







Languages
Pages
Legal


Chapter 31
Post-transcriptional Processing
(pages 1057-1066)
Learning objectives: Understand the following
• What are capping, polyadenylation, splicing?• What is the functional importance of each process?• Why does each process work on transcripts produced by RNAP II?

Post-transcriptional Processing of mRNA in Eukaryotes
• Translation closely follows transcription in prokaryotes
• In eukaryotes, these processes are separated - transcription in nucleus, translation in cytoplasm
• On the way from nucleus to cytoplasm, the mRNA is converted from a "primary transcript" to "mature mRNA"

Post-transcriptional Modifications of mRNA in Eukaryotes
1) 5’ end = capping
2) 3’ end = polyadenylation
3) internal = splicing
Order
of
events
pre-mRNA
(heteronuclear RNA)
Mature mRNA
(gets translated)
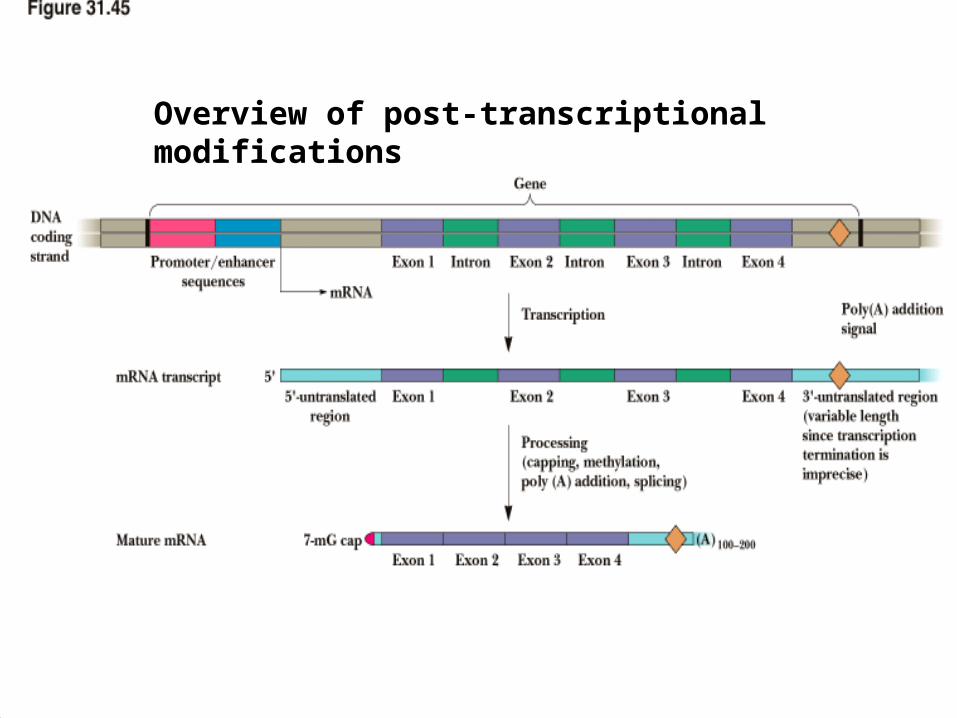
Overview of post-transcriptional modifications

Capping and Methylation
• Primary transcripts (pre-mRNAs or heterogeneous nuclear RNA) are usually first "capped" by a guanylyl group
• This is a modification of the 5’ triphosphate of the mRNA
• The reaction is catalyzed by the enzymes phosphohydrolase, guanylyl transferase and methyl transferases

Capping and Methylation
• Starting with the 5’ triphosphate of the mRNA the gamma phosphate group is hydrolyzed off.
O-P-O-P-O-P-OCH2
5’
O
O O O
O O O
- - -
OHO
base
RNA chain
O-P-O-P-OCH2
5’
O
O O
O O- -
OHO
base
RNA chainPi
phosphohydrolase
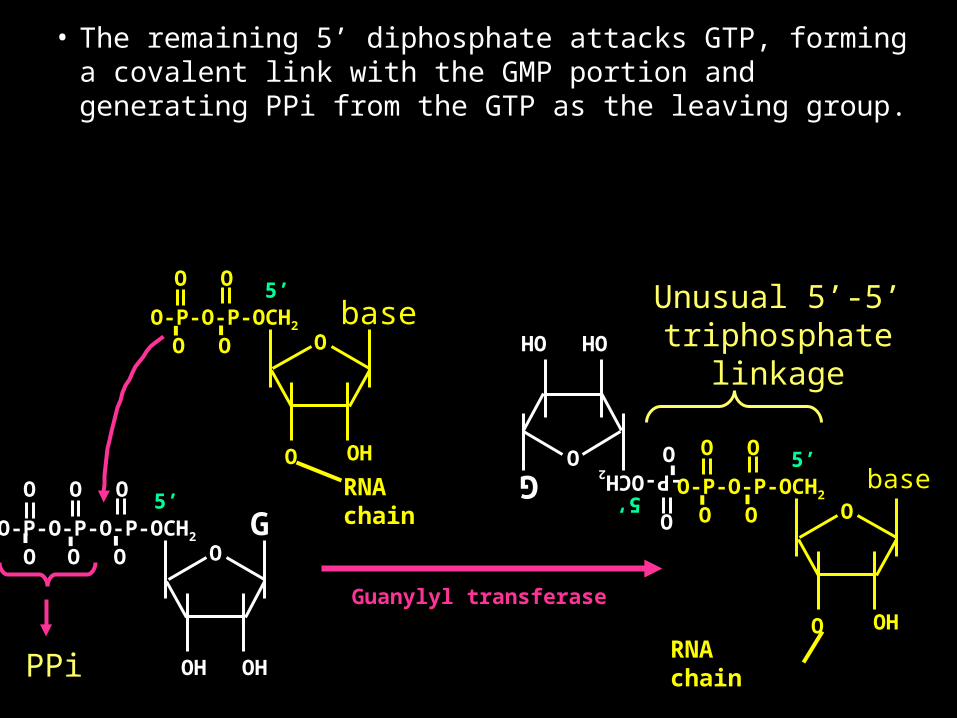
• The remaining 5’ diphosphate attacks GTP, forming a covalent link with the GMP portion and generating PPi from the GTP as the leaving group.
O-P-O-P-OCH2
5’
O
O O
O O
- -
OHO
base
RNA chain
Guanylyl transferase
PPi
O-P-O-P-O-P-OCH2
O
O O O
O O O
- - -
OHOH
G5’
RNA chain
O-P-O-P-OCH2
5’
O
O O
O O
- -
OHO
base-P-OCH2
O
O
O
-
OH OH
G5’
Unusual 5’-5’ triphosphate linkage

Hydrolyzed
Attack
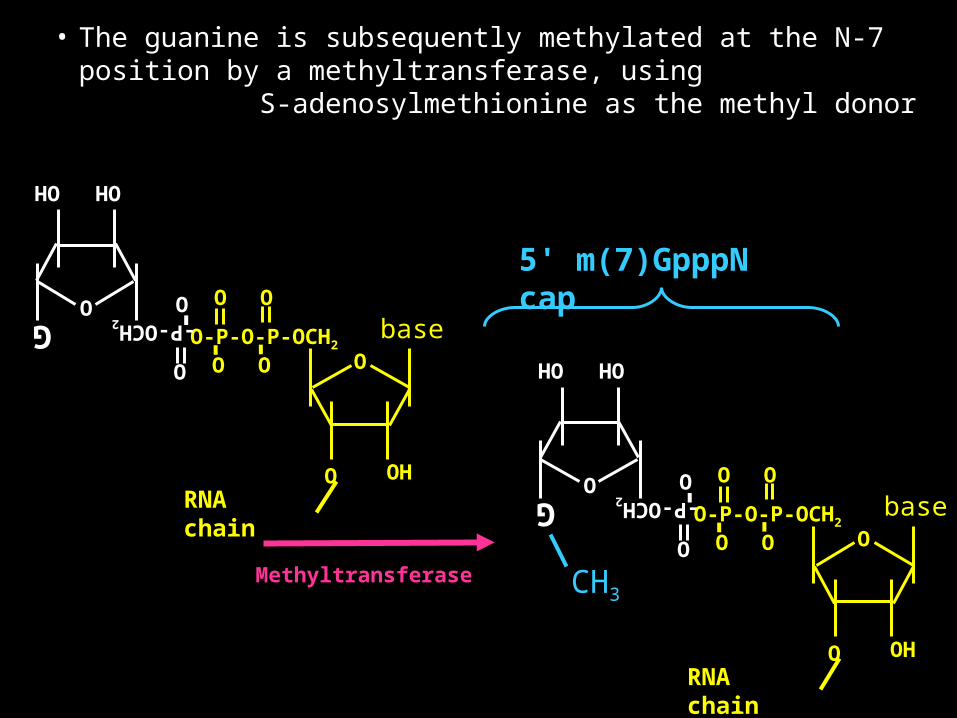
• The guanine is subsequently methylated at the N-7 position by a methyltransferase, using S-adenosylmethionine as the methyl donor
Methyltransferase
RNA chain
O-P-O-P-OCH2
O
O O
O O
- -
OHO
base-P-OCH2
O
O
O
-
OH OH
G
RNA chain
O-P-O-P-OCH2
O
O O
O O
- -
OHO
base-P-OCH2
O
O
O
-
OH OH
G
CH3
5' m(7)GpppN cap

• All eukaryotic mRNAs have 5’-N7 methylguanosine CAP structure.
• This is referred to as a “CAP 0” structure
• Additional methyl groups are added in the majority of mRNAs:
• CAP1 structure has a second methylation (not found in yeast mRNAs)
• CAP2 structure has a third methylation (found mainly in vertebrate mRNAs)
Capping and Methylation

CAP 0
CAP 1
CAP 2
Additionalmethylations are possible

Capping and Methylation summary
• Primary transcripts (pre-mRNAs or heterogeneous nuclear RNA) are "capped" by a guanylyl group
• The reaction is catalyzed by three different enzymes
• The capping G residue is methylated at 7-position
• Additional methylations occur at 2'-O positions of next two residues and at 6-amino of the first adenine

Why have a Cap?
• The CAP structure promotes stability of the mRNA (prevents degradation by 5’ exonucleases)
• The CAP structure promotes translation of the mRNA
• A ribosomal protein subunit interacts with the CAP and recruits mRNA to the ribosome

Why are only mRNAs Capped?
• The CAP structure is only added to transcripts synthesized by RNAP II (i.e. mRNA transcripts)
• This is because the capping enzymes bind to a part of RNAP II that is unique to that enzyme (not found in RNAP I or RNAP III)
• Capping enzymes bind to the CTD of RNAP II

Review of CTD
RPB1
RPC1
’
RPB1 = largest subunit of RNAP IIRPC1 = largest subunit of RNAP III
•Diagram shows an alignment of amino acids from the largest subunits of RNAP II and III from yeast, and from E. coli.
•The boxes are domains of high amino acid similarity
•RNAPII largest subunit has a C-terminal extension - the CTD!!

The sequence of the CTD is an unusual 7 amino-acid repeat with an “extended” structure
26
52
• deletion studies showed that yeast require at least 13 repeats to survive
• therefore the CTD is essential for RNAP II function
• the CTD has a lot of S, T amino acids which can be phosophorylated
• it is known that RNAP II in an initiation complex has a non-phosphorylated CTD
• elongating RNAP II has a phosphorylated CTD

Why are only mRNAs Capped?
• Capping enzymes bind to the CTD of RNAP II
• They bind only to the elongating form of the CTD( I.e. the phosphorylated CTD)
• As soon as the transcript emerges from RNAP II it is in contact with the capping enyzmes

3'-Polyadenylation
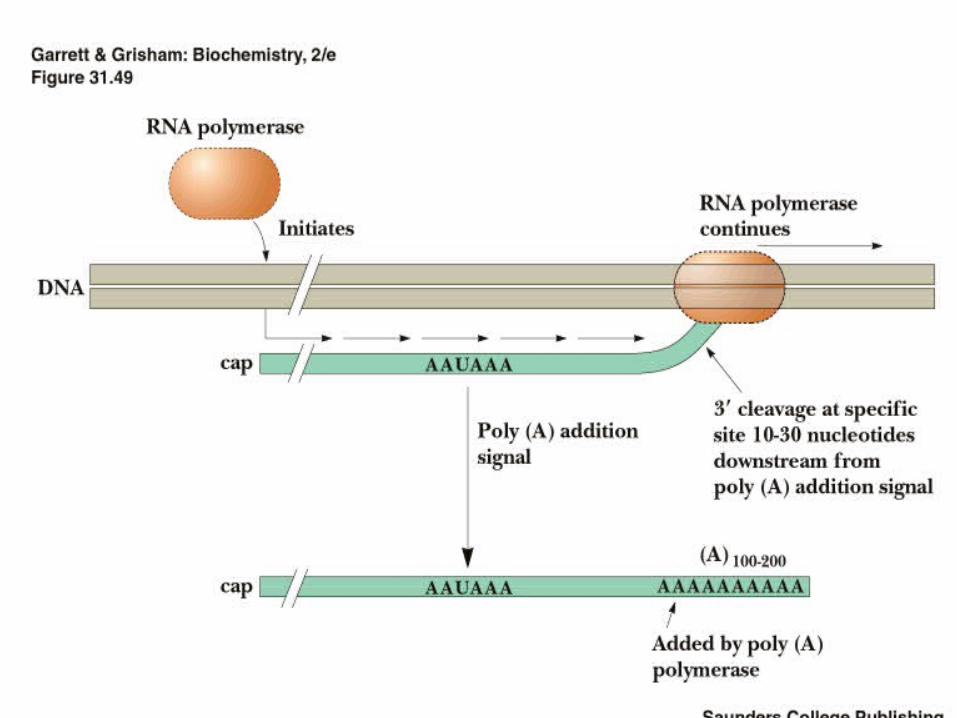
A poly(A) tail is added to the 3’ end of the transcript (defines the 3’ end) in 2 steps…
1) cleavage: the RNA is cut 10-30 nucleotides downstream of a specific sequence in the 3’UTR
2) addition of A’s (100-200 are added) to generate a poly(A) tail
Coding sequence of the geneTATA box +1
25-30 bpmRNA
Translated region5’UTR 3’UTR
ATG Stop

mRNA
5’ CAP
AAUAAA
CPSF
1) CPSF = “cleavage/polyadenylation specificity factor” (3 subunits) recognizes the AAUAAA sequence in the RNA 3’UTR and binds to it
2) Recruits CFI and CFII = “cleavage factors”
3) Recruits PAP = “poly(A) polymerase”
CFICFII PAP
A ribonucleoprotein complex

mRNA
5’ CAP
AAUAAA
CPSF
• The RNA is cleaved 10-30 nucleotides downstream of the AAUAAA by CFI/II• This generates a 3’OH to which PAP adds A residues. It doesn’t copy from a template - just needs a 3’OH of an RNA• The poly(A) tail is bound by poly(A)-binding protein
CFICFII PAP
AAUAAAAAAAAAAAAAAAA100-200

Why have a Poly(A) tail?• The poly(A) tail promotes stability of the mRNA (prevents
degradation by 3’ exonucleases)
• The poly(A) tail promotes translation of the mRNA
• A ribosomal protein subunit that interacts with the CAP structure is stimulated in CAP binding by poly(A) binding protein
• Therefore the CAP and the poly(A) tail work synergistically to recruit mRNA to the ribosome

Why are only mRNAs polyadenylated?
• The poly(A) tail is only added to transcripts synthesized by RNAP II (I.e. mRNA transcripts)
• This is because the essential polyadenylation factor CPSF binds to the CTD of RNAP II
• Therefore, only transcripts made by RNAP II are in close proximity to CPSF which starts off the polyadenylation process

3'-PolyadenylationSummary
• Termination of transcription occurs only after RNA polymerase has transcribed past a consensus AAUAAA sequence - the poly(A)+ addition site
• 10-30 nucleotides past this site, a string of 100 to 200 adenine residues are added to the mRNA transcript - the poly(A)+ tail
• poly(A) polymerase adds these A residues
• Function still not completely worked out, but poly(A) tail increases stability of the mRNA
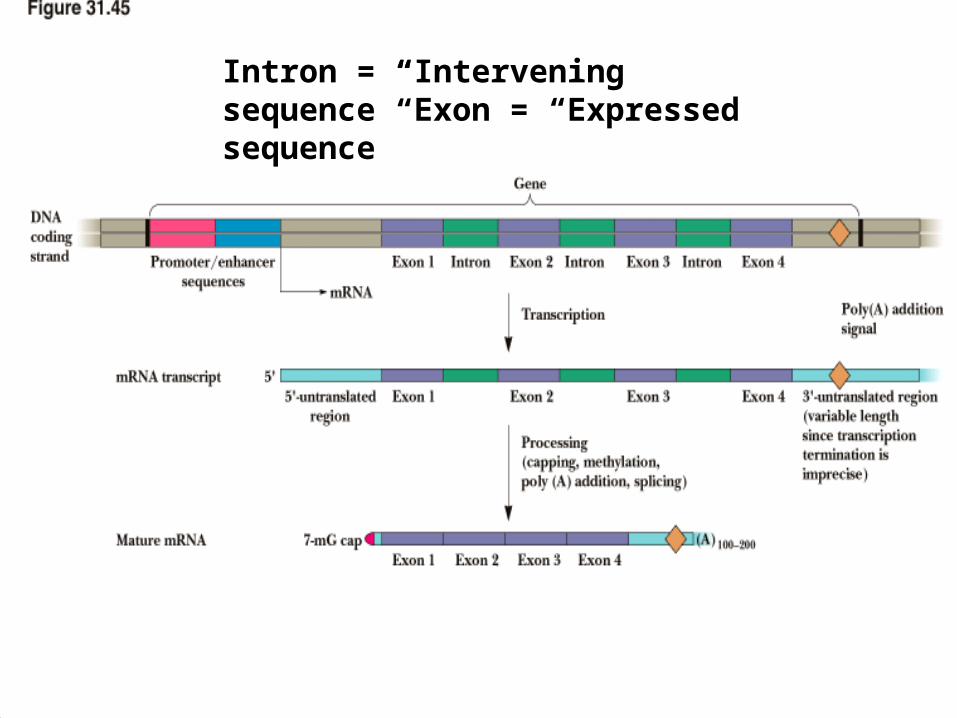
Intron = “Intervening sequence” Exon = “Expressed sequence”

Eukaryotic Genes are Split
• Introns intervene between exons
• Examples: actin gene has 309-bp intron separates first three amino acids and the other 350 or so
• But chicken pro-alpha-2 collagen gene is 40-kbp long, with 51 exons of only 5 kbp total.
• The exons range in size from 45 to 249 bases
• Mechanism by which introns are excised and exons are spliced together is complex and must be precise

Organization of the mammalian DHFR gene

Splicing of Pre-mRNA
Capped, polyadenylated RNA, in the form of a RNP complex, is the substrate for splicing
• In "splicing", the introns are excised and the exons are sewn together to form mature mRNA
• Splicing occurs only in the nucleus • The 5'-end of an intron in higher eukaryotes is
always GU and the 3'-end is always AG • All introns have a "branch site" 18 to 40
nucleotides upstream from 3'-splice site
• Branch site is essential to splicing
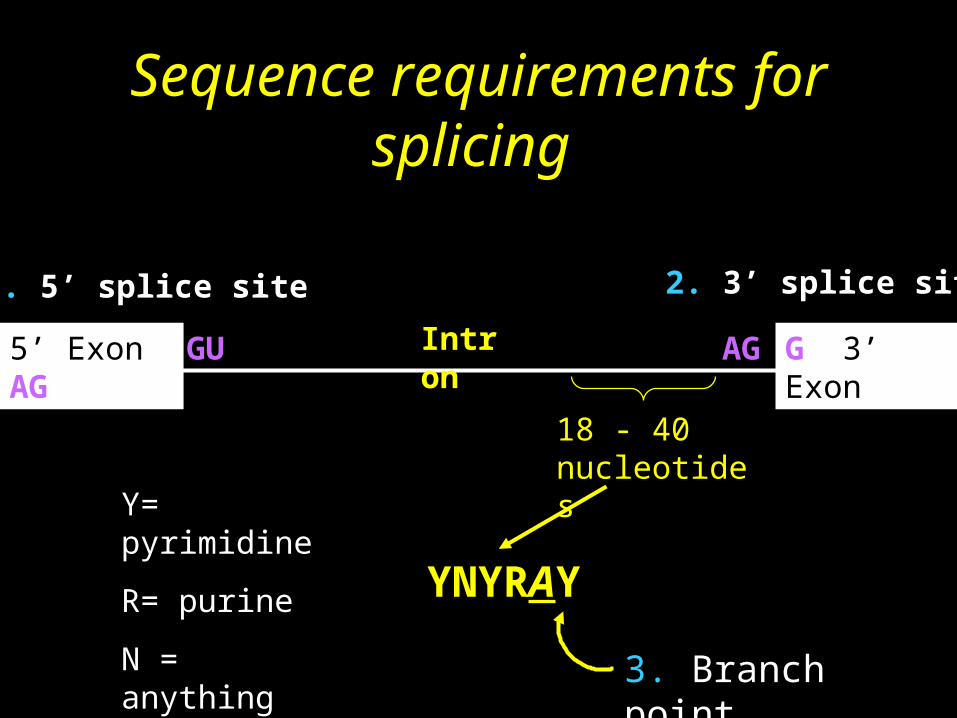
Sequence requirements for splicing
G 3’ Exon5’ Exon AG GU AGIntron
18 - 40 nucleotides
3. Branch point
YNYRAAY
Y= pyrimidine
R= purine
N = anything
1. 5’ splice site 2. 3’ splice site

Splicing occurs through two Transesterification reactions
P
O
-O
O -
O
-
X
YRN
A C
hai
n
+ R - OH P
O
-O
O -
O
-
R
Y
X - OH +
• In a transesterification reaction a phosphodiester bond is transferred to a different hydroxyl group
• There is no hydrolysis and no energy loss
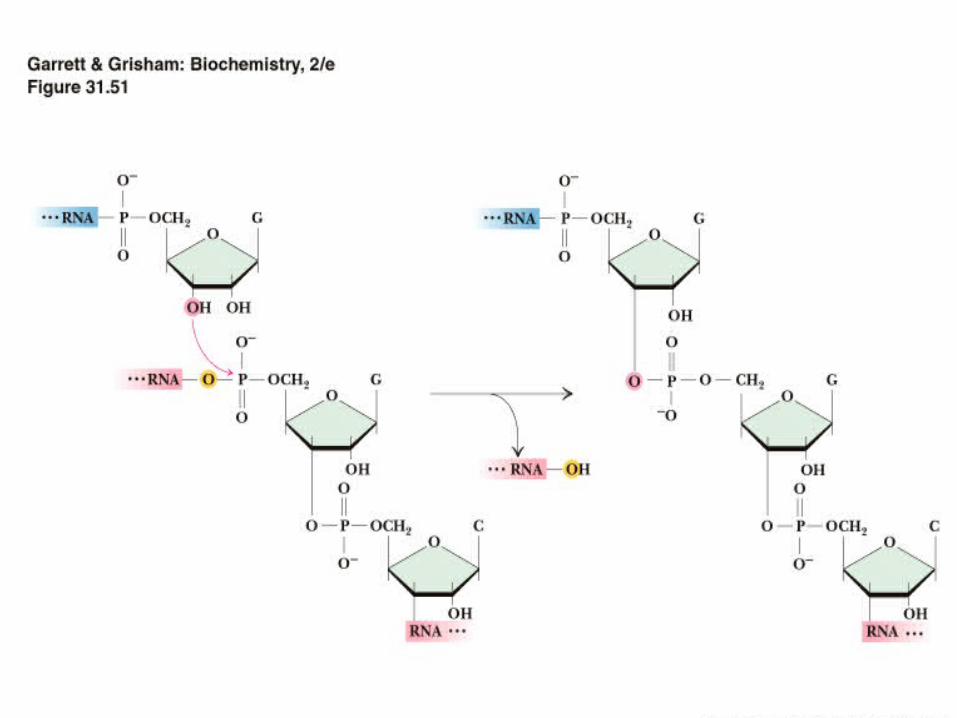


The Branch site and Lariat Summary
• Branch site is usually YNYRAY, where Y = pyrimidine, R = purine and N is anything
• The "lariat" a covalently closed loop of RNA is formed by attachment of the 5'-P of the intron's invariant 5'-G to the 2'-OH at the branch A site
• The exons then join, excising the lariat.
• The lariat is unstable; the 2'-5' phosphodiester is quickly cleaved and intron is degraded in the nucleus.

The Importance of snRNP
• Small nuclear ribonucleoprotein particles - snRNPs, pronounced "snurps" - are involved in splicing
• A snRNP consists of a small RNA (100-200 bases long) and about 10 different proteins
• snRNPs and pre-mRNA form the spliceosome
• Spliceosome is the size of ribosomes, and its assembly requires ATP

Spliceosome
5 snRNPs (“snurps”)
snRNA ~ 10 different proteins
•Small (100-200 ntds)
•Rich in U’s
•U1, U2, U4, U5, U6 snRNAs
U1, U2, U4, U5, U6 snRNPs

snRNAs
• snRNAs are very important in splicing
• mediate the sequence specificity and the accuracy of the splicing reaction
• fold into secondary structures (stem-loops) that are important for catalysis
•In snRNPs it is the RNA part that is catalytic, not the protein part
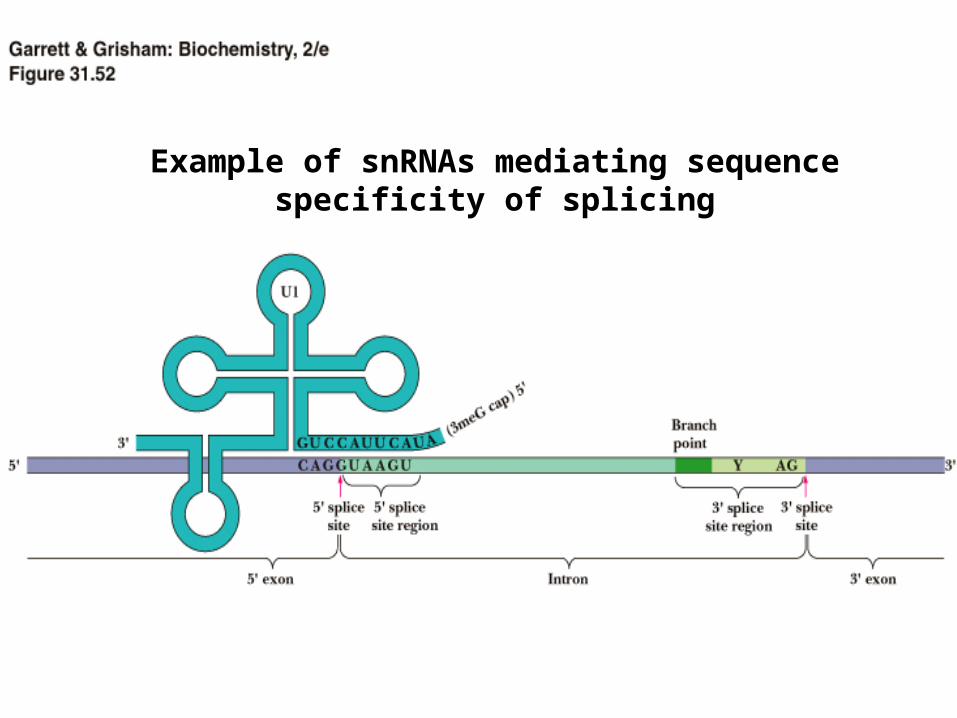
Example of snRNAs mediating sequence specificity of splicing

Assembly of the Spliceosome
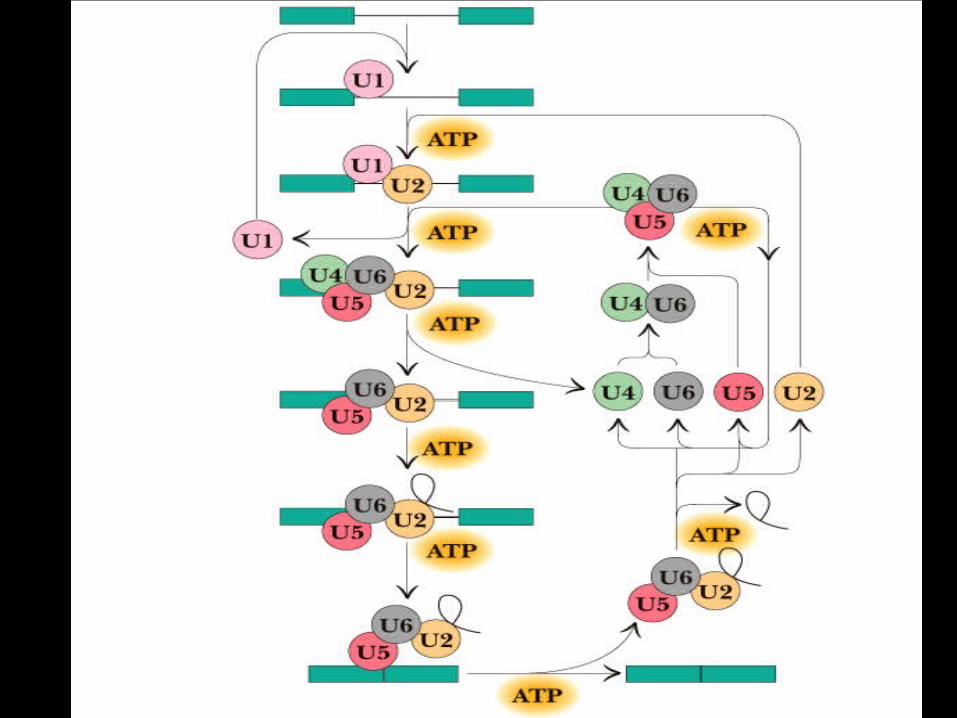
1) U1 snRNP binds to the 5’ splice junction
• This helps to recruit
2) U2 snRNP binds to the intron branch point
• This helps to recruit:
3) A trimeric snRNP composed of U4/U6-U5
• U4 and U6 RNAs are base-paired together to keep U6 RNA in the correct format for binding
• U6 RNA binds to the 5’ splice site and replaces U1

Assembly of the Spliceosome (continued)
4) U6 RNA now peels off the U4 RNA (it leaves the spliceosome)
5) This allows U6 RNA to pair with U2 RNA
• This brings the 5’ splice junction right next to the branch point
6) Catalysis!! The first transesterification occurs
• This generates the intron lariat + free 3’OH of the 5’ exon
• The free 5’exon doesn’t diffuse away since it is held in place by interactions with U5 snRNP

Assembly of the Spliceosome (continued)
7) The second transesterification occurs
• This generates a free lariat structure which is rapidly degraded, plus the spliced exons.
8) Now the spliceosome components all disassemble. And the U6 snRNP reassociates with a U4sNRNP for delivery to the next spliceosome.
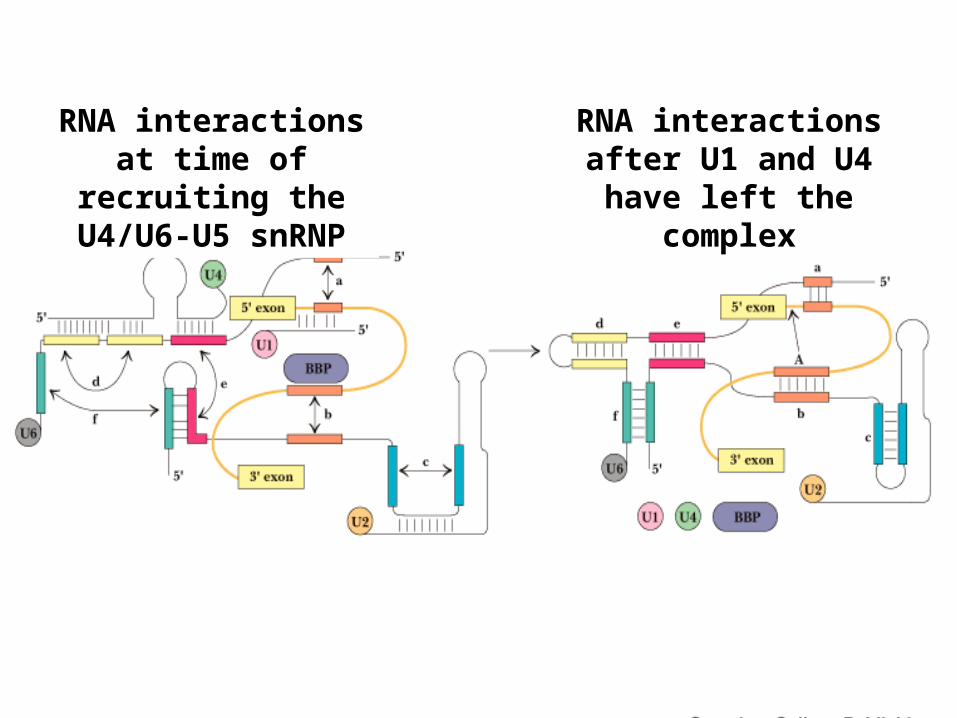
RNA interactions at time of recruiting
the U4/U6-U5 snRNP
RNA interactions after U1 and U4 have
left the complex
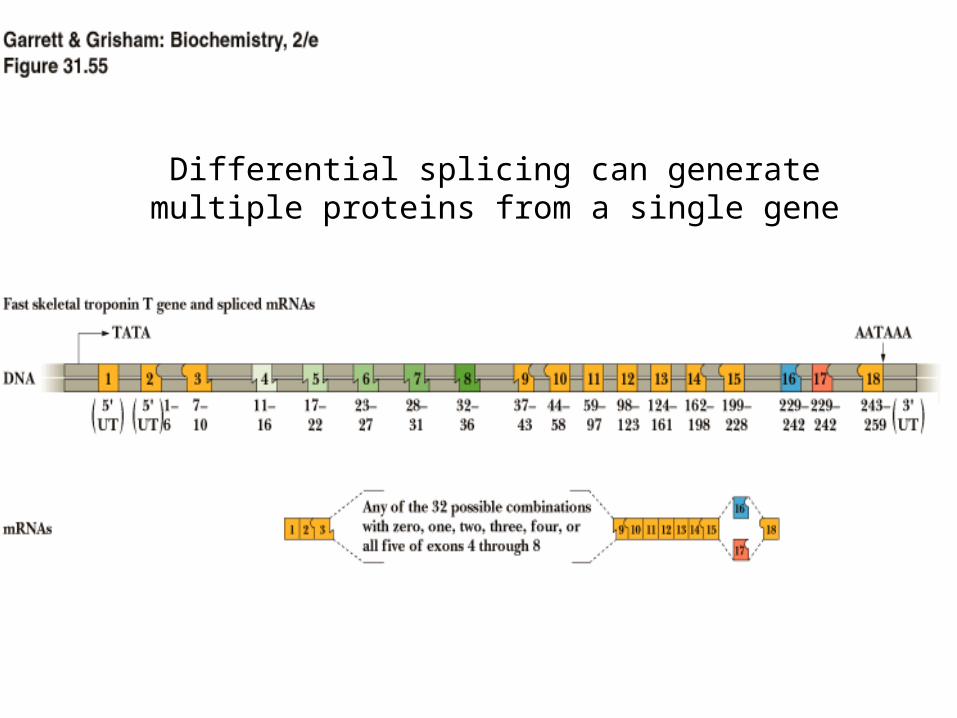
Differential splicing can generate multiple proteins from a single gene

Why are only mRNAs spliced by snRNPs?
• The CTD of RNAP II is required for splicing
• Therefore it is proposed that splicing factors from the spliceosome interact with the CTD
• This ensures that transcripts emerging from RNAPII are in the correct location for interaction with the splicing machinery

We have now finished Chapter 31!!
For next class please read:
Chapter 32 sections 1 and 2Pages 1069-1074
Elucidation of the genetic code
Top Related