







Languages
Pages
Legal


ArrestinexperimentsTheterminationstepinGPCRsignalinginvolvesatwo-stepmechanism-arapidphosphorylationofthereceptorfollowedbythearrestinbindingthatstericallyblocksfurtherinteractionofG-proteinwiththereceptor,resultinginterminationoftheprimarysignalingevent.WehavepreviouslyreportedcrystalstructuresandSAXSstudiesonconstitutivelyactivevariantsofvisualarrestin(p44-anaturallyoccurringtruncatedvariant;andR175E-achargereversalarr-1mutantinthepolarcore).Currently, we are investigating the effect of synthetic peptides that mimic thephosphorylated C-terminus of rhodopsin on arr-1 variants. Our preliminary resultsusing size-exclusion chromatography (SEC) show complex formation and a shift inelutionprofile,suggestingachangeinoligomericstate.IntheSAXSexperimentonBM29wehaveinvestigatedstructuralchangesuponbindingofasyntheticpeptidethatactivatesarrestines.Belowisasummaryofthekeyresults:Ascontrolswehavemeasuredtheproteinsp44andR175Ewithoutthepeptidebeingpresent(fig1).Thecontrolexperimentsdemonstratedthatbothproteinsarepresentinsolution.P44showsabsenceofaggregation,R175Ehadasmallcorrectableamountofaggregation.
Figure1:SAXSdataofp44andR175Eintheabsenceofsyntheticpeptide.Theredsolidlinearecalculatedtheoreticalscatteringcurvesbasedontheavailablecrystalstructuresofp44andR175E,respectively.
0 0.1 0.2 0.3 0.4 0.5
0.1
1
10
100
I(q)
3UGU
P44
0 0.1 0.2 0.3 0.4 0.5
q / Å-10
0.01
0.02
0.03
0.04
0.05
0.06
I(q)*
q2
0 0.1 0.2 0.3 0.4 0.5
0.1
1
10
100
I(q)
R175E
0 0.1 0.2 0.3 0.4 0.5
q / Å-10
0.02
0.04
0.06
0.08
0.1
I(q)*
q2

Inasecondstepweaddedthesyntheticpeptidetothep44andR175EsolutionsandrecordedSAXSdata.Theexperimentaldataisshowninfig.2.
Figure2:MeasuredSAXSdataofp44andR175inthepresenceofpeptide.Thesolidlinesaremodelsofdimericproteins.
Themeasuredforwardscatteringdemonstratesdirectlythatpeptidebindinginducestheassemblyofp44andR175Edimers.Possibleassembliesofdimershavebeenscreenedandwecoulddeterminetopossibledimerisationassemblies,whichareactuallyverysimilarintheiroverallshape,butdifferintherotationofthemonomersaroundthelongaxis.

KineticexperimentsofLOVproteinsafterlight-excitationOur group members have determined crystal structures of, PpSB1-LOV, DsLOV ofDinoroseobacter shibae DFL12T, and more recently, dark state crystal structure ofPpSB1-LOV. Comparison of the two states reveal remarkably large conformationalchanges including~11Åmovementof theC-terminalhelix Jα,disruptionofhydrogenbondsinthedimerinterface,anda~29°rotationofchainBrelativetochainA.Here,wewouldliketoperformtime-resolvedkineticmeasurementsofdarkrecoveryofPpSB1-LOVproteinvariantsusingSAXS.WehaveperformedkineticSAXSexperimentsofPbSB1-R66I.TheproteinsolutionwasilluminatedbybluelightandtherecoveringprocessinthedarkwasfollowedbykineticSAXSexperiments.ThedifferenceSAXSspectrum(lightilluminatedatt=0minusdarkatt)atdifferenttimesisshowninfigure3.Acharacteristicdropisfoundat0.05Å-1,whichwasalsoobservedinstaticSAXSexperimentperformedbeforeonBM29.
Figure3:DifferenceSAXSspectrumoflight-illuminatedR66Iminusproteininthedarkatdifferenttime-points.
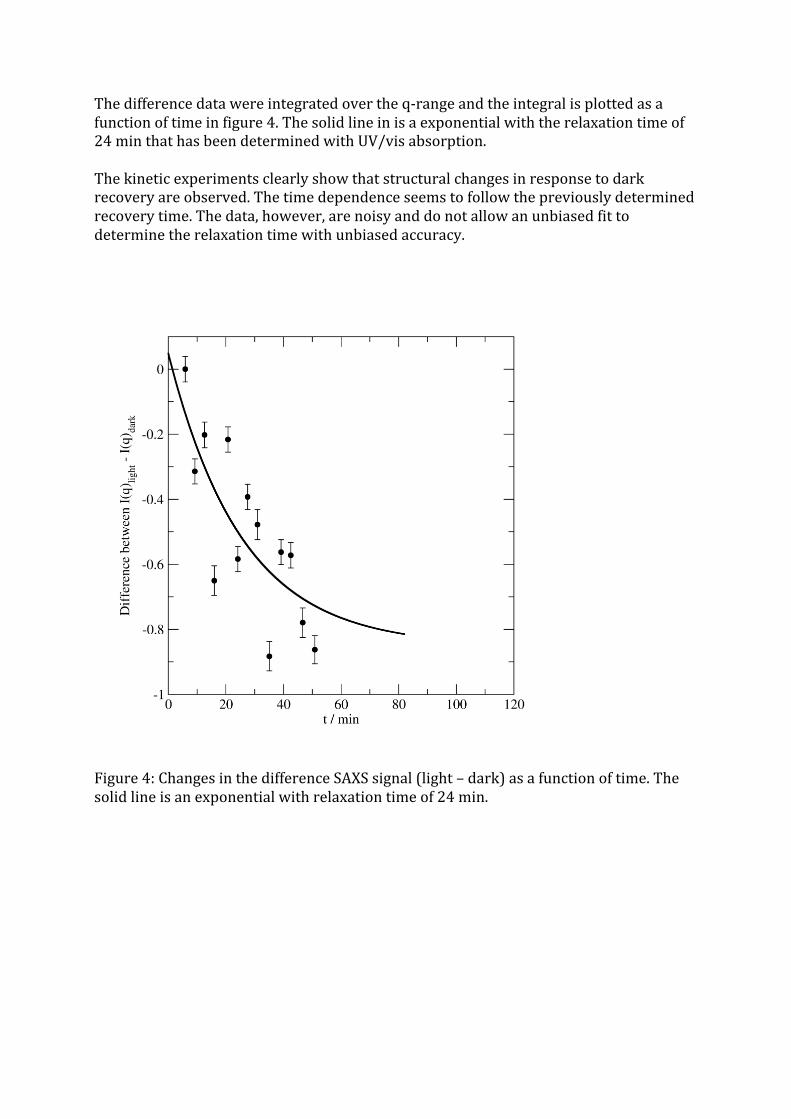
Thedifferencedatawereintegratedovertheq-rangeandtheintegralisplottedasafunctionoftimeinfigure4.Thesolidlineinisaexponentialwiththerelaxationtimeof24minthathasbeendeterminedwithUV/visabsorption.Thekineticexperimentsclearlyshowthatstructuralchangesinresponsetodarkrecoveryareobserved.Thetimedependenceseemstofollowthepreviouslydeterminedrecoverytime.Thedata,however,arenoisyanddonotallowanunbiasedfittodeterminetherelaxationtimewithunbiasedaccuracy.
Figure4:ChangesinthedifferenceSAXSsignal(light–dark)asafunctionoftime.Thesolidlineisanexponentialwithrelaxationtimeof24min.
Top Related