
Variability in the Degree of Expression of Phosphorylated...
12
Featured Article Variability in the Degree of Expression of Phosphorylated IB in Chronic Lymphocytic Leukemia Cases With Nodal Involvement Antonia Rodrı ´guez, 1 Nerea Martı ´nez, 1 Francisca I. Camacho, 1 Elena Ruı ´z-Ballesteros, 2 Patrocinio Algara, 2 Juan-Fernando Garcı ´a, 1 Javier Mena ´rguez, 3 Toma ´s Alvaro, 5 Manuel F. Fresno, 6 Fernando Solano, 4 Manuela Mollejo, 2 Carmen Martin, 1 and Miguel A. Piris 1 1 Molecular Pathology Program, Centro Nacional de Investigaciones Oncolo ´gicas, Madrid, Spain; 2 Department of Genetics and Pathology, Hospital Virgen de la Salud, Toledo, Spain; 3 Department of Pathology, Hospital General Universitario Gregorio Maran ˜o ´n, Madrid, Spain; 4 Department of Hematology, Hospital Nuestra Sen ˜ora del Prado, Talavera de la Reina, Toledo, Spain; 5 Department of Pathology, Hospital Verge de la Cinta, Tortosa, Spain; and 6 Department of Pathology, Hospital Central de Asturias, Oviedo, Spain ABSTRACT Purpose: Based on previous preliminary observations, we hypothesize that the molecular and clinical variability of chronic lymphocytic leukemia (CLL) reflects differences in the degree of nuclear factor (NF)-B activation, as deter- mined by the expression of phosphorylated IB (p-IB). Experimental Design: The expression profile (mRNA and protein expression) was analyzed with the Centro Na- cional de Investigaciones Oncolo ´gicas Oncochip, a cDNA microarray containing 6386 cancer-related genes, and a tis- sue microarray (TMA). The results were correlated with the IgV H mutational status, ZAP-70 expression, cytogenetic al- terations, and clinical outcome. Results: We found correlations between the presence of p-IB, a surrogate marker of NF-B activation, and changes in the expression profile (mRNA and protein ex- pression) and clinical outcome in a series of CLL cases with lymph node involvement. Activation of NF-B, as deter- mined by the expression of p-IB, was associated with the expression of a set of genes comprising key genes involved in the control of B-cell receptor signaling, signal transduction, and apoptosis, including SYK, LYN, BCL2, CCR7, BTK, PIK3CD, and others. Cases with increased expression of p-IB showed longer overall survival than cases with lower expression. A Cox regression model was derived to estimate some parameters of prognostic interest: IgV H mutational status, ZAP-70, and p-IB expression. The multivariate analysis disclosed p-IB and ZAP-70 expression as inde- pendent prognostic factors of survival. Conclusions: A variable degree of activation of NF-B, as determined by the expression of p-IB, is an identifiable event in CLL, and is correlated with changes in the expres- sion profile and overall survival. INTRODUCTION Chronic lymphocytic leukemia/small lymphocytic lym- phoma (CLL/SLL) is a lymphoproliferative disorder character- ized by the slow proliferation and accumulation of monoclonal mature CD5-positive B lymphocytes in peripheral blood, bone marrow, lymph nodes, and other tissues. Ninety percent of chronic lymphocytic leukemias are CLLs, and this is the most frequent adult leukemia in Western countries. Although CLL is a disease that is considered to be incur- able with currently available therapy, its clinical course is het- erogeneous; some patients have a more stable disease and die after many years from unrelated causes, whereas others progress very quickly and die within a few years. This variability has stimulated the search for prognostic markers with which to predict the outcome of patients and to allow treatments to be adapted to the specific risk. In addition to the Rai and Binet clinical staging systems (1, 2), specific cytogenetic [11q and 17p deletions, 12 trisomy, 13q14 deletion (3)], molecular events [IgV H mutational index (4 – 6)], and the expression level of CD38 and ZAP70 (5–11) have been found to be associated with different survival probabilities. The nuclear factor (NF)-B protein family, p105/p50, p100/p52, Rel A (p65), c-Rel, and Rel B play an important role in differentiation and survival in normal B cells and presumably also in neoplastic B cells. NF-B proteins are present in the cytoplasm bound to IB proteins, which are inhibitory mole- cules that sequester NF-B dimers in the cytoplasm. After B-cell activation and subsequent phosphorylation, ubiquitina- tion and proteasome degradation of IB, NF-B translocate to the nucleus, making possible the transcription of its target genes. NF-B activation after induction by CD40-CD154 ligation has been described in CLL cells (12) and is correlated with Received 4/19/04; revised 7/2/04; accepted 7/9/04. Grant support: This study was supported by grants from the Comu- nidad Auto ´ noma de Madrid (CAM 08.1/0011/2001.1), the Ministerio de Sanidad y Consumo (FIS 01– 0035), and the Ministerio de Ciencia y Tecnologı ´a (SAF 2001–0060), Spain. F. Camacho is supported by an Olivia Roddom grant from the Spanish Association for Cancer Research (AECC). A. Rodrı ´guez is supported by a grant from the Ministerio de Sanidad y Consumo (FIS 02–9355). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Note: Supplementary data for this article can be found at Clinical Cancer Research Online (http://clincancerres.aacrjournals.org) Requests for reprints: Miguel Angel Piris, Molecular Pathology Pro- gram, Centro Nacional de Investigaciones Oncolo ´gicas, C/Melchor Fer- na ´ndez Almagro 3, E-28029 Madrid, Spain. Phone: 34-91-224-69-00; Fax: 34-91-224-69-23; E-mail: [email protected]. ©2004 American Association for Cancer Research. 6796 Vol. 10, 6796 – 6806, October 15, 2004 Clinical Cancer Research Cancer Research. on November 7, 2018. © 2004 American Association for clincancerres.aacrjournals.org Downloaded from
Transcript of Variability in the Degree of Expression of Phosphorylated...

Featured Article
Variability in the Degree of Expression of Phosphorylated I�B� inChronic Lymphocytic Leukemia Cases With Nodal Involvement
Antonia Rodrıguez,1 Nerea Martınez,1
Francisca I. Camacho,1 Elena Ruız-Ballesteros,2
Patrocinio Algara,2 Juan-Fernando Garcıa,1
Javier Menarguez,3 Tomas Alvaro,5
Manuel F. Fresno,6 Fernando Solano,4
Manuela Mollejo,2 Carmen Martin,1 andMiguel A. Piris1
1Molecular Pathology Program, Centro Nacional de InvestigacionesOncologicas, Madrid, Spain; 2Department of Genetics and Pathology,Hospital Virgen de la Salud, Toledo, Spain; 3Department ofPathology, Hospital General Universitario Gregorio Maranon,Madrid, Spain; 4Department of Hematology, Hospital Nuestra Senoradel Prado, Talavera de la Reina, Toledo, Spain; 5Departmentof Pathology, Hospital Verge de la Cinta, Tortosa, Spain;and 6Department of Pathology, Hospital Central de Asturias,Oviedo, Spain
ABSTRACTPurpose: Based on previous preliminary observations,
we hypothesize that the molecular and clinical variability ofchronic lymphocytic leukemia (CLL) reflects differences inthe degree of nuclear factor (NF)-�B activation, as deter-mined by the expression of phosphorylated I�B� (p-I�B�).
Experimental Design: The expression profile (mRNAand protein expression) was analyzed with the Centro Na-cional de Investigaciones Oncologicas Oncochip, a cDNAmicroarray containing 6386 cancer-related genes, and a tis-sue microarray (TMA). The results were correlated with theIgVH mutational status, ZAP-70 expression, cytogenetic al-terations, and clinical outcome.
Results: We found correlations between the presence ofp-I�B�, a surrogate marker of NF-�B activation, and
changes in the expression profile (mRNA and protein ex-pression) and clinical outcome in a series of CLL cases withlymph node involvement. Activation of NF-�B, as deter-mined by the expression of p-I�B�, was associated with theexpression of a set of genes comprising key genes involved inthe control of B-cell receptor signaling, signal transduction,and apoptosis, including SYK, LYN, BCL2, CCR7, BTK,PIK3CD, and others. Cases with increased expression ofp-I�B� showed longer overall survival than cases with lowerexpression. A Cox regression model was derived to estimatesome parameters of prognostic interest: IgVH mutationalstatus, ZAP-70, and p-I�B� expression. The multivariateanalysis disclosed p-I�B� and ZAP-70 expression as inde-pendent prognostic factors of survival.
Conclusions: A variable degree of activation of NF-�B,as determined by the expression of p-I�B�, is an identifiableevent in CLL, and is correlated with changes in the expres-sion profile and overall survival.
INTRODUCTIONChronic lymphocytic leukemia/small lymphocytic lym-
phoma (CLL/SLL) is a lymphoproliferative disorder character-ized by the slow proliferation and accumulation of monoclonalmature CD5-positive B lymphocytes in peripheral blood, bonemarrow, lymph nodes, and other tissues. Ninety percent ofchronic lymphocytic leukemias are CLLs, and this is the mostfrequent adult leukemia in Western countries.
Although CLL is a disease that is considered to be incur-able with currently available therapy, its clinical course is het-erogeneous; some patients have a more stable disease and dieafter many years from unrelated causes, whereas others progressvery quickly and die within a few years. This variability hasstimulated the search for prognostic markers with which topredict the outcome of patients and to allow treatments to beadapted to the specific risk. In addition to the Rai and Binetclinical staging systems (1, 2), specific cytogenetic [11q and 17pdeletions, 12 trisomy, 13q14 deletion (3)], molecular events[IgVH mutational index (4–6)], and the expression level ofCD38 and ZAP70 (5–11) have been found to be associated withdifferent survival probabilities.
The nuclear factor (NF)-�B protein family, p105/p50,p100/p52, Rel A (p65), c-Rel, and Rel B play an important rolein differentiation and survival in normal B cells and presumablyalso in neoplastic B cells. NF-�B proteins are present in thecytoplasm bound to I�B� proteins, which are inhibitory mole-cules that sequester NF-�B dimers in the cytoplasm. AfterB-cell activation and subsequent phosphorylation, ubiquitina-tion and proteasome degradation of I�B�, NF-�B translocate tothe nucleus, making possible the transcription of its target genes.
NF-�B activation after induction by CD40-CD154 ligationhas been described in CLL cells (12) and is correlated with
Received 4/19/04; revised 7/2/04; accepted 7/9/04.Grant support: This study was supported by grants from the Comu-nidad Autonoma de Madrid (CAM 08.1/0011/2001.1), the Ministerio deSanidad y Consumo (FIS 01–0035), and the Ministerio de Ciencia yTecnologıa (SAF 2001–0060), Spain. F. Camacho is supported by anOlivia Roddom grant from the Spanish Association for Cancer Research(AECC). A. Rodrıguez is supported by a grant from the Ministerio deSanidad y Consumo (FIS 02–9355).The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.Note: Supplementary data for this article can be found at ClinicalCancer Research Online (http://clincancerres.aacrjournals.org)Requests for reprints: Miguel Angel Piris, Molecular Pathology Pro-gram, Centro Nacional de Investigaciones Oncologicas, C/Melchor Fer-nandez Almagro 3, E-28029 Madrid, Spain. Phone: 34-91-224-69-00;Fax: 34-91-224-69-23; E-mail: [email protected].
©2004 American Association for Cancer Research.
6796 Vol. 10, 6796–6806, October 15, 2004 Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from
Bcl-2 expression (13). The presence of activated NF-�B hasbeen shown to be activated by cytokines (14) and regulated byAPRIL and BAFF genes (15). Electrophoretic mobility shiftanalysis studies have confirmed the presence of NF-�B activa-tion in a subset of CLL cases and have also revealed a subset ofcases lacking this (16). Nevertheless, the real incidence of thisphenomenon and its precise features have yet to be fully eluci-dated.
Our previous preliminary observations led us to hypothe-size that the clinical variability of CLL reflects, at least partially,differences in the degree of NF-�B activation of CLL, a cardinal
feature in most lymphoid malignancies. Here we have investi-gated the correlation of the presence of phosphorylated I�B�(p-I�B�), a marker of NF-�B activation (17, 18), with changesin the expression profile and clinical outcome of CLL cases.
To achieve this, we used the Centro Nacional de Investi-gaciones Oncologicas (CNIO) Oncochip, a cDNA microarray,containing 6386 cancer-related genes, and a tissue microarray(TMA), containing 96 cores corresponding to 38 cases andcontrols. To facilitate simultaneous analysis using a cDNAmicroarray and a TMA, we restricted the analysis to CLL caseswith nodal involvement, selected for being at Rai stage � 1.
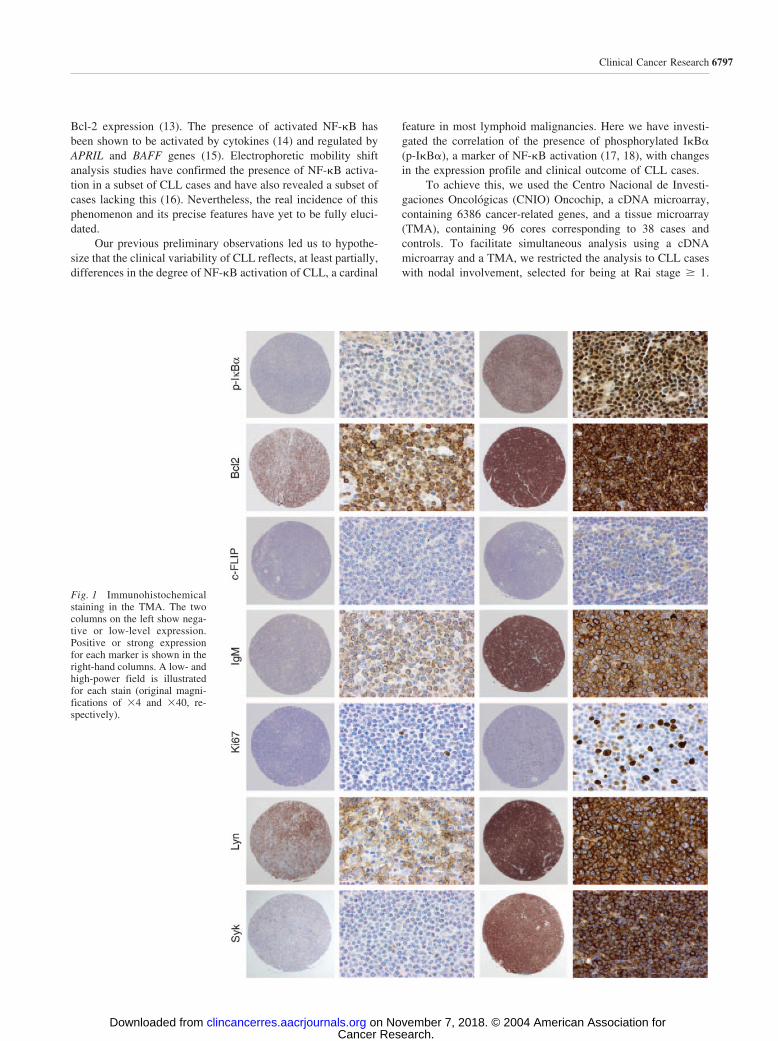
Fig. 1 Immunohistochemicalstaining in the TMA. The twocolumns on the left show nega-tive or low-level expression.Positive or strong expressionfor each marker is shown in theright-hand columns. A low- andhigh-power field is illustratedfor each stain (original magni-fications of �4 and �40, re-spectively).
6797Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

This meant that we could also use frozen, unmanipulated diag-nostic lymph nodes, the NF-�B activation status of which moreclosely reflects the in vivo status. These more advanced CLLcases are of additional interest because they require furtherclinical surveillance and early treatment and because auxiliaryprognostic variables could be identified.
MATERIALS AND METHODSPatients and Samples. Cases were selected for study
based on the availability of frozen lymph node tissue at themoment of diagnosis of CLL. Frozen samples were supplied bythe Hospital Virgen de la Salud, Toledo; Hospital GregorioMaranon, Madrid; Hospital Doce de Octubre, Madrid; HospitalCentral de Asturias, Oviedo; Hospital Verge de la Cinta, Tor-tosa; and the Tumor Bank Network of the CNIO. Diagnosis ofCLL was based on the National Cancer Institute-sponsoredWorking Group guidelines and the World Health OrganizationClassification criteria (19, 20).
Lymph node biopsies were done to assess CLL involve-ment at diagnosis. Lymph node tissue was frozen and paraffin-embedded following standard procedures. Expert pathologistsdid blind evaluations of morphology and immunohistochemistryprevious to entering into the study. Frozen reactive lymph nodesamples were used as controls.
Forty-one cases were found to be suitable for study in accord-ance with the established criteria. Clinical data were obtained frommedical records. The series comprised 25 males and 16 females.The mean age at diagnosis was 67 years, with only two patientsunder 50 (37 and 39) years of age. At the moment of diagnosis, allpatients had lymph node involvement, in addition to peripheralblood lymphocytosis corresponding to a stage �1 on the Raiscoring system. According to the Binet classification, 10 cases(24%) were stage A, 23 (56%) cases were stage B, five (12%) caseswere stage C, and three (7%) cases could not be assigned acategory. The average follow-up after diagnosis was 52 months,with a maximum of 132 months. Criteria to consider progressivedisease and to determine need for treatment were based on NCIguidelines (20). First-line treatment was Chlorambucil � Predni-sone in 19 patients, CHOP (cyclophosphamide, doxorubicin, vin-cristine, and prednisone) or CHOP-like regimen in seven patientsand nucleotide analogous-based therapy in four patients. Their
mean survival-free disease time was 30 months (12–68 months).Progression with or without treatment occurred in 28 cases, with amean and median progression-free survival of 30 and 39 months,respectively. The mean overall survival (OS) in this series was 52months (median, 46 months). This was shorter than that of otherseries, perhaps because of the disease being more advanced at thetime of its diagnosis.
In most cases, CLL was considered the main cause ofdeath, although four patients developed secondary gastrointes-tinal and breast carcinoma.
Tissue Microarray and Immunohistochemistry. ATMA block was constructed with a Tissue Arrayer device (BeecherInstruments, MD) with two 1-mm-diameter cylinders from twoselected areas of each case and from appropriate controls. Samplecores were obtained from paraffin-embedded lymph node blockscorresponding to the initial diagnosis, which were available in 38samples of the study. Paraffin sections were examined to select theinvolved area. The resulting TMA contained 96 cores from casesand controls. Antibodies against Syk (Santa Cruz, CA), Lyn (SantaCruz Biotechnology), BclII (DAKO, Copenhagen, Denmark),p-I�B� (Cell Signaling, Beverly, MA), Ki67 (DAKO), FLIP (Ab-cam, Cambridge, United Kingdom), and IgM (DAKO) were usedto determine protein expression. The immunohistochemical proto-col was carried out as described previously (21).
All hybridized TMAs were scanned with the BLISS system(Bacus Laboratories, Inc, Lombard, IL), which uses a three charge-coupled device chip for the three colors red, green, and blue sensoroptically coupled to a microscope. Tissue microarrays were quan-titatively scored on a scale of 0 to 100 with TMAscore v.1.0 imageanalysis software (Bacus Laboratories, Inc., Lombard, IL), whichuses WebSlide v.1.0 (Bacus Laboratories, Inc.) virtual slides.
ZAP-70 expression (Cell Signaling) was classified as pos-itive or negative after visual inspection and comparison with thelevel of expression of reactive T cells.
Microarray Assays. RNA isolation and amplificationfor microarray experiments were performed as described previ-ously (22). Universal Human Reference RNA (Stratagene) wasused as a reference. From each amplified RNA product, 2.5 �gwas labeled with cyanine 3-conjugated dUTP (Cy3) in the caseof the samples and with cyanine 5-conjugated dUTP (Cy5) inthe case of the reference RNA.
Hybridizations were done in the CNIO OncoChip (22). Hy-bridized slides were scanned with the Scanarray 5000 XL (GSILumonics Kanata, Ontario, Canada) or with the Agilent G2565AAMicroarray Scanner System (Agilent, Palo Alto, CA), and images
Table 1 Description of the expression of the markers analyzed inthe TMA
Median Range Visual evaluation
Syk 27.25 0.31–94.96 Strongly heterogeneous cytoplasmicexpression.
Lyn 63.05 20.46–82.61 Cytoplasmic expression, moderatevariability.
Bcl2 54.86 17.35–77.89 Cytoplasmic, strong expression.p-I�B� 2.63 0.01–65.43 Cytoplasmic and nuclear
expression, highly variable. Twomajor groups defined.
Ki67 1.79 0.02–9.3 Nuclear expression, stronger inproliferation centers.
c-Flip 0.02 0–1.24 Surface expression, low intensity.IgM 51.76 0.26–82.63 Variable cytoplasmic-surface
expression.
Table 2 Correlation between markers analyzed in the TMA
Syk Lyn Bcl2
LynPearson 0.543P �0.001
Bcl2Pearson 0.394 0.661P �0.05 �0.001
p-I�B�Pearson 0.627 0.409 0.54P �0.001 �0.05 �0.001
6798 Differences in NF-�B Activation in CLL
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from
were analyzed with the GenePix 4.0 program (Axon InstrumentsInc., Union City, CA).
To adjust for differences in the red and green labeling,Cy3/Cy5 ratios were normalized with the Diagnosis and Nor-malization of Microarray Data (DNMAD) tool based on stand-ard print-tip loess.7
To organize patterns on a symmetrical scale, ratios werelog-transformed (base 2), and duplicated spots on the OncoChip
were averaged to the median. Only genes that had data presentin �70% of cases were considered in the subsequent analysis.To obtain the expression profile of each tumor we referred theratios of the tumors to the mean of ratios of reactive lymphnodes from four different patients.
Cluster analysis was done with the Gene Cluster program(23). Student’s t test was used to identify genes differentiallyexpressed in the classes defined by p-I�B� expression.8
7 http://dnmad.bioinfo.cnio.es. 8 http://bioinfo.cnio.es/cgi-bin/tools/multest/multest.cgi.
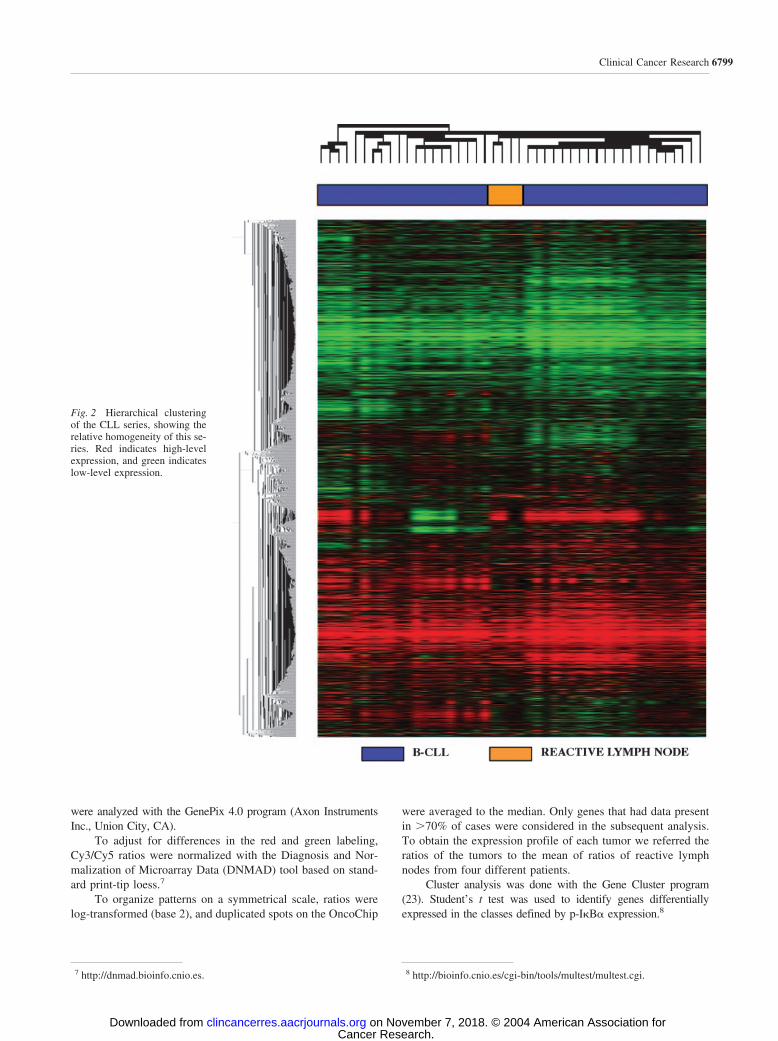
Fig. 2 Hierarchical clusteringof the CLL series, showing therelative homogeneity of this se-ries. Red indicates high-levelexpression, and green indicateslow-level expression.
6799Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

Definition of Functional Signatures. All genes printedon the CNIO Oncochip array were analyzed with the GeneOntology-based application FATIGO, which is part of theGEPAS suite (24).9 Genes that remained unclassified byFATIGO were sought in the GENECARDS database10 andbiological functions assigned. Genes were grouped in differentcategories modified in accordance with published observationsabout pathogenesis of B-cell lymphoma as NF-�B, B-cell re-ceptor (BCR) signaling and lymph node-homing categories.Functional signatures for cell growth and maintenance (806clones), apoptosis (146 clones), NF-�B pathway (167 clones),BCR signaling (54 clones), cell communication (808 clones),lymph node-homing (13 clones) and metabolism (1409 clones)were included here.
VH Gene Analysis. Rearranged IgVH genes were ampli-fied with a semi-nested PCR method following standard pub-lished procedures (25). A mixture of six framework 1 (FR1) VH
family-specific primers and two consensus primers for the JH
gene in the first round of PCR and JH internal primers in thesecond round were used.
The direct-sequencing procedure was done with an ABIPRISM 310 or 3700 Genetic Analyzer (Applied Biosystems,Weiterstadt, Germany) following the manufacturer’s procedure.Mutations were identified by comparison with the germlinesequence (immunoglobulin BLAST11 and V BASE sequencedirectories12).
A sequence was considered mutated when there was �98%homology with the closest germline VH genes.
Fluorescence In situ Hybridization Analysis. Fluores-cence in situ hybridization analysis was done with LSI p53/LSIATM and LSI D13S319/LSI 13q34/CEP 12 Multi-color ProbeSets, from Vysis (Downers Grove, IL). LSI p53 (17p13.1)covers the entire p53 gene, LSI D13S319, detects the 13q14.3region, LSI 13q34 detects a region near the subtelomere ofchromosome 13q, and CEP 12 detects the � satellite, centro-meric region of chromosome 12. LSI ATM detects the 11q22.3region of chromosome 11, which includes the entire ataxiatelangiectasia-mutated gene (ATM).
Hybridizations on the tissue microarray were done as de-scribed in Moreno-Bueno et al. (26). The cutoff value fordeletions was taken as the mean plus three times the SD of theproportion of cells with one signal in the three normal non-tumoral samples. This value corresponded to 25% of cells.Tumor samples with �25% of cells with one signal wereconsidered to be subject to genomic loss involving the targetedgenomic region. The cutoff value for trisomy was establishedwith the same three normal nontumoral samples. Gain or am-plification was defined as the presence (in �5% of tumor cells)of three distinctive signals.
Statistical Analysis. Relationships between the levels ofexpression of the different markers were explored with the
Pearson correlation. The significance of associations betweencharacteristics was determined with Fisher’s exact test.
The mean expression for each value was compared toidentify genes discriminating between groups established on thebasis of the presence or absence of a condition by Student’s ttest.13 The associated P value is presented as the adjusted Pvalue derived from procedures that control the false discoveryrate. Genes with adjusted P values using False Discovery Rate(FDR)_indep �0.2 are shown.
Cases were divided into two groups with values of p-I�B�greater than and less than the median of the series, respectively.
Cox proportional hazard models were used to evaluate thepossible associations between OS and a number of variables. OScurves were derived with the Kaplan-Meier method. Statisticalsignificance of associations between individual variables andOS was determined with the log-rank test.
Statistical analyses were done with the SPSS (SPSS Inc.,Chicago, IL) program and with tools for random permutationtests.14
RESULTSTissue Microarray Data. Captured images of expres-
sion of Syk, Lyn, Bcl2, p-I�B�, Ki67, c-FLIP, and IgM areshown in Fig. 1. Cases were divided into two groups withrespect to the median value of protein detection in the wholeseries (Table 1). For p-I�B�, this threshold indeed separates twogroups exhibiting distinct low and high levels of expression.
Levels of expression for the markers showed that p-I�B�expression was closely correlated with the level of expression ofSyk, Lyn, and Bcl2. Details of the results are given in Table 2.Other significant associations are also described in this table.
cDNA Microarray Data. Hierarchical clustering (Stan-ford University Cluster) of 41 CLL samples and four reactivelymph nodes analyzed by cDNA microarrays revealed relativehomogeneity of all of the CLL cases. Reactive lymph node caseswere grouped together (Fig. 2). The reproducibility of the tech-nique was confirmed by repeating the hybridization of foursamples and finding that parallel hybridizations clustered to-gether (supplementary information).
Two-fold over- or underexpressed clones with respect toreactive lymph nodes in at least 50% of CLL cases were con-sidered to make up the CLL signature. This includes 213 genes,comprising 114 down-regulated and 99 up-regulated genes.Genes were grouped according to functional signatures, payingparticular attention to the functional clusters of NF-�B, BCRsignaling and lymph-node homing. The results of the expressionprofiling of these gene sets are detailed in Tables 3 and 4.
Relevant up-regulated genes and proteins related withBCR- and NF-�B-signaling included LYN, BTK, CCR7, BCL2,BAFF and other genes. CCR7, a well characterized NF-�Btarget (27), is a major receptor providing the triggering signalsfor B-cell entry into lymph nodes (28), which is consistent withthe fact that the CLL cells analyzed here were derived frominvolved lymph nodes.
9 http://fatigo.bioinfo.cnio.es.10 http://bioinformatics.weizmann.ac.il/cards.11 http://www.ncbi.nlm.nih.gov/igblast.12 http://www.mrc-cpe.cam.ac.uk/vbase-ok.
13 http://bioinfo.cnio.es/cgi-bin/tools/multest/multest.cgi.14 http://bioinformatica.cnio.es/.
6800 Differences in NF-�B Activation in CLL
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

Lyn is a member of the Src-family protein tyrosine kinasesthat, along with Blk and Fyn, play an essential role in pre-B-cellreceptor-mediated NF-�B activation and B-cell development(29).
Btk has been shown to couple I�B kinase, I�B�, andNF-�B to the BCR, linking these key pathways (30); addition-ally, BTK directly regulates bcl-x expression by transcriptionalcontrol in response to BCR engagement (31).
Increased Bcl2 expression was also observed in this series,confirming previous observations (32) that linked Bcl2 expres-sion to NF-�B activation in CLL cells (33).
BAFF, a member of the tumor necrosis factor superfamilyis involved in normal and leukemic B-cell survival and differ-entiation. BAFF binds to B cells and through the B-cell co-receptor complex promotes NF-�B activation (15, 34).
Genes found to be up-regulated in the lymph node-hominggroup included CCR7 and SELL. SELL (L-selectin, CD62L) is acell surface adhesion protein that mediates the adherence oflymphocytes to endothelial cells of high endothelial venules inperipheral lymph nodes, the expression of which was found tobe increased in a subset of CLL patients (32). Interestingly, lossof L-selectin from the cell surface inhibited the migration of
activated lymphocytes to lymph nodes and resulted in theirpreferential migration to the spleen (35).
ZAP-70 Expression. ZAP-70 could be evaluated in 35cases of the complete series. Expression was positive in 20(57%) cases and negative in the remaining 15 (43%) cases. Inthe light of recent results suggesting that this expression is asurrogate of IgVH mutational status (8, 10, 36), we investigatedthis correlation in our series. We found that 15 of the 20 positivecases, but only three of the15 negative cases, belonged to theunmutated group. This association is significant (P � 0.05).
Mutational Status. DNA was obtained from 40 cases ofthe complete series. IgVH gene rearrangement was identified in36 patients. Forty-seven sequences were amplified, with doubleor triple rearrangement in seven and two cases, respectively.Only four of the nine multiple rearrangements showed differ-ences in mutational status with respect to VH family usage. Thespectrum of VH family usage was as follows: VH 1 and VH 3both in 16 cases, VH 4 in 11 cases, VH 2 in 2 cases, and VH 5and VH 6 both in 1 case. Distribution of different subfamilyusage was similar to the described, with a more frequent repre-sentation of VH1.69 (10 sequences) (37, 38). Taking a �2%variability with respect to the germline VH sequence, 18 IgVH
sequences from 17 patients were considered as mutated, and 29sequences from 19 patients had a low mutational index. Per-centages of mutated (48%) and unmutated (52%) cases werecalculated based on 36 rearrangements. All cases with VH1.69subfamily usage were unmutated.
FISH Analysis. To identify chromosomal abnormalitiesand assess their involvement in the findings in this series, we didFISH analysis on TMA cores, thereby investigating deletions in11q22 (ATM), 17p13 (p53), 13q14 regions, and 12 trisomy. Thedistribution of chromosomal abnormalities was as follows:11q22 deletion was present in five cases of 32 assessable cores(15%); only one case featured deletion of 17p13 out of 30assessable cores (3%); 13q14 deletion was found in four of 28assessable cores (14%); finally, only one of the 32 assessablecases was positive for trisomy of chromosome 12 (3%). Twocases had double chromosomal abnormalities, both with 11q22and 13q14 deletions. All cases with 11q22 deletion were fromthe high-level p-I�B� expression group.
Correlation with p-I�B�. CLL cases were divided intotwo groups according to the median value (2.63) of p-I�B�protein detection in the whole series. Employing previously
Table 3 CLL signature genes divided into functional groups
Function %Expression
ratio Genes
NF-�B 5 �2-fold 6�2-fold 6
BCR signaling 3 �2-fold 2�2-fold 4
Apoptosis 7 �2-fold 9�2-fold 6
Cell growth and maintenance 14 �2-fold 19�2-fold 11
Metabolism 28 �2-fold 36�2-fold 24
Lymph-node homing 2 �2-fold 3�2-fold 2
Cell communication 27 �2-fold 37�2-fold 21
Other or unknown function 14 �2-fold 19�2-fold 14
NOTE. Functional groups are defined by Gene Ontology and levelof deregulation (�2-fold up- or down-regulated in at least 50% ofsamples). Some genes were found in two or more groups.
Table 4 Expression-profiling genes upregulated in NF-�B and BCR groups
Gene symbol Gene description N-fold (average)
NF-�B genesBCL2 B-cell CLL/lymphoma 2 3.091CCR7 chemokine (C-C motif) receptor 7 2.715LYN v-yes-1 Yamaguchi sarcoma viral-related oncogene homolog 2.495TLE1 transducin-like enhancer of split 1, homolog of Drosophila E(sp1) 2.269IL1A interleukin 1, alpha 1.291BAFF B-cell activating factor of the TNF family 0.9
BCR genesBCL2 B-cell CLL/lymphoma 2 3.091BTK Bruton agammaglobulinemia tyrosine kinase 2.846LYN v-yes-1 Yamaguchi sarcoma viral-related oncogene homolog 2.495GAB1 GRB2-associated binding protein 1 1.967
6801Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

defined functional signatures, Student’s t test was used to iden-tify genes that were differentially expressed in these two groups.In total, 894 genes with the best adjusted P value (see Materialand Methods) were differentially expressed (Fig. 3A and B). Themost characteristic genes of the high-level p-I�B� expressiongroup are listed in Tables 5 and 6.
Among the genes to be deregulated in the cases withNF-�B activation, some of the most noticeable were PIK3CD,LYN, IRAK1, IGF1, FAF1, BAFF, RBBP2, HDAC6, CDC16,PMSCL2, PPAP2B, SELL, and MALT1.
PIK3CD codes for the p110� catalytic subunit of the phos-phoinositide 3-kinase (PI3K) family, preferentially expressed byleukocytes, which plays a role, after antigen-receptor triggeringand SYK activation, in protein kinase C-- and AKT/PKB-dependent B-cell survival (39).
IGF1 and its receptor, IGF1-R, are key regulators of cellgrowth and development. After activation, they induce a cas-cade of tyrosine phosphorylations, leading to the eventual acti-vation of Akt and up-regulation of the antiapoptotic proteinBcl-xL (40).
FAF1 is a member of the death-inducing signaling complexin Fas-mediated apoptosis and has also been shown to be able toplay a role as an NF-�B activity suppressor through cytoplasmicretention of p65, via physical interaction (41).
BAFF expression was greater in cases with NF-�B activa-tion, thereby confirming the proposed role of BAFF as a pro-moter of NF-�B activation in leukemic B-cells (15, 34).
It is of particular note that the increased expression of RBBP2and HDAC6, a binding partner for NF-�B p50 and p65 (42),associates NF-�B activation in CLL with histone acetylation.
Fig. 3 Student’s t test analysis.A, differentially expressed genesidentified in cases with high ver-sus low levels of expression ofphosphorylated I�B�. B, detailshowing the most closely asso-ciated genes with highest Pvalue within each group. Redindicates high-level expression,green indicates low-level ex-pression, and missing values arein gray.
6802 Differences in NF-�B Activation in CLL
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

In contrast, cases with a low level of NF-�B activationexpress CUL3, PTH, CARD12, TRAF3, MADH4, GAD2,DNAI1, UBE2D3, GTF2B, HEAB, POL2RG, G3BP, and ARG1.These findings suggest abnormalities in the ubiquitination andprotein degradation in this group of cases.
Associations of p-I�B� expression with VH mutationalstatus and chromosomal abnormalities were sought using Fish-er’s exact test. There was no substantial relationship between thelevel of p-I�B� expression and IgVH mutational index, althoughwe observed that high levels of p-I�B� were associated with11q22 (ATM) deletion (P � 0.05).
Survival Analysis. Using the Kaplan-Meier method toassess the prognostic importance of p-I�B� in the OS of CLL,we found that cases with a high level of expression had bettersurvival probability than cases with a low level of expression(P � 0.05) (Fig. 4). The five-year OS probability was 40 and60% in the low- and high-level p-I�B� expression groups,respectively.
Kaplan-Meier analysis of IgVH mutational status revealeda trend toward longer survival in the mutated group. No relationwas found between chromosomal abnormalities and OS in thisseries.
A Cox proportional hazards regression model was derived
to estimate a number of variables of potential prognostic inter-est, such as IgVH mutational status, ZAP-70, and p-I�B� ex-pression. This multivariate analysis showed ZAP-70 expressionand p-I�B� to be independent prognostic factors of survival(P � 0.01; Table 7).
DISCUSSIONResults from our study show that activation of NF-�B, as
determined by the expression of p-I�B�, is a relatively frequentphenomenon in CLL with lymph node involvement. The inten-sity of this expression parallels NF-�B activation and oscillatesalong the series. Additionally, the expression of p-I�B� islinked with the expression of a large set of key genes involvedin the control of B-cell signaling, signal transduction and apo-ptosis.
CLL cells are capable of responding to external cell pro-liferation signals, such as microenvironmental stimuli fosteringCD40 stimulation (43), and anti-apoptotic signals after antigen-receptor triggering. BCR complex is formed by surface IgM thatis non-covalently associated with CD79a/CD79b signal trans-duction units. Following sIg cross-linking, it activates the BCRsignaling pathway downstream. In normal B-lymphocytes, this
Fig. 3 Continued
6803Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

leads to proliferation, differentiation, apoptosis, survival, andimmune tolerance, depending on the B-cell developmental stageand cell environment. Molecules downstream of BCR includemembers of the Src kinase family (Lyn, Fyn, and Blk; ref. 29),the Tec kinase family (Btk; ref. 44), Syk tyrosine kinase (45,46), members of the protein kinase C family of serine-threoninekinases (29), PI3K; ref. 39), the mitogen-activated protein ki-nase family (45), the antiapoptotic NF-�B complex (29), andZAP-70 (47) in the case of CLL cells. Activation of B-cellco-receptor complex (CD21, CD19 and CD81) by the BAFFgene (34) and tumor necrosis factor receptor CD40-CD154interactions amplify BCR signaling (48).
This study, including the mRNA and protein expressionlevel of a majority of these genes, suggests the possible signif-icance of BCR signaling in the activation of NF-�B and high-lights the expression of a selected number of genes playing acritical role in this process, including SYK, IRAK1, and PI3K.Interestingly, this phenomenon seems to involve preferentially agroup of CLL cases, thus further increasing the biological het-erogeneity of CLL.
IRAK1 is a key TRAF6-interacting protein that mediatesNF-�B activation downstream of tumor necrosis factor andToll/IL-1R-containing receptors (49). In fact, in this CLL series,the expression of both IRAK1 and IL1 appears to be closelyrelated to that of phosphorylated I�B�.
This study also underlines the relevance of the PI3K path-way in the molecular pathogenesis of CLL cells. Although themolecular details of the recruitment, activation, and signaling ofPI3Ks by the BCR are not fully understood, it is clear that theyare at least partially regulated by BCR- and Syk-dependentphosphorylation of CD19 and other B-cell PI3K adaptor mole-cules (39). Here, the results highlight a simultaneous expressionof a large set of the key molecules in this process in NF-�Bactivated CLL cases, including SYK, IRAK1, IL1, PIK3CD, andPIK3CB. Studies at the protein level have confirmed thesefindings; these observations also extend to the expression ofBclII and Lyn.
In this study, Bcl2 protein expression level was closelyrelated with p-I�B�, Syk, and other molecules involved in theBCR signaling that leads to NF-�B activation. Indeed, the highlevel of BclII protein expression is routinely observed in thediagnosis of small lymphocytic neoplasms and may be seen inbone marrow and lymph node examinations. Thus, increasedBcl2 protein expression is a common and substantial finding inthe major types of small lymphocytic neoplasms, includingmantle cell lymphoma, follicular lymphoma, and CLL. Theroutes leading to Bcl2 expression, nevertheless, differ in distinctneoplasms (21).
Variation of NF-�B activation in CLL is not an entirelynovel observation, because electrophoretic mobility shift anal-ysis data showing variability in tumoral cells and cell lines havebeen already reported (16). Phosphorylated I�B� has been usedas a surrogate for the activation of NF-�B, because it constitutesa key step in the release of NF-�B subunits and eventual nucleartranslocation (17, 18). Moreover, a variable response to BCRsignaling in CLL cells has been reported previously (33, 50, 51)and linked to the presence of decreased levels of functionallyrelevant adhesion molecules, cell-signaling receptors, and11q22-q23 deletion (16). Gene expression profiling studies (52,53) have also showed the presence of an IgVH mutational status
Fig. 4 Kaplan-Meier curve showing association between p-I�B� ex-pression and OS.
Table 5 Differentially expressed genes in cases with highp-I�B� expression
Function Total genesExpression
ratio (median) Genes
NF-�B 33 �2-fold 1�2-fold 3
BCR signaling 12 �2-fold 0�2-fold 3
Apoptosis 44 �2-fold 2�2-fold 2
Cell growth and maintenance 179 �2-fold 6�2-fold 5
Metabolism 415 �2-fold 13�2-fold 14
Lymph node-homing 4 �2-fold 1�2-fold 1
Cell communication 177 �2-fold 7�2-fold 9
Other or unknown function 288 �2-fold 13�2-fold 12
NOTE. Genes are divided into functional groups, as defined byGene Ontology. Genes are considered up- or down-regulated if themedian gene expression was �2-fold or �2-fold, respectively.
Table 6 NF-�B genes associated with high level ofp-I�B� expression
Genesymbol Gene description
N-fold(average)
IRAK1 Interleukin-1 receptor-associated kinase 1 1.519PAWR PRKC, apoptosis, WT1, regulator 1.295IGF1 Insulin-like growth factor 1 (somatomedin C) 1.125BAFF B-cell activating factor of the TNF family 1.250FAF1 Fas (TNFRSF6)-associated factor 1 0.998
6804 Differences in NF-�B Activation in CLL
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

gene signature, composed of genes that have various expres-sions among mutated and unmutated cases, thus underlining therelevance of BCR signaling to the generation of the cellularexpression profile.
The data from this study reveal the variable degree ofexpression of p-IkB�, a surrogate of NF-�B activation, at leastin CLL cases with nodal involvement, linking this heterogeneitywith the expression of genes involved in the BCR signaling andNF-�B activation. Whether this finding is also relevant in CLLcases with early clinical stages remains to be clarified.
If this finding is confirmed in larger CLL series with earlyor advanced stages, p-I�B� expression could be added to the listof biological variables predicting clinical behavior that alreadyincludes IgVH somatic mutation, ZAP-70 expression, and othercytogenetic alterations. Data here obtained do not provide anexplanation for the more adverse clinical course in cases lackingNF-�B activation; a reason for this phenomenon would be givenby the presence of alternative cytogenetic or molecular alter-ations. This contrasts with the situation observed in multiplemyeloma, where an increased NF-�B activity is associated withenhanced tumor cell survival (54).
The study has also identified a large set of molecules thatcould be functionally assayed in the future with the aim ofidentifying therapeutic targets.
ACKNOWLEDGMENTSWe thank the CNIO Tumor Bank for providing most of the samples
analyzed. The authors acknowledge the technical assistance of theGenomics and Histology Units and the Molecular Pathology Program.We also thank Juan C. Cigudosa for carrying out FISH analysis andRamon Diaz for statistical advice.
REFERENCES1. Binet JL, Lepoprier M, Dighiero G, et al. A clinical staging systemfor chronic lymphocytic leukemia: prognostic significance. Cancer(Phila) 1977;40:855–64.2. Rai KR, Montserrat E. Prognostic factors in chronic lymphocyticleukemia. Semin Hematol 1987;24:252–6.3. Dohner H, Stilgenbauer S, Benner A, et al. Genomic aberrations andsurvival in chronic lymphocytic leukemia. N Engl J Med 2000;343:1910–6.4. Hamblin TJ, Davis Z, Gardiner A, Oscier DG, Stevenson FK. Un-mutated Ig V(H) genes are associated with a more aggressive form ofchronic lymphocytic leukemia. Blood 1999;94:1848–54.5. Hamblin TJ, Orchard JA, Ibbotson RE, et al. CD38 expression andimmunoglobulin variable region mutations are independent prognosticvariables in chronic lymphocytic leukemia, but CD38 expression mayvary during the course of the disease. Blood 2002;99:1023–9.6. Damle RN, Wasil T, Fais F, et al. Ig V gene mutation status andCD38 expression as novel prognostic indicators in chronic lymphocyticleukemia. Blood 1999;94:1840–7.7. Ibrahim S, Keating M, Do KA, et al. CD38 expression as an impor-tant prognostic factor in B-cell chronic lymphocytic leukemia. Blood2001;98:181–6.
8. Wiestner A, Rosenwald A, Barry TS, et al. ZAP-70 expressionidentifies a chronic lymphocytic leukemia subtype with unmutatedimmunoglobulin genes, inferior clinical outcome, and distinct geneexpression profile. Blood 2003;101:4944–51.
9. Krober A, Seiler T, Benner A, et al. V(H) mutation status, CD38expression level, genomic aberrations, and survival in chronic lympho-cytic leukemia. Blood 2002;100:1410–6.
10. Crespo M, Bosch F, Villamor N, et al. ZAP-70 expression as asurrogate for immunoglobulin-variable-region mutations in chroniclymphocytic leukemia. N Engl J Med 2003;348:1764–75.
11. Durig J, Nuckel H, Cremer M, et al. ZAP-70 expression is aprognostic factor in chronic lymphocytic leukemia. Leukemia 2003;17:2426–34.
12. Furman RR, Asgary Z, Mascarenhas JO, Liou HC, Schattner EJ.Modulation of NF-kappa B activity and apoptosis in chronic lympho-cytic leukemia B cells. J Immunol 2000;164:2200–6.
13. Viatour P, Bentires-Alj M, Chariot A, et al. NF- kappa B2/p100induces Bcl-2 expression. Leukemia 2003;17:1349–56.
14. Zaninoni A, Imperiali FG, Pasquini C, Zanella A, Barcellini W.Cytokine modulation of nuclear factor-kappaB activity in B-chroniclymphocytic leukemia. Exp Hematol 2003;31:185–90.
15. Kern C, Cornuel JF, Billard C, et al. Involvement of BAFF andAPRIL in the resistance to apoptosis of B-CLL through an autocrinepathway. Blood 2004;103:679–88.
16. Sembries S, Pahl H, Stilgenbauer S, Dohner H, Schriever F. Re-duced expression of adhesion molecules and cell signaling receptors bychronic lymphocytic leukemia cells with 11q deletion. Blood 1999;93:624–31.
17. Sun Q, Matta H, Chaudhary PM. The human herpes virus 8-en-coded viral FLICE inhibitory protein protects against growth factorwithdrawal-induced apoptosis via NF-kappa B activation. Blood 2003;101:1956–61.
18. Liu L, Eby MT, Rathore N, Sinha SK, Kumar A, Chaudhary PM.The human herpes virus 8-encoded viral FLICE inhibitory proteinphysically associates with and persistently activates the Ikappa B kinasecomplex. J Biol Chem 2002;277:13745–51.
19. Jaffe ES Harris NL, Stein H, Vardiman JW. Pathology andgenetics of tumours of haematopoietic and lymphoid tissues. Worldhealth organization classification of tumours. Lyon, Fance: IARCPress; 2001.
20. Cheson BD, Bennett JM, Grever M, et al. National Cancer Institute-sponsored Working Group guidelines for chronic lymphocytic leuke-mia: revised guidelines for diagnosis and treatment. Blood 1996;87:4990–7.
21. Martinez N, Camacho FI, Algara P, et al. The molecular signatureof mantle cell lymphoma reveals multiple signals favoring cell survival.Cancer Res 2003;63:8226–32.
22. Tracey L, Villuendas R, Ortiz P, et al. Identification of genesinvolved in resistance to interferon-alpha in cutaneous T-cell lym-phoma. Am J Pathol 2002;161:1825–37.
23. Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysisand display of genome-wide expression patterns. Proc Natl Acad SciUSA 1998;95:14863–8.
24. Herrero J, Al-Shahrour F, Diaz-Uriarte R, et al. GEPAS: a web-based resource for microarray gene expression data analysis. NucleicAcids Res 2003;31:3461–7.
25. Algara P, Mateo MS, Sanchez-Beato M, et al. Analysis of theIgV(H) somatic mutations in splenic marginal zone lymphoma definesa group of unmutated cases with frequent 7q deletion and adverseclinical course. Blood 2002;99:1299–304.
26. Moreno-Bueno G, Sanchez-Estevez C, Cassia R, et al. Differentialgene expression profile in endometrioid and nonendometrioid endome-trial carcinoma: STK15 is frequently overexpressed and amplified innonendometrioid carcinomas. Cancer Res 2003;63:5697–702.27. Mathas S, Lietz A, Janz M, et al. Inhibition of NF-kappaB essen-tially contributes to arsenic-induced apoptosis. Blood 2003;102:1028–34.
Table 7 Multivariate Cox hazard ratios of Zap-70 and p-I�B�expression
Hazard ratio 95% CI P
Zap-70 6.45 1.84–22.64 �0.01p-I�B� 10.3 2.56–41.5 �0.01
6805Clinical Cancer Research
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

28. Okada T, Ngo VN, Ekland EH, et al. Chemokine requirements forB cell entry to lymph nodes and Peyer’s patches. J Exp Med 2002;196:65–75.29. Saijo K, Schmedt C, Su IH, et al. Essential role of Src-familyprotein tyrosine kinases in NF-kappaB activation during B cell devel-opment. Nat Immunol 2003;4:274–9.30. Petro JB, Khan WN. Phospholipase C-gamma 2 couples Bruton’styrosine kinase to the NF-kappaB signaling pathway in B lymphocytes.J Biol Chem 2001;276:1715–9.31. Petro JB, Castro I, Lowe J, Khan WN. Bruton’s tyrosine kinasetargets NF-kappaB to the bcl-x promoter via a mechanism involvingphospholipase C-gamma2 following B cell antigen receptor engage-ment. FEBS Lett 2002;532:57–60.32. Korz C, Pscherer A, Benner A, et al. Evidence for distinct patho-mechanisms in B-cell chronic lymphocytic leukemia and mantle celllymphoma by quantitative expression analysis of cell cycle and apopto-sis-associated genes. Blood 2002;99:4554–61.33. Bernal A. Survival of leukemic B cells promoted by engagement ofthe antigen receptor. Blood 2001;98:3050–7.34. Hase H, Kanno Y, Kojima M, et al. BAFF/BLyS can potentiateB-cell selection with the B-cell co-receptor complex. Blood 2003.35. Venturi GM, Tu L, Kadono T, et al. Leukocyte migration is regu-lated by L-selectin endoproteolytic release. Immunity 2003;19:713–24.36. Orchard JA, Ibbotson RE, Davis Z, et al. ZAP-70 expression andprognosis in chronic lymphocytic leukaemia. Lancet 2004;363:105–11.37. Fais F, Ghiotto F, Hashimoto S, et al. Chronic lymphocytic leuke-mia B cells express restricted sets of mutated and unmutated antigenreceptors. J Clin Investig 1998;102:1515–25.38. Kipps TJ, Tomhave E, Pratt LF, Duffy S, Chen PP, Carson DA.Developmentally restricted immunoglobulin heavy chain variable re-gion gene expressed at high frequency in chronic lymphocytic leukemia.Proc Natl Acad Sci USA 1989;86:5913–7.39. Okkenhaug K, Vanhaesebroeck B. PI3K in lymphocyte develop-ment, differentiation and activation. Nat Rev Immunol 2003;3:317–30.40. Sekharam M, Zhao H, Sun M, et al. Insulin-like growth factor 1receptor enhances invasion and induces resistance to apoptosis of coloncancer cells through the Akt/Bcl-x(L) pathway. Cancer Res 2003;63:7708–16.41. Park MY, Jang HD, Lee SY, Lee KJ, Kim E. Fas associated factor1 inhibits NF-kB activity by interfering with nuclear translocation ofRelA (p65) subunit of NF-kB. J Biol Chem 2003.
42. Zhang W, Kone BC. NF-kappaB inhibits transcription of the H()-K()-ATPase alpha(2)-subunit gene: role of histone deacetylases. Am JPhysiol Renal Physiol 2002;283:F904–11.
43. Granziero L, Ghia P, Circosta P, et al. Survivin is expressed onCD40 stimulation and interfaces proliferation and apoptosis in B-cellchronic lymphocytic leukemia. Blood 2001;97:2777–83.
44. Khan WN. Regulation of B lymphocyte development and activationby Bruton’s tyrosine kinase. Immunol Res 2001;23:147–56.
45. Kawauchi K, Ogasawara T, Yasuyama M. Activation of extracel-lular signal-regulated kinase through B-cell antigen receptor in B-cellchronic lymphocytic leukemia. Int J Hematol 2002;75: 508–13.46. Lanham S, Hamblin T, Oscier D, Ibbotson R, Stevenson F, Pack-ham G. Differential signaling via surface IgM is associated with VHgene mutational status and CD38 expression in chronic lymphocyticleukemia. Blood 2003;101:1087–93.47. Chen L, Widhopf G, Huynh L, et al. Expression of ZAP-70 isassociated with increased B-cell receptor signaling in chronic lympho-cytic leukemia. Blood 2002;100:4609–14.48. Haxhinasto SA, Bishop GA. Synergistic B cell activation by CD40and the B cell antigen receptor: Role of BCR-mediated kinase activationand TRAF regulation. J Biol Chem 2003.49. Suzuki N, Suzuki S, Duncan GS, et al. Severe impairment ofinterleukin-1 and Toll-like receptor signalling in mice lacking IRAK-4.Nature (Lond) 2002;416:750–6.50. Zupo S, Cutrona G, Mangiola M, Ferrarini M. Role of surface IgMand IgD on survival of the cells from B-cell chronic lymphocyticleukemia. Blood 2002;99:2277–8.51. Hivroz C, Geny B, Brouet JC, Grillot-Courvalin C. Alteredsignal transduction secondary to surface IgM cross-linking on B-chronic lymphocytic leukemia cells. Differential activation of thephosphatidylinositol-specific phospholipase C. J Immunol 1990;144:2351– 8.52. Rosenwald A, Alizadeh AA, Widhopf G, et al. Relation of geneexpression phenotype to immunoglobulin mutation genotype in B cellchronic lymphocytic leukemia. J Exp Med 2001;194:1639–47.53. Klein U, Tu Y, Stolovitzky GA, et al. Gene expression profiling ofB cell chronic lymphocytic leukemia reveals a homogeneous phenotyperelated to memory B cells. J Exp Med 2001;194:1625–38.54. Berenson JR, Ma HM, Vescio R. The role of nuclear factor-kappaBin the biology and treatment of multiple myeloma. Semin Oncol 2001;28:626–33.
6806 Differences in NF-�B Activation in CLL
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from

2004;10:6796-6806. Clin Cancer Res Antonia Rodríguez, Nerea Martínez, Francisca I. Camacho, et al. Involvement
in Chronic Lymphocytic Leukemia Cases With NodalαBκVariability in the Degree of Expression of Phosphorylated I
Updated version
http://clincancerres.aacrjournals.org/content/10/20/6796
Access the most recent version of this article at:
Cited articles
http://clincancerres.aacrjournals.org/content/10/20/6796.full#ref-list-1
This article cites 49 articles, 30 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/10/20/6796.full#related-urls
This article has been cited by 6 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/10/20/6796To request permission to re-use all or part of this article, use this link
Cancer Research. on November 7, 2018. © 2004 American Association forclincancerres.aacrjournals.org Downloaded from


















