-trnF). In addition,the previously published ITS data set ofPLUNKETT et al. (2001) was expanded by theaddition of seven new sequences. Leaf tissue wascollected from 29 accessions (representing 19 ofthe 26 species of Tieghemopanax) and dried in sil-ica gel or preserved in a CTAB/salt solution (seeTable 1). Total DNAs were extracted using theCTAB method of DOYLE & DOYLE (1987) asmodified by SOLTIS et al. (1991) or the DNeasyPlant Mini kit (QIAGEN Inc.). Oligonucleotideprimers were obtained on the basis of previouslypublished studies for ITS (WHITE et al. 1990;DOWNIE & KATZ-DOWNIE 1996; WEN &ZIMMER 1996), the 5S region (UDOVICIC et al.1995) and trnL-trnF (TABERLET et al. 1991,although their primer “f ”was modified as follows:5´—AAC TGG TGA CAC GAG GAT TTTCAG—3´).
For the trnL-trnF and ITS regions, each PCRreaction included 0.4 µL of unquantified tem-plate DNA, 0.2 µL Taq DNA polymerase (QIA-GEN), 5 µL 10X Taq buffer (supplied withenzyme), 2 µL of 25 mM MgCl2, 2 µL DMSO,1 µL of a 5 µM solution of each primer, 4 µL of10 mM dNTPs and ultrapurified water to a finalvolume of 50 µL. Temperature-cycler parametersfor the trnL-trnF regions were set at 94°C(1 min.), 49°C (1 min.), and 72°C (2 min.) fordenaturing, annealing, and extension steps,respectively (35 cycles). For ITS, 40 cycles wereused, each cycle including 94°C (30 sec.), 48°C(1 min.), and 68°C (1 min.). In amplifications ofthe 5S spacer, an identical PCR recipe was usedexcept that 8 µL of 10X buffer was added (ratherthan 5 µL). In addition, temperature cycler para-meters for the 5S amplifications were set at 93°C(10 sec.), 60°C (10 sec.), and 72°C (12 sec.) fordenaturing, annealing, and extension steps,respectively (31 cycles). These modifications of
PCR parameters for 5S preferentially amplified asingle repeat unit (rather than 2 or more units ofthis tandemly repeated gene and its spacer), thuseliminating the multiple-banding patternsobserved in early PCR trials (see also UDOVICIC
et al. 1995; HENEGARIU et al. 1997). PCR ampli-cons were purified using the QIAquick PCRcleanup kit or the agarose gel extraction kit (bothQIAGEN Inc.). For the 5S spacer, sequenceswere obtained from complimentary strands usingjust two sequence reactions. For the trnL-trnFregion, internal primers (trnL-trnF “d” and “e”)were also necessary in most taxa to acquire com-plete sequences. Internal primers (ITS 3 and ITS5.8S) were also necessary to obtain complete ITSsequences. Modified dideoxy-chain terminationsequences (SANGER 1977) were performed usingthe BigDye Terminator cycle sequencing kit (PEApplied Biosystems) according to the manufac-turer’s instructions, followed by DyeEx Spin Kit(QIAGEN) purification and electrophoresis onan ABI Prism 377 automated sequencer. DNAsequence data were assembled and edited usingthe Sequencher (version 3.1) computer program(GeneCodes Corp.).
Despite efforts to eliminate multiple bands in5S rDNA amplicons, several accessions of Polysciasdioica (e.g., Lowry 4771, 4800, 4804) and thesingle accessions of P. elegans (Plunkett 1495),P. cissodendron (Lowry 4749), and P. pancheri(Lowry 4793) yielded two or more amplificationproducts. These bands did not express the typicalmultiple repeat pattern observed prior to opti-mization of PCR for 5S, suggesting the presenceof two or more 5S spacer size variants in thesesamples. PCR products from two of these acces-sions, P. dioica (Lowry 4804) and P. elegans(Plunkett 1495), were separated in 2% TBEagarose, excised, and gel-cleaned. Each band wasthen cloned into the pCR2.1-TOPO vectorusing the TOPO TA Cloning kit (InvitrogenCorp.) and sequenced independently to deter-mine sequence homology with 5S spacers fromother species. The remaining accessions in thestudy, including all other P. dioica samples (Lowry4647, 4712, 4713, 4781, and 5099), yielded sin-gle-banded amplicons in PCR reactions targetingthe 5S spacer region, allowing for direct sequenc-ing of the PCR products.
Polyscias sect. Tieghemopanax (Araliaceae)
27ADANSONIA, sér. 3 • 2001 • 23 (1)
Eibl J.M., Plunkett G.M. & Lowry II P.P.
28 ADANSONIA, sér. 3 • 2001 • 23 (1)
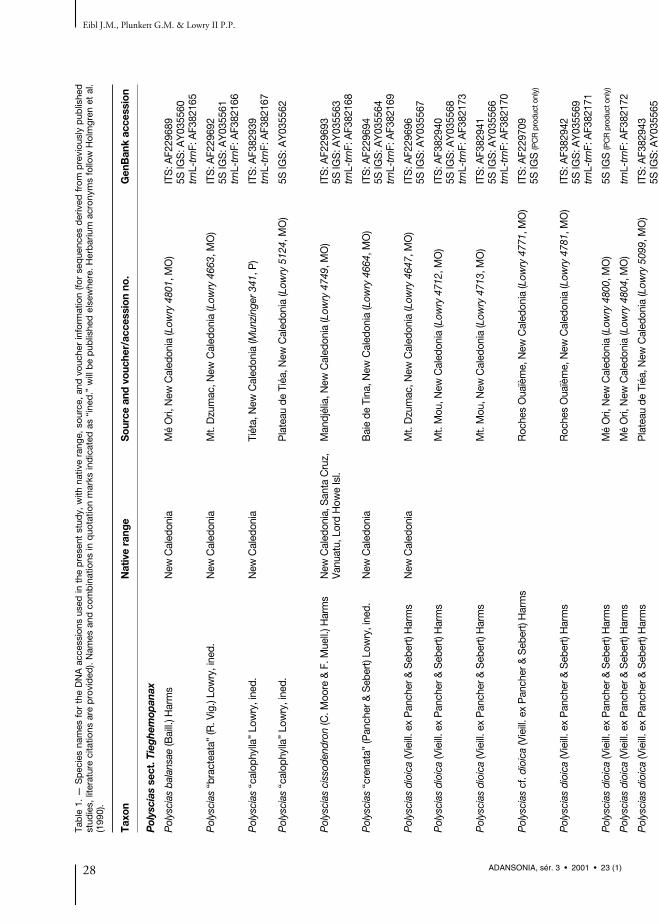
Tab
le 1
.—S
pec
ies
nam
es f
or t
he D
NA
acc
essi
ons
used
in t
he p
rese
nt s
tud
y, w
ith n
ativ
e ra
nge,
sou
rce,
and
vou
cher
info
rmat
ion
(for
seq
uenc
es d
eriv
ed f
rom
pre
viou
sly
pub
lishe
dst
udie
s, li
tera
ture
cita
tions
are
pro
vid
ed).
Nam
es a
nd c
omb
inat
ions
in q
uota
tion
mar
ks in
dic
ated
as
“ine
d.”
will
be
pub
lishe
d e
lsew
here
. Her
bar
ium
acr
onym
s fo
llow
Hol
mgr
en e
t al
.(1
990)
.
Taxo
nN
ativ
e ra
nge
Sou
rce
and
vouc
her/
acce
ssio
n no
.G
enB
ank
acce
ssio
n
Pol
ysci
asse
ct. T
iegh
emop
anax
Pol
ysci
as b
alan
sae
(Bai
ll.) H
arm
s N
ew C
aled
onia
M
é O
ri, N
ew C
aled
onia
(Low
ry 4
801,
MO
) IT
S: A
F229
689
5S IG
S: A
Y03
5560
trnL
-trn
F: A
F382
165
Pol
ysci
as“b
ract
eata
” (R
. Vig
.) Lo
wry
, ine
d.
New
Cal
edon
ia
Mt.
Dzu
mac
, New
Cal
edon
ia (L
owry
466
3, M
O)
ITS
: AF2
2969
25S
IGS
: AY
0355
61tr
nL-t
rnF:
AF3
8216
6
Pol
ysci
as “
calo
phyl
la”
Low
ry, i
ned.
New
Cal
edon
ia
Tiét
a, N
ew C
aled
onia
(Mun
zing
er 3
41, P
) IT
S: A
F382
939
trnL
-trn
F: A
F382
167
Pol
ysci
as “
calo
phyl
la”
Low
ry, i
ned.
Pla
teau
de
Tiéa
, New
Cal
edon
ia (L
owry
512
4, M
O)
5S IG
S: A
Y03
5562
Pol
ysci
as c
isso
dend
ron
(C. M
oore
& F
. Mue
ll.) H
arm
s N
ew C
aled
onia
, San
ta C
ruz,
M
andj
élia
, New
Cal
edon
ia (L
owry
474
9, M
O)
ITS
: AF2
2969
3Va
nuat
u, L
ord
How
e Is
l. 5S
IGS
: AY
0355
63tr
nL-t
rnF:
AF3
8216
8
Pol
ysci
as “
cren
ata”
(Pan
cher
& S
eber
t) Lo
wry
, ine
d.
New
Cal
edon
ia
Bai
e de
Tin
a, N
ew C
aled
onia
(Low
ry 4
664,
MO
) IT
S: A
F229
694
5S IG
S: A
Y03
5564
trnL
-trn
F: A
F382
169
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
s N
ew C
aled
onia
Mt.
Dzu
mac
, New
Cal
edon
ia (L
owry
464
7, M
O)
ITS
: AF2
2969
65S
IGS
: AY
0355
67
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
sM
t. M
ou, N
ew C
aled
onia
(Low
ry 4
712,
MO
) IT
S: A
F382
940
5S IG
S: A
Y03
5568
trnL
-trn
F: A
F382
173
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
sM
t. M
ou, N
ew C
aled
onia
(Low
ry 4
713,
MO
) IT
S: A
F382
941
5S IG
S: A
Y03
5566
trnL
-trn
F: A
F382
170
Pol
ysci
ascf
. dio
ica
(Vie
ill. e
x P
anch
er &
Seb
ert)
Har
ms
Roc
hes
Oua
ïèm
e, N
ew C
aled
onia
(Low
ry 4
771,
MO
) IT
S: A
F229
709
5S IG
S (P
CR
pro
duct
onl
y)
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
sR
oche
s O
uaïè
me,
New
Cal
edon
ia (L
owry
478
1, M
O)
ITS
: AF3
8294
25S
IGS
: AY
0355
69tr
nL-t
rnF:
AF3
8217
1
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
sM
é O
ri, N
ew C
aled
onia
(Low
ry 4
800,
MO
) 5S
IGS
(PC
R p
rodu
ct o
nly)
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
sM
é O
ri, N
ew C
aled
onia
(Low
ry 4
804,
MO
) tr
nL-t
rnF:
AF3
8217
2
Pol
ysci
as d
ioic
a(V
ieill
. ex
Pan
cher
& S
eber
t) H
arm
s
Pla
teau
de
Tiéa
, New
Cal
edon
ia (L
owry
509
9, M
O)
ITS
: AF3
8294
35S
IGS
: AY
0355
65

Polyscias sect. Tieghemopanax (Araliaceae)
29ADANSONIA, sér. 3 • 2001 • 23 (1)
Pol
ysci
as “
dzum
acen
sis”
Low
ry, i
ned.
N
ew C
aled
onia
M
t. D
zum
ac, N
ew C
aled
onia
(Low
ry 4
650,
MO
) IT
S: A
F229
697
5S IG
S: A
Y03
5570
trnL
-trn
F: A
F382
174
Pol
ysci
as “
jaffr
ei”
Low
ry, i
ned.
N
ew C
aled
onia
K
opét
o M
assi
f, N
ew C
aled
onia
(Low
ry 4
740,
MO
) IT
S: A
F229
700
5S IG
S: A
Y03
5571
trnL
-trn
F: A
F382
176
Pol
ysci
as jo
skei
Gib
bsFi
ji O
vala
u, F
iji (A
.C. S
mith
758
4, U
S)
ITS
: AF3
8294
45S
IGS
: AY
0355
72tr
nL-t
rnF:
AF3
8217
7
Pol
ysci
as “
leca
rdii”
(R. V
ig.)
Low
ry, i
ned.
N
ew C
aled
onia
M
andj
élia
, New
Cal
edon
ia (L
owry
475
4, M
O)
ITS
: AF2
2970
15S
IGS
: AY
0355
73tr
nL-t
rnF:
AF3
8217
8
Pol
ysci
as m
icro
botr
ys(B
aill.
) Har
ms
New
Cal
edon
ia
Mé
Ori,
New
Cal
edon
ia (L
owry
480
2, M
O)
ITS
: AF2
2970
45S
IGS
: AY
0355
74tr
nL-t
rnF:
AF3
8218
1
Pol
ysci
as “
nitid
a” L
owry
, ine
d.N
ew C
aled
onia
Sar
ram
éa, N
ew C
aled
onia
(Low
ry 4
717A
, MO
) IT
S: A
F382
945
5S IG
S: A
Y03
5575
trnL
-trn
F: A
F382
179
Pol
ysci
as “
noth
isii”
Low
ry, i
ned.
N
ew C
aled
onia
N
ear P
ouem
bout
, New
Cal
edon
ia (L
owry
473
9, M
O)
ITS
: AF2
2970
75S
IGS
: AY
0355
76tr
nL-t
rnF:
AF3
8218
0
Pol
ysci
as p
anch
eri(
Bai
ll.) H
arm
s N
ew C
aled
onia
P
lain
e de
s La
cs, N
ew C
aled
onia
(Low
ry 4
693,
MO
) IT
S: A
F229
710
5S IG
S: A
Y03
5577
trnL
-trn
F: A
F382
182
Pol
ysci
as “
rega
lis”
Ber
nard
i ex
Low
ry, i
ned.
N
ew C
aled
onia
P
late
au d
e D
ogny
, New
Cal
edon
ia (L
owry
472
0, M
O)
ITS
: AF2
2971
25S
IGS
: AY
0355
78tr
nL-t
rnF:
AF3
8218
3
Pol
ysci
as s
ambu
cifo
lia(S
ieb.
ex
DC
.) H
arm
s A
ustr
alia
C
ultiv
ated
, Aus
tral
ian
Nat
l. B
ot. G
arde
n IT
S: A
F229
713
(acc
essi
on n
o.89
0062
4, v
ouch
er D
avie
s 12
72, C
BG
) 5S
IGS
: AY
0355
79tr
nL-t
rnF:
AF3
8218
4
Pol
ysci
as “
scop
olia
e” (B
aill.
) Low
ry, i
ned.
N
ew C
aled
onia
M
t. D
zum
ac, N
ew C
aled
onia
(Low
ry 4
638,
MO
) IT
S: A
F229
715
5S IG
S: A
Y03
5580
trnL
-trn
F: A
F382
185
Pol
ysci
as “
subi
ncis
a”(R
. Vig
.) Lo
wry
, ine
d.
New
Cal
edon
ia
Kou
mac
to T
iéba
ghi,
New
Cal
edon
ia (M
unzi
nger
361
, P)
5S IG
S: A
Y03
5581
trnL
-trn
F: A
F382
186
Pol
ysci
as “
subi
ncis
a”(R
. Vig
.) Lo
wry
, ine
d.
Man
djél
ia, N
ew C
aled
onia
(Low
ry 4
750,
MO
) IT
S: A
F229
718
Pol
ysci
as “
veill
onii”
Low
ry, i
ned.
N
ew C
aled
onia
M
t. O
uin,
New
Cal
edon
ia (L
owry
465
8, M
O)
ITS
: AF2
2972
15S
IGS
: AY
0355
82tr
nL-t
rnF:
AF3
8218
7

Eibl J.M., Plunkett G.M. & Lowry II P.P.
30 ADANSONIA, sér. 3 • 2001 • 23 (1)
Tab
le 1
.(co
ntd
).
Oth
er P
olys
cias
Pol
ysci
as “
abra
ham
iana
” Lo
wry
, ine
d.M
adag
asca
r A
ndas
ibe,
Mad
agas
car (
Laba
t 306
4, P
) IT
S: A
F229
686
Pol
ysci
as “
abra
ham
iana
” Lo
wry
, ine
d.A
ndas
ibe,
Mad
agas
car (
Laba
t 306
5, P
) 5S
IGS
: AY
0355
57tr
nL-t
rnF:
AF3
8214
8
Pol
ysci
as a
lber
sian
aH
arm
s Ta
nzan
ia
W U
sam
bara
Mts
., Ta
nzan
ia (L
owry
515
3, M
O)
ITS
: AF2
2968
7
Pol
ysci
as a
ustr
alia
na(F
. Mue
ll) P
hilip
son
Aus
tral
ia (Q
ueen
slan
d),
Nea
r Bel
lend
en K
er P
ark,
nor
ther
n Q
ueen
slan
d,
ITS
: AF2
2968
8N
ew G
uine
aA
ustr
alia
(Plu
nket
t 150
0, M
O)
Pol
ysci
as b
aret
iana
Ber
nard
i M
adag
asca
r M
asoa
la P
enin
sula
, Mad
agas
car (
Arid
y 28
1, M
O)
ITS
: AF2
2969
05S
IGS
: AY
0355
58tr
nL-t
rnF:
AF3
8214
9
Pol
ysci
as b
elle
nden
kere
nsis
(F.M
. Bai
ley)
Phi
lipso
n A
ustr
alia
(Que
ensl
and)
M
t. B
artle
Fre
re, n
orth
ern
Que
ensl
and,
Aus
tral
ia
ITS
: AF2
2969
1(P
lunk
ett 1
538,
MO
)
Pol
ysci
as c
umin
gian
a(C
. Pre
sl.)
Fern
.-Vi
ll.
Mal
esia
; wid
ely
cult.
P
o R
iver
, Bet
wee
n P
onér
ihou
en a
nd H
ouaï
lou,
IT
S: A
F229
695
New
Cal
edon
ia (L
owry
478
8, M
O)
Pol
ysci
as e
lega
ns(C
. Moo
re &
F. M
uell.
) Har
ms
Aus
tral
ia (Q
ueen
slan
d),
Sou
th M
issi
on B
each
, nor
ther
n Q
ueen
slan
d,IT
S: A
F229
698
New
Gui
nea
Aus
tral
ia (P
lunk
ett 1
495,
MO
) tr
nL-t
rnF:
AF3
8217
5
Pol
ysci
as fu
lva
(Hie
ron.
) Har
ms
Trop
ical
Afri
ca
Bvu
mba
Mts
., Zi
mba
bwe
(Low
ry 4
806,
MO
) IT
S: A
F229
699
5S IG
S: A
Y03
5559
trnL
-trn
F: A
F382
150
Pol
ysci
as fr
utic
osa
(L.)
Har
ms
Unk
now
n, li
kely
Mel
anes
ia;
Mitc
hell
& W
agst
aff (
1997
)IT
S: U
6319
1 w
idel
y cu
lt.
Pol
ysci
ascf
. gui
lfoyl
ei(W
. Bul
l) L.
H. B
aile
y U
nkno
wn,
like
ly M
elan
esia
; C
ultiv
ated
, Hon
olul
u B
otan
ical
Gar
den
ITS
: AF2
2968
5
wid
ely
cult.
(P
lunk
ett 1
357,
WS
)
Pol
ysci
as m
acgi
llivr
ayi(
See
m.)
Har
ms
Aus
tral
ia (Q
ueen
slan
d),
Cap
e K
imbe
rly, n
orth
ern
Que
ensl
and,
Aus
tral
ia
ITS
: AF2
2970
2
E M
ales
ia, S
olom
on Is
l.,
(Plu
nket
t 153
6, M
O)
Mic
rone
sia
Pol
ysci
as m
ayot
tens
isLo
wry
, O. P
asca
l & L
abat
Com
oro
Isl.
Saz
iley,
May
otte
, Com
oros
Isl.
(Lab
at 2
935,
MO
) IT
S: A
F229
703
Pol
ysci
as m
ollis
(Ben
th.)
Har
ms
Aus
tral
ia (Q
ueen
slan
d)
Bel
lend
en K
er P
ark,
nor
ther
n Q
ueen
slan
d, A
ustr
alia
ITS
: AF2
2970
5
(Plu
nket
t 150
7, M
O)
Pol
ysci
as m
urra
yi(F
. Mue
ll.) H
arm
s A
ustr
alia
(Que
ensl
and)
B
elle
nden
Ker
Par
k, n
orth
ern
Que
ensl
and,
Aus
tral
ia
ITS
: AF2
2970
6
(Plu
nket
t 150
5, M
O)
Pol
ysci
as “
orie
ntal
is”
Low
ry, i
ned.
M
adag
asca
r A
nkiri
ndro
, Mad
agas
car (
Sch
atz
3925
, MO
) IT
S: A
F229
708
Pol
ysci
as p
urpu
rea
C.T
. Whi
te
Aus
tral
ia (Q
ueen
slan
d)
Licu
ala
Sta
te F
ores
t Par
k, n
orth
ern
Que
ensl
and,
IT
S: A
F229
711
A
ustr
alia
(Plu
nket
t 149
6, M
O)
Pol
ysci
as“s
chat
zii”
Low
ry, i
ned.
M
adag
asca
r A
nkiri
ndro
, Mad
agas
car (
Sch
atz
3898
, MO
) IT
S: A
F229
714
Pol
ysci
as s
cute
llaria
(Bur
m. f
.) Fo
sber
g U
nkno
wn,
like
ly M
elan
esia
; C
ultiv
ated
, Fle
cker
Bot
anic
al G
arde
n, C
airn
es,
ITS
: AF2
2971
6
wid
ely
cult.
Q
ueen
slan
d, A
ustr
alia
(Plu
nket
t 149
1, M
O)

Polyscias sect. Tieghemopanax (Araliaceae)
31ADANSONIA, sér. 3 • 2001 • 23 (1)
Pol
ysci
as s
essi
liflo
raM
arai
s R
éuni
on
Col
de
Béb
our,
Réu
nion
(Low
ry 4
981,
MO
) IT
S: A
F229
717
Pol
ysci
as ta
hitie
nsis
(Nad
aud)
Har
ms
Tahi
ti Ta
hiti
(Flo
renc
e 12
911,
MO
) IT
S: A
F229
719
Pol
ysci
as te
nnan
tiiB
erna
rdi
Mad
agas
car
And
asib
e, M
adag
asca
r (La
bat 3
074,
P)
ITS
: AF2
2972
0
Oth
er A
ralia
ceae
Api
opet
alum
vel
utin
umB
aill.
N
ew C
aled
onia
M
t. M
ou, N
ew C
aled
onia
(Low
ry 4
700,
MO
) IT
S: A
F229
742
Ara
lia n
udic
aulis
L.
Nor
th A
mer
ica
Wen
& Z
imm
er (1
996)
IT
S: U
4167
4tr
nL-t
rnF:
AF3
8215
7
Art
hrop
hyllu
m “
mac
keei
”Lo
wry
, ine
d.
New
Cal
edon
ia
Hau
te Y
até,
New
Cal
edon
ia (L
owry
467
0, M
O)
ITS
: AF2
2973
6tr
nL-t
rnF:
AF3
8215
8
Del
arbr
ea p
arad
oxa
Viei
ll. s
sp. p
arad
oxa
Mal
esia
, Sol
omon
Isl.,
P
onan
dou
R.,
New
Cal
edon
ia (L
owry
476
6, M
O)
ITS
: AF2
2975
0Va
nuat
u, N
ew C
aled
onia
, N
orfo
lk Is
l.
Del
arbr
ea p
arad
oxa
Viei
ll. s
sp. p
arad
oxa
Kat
rikoi
n, N
ew C
aled
onia
(Low
ry 4
791,
MO
) tr
nL-t
rnF:
AF3
8215
2
Gas
toni
a cu
tispo
nga
Lam
. R
éuni
on
Cul
t., U
niv.
de
la R
éuni
on, R
éuni
on (L
owry
497
6, M
O)
ITS
: AF2
2972
2tr
nL-t
rnF:
AF3
8216
1
Gas
toni
a du
plic
ata
Thou
ars
ex B
aill.
M
adag
asca
r M
asoa
la P
enin
sula
, Mad
agas
car (
Arid
y 29
9, M
O)
trnL
-trn
F: A
F382
163
Gas
toni
a ro
drig
uesi
ana
Mar
ais
Rod
rigue
s Is
l. C
ultiv
ated
, Roy
al B
otan
ic G
arde
n K
ew
ITS
: AF2
2972
3(a
cc. 6
62-8
6.06
150)
tr
nL-t
rnF:
AF3
8216
2
Gas
toni
a sp
ecta
bilis
(Har
ms)
Phi
lipso
n N
ew G
uine
a, S
olom
on Is
l.,
Mt.
Isle
y, E
dmon
ton,
Que
ensl
annd
, Aus
tral
iatr
nL-t
rnF:
AF3
8216
4
Aus
tral
ia
(Plu
nket
t 153
7, M
O)
Mac
kinl
aya
mac
rosc
iade
a(F
. Mue
ll.) F
. Mue
ll.
Aus
tral
ia (Q
ueen
slan
d)
Cul
tivat
ed, H
untin
gton
Bot
anic
al G
arde
n IT
S: A
F229
744
S
an M
arin
o, C
alifo
rnia
(Plu
nket
t 136
5, W
S)
Mer
yta
bala
nsae
Bai
ll.
New
Cal
edon
ia
Pla
teau
de
Dog
ny, N
ew C
aled
onia
(Low
ry 4
733,
MO
) tr
nL-t
rnF:
AF3
8216
0
Mer
yta
denh
amii
See
m.
New
Cal
edon
ia
Mé
Ori,
New
Cal
edon
ia (L
owry
479
3, M
O)
ITS
: AF2
2972
5
Mer
yta
“lec
ardi
i”(R
. Vig
.) Lo
wry
, ine
d.
New
Cal
edon
ia
Riv
ière
Ble
ue P
ark,
New
Cal
edon
ia (L
owry
467
8, M
O)
ITS
: AF2
2972
4
Mer
yta
“ped
uncu
lata
”Lo
wry
, ine
d.
New
Cal
edon
iaR
iviè
re B
leue
Par
k, N
ew C
aled
onia
(Low
ry 4
756,
MO
) tr
nL-t
rnF:
AF3
8215
9
Mer
yta
sinc
lairi
i(H
ook.
f.) S
eem
.N
ew Z
eala
nd
Mitc
hell
& W
agst
aff (
1997
) IT
S: U
6319
4
Myo
doca
rpus
cra
ssifo
lius
Dub
ard
& R
. Vig
. N
ew C
aled
onia
Mt.
Dzu
mac
, New
Cal
edon
ia (L
owry
464
1, M
O)
ITS
: AF2
2975
1
Myo
doca
rpus
cra
ssifo
lius
Dub
ard
& R
. Vig
.M
t. M
ou, N
ew C
aled
onia
(Low
ry 4
704,
MO
) tr
nL-t
rnF:
AF3
8215
1
Opl
opan
ax h
orrid
us (J
. Sm
ith) M
iq.
Nor
th A
mer
ica
Pac
ific
Nor
thw
est,
Nor
th A
mer
ica
(Sou
le 3
821,
WS
) tr
nL-t
rnF:
AF3
8215
5
Ore
opan
ax s
ande
rianu
s H
emsl
. M
esoa
mer
ica
Cul
tivat
ed, M
isso
uri B
otan
ical
Gar
den,
no.
873
066
trnL
-trn
F: A
F382
156
(P
lunk
ett 1
343,
WS
)
Pse
udos
ciad
ium
bal
ansa
e B
aill.
N
ew C
aled
onia
M
t. M
ou, N
ew C
aled
onia
(Low
ry 4
714,
MO
) IT
S: A
F229
760
Sch
effle
ra a
ctin
ophy
lla(E
ndl.)
Har
ms
Aus
tral
asia
C
ultiv
ated
, New
Yor
k B
otan
ical
Gar
den
trnL
-trn
F: A
F382
153
(P
lunk
ett 1
316,
WS
)
Sch
effle
ra tr
eves
ioid
es H
arm
s S
. Chi
na, V
iet N
am
Fan
Si P
an M
tn.,
Viet
nam
(Low
ry 4
920,
MO
) IT
S: A
F229
732
trnL
-trn
F: A
F382
154

Sequence alignments were derived manually.Pairwise distances were calculated for each data setwith PAUP* (vers. 4; D. SWOFFORD, SmithsonianInst.) using the TAJIMA-NEI DNA distance algo-rithm (TAJIMA & NEI 1984). Treating gaps asmissing data, the data sets were analyzed usingPAUP* by maximum parsimony (MP) to inferphylogenetic relationships. Because our samplingfor each marker did not completely overlap, sepa-rate analyses were first performed for each of thethree data sets. For the ITS data, we sampledwidely throughout Araliaceae, following the find-ings of PLUNKETT et al. (2001) and WEN et al.(2001) in choosing outgroups. An initial searchusing 100 replicates (saving no more than 100 treesper replicate) was conducted because preliminarysearches yielded many tens of thousands of mostparsimonious trees. The strict consensus treebased on these 100 replicates was used as a topo-logical constraint (saving only those trees notagreeing with the constraint) for an additional1,000 replicates, each of which was aborted ifmore than 1,000 trees were found. After findingno additional topologies of equal or shorterlength, the shortest trees resulting from the firstsearch were loaded as starting trees and swappedto completion saving no more than 10,000 trees.For both the trnL-trnF and 5S data sets, heuristicsearches (each with 100 random addition repli-cates) were performed. Because the number ofmost parsimonious trees recovered was relativelylow, it was unnecessary to limit the number oftrees saved. Outgroup selection for these two datasets was based on results of the ITS analyses(herein, PLUNKETT et al. 2001, and WEN et al.2001).
Previous studies (e.g., OLMSTEAD & SWEERE
1994; SOLTIS et al. 1998) have found that analy-ses of combined data sets can provide increasedphylogenetic signal, resulting in both higherinternal support (e.g., bootstrap percentages) anddecreased computer run times. However, treat-ment of multiple data sets remains controversial(see DE QUEIROZ et al. 1995; CANNATELLA et al.1998). Biological sources of incongruence (e.g.,non-dichotomous patterns of diversification, lin-eage sorting of ancestral polymorphisms, and rateheterogeneity in sequence evolution) are amongthe potential problems that may produce mis-
leading results in phylogenetic analyses based oncombined matrices. Conversely, such incongru-ence may also result from sampling error, eitheramong taxa or among characters. There is littleagreement in the literature regarding the bestmethods of testing for congruence (reviewed inJOHNSON & SOLTIS 1998), or even the basic cri-terion that should be tested, although most work-ers agree that a null hypothesis of congruence ordata homogeneity must be explicitly rejected(e.g., BULL et al. 1993; MASON-GAMER &KELLOGG 1996; JOHNSON & SOLTIS 1998). Toassess congruence, we constructed separate datasets for each of the three markers using an identi-cal set of 24 taxa (to eliminate taxon samplingerror). Each of these was analyzed separately andin a single combined analysis. Congruence wasestimated using the incongruence test (IMF) ofMICKEVITCH & FARRIS (1981), calculated manu-ally, and the partition homogeneity test of FARRIS
et al. (1995) as implemented in PAUP*. Resultsfrom these tests (see below) suggest that the datasets are combinable. Therefore, all pairwise com-binations were assembled, maximizing taxonsampling in each case. Combined data sets wereanalyzed in the same manner as the separateanalyses of trnL-trnF and 5S.
Confidence in individual clades for all treeswas estimated using bootstrap analyses (100 repli-cates, saving no more than 1,000 trees per repli-cate) (FELSENSTEIN 1985). Fitness indices andnucleotide compositions for each data set wereassessed using MacClade (MADDISON &MADDISON 1992) and/or PAUP*. In addition,distance analyses based on TAJIMA-NEI estima-tions were used to determine relative levels ofvariation among a representative sample ofsequences from the ITS, trnL-trnF, and 5S matrices.
RESULTS
Matrix comparisonsITS sequence data represented the entire ITS1
and ITS2 non-coding regions and the intervening5.8S coding region from 58 taxa. This matrix re-quired approximately 41 alignment gaps andcomprised an aligned length of 666 bp, 363 ofwhich were constant, 109 variable in only one
Eibl J.M., Plunkett G.M. & Lowry II P.P.
32 ADANSONIA, sér. 3 • 2001 • 23 (1)
taxon, and 194 potentially informative (Table 2).The trnL-trnF data matrix (40 taxa) representedthe entire length of both the trnL intron and thetrnL-trnF intergenic spacer (plus flanking se-quences from the adjacent coding regions). Thesesequences required approximately 27 alignmentgaps, resulting in an aligned length of 952 bp, 840of which were constant, 56 variable in onlyone taxon, and 56 potentially informative.Representing the entire non-transcribed spaceralong with short flanking regions of the gene, the5S rDNA matrix required approximately 17 gapsto align the matrix of 26 sequences. After align-ment, these sequences were 301 bp long, contain-ing 199 constant characters, 51 characters variablein only one taxon, and 51 characters potentiallyinformative for parsimony. Table 2 also providesother metrics pertaining to the data sets and parsi-mony trees (i.e., lengths of the shortest trees, val-ues of the consistency and retention indices, andnucleotide composition).
Rates of sequence evolution, based on TAJIMA-NEI pairwise distances, were greatest in the ITSdata set, which contained the broadest samplingof araliad taxa outside of the Tieghemopanaxgroup. Overall, the highest level of variation wasbetween sequences of the outgroup taxaMackinlaya macrosciadea and Myodocarpus crassi-folius (38.2%). In the same data set, the greatestvariation among species currently assigned to the
genus Polyscias was 10.4% (between P. fruticosaand P. fulva). Among the species of sectionTieghemopanax this value was 6.3% (betweenP. sambucifolia and P. joskei). The lowest level ofsequence variation among all taxa in the ITS dataset was identity, found in two pairs of P. dioicasamples. In the trnL-trnF data set, the greatestlevel of variation was 7.4% between sequences ofMyodocarpus crassifolius and Gastonia rodrigues-iana. Across Polyscias, the greatest variation was1.4% between P. fulva and P. joskei; within sec-tion Tieghemopanax, this value was 1.3%,between P. sambucifolia and P. joskei. Several pairsof taxa both within and outside of sectionTieghemopanax shared values of identity. In the5S spacer data set, the greatest overall levels ofvariation were between P. fulva and P. “bracteata”1
(19.9%), and within section Tieghemopanaxbetween P. sambucifolia and P. “bracteata”(13.7%). Four species pairs shared values of iden-tity. Comparing only taxa found across all threedata sets (the same 24 taxa used for the congru-ence tests), ITS sequences and 5S spacer sequen-ces have evolved on average at rates 6.2 and16.4 times greater (respectively) than trnL-trnF.Although trnL-trnF provides few variable charac-ters, they are highly consistent. For example, in
Polyscias sect. Tieghemopanax (Araliaceae)
33ADANSONIA, sér. 3 • 2001 • 23 (1)
1. Names in quotation marks and indicated as “ined.” in Table 1 willbe published elsewhere.
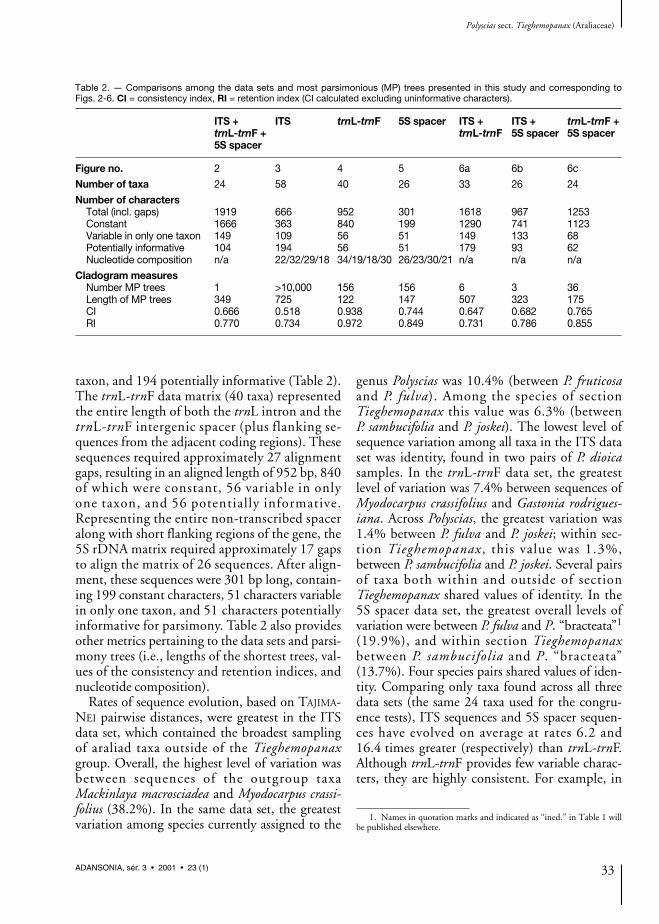
Table 2. — Comparisons among the data sets and most parsimonious (MP) trees presented in this study and corresponding toFigs. 2-6. CI = consistency index, RI = retention index (CI calculated excluding uninformative characters).
ITS + ITS trnL-trnF 5S spacer ITS + ITS + trnL-trnF +trnL-trnF + trnL-trnF 5S spacer 5S spacer5S spacer
Figure no. 2 3 4 5 6a 6b 6c
Number of taxa 24 58 40 26 33 26 24
Number of charactersTotal (incl. gaps) 1919 666 952 301 1618 967 1253Constant 1666 363 840 199 1290 741 1123Variable in only one taxon 149 109 56 51 149 133 68Potentially informative 104 194 56 51 179 93 62Nucleotide composition n/a 22/32/29/18 34/19/18/30 26/23/30/21 n/a n/a n/a
Cladogram measuresNumber MP trees 1 >10,000 156 156 6 3 36Length of MP trees 349 725 122 147 507 323 175CI 0.666 0.518 0.938 0.744 0.647 0.682 0.765RI 0.770 0.734 0.972 0.849 0.731 0.786 0.855

Eibl J.M., Plunkett G.M. & Lowry II P.P.
34 ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig.
1(a
-c).
—S
tric
t co
nsen
sus
tree
s re
sulti
ng fr
om s
epar
ate
anal
yses
of I
TS, t
rnL-
trnF
, and
5S
sp
acer
dat
a, e
ach
with
an
iden
tical
24-
taxo
n sa
mp
ling.
Val
ues
alon
g no
des
are
boo
t-st
rap
per
cent
ages
; nod
es w
ith d
ashe
d b
ranc
hes
have
boo
tstr
aps
bel
ow 7
0%. C
lad
es c
onsi
sten
tly r
ecov
ered
in th
ese
tree
s (a
nd in
Fig
. 2) a
re la
bel
ed A
-E, a
lthou
gh c
lad
e C
in th
e IT
Str
ee a
nd c
lad
e E
in
the
trnL
-trn
F w
ere
left
unr
esol
ved
(d
ashe
d b
rack
ets)
. P
lace
men
t of
Pol
ysci
as s
amb
ucifo
lia a
nd P
. m
icro
bot
rys
(dis
cuss
ed i
n te
xt)
are
den
oted
by
an a
ster
isk.
Out
grou
ps
ind
icat
ed b
y “O
G.”
— (a
) ITS
ana
lysi
s: s
tric
t con
sens
us o
f 94
mos
t par
sim
onio
us (M
P) t
rees
; tre
e le
ngth
= 1
66 s
tep
s; c
onsi
sten
cy in
dex
= 0
.600
; ret
entio
n in
dex
= 0
.714
.—(b
) trn
L-tr
nF a
naly
sis:
str
ict
cons
ensu
s of
3 M
P t
rees
; tre
e le
ngth
= 2
9 st
eps;
con
sist
ency
ind
ex =
0.9
23; r
eten
tion
ind
ex =
0.9
68.—
(c) 5
S-s
pac
er a
naly
sis:
str
ict
cons
ensu
s of
54
MP
tree
s; t
ree
leng
th =
146
ste
ps;
con
sist
ency
ind
ex =
0.5
56; r
eten
tion
ind
ex =
0.6
64.

the 24-taxon data set, there were only 12 parsi-mony informative trnL-trnF characters, but onlyone of these was homoplasious, yielding veryhigh consistency values for the resulting trees(CI = 0.923, RI = 0.968). Thus, while at thelower limit of the phylogenetic utility of trnL-trnF, these data appear to make a significantcontribution to the present study.
Congruence, resolution, and bootstrap supportTests for congruence among the three 24-taxon
data sets suggest that the data are homogeneousand therefore combinable. For the three-matrixcomparison, the partition homogeneity test(FARRIS et al. 1995) yielded a result of p = 0.22(only values of p ≤ 0.05 indicate significantheterogeneity). In pairwise comparisons of trnL-trnF vs. ITS and trnL-trnF vs. 5S spacers, p = 1(no heterogeneity). However, in the comparisonof the ITS versus 5S spacer data, results suggestedsignificant heterogeneity (p = 0.04). The incon-gruence test of MICKEVITCH & FARRIS (1981)also suggested no incongruence in pairwise com-parisons of either nuclear set to the plastid dataset (IMF = 0 in both cases). Some incongruencewas found between the two nuclear sets, but thiswas rather low (IMF = 12.7%).
Although evidence for incongruence is equivo-cal, we followed the protocol of BULL et al.(1993) and SEELANAN et al. (1997) in an attemptto investigate the sources of the possible incon-gruence between the two nuclear data sets. Themost common biological source of incongruencein plant groups is reticulate evolution involvinghybridization and/or polyploid speciation, result-ing in the lateral transfer of genes or genomes.Our results do suggest that reticulation mayexplain at least in part the evolutionary origin ofPolyscias dioica, a finding supported by theunusually broad morphological variability foundin this species. Examination of all cladograms,however, indicates that possible reticulationinvolving P. dioica is not the source of incongru-ence in our data. In fact, all three separate analy-ses (Fig. 1) agree in resolving a single cladecomprising all samples of P. dioica and the singlesample of P. cissodendron. Apart from P. dioica(and P. cissodendron), evidence for reticulation insection Tieghemopanax is likewise scant.
We visually inspected the cladograms (Fig. 1)to identify other potentially problematic taxa insection Tieghemopanax. Only the placements ofP. microbotrys and P. sambucifolia differedmarkedly from one analysis to another, butremoval of these taxa did not improve the parti-tion homogeneity score. Although combining thedata sets based on the two nuclear markers mayseem imprudent, results from this analysis indi-cate that incongruence between ITS and 5S spac-ers is likely due to sampling error rather than anunderlying biological cause. Apart from poor res-olution, examination of well-supported clades(e.g., bootstrap = 70%) reveals no disagreementamong the separate trees (Fig. 1). When thesedata sets are combined, a single most parsimo-nious tree is recovered (identical in topology toFig. 2) in which all of the well-supported cladesare again resolved (with a single exception: aminor re-arrangement of terminal taxa in theclade labeled “A” in the 5S spacer tree; cf.Figs. 1c, 2). In all but three cases, bootstrap sup-port for these same clades is higher in the com-bined tree than in the separate trees. Congruencebetween ITS and 5S spacer matrices is furthersupported by increases in two standard measuresof homoplasy, the consistency (CI) and retention(RI) indices. For the separate (24-taxon) datasets, the CI and RI were (respectively) 0.6 and0.714 for ITS, and 0.556 and 0.664 for the 5Sspacers, but the combined (ITS + 5S spacer)analysis yielded higher values (CI = 0.644, RI =0.745), indicating lower levels of homoplasy.Overall, these results suggest that the phyloge-netic signal present in each of the separate datasets is additive when combined, effectively reduc-ing levels of random homoplasy or “backgroundnoise” (see CHIPPENDALE & WIENS 1994;OLMSTEAD & SWEERE 1994; MASON-GAMER &KELLOGG 1996). Therefore, we present both sep-arate and combined trees based on all availabledata for a total evidence approach to assessingrelationships (KLUGE & WOLFE 1993).
Phylogenetic relationshipsThe 24-taxon data matrices (and their trees;
Fig. 1) were designed to test congruence amongthe three separate data sets. Since these matricesrepresented reductions of the full data sets in each
Polyscias sect. Tieghemopanax (Araliaceae)
35ADANSONIA, sér. 3 • 2001 • 23 (1)

the maximum limit of 10,000 trees was met, butno topologies of equal or shorter length could befound that did not agree with the strict consensus
Eibl J.M., Plunkett G.M. & Lowry II P.P.
36 ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig. 2. — The single most parsimonious tree resulting from the combined analysis of ITS + trnL-trnF + 5S spacer sequences, basedon the identical 24-taxon sampling as used in Fig. 1; tree length = 349 steps; consistency index = 0.663; retention index = 0.770.Values along nodes are bootstrap percentages; nodes with dashed branches have bootstraps below 70%. Identical or near identicalclades (labeled A-E) from Fig. 1 are also labeled, as is the placement of Polyscias sambucifolia and P. microbotrys (asterisks), as dis-cussed in text. Outgroups indicated by “OG.”
case, we shall hereafter discuss only those resultsthat are based on the full data sets (Figs. 3-5) foreach separate analysis. In the analysis of ITS data,

tree, suggesting that this strict tree is a reasonableestimation of all shortest length trees. For theremaining analyses, fewer than 10,000 trees werefound, allowing all analyses to be completedwithout tree limits. Analyses of the trnL-trnF and5S matrices each yielded 156 most parsimonioustrees. The combined analyses of ITS + trnL-trnF,ITS + 5S, and trnL-trnF + 5S yielded 6, 3, and36 most parsimonious trees, respectively. In thecombined analyses of all three data sets, a single,fully-resolved tree was found.
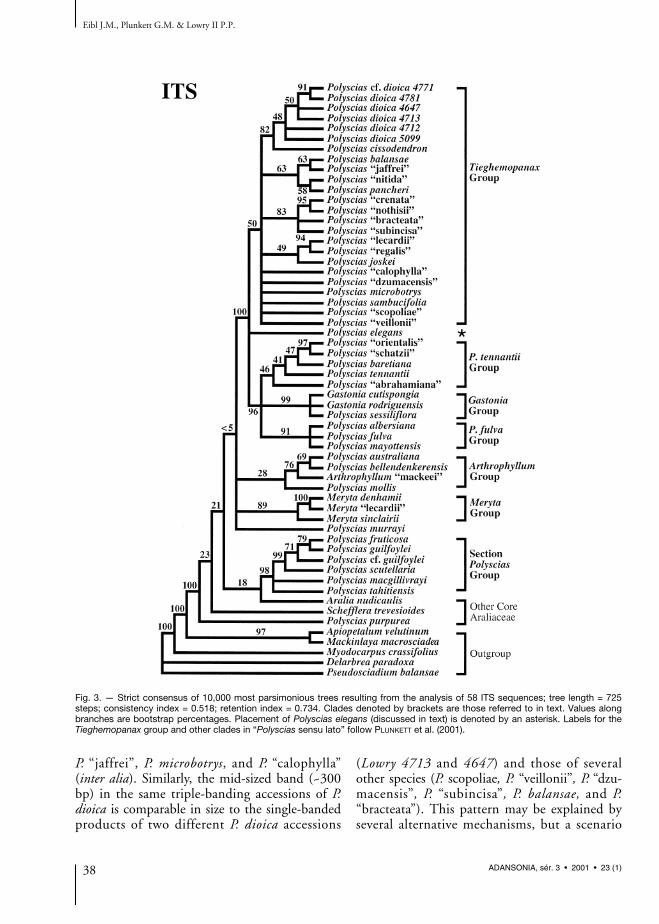
The strict consensus resulting from analysis ofITS data (Fig. 3) includes a broad samplingof species from throughout Araliaceae. Thistree depicts the same major clades recoveredby PLUNKETT et al. (2001), including theTieghemopanax, P. tennantii, Gastonia, P. fulva,Arthrophyllum, Meryta, and Polyscias sectionPolyscias groups, which together comprise the“Polyscias sensu lato” clade. The recovery of mostof these same clades in the parsimony analysis ofthe separate trnL-trnF data set helps to demon-strate the congruence of ITS and trnL-trnFtopologies (see Figs. 3, 4). Congruence betweentopologies based on all three separate analyses(ITS, trnL-trnF, 5S) is evident in the recovery ofmany identical or near identical clades, such asthe sister relationship between sectionTieghemopanax and Polyscias baretiana andbetween the P. “lecardii”/P. “regalis” species pair,among many others (Figs. 3-5). Other relation-ships demonstrate the congruence (or at least anabsence of conflict) between these topologieswhen polytomies are taken into consideration.For example, the P. “scopoliae”/P. “veillonii”species pair is well resolved in trees based on sepa-rate trnL-trnF and 5S data (Figs. 4, 5); becausethis relationship is left unresolved in the treesbased on ITS sequences (Fig. 3), the ITS data setneither confirms nor refutes it.
Topologies resulting from the combined dataset (ITS + trnL-trnF; Fig. 6a) confirm the mono-phyly of section Tieghemopanax, supported by abootstrap value of 63%. However, outgroupsampling in this pairwise combined tree waslimited, and bootstrap values supporting themonophyly of section Tieghemopanax in the sep-arate trees are either lower (50% in the ITS tree;Fig. 3) or the clade fails to resolve (trnL-trnF
tree, although the clade comprising Tieghe-mopanax species + P. elegans is supported by abootstrap of 89%; Fig. 4). These trees also helpto confirm the paraphyly of Polyscias sensu lato(PLUNKETT et al. 2001), with taxa from severalother genera appearing within the broad Polysciasclade (Figs. 3, 4, 6a).
Banding patterns and sequence length varia-tion in 5S spacer sequences of Tieghemopanaxaccessions
Multiple-banding patterns of 5S PCR ampli-cons were observed in several accessions of taxain the Tieghemopanax group, principally in some(but not most) accessions of P. dioica (Lowry4771, 4800, 4804) and a single accession eachof P. elegans (Plunkett 1495), P. cissodendron(Lowry 4749), and P. pancheri (Lowry 4793).The remaining Tieghemopanax accessions(including all other P. dioica samples) yieldedonly a single 5S PCR product. Individual bandsfrom P. dioica (Lowry 4804) and P. elegans(Plunkett 1495) were gel-purified and cloned,but attempts to sequence these cloned productsfailed to produce clean sequence data. Therefore,these samples were not included in any of thephylogenetic analyses. Nevertheless, the resulting“dirty” sequences were compared to all sequencesdeposited in GenBank to assess their homologyto the 5S rDNA region. The highest scoresreturned were in all cases for matches withangiosperm 5S sequences (from the flankingcoding regions).
In general, length variability of 5S spacerproducts was very high throughout Tieghe-mopanax, ranging from ~220 bp to ~360 bp.Thus, despite the difficulties in obtaining usablesequence data from samples yielding multiplePCR bands, a visual inspection and comparisonof these sizes may provide some insights. Forexample, two of the bands in the triple-bandedproducts resulting from three P. dioica samples(accessions Lowry 4771, 4800, and 4804) arevisually similar in size to the single-bandedamplicons resulting from other accessions ofP. dioica and several other species. The largest ofthese bands (~350 bp) is similar in size to thesingle band resulting from two P. dioicaaccessions (Lowry 4712 and 4781) and from
Polyscias sect. Tieghemopanax (Araliaceae)
37ADANSONIA, sér. 3 • 2001 • 23 (1)
P. “jaffrei”, P. microbotrys, and P. “calophylla”(inter alia). Similarly, the mid-sized band (~300bp) in the same triple-banding accessions of P.dioica is comparable in size to the single-bandedproducts of two different P. dioica accessions
(Lowry 4713 and 4647) and those of severalother species (P. scopoliae, P. “veillonii”, P. “dzu-macensis”, P. “subincisa”, P. balansae, and P.“bracteata”). This pattern may be explained byseveral alternative mechanisms, but a scenario
Eibl J.M., Plunkett G.M. & Lowry II P.P.
38 ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig. 3. — Strict consensus of 10,000 most parsimonious trees resulting from the analysis of 58 ITS sequences; tree length = 725steps; consistency index = 0.518; retention index = 0.734. Clades denoted by brackets are those referred to in text. Values alongbranches are bootstrap percentages. Placement of Polyscias elegans (discussed in text) is denoted by an asterisk. Labels for theTieghemopanax group and other clades in “Polyscias sensu lato” follow PLUNKETT et al. (2001).

involving hybridization either among variableP. dioica individuals or between P. dioica andother members of section Tieghemopanax couldproduce these results. Future work is needed toaddress the issue more fully.
DISCUSSION
The monophyly of Polyscias sect. Tieghemopanaxwas confirmed by the combined analysis of ITS +trnL-trnF data (Fig. 6a; bootstrap 63%) and in
Polyscias sect. Tieghemopanax (Araliaceae)
39ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig. 4. — Strict consensus of 156 most parsimonious trees resulting from the analysis of 40 trnL-trnF sequences; tree length = 122steps; consistency index = 0.938; retention index = 0.972. Values along branches are bootstrap percentages. Placement ofPolyscias elegans (discussed in text) is denoted by an asterisk. Labels for the Tieghemopanax group and other clades in “Polysciassensu lato” follow PLUNKETT et al. (2001).

the ITS tree (Fig. 3; bootstrap 50%). In thetrnL-trnF tree (Fig. 4), the lineages of sectionTieghemopanax form a large polytomy with P. ele-gans, supported by a bootstrap of 89%. Thus,despite the paraphyly evident in Polyscias as awhole, section Tieghemopanax appears to form amonophyletic group that is both morphologicallyand geographically coherent. The relationshipbetween these New Caledonian species and twoAustralian taxa examined, P. elegans and P. sam-bucifolia, are poorly resolved in the separateanalyses of ITS and trnL-trnF (Figs. 3, 4), but thecombined analysis of these two data sets (Fig. 6a)provides a clearer picture of relationships andaffords important insights into the biogeographichistory of Tieghemopanax. Specifically, theAustralian P. elegans is sister to the entireTieghemopanax group, a result supported by abootstrap of 86%. This finding is mirrored inmany other major clades of Polyscias sensu lato(e.g., the section Polyscias and Arthrophyllumgroups of PLUNKETT et al. 2001), and is consis-tent with the idea that the entire genus Polysciasoriginated in Australasia. Specifically, it seemsl ike l y tha t the common ance s to r o fTieghemopanax arrived in New Caledoniathrough a single long-distance dispersal eventafter the island’s separation from Australia.However, because we have no reliable estimate forthe age of these lineages, we cannot rule out amore ancient vicariance event. Multiple dispersalsfrom Australia are also conceivable, although thisscenario would be less parsimonious than either asingle dispersal or an ancient vicariance.
Within Tieghemopanax, only four taxa occuroutside New Caledonia. Three species (P. joskei,P. schmidii, and P. sambucifolia) are found exclu-sively in other nearby archipelagos, and thefourth taxon, P. cissodendron, is native to NewCaledonia but is also found on islands both tothe north and south. Relationships among thesespecies and the New Caledonian endemics pro-vide preliminary insights into the biogeographicrelationships within section Tieghemopanax.Polyscias sambucifolia is endemic (but widelyranging) in eastern Australia. Its placement in themolecular trees is somewhat unstable (cf. Figs. 1-6),and even in those cladograms where its positionis resolved, bootstrap support is very weak. Two
placements, however, recur in several trees: assister to all remaining members of sectionTieghemopanax (Figs. 1c, 5, 6c) or as sister to theP. dioica + P. cissodendron clade (Figs. 1a, 2, 6a-b).Two alternative scenarios can thus be invoked forP. sambucifolia: either it represents an earlybranching lineage of the Tieghemopanax cladethat remained in Australia (as did P. elegans), or itarrived there through secondary long-distancedispersal, most likely from New Caledonia.Without a broader consensus, we are unable todistinguish between these two hypotheses. Forthe remaining species, however, our results pro-vide a clearer picture. For example, P. cissoden-dron, the most widespread member of the group,ranges from the Santa Cruz islands (southernSolomon Islands) in the north through severalislands of Vanuatu (Vanua-Lava, Espiritu Santo,Erromanga, Tanna, and Aneityum) across NewCaledonia, and thence farther south to LordHowe Island (LOWRY 1989). Apart from NewCaledonia, which is continental in origin, all ofthese are relatively young volcanic islands (lessthan 30 MY old, and most younger than 7 MYold; see MUELLER-DOMBOIS & FOSBERG 1998and references therein). The age of these islands,coupled with the position of P. cissodendron in themolecular trees as sister to or nested within aclade of the New Caledonian endemic P. dioica,suggests a relatively recent series of dispersalsboth north and south from New Caledonia to theother islands.
The origin of the Fijian endemic P. joskei islikewise probably the result of long-distance dis-persal. Although the major islands of Viti Levuand Vanua Levu, where this P. joskei occurs, havebeen emergent for perhaps 30 MY, they havenever been in close proximity to New Caledonia.The placement of P. joskei as sister to the NewCaledonian endemic clade of P. “lecardii” +P. “regalis” (in all trees resolving the placement ofP. joskei; Figs. 1a, 2, 3, 6a-b), nested well withinthe Teighemopanax group, can be explained onlyby dispersal. Morphologically P. joskei closelyresembles P. “lecardii” and P. “regalis”, with whichit shares a number of distinctive characters. Forthe last species, P. schmidii, we have no moleculardata. On the basis of its morphology, however, itappears that this Vanuatu endemic is most closely
Eibl J.M., Plunkett G.M. & Lowry II P.P.
40 ADANSONIA, sér. 3 • 2001 • 23 (1)

related to P. “nitida” or P. “nothisii”, bothendemic to New Caledonia (LOWRY 1989), sug-gesting a third independent dispersal from NewCaledonia to nearby island systems. Perhaps whatis most surprising is the apparent rarity of suc-cessful dispersal events from New Caledonia toother islands involving members of theTieghemopanax group. It appears that P. cissoden-dron has experienced several independentdispersals recently enough that there is little ifany morphological differentiation among thewidely distributed populations (LOWRY 1989).Yet, this species has no apparent traits that mightsuggest an increased capacity for dispersal ascompared to other members of sectionTieghemopanax. In-depth studies of both the phy-logeny and natural history (e.g., identification ofanimal dispersers and their migration patterns)will be needed to address this issue.
In addition to broad-scale implications, themolecular analyses also provide a unified picture ofrelationships within section Tieghemopanax. Inparticular, nearly every tree resolves a series of iden-tical species pairs, each of which is also supportedby several morphological characters. Examinationof each species pair offers useful insights into themorphological characters that unite and distin-guish these species and the possible modes throughwhich speciation may have occurred.
The Polyscias “lecardii”/Polyscias “regalis”species pair
This species pair is characterized by inflores-cence features (flowers arranged in racemules)and a very similar leaf morphology (ovate to sub-orbicular, subcoriaceous leaflets having thickenedmargins). Leaflets of both species typically turn adistinctive orange-brown color when dried.Polyscias “lecardii”, however, has glabrous ovaries,calyces that form a low crown or rim, and flowerswith distinct short pedicels, whereas P. “regalis”has densely puberulent ovaries, calyces with fiveirregular, scarious, laciniate teeth, and sessileflowers. The sister-group relationship betweenP. “lecardii” and P. “regalis” is well resolved ineach of the seven analyses, with bootstrap supportranging from 63-100% (Figs. 2-6). Bootstrap val-ues from 94-100% were found in the separate
ITS analysis and in three of the combined analy-ses (Figs. 2, 3, 6a-b). These two species occur ingeographically disjunct regions (separated by c.80-100 km). Populations of P. “lecardii” grow onnon-ultramafic substrates in several parts of NewCaledonia at middle to high elevations (600-1000 m). Conversely, P. “regalis” populations areknown from only two locations very near oneanother (i.e., Plateau de Dogny and Koindé, bothin south central New Caledonia), also occurringon non-ultramafic soils. In analyses includingITS data (Figs. 1a, 2, 3, 6a-b), P. joskei from Fiji issister to the P. “lecardii”/ P. “regalis” species pair.Although bootstrap support for this relationshipsis generally low (65% or lower), P. joskei sharesmost of the distinctive features mentioned above.It closely resembles P. “lecardii”, but can be dis-tinguished by its thinner, membranaceous leafletswith more distinct venation, stouter secondaryinflorescence axes, and styles that are longer infruit.
The Polyscias “scopoliae”/Polyscias “veillonii”species pair
These species share several features, including adioecious mating system, young inflorescencessurrounded by large, caducous, cataphyll-likebracts, leaflets that are nearly succulent (obscur-ing venation on the adaxial surface), sparsebranching, and similar stature (up to 4 m high).Polyscias “scopoliae”, however, has unifoliolateleaves, whereas those of P. “veillonii” are three toseven foliolate. Bootstrap values ranging from 83-100% (Figs. 2, 4, 5, 6a-c) offer significant supportfor the sister relationship between P. “scopoliae”and P. “veillonii”. The species in this pair are dis-tinctly isolated for each other, with P. “scopoliae”restricted to dry, low elevation maquis vegetationon ultramafic substrates of the lower Tontoutaand Dumbéa river valleys and on Mt. Dzumac(elevations up to c. 350 m). Polyscias “veillonii”also occurs on ultramafics, but ranges from 500-1,300 m elevation in more mesic conditions onseveral mountains (e.g., Mt. Humboldt, Mt.Dzumac, Montagne des Sources, Mt. Mou, andProkoméo), with one population at 250 m inthe upper Yaté area. The valleys where P. “scopo-liae” occurs are situated just below the mountainslopes on which populations of P. “veillonii” are
Polyscias sect. Tieghemopanax (Araliaceae)
41ADANSONIA, sér. 3 • 2001 • 23 (1)

found, separated in some cases by just a few kilo-meters. Despite their geographical proximity,these two species are clearly distinguished by ele-vation (low vs. high), ecology (dry vs. more mesicconditions) and morphology (unifoliolate vs. 5- to 7-foliolate leaves).
The Polyscias “bracteata”/Polyscias “subincisa”species pair
Both of these taxa are characterized by flowersborne in heads and inflorescences with small, per-sistent bracts. Polyscias “bracteata” can be distin-guished by its hermaphroditic flowers, dense,
Eibl J.M., Plunkett G.M. & Lowry II P.P.
42 ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig. 5. — Strict consensus of 156 most parsimonious trees resulting from the analysis of 26 5S spacer sequences; tree length = 147steps; consistency index = 0.744; retention index = 0.849. Values along branches are bootstrap percentages.

spherical heads with (15-)20-30 flowers, stoutpeduncles, and large leaves with only two leafletsper node along the rachis, whereas P. “subincisa”has a dioecious mating system, open, hemispheri-cal heads with 5-10(-12) flowers, slender pedun-cles, and smaller leaves that often have reducedsecondary leaflets at each rachis node. The sister-group relationship of these species is resolvedonly in trees based on the separate 5S analysis andthree of the four combined analyses (ITS + 5S,trnL-trnF + 5S, and ITS + trnL-trnF + 5S), butbootstrap support in these trees was 100% in allcases (Figs. 2, 5, 6b-c). Polyscias “bracteata” iswidespread in maquis vegetation and disturbedforest and forest edges, occurring mostly on ultra-mafic substrates, although it is also known tooccur on various other substrates in the north-eastern part of New Caledonia and in the adja-cent Loyalty Islands from low to fairly highelevations (up to 1000 m). Also found in maquis,P. “subincisa” grows mostly on ultramafic sub-strates but is concentrated in the northwesternregion of the island. Like P. “bracteata,” P. “subin-cisa” can also occur on other substrates, but islimited to elevations below 200 m. These speciesshare little or no range overlap.
The Polyscias “crenata”/Polyscias “nothisii”species pair
Both species in this pair are dioecious andshare several morphological characters such asdensely puberulent inflorescences and large leaveswith numerous thin leaflets. In addition, individ-uals of these species are branched trees (5-8 mtall) with similar overall habits. Polyscias “crenata”differs in having puberulent petioles, sessilefemale flowers, and sub-ovoid fruits that are ellip-tical in cross-section, whereas P. “nothisii” hasglabrous petioles, female flowers that are dis-tinctly pedicellate, and fruits that are stronglyflattened laterally. The sister relationship betweenP. “crenata” and P. “nothisii” is well supported inall strict trees (Figs. 2, 3, 5, 6) except that of thetrnL-trnF data (Fig. 4). Bootstrap values rangedfrom 95-100%. Polyscias “crenata” is known tooccur in sclerophyllous and transition forests inwestern New Caledonia on calcareous substratesat elevations ranging from sea level to 500 m.Polyscias “nothisii” has a similar ecological range,
but it is known from only 3 localities at lowelevation (less than 50 m) in western NewCaledonia.
The Polyscias balansae/Polyscias “jaffrei” spe-cies pair
Shared features between these species includetheir habit (monocaulous to sparsely branchedtreelets 1-2 m tall), a dioecious mating system,and similar leaf morphology (subcoriaceousleaflets that are ovate to sub-orbicular in shape).Polyscias balansae differs in having hermaphro-ditic flowers in umbellules and male flowers thatare 5-merous, whereas P. “jaffrei” has herma-phroditic flowers in racemules and male flowersthat are 4-merous. The sister-group relationshipbetween these species is well resolved in only twostrict trees (Figs. 3, 6a) with only moderate boot-strap support (63% in both cases). In other trees,these species come out together but in largerclades of five taxa. These species are geographi-cally disjunct, with P. balansae occurring just tothe south of the southern limit of P. “jaffrei”.
The Polyscias dioica/Polyscias cissodendronspecies pair
These two species share a dioecious mating sys-tem, flowers in umbellules, fruits that are 2-5 mmlong, slender pedicels 0.4-0.7 mm long, calycesthat form a low crown or rim, and inflorescencebracts 1-4 mm long. Polyscias dioica differs inhaving coriaceous to subcoriaceous leaves, entireto coarsely dentate and often minutely revoluteleaflet margins, persistent inflorescence bracts,fruits (3.5-)4-5 mm long, and styles of femaleflowers and fruits united for less than half theirlength, whereas P. cissodendron has papyraceousleaves, coarsely crenulate leaflet margins,caducous inflorescence bracts, fruits 2-3(-3.3)mm long, and styles of female flowers and fruitsunited for more than half of their length. Therelationship between P. dioica and P. cissodendronis supported by all strict trees, with bootstrap val-ues between 64% and 99% (Figs. 2-6). Sometrees show a sister-group relationship betweenthe two species (Figs. 3, 6a) whereas others nestP. cissodendron within the P. dioica clade (Figs. 2,5, 6b-c). These species are again largely separatedgeographically, but this separation is due primar-
Polyscias sect. Tieghemopanax (Araliaceae)
43ADANSONIA, sér. 3 • 2001 • 23 (1)

Eibl J.M., Plunkett G.M. & Lowry II P.P.
44 ADANSONIA, sér. 3 • 2001 • 23 (1)
Fig.
6 (a
-c).
—S
tric
t co
nsen
sus
tree
s re
sulti
ng fr
om p
airw
ise
com
bin
atio
n of
the
thr
ee d
ata
sets
in w
hich
max
imum
ove
rlap
of t
axon
sam
plin
g w
as e
mp
loye
d; v
alue
s al
ong
bra
nche
sar
e b
oots
trap
per
cent
ages
.—(a
) Str
ict
cons
ensu
s of
6 m
ost
par
sim
onio
us (M
P) t
rees
res
ultin
g fr
om a
naly
sis
of 3
3 IT
S +
trn
L-tr
nF s
eque
nces
; tre
e le
ngth
= 5
07 s
tep
s; c
onsi
sten
cyin
dex
= 0
.647
; ret
entio
n in
dex
= 0
.731
.—(b
) Str
ict
cons
ensu
s of
3 M
P t
rees
res
ultin
g fr
om a
naly
sis
of 2
6 IT
S +
5S
sp
acer
seq
uenc
es; t
ree
leng
th =
323
ste
ps;
con
sist
ency
ind
ex =
0.68
2; r
eten
tion
ind
ex =
0.7
86.
—(c
) Str
ict
cons
ensu
s of
36
MP
tre
es r
esul
ting
from
ana
lysi
s of
24
trnL
-trn
F +
5S
sp
acer
seq
uenc
es;
tree
leng
th =
175
ste
ps;
con
sist
ency
ind
ex =
0.76
5; r
eten
tion
ind
ex =
0.8
55.

ily to edaphic constraints. Polyscias dioica is wide-spread across the island (except in the northeast-ern regions) but occurs almost exclusively onultramafic substrates at low to high elevation. Incontrast, P. cissodendron is restricted to non-ultra-mafic substrates in the northeastern and north-central regions of New Caledonia at elevationsranging from sea level to 750 m. As noted above,P. cissodendron is the only New Caledonian mem-ber of the Tieghemopanax group that is notendemic, with populations occurring in thesouthern Solomon Islands and Vanuatu to thenorth, and on Lord Howe Island to the south(LOWRY et al. 1986; LOWRY 1989).
Each of the species pairs described aboveexhibits one or more characteristics pointingeither to an allopatric (MAYR 1963) or adaptivedivergence (TEMPLETON 1981) model of specia-tion. In each pair, both molecular and morpho-logical features help to confirm a commonancestry. Several different forms of geographicisolation, however, are evident in these pairs. Forexample, P. “lecardii” and P. “regalis” are sepa-rated both geographically and edaphically.Polyscias dioica and P. cissodendron are geographi-cally sympatric (or at least parapatric) in north-central New Caledonia, but are separatedprimarily on the basis of edaphic conditions.Both P. “scopoliae” and P. “veillonii” have nearlyidentical geographic distributions (the southernregion of New Caledonia) and soil requirements,but occur at different elevations with little or nooverlap. Three other species pairs, P. “bracteata”/P. “ subincisa”, P. “crenata”/ P. “nothisii”, andP. balansae/P. “jaffrei”, are geographically disjunct.In general, therefore, a paradigm involving isola-tion and divergence appears to be the predomi-nant mode of speciation within sectionTieghemopanax, and this pattern may reflect amore generalized syndrome for many taxa in theNew Caledonian flora. In contrast, the strikingmorphological variability and widespread distrib-ution of P. dioica, in conjunction with the multi-ple banding patterns and length variationobserved in the 5S PCR amplicons, seem to sug-gest an alternative mode of speciation for thisproblematic taxon.
Possible evolutionary scenarios in Polysciasdioica
The multiple banding patterns evident in thePCR products of the 5S region from severalaccessions of P. dioica suggest that this morpho-logically variable species also exhibits consider-able heterogeneity in the length and copynumber of the 5S repeat unit. Simple assessmentof the sizes of 5S spacer PCR products from10 accessions of P. dioica and from other speciesillustrates the great length variability of thissequence across the species of sectionTieghemopanax (see Results). Among the samplesof P. dioica, both single and triple banded PCRproducts were found; a similar result (data notdiscussed here) was observed in amplifications ofthe trnL-trnF marker in P. dioica, where both sin-gle- and double-banded amplicons were seenamong different accessions. In both cases, multi-ple bands may have resulted from non-specificamplification of an independent sequence. In atleast one case (P. dioica, Lowry 4804), however,homology of each of the three bands has beenestablished with 5S coding-region sequencesfrom other angiosperm taxa. This suggests thatmultiple (possibly unlinked) copies of the 5Sregion may be present in some (but not all) indi-viduals of P. dioica.
This result may be explained by several alterna-tive hypotheses, including gene duplication, het-erogeneity among repeat units at the same locus,or hybridization among different genotypes ofP. dioica (or alternatively between P. dioica andother species). Given the information presentedherein (and also in EIBL 2000), we speculate thathybridization is the most likely explanation. First,two of the three size variants found among someaccessions of P. dioica are of similar size to thosefound in single banded accessions (of bothP. dioica and other species). It is relatively easy toenvision an additive pattern resulting from such ahybridization (although this does not explain thesmallest product of ~200 bp, which was found innone the single-banded accessions). Secondly,P. dioica exhibits levels of morphological variationand a geographic range far exceeding those ofother species in section Tieghemopanax, a resultconfirmed by recent morphometric analysis ofthe group (EIBL 2000; EIBL et al. subm.). Polyscias
Polyscias sect. Tieghemopanax (Araliaceae)
45ADANSONIA, sér. 3 • 2001 • 23 (1)

dioica may thus represent the remnants of a com-plex of closely related species that were once mor-phologically and perhaps geographically andecologically distinct, but which have relativelyrecently experienced a breakdown in reproductivebarriers leading to hybridization and the resultingpattern of morphological variability seen today. Athorough population-level sampling of geneticmarkers capable of discriminating among thesealternative explanations is required to test thisscenario. It is clear, however, that the generalmode of divergent speciation so evident amongother members of section Tieghemopanax is not atwork in P. dioica. Because of its geographic distri-bution and the variety of its diversification pat-terns, Tieghemopanax appears to represent amodel system for evaluating modes of speciationin New Caledonia. Future studies are currentlyunderway to take advantage of these opportuni-ties and to examine several potentially parallelcases in genera of other families.
Acknowledgments
The authors thank J. ARIDY, J. FLORENCE, J.-N. LABAT,J. MUNZINGER, and many botanical gardens andherbaria (cited in Table 1) for kindly providing leafand/or herbarium material; the staffs of IRD Nouméaand CSIRO (Atherton and Canberra) for assistance inNew Caledonia and Australia, respectively;W. EGGLESTON for laboratory assistance; N. VU forsupplying unpublished sequences; and J. WEN andD.G. FRODIN for helpful suggestions to improve thefinal manuscript. Support for field work was providedby grants from the National Geographic Society(5793-96 to GMP) and the National ScienceFoundation (DEB-9627072 to PPL as Co-PI); labora-tory work was supported by grants from the NationalScience Foundation (DEB-9981641 to GMP andPPL) and the Thomas F. JEFFRESS and Kate MILLERJEFFRESS Memorial Trust (J-506 to GMP).
REFERENCES
BERNARDI L. 1971. — Araliacearum Madagascariae etComores propositum. 2. Revisio et taxa novaPolysciadum. Candollea 26: 13-89.
BERNARDI L. 1979. — The New Caledonian genera ofAraliaceae and their relationships with those ofOceania and Indonesia: 315-325, in LARSEN K. &HOLM-NIELSEN L.B. (eds.), Tropical Botany.
Academic Press, London, New York & SanFrancisco.
BULL J.J., HUELSENBECK J.P., CUNNINGHAM C.W.,SWOFFORD D.L. & WADDELL P.J. 1993. —Partitioning and combining data in phylogenetic
analysis. Syst. Biol. 42: 384-397.CANNATELLA D.C., HILLIS D.M., CHIPPINDALE P.T.,
WEIGT L., RAND A.S. & RYAN M.J. 1998. —Phylogeny of frogs of the Physalaemus pustulosus
species group, with an examination of data incon-gruence. Syst. Biol. 47: 311-355.
DE QUEIROZ A., DONOGHUE M.J. & JUNHYONG K.1995. — Separate versus combined analysis of phy-logenetic evidence. Ann. Rev. Ecol. Syst. 26: 657-681.
DOWNIE S.R. & KATZ-DOWNIE D.S. 1996. — A mol-ecular phylogeny of Apiaceae subfamily Apioideae:evidence from nuclear ribosomal DNA internaltranscribed spacer sequences. Amer. J. Bot. 83: 234-251.
DOYLE J.J. & DOYLE J.L. 1987. — A rapid DNA iso-lation procedure for small quantities of fresh tissue.Phytochem. Bull. 19: 11-15.
EIBL J.M. 2000. — Molecular and morphologicalsystematics of Polyscias section Tieghemopanax(Araliaceae). Master’s thesis, Virginia CommonwealthUniversity, Richmond, Virginia.
FARRIS J.S., KÄLLERSJÖ M., KLUGE A.G. & BULT C.1995. —Testing significance of incongruence.Cladistics 10: 315-319.
FELSENSTEIN J. 1985. — Confidence limits on phylo-genies: an approach using the bootstrap. Evolution39: 783-791.
FORSTER J.R. & FORSTER G. 1776. — CharacteresGenerum Plantarum. London.
GUILLAUMIN A. — 1948. Flore analytique et synoptiquede la Nouvelle-Calédonie. Phanérogames. Office de laRecherche Scientifique Coloniale, Paris.
HARMS H. 1894-97. — Araliaceae: 1-62, in ENGLER A.and PRANTL K. (eds.), Die Natürlichen Pflan-zenfamilien III, Vol. 8. W. Engelmann, Leipzig.
HENEGARIU O., HEEREMA N.A., DLOUHY S.R.,VANCE G.H. & VOGT P.H. 1997. — MultiplePCR: critical parameters and step-by-step protocol.BioTechniques 23: 504-511.
HOLMGREN P.K., HOLMGREN N.H. & BARNETT L.C.1990. —Index herbariorum. New York BotanicalGarden, Bronx, New York.
HUTCHINSON J. 1967. — The genera of floweringplants. Vol. 2. Oxford University Press, London.
JAFFRÉ T. 1976. — Composition chimique et condi-tions d’alimentation minérale des plantes surroches ultrabasiques (Nouvelle-Calédonie). Cah.ORSTOM, Sér. Biol. 9: 53-63.
JAFFRÉ T., MORAT Ph., VEILLON J.-M. & MACKEEH.S. 1987. — Changements dans la végétation dela Nouvelle-Calédonie au cours du Tertiaire: lavégétation et la flore des roches ultrabasiques. Bull.Mus. Natl. Hist. Nat., B, Adansonia 9: 365-391.
Eibl J.M., Plunkett G.M. & Lowry II P.P.
46 ADANSONIA, sér. 3 • 2001 • 23 (1)

JAFFRÉ T., MORAT Ph. & VEILLON J.-M. 1993. —Etude floristique et phytogéographique de la forêt
sclérophylle de Nouvelle-Calédonie. Bull. Mus.Natl. Hist. Nat., B, Adansonia 15: 107-146.
JOHNSON L.A. & SOLTIS D.E. 1998. — Assessingcongruence: empirical examples from moleculardata: 297-348, in SOLTIS D.E., SOLTIS P.S. &DOYLE J.J. (eds.), Molecular Systematics of Plants II:DNA Sequencing. Klewer Publishers, Boston,Massachusetts.
KLUGE A.G. & WOLF A.J. 1993. — Cladistics: What’sin a Word? Cladistics 9: 183-199.
KRONKE L.W. 1996. — Plate tectonic development ofthe western and southwestern Pacific: mesozoic tothe present: 19-34 in KEATS A. & MILLER S.E.(eds.), The origin and evolution of Pacific islandbiotas, New Guinea to eastern Polynesia, patterns andprocesses. SPB Academic Publ., Amsterdam.
LOWRY II P.P. 1989. — A revision of Araliaceae fromVanuatu. Bull. Mus. Natl. Hist. Nat., B, Adansonia11: 117-155.
LOWRY II P.P. 1998. — Diversity, endemism, andextinction in the flora of New Caledonia: a review:181-206, in PENG C.-I & LOWRY II P.P. (eds.),Proceedings of the International Symposium on Rare,Threatened, and Endangered Floras of Asia and thePacific. Academia Sinica, Taipei.
LOWRY II P.P., DAVIES F.G. & GREEN P.S. 1986. —The distribution and synonymy of Polyscias cisso-
dendron (Araliaceae). Kew Bull. 41: 804.MABBERLEY D.J. 1997. — The Plant-Book: a portable
dictionary of the higher plants. Cambridge UniversityPress, New York.
MADDISON W.P. & MADDISON D.R. 1992. —MacClade: analysis of phylogeny and character evolu-
tion , version 3.01. Sinauer Associates Inc.,Sunderland, Massachusetts.
MAYR E. 1963. — Animal species and evolution.Harva rd Unive r s i t y P re s s , Cambr idge ,Massachusetts.
MASON-GAMER R.J. & KELLOGG E.A. 1996. —Testing for phylogenetic conflict among molecular
data sets in the tribe Triticeae (Gramineae). Syst.Biol. 45: 524-545.
MICKEVICH M.F. & FARRIS J. S. 1981. — The impli-cations of congruence in Menidia. Syst. Zool. 30:351-370.
MIQUEL F.A.G. 1855. — Plantae Junghuhnianae. Vol.4. A.W. Sythoff, Leiden.
MIQUEL F.A.G. 1856. — Araliacearum indicarumgenera et species aliquot novae. Bonplandia 4: 137-139.
MORAT Ph. 1993. — Our knowledge of the flora ofNew Caledonia: endemism and diversity in relationto vegetation type and substrates. Biodiv. Letters 1:72-81.
MORAT Ph., VEILLON J.-M & MACKEE H.S. 1984. —Floristic relationships of New Caledonian rain for-
est phanerogams: 71-128, in RADOVSKY F.J., RAVENP.H. & SOHMER S.H. (eds.), Biogeography of the
Tropical Pacific. Assoc. Syst. Collections, Lawrence,Kansas and Bernice P. Bishop Museum, Honolulu,Hawaii.
MORAT Ph., JAFFRÉ T., VEILLON J.-M. & MACKEEH.S. 1986. — Affinités floristiques et considéra-tions sur l’origine des maquis miniers de laNouvelle-Calédonie. Bull. Mus. Natl. Hist. Nat., B,Adansonia 8: 133-182.
MORAT Ph., JAFFRÉ T. & VEILLON J.-M. 2001. —The flora of New Caledonia’s calcareous substrates.Adansonia, sér. 3, 23: 109-127.
MUELLER-DOMBOIS D. & FOSBERG F.R. 1998. —Vegetation of the tropical Pacific islands. Sprenger-
Verlag, New York.OLMSTEAD R.G. & SWEERE J.A. 1994. — Combining
data in phylogenetic systematics: an empiricalapproach using three molecular data sets in theSolanaceae. Syst. Biol. 43: 467-481.
PHILIPSON W.R. 1978. — A synopsis of the Malesianspecies of Polyscias (Araliaceae). Blumea 24: 169-172.
PHILIPSON W.R. 1979. — Araliaceae: 1-105. FloraMalesiana, ser. 1, 9.
PLUNKETT G.M., LOWRY II P.P. & BURKE M.K.2001. — The phylogenetic status of Polyscias(Araliaceae) based on nuclear ITS sequence data.Ann. Missouri Bot. Gard. 88: 213-230.
SANGER F., NICKLEN S. & COULSON A.R. 1977. —DNA sequencing with chain-terminating
inhibitors. Proc. Natl. Acad. Sci., USA 74: 5463-5467.
SCHLESSMAN M.A., LLOYD D.G. & LOWRY II P.P.1990. — Evolution of sexual systems in NewCaledonian Araliaceae. Mem. New York Bot. Gard.55: 105-117.
SCHLESSMAN M.A., PLUNKETT G.M., LOWRY II P.P.& LLOYD D.G. 2001. — Sexual systems in NewCaledonian Araliaceae: a preliminary phylogeneticreappraisal. Edinburgh J. Bot. 58: 221-228.
SEELANAN T., SCHNABEL A. & WENDEL J.F. 1997. —Congruence and consensus in the cotton tribe(Malvaceae). Syst. Bot., 22: 259-290.
SMITH A.C. 1985. — Flora Vitiensis Nova. Vol. 3.Pacific Tropical Bot. Garden, Lawai, Hawaii.
SMITH A.C. & STONE B.C. 1968. — Studies of PacificIsland plants. XIX. The Araliaceae of the NewHebrides, Fiji, Samoa, and Tonga. J. Arnold Arbor.49: 431-493.
SOLTIS D.E. & SOLTIS P.S. 1998. — Choosing anapproach and an appropriate gene for phylogeneticanalysis: 1-42, in SOLTIS D.E., SOLTIS P.S. &DOYLE J.J. (eds.), Molecular Systematics of Plants II:DNA Sequencing. Kluwer Publishers, Boston,Massachusetts.
SOLTIS D.E., SOLTIS P.S., COLLIER T.G. &EDGERTON M.L. 1991. — The Heuchera group(Saxifragaceae): evidence for chloroplast transferand paraphyly. Amer. J. Bot. 78: 1091-1112.
SOLTIS D.E., SOLTIS P.S., MORT M.E., CHASE M.W.,SAVOLAINEN V., HOOT S.B. & MORTON C.M.
Polyscias sect. Tieghemopanax (Araliaceae)
47ADANSONIA, sér. 3 • 2001 • 23 (1)

1998. —Inferring complex phylogenies using parsi-mony: an empirical approach using three largeDNA data sets for angiosperms. Syst. Biol. 47: 32-42.
STONE B.C. 1965a. — Notes on the type species ofPolyscias J.R. & G. Forst. (Araliaceae). Taxon 14:281-285.
STONE B.C. 1965b. — Notes on Polyscias (Araliaceae)from Micronesia. Micronesica 2: 51-59.
TABERLET P., GIELLY L., PAUTOU G. & BOUVET J.1991. — Universal primers for amplification ofthree non-coding regions of chloroplast DNA.Plant Mol. Biol. 17: 1105-1109.
TAJIMA F. & NEI M. 1984. — Estimation of evolu-tionary distance between nucleotide sequences.Mol. Biol. Evol. 1: 269-285.
TEMPLETON A.R. 1981. — Mechanisms of speciation— a population genetic approach. Ann. Rev. Ecol.Syst. 12: 23-48.
UDOVICIC F., MCFADDEN G.I. & LADIGES P.Y.1995. — Phylogeny of Eucalyptus and Angophora
based on 5S rDNA spacer sequence data. Mol.Phylogen. Evol. 4: 247-256.
VIGUIER R. (1910-13) 1925. — Contribution àl’étude de la flore de la Nouvelle-Calédonie:Araliacées. J. Bot. (Morot) II, 3: 38-101 (dated1910-13 but not distributed until 1925; cf.GREEN M.L., Bull. Misc. Inform. 1928: 155-156,1928).
VIGUIER R. 1905. — Sur les Araliacées du groupe desPolyscias. Bull. Soc. Bot. France 52: 285-315.
WEN J. & ZIMMER E.A. 1996. — Phylogeny and bio-geography of Panax L. (the ginseng genus,Araliaceae): inferences from ITS sequences ofnuclear ribosomal DNA. Mol. Phylogen. Evol. 6:167-177.
WHITE T.J., BRUNS T., LEE S. & TAYLOR J. 1990. —Amplification and direct sequencing of fungal ribo-somal RNA genes for phylogenetics: 315-322, inINNIS M.A. (ed.), PCR Protocols: A guide to methodsand applications. Academic Press, San Diego,California.
Manuscript received 28 February 2001;revised version accepted 30 April 2001.
Eibl J.M., Plunkett G.M. & Lowry II P.P.
48 ADANSONIA, sér. 3 • 2001 • 23 (1)
Top Related