
Unraveling functional significance of natural variations ... · PDF fileof a human galectin by...
6
Unraveling functional significance of natural variations of a human galectin by glycodendrimersomes with programmable glycan surface Shaodong Zhang a , Ralph-Olivier Moussodia a , Sabine Vértesy b , Sabine André b , Michael L. Klein c,1 , Hans-Joachim Gabius b , and Virgil Percec a,1 a Roy & Diana Vagelos Laboratories, Department of Chemistry, University of Pennsylvania, Philadelphia, PA 19104-6323; b Institute of Physiological Chemistry, Faculty of Veterinary Medicine, Ludwig-Maximilians-University Munich, 80539 Munich, Germany; and c Institute of Computational Molecular Science, Temple University, Philadelphia, PA 19122 Contributed by Michael L. Klein, March 31, 2015 (sent for review March 5, 2015; reviewed by Ricardo Riguera Vega and Peter H. Seeberger) Surface-presented glycans (complex carbohydrates) are docking sites for adhesion/growth-regulatory galectins within cell–cell/ matrix interactions. Alteration of the linker length in human galectin- 8 and single-site mutation (F19Y) are used herein to illustrate the potential of glycodendrimersomes with programmable glycan displays as a model system to reveal the functional impact of nat- ural sequence variations in trans recognition. Extension of the linker length slightly reduces lectin capacity as agglutinin and slows down aggregate formation at low ligand surface density. The mutant protein is considerably less active as agglutinin and less sensitive to low-level ligand presentation. The present results suggest that mimicking glycan complexity and microdomain occur- rence on the glycodendrimersome surface can provide key insights into mechanisms to accomplish natural selectivity and specificity of lectins in structural and topological terms. adhesion | agglutination | glycobiology | membrane mimic | self-assembly T he sociology of cells critically depends on selective inter- actions between cells and the extracellular matrix. Taking advantage of the capacity of carbohydrates to store biological information, glycans are a versatile means to generate the re- quired cell-surface recognition (1–3). These molecular signals are docking sites for tissue receptors (lectins), which translate the glycan-based information into adhesion and bridging, often triggering outside-in signaling (3–6). Key insights into these processes have been obtained by teaming up synthetic chemistry, which delivers a defined custom-made matrix, with bioassays using cells (2, 7, 8). Questions on sugar specificity and density leading to the detection of threshold phenomena were resolved this way for hepatocyte and macrophage adhesion (7, 9). Ex- tending this conceptual approach to cell models, we recently introduced glycodendrimersomes with programmable glycan dis- play and established their bioactivity by testing lectins for their capacity in bridging (trans interactions) (10, 11). Having thus validated design and suitability for application of the test system, it can now be applied to resolve questions on physiological structure–activity correlation for tissue lectins, thereby merging supramolecular chemistry with biomedicine. The present study focuses on two such issues for the class of adhesion/growth regulatory galectins (5, 12, 13). One mode of structural design for cross-linking is the tandem- repeat display of two-carbohydrate recognition domains (CRDs) connected by a linker peptide shown for galectin-8 (Gal-8) in Fig. 1. As expected, this bivalent protein is a potent mediator of cell adhesion, referred to as a “novel type of matricellular pro- tein” (14–16). Physiologically, alternative splicing facilitates its occurrence in two isoforms with different linker lengths, termed 8S and 8L (Fig. 1; for details of amino acid sequences, see Fig. S1) (17, 18). Because the functional impact of this change has not yet been defined, the expression pattern appears to be stochastic in nature (17). Notably, the presence of Gal-8 is conserved in other branches of the phylogenetic tree, with fur- ther variation in the linker length, e.g., in the chicken. This ortholog, i.e., Gal-8S (CG-8S), has a linker of only nine amino acids (Fig. 1) (19). Because the linker is susceptible to proteolytic cleavage (20), the two CRDs can be separated from each other and present as free proteins. Equally interesting is the fact that Gal-8 is the first case of a single amino acid polymorphism in the coding region with medical relevance, i.e., associated with rheu- matoid arthritis (21). The F19Y substitution causes a displace- ment of ∼1.5 Å of the positions of N-terminal amino acids 11–15 and a shift of the β-strand F0 in the vicinity of the linker, with impact on thermal stability and enthalpic/entropic contributions to ligand binding (22). Thus, questions on relative bioactivity levels depending on linker length and sugar density on the glycodendrimersomes can be answered on a proof-of-principle basis, as can be done for the natural variant (F19Y). The re- spective experiments reveal a conspicuous significance of the examined parameter changes and complete switch-off by linker cleavage. Results and Discussion Topological Aspects of Ligand Reactivity. Distinct topological modes of ligand presentation have been assessed by their reactivity to Gal-8S; this can lead to bridging between glycodendrimersomes, which experimentally is followed by an increase in absorbance at Significance Lectins are endogenous sugar receptors involved in diverse physiological and disease-associated processes. The functional consequences of naturally occurring single-nucleotide poly- morphism and alternative splicing in lectins has been explored using glycodendrimersomes, a versatile test system with pro- grammable glycan (complex carbohydrates) display. Importantly, glycodendrimersomes facilitate quantitative determination of lectin-mediated cross-linking, a hallmark of their activity. Thresh- old and kinetic effects measured for a human galectin associated with autoimmune disease document the sensitivity of the test system and highlight its potential as a new and highly versatile supramolecular sensor for biomedical applications. Author contributions: H.-J.G. and V.P. designed research; S.Z., R.-O.M., S.V., and S.A. performed research; S.Z., S.A., M.L.K., H.-J.G., and V.P. analyzed data; and S.Z., S.A., M.L.K., H.-J.G., and V.P. wrote the paper. Reviewers: R.R.V., University of Santiago de Compostela; and P.H.S., Max Planck Institute of Colloids and Interfaces. The authors declare no conflict of interest. Freely available online through the PNAS open access option. 1 To whom correspondence may be addressed. Email: [email protected] or percec@sas. upenn.edu. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1506220112/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1506220112 PNAS | May 5, 2015 | vol. 112 | no. 18 | 5585–5590 CHEMISTRY
Transcript of Unraveling functional significance of natural variations ... · PDF fileof a human galectin by...

Unraveling functional significance of natural variationsof a human galectin by glycodendrimersomes withprogrammable glycan surfaceShaodong Zhanga, Ralph-Olivier Moussodiaa, Sabine Vértesyb, Sabine Andréb, Michael L. Kleinc,1,Hans-Joachim Gabiusb, and Virgil Perceca,1
aRoy & Diana Vagelos Laboratories, Department of Chemistry, University of Pennsylvania, Philadelphia, PA 19104-6323; bInstitute of Physiological Chemistry,Faculty of Veterinary Medicine, Ludwig-Maximilians-University Munich, 80539 Munich, Germany; and cInstitute of Computational Molecular Science,Temple University, Philadelphia, PA 19122
Contributed by Michael L. Klein, March 31, 2015 (sent for review March 5, 2015; reviewed by Ricardo Riguera Vega and Peter H. Seeberger)
Surface-presented glycans (complex carbohydrates) are dockingsites for adhesion/growth-regulatory galectins within cell–cell/matrix interactions. Alteration of the linker length in human galectin-8 and single-site mutation (F19Y) are used herein to illustratethe potential of glycodendrimersomes with programmable glycandisplays as a model system to reveal the functional impact of nat-ural sequence variations in trans recognition. Extension of thelinker length slightly reduces lectin capacity as agglutinin andslows down aggregate formation at low ligand surface density.The mutant protein is considerably less active as agglutinin andless sensitive to low-level ligand presentation. The present resultssuggest that mimicking glycan complexity and microdomain occur-rence on the glycodendrimersome surface can provide key insightsinto mechanisms to accomplish natural selectivity and specificityof lectins in structural and topological terms.
adhesion | agglutination | glycobiology | membrane mimic | self-assembly
The sociology of cells critically depends on selective inter-actions between cells and the extracellular matrix. Taking
advantage of the capacity of carbohydrates to store biologicalinformation, glycans are a versatile means to generate the re-quired cell-surface recognition (1–3). These molecular signalsare docking sites for tissue receptors (lectins), which translatethe glycan-based information into adhesion and bridging, oftentriggering outside-in signaling (3–6). Key insights into theseprocesses have been obtained by teaming up synthetic chemistry,which delivers a defined custom-made matrix, with bioassaysusing cells (2, 7, 8). Questions on sugar specificity and densityleading to the detection of threshold phenomena were resolvedthis way for hepatocyte and macrophage adhesion (7, 9). Ex-tending this conceptual approach to cell models, we recentlyintroduced glycodendrimersomes with programmable glycan dis-play and established their bioactivity by testing lectins for theircapacity in bridging (trans interactions) (10, 11). Having thusvalidated design and suitability for application of the test system,it can now be applied to resolve questions on physiologicalstructure–activity correlation for tissue lectins, thereby mergingsupramolecular chemistry with biomedicine. The present studyfocuses on two such issues for the class of adhesion/growthregulatory galectins (5, 12, 13).One mode of structural design for cross-linking is the tandem-
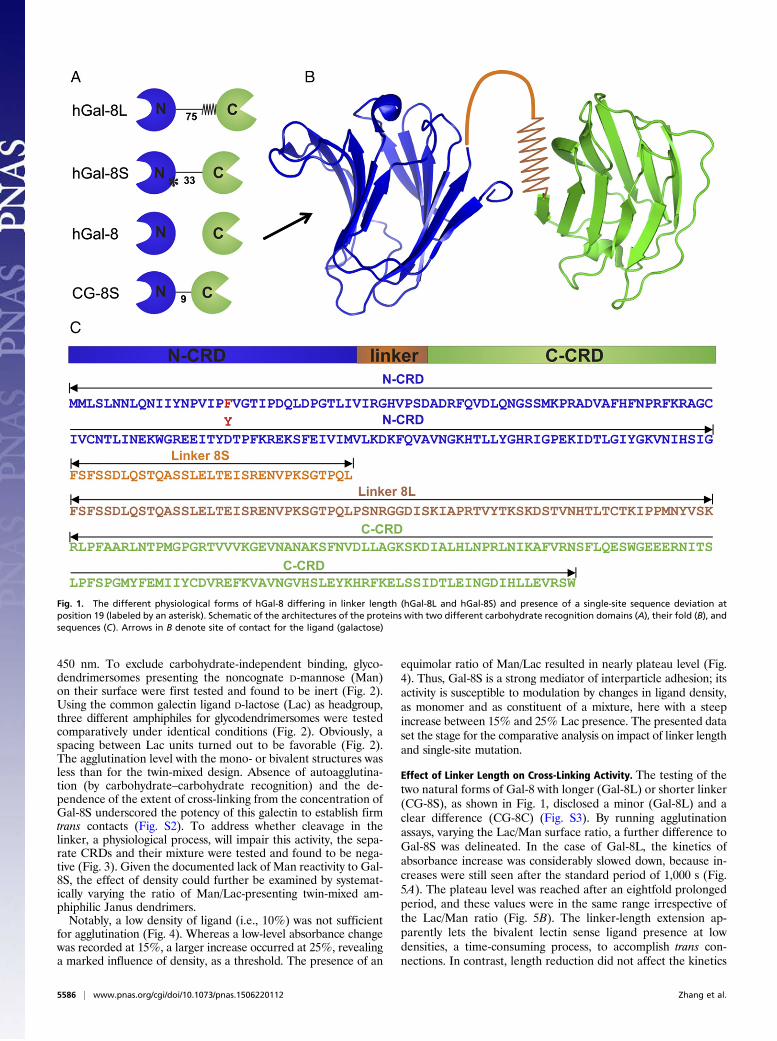
repeat display of two-carbohydrate recognition domains (CRDs)connected by a linker peptide shown for galectin-8 (Gal-8) inFig. 1. As expected, this bivalent protein is a potent mediator ofcell adhesion, referred to as a “novel type of matricellular pro-tein” (14–16). Physiologically, alternative splicing facilitates itsoccurrence in two isoforms with different linker lengths, termed8S and 8L (Fig. 1; for details of amino acid sequences, see Fig.S1) (17, 18). Because the functional impact of this changehas not yet been defined, the expression pattern appears to bestochastic in nature (17). Notably, the presence of Gal-8 is
conserved in other branches of the phylogenetic tree, with fur-ther variation in the linker length, e.g., in the chicken. Thisortholog, i.e., Gal-8S (CG-8S), has a linker of only nine aminoacids (Fig. 1) (19). Because the linker is susceptible to proteolyticcleavage (20), the two CRDs can be separated from each otherand present as free proteins. Equally interesting is the fact thatGal-8 is the first case of a single amino acid polymorphism in thecoding region with medical relevance, i.e., associated with rheu-matoid arthritis (21). The F19Y substitution causes a displace-ment of ∼1.5 Å of the positions of N-terminal amino acids 11–15and a shift of the β-strand F0 in the vicinity of the linker, withimpact on thermal stability and enthalpic/entropic contributionsto ligand binding (22). Thus, questions on relative bioactivitylevels depending on linker length and sugar density on theglycodendrimersomes can be answered on a proof-of-principlebasis, as can be done for the natural variant (F19Y). The re-spective experiments reveal a conspicuous significance of theexamined parameter changes and complete switch-off by linkercleavage.
Results and DiscussionTopological Aspects of Ligand Reactivity. Distinct topological modesof ligand presentation have been assessed by their reactivity toGal-8S; this can lead to bridging between glycodendrimersomes,which experimentally is followed by an increase in absorbance at
Significance
Lectins are endogenous sugar receptors involved in diversephysiological and disease-associated processes. The functionalconsequences of naturally occurring single-nucleotide poly-morphism and alternative splicing in lectins has been exploredusing glycodendrimersomes, a versatile test system with pro-grammable glycan (complex carbohydrates) display. Importantly,glycodendrimersomes facilitate quantitative determination oflectin-mediated cross-linking, a hallmark of their activity. Thresh-old and kinetic effects measured for a human galectin associatedwith autoimmune disease document the sensitivity of the testsystem and highlight its potential as a new and highly versatilesupramolecular sensor for biomedical applications.
Author contributions: H.-J.G. and V.P. designed research; S.Z., R.-O.M., S.V., and S.A.performed research; S.Z., S.A., M.L.K., H.-J.G., and V.P. analyzed data; and S.Z., S.A.,M.L.K., H.-J.G., and V.P. wrote the paper.
Reviewers: R.R.V., University of Santiago de Compostela; and P.H.S., Max Planck Instituteof Colloids and Interfaces.
The authors declare no conflict of interest.
Freely available online through the PNAS open access option.1To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1506220112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1506220112 PNAS | May 5, 2015 | vol. 112 | no. 18 | 5585–5590
CHEM
ISTR
Y
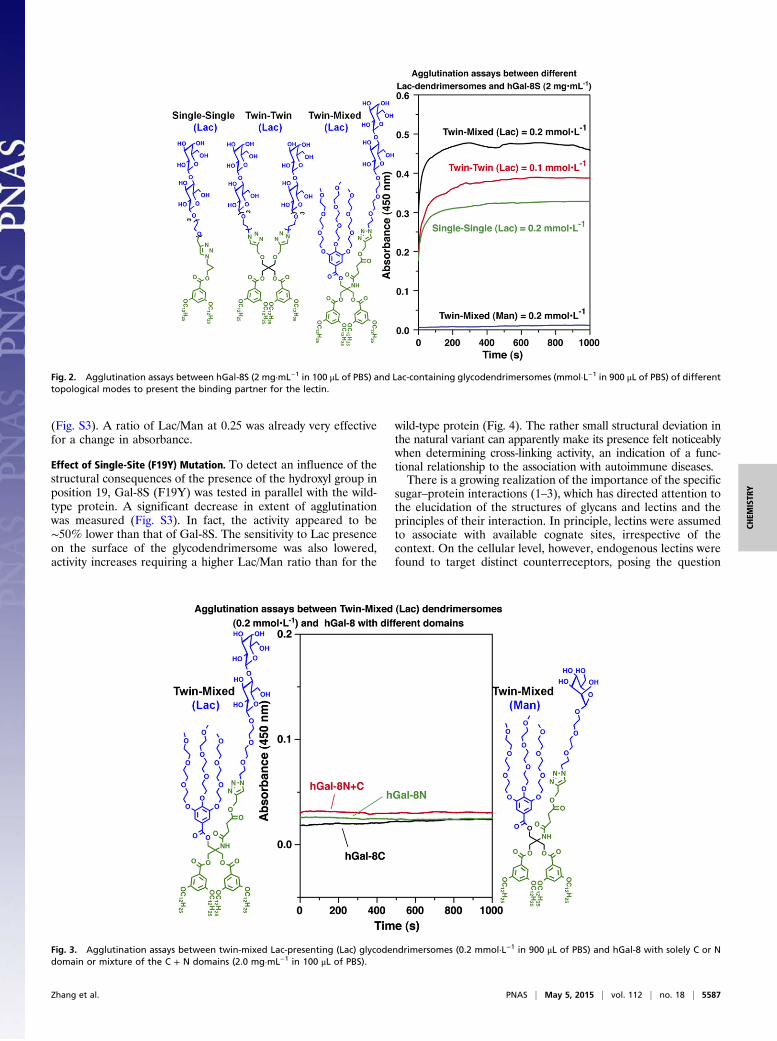
450 nm. To exclude carbohydrate-independent binding, glyco-dendrimersomes presenting the noncognate D-mannose (Man)on their surface were first tested and found to be inert (Fig. 2).Using the common galectin ligand D-lactose (Lac) as headgroup,three different amphiphiles for glycodendrimersomes were testedcomparatively under identical conditions (Fig. 2). Obviously, aspacing between Lac units turned out to be favorable (Fig. 2).The agglutination level with the mono- or bivalent structures wasless than for the twin-mixed design. Absence of autoagglutina-tion (by carbohydrate–carbohydrate recognition) and the de-pendence of the extent of cross-linking from the concentration ofGal-8S underscored the potency of this galectin to establish firmtrans contacts (Fig. S2). To address whether cleavage in thelinker, a physiological process, will impair this activity, the sepa-rate CRDs and their mixture were tested and found to be nega-tive (Fig. 3). Given the documented lack of Man reactivity to Gal-8S, the effect of density could further be examined by systemat-ically varying the ratio of Man/Lac-presenting twin-mixed am-phiphilic Janus dendrimers.Notably, a low density of ligand (i.e., 10%) was not sufficient
for agglutination (Fig. 4). Whereas a low-level absorbance changewas recorded at 15%, a larger increase occurred at 25%, revealinga marked influence of density, as a threshold. The presence of an
equimolar ratio of Man/Lac resulted in nearly plateau level (Fig.4). Thus, Gal-8S is a strong mediator of interparticle adhesion; itsactivity is susceptible to modulation by changes in ligand density,as monomer and as constituent of a mixture, here with a steepincrease between 15% and 25% Lac presence. The presented dataset the stage for the comparative analysis on impact of linker lengthand single-site mutation.
Effect of Linker Length on Cross-Linking Activity. The testing of thetwo natural forms of Gal-8 with longer (Gal-8L) or shorter linker(CG-8S), as shown in Fig. 1, disclosed a minor (Gal-8L) and aclear difference (CG-8C) (Fig. S3). By running agglutinationassays, varying the Lac/Man surface ratio, a further difference toGal-8S was delineated. In the case of Gal-8L, the kinetics ofabsorbance increase was considerably slowed down, because in-creases were still seen after the standard period of 1,000 s (Fig.5A). The plateau level was reached after an eightfold prolongedperiod, and these values were in the same range irrespective ofthe Lac/Man ratio (Fig. 5B). The linker-length extension ap-parently lets the bivalent lectin sense ligand presence at lowdensities, a time-consuming process, to accomplish trans con-nections. In contrast, length reduction did not affect the kinetics
Fig. 1. The different physiological forms of hGal-8 differing in linker length (hGal-8L and hGal-8S) and presence of a single-site sequence deviation atposition 19 (labeled by an asterisk). Schematic of the architectures of the proteins with two different carbohydrate recognition domains (A), their fold (B), andsequences (C). Arrows in B denote site of contact for the ligand (galactose)
5586 | www.pnas.org/cgi/doi/10.1073/pnas.1506220112 Zhang et al.
(Fig. S3). A ratio of Lac/Man at 0.25 was already very effectivefor a change in absorbance.
Effect of Single-Site (F19Y) Mutation. To detect an influence of thestructural consequences of the presence of the hydroxyl group inposition 19, Gal-8S (F19Y) was tested in parallel with the wild-type protein. A significant decrease in extent of agglutinationwas measured (Fig. S3). In fact, the activity appeared to be∼50% lower than that of Gal-8S. The sensitivity to Lac presenceon the surface of the glycodendrimersome was also lowered,activity increases requiring a higher Lac/Man ratio than for the
wild-type protein (Fig. 4). The rather small structural deviation inthe natural variant can apparently make its presence felt noticeablywhen determining cross-linking activity, an indication of a func-tional relationship to the association with autoimmune diseases.There is a growing realization of the importance of the specific
sugar–protein interactions (1–3), which has directed attention tothe elucidation of the structures of glycans and lectins and theprinciples of their interaction. In principle, lectins were assumedto associate with available cognate sites, irrespective of thecontext. On the cellular level, however, endogenous lectins werefound to target distinct counterreceptors, posing the question
Fig. 2. Agglutination assays between hGal-8S (2 mg·mL−1 in 100 μL of PBS) and Lac-containing glycodendrimersomes (mmol·L−1 in 900 μL of PBS) of differenttopological modes to present the binding partner for the lectin.
Fig. 3. Agglutination assays between twin-mixed Lac-presenting (Lac) glycodendrimersomes (0.2 mmol·L−1 in 900 μL of PBS) and hGal-8 with solely C or Ndomain or mixture of the C + N domains (2.0 mg·mL−1 in 100 μL of PBS).
Zhang et al. PNAS | May 5, 2015 | vol. 112 | no. 18 | 5587
CHEM
ISTR
Y

of the underlying causes for the selectivity (23, 24). For exam-ple, suitably glycosylated T-cell glycoproteins CD7, CD43, andCD45; laminin or carcinoembryonic antigen on colon carcinomacells; and the α5β1-integrin (fibronectin receptor) upon tumorsuppressor expression in pancreatic carcinoma cells are the
main binding partner and the functional counter receptors forGal-1/Gal-3 (25–27). Finding out why, despite the abundanceof cell surface β-galactosides, tissue galectins exhibit this pref-erence poses a major challenge. Obviously, model systems withprogrammable surface properties will be valuable to resolve this
Fig. 4. Agglutination assays between hGal-8S (2 mg·mL−1 in 100 μL of PBS) and mixtures of twin-mixed Man-containing (Man) and Lac-containing (Lac)glycodendrimersomes to produce different surface compositions (Man + Lac = 0.2 mmol·L−1 in 900 μL of PBS).
Fig. 5. Agglutination assays between hGal-8L (2 mg·mL−1 in 100 μL of PBS) and mixtures of twin-mixed Man-containing (Man) and Lac-containing (Lac)glycodendrimersomes to produce different surface compositions (Man + Lac = 0.2 mmol·L−1 in 900 μL of PBS) for 1,500 s (A) and 8,000 s (B).
5588 | www.pnas.org/cgi/doi/10.1073/pnas.1506220112 Zhang et al.
conundrum. Equally important is the fact that the in-depth study ofstructural features of endogenous lectins has delineated physio-logical variations. If a single-nucleotide polymorphism, one routeto such changes on the genetic level that is seen in populations,causes occurrence of an early stop and thus a defective receptor,then the emerging loss of a function is readily interpreted. Thiscase is possible for occurrence of Y238X in the gene for theβ-glucan receptor dectin-1 and its association to fungal infections inpatients (28, 29). At this early stage of analyzing such variants, casestudies on single-nucleotide polymorphisms have either revealedrelevance for quaternary structure (for a serum collectin) andprocessing by proteolysis (for Gal-3) or no effect (for E-selectin)(30–32). A readily applicable robust functional assay would con-siderably help track down implications of the sequence variation. Inthis study, we apply a highly versatile chemical platform with cell-like features to investigate whether and how the length of the linkerin a tandem repeat-type galectin and the natural single-site muta-tion affect the protein’s activity as mediator of trans contacts.Following the demonstration of carbohydrate/protein concen-
tration dependence, wild-type Gal-8S is shown to be a potentagglutinin of Lac-presenting glycodendrimersomes. The linker ofthis lectin, shown by NMR spectroscopy to be rather unstructuredwhen connecting the two CRDs of Gal-1 (33), appears to presentthe two binding sites of Gal-8S capable for bridging vesicles.When probing the mode of Lac presentation, a hybrid with onlyone sugar headgroup in a bivalent setting accessible on the gly-codendrimersome surface results in higher signals than a denserLac presentation by the mono- or bivalent analogs. Extension ofthe linker, mechanistically by alternative splicing, leads to re-duced extent of agglutination. Mimicking glycome complexity insimple terms by mixing Lac/Man-presenting building blocks, twofurther differences were disclosed with linker-length extension:(i) diminution of extent of agglutination at 100% Lac pre-sentation and (ii) retarded absorbance increase at low-level Lacpresentation. It is also interesting to note that structural mani-festation of the single-nucleotide polymorphism significantly im-pairs agglutination activity, at full-scale Lac presentation and in
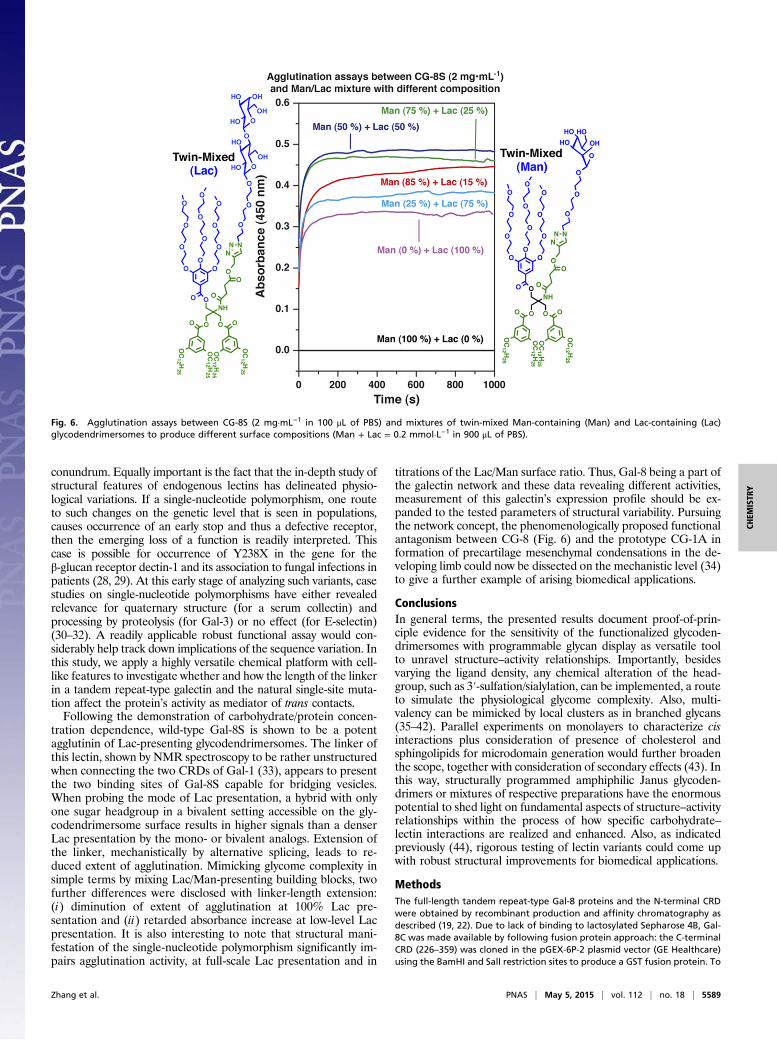
titrations of the Lac/Man surface ratio. Thus, Gal-8 being a part ofthe galectin network and these data revealing different activities,measurement of this galectin’s expression profile should be ex-panded to the tested parameters of structural variability. Pursuingthe network concept, the phenomenologically proposed functionalantagonism between CG-8 (Fig. 6) and the prototype CG-1A information of precartilage mesenchymal condensations in the de-veloping limb could now be dissected on the mechanistic level (34)to give a further example of arising biomedical applications.
ConclusionsIn general terms, the presented results document proof-of-prin-ciple evidence for the sensitivity of the functionalized glycoden-drimersomes with programmable glycan display as versatile toolto unravel structure–activity relationships. Importantly, besidesvarying the ligand density, any chemical alteration of the head-group, such as 3′-sulfation/sialylation, can be implemented, a routeto simulate the physiological glycome complexity. Also, multi-valency can be mimicked by local clusters as in branched glycans(35–42). Parallel experiments on monolayers to characterize cisinteractions plus consideration of presence of cholesterol andsphingolipids for microdomain generation would further broadenthe scope, together with consideration of secondary effects (43). Inthis way, structurally programmed amphiphilic Janus glycoden-drimers or mixtures of respective preparations have the enormouspotential to shed light on fundamental aspects of structure–activityrelationships within the process of how specific carbohydrate–lectin interactions are realized and enhanced. Also, as indicatedpreviously (44), rigorous testing of lectin variants could come upwith robust structural improvements for biomedical applications.
MethodsThe full-length tandem repeat-type Gal-8 proteins and the N-terminal CRDwere obtained by recombinant production and affinity chromatography asdescribed (19, 22). Due to lack of binding to lactosylated Sepharose 4B, Gal-8C was made available by following fusion protein approach: the C-terminalCRD (226–359) was cloned in the pGEX-6P-2 plasmid vector (GE Healthcare)using the BamHI and SalI restriction sites to produce a GST fusion protein. To
Fig. 6. Agglutination assays between CG-8S (2 mg·mL−1 in 100 μL of PBS) and mixtures of twin-mixed Man-containing (Man) and Lac-containing (Lac)glycodendrimersomes to produce different surface compositions (Man + Lac = 0.2 mmol·L−1 in 900 μL of PBS).
Zhang et al. PNAS | May 5, 2015 | vol. 112 | no. 18 | 5589
CHEM
ISTR
Y

substitute the amino acids of the BamHI cleavage site, the QuikChange Site-Directed Mutagenesis protocol (Stratagene) was used. Protein was expressedin Escherichia coli cells BL21 (DE3) pLysS (Promega) induced by addition ofisopropylβ-D-1-thiogalactopyranoside (100 μM) followed by overnight in-cubation at 22 °C. Cells were collected by centrifugation and lysed by soni-fication. The fusion protein was purified using glutathione Sepharose 4B (GEHealthcare) and the GST-tag was removed by proteolysis with human rhi-novirus 3C protease followed by separation of Gal-8C from GST and theprotease. All proteins were ascertained for purity by 1D and 2D gel electro-phoresis and mass-spectrometric analysis (peptic fingerprint) and for activityby hemagglutination, solid-phase, and cell growth assays as described (19, 22).
Glycodendrimersomes self-assembled from amphiphilic Janus dendrimersof different topology for ligand presentation (11, 44) were prepared by theinjection method as follows (45, 46). A stock solution was prepared by dis-solving the required amount of amphiphilic Janus glycodendrimers in freshlydistilled THF. Glycodendrimersomes were then generated by injection of100 μL of the stock solution into 2 mL Millipore water or PBS or Hepesbuffer, followed by 5-s vortexing to give the final concentration of glyco-dendrimers of 0.5–1 mg × mL−1 in water or in buffer. Any exceptions to thisprotocol are noted in the respective figure captions.
Agglutination assays of glycodendrimersomes and lectins were moni-tored in 1.50 mL semimicro disposable cuvettes at 23 °C at single wavelengthλ = 450 nm by using a Shimadzu UV-VIS Spectrophotometer UV-1601 withShimadzu/UV Probe software with kinetic mode. The solution of lectin(100 μL) was injected into 900 μL of solution of glycodendrimersomes. Themixture was shaken for 2 s before recording the absorbance change in time.The agglutination assays were carried out in PBS (1×) buffer. The concen-tration of lectins and glycodendrimer solutions can be found in the captionof each figure. The solutions of glycodendrimersome were freshly preparedvia injection method from THF solution into PBS (1×) buffer. The samebuffer was used to bring the lectins into solutions that were used im-mediately. The solutions of lectin were kept at 0 °C during the aggluti-nation assays.
ACKNOWLEDGMENTS. We thank Prof. B. Friday and Dr. G. Notelecs forinspiring discussions. Financial support was provided by National ScienceFoundation Grants DMR-1066116 and DMR-1120901 (to V.P.) and DMR-1120901 (to M.L.K.), the P. Roy Vagelos Chair at the University of Pennsylvania(to V.P.), and the European Commission Seventh Framework ProgrammeGrant 317297; GLYCOPHARM (to H.-J.G.).
1. Roseman S (1970) The synthesis of complex carbohydrates by multiglycosyltransferasesystems and their potential function in intercellular adhesion. Chem Phys Lipids 5(1):270–297.
2. Brandley BK, Schnaar RL (1986) Cell-surface carbohydrates in cell recognition andresponse. J Leukoc Biol 40(1):97–111.
3. Gabius HJ, ed (2009) The Sugar Code. Fundamentals of Glycosciences (Wiley, Wein-heim, Germany).
4. Barondes SH (1984) Soluble lectins: A new class of extracellular proteins. Science223(4642):1259–1264.
5. Lis H, Sharon N (1998) Lectins: Carbohydrate-specific proteins that mediate cellularrecognition. Chem Rev 98(2):637–674.
6. André S, Kaltner H, Manning JC, Murphy PV, Gabius HJ (2015) Lectins: Getting familiarwith translators of the sugar code. Molecules 20(2):1788–1823.
7. Weigel PH, et al. (1979) Adhesion of hepatocytes to immobilized sugars. A thresholdphenomenon. J Biol Chem 254(21):10830–10838.
8. Schnaar RL (1984) Immobilized glycoconjugates for cell recognition studies. Anal Bio-chem 143(1):1–13.
9. Largent BL, Walton KM, Hoppe CA, Lee YC, Schnaar RL (1984) Carbohydrate-specificadhesion of alveolar macrophages to mannose-derivatized surfaces. J Biol Chem259(3):1764–1769.
10. Percec V, et al. (2013) Modular synthesis of amphiphilic Janus glycodendrimers andtheir self-assembly into glycodendrimersomes and other complex architectures withbioactivity to biomedically relevant lectins. J Am Chem Soc 135(24):9055–9077.
11. Zhang S, et al. (2014) Mimicking biological membranes with programmable glycanligands self-assembled from amphiphilic Janus glycodendrimers. Angew Chem Int Ed53(41):10899–10903.
12. Hirabayashi J (1997) Recent topics on galectins. Trends Glycosci Glycotechnol 9(45):1–182.
13. Liu FT, Yang RY, Hsu DK (2012) Galectins in acute and chronic inflammation. Ann N YAcad Sci 1253:80–91.
14. Levy Y, et al. (2001) Galectin-8 functions as a matricellular modulator of cell adhesion.J Biol Chem 276(33):31285–31295.
15. Nishi N, et al. (2003) Galectin-8 modulates neutrophil function via interaction withintegrin alphaM. Glycobiology 13(11):755–763.
16. Zick Y, et al. (2004) Role of galectin-8 as a modulator of cell adhesion and cell growth.Glycoconj J 19(7-9):517–526.
17. Gopalkrishnan RV, et al. (2000) Molecular characterization of prostate carcinomatumor antigen-1, PCTA-1, a human galectin-8 related gene. Oncogene 19(38):4405–4416.
18. Bidon N, et al. (2001) Two messenger RNAs and five isoforms for Po66-CBP, a galectin-8 homolog in a human lung carcinoma cell line. Gene 274(1-2):253–262.
19. Kaltner H, et al. (2009) Unique chicken tandem-repeat-type galectin: Implications ofalternative splicing and a distinct expression profile compared to those of the threeproto-type proteins. Biochemistry 48(20):4403–4416.
20. Nishi N, Itoh A, Shoji H, Miyanaka H, Nakamura T (2006) Galectin-8 and galectin-9 arenovel substrates for thrombin. Glycobiology 16(11):15C–20C.
21. Pál Z, et al. (2012) Non-synonymous single nucleotide polymorphisms in genes forimmunoregulatory galectins: Association of galectin-8 (F19Y) occurrence with auto-immune diseases in a Caucasian population. Biochim Biophys Acta 1820(10):1512–1518.
22. Ruiz FM, et al. (2014) Natural single amino acid polymorphism (F19Y) in human ga-lectin-8: Detection of structural alterations and increased growth-regulatory activityon tumor cells. FEBS J 281(5):1446–1464.
23. Varki A (1994) Selectin ligands. Proc Natl Acad Sci USA 91(16):7390–7397.24. Gabius HJ, André S, Jiménez-Barbero J, Romero A, Solís D (2011) From lectin structure
to functional glycomics: Principles of the sugar code. Trends Biochem Sci 36(6):298–313.
25. Ohannesian DW, et al. (1995) Carcinoembryonic antigen and other glycoconjugatesact as ligands for galectin-3 in human colon carcinoma cells. Cancer Res 55(10):2191–2199.
26. Pace KE, Baum LG (1997) Induction of T lymphocyte apoptosis: A novel function forgalectin-1. Trends Glycosci Glycotechnol 9(45):21–30.
27. Amano M, et al. (2012) Tumour suppressor p16(INK4a) - anoikis-favouring decrease inN/O-glycan/cell surface sialylation by down-regulation of enzymes in sialic acid bio-synthesis in tandem in a pancreatic carcinoma model. FEBS J 279(21):4062–4080.
28. Ferwerda B, et al. (2009) Human dectin-1 deficiency and mucocutaneous fungal in-fections. N Engl J Med 361(18):1760–1767.
29. Marakalala MJ, Kerrigan AM, Brown GD (2011) Dectin-1: A role in antifungal defenseand consequences of genetic polymorphisms in humans. Mamm Genome 22(1-2):55–65.
30. Eisen DP, Osthoff M (2014) If there is an evolutionary selection pressure for the highfrequency of MBL2 polymorphisms, what is it? Clin Exp Immunol 176(2):165–171.
31. Balan V, et al. (2008) Racial disparity in breast cancer and functional germ line mu-tation in galectin-3 (rs4644): A pilot study. Cancer Res 68(24):10045–10050.
32. Preston RC, et al. (2014) Implications of the E-selectin S128R mutation for drug dis-covery. Glycobiology 24(7):592–601.
33. Vértesy S, et al. (2015) Structural significance of galectin design: Impairment of ho-modimer stability by linker insertion and partial reversion by ligand presence. ProteinEng Des Sel, 10.1093/protein/gzv014.
34. Bhat R, et al. (2011) A regulatory network of two galectins mediates the earliest stepsof avian limb skeletal morphogenesis. BMC Dev Biol 11(1):6.
35. Ruff Y, et al. (2010) Glycodynamers: Dynamic polymers bearing oligosaccharidesresidues—generation, structure, physicochemical, component exchange, and lectinbinding properties. J Am Chem Soc 132(8):2573–2584.
36. Burke SD, Zhao Q, Schuster MC, Kiessling LL (2000) Synergistic formation of solublelectin clusters by a templated multivalent saccharide ligand. J Am Chem Soc 122(18):4518–4519.
37. Munoz EM, Correa J, Riguera R, Fernandez-Megia E (2013) Real-time evaluation ofbinding mechanisms in multivalent interactions: A surface plasmon resonance kineticapproach. J Am Chem Soc 135(16):5966–5969.
38. Ponader D, et al. (2014) Carbohydrate-lectin recognition of sequence-defined heter-omultivalent glycooligomers. J Am Chem Soc 136(5):2008–2016.
39. Grogan MJ, Pratt MR, Marcaurelle LA, Bertozzi CR (2002) Homogeneous glycopep-tides and glycoproteins for biological investigation. Annu Rev Biochem 71(1):593–634.
40. Kiessling LL, Splain RA (2010) Chemical approaches to glycobiology. Annu Rev Bio-chem 79:619–653.
41. Miura Y (2012) Design and synthesis of well-defined glycopolymers for the control ofbiological functionalities. Polym J 44(7):679–689.
42. Zhang Q, Haddleton DM (2013) Synthetic glycopolymers: Some recent developments.Adv Polym Sci 262:39–60.
43. Horan N, Yan L, Isobe H, Whitesides GM, Kahne D (1999) Nonstatistical binding of aprotein to clustered carbohydrates. Proc Natl Acad Sci USA 96(21):11782–11786.
44. Zhang S, et al. (2015) Dissecting molecular aspects of cell interactions using glyco-dendrimersomes with programmable glycan presentation and engineered humanlectins. Angew Chem Int Ed 54(13):4036–4040.
45. Percec V, et al. (2010) Self-assembly of Janus dendrimers into uniform dendrimersomesand other complex architectures. Science 328(5981):1009–1014.
46. Peterca M, Percec V, Leowanawat P, Bertin A (2011) Predicting the size and propertiesof dendrimersomes from the lamellar structure of their amphiphilic Janus den-drimers. J Am Chem Soc 133(50):20507–20520.
5590 | www.pnas.org/cgi/doi/10.1073/pnas.1506220112 Zhang et al.


















