
University of Groningen Behavioral and neuroimaging ... fileChapter 8: fMRI study in Parkinson’s...
33
University of Groningen Behavioral and neuroimaging studies on language processing in Dutch speakers with Parkinson's disease Colman, Katrien Suzanne François IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2011 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Colman, K. S. F. (2011). Behavioral and neuroimaging studies on language processing in Dutch speakers with Parkinson's disease. Groningen: s.n. Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 08-05-2019
Transcript of University of Groningen Behavioral and neuroimaging ... fileChapter 8: fMRI study in Parkinson’s...

University of Groningen
Behavioral and neuroimaging studies on language processing in Dutch speakers withParkinson's diseaseColman, Katrien Suzanne François
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2011
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Colman, K. S. F. (2011). Behavioral and neuroimaging studies on language processing in Dutch speakerswith Parkinson's disease. Groningen: s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 08-05-2019

Chapter 8: fMRI study in Parkinson’s Disease patients
121
Chapter 8 Differential activation of the neural language network in Parkinson’s disease patients compared to healthy subjects: an fMRI study
8.1 Introduction
As has been described in the introductory chapters of this thesis (see Chapter 1 and Chapter 2) Parkinson’ disease (PD) is a progressive neurodegenerative disease that results in degeneration of dopamine producing neurons in the substantia nigra pars compacta (SNc) and in the ventral tegmental area (VTA). Since the SNc and VTA neurons mainly project to the striatum, this cell degeneration leads to a reduction of dopamine in the striatum, which is a part of the basal ganglia (BG) complex. PD hinders the efficient functioning of the cortico‐striato‐cortical circuits, which connect the BG with the cerebral cortex (see Figures 2.2 and 2.3 in Chapter 2). Five different circuits have been described (Alexander, DeLong, & Strick, 1986), of which three incorporate non‐motor areas of the frontal cortex including the dorsolateral prefrontal cortex (DLPFC), the lateral orbitofrontal cortex, and the anterior cingulate (ACC)/medial orbitofrontal cortices. However, in anatomical studies of non‐human primates, all frontal regions examined have been found to receive inputs from BG thalamo‐cortical circuits (Middleton & Strick, 2002). It is therefore not unlikely that the left inferior frontal gyrus (LIFG), which is known to be involved in sentence processing, also forms part of a circuit connected with the BG that is used while, for example, comprehending a sentence (Ullman, 2006). Recently Snijders, Petersson, & Hagoort (2010) reported the results from a psychophysical interactions analysis, showing that the striatum was functionally connected to LIFG for sentence‐ambiguous versus sentence‐unambiguous conditions.
Accumulating evidence for the involvement of the cortico‐striato‐cortical circuits in sentence comprehension comes in part from studies in PD patients in whom these circuits are disturbed. The results of off‐line tasks such as spoken sentence to picture matching and grammaticality judgment demonstrated that non‐canonical structures such as sentences with object relative clauses and verbs in the passive voice were most vulnerable in individuals with PD (for a review see Chapter 3). Like non‐canonical sentences, sentences in the passive voice were found to be impaired in PD (Lieberman, et al., 1992). Moreover, problems with the passives in PD have also been described when comprehension of the passive voice was complicated by additional factors such as 1) length combined with sentence picture matching (see Chapter 5), 2) when real‐world knowledge could not be used to understand the sentence (Hochstadt, Nakano, Lieberman, & Friedman, 2006) or 3) when the passive sentences contained a center‐embedded relative clause compared to a final relative clause (Hochstadt et al., 2006). In a recent eye‐tracking experiment by Hochstadt (2009), some PD patients showed deviant eye blinking patterns on passive sentences. These

Chapter 8: fMRI study in Parkinson’s Disease patients
122
patients initially looked toward the reversed roles distracter item, which depicted the subject noun phrase (NP) as the ‘agent’, and not as the ‘patient’.
The results of studies on comprehension in PD patients have suggested that impaired executive control functions (e.g., Cools, Barker, Sahakian, & Robbins, 2001; Owen, Doyon, Dagher, Sadikot, & Evans, 1998) underlie the observed impaired comprehension of non‐canonical sentences (e.g., see Chapter 5; Colman, Koerts, Van Beilen, Leenders, & Bastiaanse, 2006; Geyer & Grossman, 1994; Grossman et al., 1993; Grossman, Carvell, Stern, Gollomp, & Hurtig, 1992a; Hochstadt, 2009; Hochstadt et al., 2006; Kemmerer, 1999). Specifically for Dutch passives, our correlation analyses suggested that set shifting and the digit span backward score correlated with PD patients’ impairments for the comprehension of Dutch passives (Chapter 5). These results were in line with Hochstadt et al., (2006) who reported for English that Reading Span and mean Voice Onset Time separation [related to articulatory rehearsal in working memory (WM)] were associated with passive sentence comprehension across a range of sentences.
According to Graybiel (1995a) the BG are “critical structures involved in action planning – both motor actions and mental actions” (p. 62). As many human behaviors, language is a sequencing time‐related phenomenon. It can thus be concluded that sequential processing is highly relevant to both motor and language processing. To comprehend a sentence, it is essential to determine ‘who did/is doing what to whom’. A hierarchical representation of the thematic roles (i.e., the argument structure) needs to be extracted from a serial input. In derived order sentences, such as passives, it can be more difficult to decode the syntactic structure and to assign the thematic roles (see Chapter 1 for a detailed discussion of word order in Dutch). Since PD patients are known to have problems with both the comprehension of passive sentences (Chapter 5; Hochstadt, 2009; Hochstadt et al., 2006; Lieberman et al., 1992) and to have impaired sequencing of actions (Godbout & Doyon, 2000) it was expected to find a correlation between complex sequencing abilities and the comprehension of syntactically more complex passive sentences in our off‐line sentence comprehension experiment (see Chapter 5). However, no association between the complex sequencing task and the comprehension of passive sentences was found. One explanation for the lack of an association between the complex sequencing task and the comprehension of passive sentences in PD is that a large number of participants were unable to perform the complex sequencing task, leading to too low a variance to measure the effects of sequencing ability. Therefore, in this functional Magnetic Resonance Imaging (fMRI) study, we aim at evaluating more directly the recruitment of the BG and inferior frontal gyrus (IFG) as parts of the cortico‐striato‐cortical circuits during the sequential processing of canonical sentences in which the sentence components are in base position (i.e., active sentences) as compared to non‐canonical sentences in which the components are in derived position (i.e., passive sentences).
The current literature suggests that in addition to the deficits in comprehension of non‐canonical sentences, verb production is impaired as part of the neurodegenerative process associated with PD (Bertella et al., 2002; Boulenger et al., 2008; Crescentini et al.; 2008; Péran et al., 2003). The evidence for verb processing impairments in PD has led to the hypothesis that dysfunction of the motor system has an impact on verb production (Péran et al., 2003, 2009). Péran et al. (2009) explored the relationship between the motor deficit in PD patients and brain activation in noun compared to verb generation tasks. They observed

Chapter 8: fMRI study in Parkinson’s Disease patients
123
a relationship between increased activation during task performance and increased motor‐frontal dysfunction in a network of cortical areas, including Broca's area. This increased activity in Broca's area was suggested to represent the increased mental simulation of action required to overcome the disturbed motor representations. In our own off‐line verb production study, the score on the verb production ability‐scale correlated significantly with set‐switching and WM. We suggested that because of failing automaticity, PD patients rely more on the cortically represented executive functions. This is in line with conclusions for receptive language impairments in PD. Unfortunately, due to the disturbed intimate relation between the BG and the frontal cortex, these executive functions are also dysfunctional. Thus, based on the literature reviewed in Chapter 3 and our own verb production study (see Chapter 4), we can conclude that dysfunctional cortico‐striato‐cortical circuits influence verb processing and that the BG, are critical in verb processing too. Numerous verb generation studies have been conducted before (see review Chapter 3), but the current study is the first to investigate the neural activation of verb representations during sentence comprehension in PD using fMRI.
As described in Chapter 6, language‐related Event Related Potentials (ERP) components have been identified upon the manipulation of both semantic or syntactic aspects of sentence components (for a detailed overview see Kutas & Van Petten, 1994; Osterhout & Holcomb, 1995). The model of comprehension by Friederici (2002) states that early access to lexico‐syntactic structure (early left anterior negativity or ELAN) is distinguished from later processing of thematic roles and semantics (N400) and a terminal step of syntactic integration and re‐processing (P600).20 PD or lesions to the BG influence these language‐related ERP components (Friederici, Von Cramon & Kotz, 1999; Friederici et al., 2003; Kotz et al., 2002, 2003, see also Chapters 6 and 7). The violation studies by Friederici and colleagues in PD (and BG‐lesioned) patients showed an intact ELAN, but a reduced P600 in response to word category errors [e.g., *Die Hose wurde am gebügelt/ ‘*The pants was on ironed’ vs. Die Bluse wurde am Freitag gebügelt/ ‘The blouse was on Friday ironed’21]. Reduction of the P600 effect in patients with PD was seen as evidence for the involvement of the BG in late, controlled syntactic integration processes, rather than early automatic structure building processes (Friederici et al., 1999, 2003; Kotz et al., 2002, 2003). However, the P600 effect has not only been reported in response to word category violations, but also for a range of syntactic violations (Friederici et al., 1996; Gunter, Friederici, & Schriefers; 2000; Hagoort et al, 1993; Neville et al., 1991; Osterhout, Holcomb, & Swinney, 1994; Osterhout & Mobley, 1995; Osterhout, McKinnon, Bersick, & Corey, 1996; Osterhout, 1997; Osterhout, Bersick, & McLaughlin, 1997; Vos, Gunter, Kolk, & Mulder, 2001) and has in addition been observed in sentences when a preferred syntactic analysis can no longer be maintained, like in syntactic ambiguous sentences (Friederici, Hahne, & Mecklinger, 1996; Frisch, Schlesewsky, Saddy, & Alpermann, 2002; Kaan & Swaab, 2003; Mecklinger, Schriefers, Steinhauer, & Friederici, 1995; Osterhout & Holcomb, 1992, 1993), or when syntactic complexity is increased (Friederici, Hahne, & Saddy, 2002; Kaan, Harris, Gibson, & Holcomb, 2000; Kaan & Swaab, 2003). Regardless of its interpretation, the P600 is elicited exclusively in sentence contexts 20 Important to note is that the qualitatively distinction of syntactic ((E)LAN/P600) versus semantic (N400) processing has recently been shown to be untenable (for reviews see Bornkessel‐Schlesewsky & Schlesewsky, 2008 and Kuperberg, 2007). The serial models of language processing and the constraint‐based models have different interpretations of the P600, but this discussion is beyond the scope of this thesis (see for a review Kuperberg, 2007). 21 A literal English translation of the sentence is given.

Chapter 8: fMRI study in Parkinson’s Disease patients
124
and must thus be seen as a function of expectancy and integration of semantic and syntactic information (Kotz, Schwartze, & Schmidt‐Kassow, 2009). Of great importance for our research is the fact that all of the linguistic phenomena that elicit a P600 are known to be impaired in PD patients (see Chapter 6 for a more detailed discussion of the ERP components).
In order to specify more PD patients’ impaired underlying pattern of neurological activation, we used two different grammatical violation types that are supposed to elicit a P600. The first type was a violation of the morphosyntactic relation between the auxiliary and past participle of the main verb as illustrated in the Dutch sentence in (1)22:
(1) *De juwelier heeft de poelier tipt ‘*The jeweler has the poulterer tips off’
Morphosyntactic violations such as inflectional violation elicit a LAN effect occurring between 300 and 500 msec (Gross et al., 1998; Gunter et al., 2000; Hahne & Jescheniak, 2001; Penke et al., 1997; Vos et al., 2001). Based on the functional interpretation of the P600, it is reasonable to assume that a violated past participle cannot be integrated into the sentence structure that is initially built up during parsing and, as such, also elicits a P600 effect.
The second type of violation used in the present study was a violation of the verb‐argument structure that violates both the syntactic and semantic aspects of sentence processing. The Dutch example in (2) illustrates that the intransitive verb proesten ‘to snort’ can only entail one argument, an agent, and thus cannot take a patient.
(2) *De juwelier heeft de poelier geproest ‘*The jeweler has the poulterer snorted’
Friederici and Frisch (2000) found a biphasic N400‐P600 pattern for this type of violations in active sentences. The authors argued that the N400 reflected the semantic/thematic problems that arise when an argument NP is not provided with a thematic role by the verb, whereas the P600 was seen as reflecting the fact that the sentence structure (transitive) is not licensed by the (intransitive) verb.
In the first part of the fMRI project (Chapter 7), we investigated only healthy subjects and reported evidence for the contributions of the IFG and BG in comprehending written sentence materials in which variables of canonicity (i.e., word order) and grammaticality were factorially manipulated. The combination of region of interest (ROI) analyses in subparts of the BG and IFG and a factorial whole brain analysis revealed a main effect of word order in the right pallidum [and to a lesser extent in the putamen (Put)], in which more activation was elicited by active sentences than passive sentences. It was suggested that this output nucleus supported facilitation of heuristic routines in the comprehension of healthy subjects. The factorial whole brain analysis showed increased activation in two small clusters of voxels situated in the right Put and the lateral aspect of the left superior frontal gyrus (SFG, BA 6, Montreal Neurological Institute (MNI) coordinates ‐20, 6, 62) for active sentences
22 See introduction of chapter 7 for more details on the construction of the two used violation types, namely the inflectional violation and the violation of the verb‐argument structure.

Chapter 8: fMRI study in Parkinson’s Disease patients
125
versus passives. This activation might be related to the processing of basic action order representation in active sentences. Our findings that the effect of grammaticality activated the left middle temporal gyrus (MTG, Brodmann Area (BA) 21/39) in healthy subjects is consistent with the suggestion of Hickok and Poeppel (2005, 2007), who claimed that the left posterior MTG (BA 21) stores lexical representations. Furthermore, the activation in the right parietal cortex might be related to short‐term storage of this semantic information and is related to the N400 effect over the parietal cortex following errors of semantic integration during ERP studies. The pars opercularis (BA 44) and the pars triangularis (BA 45) of the IFG showed similar interaction effects between grammaticality and canonicity. For the right BA 45 and to a lesser extent the pars orbitalis of the IFG (BA 47), however, a main effect of grammaticality was found. Possibly, these areas are involved in a semantic processing strategy used by the participants.
These data indicate that the BG play a role in processing word order and that the RH is prominently activated in elderly non‐brain‐damaged subjects (NBDS). Additionally, simultaneous activation of areas in the fronto‐parieto‐temporal cortex was explained as being supportive to these activations during silent reading of sentences for comprehension. The literature discussed in this introduction on PD patients’ impairments in language processing and our data in NBDS give us a solid basis to formulate the central research question of the second part of our fMRI project: what are the consequences of PD patients’ striatal dopamine depletion on the patterns of activation during the comprehension of sentences in which canonicity and grammaticality were factorially manipulated?
We formulated here three predictions regarding activation patterns in PD patients. First, since past research indicated that PD patients have problems with sequencing of actions (e.g., Godbout & Doyon, 2000) it is not expected to find a normal pattern of word order (i.e., canonicity) in the right BG of PD patients during the comprehension of active sentences. In our previous report on the results of healthy subjects, activation for actives versus passives was found in the right Put and right pallidum or globus pallidus (GP). As Owen, Doyon, Dagher, Sadikot and Evans (1998) proposed, PD patients’ impairments in cognition are related to disturbed BG outflow. Consequently, impaired cognitive functions in PD are explained as being the result of the disturbed BG outflow of the right GP, which has in turn an impact on information transfer to the frontal cortex. The PD patients in this study will not be able to detect word order information in sentence context and as a result, PD patients will not show activation in the right globus pallidus during the processing of active sentences compared to passive sentences and this is in contrast to what was previously found for the NBDS (Chapter 7). Moreover, if the deficit of PD patients is located in the detection of word order in sentence context, these patients should demonstrate a different activation pattern compared to healthy control (HC) subjects due to different processing strategies while processing the derived order sentences (i.e., the non‐canonical passive sentences). In the literature a strong hypothesis exists on the involvement of the left pars opercularis of the IFG (BA 44) in grammatical complexity (Friederici, Opitz, & Von Cramon, 2000; Kuperberg et al., 2003). Second, concerning the processing of violations in PD we only expect to find increased activation for the inflectional violations, since these violations can be detected in a superficial way during the highly automatic first‐pass parsing processes solely based on the striking visual characteristics of the verb segment, namely word length. See for an example of the inflectional violation the Dutch sentence in (1).

Chapter 8: fMRI study in Parkinson’s Disease patients
126
Finally, only one fMRI study by Grossman et al. (2003) evaluated sentence comprehension in PD patients compared to HC subjects. In line with the findings of Grossman et al. (2003), it was expected to find compensatory cortical activation in the PD group when comparing the groups directly.
8.2 Methods
8.2.1 Subjects
Sixteen idiopathic PD patients participated in this fMRI experiment, but one patient had to be excluded due to movement artifacts and hence the analyses were done over 15 subjects. Relevant medical and demographic information for this group is given in Table 8.1. PD patients were diagnosed according to the criteria of the United Kingdom Parkinson’s Disease Society Brain Bank. Right before entering the scanner, the PD patients were assessed with the Unified Parkinson’s Disease Rating Scale, part III (UPDRS; Fahn & Elton, 1987, see appendix B). The UPDRS score was used to estimate the Hoehn and Yahr stage (H&Y stage, Hoehn & Yahr, 1967, see appendix A). According to the H&Y disability staging criteria, the motor disability of PD patients included in this study ranged from mild to moderate. PD patients were assessed while they were on medication. A Levodopa Equivalent Daily Dose score (LEDD‐score) was obtained for each individual patient. The LEDD‐score was calculated according to the formula: regular Levodopa dose x 1 + slow release Levodopa x .75 + bromocriptine x 10 + apomorphine x 10 + ropinirole x 20 + pergolide x 100 + pramipexole x 100 + [regular Levodopa dose + (slow release Levodopa x .75)] x .2 if taking entacapone (Esselink et al., 2004).
The PD patients were compared with 15 HC participants. The healthy elderly subjects in this experiment were the same 15 participants described in the experiment described in Chapter 7 of this thesis. The characteristics of the HC subjects and PD patients are summarized in Table 8.1. One HC subject out of 16 scanned persons was excluded from further analyses due to movement artifacts. All HC subjects were recruited from the Groningen community and were matched on age (t(28)= 1.353, p > .05) and education (t(28)= ‐.235, p > .05) with the PD patients (see Table 8.1). HC subjects were screened for a history of neurological or psychiatric conditions prior to inclusion to the study and only subjects with no such history were included.
All participants were also assessed for the presence of dementia and depression. Exclusion criteria were dementia (Mini‐Mental State Examination (MMSE) < 25; Folstein, Folstein, & McHugh, 1975) and depression (Montgomery‐Åsberg Depression Rating Scale (MADRS); Montgomery & Åsberg, 1979) ≥ 18 (Leentjens et al., 2000). Patients and HC were all native speakers of Dutch, who reported no premorbid language difficulties (such as dyslexia) and had self‐reported normal or corrected‐to‐normal vision and hearing. Handedness was determined using the Dutch version of the Edinburgh Handedness Inventory (Van Strien, 1992). All participants were strongly right‐handed (PD: mean = 9.8, SD = .56; HC: mean = 10).
Chapter 8: fMRI study in Parkinson’s Disease patients
127
The Medical Ethical Committee of the University Medical Center Groningen (UMCG) approved this study. After explanation of the protocol, all participants gave their written informed consent according to the Declaration of Helsinki. The participants were checked for absence of bodily ferromagnetic materials and other MRI incompatibility. During scanning, the standard MRI safety regulations were followed.
Table 8.1: Demographic and clinical features [Mean (SD)] of the PD patients and HC subjects.
PD patients (n= 15)
HC subjects (n= 15)
Gender (M:F) 10:5 9:6 Handedness score 9.8 (.56) 10 Age in years 61.73 (8.75) 57.33 (9.06) Education in years 15 (4.74) 15.4 (4,58) MMSE score 28 (1) 28.33 (.72) MADRS score 5.47 (2.3) 4.2 (2.54) Trail index 1.94 (.46) 2.02 (.51) Reading Span score 56.15 (11.01) 57.43 (9.16) UPDRS part III 22 (9.81) n/a H&Y staging 2.07 (.73) n/a LEDD‐score 575.61 (291.61) n/a
Note: See for the abbreviations the List of Abbreviations.
8.2.2 The fMRI experiment
8.2.2.1 Materials and methods
Examples of the six critical sentence conditions for the present study are shown in Table 8.2. The construction of the sentence materials was described in detail in the Materials section in Chapter 7. The procedures used for stimulus presentation and the MRI characteristics are the same as in the event related fMRI design conducted in a group with healthy elderly subjects. For details on these procedures and the MRI characteristics, see the section on Experimental procedure and Image acquisition of Chapter 7.

Chapter 8: fMRI study in Parkinson’s Disease patients
128
Table 8.2: Examples of the experimental materials in Dutch
Grammaticality Canonicity
Active Passive
N De juwelier/ heeft/ de poelier/ getipt. The jeweler has the poulterer tipped off.
De pater/ wordt door/ de militair/ getipt. The father is by the soldier tipped off.
I De juwelier/ heeft/ de poelier/ tipt. The jeweler has the poulterer tips off.
De pater/ wordt door/ de militair/ tipt. The father is by the soldier tips off.
VA De juwelier/ heeft/ de poelier/ geproest. The jeweler has the poulterer snorted.
De pater/ wordt door/ de militair/ geproest. The father is by the soldier snorted.
CS Vm gthsv/ kcrtf/ pg btcpkh/ bcpfhsvhn.
Vm mglbsfv/ vsntf hmcg/ pg vbntjsg/ kjgpfvbgsd.
Note: The accompanying literal English translation is given. Each segment border is indicated with a vertical line. For the abbreviations see the List of Abbreviations.
Overall, in the experiment two within‐subject factors and a between‐subject factor of group (PD patients vs. HC subjects) were crossed in a 2x2x3 factorial design resulting in six experimental conditions. The first within‐subject factor was canonicity with two levels: active vs. passive voice. The second within subject factor was grammaticality with three levels: no violation (N), inflectional violation (I) and verb‐argument structure violation (VA) condition. All subjects read one out of three experimental lists, containing per list a total of 240 sentences with 40 sentences in each experimental category, 120 Consonant Strings (CS) and 90 filler sentences to balance the amount of correct and violated sentences.

Chapter 8: fMRI study in Parkinson’s Disease patients
129
8.2.2.2 Data analyses
The fMRI data analyses up until the statistics (file conversion, preprocessing and modeling) were identical to those used in the mixed block/event‐related fMRI design described in Chapter 7.
Regions of interest analysis
On the basis of the hypothesis that the frontal cortex and the BG are differently involved in sentence processing, at least with regard to the processing of violations, we analyzed six ROIs, in both the left and right hemisphere (LH and RH respectively):
1. Caudate nucleus (Caud) 2. Putamen (Put) 3. Globus pallidus (GP) or Pallidum 4. BA 44 or pars opercularis of the IFG 5. BA 45 or pars triangularis of the IFG 6. BA 47 or pars orbitalis of the IFG
We have chosen to subdivide the BG complex and the IFG into subparts in order not to miss significant effects that are limited to one subarea. Moreover, although many cognitive studies focused on the striatum, involvement of the BG in cognitive functioning is not limited to the striatum. According to Owen et al. (1998), the cognitive impairments in PD are due to disturbed BG outflow, particularly of the right GP, which has in turn an impact on the information transfer to the frontal cortex. Therefore, we chose to include both input nuclei (i.e., Caud and Put) and principal output nuclei (i.e., GP) of the cortico‐striato‐cortical circuit in our ROI analysis. In order to investigate whether distinct subregions within the IFG contribute differentially to sentence processing, all three IFG subregions were included in the anatomical ROI analysis.
ROI definition and signal extraction was performed using MARSBAR (Brett et al., 2002). These ROIs were defined anatomically using the Anatomical Automatic Labeling (AAL) map of the MNI brain (Tzourio‐Mazoyer et al., 2002). In addition, three extra functional ROIs were defined within the LH. On the basis of our previous findings in a group of elderly subjects (Chapter 7), we had the a priori hypothesis that additional areas in the superior precentral gyrus (BA 6) and the superior and middle temporal gyri (BA 22 and BA 21 respectively) would be involved in comprehension of our sentence materials. These functional ROIs were defined using MARSBAR (Brett et al., 2002), based on the effects of interest activation during reading of the complete sentences compared with looking at the CS (p < .05, False Discovery Rate, FDR).
For all the ROIs, average activation values per condition were extracted for each participant. Before proceeding with further statistical analyses, the necessary assumptions to use repeated measures ANOVA were evaluated. First, the effects of heterogeneous variability of the data were removed, by transforming the raw data extracted for each anatomical and functional ROI into z‐values. Secondly, normality of the distribution of the z‐values was analyzed with the Kolmogorov‐Smirnov Test implemented in the statistical Package for the Social Sciences (SPSS 16.0). This was done separately for each ROI area, within groups, and for each separate condition. All variables were found to be normally distributed. Finally, we

Chapter 8: fMRI study in Parkinson’s Disease patients
130
tested for violations of sphericity, using Mauchly’s Test of Sphericity. If the assumption of sphericity was violated, the output for the Greenhouse‐Geisser correction factor was used. Using SPSS, for the anatomical ROIs, a series of repeated measures (2x2x2x3) ANOVAs with group (PD patients vs. HC subjects) as a between‐subject factor and the factors hemisphere (LH vs. RH), canonicity (active vs. passive) and grammaticality (no violation, inflectional violation and verb‐argument structure violation) were performed. For the functional ROIs a similar analysis was conducted. However, since the functional ROIs were all located in the LH, the within‐subject factor of hemisphere was not included.
Whole brain analysis
In addition to the ROI analysis, we conducted a whole brain analysis. At the first level, individual subject analyses specifying contrasts between reading of the full sentences and viewing whole CS were calculated. The contrast (difference in β) images of the first‐level analysis were then used for the second‐level group statistics, in which a one sample t‐test was used to determine the areas that responded more strongly to sentences as compared to looking at CS. The statistical parametric maps (SPM {T14}) were thresholded at T ≥ 3.79 (p < .001, uncorrected) with a cluster extent threshold of 20. Only supra‐threshold clusters of voxels that survived at an α level of p < .05, corrected for multiple comparisons, are reported.
The contrast images of the individual‐level analysis were also used for the second‐level group statistics (two sample t‐test) to reveal where HC subjects had more activation than PD patients and vice versa. The statistical parametric maps (SPM {T}) were thresholded at T > 3.41 at the voxel level (p < .001, uncorrected). Only supra‐threshold clusters of voxels that survived at p < .05, corrected for multiple comparisons, are reported. Furthermore, using a mask we revealed also the activation that the PD patients had in common with the HC subjects. The mask included only activated voxels in the HC group that were at a p < .01 level, uncorrected for multiple comparisons (T > 2.62), and in a cluster that exceeded a threshold of 300 voxels to avoid spike activations in each contrast.
A within‐subjects factorial design matrix was employed to test the main effect of canonicity, the main effect of grammaticality and their interaction. First for each participant, at the first level, six contrasts were generated by contrasting the activation of the critical verb segment of each of the six conditions with the activation of viewing the fourth segment of the CS. The main effect of canonicity (2 levels) was tested using a t‐test. An F‐test was used for the main effect of grammaticality (3 levels) and for the interaction effect. In general, the maxima of suprathreshold regions are localized and reported in MNI coordinates as used in SPM5. Brain regions are identified using MRIcron software (www.sph.sc.edu/comd/rorden/mricron). An anatomical description and the corresponding BAs are also reported.

Chapter 8: fMRI study in Parkinson’s Disease patients
131
8.2.3 Behavioral and cognitive testing
After the scanning procedure, all participants were tested with a violation detection task (see also Chapter 7). In addition, set‐switching abilities and WM were assessed. In the following, the violation detection, the behavioral and cognitive tests are discussed.
8.2.3.1 Violation detection task
Each subject judged a list consisting of 90 sentences, 36 of which are test sentences (six of each of the six experimental conditions). The participants were assigned to one of the two lists of items that she/he had not seen in the fMRIscanner. Experimental presentation and response collection was conducted using the E‐prime software (Psychology Software Tools Inc., 2001). The participants were instructed to read the written sentences on the screen of a laptop and to judge whether the presented sentence made sense. They were asked to press a red button on the keyboard of a laptop as soon as a sentence stopped making sense. They were asked to press a green button on the keyboard at a sentence final prompt (a question mark) if no error was encountered. Participants were asked to respond as quickly and accurately as possible when a violation was encountered and to wait for the question mark to affirm that the sentence was correct. They started each new trial by pressing the space bar. Prior to the behavioral experiment, a practice session was conducted on correct and unrelated violated sentences.
To analyze the accuracy data, a repeated measures ANOVA was performed, including the between‐subject factor group (PD patients vs. HC subjects) and the within‐subject factors canonicity (active vs. passive) and grammaticality (no violation, inflectional violation and verb‐argument structure violation). Individual mean Reaction Times (RTs) per condition were obtained to calculate group means per condition. Unfortunately, too many subjects, especially in the PD group, failed to react within the allowed response time, resulting in unreliable mean RT’s. For this reason, no further non‐parametric analysis was performed.
8.2.3.2 Cognitive set‐switching
The Trail Making Test parts A and B (TMT A & B; Reitan, 1992) were administered to assess cognitive set‐switching. The target measure of the Trailmaking was the performance on part B corrected for psychomotor speed (by dividing it by the performance on part A), the B/A index.
8.2.3.3 Verbal working memory
A computerized version of the Reading Span Task in Dutch (Van den Noort et al., 2005) was used in order to measure both the reading span and the RTs. The test started with an instruction on the screen and two practice trials after which the experimental material was presented. The participants completed five series of 20 sentences (100 sentences in total), that were presented in different set sizes (2, 3, 4, 5 or 6 sentences) in random order. The participants were instructed to read the sentences aloud as fast as possible and subsequently press the space bar, to start the next sentence. The word ‘herinner’ (recall) was presented, to trigger the participant to recall the last word of each sentence in the set. The order of recall was free. The total number of remembered words were collected.

Chapter 8: fMRI study in Parkinson’s Disease patients
132
8.3 Results
8.3.1 Behavioral and cognitive testing
8.3.1.1 Cognitive set‐switching
One of the HC subjects did not understand the instructions for the task and was therefore excluded from the analyses. As can be seen in Table 8.1, HC subjects had a mean Trail index of 2.02 (.51) and the PD patients had a mean Trail index of 1.94 (.46). These scores did not differ (t(27) = ‐.446, p > .05).
8.3.1.2 Verbal working memory
The Reading Span Task (Van den Noort et al., 2005) was aborted in one HC subject and two PD patients due to communicated frustration. The results of these subjects were not included in the analyses. As summarized in Table 8.1, the PD patients remembered on average 56.15 (11.01) words in total. The HC subjects remembered on average 57.43 (9.16) words in total; the difference between the groups was not significant (t(25) = ‐.328, p > .05).
8.3.1.3 Violation detection task
The descriptive data of the accuracy and RTs for the groups are shown in Table 8.3. The repeated measures ANOVA for the accuracy data revealed no significant effects. For the latency data, including the PD patients’ performance, only the descriptive data are given, since not enough valid data were available in the PD group to process further statistics.
Table 8.3: Mean (SD) accuracy and latency to respond to stimuli during behavioral task for the PD patients and HC subjects
PD patients HC subjects
Sentence condition
Mean (SD) % correct
Mean (SD) msec latency
Mean (SD) % correct
Mean (SD) msec latency
AN 83.33 (24.46) 846.33 (280.21) 94.44 (12.06) 568.08 (138.48) PN 78.57 (26.50) 884.14 (431.61) 91.11 (8.61) 713.25 (366.16) AI 88.10 (17.82) 1311.08 (366.11) 92.22 (15.26) 989.58 (334.82) PI 90.48 (12.60) 1267.29 (180.89) 95.56 (11.73) 966.83 (322.44) A‐VA 86.90 (21,86) 1671.93 (491.80) 84.44 (22.24) 1169.25 (474.35) P‐VA 84.52 (20.11) 1439.67 (128.55) 85.56 (30.12) 1485.17 (80.14)
Note: See for the abbreviations the List of Abbreviations.
In the HC group, the detection of a verb‐argument structure violation in a passive sentence requires the most time [1485.17 (80.14)], but the active sentences without a violation are understood relatively quickly [568.08 (138.48)]. The available RT data of the PD patients, however, revealed slower responses when processing active sentences with a verb‐argument structure violation [1671.93 (491.80)], and similar to the HC subjects the fastest for active sentences without a violation [846.33 (280.21)].

Chapter 8: fMRI study in Parkinson’s Disease patients
133
8.3.2 Functional Imaging results
8.3.2.1 ROI based analyses
Anatomical ROI analyses
For each ROI an ANOVA was calculated separately, including the between‐subject factor group (PD patients vs. HC subjects) and the within‐subjects factors hemisphere (LH vs. RH), canonicity (active vs. passive) and grammaticality (no violation, inflectional violation and verb‐argument structure violation). In the following, we will discuss the results for the BG and the IFG regions in separate sections.
Basal ganglia
As predicted, the results revealed no significant effects for the Caud, which is consistent with what was found in our previous report on HC subjects in Chapter 7.
Based on the elderly HC subject data, there is a hypothesis that there should be an effect of canonicity in the right BG (pallidum and to a lesser extent Put ), since the factorial whole brain analysis revealed an increased activation for active sentences compared to passive sentences in these subregions of the BG.
For the Put, when the groups were analyzed together, a significant interaction hemisphere x canonicity (F(1, 28) = 8.756; p < .01), was revealed which means that the canonicity effect is different in the LH and RH. In addition, the Put also showed a marginal interaction of hemisphere x canonicity x group (F(1, 28) = 3.733; p = .064) and of hemisphere x grammaticality x group (F(2, 56) = 2.685; p = .077). Furthermore, we looked at the main effects and interaction for each hemisphere separately. The ANOVA for the right Put revealed a trend to a main effect of canonicity (F(1, 28) = 2.929; p = .098). Consistent with the previous findings, the processing of active sentences activated the right Put more than processing of the passive sentences. However, in contrast to our expectations, no effect of group was found in the right Put , which is against our hypothesis on a reduced canonicity effect in PD patients. However, the left Put showed a significant interaction for grammaticality x group (F(2, 56) = 3.962; p < .05), suggesting that each group activated the left Put differently according to the type of violation. As illustrated in Figure 8.1 the inflectional violated sentences activated the left Put more in the PD patients.

Chapter 8: fMRI study in Parkinson’s Disease patients
134
Figure 8.1: Left Putamen: mean activation in HC and PD subjects for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean.
For the pallidum or GP, a main effect of grammaticality (F(2, 56) = 3.562; p < .05) was evident. Also an interaction of canonicity x group (F(1, 28) = 7.056; p < .05) was present, suggesting that the groups activated the pallidum differently for active compared to passive sentences. In addition, a trend to a hemisphere x group effect (F(1, 28) = 3.679; p = .065) and a trend to a hemisphere x grammaticality effect (F(2, 56) = 3.059; p = .055) was found. To further explore these effects in the PD group, an ANOVA with factors hemisphere x canonicity x grammaticality was conducted in this group. In our previous ROI analysis for the HC subjects, a main effect of canonicity and an interaction effect of hemisphere x canonicity was found. Conversely, for the PD patients, a main effect of grammaticality was found (F(2, 28) = 4.141; p < .05). In addition, a trend to a hemisphere x grammaticality interaction (F(2, 28) = 2.691; p = .085) and a marginal trend to a canonicity x grammaticality interaction (F(2, 28) = 2.596; p = .092) was found. Figure 8.2 illustrates the grammaticality effect in the PD patients’ bilateral pallidum, showing more activation for the sentences containing an inflectional violation compared to the other types of sentences.

Chapter 8: fMRI study in Parkinson’s Disease patients
135
Figure 8.2: Left and right pallidum: mean activation in HC and PD subjects for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean.
Inferior frontal gyrus
For the IFG pars opercularis (BA 44), a main effect of grammaticality (F(2, 56) = 5.343; p < .01), an interaction effect of grammaticality x group (F(2, 56) = 8.277; p = .001), an interaction effect of canonicity x grammaticality x group (F(2, 56) = 4.188; p < .05) and an interaction effect of hemisphere x canonicity x grammaticality (F(2, 56) = 3.677; p < .05) were found. For the elderly HC subjects an interaction effect of canonicity x grammaticality was already demonstrated (see Chapter 7) and was attributed to the high activation of the active sentences with no violation (AN) in the IFG pars opercularis (BA 44). In contradiction to this finding, the PD group evidenced a strong main effect of grammaticality (F(2, 28) = 16.962; p < .001), providing further evidence that the PD patients activated bilaterally the IFG pars opercularis to read inflectional violated sentences, independent of the factor of canonicity (see Figure 8.3).
Based on the described hemisphere interaction above, we also looked at the main and interaction effects for each hemisphere separately. In the LH a complex pattern of results was evident: a main effect of grammaticality (F(2, 56) = 6.148; p < .01), an interaction of grammaticality x group (F(2, 56) = 6.377; p < .01), an interaction of canonicity x grammaticality x group (F(2, 56) = 3.833; p < .05) and a trend to an interaction of canonicity x grammaticality (F(2, 56) = 2.806; p = .069). The PD patients showed clearly increased activation for inflectional violations (independent of canonicity) while the HC subjects showed increased activation for the non‐violated sentences only in the passive condition. For the RH, an interaction effect of

Chapter 8: fMRI study in Parkinson’s Disease patients
136
grammaticality x group (F(2, 56) = 5.249; p < .01) and also a trend to a main effect of grammaticality was present (F(2, 56) = 2.789; p = .070). Figure 8.3 demonstrates that the PD patients showed similar patterns in the left and right BA 44, which was not seen in the HC group.
Figure 8.3: Left and right IFG pars opercularis (BA 44): mean activation in HC and PD subjects for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean. Note: Abbreviations not mentioned in the text: LNoViol; left no violation, LInflViol; left inflectional violation; LArgViol, left verb‐argument violation; RNoViol; right no violation, RInflViol; right inflectional violation; RArgViol, right verb‐argument violation.
For the more anterior IFG pars triangularis (BA 45), the global ANOVA showed a main effect of grammaticality (F(2, 56) = 3.407; p < .05), an interaction effect of grammaticality x group (F(2, 56) = 10.272; p = .000) and an interaction of hemisphere x grammaticality x group (F(2, 56) = 3.198; p < .05). Finally, also an interaction canonicity x grammaticality (F(2, 56) = 3.269; p < .05) was evident. The post hoc repeated measure ANOVAs performed for each group separately exposed for the HC subjects, a trend to a main effect of grammaticality (F(2, 28) = 3.186; p = .057), a significant interaction hemisphere x grammaticality (F(2, 28) = 3.501; p < .05) and an interaction canonicity x grammaticality (F(2, 28) = 4.567; p < .05) (see also Chapter 7). For the PD group (F(2, 28) = 11.208; p < .001) only a significant main effect of grammaticality was evident. Thus, comparable to what was found for the pars opercularis of the IFG (BA 44), the PD patients showed a significant main effect of grammaticality, attributable to the stronger activation for the inflectional violation condition.
Chapter 8: fMRI study in Parkinson’s Disease patients
137
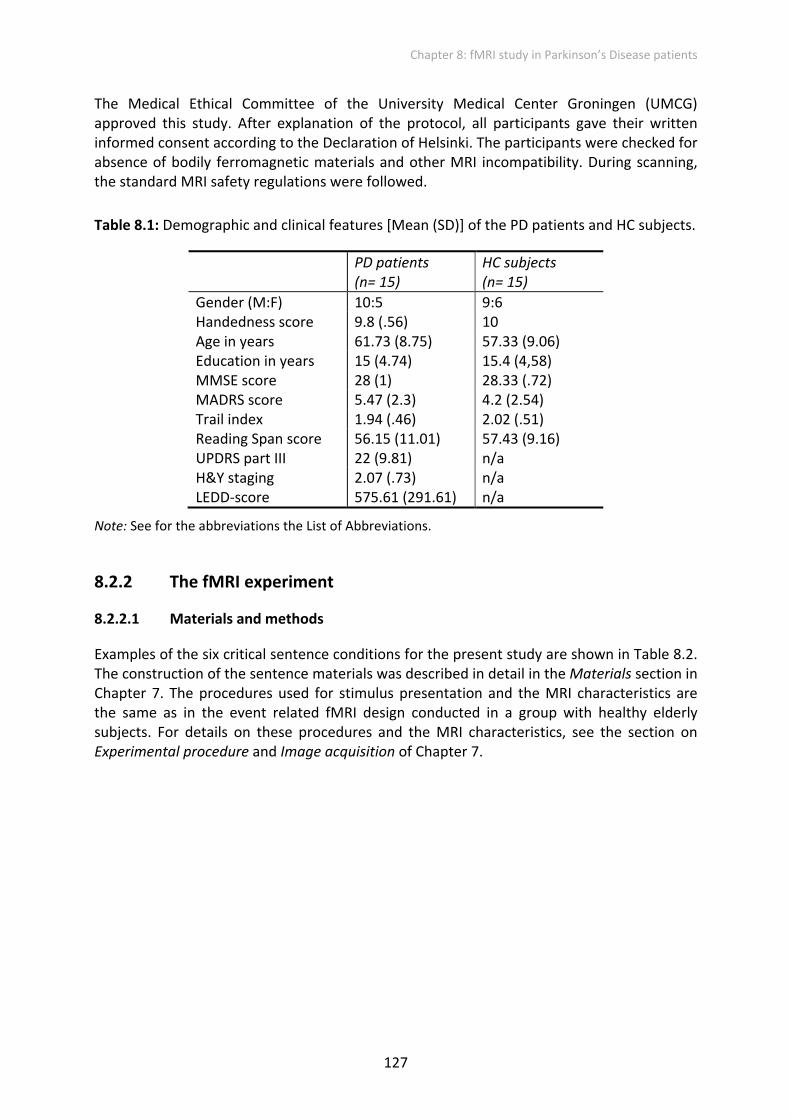
Furthermore, the ANOVAs for the left and right IFG pars triangularis separately with factors canonicity x grammaticality x group revealed a significant grammaticality x group interaction for the LH only (F(2, 56) = 5.924; p < .01). For the RH a main effect of grammaticality (F(2, 56) = 4.506; p < .05) and an interaction of grammaticality x group (F(2, 56) = 11.664; p < .001) was found. Figure 8.4 illustrates that the groups revealed a different pattern of activation depending on the grammaticality type. Furthermore, Figure 8.4 also demonstrates that the PD patients showed a similar pattern of activation in the right and left IFG pars triangularis. For the HC subjects the pattern is less consistent between the two hemispheres.
Figure 8.4: Left and right pars triangularis of the IFG (BA 45): mean activation in HC and PD subjects for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean. Note: Abbreviations not mentioned in the text: LNoViol; left no violation, LInflViol; left inflectional violation; LArgViol, left verb‐argument violation; RNoViol; right no violation, RInflViol; right inflectional violation; RArgViol, right verb‐argument violation.
Finally, for the IFG pars orbitalis (BA 47) the ANOVA yielded a significant interaction effect for grammaticality x group (F(2, 56) = 5.512; p < .01).
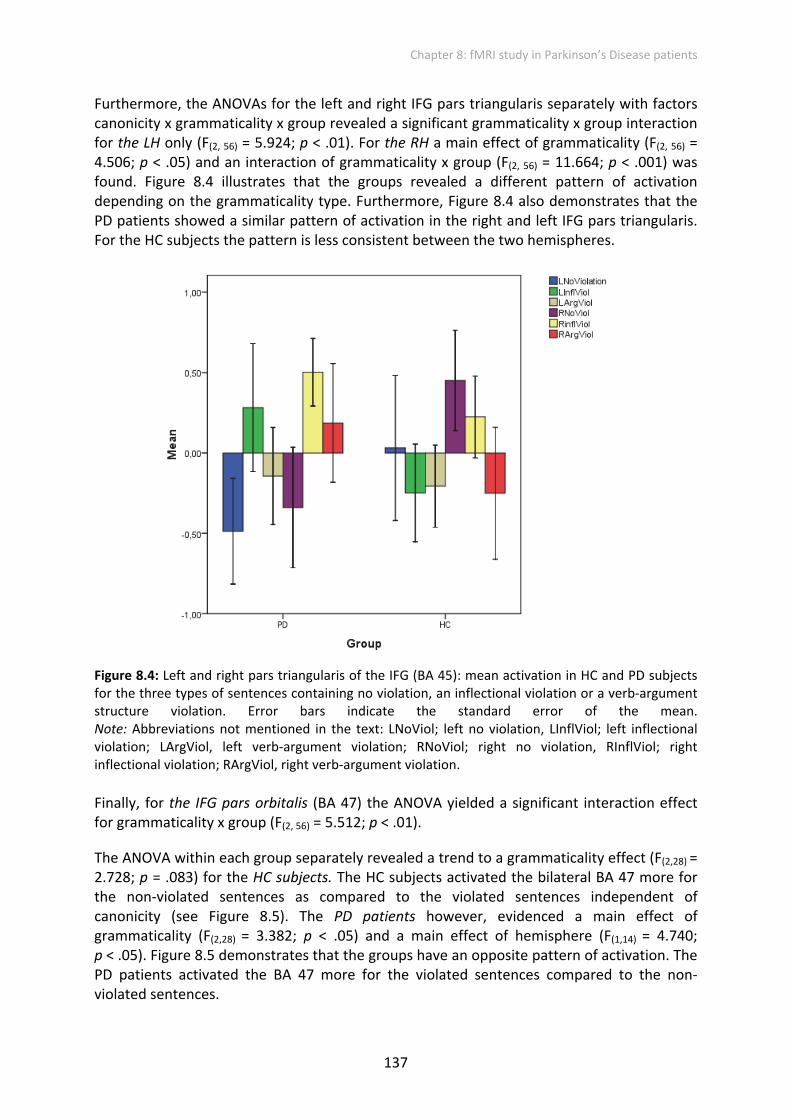
The ANOVA within each group separately revealed a trend to a grammaticality effect (F(2,28) = 2.728; p = .083) for the HC subjects. The HC subjects activated the bilateral BA 47 more for the non‐violated sentences as compared to the violated sentences independent of canonicity (see Figure 8.5). The PD patients however, evidenced a main effect of grammaticality (F(2,28) = 3.382; p < .05) and a main effect of hemisphere (F(1,14) = 4.740; p < .05). Figure 8.5 demonstrates that the groups have an opposite pattern of activation. The PD patients activated the BA 47 more for the violated sentences compared to the non‐violated sentences.
Chapter 8: fMRI study in Parkinson’s Disease patients
138
Figure 8.5: Bilateral pars orbitalis of the IFG (BA 47): mean activation in the in HC and PD subjects for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean.
Functional ROI analyses
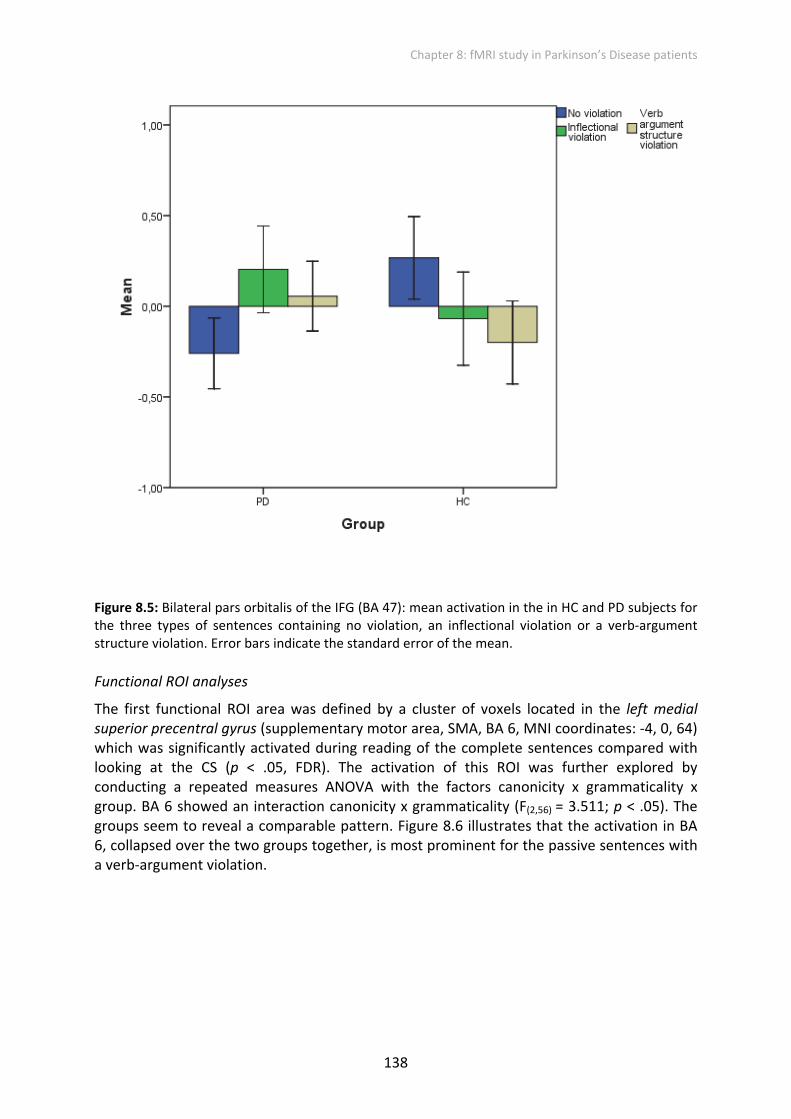
The first functional ROI area was defined by a cluster of voxels located in the left medial superior precentral gyrus (supplementary motor area, SMA, BA 6, MNI coordinates: ‐4, 0, 64) which was significantly activated during reading of the complete sentences compared with looking at the CS (p < .05, FDR). The activation of this ROI was further explored by conducting a repeated measures ANOVA with the factors canonicity x grammaticality x group. BA 6 showed an interaction canonicity x grammaticality (F(2,56) = 3.511; p < .05). The groups seem to reveal a comparable pattern. Figure 8.6 illustrates that the activation in BA 6, collapsed over the two groups together, is most prominent for the passive sentences with a verb‐argument violation.

Chapter 8: fMRI study in Parkinson’s Disease patients
139
Figure 8.6: Left superior precentral gyrus (BA 6): mean activation in the in the HC and PD subjects together for the three types of sentences containing no violation, an inflectional violation or a verb‐argument structure violation. Error bars indicate the standard error of the mean. Note: See for abbreviations the List of Abbreviations.
The second functional area was also based on a cluster, which activated in response to sentence reading and was located in the left middle temporal gyrus (left MTG, BA 21, MNI coordinates: 48, ‐46, 12). The ANOVA yielded a main effect of canonicity (F(1,28) = 6.583; p < .05) with greater activation for passives and a trend to an interaction of canonicity x group (F(1,28) = 3.692; p = .065). The HC subjects activated this area to process the passive sentences, a pattern that is not visible in the PD group (see Figure 8.7).

Chapter 8: fMRI study in Parkinson’s Disease patients
140
Figure 8.7: Left middle temporal gyrus (BA 21): mean activation in the in HC and PD subjects for all sentence conditions. Error bars indicate the standard error of the mean. Note: See for abbreviations the List of Abbreviations.
The last functional area was located in the left superior temporal gyrus (left STG, BA 22). This area showed only a trend to an interaction for grammaticality x group (F(2,56) = 2.660; p = .079).
In summary, there are four main observations from the ROIs analyses to note and to discuss in more detail in the discussion section of this chapter.
First a main effect of canonicity was found in the pallidum of the HC group, but not for the PD group. In the HC group, the right pallidum was more activated for the active sentences compared to the passive sentences. At the same time, HC subjects recruited more the left MTG (BA 21) to read the passive sentences as compared to the active sentences, a pattern that is not evident in the PD group.
Second, our results indicate a prominent grammaticality effect in the PD patients, occurring in the pallidum, the left Put , the pars opercularis (BA 44), triangularis (BA 45) and orbitalis (BA 47) of the IFG. This pattern is mainly attributable to the stronger activation for the inflectional violation. In the HC subjects, the effect of grammaticality in these regions is not as prominent as in the PD patients.

Chapter 8: fMRI study in Parkinson’s Disease patients
141
Third, in the pars orbitalis of the IFG (BA 47) the two groups showed an opposite pattern of activation. PD patients relied more on this inferior frontal area for the violated sentences than for the non‐violated sentences. In contrast, the HC subjects activated more the IFG pars orbitalis (BA 47) for the non‐violated sentences compared to the violated sentences.
Finally, in the left BA 6, PD patients show a pattern similar to the HC subjects. BA 6 was the sole area where the effect of the verb‐argument structure violation in the passive sentences (P‐VA) was visible.
8.3.2.2 Whole Brain analyses
Ultimately, the goal of this study was to compare the activations found for PD patients compared to the activations of HC subjects during comprehension of written sentences. In the following, we will first describe the activations of the PD patients in response to the subtraction of looking at the CS from reading of the sentences (whole sentence duration) and then refer to the similarities and differences compared to the HC subjects.
Overall, in the PD group, subtracting the looking at the CS from the reading of the sentences resulted exclusively in LH activations. More particularly the left IFG pars triangularis (BA 45), the left precentral gyrus (BA 6) and the left STG (BA 22). Only the activation in the left BA 22 was found to be in common with the HC subjects (see Figure 8.8, Table 8.4).
A. Reading sentences versus looking at CS: left and right lateral view
B. Reading sentences versus looking at CS: anterior and posterior view
Figure 8.8: For the PD patients, areas activated in whole‐brain analysis for reading of sentences vs. looking at CS overlaid on a template brain surface. A: left and right lateral view; B: anterior and posterior view. The results are thresholded at p < .05, corrected for multiple comparisons.
L R
P A

Chapter 8: fMRI study in Parkinson’s Disease patients
142
Table 8.4: Results of whole‐brain analyses comparing reading of sentences versus looking at CS (whole sequence duration) for PD patients and HC subjects
PD patients HC subjects
Region BA Side MNI Coordinates
T‐value
kE MNI Coordinates
T‐value
kE
x y z x y z SMA 6 L ‐8 4 58 8.76 661
Precentral gyrus 6 L ‐36 ‐10 64 5.08 61 Cingulate gyrus 24 R 2 18 40 5.78 96 Post‐& precentral
gyrus 3,46
L ‐58 ‐22 44 8.09 1579
Postcentral gyrus 3 R 54 ‐24 42 8.04 220 Inferior frontal
gyrus 45 L ‐50 30 2 8.35 71
Inferior frontal gyrus
47 L ‐44 32 ‐6 6.48 69
Superior temporal gyrus
22 L ‐60 ‐42 12 9.19 42
Superior temporal gyrus
22 L ‐58 ‐38 4 8.92 1044
Middle temporal gyrus
21 R 52 ‐24 ‐6 6.12 290
Middle temporal gyrus
21 R 54 ‐2 ‐16 5.91 100
Superior parietal lobe
5 R 22 ‐46 68 5.16 81
Superior parietal lobe
7 L ‐26 ‐60 46 5.10 57
Superior parietal lobe
7 R 30 ‐60 36 6.81 194
Precuneus 7 R 10 ‐70 34 5.13 94 Lingual gyrus 18 R 12 ‐88 ‐2 7.29 468
Ventral pallidum ‐ R 18 ‐4 ‐8 6.24 72 Cerebellum ‐ R 20 ‐58 ‐20 5.37 60
Note: The T‐value is of the maximally activated voxel of which the corresponding MNI‐coordinates are given. Results are corrected for multiple comparisons at p < .05. See for the abbreviations the List of Abbreviations.

Chapter 8: fMRI study in Parkinson’s Disease patients
143
The mask analysis confirmed that PD patients and HC subjects had activation in the left STG (BA 22) in common, but also in the left precentral gyrus (BA 6). HC subjects showed more activation in the left SMA (BA 6) and the right superior parietal lobe (SPL, BA 7) however, these activations were only in the direction of a trend (see Figure 8.9, Table 8.5). Against our expectations, the PD patients did not show greater cortical activation than the HC subjects did.
Reading sentences versus looking at CS: HC > PD
Figure 8.9: Increased activation for HC subjects versus PD patients in whole‐brain analysis to overlaid on several axial slices of the brain. The results are thresholded at p < .05, corrected for multiple comparisons.
Table 8.5: Activated areas during reading of sentences versus looking at CS (whole sequence duration) for HC subjects compared to PD patients.
Region Side HC > PD HC = PD
MNI Coordinates
T‐value MNI Coordinates
T‐value
x y z x y z SMA BA 6 L ‐8 4 58 *5.18 Precentral gyrus BA 6 L ‐36 ‐10 64 5.08 Precentral gyrus BA 6 L ‐38 ‐6 44 *4.03 Superior parietal lobe BA 7 R 20 ‐50 56 *4.41 Superior temporal gyrus BA 22 L ‐60 ‐42 12 9.19
Note: pcorr < .05 (at the voxel level);* trend. See for abbreviations in List of Abbreviations.
Comparing all the passive versus all the active sentences in the PD group showed a significant activation at the cluster level in the lingual gyrus (BA 18, T = 3.92, pcorrected < .05, peak MNI coordinates 16, ‐88, ‐10, kE = 100), but in the RH rather than in the LH, as in the HC subjects. Furthermore, compared to HC subjects, PD patients showed significant increased activation for passive versus active sentences in the left medial/superior frontal gyrus (BA 10, T = 4.52, pcorrected < .05, peak MNI coordinates ‐8, 64, 10, kE = 95) (see Figure 8.10 and Table 8.6). In contrast to the HC subjects (see Chapter 7, Table 7.8 and Figure 7.7A & B), no significant activations above the threshold were observed for the main effect of grammaticality or the interaction effect of canonicity x grammaticality in the PD group.
65 60 55

Chapter 8: fMRI study in Parkinson’s Disease patients
144
Main effect of canonicity: P > A
Figure 8.10: For the PD patients, areas activated in whole‐brain analysis to main effect of canonicity overlaid on several axial slices of the brain. The results are thresholded at p < .05, corrected for multiple comparisons.
Table 8.6: Activated areas during reading of endpoints of passive sentences versus reading of active sentences for PD patients and HC subjects
Region Side PD patients HC subjects
MNI Coordinates
T‐value MNI Coordinates
T‐value
x y z x y z Lingual gyrus BA 18 L ‐24 ‐84 ‐10 4.91 Lingual gyrus BA 18 R 16 ‐88 ‐10 3.92 Medial/superior frontal gyrus
BA 10 L ‐8 64 10 4.52
Note: pcorr < .05; T ≥ 3.21 (at the cluster level); cluster threshold of 20 voxels. For abbreviations see the List of Abbreviations.

Chapter 8: fMRI study in Parkinson’s Disease patients
145
8.4 Discussion
The goal of this fMRI study was to evaluate the patterns of activation during the comprehension of sentences in which canonicity and grammaticality were manipulated in patients with PD compared to healthy older adults.
8.4.1 Alterations in canonicity processing in PD
In our previous study of HC subjects (see Chapter 7) a main effect of canonicity was evident in the right pallidum, in which more activation was found for the active sentences compared to the passive sentences. It was suggested that the output nucleus supported facilitation of heuristic routines in the comprehension of canonical sentences (i.e., active voice). In the same study, the factorial whole brain analysis showed an increased activation in the right putamen. In the current study, when the groups were analyzed together, a trend to a main effect of canonicity was found in the right putamen , which is consistent with previous whole brain findings of only HC subjects (see Chapter 7). In contrast to our expectations, no interaction effect with group in the right putamen was found, meaning that PD patients did not show an explicit reduced word order effect in their right putamen compared to HC subjects. However, in line with our expectations, a main effect of canonicity was found in the right pallidum for the HC subjects only, suggesting that PD patients fail to exploit sequential syntactic information and, if our interpretation is correct, possibly lose their capacity to use heuristic routines in the comprehension of the canonical active sentences.
Moreover, PD patients do not engage their left MTG (BA 21) to process passive sentences, since a canonicity effect was only evident in the BA 21 for the HC subjects. Activation in the left posterior middle temporal gyrus has been implicated in the storage of lexical representations (Hickok & Poeppel, 2004).
Although PD patients have a reduced sensitivity to word order variations via the normal system, they did show an increased recruitment of the left medial/superior prefrontal cortex (PFC, BA 10) during the reading of passive sentences compared to active sentences. This increased recruitment of the left medial PFC (BA 10) was found in addition to common activation with HC subjects in a visual area (Lingual gyrus, BA 18) and possibly reflects a compensatory cortical mechanism. In the following, three possible explanations for the involvement of the PFC in the processing of passive sentences in PD patients are put forward.
One of the central issues in this thesis was to examine the degree to which deficits in language processing in PD can be explained by deficits independent of language. As described in Chapter 5, PD patients’ impaired comprehension of passive sentences was associated with their poor set‐switching abilities, inhibition and WM scores. The PFC has since long been associated with WM, (for a review see Wager & Smith, 2003). The PD patients participating in the fMRI experiment performed as good as the HC subjects on the verbal WM span tasks (i.e., Reading Span Task in Dutch, Van den Noort et al., 2005). Traditional theories of WM have distinguished between storage and executive components (Baddeley, 1986), with the former responsible for maintaining information online and the

Chapter 8: fMRI study in Parkinson’s Disease patients
146
latter responsible for its manipulation (i.e., the execution of control). Carpenter et al. (1994) hypothesized that WM load is directly related to sentence complexity. Higher sentence complexity is found if roles occur in non‐canonical order (such as in passives and object‐relatives). King and Just (1991) demonstrated that the object‐relatives (e.g., The reporter that the senator attacked admitted the error) imposed a higher demand on verbal WM than the subject‐relatives (e.g., The reporter that attacked the senator admitted the error). This leads to a first explanation. Evidence from neuroimaging literature points to an engagement of the PFC in tasks relying more on controlled processing (Baker et al., 1996; Banich et al., 2000; Cohen et al., 1997; Frith et al., 1991; MacDonald et al., 2000; Smith & Jonides, 1999). Thus the PD patients possibly relied on the executive control component of WM allocated in the PFC. The results of our behavioral sentence comprehension study (Chapter 5) are compatible with this idea. The backward digit span (testing merely manipulative aspects of WM) of PD patients was correlated to comprehension of passive sentences, suggesting that PD patients who had problems with the manipulative aspects of WM also had more difficulties processing passive sentences. Given this behavioral evidence, the findings from this present fMRI study and the fact that the PD patients did not show reading span deficits suggests that WM allocated in the PFC contributes to comprehension of the passive sentences in PD patients. Furthermore, PD patients’ reliance on WM during the comprehension of passive sentences can be interpreted as a compensatory mechanism (see Chapters 4 and 5 and Colman et al., 2009; Koerts et al., 2009; Marié et al., 2007).
Secondly, examining ambiguity resolution, Stowe, Paans, Wijers, and Zwarts (2004) reported a similar left medial prefrontal area as the current study, which they linked to supporting higher‐level semantic processes involved in evaluation of plausibility. Indeed, this PFC area is part of a distributed neural system underlying the Theory of Mind (ToM)23 mechanism (Walter, Adenzato, Ciaramidaro, Enrici, Pia, & Bara, 2004). In a study that investigated the representation of abstract person‐based knowledge for actions, Mason, Banfield and Macrae also (2004) reported activity in the medial PFC for action judgments about people and not about animals, which is indicative of ToM reasoning. The present findings are supportive of the viewpoint of plausibility of the action in the sentence context. When lying in the scanner, PD patients appeared to reason about human behavior associated with the verb, instead of relying on the stored lexical representations. Although we did not explicitly ask subjects to judge actions expressed by the sentence final verb, patients might still have done this covertly. However, why did the PD patients rely more on this plausibility strategy when processing passives than when processing actives? The answer lies in the findings of Traxler et al. (2005), who monitored participants’ eye movements while they read reversible object and subject relative sentences similar to the ones studied in King and Just (1991). In contrast to King and Just’s (1991) results, Traxler et al.’s eye tracking results provided no evidence that WM capacity was associated to the difference between object and subject relatives. However, they found that participants with higher WM capacity were better able to make use of semantic information to reduce the difficulty of object relatives. For the present study, PD patients could only have relied on WM in the syntactically more complex passive sentences.
23 Theory of Mind (ToM) has been defined as our ability to explain and predict other people’s behavior by attributing independent mental states to them (Baron‐Cohen, 1995).

Chapter 8: fMRI study in Parkinson’s Disease patients
147
The anterior part of the left prefrontal cortex has also been described as being involved in producing coherent sequences of actions tested with action event scripts (Sirigu et al., 1998). Sirigu et al. (1998) demonstrated that patients with lesions in the prefrontal cortex had difficulties when a sequence was related to the semantic aspects of action order in a script, as opposed to the syntactic aspects of ordering words in a sentence. For the present results, this implies that the exclusive activation of the intact prefrontal cortex in PD patients for passive sentences reflects a semantic strategy for dealing with word order information. However, many non‐demented PD patients have a relative sparing of their lexical semantic and world knowledge (Hochstadt, 2009; Hochstadt et al., 2006). Several studies evidenced that PD patients have higher sentence comprehension scores when the use of syntax can be circumvented by relying on the semantic information from the individual words in the sentence (Grossman, 1999; Grossman et al., 1991, 1992, 2000, 2001; Grossman, Lee, et al., 2002; Grossman, Zurif, et al., 2002; Kemmerer, 1999; Lieberman et al., 1990, 1992; Natsopoulos et al., 1991, 1993; Skeel et al., 2001). This suggests that comprehension in PD patients was intact at the word level, but meaning at the sentence level was disturbed under conditions in which word order was essential for comprehension. In addition, it has been hypothesized that during sentence processing, the BG contribute to a selective gating (or filtering) of relevant information into WM by inhibiting or disinhibiting the prefrontal cortex (Frank, Loughry, & O’Reilly, 2001; McNab & Klingberg, 2008). According to this hypothesis, PD patients would have disturbed filtering of lexical‐syntactic information into WM, which might in turn complicate their sentence comprehension for complex sentences.
However, the Sirigu’s semantic strategy cannot have been very successful since only semantically reversible sentence segments were used during this experiment. By using semantically reversible sentences we aimed at avoiding the use of semantic strategies, such as those successfully used by individuals with agrammatic aphasia (Caramazza & Zurif, 1976).
To summarize, the lack of an effect of canonicity in the globus pallidus of the PD patients in the present study illustrates the disturbed involvement of the output nucleus in word order processing during sentence comprehension of active sentences. The data furthermore show that the PD patients in this study did not show WM impairments, which makes it more likely that WM allocated in the PFC was engaged to process specifically the passive sentences in a semantic way. However, due to a disturbance in the filtering of relevant semantic information, comprehension can still be disturbed.
8.4.2 Disproportionate focus on the inflectional violation in PD
Reduced or absent P600 effects in patients with PD or BG lesions were seen as evidence for the involvement of the BG in late, controlled syntactic integration processes, rather than early automatic structure building processes (Friederici et al., 1999, 2003; Kotz et al., 2002, 2003). Based on this model, PD causes deficits in the late controlled processes that are used for late stage syntactic integration and reanalysis processes when there is a conflict between lexical‐semantic and syntactic information. However, Hochstadt (2009) interpreted the reduced P600 effect in PD differently and suggested that “the reduced P600 might reflect difficulty with not just syntactic integration but integration with context more broadly” (p. 2001). Rather than having deficits in monitoring conflict, PD patients might not experience the conflict to begin with.

Chapter 8: fMRI study in Parkinson’s Disease patients
148
Similar to what was found for the HC subjects (Chapter 7), for the PD patients a main effect of grammaticality was evident in subparts of the IFG. More specifically, an effect of grammaticality was found for the bilateral pars opercularis (BA 44), pars triangularis (BA 45) and pars orbitalis (BA 47) of the IFG. Unlike the matched control group, however, PD patients showed a main effect of grammaticality in the pallidum and the left putamen. In the PD group, the grammaticality effect in all these regions was based on the higher activation for the sentences containing an inflectional violation compared to the other types of violation. Two possible underlying mechanisms are proposed for this disproportionate focus on the inflectional violation:
First, in on‐ and off‐line behavioral studies, PD patients showed reduced sensitivity to sentence anomalies (Grossman et al., 1992 and Grossman et al., 2000). Similarly, PD patients were described as processing sentences “on a more superficial level” than controls (Whiting et al., 2005). Given these earlier findings, one possibility is that the patients noticed the inflectional violation solely on the basis of the physical visual features of the shorter verb segment during the highly automatic first‐pass parsing processes. Consequently, increased activation in the cortical and subcortical parts of the cortico‐straito‐cortical circuits is only seen for the inflectional violations.
Alternatively, the PD patients may not have been able to shift their attention to a subsequent integration processes of the violated verb form into the sentence. This suggestion is based on the broader impairment of PD patients in switching attention between cognitive sets (e.g., Cools et al., 2001; Green et al., 2002; Muslimovic et al., 2005). However, in this study the switching capacities as measured by the TMT index of PD patients and the age‐ and education‐matched controls were equivalent. Therefore, poor attentional shifting is not a likely explanation.
To conclude, the precise reason why PD patients focus their attention on the short verb segment is still an issue of speculation and needs further investigation.

Chapter 8: fMRI study in Parkinson’s Disease patients
149
8.4.3 Opposite patterns of activation in BA 47 for the participant groups
A clear grammaticality effect independent of canonicity was found in the bilateral BA 44 and BA 45 of the PD patients and was due to increased activation for the inflectional violation independent of canonicity. In the HC subjects it is clear that grammaticality and canonicity interacted in both BA 44 and BA 45, with larger effects of violation for the active sentences than the passive sentences. The IFG (BA 44/45) has also been found to be significantly activated by grammatical violations in other neuroimaging studies. Friederici, Opitz and Von Cramon (2000) suggested that left IFG activation was modulated by the grammaticality of the sentence.
In the BA 47, a clear opposite pattern of activation was evident for the two groups. PD patients apparently relied more on this inferior frontal area for the violated sentences than for the non‐violated sentences. HC subjects activated the BA 47 more for the non‐violated sentences as compared to the violated sentences.
There is strong support for a role of the pars orbitalis of the left IFG (BA 47) in semantic processing across a range of tasks (Dapretto & Bookheimer, 1999; Gold & Buckner, 2002; McDermott et al., 2003; Poldrack et al., 1999; for a review, see Bookheimer, 2002). Moreover, Badre and Wagner (2007) proposed different cognitive functions for sub‐regions of the IFG. The left BA 45 may support a domain‐general selection process among active representations that operates post‐retrieval, whereas the left BA 47 may support controlled access to stored conceptual representations. In the fMRI study of Baumgaertner, Weiller and Büchel (2002), an increased activation was found in the lower part of the left IFG (BA 47) for sentence completions that violated contextual constraints compared to expected sentence completions. Their findings were consistent with an essential role for the left inferior frontal cortex in a semantic executive system that contributes to the on‐line retrieval of semantic information (Demb et al., 1995; Gabrieli et al., 1998; Poldrack et al., 1999; Thompson‐Schill et al., 1997). Baumgaertner et al. (2002) argued that processing of the semantic anomalies involved executive functions such as “monitoring semantic representations” (Baumgaertner et al., 2002, p. 743).
In the present study, task instructions required subjects to comprehend the meaning of the sentences. In the course of processing the subsequent sentence segments, more lexical semantic information becomes available and generates expectations with respect to characteristics of subsequent words (e.g., Kutas & Hillyard, 1984). A word that corresponds well to expectations should be easy to integrate into the overall sentence intention. Subjects activate the next sentence element based on probabilistic factors that are the result of previous experiences with the occurring language elements (MacDonald & Christiansen, 2002). An unexpected word is more difficult to integrate and this will be associated with increased processing cost (see also Hagoort, 2003, 2005). Applying this assumption to the violation paradigm used in our study, processing of the morphosyntactic violation of the sentence final word (i.e., the participle) is supposed to increase the demand on and recruiting of cognitive resources necessary to integrate the word compared to normal sentences. It is suggested that the HC subjects were able to detect the violation and subsequently called off the sentence integration on a syntactic basis in case of the violated sentences. However, for the non‐violated sentences, the HC subjects semantically integrated

Chapter 8: fMRI study in Parkinson’s Disease patients
150
the participle of the verb in the sentence, which explains their activation of the semantic executive system supported by BA 47. Compared to the HC subjects, PD patients apparently have difficulties with the monitoring processes in reaction to the violation in the sentence. Instead, PD patients try to integrate the violated participle in the sentence relying on their semantic executive system, which produces some difficulty and greater activation of this area.
8.4.4 Verb argument processing in passive sentences in BA 6
The functional ROI analysis showed that the left medial aspect of BA 6 (SMA, MNI ‐4, 0, 64) showed an interaction of canonicity x grammaticality. The groups demonstrated a similar pattern in the left SMA. Activation was most prominent for the verb‐argument violations in the passive voice. Note that the HC subjects showed activation in a similar left BA 6 area (SMA, MNI ‐8, 4, 58) for reading sentences versus looking at CS (see Table 8.4). Interestingly, this SMA area was also found to be activated more by the HC subjects than by the PD patients (see Table 8.5). There is a body of convergent data about a critical role of the SMA in speech production, suggesting that it aids in particular initiating speech sounds (e.g., Ziegler, Kilian, & Deger, 1997). Pickard and Strick (1996) reviewed the functions of the SMA and concluded that the SMA comprises at least two subregions that can be distinguished on the basis of cytoarchitecture, connectivity, and function: the pre‐SMA located rostral to the vertical line passing through the anterior commissure (vertical commissure anterior line, VCA line), and the SMA‐proper located caudally. Based on the coordinates found in the present study, the coordinate of the functional ROI is associated with pre‐SMA. This extensive review on SMA by Picard and Strick (1996) suggests a role in planning for the pre‐SMA and in motor performance for the SMA proper. Picard and Strick (1996) also included studies that reported pre‐SMA activation for complex verbal tasks, like for example silent verb generation (Wise et al., 1991), silent rehearsal of a sequence of letters (Paulesu et al., 1993) and phonological discrimination of letter sounds (Sergent et al., 1992).
Two possible explanations are raised for the pre‐SMA activation in the current study: First, this activation in these complex verbal tasks is related to subvocalization, which probably occurred during task performance. Secondly, it is possible that the activations in the pre‐SMA during silent verbal tasks were partly due to the active inhibition of speech production, since pre‐SMA activation has also been found during the Stroop conflict task that requires inhibition of habitual responses (Pardo et al., 1990). Based on this latter view, we hypothesize that during the reading of violated sentences and in case of the passive voice, basic word order needs to be inhibited. The HC subjects in our study rely on the pre‐SMA during the sequential reading process of the sentence segments, which the PD patients clearly did less.
For the factorial whole brain analysis in exclusively the HC subjects (Chapter 7), the lateral aspect of the left SFG (BA 6, MNI ‐20, 6, 62) was activated more for the active sentences than for the passives. This part of the BA 6 is considered to be part of the premotor cortex and is traditionally defined as being involved in preparation and organization of movements and actions (Wise, 1985). However, the area is also known to be involved in cognitive functioning like abstract sequencing (Schubotz & Von Cramon, 2002; 2004). According to sensory‐motor theories of conceptual knowledge, sensory‐motor information about actions

Chapter 8: fMRI study in Parkinson’s Disease patients
151
is not only accessed during the comprehension of perceived actions but also during the comprehension of action words (Martin & Chao, 2001; Pulvermüller, 2005; Rizzolatti et al., 2001). Den Ouden, Fix, Parrish and Thompson (2009) reported clear LH activation in the precentral gyrus (BA 6), middle frontal gyrus (BA 6, 9 and 46) and in the IFG (BA 44 and 45) for the production (rather than perception) of transitive verbs compared to intransitive verbs. For the present study, the direct coupling between action representations of verb meaning and (pre)motor areas might explain the consistent activation of these action‐related areas during the processing of verb‐argument violation in the participant groups.
8.4.5 Compensatory cortical activation in PD patients
Increased activation of other brain areas than the striatum may help patients with PD to maintain task performance (Grossman et al., 2003). Our intergroup analysis contrasting all sentences versus CS showed no greater activation for the PD patients than for the HC subjects, which is in contradiction to the expected compensatory cortical activation previously found in a direct contrast of both groups by Grossman et al. (2003). However, similarly to what Grossman et al. (2003) found, in our study, common activation in both groups was evident in the left precentral gyrus (BA 6) and the left superior temporal gyrus (BA 22). In addition, where Grossman et al. (2003) reported less medial prefrontal activation (BA 32/10) in PD patients for all sentence conditions, we found increased activation in the PD patients for this prefrontal area during the reading of passive sentences compared to active sentences.
8.4.6 Suggestions for future research
Functional connectivity analysis can be used to examine the degree of collaboration between language‐specific cortical areas and the BG, when processing is compared of sentences with and without violation.
Additional studies will be required to unravel the effects of pharmacological manipulation in PD patients on their processing of sentences with morphosyntactic violations. Evidence from behavioral studies in the domain of semantic priming suggested earlier that the effects of pharmacological manipulation on semantic networks may have implications on language perception (Angwin et al., 2004; Copland et al., 2003; Kischka et al., 1996). Moreover, using functional connectivity analysis, Tivarusa, Hilliera, Schmalbrock and Beversdorf (2008) studied whether Levodopa (L‐dopa) medication administered in HC subjects affected the interaction between the components of the language network involved in semantic and phonological categorization tasks. In contradiction to the findings of the behavioral studies (Angwin et al., 2004; Copland et al., 2003; Kischka et al., 1996), no significant drug effect was found. Not finding an effect might have been due to methodological differences between the studies, since the categorization task implied more controlled and less automatic processing than the semantic priming tasks.

Chapter 8: fMRI study in Parkinson’s Disease patients
152
8.5 Conclusions
This study provided evidence that PD patients have a disturbance in processing word order during sentence comprehension, since no effect of canonicity was found in either their pallidum or MTG. However, they recruited additional PFC areas when processing passive sentences: the BA 10 for passives compared to actives, and the BA 6 for processing verb argument structure violation in the passives compared to the actives. In addition, PD patients had a deviant activation pattern with violations in the pallidum and the IFG pars orbitalis (BA 47), which was not found in the HC subjects. PD patients also relied more on the BA 47 to process inflectional violated sentences compared to other sentences, which adds evidence to their difficulties with detecting violations and their subsequent reliance on the semantic executive system to ultimately understand the violated sentences. Our results are in line with the suggestion of Hochstadt (2009) that PD patients have deficits in the late phase of sentence processing, in which monitoring of the violation is lacking. In other words, PD patients might not experience the violations in the first place.
Clearly, our suggestions are admittedly tentative and data that are more experimental are needed.







![Haptic fMRI : Combining Functional Neuroimaging with ... · uses electromagnetic motors to enable high-fidelity haptic rendering ... roimaging technique [1], [2], ... the minimum](https://static.fdocuments.us/doc/165x107/5b396d3e7f8b9a4b0a8cbc7d/haptic-fmri-combining-functional-neuroimaging-with-uses-electromagnetic.jpg)










