
Xenobiotic metabolism in the isolated conceptus
13
Toxic. in Vitro Vol. 4, No. 4/5, pp. 480-492, 1990 0887-2333/90$3.00 + 0.00 Printed in Great Britain.All rights reserved Copyright © 1990Pergamon Pressplc XENOBIOTIC METABOLISM IN THE ISOLATED CONCEPTUS R. BECHTER and G. D. C. TERLOUW Toxicology Department, Drug Safety Assessment, Sand0z Ltd, CH-4002, Basle, Switzerland Abstract--The development of culture systems using either pre- or post-implantation embryos has made it possible to study the metabolizing capacity of the isolated conceptus in vitro. In the rodent pre-implantation embryo and post-implantation conceptus (embryo and its membranes), constitutive levels and inducibility of different enzyme systems involved in drug metabolism have been shown in vitro to lead to the formation of embryotoxic metabolites of different xenobiotics. This indicated the presence of enzyme systems during early organogenesis. For example, using the rat post-implantation embryo culture, we could show that incubation with the lipoxygenaseinhibitor N-hydroxy-N-methyl-7-propoxy-2- naphthalenethanamine (QAB) led to high levels of the main in vivo metabolite 7-propoxy-naphthalene-2- ylacetic acid (QAA) and two as yet unidentified products, M5 and M6, in the conceptus. QAB was not found in tissues and QAA itself did not enter the embryonic compartments. In addition, accumulation in tissue was dependent on the time and duration of exposure. It started at 10.5 days of development. A similar metabolite pattern was obtained after yolk-sac tissue had been cultured alone, which suggests metabolizing capacity of mainly the yolk-sac tissue. The enzyme reactions involved might have included oxidative N-demethylation and oxidative deamination, probably also including the formation of reactive intermediate metabolites. In conclusion, our data demonstrate that not only maternal metabolism may play an important role in the toxic action of xenobiotics, but also the metabolizing capacity of the conceptus itself may be crucial, since the formation of (intermediate, highly reactive) metabolites takes place at the target site. Introduction In the late 1970s and early 1980s the drug metaboliz- ing capacities (Phase I and Phase II reactions) of the non-human mammalian embryo were believed to be absent or very limited (Dutton, 1978 and 1982; Neims et al., 1976; Pelkonen, 1978 and 1980). The lack of information about the development of these enzy- matic systems during the embryonic phase was partly due to the lack of appropriate biological and ana- lytical tools at that time. However, Filler and Lew (1981) demonstrated the onset of mixed function oxidase activities at the mouse blastocyst stage after in vitro development from the 2-cell stage onwards. They showed differential inducibility with tetra- chlorodibenzo-p-dioxin of the cytochrome P-450- dependent metabolism of benzo[a]pyrene (BaP) in mouse embryo cultures of a non-responsive and a responsive strain by using high-performance liquid chromatography (HPLC) to determine the metabolic products of BaP. Galloway et al. (1980) observed that the frequency of sister chromatid exchange increased in post-implantation mouse embryos of BaP-responsive strains in comparison with non- responsive strains after the culture of 7.5- and 8.5-day-old embryos in BaP. Pederson et al. (1985) also demonstrated, using the same technique, Abbreviations: AAF = 2-acetylaminofluorene; BaP = benzo- [a]pyrene; DPH = diphenylhydantoin; HPLC = high- performance liquid chromatography; MC = 3-methyl- cholanthrene; PB = phenobarbital; PO = phenoxazone (Resorufine); QAA = 7-propoxy-naphthalene-2-ylacetic acid; QAB = N-hydroxy-N-methyl-7-propoxy-2-naph- thalenethanamine; TMAH = tetramethylammonium hy- drogen sulphate monohydrate. cytochrome-P-450-dependent enzyme activities in embryonic and extra-embryonic tissues of these post- implantation embryos in culture. The compound 2-acetylaminofluorene (AAF) has been shown to require bioactivation to exert its embryotoxic action in rat embryos in vitro (Juchau et al., 1985). Growth, differentiation and morphology of 10-day-old rat embryos were not affected after direct exposure to AAF in vitro (Faustman-Watts et aL, 1983). After pre-exposure of pregnant dams to 3- methylcholanthrene (MC) but not to phenobarbital (PB) prior to the excision and culture of the 10-day- old embryos, AAF produced a high proportion of embryos with specific morphological anomalies (Faustman-Watts et al., 1984; Juchau et al., 1985). The inclusion of carbon monoxide in the cultures dramatically decreased the frequency of embryos with anomalies (Faustman-Watts et al., 1984 and 1986). The results of these studies suggest the presence of MC but not PB-inducible cytochrome- P-450 isozymes in these early post-implantation rat embryos. A variety of phenoxazone ethers has been used as specific probe substrates for the detection of a variety of cytochrome-P-450 isozymes (Burke and Mayer, 1983; Guengerich et al., 1982; Klotz et al., 1984; Lubet et al., 1985). Through a combination of specific antibody probes, selective inhibitors and in utero induction, Yang et al. (1988) concluded that at least four functional cytochrome-P-450 isozymes (three constitutive and one MC inducible) were present in tissues (embryo proper, yolk-sac and ectoplacental cone) of the 1l-day-old rat conceptus. We could show that, in 9.5- to l l.5-day-old concepti in vitro, N-hydroxy-N-methyl-7-propoxy-2- 480
Transcript of Xenobiotic metabolism in the isolated conceptus

Toxic. in Vitro Vol. 4, No. 4/5, pp. 480-492, 1990 0887-2333/90 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1990 Pergamon Press plc
XENOBIOTIC METABOLISM IN THE ISOLATED CONCEPTUS
R. BECHTER and G. D. C. TERLOUW Toxicology Department, Drug Safety Assessment, Sand0z Ltd, CH-4002, Basle, Switzerland
Abstract--The development of culture systems using either pre- or post-implantation embryos has made it possible to study the metabolizing capacity of the isolated conceptus in vitro. In the rodent pre-implantation embryo and post-implantation conceptus (embryo and its membranes), constitutive levels and inducibility of different enzyme systems involved in drug metabolism have been shown in vitro to lead to the formation of embryotoxic metabolites of different xenobiotics. This indicated the presence of enzyme systems during early organogenesis. For example, using the rat post-implantation embryo culture, we could show that incubation with the lipoxygenase inhibitor N-hydroxy-N-methyl-7-propoxy-2- naphthalenethanamine (QAB) led to high levels of the main in vivo metabolite 7-propoxy-naphthalene-2- ylacetic acid (QAA) and two as yet unidentified products, M5 and M6, in the conceptus. QAB was not found in tissues and QAA itself did not enter the embryonic compartments. In addition, accumulation in tissue was dependent on the time and duration of exposure. It started at 10.5 days of development. A similar metabolite pattern was obtained after yolk-sac tissue had been cultured alone, which suggests metabolizing capacity of mainly the yolk-sac tissue. The enzyme reactions involved might have included oxidative N-demethylation and oxidative deamination, probably also including the formation of reactive intermediate metabolites. In conclusion, our data demonstrate that not only maternal metabolism may play an important role in the toxic action of xenobiotics, but also the metabolizing capacity of the conceptus itself may be crucial, since the formation of (intermediate, highly reactive) metabolites takes place at the target site.
Introduction
In the late 1970s and early 1980s the drug metaboliz- ing capacities (Phase I and Phase II reactions) of the non-human mammalian embryo were believed to be absent or very limited (Dutton, 1978 and 1982; Neims et al., 1976; Pelkonen, 1978 and 1980). The lack of information about the development of these enzy- matic systems during the embryonic phase was partly due to the lack of appropriate biological and ana- lytical tools at that time. However, Filler and Lew (1981) demonstrated the onset of mixed function oxidase activities at the mouse blastocyst stage after in vitro development from the 2-cell stage onwards. They showed differential inducibility with tetra- chlorodibenzo-p-dioxin of the cytochrome P-450- dependent metabolism of benzo[a]pyrene (BaP) in mouse embryo cultures of a non-responsive and a responsive strain by using high-performance liquid chromatography (HPLC) to determine the metabolic products of BaP. Galloway et al. (1980) observed that the frequency of sister chromatid exchange increased in post-implantation mouse embryos of BaP-responsive strains in comparison with non- responsive strains after the culture of 7.5- and 8.5-day-old embryos in BaP. Pederson et al. (1985) also demonstrated, using the same technique,
Abbreviations: AAF = 2-acetylaminofluorene; BaP = benzo- [a]pyrene; DPH = diphenylhydantoin; HPLC = high- performance liquid chromatography; MC = 3-methyl- cholanthrene; PB = phenobarbital; PO = phenoxazone (Resorufine); QAA = 7-propoxy-naphthalene-2-ylacetic acid; QAB = N-hydroxy-N-methyl-7-propoxy-2-naph- thalenethanamine; TMAH = tetramethylammonium hy- drogen sulphate monohydrate.
cytochrome-P-450-dependent enzyme activities in embryonic and extra-embryonic tissues of these post- implantation embryos in culture.
The compound 2-acetylaminofluorene (AAF) has been shown to require bioactivation to exert its embryotoxic action in rat embryos in vitro (Juchau et al., 1985). Growth, differentiation and morphology of 10-day-old rat embryos were not affected after direct exposure to AAF in vitro (Faustman-Watts et aL, 1983). After pre-exposure of pregnant dams to 3- methylcholanthrene (MC) but not to phenobarbital (PB) prior to the excision and culture of the 10-day- old embryos, AAF produced a high proportion of embryos with specific morphological anomalies (Faustman-Watts et al., 1984; Juchau et al., 1985). The inclusion of carbon monoxide in the cultures dramatically decreased the frequency of embryos with anomalies (Faustman-Watts et al., 1984 and 1986). The results of these studies suggest the presence of MC but not PB-inducible cytochrome- P-450 isozymes in these early post-implantation rat embryos.
A variety of phenoxazone ethers has been used as specific probe substrates for the detection of a variety of cytochrome-P-450 isozymes (Burke and Mayer, 1983; Guengerich et al., 1982; Klotz et al., 1984; Lubet et al., 1985). Through a combination of specific antibody probes, selective inhibitors and in utero induction, Yang et al. (1988) concluded that at least four functional cytochrome-P-450 isozymes (three constitutive and one MC inducible) were present in tissues (embryo proper, yolk-sac and ectoplacental cone) of the 1 l-day-old rat conceptus.
We could show that, in 9.5- to l l.5-day-old concepti in vitro, N-hydroxy-N-methyl-7-propoxy-2-
480
naphthalinethanamine (QAB) was embryotoxic at concentrations higher than 3.9 #M, whereas its main in vivo metabolite 7-propoxy-naphtha!ine-2-ylacetic acid (QAA) did not lead to embryotoxicity up to the highest concentration of 115 #M tested (Bechter and Brouillard, 1988). Determinations of medium and tissue levels of the parent compound and its metab- olites by HPLC revealed that no QAB was present in the tissues of the conceptus. QAA and two as yet unidentified products, M5 and M6, were found at high concentrations in the embryo proper, yolk-sac tissue and the ectoplacental cone. Direct exposure to QAA did not lead to substantial tissue levels of either QAA or the other products of QAB. These results suggested that the conceptus metabolized QAB and accumulated its metabolites. Preliminary experiments in which concepti were exposed in vitro to QAB for different lengths of time revealed that the conceptus metabolized QAB and accumulated its metabolites only when the conceptus was 10.5 days or older (Terlouw et al., 1989).
The aim of the present study was to further characterize the onset of the capacity of the isolated conceptus to metabolize QAB in vitro and to accumu- late its products. Furthermore, an attempt was made to identify the tissue(s) of the conceptus responsible for this metabolic activity.
3 . 6
Materials and Methods
4 . 0 45 -
Animals. Female Han Wistar rats (Kfm:WSA, Kleintierfarm Mad6rin, Fiillinsdorf, Switzerland), 10-11 wk old (180-230 g), were used for study. The animals had free access to rat pellets (No. 24.343.7, Klingenthalmiihle AG, Kaiseraugst, Switzerland) and bottled tap-water and were kept in an air- conditioned animal room at a temperature of 22-25°C and a relative humidity of not less than 40%. Artificial light was provided for a 12-hr light/dark cycle. Two females were mated with one Han Wistar male overnight. The next morning (day
0 of gestation) the vaginal smears were checked for the presence of sperm to determine whether copulation had occurred.
Whole embryo culture. Embryos (early somite stage) within the intact visceral yolk-sac and amnion were explanted from the uteri of 9.5-day pregnant dams. The embryos were cultivated in heat-inacti- vated, pure male rat serum for 48 hr with or without a drug activating system according to the method of New (1978) as described in Bechter and Brouillard (1988). The concentration of QAB used in the whole embryo cultures was 30 #g/ml (115/tu).
The concepti were exposed to the compound at different culture times for various lengths of time (Fig. 1). After the concepti had been washed in phosphate buffered saline fresh medium was added. At least 8 embryos were cultured in each group. Control cultures received only 100/tl of the gelatine solution and, where appropriate, the activating system. At the end of the 48 hr culture period the diameter of the yolk-sac, its vascularization and circulation, the embryonic heart beat and rotation to concave position were evaluated under a dissecting microscope. Crown-rump and head lengths were taken as indicators of embryonic growth. The degrees of differentiation were evaluated by using the slightly modified scoring system of Brown and Fabro (1981), and by counting the somites. Embryonic features, including flexion, heart, fore, mid and hindbrain, otic, optic and olfactory systems, branchial bars, mandibular and maxillary processes, fore and hind- limbs and somite formation were examined and anomalies recorded.
Yolk-sac culture. Embryos were explanted either from the uteri of 10.5-day or 11.5-day pregnant dams. After having removed the decidua, trophoblastic remnants, Reichert's membrane, and the parietal yolk-sac, the visceral yolk-sac was separated from the embryo and ectoplacental cone, and cultivated in heat-inactivated, pure male rat serum for 24 and 48 hr. Four yolk-sacs per culture flask (30 ml) were
3 . 4
3 .2
3 . 0
37 • 20"
Contro L
Q A B e x p o s u r e t i m e (h)
4 3 -
0 0 -
8 0 -
41 - 60 "
5 9 - 4 0 "
3 5 , 0 0 -48
3 . 8
0 - 1 5 t 5 - 24 2 4 - 4 8 15 -48 0 - 4 8 °
Xenobiotic metabolism in isolated conceptus 481
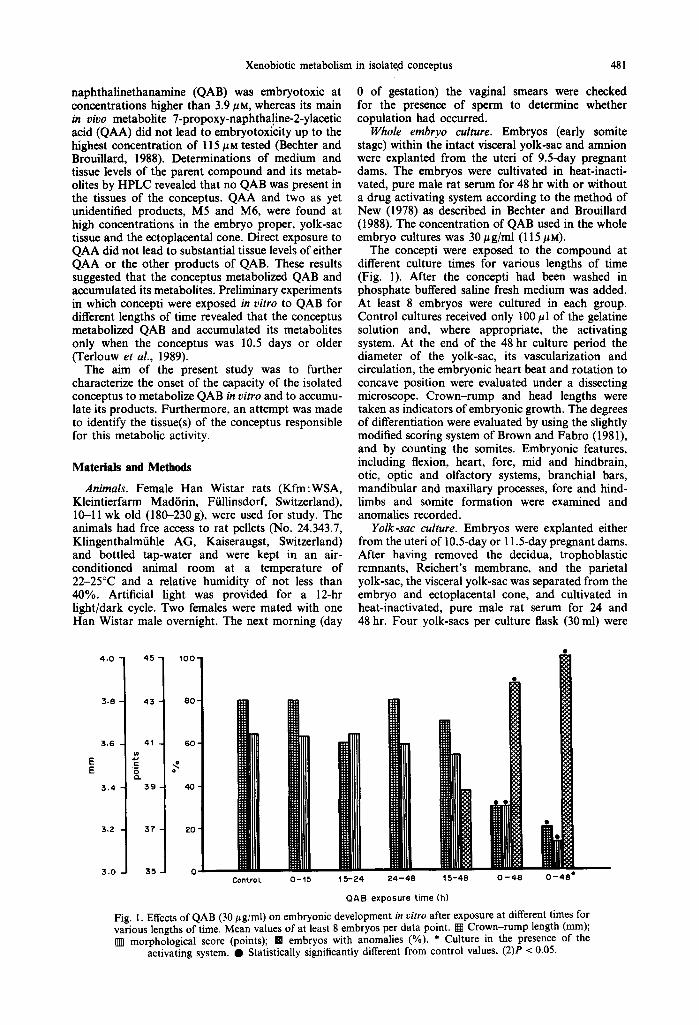
Fig. I. Effects of QAB (30 #g/ml) on embryonic development in vitro after exposure at different times for various lengths of time. Mean values of at least 8 embryos per data point. I~ Crown-rump length (mm); [] morphological score (points); B embryos with anomalies (%). * Culture in the presence of the
activating system. • Statistically significantly different from control values, (2)P < 0.05.

482 R. BECHTER and G. D. C. TEgLOUW
grown in 5 ml medium at 37°C using a roller bottle apparatus (Her/ius, type B5060 EK/CO2) and inter- mittent gassing with an oxygen, nitrogen and carbon dioxide mixture. Cultivation took place both in the presence and absence (control) of the test compound. The concentrations used in the yolk-sac cultures were 30 and 60/~g/mi (115 and 230 #U). Drug metabolism was induced in a part of the experiments by the addition of 30/~1 of a rat liver microsomal prep- aration containing approximately 40 mg protein/ml, Aroclor 1254-induced, according to Maron and Ames (1983), and NADPH (5 x l0 -4 M) to each culture flask. Control cultures received only 100 gl of the gelatine solution and, where appropriate, the activating system.
Test compounds. The test compound QAB was obtained from the Internal Drug Center, Sandoz Ltd, and QAA was provided by Dr M. Strasser, Sandoz Ltd. The compounds were dissolved or suspended in a 2% gelatine solution immediately before use; 100 #l of the solutions/suspensions were then added to each culture flask, which contained 5 ml of medium.
Determination o f QAB and its metabolites in medium and tissue homogenates. Medium samples (100/zl) from each whole embryo culture and yolk- sac culture were taken at different time points during the culture period, and immediately frozen and stored at - 20°C until analysis. At the end of the culture period of the whole embryos, after the yolk-sac diameters had been measured and the embryos had been evaluated, the tissues (embryo proper, yolk-sac and ectoplacental cone) were frozen and stored indi- vidually. They were further evaluated as described by Bechter and Brouillard (1988). Tissue samples from the yolk-sac cultures were taken after 24 and 48 hr of cultivation, frozen, and stored individually until further evaluation. QAB and its metabolites/ breakdown products were extracted from the medium samples and spiked serum samples by ultrasonication (MSE Sonoprep 150 Ultrasonic Disintegrator) of 100/11 of medium with 900/zl of HPLC mobile phase (70% acetonitrile/30% of a 0.2% aqueous solution of tetramethylammonium hydrogen sulphate mono- hydrate (TMAH)) at an amplitude of 10/~ for 2 x 20sec. After 5min sedimentation time the samples were centrifuged (Eppendorf Centrifuge 5415, 5 min at 14,000 rpm/min), 100/~l of the super- natant was diluted to l ml and then 10#l was analysed. 300/~l of HPLC mobile phase was added to the tissues. They were then homogenized by ultrason- ication at an amplitude of 10/~ for 2 × 20 sec. After another 700/~1 of mobile phase was added, the hom- ogenates were centrifuged for 5 min at 14,000 rpm/ rain. Depending on the concentration of QAB and its metabolites, the supernatant was diluted 2 or 4 times; 10/~l was analysed. HPLC (Perkin-Elmer ISS-100, 410 LC Pump, LCI-100, Hewlett-Packard HP 1046A) was performed using a reverse phase system with a RP-8 precolumn (30 x 4.6mm, 5#m; Kon- tron) and a Spheri-5 RP-18 column (100 x 4.6mm, 5/lm; Kontron). 70% acetonitrile (HPLC grade S, Rathburn Chemicals Ltd, Scotland)/30% of a 0.2% aqueous solution of TMAH (Fluka Chemic AG, Switzerland) was used as mobile phase. The flow rate was 1.2 ml/min, the excitation wavelength 229 nm, and the emission wavelength 333 nm.
Electron microscopy. One yolk-sac tissue per culture flask and sampling time point was fixed and the yolk-sac prepared for electron microscopy according to the method of Schmid et al. (1985). They were examined by using a Philips 300 electron microscope. In addition, visceral yolk-sac tissues from pregnant dams were prepared on days 10.5 to 13.5 as in vivo controls.
Statistics. Statistical analyses of the differences in embryonic growth and differentiation between the control and treated groups were performed using the two-sided Student's t-test (homogeneous distri- bution) or the t-test of Welch (inhomogeneous distribution). The statistical significance of an increased incidence of embryos with anomalies, in comparison with the controls, was determined using the one-sided Fisher test. The Kruskal-Wallis mul- tiple comparison test or the Newman-Keuls multiple range test was used to test for statistical differences of embryonic parameters as well as medium and tissue levels due to the use of the different culture conditions and compounds. The level of significance was set at 0.05.
Resul t s
Whole embryo culture
The rat conceptus within its visceral yolk-sac was exposed in vitro to QAB for various lengths of time and at various time points during the 48-hr culture period (Fig. 1). Embryonic growth and differentiation were marginally impaired if the compound was pre- sent in the medium (30#g/ml) from 15 to 48 hr of culture. In this set, 38.5% of the embryos were morphologically abnormal. All embryos exposed to the compound for shorter periods of time and/or at earlier stages developed normally. Incubation of the conceptus with QAB during the entire culture period significantly inhibited embryonic growth and differentiation and led to 87.5% of embryos with anomalies. These effects were more pronounced in the presence of the S-9 mix; all embryos had morpho- logical anomalies.
QAA and M6 were found in culture medium at concentrations below 0.1/lg/ml (Fig. 2) at the end of the 48-hr drug incubation with the medium alone. Similar concentrations were determined in cultures with whole embryos incubated with QAB from 0 to 15 or 15 to 24 hr of the culture period. The concen- trations were significantly increased in the medium of QAB containing cultures of whole embryos older than 10.5 days, and they were independent of the exposure time to QAB. Addition of the hepatic drug activation system to the cultures of 0-48 hr led to an increase of medium metabolite concentrations at the end of the culture period. In contrast, M5 was present in considerable amounts in cultures of QAB and medium alone. Its concentration did not increase in cultures using whole embryos, with the exception of the experiment where the S-9 mix had been added.
In yolk-sac and embryonic tissue of concepti exposed to QAB prior to the 24-hr time point, only very small amounts of QAA, M5 and M6 were detected (Fig. 3). Exposure to QAB after this time point led to a considerable accumulation of the three products in tissue of the embryo proper, the yolk-sac

Plate 1. Morphology of normal visceral yolk-sac tissue prepared e x vivo on days 10.5 (a), 11.5 (b), 12.5 (c) and 13.5 (d) post coitum. Lysosomes (L), Microvilli (MV) at apical side, nucleus (N), mitochondrion
(M), endoplasmic reticulum (ER), lipid droplets (LD).
483
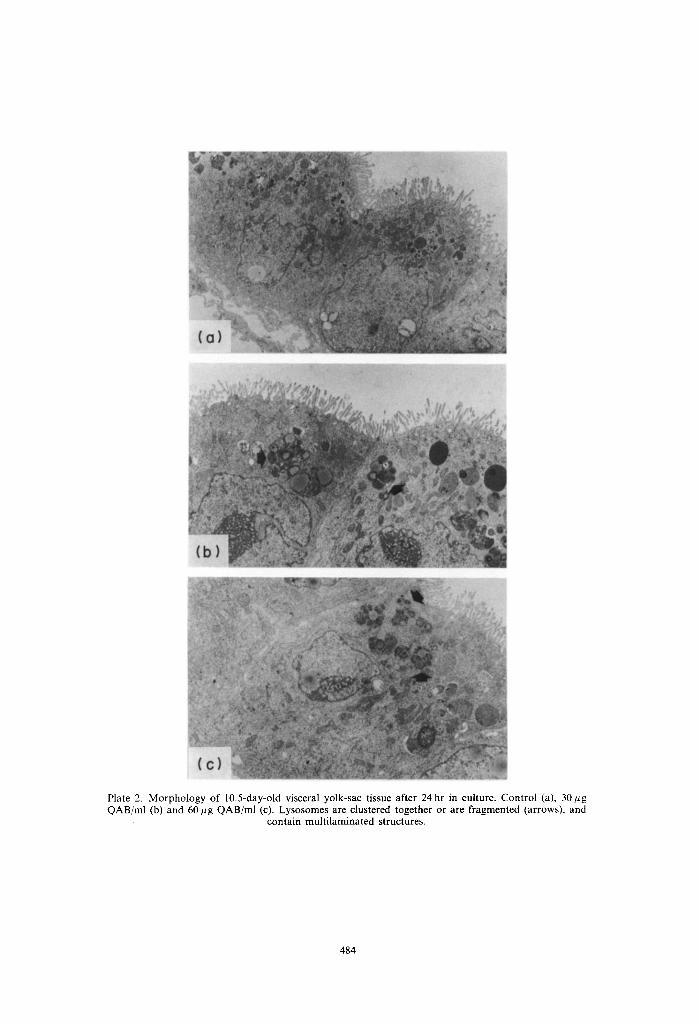
Plate 2. Morphology of 10.5-day-old visceral yolk-sac tissue after 24 hr in culture. Control (a), 30 gg QAB/ml (b) and 60 ~g QAB/ml (c). Lysosomes are clustered together or are fragmented (arrows), and
contain multi laminated structures.
484
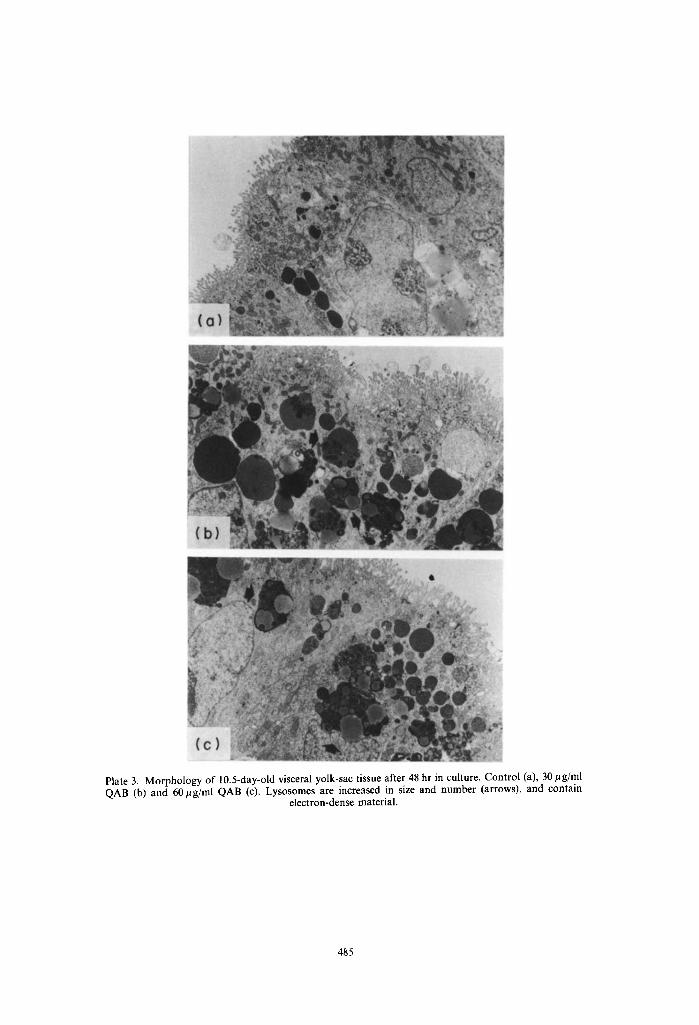
Plate 3. Morphology of 10.5-day-old visceral yolk-sac tissue after 48 hr in culture. Control (a), 30/~g/ml QAB (b) and 60/tg/ml QAB (c). Lysosomes are increased in size and number (arrows), and contain
electron-dense material.
485
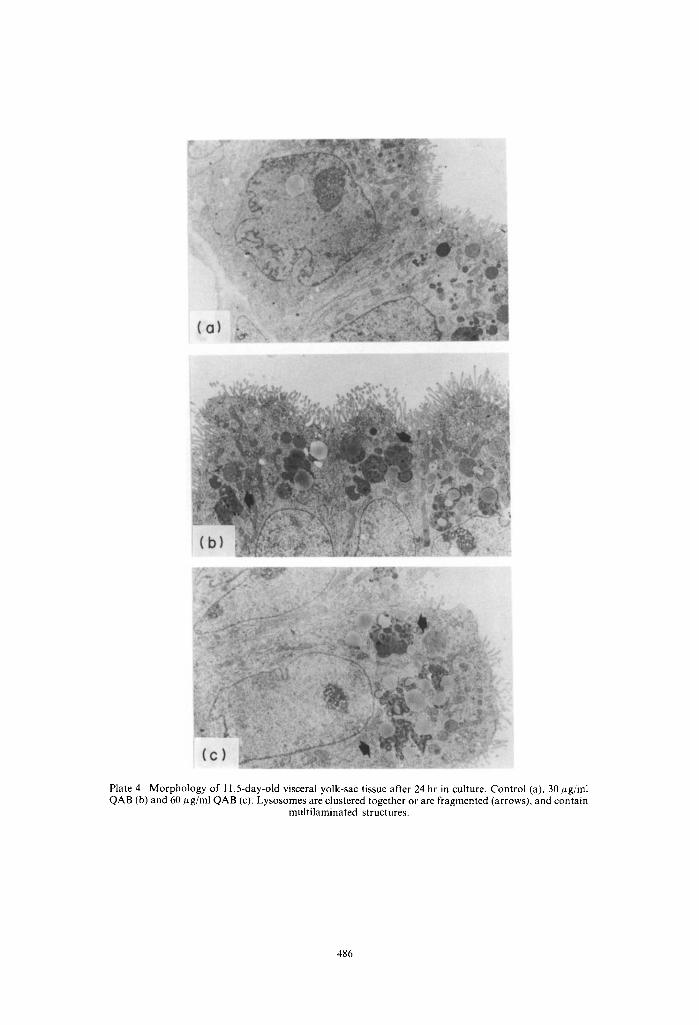
Plate 4. Morphology of 11.5-day-old visceral yolk-sac tissue after 24 hr in culture. Control (a), 30 #g/mI QAB (b) and 60 F~g/ml QAB (c). Lysosomes are clustered together or are fragmented (arrows), and contain
multilaminated structures.
486
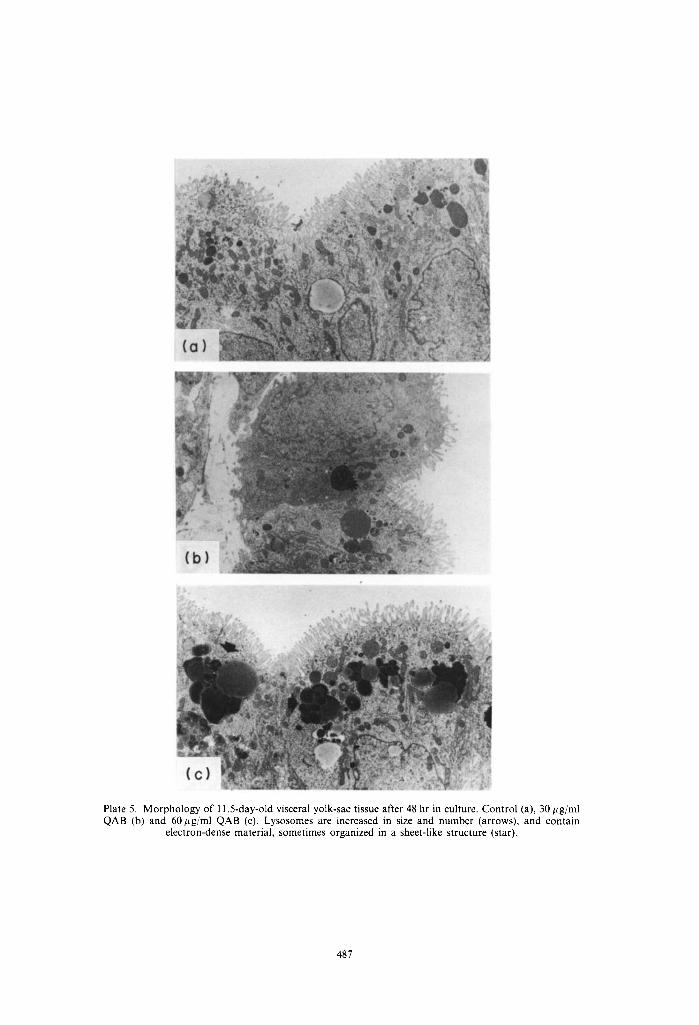
Plate 5. Morphology of 11.5-day-old visceral yolk-sac tissue after 48 hr in culture. Control (a), 30 ~g/ml QAB (b) and 60/~g/ml QAB (c). Lysosomes are increased in size and number (arrows), and contain
electron-dense material, sometimes organized in a sheet-like structure (star).
487


1.6 _
1.4
1.:>
E 1.0 --
~) 0 . 8 --
o "~" 0 . 6 -
~ 0.4
0.2
O~
01
No embryo 0 - -15
O
ooL 15- -24 2 4 - - 4 8 1 5 - - 4 8 0 - - 4 8 0 - - 4 8 ~'
QAB exposure t i m e ( h )
Fig. 2. Medium concentrations of QAA, M5 and M6 after exposure with QAB (30/~g/ml) at different times for various lengths of time. Mean values of samples of at least 3 culture bottles per data point. [ ] QAA; [] M5; [] M6. *Culture in the presence of the activating system. (3 Statistically significantly different
from the value 0-48 hr without activation, (2)P < 0.05.
1 0 0
8 0
6 0
4 0
2 0
0 0 0
0 --15
( a )
0 O0
1 5 - - 2 4
O o o
t 5 - - 2 4
o
24 -- 48 15 - - 4 8 O-- 4 8 0 -- 4 8 ~
QAB exposure ¢ i m e ( h )
2 0 (b)
~ 10-
g
0 0 0 0
0 - 1 5 24 - - 4 8 15 - - 4 8 0 - - 4 8 0 - - 4 8 *
QAB exposure t i m e ( h )
Xenobiotic metabolism in isolated conceptus 489
Fig. 3. Concentrations of QAA, M5 and M6 in tissue of the yolk-sac (a) and embryo proper (b) after exposure with QAB (30 #g/ml) at different times for various lengths of time. Mean values of at least 8 tissue samples per data point. I~ QAA; [] M5; [] M6. *Culture in the presence of the activating system.
© Statistically significantly different from the value 0-48 hr without activation, (2)P < 0.05.

490 R. BECHTER and G. D. C. TERLOUW
Table 1. Accumulation of the products QAA, M5 and M6 in the tissues of the conceptus in vitro after exposure to QAB for 48 hr without the addition of
an activating system
Tissue QAA M5 M6
Embryo proper 5* 6 27 Ectoplacental cone 4 10 60 Yolk-sac 10 46 189
*Ratio of tissue level (ltg/g wet weight) to medium concentration (/~ g/ml).
and the ectoplacental cone (Table 1). In all tissues analysed, the ratio between tissue and medium concentration was the highest for M6 followed by that for M5 and QAA. The accumulation of the three products was most pronounced in the yolk-sac tissue. Drug activation by the S-9 mix generally caused statistically significantly increased tissue accumulation.
Yolk-sac culture
The morphology of in vivo grown visceral yolk-sacs (in vivo control) is shown in Plate 1. They had one layer of cylindrical cells, with microvilli at their apical side with a high lysosome content typical for their high pynocytotic activity. The in vitro development of yolk-sac tissue of 10.5- and l l.5-day-old concepti after 24 and 48 hr was comparable with in vivo development as judged by the morphological appear- ance of the tissues (Plate 2-5). A marked increase of partly fragmented inclusion bodies, most probably lysosomes, the majority of which contained multi- laminated structures, was found after 24 hours' culture time in the presence of 30 and 60 #g QAB/ml in both 10.5- and 11.5-day-old tissues (Plates 2 and 4). The effects were only partially concentration depen- dent. These lysosomes were huge, and had increased their numbers significantly at the end of the cultures of the 10.5-day-old tissues (Plate 3). In the tissues cultured from 11.5 days onwards, the size and num- bers of these lysosomal structures did not increase during the second day of culture in comparison with the first 24 hr. The contents of the vesicles after 48 hr in culture in both the 10.5- and 11.5-day cultures were more electron-dense than those found after 24 hr in culture and in part were of a sheet-like appearance. The addition of a hepatic activation system from the start of the cultures generally led to a more pronounced effect on these lysosomal structures (data not shown).
In contrast to the yolk-sac tissue of whole embryo cultures, QAB and its products QAA and M4 were found at limited concentrations in the tissues of 10.5- and 11.5-day-old yolk-sac cultured for 24 and 48 hr (Table 2). The products M5 and especially M6 were found at concentrations as high as 4 mg/g of wet tissue. The tissue concentrations were generally higher in the presence of 60 #g QAB/ml in the culture medium than at the lower QAB level, and higher in the yolk-sac cultured from day 11.5 onwards as compared with the younger set. In 10.5-day-old yolk- sac tissue the concentrations of M5 and M6 increased during the second day of culture in comparison with the first 24 hr. In contrast, in the older tissue cultured from 11.5 to 13.5 days, the concentrations of M5 and M6 were lower or at similar levels after 48 hours'
<
"?o
"t3
~'~
<
.o
O'
N g
N
-I 41 +I 41 4~ 4P 41 4~ +I 41
- - - ~ 41 41 41 41 41 41 41 41
q 41 +l 41 41 41 41 41 +i 4l
- o~
r414141414141414141
0 41~ 41 _ 41~ 4~ 41
41414141414141414r
4r4;+l+l÷141+l~P41
4 ; + r + l + l ÷
~ ~ 2 N
d
E o~
E =
,.~ .~,
~pSo
"o~ ~ ~
. .~.~ ++ ~

Xenobiotic metabolism in isolated conceptus
Table 3. Tissue accumulation of QAB and its products QAA, M4, M5, and M6 in yolk-sac of 10.5- and 11.5-day-old concepti after 24 and 48 hours' culture in the presence of QAB with and without an external
hepatic activation system (S-9 mix). Culture time (days)
10.5-11.5 10.5-12.5 11.5-12.5 11.5-13,5 Compound S-9 mix 30* 60 30 60 30 60 30 60
QAB - t 2.6~ 1.4 - 3.3 1.6 1.3 - 2.3 + 2.1 2.4 - 2.6 3.7 11.1 - 5.2
QAA - 16.7 6.3 1.6 2.9 3.3 1.3 0.8 0.8 + 14.4 10.9 3.9 2.5 7.1 6.7 1.2 2.4
M4 - 20.0 12.5 2.7 5.7 8.6 7.5 2.0 4.8 + 6.7 7.5 6.7 4.1 8.3 10.0 3.1 6.1
M5 - 87 103 143 169 423 198 144 238 + 93 93 373 117 233 93 105 84
M6 - 125 144 190 261 112 74 164 259 + 125 142 305 185 235 299 309 262
*Medium concentration of QAB at the beginning of the culture (#g/ml). tWithout ( - ) or with ( + ) S-9 mix. STissue accumulation as ratio between mean tissue concentration (#g/g wet weight) and mean medium
concentration (/a g/ml).
491
culture than in the yolk-sac tissue cultured for 24 hr, with the exception of M5 in the 60 # g QAB/ml group. QAB did, to a small extent, accumulate in yolk-sac tissues of all ages and culture duration (Table 3). QAA and M4 accumulated to a high extent only during the first 24hr in the 10.5-day-old tissue, although appreciable accumulation was also found after the first culture day of 11.5-day-old tissue. The accumulation of these two products was much less pronounced after the second day of culture of both the 10.5- and 11.5-day-old tissues. The products M5 and M6 were accumulated heavily, and in contrast to QAA and M4, more so after the second half of the cultures of 10.5-day tissue than during the first 24-hr incubations. M5 was found at higher levels in the tissue of 11.5-day-old yolk-sacs after 24 hr than ~fter 48 hours' culture. The ratios between tissue and medium concentrations of M6 were similar after 24 and 48 hours' culture.
Discussion
Exposure of whole embryos to QAB at different time points of development and for different dur- ations in vitro led to significant embryotoxicity if the conceptus was exposed to the compound for the whole culture period of 9.5-11.5 days. Embryos exposed to the compound at 10.5 days for 24 hr did not show impaired growth and differentiation, and no morphological anomalies were found. However, ac- cumulation of QAA, M5 and M6 took place only in concepti older than 10.5 days. These results can be explained by the fact that the embryo at 9.5 days was not yet surrounded by its yolk-sac and was therefore much more susceptible to direct chemical injury than the older conceptus, which was protected from a direct action of QAB by its membranes.
Incubation of QAB with medium alone for 48 hr led to approximately 0.6/~g M5/ml at the end of the culture period. The concentration of this product in the medium after the culture of the whole embryo did not increase significantly, which suggested that it was a breakdown product of QAB. QAA and M6 levels were significantly higher in cultures with whole embryos than in medium alone at the end of the culture period when the compound QAB had been
added during the second day. In earlier experiments it was shown that QAB and QAA did not cross the embryonic membranes in significant amounts (Bechter and Brouillard, 1988). These findings suggest QAB metabolism in concepti of 10.5 days of prenatal age leading to accumulation of QAA and M6. In addition, the high tissue levels of M5 indicated additional capacities of the yolk-sac tissue to trans- port (passively or actively) this product into the tissues in vitro.
Yolk-sac tissue cultured alone generally led to a similar pattern of QAB products. In contrast to their presence in the intact yolk-sac of a whole embryo, QAB and M4 were found at low levels in this tissue after it had been cultured alone. QAA was found in tissue of cultured yolk-sac at a similar concentration as in yolk-sac tissue exposed intact for 24 hr after 10.5 days. In the isolated yolk-sac the levels of this metabolite were stable thereafter, or decreased slightly with increasing culture time. In contrast, the tissue levels of the product M5 were somewhat higher in yolk-sac cultured alone than those obtained after culture of the whole embryo. The values for M6 in isolated yolk-sac cultures had to be divided by a factor of approximately 10 (because of a different Rr-value in the two HPLC methods used) before they could be compared with the tissue concentrations after a whole embryo culture. After this correction, the levels of M6 were approximately 1.5 times higher in yolk-sac tissue cultured alone than in the exper- iments with intact embryos. These slightly different metabolite patterns between yolk-sac tissues cultured separately and those of the whole embryo, might have been due to the fact that the tissue was also exposed to the culture medium from the basal side. This could have led to an unphysiological uptake of QAB and M4 from this side as well as to leakage of products (e.g. QAA) into the medium. Despite this fact, the metabolite pattern in yolk-sac tissue either grown separately or intact, showed a similar profile, which suggests QAB metabolism and accumulation of its products.
The experiments using isolated yolk-sac tissues generally showed an increase of metabolites during the culture from day 10.5 to 12.5, whereas increasing metabolite levels were found only after the first day

492 R. BECHTER and G. D. C. TERLOUW
of culture in tissue grown from day 11.5 to 13.5. This pattern of tissue concentrations, especially of the products M5 and M6 in the cultures without the addition of the S-9 mix, was also reflected by the numbers and the size of lysosomes observed in the respective tissue under the electron microscope. Whether the contents of these vesicles consist of QAB products (free or bound to e.g. phospholipide) needs to be determined.
In conclusion, the results of the present study suggest that the yolk-sac tissue was involved in QAB metabolism and accumulation of its products. Its activity was highest in yolk-sac tissue between 10.5 and 12.5 days of prenatal age. Currently ongoing metabolite isolation and identification in our lab will lead to a better understanding of enzymatic activities involved in xenobiotic metabolism in the conceptus and their pattern of development during the early organogenic phase.
Acknowledgements--We thank A. Bianchi, W. Gehrig, M. Gianella, B. Scheim, A. Trippmacher and A. Zwahlen for their excellent technical assistance, N. Koeth for editorial help and J. Saladin for typing the manuscript.
REFERENCES
Bechter R. and Brouillard J.-F. (1988) The effects of different chemical forms of a test compound on embryo- toxicity, distribution and metabolism in vitro. Toxic. in Vitro 2, 181-188.
Brown N. A. and Fabro S. (1981) Quantitation of rat embryonic development in vitro: a morphological scor- ing system. Teratology 24, 65-78.
Burke M. D. and Mayer R. T. (1983) Differential effects of phenobarbitone and 3-methylcholanthrene induction on the hepatic microsomal metabolism and cytochrome P- 450 binding of phenoxazone and a homologous series of its n-allyl ethers (alkoxyresorufins). Chemico-Biol. Inter- actions 45, 243-258.
Dutton G. J. (1978) Developmental aspects of drug con- jugation, with special reference to glucuronidation. A. Rev. Pharmac. Toxic. 18, 17-35.
Dutton G. J. (1982) Drug metabolism and development. In The Biochemical Development of the Fetus and the Neonate. Edited by C. T. Jones. pp. 832-844. Elsevier, Amsterdam.
Faustman-Watts E., Giachelli C., Greenaway. J., Fantel A., Shepard T. and Juchau M. (1984) Methylcholanthrene (MC) or phenobarbital (PB) pretreatment of rat embryos: effects on teratogenicity in vitro. Teratology 29, 28A.
Faustman-Watts E., Giachelli C. M. and Juchau M. (1986) Carbon monoxide inhibits monooxygenation by the conceptus and embryotoxic effects of proteratogens in vitro. Toxic. appl. Pharmac. 83, 590-595.
Faustman-Watts E., Greenaway J. C., Namkung M. J., Fantel A. G. and Juchau M. R. (1983) Teratogenicity in vitro of 2-acetylaminofluorene: role of biotransformation in the rat. Teratology 27, 19-28.
Filler R, and Lew K. J. (1981) Developmental onset of mixed-function oxydase activity in preimplantation
mouse embryos. Proc. natn. Acad. Sci. U.S.A. 78, 6991-6995.
Galloway S. M., Perry P. E., Meneses J., Nebert D. W. and Pedersen R. A. (1980) Cultured mouse embryos metabolize benzo[a]pyrene during early gestation: genetic differences detectable by sister chromatid exchange. Proc. natn. Acad. Sci. U.S.A. 77, 3524-3528.
Guengerich F. P., Dannan G. A., Wright S. T., Martin M. V. and Kaminsky L. S. (1982) Purification and characterization of liver microsomal cytochromes P-450: electrophoretic, spectral, catalytic and immunochemical properties and inducibility of eight isozymes isolated from rats treated with phenobarbital or fl-naphthoflavone. Biochemistry, N.Y. 21, 6019-6030.
Juchau M. R., Giachelli C.M., Fantel A. G., Greenaway J. C., Shepard T. H. and Faustman-Watts E. M. (1985) Effects of 3-methylcholanthrene and phenobarbital on the capacity of embryos to bioactivate teratogens during organogenesis. Toxic. appl. Pharmac. 80, 137-146.
Klotz A. V., Stegman J. J. and Walsh C. (1984) An alternative 7-ethoxyresorufin O-deethylase activity assay: a continuous visible spectrophotometric method for measurement of cytochrome P-450 monooxygenase ac- tivity. Analyt. Biochem. 140, 138-145.
Lubet R. A., Mayer R. T., Cameron J. W., Nims R. W., Burke M. D., Wolfe T. and Guengerich F. P. (1985) Dealkylation of pentoxyresorufin: a rapid and sensitive assay for measuring induction of cytochrome(s) P-450 by phenobarbital and other xenobiotics in the rat. Archs Biochem. Biophys. 238, 43-48.
Maron D. M. and Ames B. N. (1983) Revised method for the Salmonella mutagenicity test. Mutation Res. 113, 173-215.
Neims A. H., Warner M., Loughnan P. M. and Aranda J. V. (1976) Developmental aspects of the hepatic cytochrome P450 monooxygenase system. A. Rev. Pharmac. Toxic. 16, 427-446.
New D. A. T. (1978) Whole-embryo culture and the study of mammalian embryos during organogenesis. Biol. Rev. 53, 81-122.
Pederson R. A., Meneses J., Spindle A., Wu K. and Galloway S. M. (1985) Cytochrome P-450 metabolic activity in embyronic and extra-embryonic tissue lineages of mouse embryos. Proc. natn. Acad. Sci. U.S.A. 82, 3311-3315.
Pelkonen O. (1978) Prenatal and neonatal development of drug and carcinogen metabolism. In The Induction of Drug Metabolism. Edited by R. W. Estabrook and E. Lindenlaub. pp. 507-516. F. K. Schattauer Verlag, Stuttgart.
Pelkonen O. (1980) Biotransformation of xenobiotics in the fetus. Pharmac. Ther. 10, 261-281.
Schmid B. P., Hauser R. E. and Donatsch. P. (1985) Effects of cyproheptadine on the rat yolk sac membrane and embyronic development in vitro. Xenobiotica 15, 695~99.
Terlouw G. D. C., Moyle J. and Bechter R. (1989) The effects of time and duration of exposure to QA 208-199 on embryotoxicity, metabolism and accumulation in vitro. Experientia 45, A67.
Yang H.-Y. L., Namkung M. J. and Juchau M. R. (1988) Cytochrome P-450-dependent biotransformation of a series of phenoxazone ethers in the rat conceptus during early organogenesis: Evidence for multiple P-450 iso- enzymes. Molec. Pharmac. 34, 67-73.


















