
Viral DNA-Synthesizing Intermediate ComplexIsolated During ...
7
JOURNAL OF VIROLOGY, Aug. 1976, p. 409415 Copyright © 1976 American Society for Microbiology Vol. 19, No. 2 Printed in U.S.A. Viral DNA-Synthesizing Intermediate Complex Isolated During Assembly of Bacteriophage bX174 HISAO FUJISAWAI AND MASAKI HAYASHI* Department of Biology, University of California, San Diego, La Jolla, California 92093 Received for publication 21 October 1975 A DNA protein complex that is a precursor of mature 4X174 phage was isolated. The complex sedimented with an S value of 50 in a sucrose gradient and contained phage DNA consisting of a replicative form molecule with an ex- tended tail of single-stranded viral DNA. The viral-strand DNA ranged from one to two genomes in length. Proteins coded on the OX174 genome as well as on the host genome were associated with the viral DNA in the 50S precursor complex. Our results indicated that both viral DNA synthesis and cleavage of the growing viral-strand DNA occurred in the 50S complex. Three different modes of phage-specific DNA synthesis have been identified in Escherichia coli cells infected with the circular, single- stranded DNA bacteriophage FX174 (for re- view, see reference 29). Immediately after its penetration into the host cell, the circular, sin- gle-stranded DNA is converted to a double- stranded form (parental replicative form [RF]). This synthesis of the complementary strand is catalyzed by host enzymes (4, 31, 32). The pa- rental RF replicates semiconservatively to pro- duce a number of progeny RF molecules. Phage gene A has been shown to be necessary for progeny RF synthesis (8, 14, 16, 32). Some of the host gene products required in this process have been identified (2, 3, 7, 12, 18, 23). Finally, single-stranded viral DNA is synthesized from RF molecules. Phage-coded proteins B, C, D, F, and G (6, 16, 20) are required for this synthesis. Gene H mutants produce aberrant short pieces of viral DNA (16, 28). In the accompanying paper (9), we show that gene A product is also required during this stage. The participation of several host proteins in this process has been indicated (3, 7, 12, 26). A particularly interesting feature of FX174 morphogenesis is that production of intact sin- gle-stranded viral DNA depends upon the func- tions of seven phage-coded genes, which include genes specifying virion structural proteins F, G, and H and nonvirion protein genes A, B, C, and D. The only known phage gene that is not involved in the maturation process is gene E (lysis) (15, 19). Previously, we analyzed protein structures that are precursors of mature phage (27, 33). We showed that F protein is found as a 9S I Permanent address: Department of Botany, Kyoto Uni- versity, Kyoto, Japan. aggregate, a pentamer of F protein, and G pro- tein is found as a 6S aggregate, also a pentamer of G protein. In the presence of functional gene B, the 9S and 6S structures aggregate into a 12S complex, presumably the structural unit of one of the 12 vertexes of the icosahedral virion. The gene B protein is not found in mature phage and may function catalytically. The 12S structures are then assembled into the protein shell of the virion. However, concomitant sin- gle-stranded DNA synthesis is required during the 4X174 assembly process, implying that vir- ion assembly or encapsulation is intimately coupled with viral DNA synthesis. Synthesis of single-stranded DNA involves a rolling-circle intermediate, which consists of an RF molecule with an extended tail of single- stranded viral DNA (5, 11, 17, 30). This inter- mediate form of DNA sediments as a heteroge- neous band in sucrose gradients between 16S and 25S, depending upon the length of the sin- gle-stranded tail. The coupling between single- stranded DNA synthesis and phage assembly suggests the interaction of phage (and host) proteins with the rolling-circle intermediate DNA-protein complex as an obligatory step in the production of a mature virion. In this paper we have identified a precursor of phage particles that has an S value of 50 in sucrose gradients (50S complex). The 50S com- plex contained rolling-circle, replicating inter- mediate DNA with single-stranded tails of viral DNA. Protein analysis indicated the presence of several phage-coded proteins as well as some host proteins in the complex. Kinetic analysis of the flow of a radioactive precursor of DNA into RF, the 50S complex, and mature phage indicated that viral DNA synthesis took place in the 50S complex. Our 409
Transcript of Viral DNA-Synthesizing Intermediate ComplexIsolated During ...

JOURNAL OF VIROLOGY, Aug. 1976, p. 409415Copyright © 1976 American Society for Microbiology
Vol. 19, No. 2Printed in U.S.A.
Viral DNA-Synthesizing Intermediate Complex IsolatedDuring Assembly of Bacteriophage bX174
HISAO FUJISAWAI AND MASAKI HAYASHI*Department ofBiology, University of California, San Diego, La Jolla, California 92093
Received for publication 21 October 1975
A DNA protein complex that is a precursor of mature 4X174 phage wasisolated. The complex sedimented with an S value of 50 in a sucrose gradient andcontained phage DNA consisting of a replicative form molecule with an ex-tended tail of single-stranded viral DNA. The viral-strand DNA ranged fromone to two genomes in length. Proteins coded on the OX174 genome as well as onthe host genome were associated with the viral DNA in the 50S precursorcomplex. Our results indicated that both viral DNA synthesis and cleavage ofthe growing viral-strand DNA occurred in the 50S complex.
Three different modes of phage-specific DNAsynthesis have been identified in Escherichiacoli cells infected with the circular, single-stranded DNA bacteriophage FX174 (for re-view, see reference 29). Immediately after itspenetration into the host cell, the circular, sin-gle-stranded DNA is converted to a double-stranded form (parental replicative form [RF]).This synthesis of the complementary strand iscatalyzed by host enzymes (4, 31, 32). The pa-rental RF replicates semiconservatively to pro-duce a number of progeny RF molecules. Phagegene A has been shown to be necessary forprogeny RF synthesis (8, 14, 16, 32). Some ofthe host gene products required in this processhave been identified (2, 3, 7, 12, 18, 23). Finally,single-stranded viral DNA is synthesized fromRF molecules. Phage-coded proteins B, C, D, F,and G (6, 16, 20) are required for this synthesis.Gene H mutants produce aberrant short piecesof viral DNA (16, 28). In the accompanyingpaper (9), we show that gene A product is alsorequired during this stage. The participation ofseveral host proteins in this process has beenindicated (3, 7, 12, 26).A particularly interesting feature of FX174
morphogenesis is that production of intact sin-gle-stranded viral DNA depends upon the func-tions ofseven phage-coded genes, which includegenes specifying virion structural proteins F,G, and H and nonvirion protein genes A, B, C,and D. The only known phage gene that is notinvolved in the maturation process is gene E(lysis) (15, 19).
Previously, we analyzed protein structuresthat are precursors of mature phage (27, 33).We showed that F protein is found as a 9S
I Permanent address: Department of Botany, Kyoto Uni-versity, Kyoto, Japan.
aggregate, a pentamer of F protein, and G pro-tein is found as a 6S aggregate, also a pentamerof G protein. In the presence of functional geneB, the 9S and 6S structures aggregate into a12S complex, presumably the structural unit ofone of the 12 vertexes of the icosahedral virion.The gene B protein is not found in maturephage and may function catalytically. The 12Sstructures are then assembled into the proteinshell of the virion. However, concomitant sin-gle-stranded DNA synthesis is required duringthe 4X174 assembly process, implying that vir-ion assembly or encapsulation is intimatelycoupled with viral DNA synthesis.
Synthesis of single-stranded DNA involves arolling-circle intermediate, which consists ofanRF molecule with an extended tail of single-stranded viral DNA (5, 11, 17, 30). This inter-mediate form of DNA sediments as a heteroge-neous band in sucrose gradients between 16Sand 25S, depending upon the length of the sin-gle-stranded tail. The coupling between single-stranded DNA synthesis and phage assemblysuggests the interaction of phage (and host)proteins with the rolling-circle intermediateDNA-protein complex as an obligatory step inthe production of a mature virion.
In this paper we have identified a precursorof phage particles that has an S value of 50 insucrose gradients (50S complex). The 50S com-plex contained rolling-circle, replicating inter-mediate DNA with single-stranded tails of viralDNA. Protein analysis indicated the presenceof several phage-coded proteins as well as somehost proteins in the complex.
Kinetic analysis of the flow of a radioactiveprecursor of DNA into RF, the 50S complex,and mature phage indicated that viral DNAsynthesis took place in the 50S complex. Our
409

410 FUJISAWA AND HAYASHI
results also suggested that cleavage of thegrowing viral-strand DNA occurred in the 50Scomplex and that the complex must contain anendonuclease activity responsible for the site-specific cleavage. We identified this endonucle-ase as the gene A product in the accompanyingpaper (9).
MATERIALS AND METHODS
Bacteria and bacteriophages. E. coli HF4704 andCR61.3 were used as nonpermissive and permissivehosts for amber mutants, respectively. Nll is anamber mutant of the lysis gene E and was used aswild-type phage.Media and buffer. HF complete medium and HF
basal medium were described by Gelfand and Ha-yashi (10). For experiments involving labeling with'Hithymidine, the thymidine content of HF com-
plete medium was decreased to 0.5 ,ug/ml. EDTAbuffer was 0.1 M NaCl-10 mM phosphate buffer-5mM EDTA (pH 7.4).
Preparation of the infected cell extracts. E. coliHF4704 was grown to a density of 5 x 108 cells/ml inHF complete medium at 37°C. Cells were treatedwith mitomycin C (50 ,ug/ml) for 20 min in the darkwithout aeration to reduce host DNA synthesis (19).After mitomycin treatment the cells were sedi-mented, washed once with HF basal medium, andsuspended in 0.1 volume of HF basal medium. Cellswere then infected with phage at a multiplicity ofinfection of 7. After a 10-min adsorption period at37°C, prewarmed HF complete medium (37°C) con-taining 0.5 ,tg of thymidine per ml was added tobegin the infection. Infected cells were kept at 37°C.At the desired time, the infection was stopped bypouring the culture over 0.5 volume of frozen HFbasal medium containing 10 mM NaCN. The cellswere sedimented, suspended in 0.1 volume of EDTAbuffer containing 100 ,ug of lysozyme per ml, andincubated for 30 min at 0°C. A few drops of chloro-form and MgCl2 (final concentration, 5 mM) wereadded to the incubation mixture. After 1 min ofwarming to 37°C, the mixture was allowed to standfor 30 min at 0°C. Debris was removed from theextracts by centrifuging the mixture at 12,000 rpmfor 20 min at 4°C.
Centrifugation. The extracts were centrifugedthrough a 5 to 30% linear sucrose gradient in EDTAbuffer with a 55% CsCl bed at the bottom in an SW41rotor (at 39,000 rpm) for 150 min at 5°C. Fractionswere collected into polypropylene tubes. Aliquots ofsamples were spotted onto filter paper, and radioac-tivity was determined after the aliquots werewashed with cold 5% trichloroacetic acid-95%ethanol (28).
For further analysis of [3H]DNA the collectedfractions were treated with 0.2% sodium dodecylsulfate at 65°C for 15 min and incubated with 1 mg ofPronase per ml overnight at 30°C.
For neutral sedimentation analysis, [3H]DNAwas centrifuged through a 5 to 20% linear sucrosegradient in 5 mM EDTA-3 mM NaOH (pH 10.0) (28)in an SW50.1 rotor at 49,000 rpm for 180 or 200 minat 20°C and, for alkaline sedimentation analysis,
through a 5 to 20% linear sucrose gradient in 0.2 MNaOH-5 mM EDTA in an SW50.1 rotor at 49,000rpm for 210 min at 20°C. A mixture of['4C]thymidine-labeled RFI and RFII was added asmarkers. RFII was prepared from RFI by introduc-ing a single-stranded break with pancreatic DNase(13). After centrifugation, fractions were collected inscintillation vials or in polypropylene tubes. Radio-activity was then determined by mixing with TritonX-100 scintillation fluid as described by Noll (24).
SI endonuclease digestion. S1 endonuclease iswell characterized as an endonuclease specific forsingle-stranded DNA (1). S1 nuclease digestion wasperformed as follows. Appropriate fractions fromneutral sucrose gradients were mixed with an equalvolume of enzyme solution containing 2 x 10:1 U ofS1 endonuclease per ml in 0.1 M sodium acetate (pH5.0) and incubated at 37°C for 45 min. Sedimentationanalysis of S1 nuclease-treated [3H]DNA was per-formed as described above.
Acrylamide gel electrophoresis. Acrylamide gelelectrophoresis of proteins was performed accordingto method 2 of Siden and Hayashi (27).
Materials. Pronase and S1 endonuclease werepurchased from Calbiochem. Radioactive materialswere from New England Nuclear Corp.
RESULTS
Identification of the 50S complex as an in-termediate of phage assembly. Phage-infectedcells were incubated at 37°C for 10 min; afterthis time most of the DNA synthesized is viralstrand (data not shown). A 1-min pulse of[3H]thymidine was then administered to theculture, followed by a 4-min chase with coldthymidine. Samples were withdrawn from theculture at the end of the pulse period and afterthe 4-min chase period. Sucrose density gra-dient profiles of these lysates are shown in Fig.1. Immediately after the pulse-labeling (Fig.la), [:Hithymidine was incorporated into mate-rial with sedimentation values of 132S, 114S,50S, 30S, and 22S. The 132S peak containedinfectivity. When the 132S particle was labeledwith radioactive amino acids and analyzed bysodium dodecyl sulfate-polyacrylamide gel elec-trophresis, it was found to contain a proteinthat co-migrated with D protein, in addition toall the proteins found in mature phage parti-cles. This particle is probably the same as the140S particle reported by Weisbeek and Sins-heimer (34). The 114S particle was judged to bemature phage by its infectivity, protein compo-sition, and DNA structure. The deproteinizedmaterials from the 30S and 22S peaks sedi-mented at the positions of RFI (supercoiledclosed circular RF) and RFII (open circular RFwith at least one nick or gap in either strand)markers, respectively, when analyzed by neu-tral sucrose gradient centrifugation (see Mate-rials and Methods). Upon chase, counts incor-
J. VIROL.

DNA-SYNTHESIZING INTERMEDIATE OF 4X174 411
in
6x
0
fraction no.
FIG. 1. Sedimentation profile of [3H]DNA in'X1 74-infected cell extracts. Mitomycin C-treated
cells were infected with Nil at 37°C at a multiplicityof infection of 7. [3H]thymidine was added to thecultures at 10 min and chased at 11 min after infec-tion by adding unlabeled thymidine (200 Mg/ml) andthymine (150 pg/ml). Samples were taken at 11 min(a) and 15 min (b) after infection. Extracts were
prepared and centrifuged as described in Materialsand Methods.
porated in the 50S and RF regions disappeared,and a simultaneous increase of radioactivity inthe phage peak occurred (Fig. lb). A slightincrease of counts in the 70S region was ob-served. Analysis of the DNA in the 70S regionshowed that the 70S particle contained single-stranded DNA shorter than mature phageDNA. Therefore, the 70S particle might repre-sent a defective particle.Nature ofDNA in the 50S complex. The 50S
complex was treated with sodium dodecyl sul-fate-Pronase, and DNA was extracted by thephenol method. Purified DNA was analyzed bysucrose gradients. When the DNA of the 50Scomplex was sedimented through a neutral su-
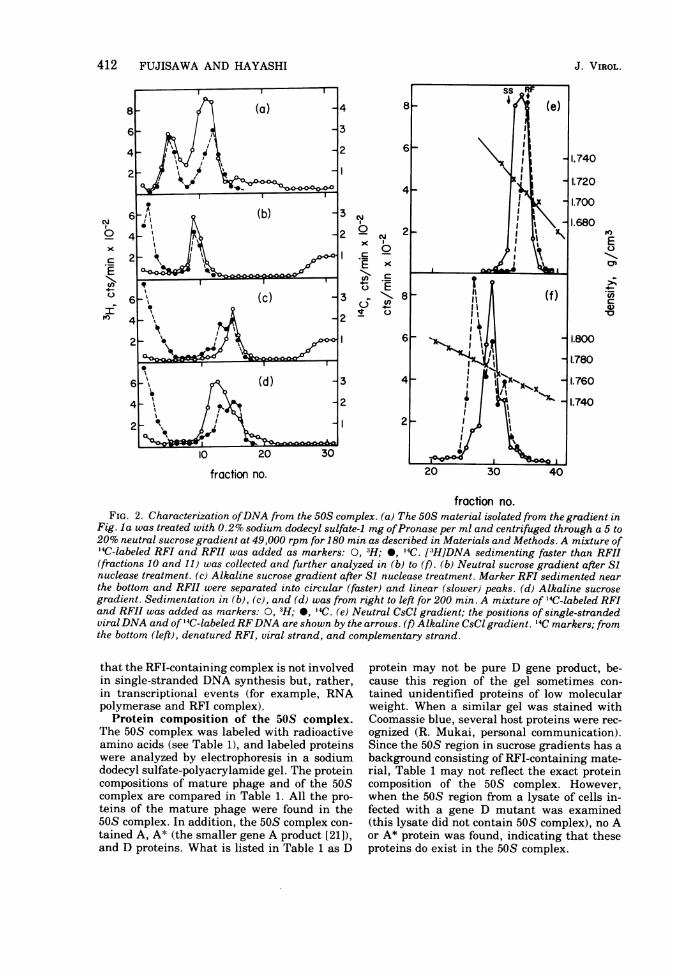
crose gradient, two peaks were observed (Fig.2a). One peak sedimented with marker RFI andthe other sedimented between markers RFI andRFII.
The DNA, isolated from the 50S complex andsedimenting between RFI and RFII (fractions10 and 11, Fig. 2a), was isolated and furtheranalyzed. (i) The DNA was treated with Siendonuclease, which is specific for single-stranded DNA. The Si-resistant structure sedi-mented with RFII with concomitant appear-ance of radioactivity on the top in a neutralsucrose gradient (Fig. 2b). (ii) When the Siendonuclease-treated sample was sedimentedthrough an alkaline sucrose gradient, the ra-dioactivity was distributed at the position cor-responding to linear phage DNA and on the topof the gradient (Fig. 2c). (iii) When the DNAwas put onto an alkaline gradient withouttreatment by Si endonuclease, the radioactiv-ity was found to sediment faster than linearphage DNA (Fig. 2d). It is to be noted that thedistribution of the radioactivity in Fig. 2d wasbroader than that in Fig. 2c, implying a moreheterogeneous population of molecules. (iv) Thedensity of the DNA was determined by equilib-rium centrifugation in neutral CsCl (Fig. 2e).Its density was found to be between double-stranded DNA (mixture of RFI and RFII; den-sity, 1.708 g/cm3) and single-stranded DNA(density, 1.724 g/cm3). When the DNA wasanalyzed by equilibrium centrifugation in alka-line cesium chloride, the radioactivity bandedmainly at the density of viral-strand DNA(tube no. 29, Fig. 2f0. A small amount of countswas found at the fractions corresponding tocomplementary strand DNA (tube no. 32, Fig.2f) and denatured RFI DNA (tube no. 26).
These experiments indicated that the DNAin the 50S complex consisted of an RFII mole-cule with an elongated tail of viral-strandDNA. The accompanying paper showed thatthe total length of the viral strand was from oneto two genomes (9).The amount of DNA isolated from the 50S
complex, which sedimented at the RFI position,differed from preparation to preparation. Whenthe DNA in each fraction of the gradient in Fig.la was analyzed, all fractions from tubes 20 to30 contained RFI. The RFI was distributedthroughout this region of the gradient in abroad band, and no significant peak containingRFI was observed. Similar preparations de-rived from B, D, F, and G mutant-infected cells(these mutants do not synthesize single-stranded DNA) were analyzed. Although thesepreparations contained neither the 50S com-plex nor the RFII molecule with single-stranded tail in 50S region, all ofthem did havethe same broad distribution of RFI found inwild-type, phage-infected cells (Fujisawa andHayashi, submitted for publication). Fromthese observations we tentatively concluded
VOL. 19, 1976
412 FUJISAWA AND HAYASHI
N
0x
cJ
"IcL00
r')
N
x b0
0
j x.)
W.'u
fraction no.
1,740
1.720
1.700
1.680'I,
E
CP
-V
1.800
1.780
1.760
1.740
fraction no.FIG. 2. Characterization ofDNA from the 50S complex. (a) The 50S material isolated from the gradient in
Fig. la was treated with 0.2% sodium dodecyl sulfate-1 mg ofPronase per ml and centrifuged through a 5 to20% neutral sucrose gradient at 49,000 rpm for 180 min as described in Materials and Methods. A mixture of"IC-labeled RFI and RFII was added as markers: 0, 3H; 0, 14C. [3H]DNA sedimenting faster than RFII(fractions 10 and 11) was collected and further analyzed in (b) to (t). (b) Neutral sucrose gradient after Sinuclease treatment. (c) Alkaline sucrose gradient after Si nuclease treatment. Marker RFI sedimented nearthe bottom and RFII were separated into circular (faster) and linear (slower) peaks. (d) Alkaline sucrosegradient. Sedimentation in (b), (c), and (d) was from right to left for 200 min. A mixture of 14C-labeled RFIand RFII was added as markers: 0, H; *, '4C. (e) Neutral CsCl gradient; the positions of single-strandedviral DNA and of 14C-labeled RFDNA are shown by the arrows. (f) Alkaline CsCl gradient. 14C markers; fromthe bottom (left), denatured RFI, viral strand, and complementary strand.
that the RFI-containing complex is not involvedin single-stranded DNA synthesis but, rather,in transcriptional events (for example, RNApolymerase and RFI complex).
Protein composition of the 50S complex.The 50S complex was labeled with radioactiveamino acids (see Table 1), and labeled proteinswere analyzed by electrophoresis in a sodiumdodecyl sulfate-polyacrylamide gel. The proteincompositions of mature phage and of the 50Scomplex are compared in Table 1. All the pro-teins of the mature phage were found in the50S complex. In addition, the 50S complex con-tained A, A* (the smaller gene A product [21]),and D proteins. What is listed in Table 1 as D
protein may not be pure D gene product, be-cause this region of the gel sometimes con-tained unidentified proteins of low molecularweight. When a similar gel was stained withCoomassie blue, several host proteins were rec-ognized (R. Mukai, personal communication).Since the 50S region in sucrose gradients has abackground consisting of RFI-containing mate-rial, Table 1 may not reflect the exact proteincomposition of the 50S complex. However,when the 50S region from a lysate of cells in-fected with a gene D mutant was examined(this lysate did not contain 50S complex), no Aor A* protein was found, indicating that theseproteins do exist in the 50S complex.
J. VIROL.

DNA-SYNTHESIZING INTERMEDIATE OF 4X174 413
TABLZ 1. Protein compositions ofphage and the 50ScompleXa
Protein Phage° 508 complexA 0.09F 1.00 1.00H 0.17 0.11A*d 0.01 0.10G 0.38 0.38D 0.97xc 0.19 0.31
a Distribution of radioactivity is expressed in rel-ative amounts, with the radioactivity of F beingtaken as 1.00.
b Radioactive amino acids used in this experimentwere [3H]leucine, [3H]lysine, and [3H]arginine (50,uCi/ml, 0.5 /hg/ml). Infected cells were labeled be-tween 10 and 20 min after infection. Cell extractswere prepared and centrifuged in a sucrose gra-dient. The mature phage fraction and the 50S frac-tions were pooled separately and analyzed by so-dium dodecyl sulfate-polyacrylamide gel electropho-resis as detailed in Materials and Methods. Gelswere sliced and counted in a scintillation counter asdescribed by Siden and Hayashi (27).
c A small-molecular-weight phage structural pro-tein (33).
d Our phage preparation, after extensive purifica-tion by sucrose gradient and CsCl density gradientcentrifugations, contained a protein co-migratingwith A* protein in sodium dodecyl sulfate-polyacryl-amide gel electrophoresis. This protein was tenta-tively assigned as A* (22).
Kinetic analysis of complex formation.Analyses of the DNA structure of the 50S com-plex suggested that elongation of viral DNAoccurred in the 50S complex (Fig. 2). Pulse andpulse-chase experiments recorded in Fig. 1 im-plied that RFII was a precursor of maturephage DNA, presumably via the 50S complex.To elucidate the relationship among RFII, the50S complex, and mature phage in the phagemorphogenetic pathway, a kinetic labeling ex-periment was performed (Fig. 3). Cells wereinfected at 30°C and [3Hlthymidine was addedto the culture 35 min after infection. At varioustimes after addition of radioactive thymidine,portions of the culture were withdrawn andlysates were prepared and analyzed by sucrosegradients. Counts incorporated into RFII, the50S complex, and mature phage were deter-mined from the sucrose gradient profiles. Ra-dioactivity appeared in RFII and the 50Scomplex immediately after addition of[3Hlthymidine and reached a plateau level in 1min. The maintenance of constant levels of ra-dioactivity in RFII and the 50S complex indi-cates that their respective pool sizes remainconstant. A lag period of 45 s was observedbefore radioactivity appeared in phage parti-
cles. Incorporation of radioactivity into phagethen continued to increase.
DISCUSSION
The 50S complex contained a DNA structureconsisting of an RF molecule with an extendedtail of single-stranded viral DNA. Such a struc-tural form ofDNA has been described by others(5, 11, 17, 30), and it is postulated to be anintermediate in the synthesis of OX174 single-stranded DNA by the rolling-circle model. Wefound that this intermediate in extracts pre-pared from wild-type, phage-infected cells sedi-mented with an S value of 50. When deprotein-ized, the DNA component of the 50S complexsedimented between RFI (21S) and RFII (16S),implying that the tailed intermediate DNA wasassociated with proteins. Analysis of the pro-tein components of the 50S complex indicated(Table 1) the presence of phage structural pro-
!Tx
.E x
.Eux-
tire, sec.
FIG. 3. Kinetic analysis of formation of phageparticles, the 50S complex, and RFII DNA in Nll-infected cells. Mitomycin-treated cells were infectedwith Nll at 30°C and labeled with [3H]thymidine at35 min after infection. Samples were taken at varioustimes after addition of [3H]thymidine, and extractswere prepared and centrifuged through neutral su-crose gradients as described in the legend to Fig. 1.Radioactive counts in the RFII, 50S, and phage re-gions were plotted as a function oftime after additionofPHithymidine.
VOL. 19, 1976

414 FUJISAWA AND HAYASHI
teins (F, G, H, X, and A*?) as well as nonstruc-tural phage proteins (A and D) and host pro-teins.
Pulse and pulse-chase experiments (Fig. 1)and kinetics of IPH]thymidine incorporation(Fig. 3) suggest that the 50S complex describedin this paper is a precursor of mature phage.RFII DNA would be a precursor of the DNA inthe 50S complex. Analysis of DNA in the 50Scomplex indicated that the elongation of viral-strand DNA took place in the complex. Theviral strand found in the 50S complex rangedfrom one to two genomes in length (9). Thisindicates that cleavage of unit length pieces ofsingle-stranded viral DNA occurs in the 50Scomplex.A possible model for viral DNA synthesis can
be envisaged from these observations. The 50Scomplex might be formed by the aggregation ofRFII with a capsomeric structure. Viral DNAsynthesis would occur and the growing single-stranded tail ofDNA would be packaged. Whenone round of DNA synthesis neared completion,cleavage would occur on the viral strand, re-sulting in the separation of RFII and a phageprecursor particle. The latter would be proc-essed into mature phage. The RFII releasedafter cleavage would enter the pool of RFII, andformation of the 50S complex would be re-peated. Alternatively, a 12S structure, whichconsists of five molecules each of F and G pro-teins, might aggregate with the growing DNAtail of an RFII molecule, forming the 50S com-plex. As viral DNA synthesis continued, more12S structures would aggregate on the elongat-ing viral DNA. When 12 of the 12S structureshave aggregated on the viral DNA, cleavagecould occur with the release of RFII and aphage precursor particle. Although we do nothave any evidence to distinguish between thesepossibilities, the fact that the 50S complex sedi-mented in a fairly homogenous manner wouldsupport the first possibility. However, bothmodels are contradictory to the fact that radio-active counts of [L'Hthymidine appeared inRFII and the 50S complex simultaneously (Fig.3). The apparent contradiction could be ex-plained by assuming that a small tail of viral-strand DNA must be synthesized on the RFIIbefore the RFII can aggregate with viral struc-tural proteins. It is also possible that the 50Scomplex, during early stages of its formation, isunstable and dissociates during the isolationprocedure, thus releasing RFII molecules withshort tails of single-stranded viral DNA. In thisrespect, Scheckman et al. (25) have reportedthat about 50% of RFII in infected cells haveshort, single-stranded tails.
ACKNOWLEDGMENTSWe are grateful to F. Fujimura for invaluable help
with the manuscript.This work was supported by grants from the National
Science Foundation (GB29170X), and the U.S. PublicHealth Service, National Institute of General Medical Sci-ences (GM12934).
LITERATURE CITED
1. Ando, T. 1966. A nuclease specific for heat-denaturedDNA isolated from a product of aspergillus oryzae.Biochim. Biophys. Acta 114:158-168.
2. Denhardt, D. T., D. H. Dressler, and A. Hathaway.1967. The abortive replication of OX-174 DNA in re-combination-deficient mutant of Escherichia coli.Proc. Natl. Acad. Sci. U.S.A. 57:813-820.
3. Denhardt, D. T., M. Iwaya, G. McFadden, and G.Schochetman. 1973. The mechanism of replication ofSX-174 single-stranded DNA. VI. Requirement forribonucleoside triphosphates and DNA polymeraseIII. Can. J. Biochem. 51:1588-1597.
4. Denhardt, D. T., and R. L. Sinsheimer. 1965. The proc-ess of infection with bacteriophage OX-174. IV. Repli-cation of the viral DNA in a synchronized infection.J. Mol. Biol. 12:647-662.
5. Dressler, D. H., and D. T. Denhardt. 1968. On themechanisms of OX single-stranded DNA synthesis.Nature (London) 219:346-351.
6. Dowell, C. E., and R. L. Sinsheimer. 1966. The processof infection with bacteriophage OX-174. IX. Studieson the physiology of three 1X-174 temperature sensi-tive mutants. J. Mol. Biol. 16:374-386.
7. Dumas, L. B., and C. A. Miller. 1974. Inhibition ofbacteriophage 4'X-174 DNA replication in dnaBmutants ofEscherichia coli C. J. Virol. 14:1369-1379.
8. Francke, B., and D. S. Ray. 1971. Cis-limited action ofthe gene-A product of bacteriophage OX-174. Proc.Natl. Acad. Sci. U.S.A. 69:475479.
9. Fujisawa, H., and M. Hayashi. 1976. Gene A product ofOX 174 is required for site-specific endonucleolyticcleavage during single-stranded DNA synthesis invivo. J. Virol. 19:416-424.
10. Gelfand, D. H., and M. Hayashi. 1969. Electrophoreticcharacterization of OX174 specific proteins. J. Mol.Biol. 44:501-516.
11. Gilbert, W., and D. Dressler. 1968. DNA replication:the rolling circle model. Cold Spring Harbor Symp.Quant. Biol. 33:483-484.
12. Greenlee, L. L. 1973. Replication of bacteriophage(X 174 in a mutant ofEscherichia coli defective in thednaE gene. Proc. Natl. Acad. Sci. U.S.A. 70-.1757-1760.
13. Hayashi, Y., and M. Hayashi. 1971. Template activitiesof the SX174 replicative allomorphic deoxyribonu-cleic acid. Biochemistry 10:4212-4218.
14. Henry, T. J., and R. Knippers. 1974. Isolation and func-tion of the gene A initiator ofbacteriophage 4X174, ahighly specific DNA endonuclease. Proc. Natl. Acad.Sci. U.S.A. 71:1549-1553.
15. Hutchison, C. A., III, and R. L. Sinsheimer. 1966. Theprocess of infection with bacteriophage FX174. X.Mutations in a (X lysis gene. J. Mol. Biol. 18:429-447.
16. Iwaya, M., and D. T. Denhardt. 1971. The mechanismof replication of 4DX174 single-stranded DNA. II. Therole of viral proteins. J. Mol. Biol. 57:159-175.
17. Knippers, R., A. Razin, R. Davis, and R. L. Sinshei-mer. 1969. The process of infection with bacterio-phage (X174. XXIX. In vivo studies of the synthesisof the single-stranded DNA. J. Mol. Biol. 45:237-263.
18. Kranias, E. G., and L. B. Dumas. 1974. Replication of
J. VIROL.

DNA-SYNTHESIZING INTERMEDIATE OF 4X174 415
bacteriophage OX174 DNA in a temperature-sensi-tive dnaC mutant of Escherichia coli V. J. Virol.13:146-157.
19. Lindqvist, B. H., and R. L. Sinsheimer. 1967. Process ofinfection with bacteriophage (X174. XIV. Studies onmacromolecular synthesis during infection with alysis-defective mutant. J. Mol. Biol. 28:87-94.
20. Lindqvist, B. H., and R. L. Sinsheimer. 1967. Theprocess of infection with bacteriophage 1'X174. XV.Bacteriophage DNA synthesis in abortive infectionswith a set of conditional lethal mutants. J. Mol. Biol.30:69-80.
21. Linney, E. A., and M. Hayashi. 1973. Two proteins ofgene A of 4X174. Nature (London) New Biol. 245:6-8.
22. Linney, E. A., and M. Hayashi. 1974. Intragenic regu-lation of the synthesis of d1X174 gene A proteins.Nature (London) 249:.345-348.
23. McFadden, G., and D. T. Denhardt. 1974. Mechanismof replication of SX-174 single-stranded DNA. IX.Requirement for the Escherichia coli dnaG protein. J.Virol. 14:1070-1075.
24. Noll, H. 1969. Polysomes: analysis of structure andfunction, p. 101-181. In J. Sargent and P. Campbell(ed.), Techniques in protein biosynthesis, vol. 2. Aca-demic Press Inc., New York.
25. Scheckman, R. W., M. Iwaya, K. Bromstrup, and D. T.Denhardt. 1971. The mechanism of replication ofOX-174 single-stranded DNA. Ifl. An enzymatic study ofthe structure of the replicative form II DNA. J. Mol.Biol. 57:177-199.
26. Schekman, R. N., and D. S. Ray. 1971. Polynucleotideligase and ODX-174 single-strand synthesis. Nature(London) New Biol. 231:170-173.
27. Siden, E. J., and M. Hayashi. 1974. Role of the gene Bproduct in bacteriophage OX-174 development. J.Mol. Biol. 89:1-16.
28. Siegel, J. E. D., and M. Hayashi. 1969. OX-174 bacterio-phage mutants which affect deoxyribonucleic acidsynthesis. Virology 4:400-407.
29. Sinsheimer, R. L. 1968. Bacteriophage OX-174 andrelated viruses. Prog. Nucleic Acid Res. Mol. Biol.8:115-168.
30. Sinsheimer, R. L., R. Knippers, and T. Komano. 1968.Stages in the replication of bacteriophage OX-174DNA in vivo. Cold Spring Harbor Symp. Quant. Biol.33:443-448.
31. Sinsheimer, R. L., B. Starman, C. Nagler, and S. Guth-rie. 1962. The process of infection with bacteriophage4X-174. I. Evidence for a "replicative form." J. Mol.Biol. 4:142-160.
32. Tessman, E. S. 1966. Mutants of bacteriophage S13blocked in infectious DNA synthesis. J. Mol. Biol.17:218-236.
33. Tonegawa, S., and M. Hayashi. 1970. Intermediates inthe assembly of OX-174. J. Mol. Biol. 48:219-242.
34. Weisbeek, P. J., and R. L. Sinsheimer. 1974. A DNA-protein complex involved in bacteriophage O1X-174particle formation. Proc. Natl. Acad. Sci. U.S.A.71:3054-3058.
VOL. 19, 1976


















