







Languages
Pages
Legal


1. Bahan Baku (prekusor)
No Nutrien Darah SUSU1. Air 91 86
2. Glukosa 0.05 trace
Lactosa 0 4.6
3. Asam amino 0.02 trace
Casein 0 2.8
- Lactoglobulin 0 0.32
- Lactalbumin 0 0.13
Immunoglobulin 2.6 0.07
Serum albumin 3.2 0.05
4. Triasilgliserol 0.06 3.7
Fosfolipida 0.25 0.035
Asam sitrat trace 0.18
5. Ca 0.01 0.13
P 0.01 0.10
K 0.025 0.15
Na 0.34 0.05
Cl 0.35 0.11
Kandungan Nutrien Dalam Darah dan Susu Sapi
BIOSINTESA AIR SUSU

Bahan Baku menurut Komponen Susu
1. Glukosa :- Bahan baku laktosa susu
- Sumber -gliserol
- Dibutuhkan dalam perubahan NADP -> NADPH
- Sumber ATP
2. Asam amino :- Bahan baku casein susu
- Sumber glukosa
3. As. lemak (asetat, -hidroksi butirat, NEFA) :- Bahan baku asam lemak susu
4. Vitamin dan Mineral :- Diabsorbsi langsung dari darah

2. Biosintesis Lemak Susu
• Terjadi di sitoplasma sel sekretori kelenjar ambing.
• Kadarnya dalam susu berfluktuasi (2,5% – 6%); sangat
dipengaruhi oleh jenis pakan, 2,5 – 6%.
• 75 – 90% lemak susu berupa trigliserida yg mengandung asam
lemak rantai pendek (C4-C14) dan rantai panjang (C > 14)
• C4 – C14 disintesis dari asetat dan β-hidroksi butirat
• C > 14 disentesis dari trigliserida plasma (NEFA) dan palmitat.

Sources of Fatty AcidsThe fatty acids used to synthesize the milk triglycerides may arisefrom two sources:
1) From breakdown of blood lipids From 40 to 60% of the milk fattyacids come from the blood. These are primarily derived from very low densitylipoproteins (VLDL), which are synthesized in the intestine or liver. VLDL arecomposed of 90 to 95% lipid (55-60% triglyceride) on the inner core and 5 to10% protein at the outer urface.
2) From de novo synthesis within the mammary epithelialcells. Synthesis of short and medium chain fatty acids in the mammary gland occurs byde novo synthesis (synthesis from the start, or synthesis of new molecules of fatty acidsfrom precursors absorbed from the blood) wich occurs in the cytoplasm of the mammaryepithelial cell.
In the ruminant, the carbon sources used for FA synthesis are acetate (the mostimportant one) and b-hydroxybutyrate (BHBA). Glucose is a carbon source for FAsynthesis in nonruminants. The reducing equivalents needed for FA synthesis come fromNADPH2 (nicotinamide adenine dinucleotide phosphate, reduced form).

Liver
Mammary Gland
Rumen Intestine
NDF & NFC
AcetateButyrate
Saturated FA+ MG
“Glucose”
NEFA
Glycerol
Milk Fats
Fatty acids
AcetateBOHB
Fatty Acids
Origin of Milk Fat
AcetateBOHB
AcetateBOHBl
BOHB = B-hydroxybutyrate
VLDL - very low density lipo-proteins
Adiposetissues
TG
NEFA = Non-Esterified Fatty acids
VLDL
Ketones
TG
MG = Monoglycerides
TG = Triglycerides
Fats Saturation + Hydrolysis
FA = Fatty acids
VLDL

The two key enzymes involved in FA synthesis in the MG are:
1. Acetyl-CoA Carboxylase, which is the rate limiting enzyme for the fatty acid synthesis pathway.
2. Fatty Acid Synthetase, which is a large complex of enzyme activities responsible for the chain elongation of the fatty acid chains.
The relative proportions of milk fatty acids derived from de novo synthesis orblood lipids is dependent on carbon chain length of the fatty acid. Typically,shorter chain fatty acids arise predominantly from de novo synthesis intheepithelial cell, while longer chain fatty acids arise directly from blood lipids. Anexample is given in the table below for the cow.
Proportional contribution of sources of fatty acids in cow milk
Fatty Acid% of FA from
De novo synthesis
% of FA from
VLDL Fatty Acid
C4 -C10 100 0
C12 80-90 10-20
C14 30-40 60-70
C16 20-30 70-80
C18 0 100

In ruminants, fermentation of dietary carbohydrate in the rumen results in formation of volatile fatty acids (VFA's):
especially, acetate, propionate, and butyrate.

Komposisi asam lemak dalam trigliserida air susu
Asam lemak Panjang C Manusia Babi Kambing Sapi
Jenuh :
Butirat
Kaproat
Kaprilat
Kaprat
Laurat
Miristat
Palmitat
Stearat
4
6
8
10
12
14
16
18
-
-
-
2
8
9
23
9
2
2
2
2
2
2
29
6
7
5
4
13
7
12
24
51
10
3
1
2
3
9
21
11
Tak jenuh :
Oleat
Linoleat
lainnya
18: 1
18: 2
-
34
7
8
35
14
12
17
3
3
31
5
4

Fatty Acid Synthesis
Summary of Fatty Acid Synthesis Reactions:
Each cycle through the malonyl-CoA pathway results in two carbons being added to the FA chain.
The total reaction is (given here for synthesis of palmitate; C16):
Acetyl-CoA + 7 Malonyl-CoA + 14 NADPH2
are catalyzed by Fatty Acid Synthetase to yield :
Palmitate + 7 CO2 + 14 NADP + 8 CoA

The FA Synthesis Pathway involves the following steps :
Activation - acetyl-CoA carboxylation
Elongation - the malonyl-CoA pathway
Condensation step
Reduction step
Dehydration step
another reduction step
The cycle is then repeated
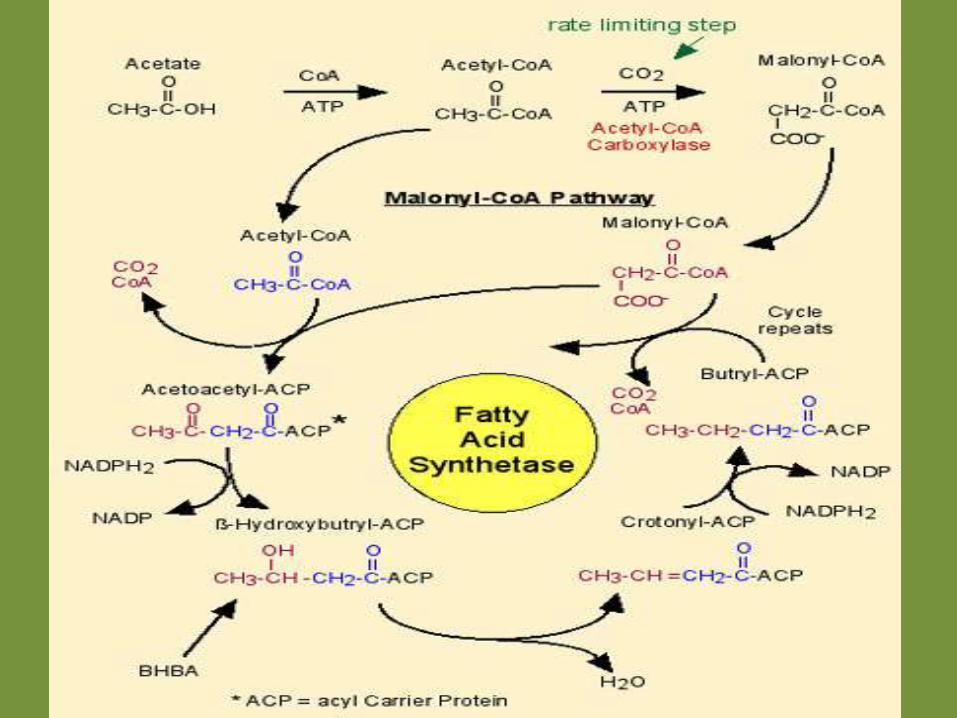


Summary of FATTY ACID SYNTHESIS• it occurs in the cytoplasm
• the intermediates are linked to ACP (acyl carrier protein)
• the enzymes of FA synthesis are linked in a complex
• elongation occurs by 2 Carbons/cycle (source of 2-Carbon units is always acetyl-CoA at the beginning of each cycle)
• the required reducing agent is NADPH2
• in general, elongation stops at C16
These two things are required for de novo FA synthesis: • a source of carbons, specifically acetyl-CoA • a source of reducing equivalents, specifically NADPH2
The origin of each of these varies among species, particularly in comparing ruminants and nonruminants.

Triglyceride Synthesis
Synthesis of triglycerides from FAs (preformed from the blood orsynthesized in the cell by de novo synthesis) occurs at thecytoplasmic surface of the smooth endoplasmic reticulum (SER).
The FAs are esterified to the hydroxyl groups of the glycerolmolecules. This occurs by a series of esterases.
There is some specificity of which carbon on the glycerol is used toesterify particular chain length FAs.
In the cow:
* C16 is found predominantly on the #2 carbon of glycerol,* C18 FAs mostly on the #3 carbon,* C4-C8 mostly on the #3 carbon, and* C10-C14 distributed among each of the glycerol carbons.

Lipid Droplet Formation
Inside the mammary epithelial cell, as the triglycerides are synthesized at the outersurface of the SER (smooth endoplasmic reticulum), they start coalescing and formmicro lipid droplets.
These micro lipid droplets bud or bleb off from the SER surface into the cytoplasm.
The micro lipid droplets may be secreted from the cells directly as very smallmilk fat globules (less than 0.5 mm), they may fuse with each other to form largerdroplets (cytoplasmic lipid droplets), and they may fuse with cytoplasmicdroplets, ultimately resulting in formation of larger milk lipid droplets.
Milk fat globules (in milk) range from less than 0.5 to greater than15 mm. Themicro and cytoplasmic lipid droplets are not surrounded by a lipid bilayermembrane but apparently are surrounded by a nonbilayer coating made ofprotein and polar lipid (gangliosides).
This surface coating prevents coalescence of the droplets with lipids in the cell,yet permits fusion between droplets. In fact, the protein and ganglioside of thiscoating, along with calcium, are involved in the fusion of the droplets.

Secretion of Milk Fat
As the large milk lipid droplets migrate to the apical surface of thecell they continue to push outward and are enveloped by the apicalmembrane of the cell.
This membrane eventually fuses at the base, releasing themembrane-bound milk fat globule, and closing the cell's apicalmembrane so ere is no hole in it. So, the milk fat globule ismembrane-surrounded and has a number of membrane-associatedproteins.
These proteins and others trapped during the process ofseparating cream from whole milk (usually by some type ofcentrifugation) are important for the whipping properties of cream.

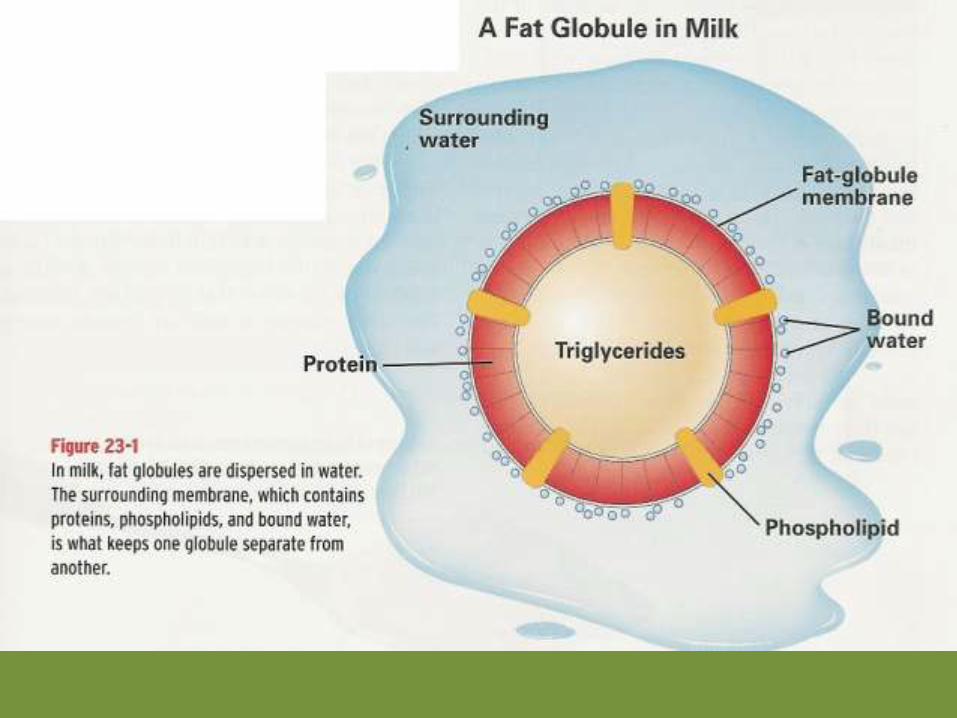
Overview of Milk Fat Synthesis
The components of milk fat synthesis are summarized in the diagrambelow.

This fat globule membrane helps to stabilize the fat globules in anemulsion within the aqueous environment of milk (remember thatcow milk is about 87% water). Lipid has a lower density thanwater, so when raw milk is centrifuged the fat rises to the top
resulting in the cream layer (see image).
Milk fat is the primary component of cream.Below milk was centrifuged to separate thecream and the skim milk (also called the plasmaphase of milk).
Homogenization is the process by which fatglobules in fluid milk are broken into sizes smallenough that they will not rise in the milk toform cream under normal milk storageconditions. This is important for processing,storage and consumption of cow milk.

3. Biosintesis Laktosa Susu
• Berlangsung di aparatus golgi sel sekretori kelenjar ambing.
• Bahan baku sintesis laktosa adalah glukosa
• Kadarnya dalam susu dipertahankan tetap yaitu 3,6 – 5,5%
• 1 molekul laktosa tersusun dari 1 mol glukosa dan 1 mol
galaktosa yang juga berasal dari glukosa. Jadi untuk sintesis 1
mol laktosa butuh 2 mol glukosa
• 80% glukosa plasma digunakan untuk sintesis laktosa, 50-60%
diubah menjadi galaktosa dahulu.

Lactose is a disaccharide composed of D-galactose and D-glucose.

• Laktosa bertanggung jawab mempertahankan keseimbangan
tekanan osmotik antara darah dan lumen susu.
• Semakin tinggi produksi laktosa semakin banyak air yg
dialirkan ke lumen susu produksi susu meningkat.
• Ketersediaan glukosa dalam sel sekretoris kelenjar ambing
sangat penting dalam peningkatan produksi susu.
• Glukosa merupakan faktor pembatas pada sapi perah
berproduksi tinggi.

Lactose Biosynthesis O OH
O
OH OH
CH2
OH
OH
OH
O
OH
CH2
OH
Galactose Glucose
Liver Secretory Cell
Glucose Glucose
Blood
CH3 CH2 CO
O -
Propionate
Glucogenic amino-acids(glutamte, aspartateserine, etc.)
Glucose-P
Galactose Galactose
Glucose
A
B
A = galactosyl transferase; B = alpha lactalbumin
LACTOSE
Golgi Apparatus
Water

Ringkasan reaksi biosentesa laktosa
ATP + glukosa Glukosa- 6P + ADPHeksokinase
Glukosa-6P Glukosa-1PPhospoglucomutase
UTP + Glukosa-1P UDP-Glukosa + PP
UDP-GlucosePyriphosporylase
UDP-Glukosa UDP-GalaktosaUDP-Galactose4-epimerase
UDP-Galaktosa+Glukosa Laktosa + UDP
Lactose synthetase

1. Pemilihan bahan pakan yang tepatdipilih bahan pakan yang banyak memberikan pasokan glukosa pada sel kelenjar ambing.
a. Bahan pakan yang menghasilkan propionat dalam rumen
b. Bahan pakan kaya pati namun lolos degradasi rumen,sehingga menghasilkan glukosa dalam usus.
Upaya meningkatkan produksi susu melalui peningkatan sintesis laktosa

2. Manipulasi Metabolisme Nutrien
• Meningkatkan uptake glukosa oleh sel kelanjer ambing(manipulasi sistem trasport glukosa)
• Pengambilan glukosa oleh sel kelenjar ambing membutuhkan IGF-I (pada sapi) dan hormon insulin (selain sapi)
• Potensi aktivitas insulin dan IGF-I dapat ditingkatkan oleh Glucoce Tolerance Factor (GTF)
• GTF adalah suatu ligan yang terdiri dari Cr3+ dan asam amino glisin, sistein, asam glutamat dan niasin
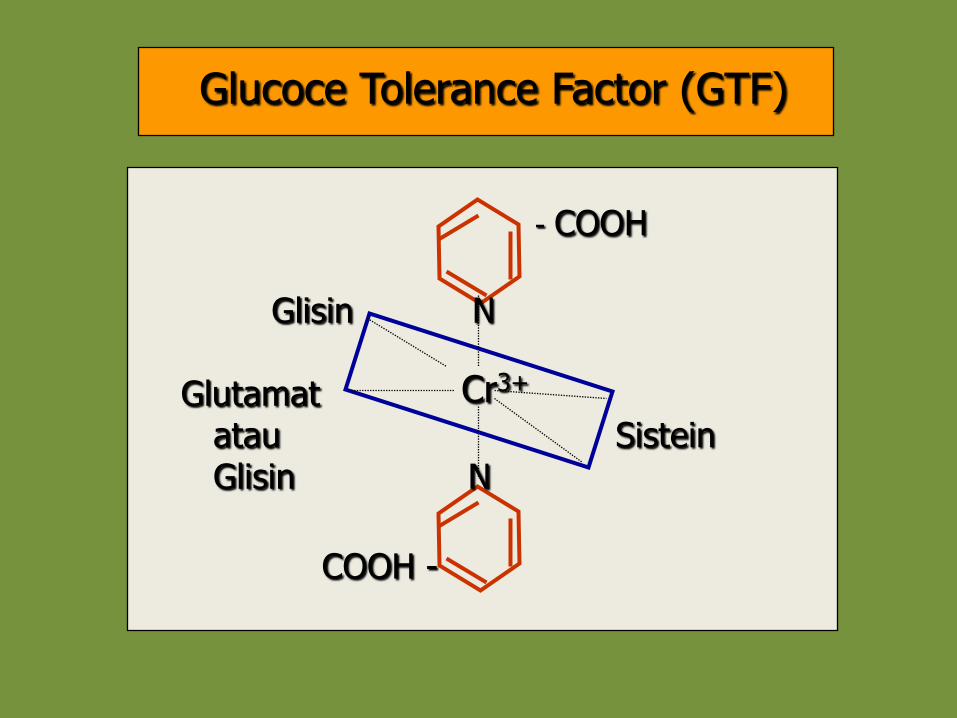
Cr3+
- COOH
Glisin N
Glutamat atau Sistein Glisin N
COOH -
Glucoce Tolerance Factor (GTF)

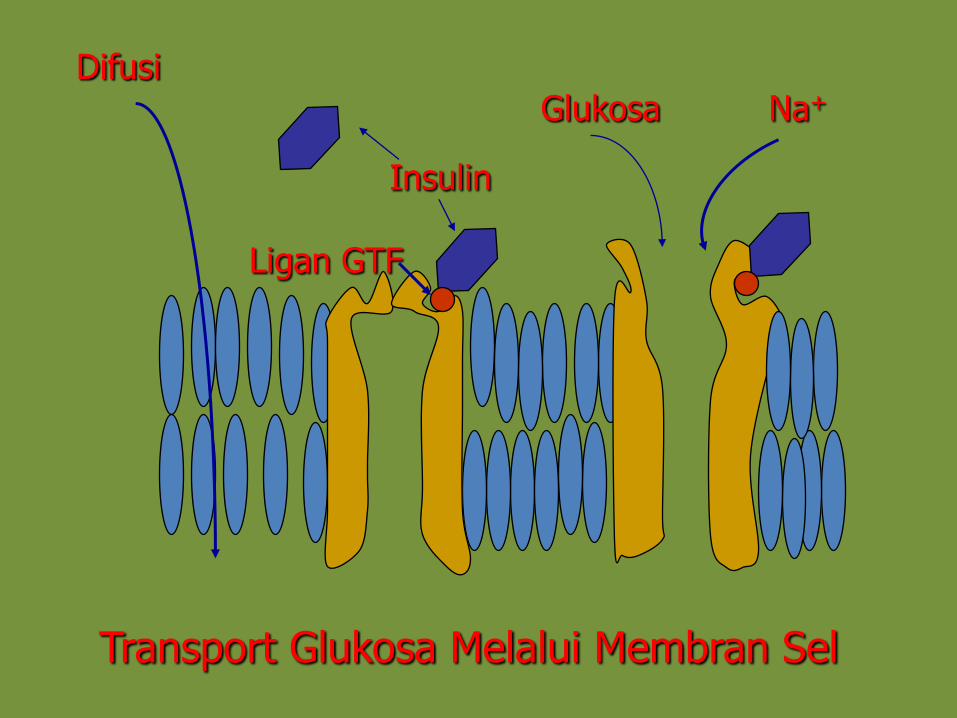
DifusiGlukosa Na+
Insulin
Ligan GTF
Transport Glukosa Melalui Membran Sel

4. Biosintesis Protein Susu (Casein)
• Terjadi di ribosom sekretoris kelenjar ambing, menggunakan
bahan baku asam-asam amino
• Kadarnya dipertahankan tetap yaitu 2,9 – 5%
• Kandungan asam amino esensial tertinggi dalam susu adalah
leusin, lisin dan valin; sedangkan asam amino non esensial
tertinggi adalah asam aspertat, asam glutamat dan prolin.
• Sintesis protein susu membutuhkan energi dalam bentuk ATP.

Ringkasan Tahapan Reaksi Sintesis Protein Susu
1. Asam amino + ATP -----------> Aminoasil-AMP +Pi
2. Aminoasil-AMP + t-RNA ----------- > Aminoasil-tRNA + AMP
3. (Aminoasil-tRNA)n + GTP ---------- > AA1–AA2–AA3 +GDP+tRNA

Kandungan AA dalam Protein air susu (g/100 g protein)
AA esensial Sapi Babi AAnon esensial Sapi Babi
Arginin
Histidin
Isoleusin
Leusin
Lisin
Metionin
Fenilalanin
Treonin
Triptofan
Valin
3.6
2.7
5.6
9.7
7.9
2.5
5.2
4.6
1.3
6.6
5.6
2.5
5.1
8.2
6.0
1.5
4.2
4.1
1.3
5.2
Alanin
As. Aspartat
Sistin
As. Glutamat
Glisin
Prolin
Serin
Tirosin
3.6
7.2
0.7
23.0
2.0
9.2
5.8
5.1
-
5.1
1.6
11.2
2.0
-
5.8
5.5




DARAH MEMBRAN SEL SEL KELENJAR AMBING
2-monogliserida
Triasilgliserol
(Kilomikron, LDL) FFA Asam lemak Lemak
susu
Gliserol -gliserol-P
NEFA
Asetat Asetat
-hidroksibutirat -hidroksibutirat
Triose-P NADP+
PPC
Glukosa 6-P NADPH
Glukosa Glukosa Glukosa-1-P-UDP Laktosa
susu
Sitrat
Isositrat -ketoglutarat
OP
NAD+ NADH
ADP ATP
P i
Asam amino Asam amino Amino Protein
Acil-AMP susu
Mineral (Ca+, P+, Cl-, K+, Mg+, Na+) Mineral
(Ca+, P+, Cl-, K+, susu
Mg+, Na+)
*NEFA = Asam lemak tak teresterifikasi ; PPC = Siklus pentosa fosfat ;
OP = Fosforilasi oksidatif
Biosintesi Komponen Susu Di Dalam Sel Sekretoris Kelenjar Ambing





Top Related