
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE … · effects and high cost, has led to the search of...
115
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS ALINE MYLENA GUEDES DA COSTA OLIVEIRA Avaliação da atividade antimalárica e citotóxica de plantas medicinais dos Biomas Caatinga e Amazônico NATAL – RN 2011
Transcript of UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE … · effects and high cost, has led to the search of...

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
ALINE MYLENA GUEDES DA COSTA OLIVEIRA
Avaliação da atividade antimalárica e citotóxica de plantas medicinais dos Biomas
Caatinga e Amazônico
NATAL – RN
2011

ALINE MYLENA GUEDES DA COSTA OLIVEIRA
Avaliação da atividade antimalárica e citotóxica de plantas medicinais dos Biomas Caatinga e Amazônico
Dissertação apresentada como requisito parcial à obtenção do título de mestre, pelo Curso de Pós-graduação em Ciências Biológicas, do Centro de Biociências da Universidade Federal do Rio Grande do Norte. Área de concentração: Biodiversidade (Parasitologia) Orientador: Prof. Dr. Valter Ferreira de Andrade Neto
NATAL – RN
2011

Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Oliveira, Aline Mylena Guedes da Costa. Avaliação da atividade antimalárica e citotóxica de plantas medicinais dos Biomas Caatinga e Amazônico. / Aline Mylena Guedes da Costa Oliveira. – Natal, RN, 2011. 114 f. : Il.
Orientador: Prof. Dr. Valter Ferreira de Andrade Neto. Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Ciências Biológicas.
1. Plantas medicinais – Dissertação 2. Atividade esquizonticida– Dissertação. 3. Citotoxicidade – Dissertação. I. Andrade Neto, Valter Ferreira de. II. Universidade Federal do Rio Grande do Norte. III. Título.
RN/UF/BSE-CB CDU 616.936

ALINE MYLENA GUEDES DA COSTA OLIVEIRA
Avaliação da atividade antimalárica e citotóxica de plantas medicinais dos Biomas Caatinga e Amazônico.
Dissertação aprovada como requisito parcial para obtenção do título de mestre no curso de Pós-graduação em Ciências Biológicas, do Centro de Biociências da Universidade Federal do Rio Grande do Norte, pela seguinte banca examinadora:
Área de concentração: Biodiversidade (Parasitologia)
APROVADA EM _15_/ 03_/ 2011
BANCA EXAMINADORA

Dedico a Deus, ser supremo, criador e presente
em todos os momentos de minha vida, e
aos meus pais pelo amor e incentivo.

AGRADECIMENTOS
À Deus, pela sua presença constante na minha vida, guiando os meus passos e
fortalecendo-me em momentos difíceis dessa constante caminhada que é a vida!
Aos meus pais, Geny e Ivanaldo, por todo amor, carinho e apoio incondicionais em
todos os momentos de minha vida. Amo muito vocês!
Ao meu orientador Válter F. de Andrade-Neto pelos importantes ensinamentos,
recebendo-me como orientanda, e nos fazendo componente de uma grande família que é o
“Labmat”. Muito obrigada!
Ao meu esposo por seu companheirismo, incentivo, paciência e compreensão, quando
em muitas vezes tive dividir a minha atenção com a pesquisa. Obrigada por você existir perto
de mim!
Ao meu irmão, não só pela ajuda no campo da informática, mas, sobretudo pela atenção,
compreensão e paciência;
Aos meus amigos, que mesmo distante não deixaram de acreditar e incentivar-me;
Aos meus colegas de laboratório Norma, Andrea, Milena, Cláudio, Gracielle, Cecília,
Valéria, Ywlliane, Lis, Bruno e em especial a Magaly por ter me auxiliado nos procedimentos
laboratoriais e mais que isso, pela sua amizade. Tenho certeza de que após esses dois anos de
convivência deixamos de serem colegas para nos tornarmos amigos!
A professora Magnólia Fernandes por ter disponibilizado seus equipamentos;
Ao professor Hugo por colocar a nossa disposição seu laboratório, equipamentos,
substâncias e não só isso, mas também pelo carinho, alegria e presteza em nos atender e
ensinar; obrigada pela sua participação neste trabalho, sua colaboração foi importantíssima!
À Leandro, doutorando, do Laboratório do professor Hugo, pelos esclarecimentos de
dúvidas e pelo auxílio nos procedimentos in vitro;
Aos professores de Bioestatística Gilberto Corso, Umberto Laino e Alexandre
Vasconcellos pelo auxílio na execução das análises estatísticas, pela amizade e apoio;
Ao funcionário Raimundo por nos atender sempre com presteza e carinho, nos
fornecendo os animais para nossa pesquisa. Muito obrigada!
E ao CNPQ, Conselho nacional de desenvolvimento científico e tecnológico pelo apoio
financeiro para a execução desse trabalho.

RESUMO
A resistência do Plasmodium falciparum aos antimaláricos usuais, bem como os seus
efeitos adversos e custo elevado, tornam necessária a busca de novos medicamentos contra a
malária. Diversos fármacos foram descobertos a partir de plantas medicinais com base na
etnofarmacologia, inclusive os antimaláricos mais usados atualmente; quinina e artemisinina.
Neste trabalho foi avaliada a atividade esquizonticida de extratos e frações de algumas plantas
medicinais dos Biomas da Caatinga e Amazônico a partir de um referencial
etnofarmacológico e de quimiossistemática. São elas: Ximenia americana, Maytenus rigida,
Sideroxylon obtusifolium, Stryphnodendro coriaceum, Bowdichia virgiliodes, Schinopis
brasiliensis e Picrolemma sprucei, sendo esta última, uma espécie amazônica. Os testes
antimaláricos de esquizonticidas sanguíneos foram feitos em camundongos Swiss infectados
com P. berghei e in vitro contra o P. falciparum. Estudos de citotoxicidade in vitro foram
realizados utilizando as linhagens celulares HeLa, CHO, 3T3, Raw e HEPG2. A excessão da
X. americana, todas as espécies apresentaram atividade antimalárica in vivo ou in vitro,
inibindo o crescimento do parasito em até 79%. Os extratos exibiram toxicidade moderada
com cinética de atividade dose-dependente. Nesse contexto, a abordagem etnofarmacológica
associada ao perfil quimiossistemático, se mostram ferramentas úteis e promissoras na busca
de novos fármacos, permitindo contribuir significativamente para o conhecimento científico
do potencial antimalárico da flora brasileira e deste modo, abrir perspectivas para o
desenvolvimento de novos antimaláricos.
Palavras-Chave: Plantas medicinais. Atividade esquizonticida. Citotoxicidade. Malária.

ABSTRACT
Resistance of Plasmodium falciparum to the usual antimalarials, as well as their adverse
effects and high cost, has led to the search of new drugs against malaria. Several of these have
been developed from medicinal plants based on ethnopharmacology, including the most
widely used antimalarials today: quinine and artemisinin. In the present study schizonticide
activity of extracts and fractions of a number of medicinal plants from the Caatinga and
Amazon biomes were assessed based on ethnopharmacological and chemosystematic
information. These included Ximenia americana, Maytenus rigida, Sideroxylon obtusifolium,
Stryphnodendro coriaceum, Bowdichia virgiliodes, Schinopis brasiliensis and Picrolemma
sprucei, the last, an Amazon species. Antimalarial tests of blood schizonticides were
conducted in Swiss mice infected with P. berghei and in vitro against P. falciparum. In vitro
cytotoxicity studies were carried out using HeLa, CHO, 3T3, Raw and HEPG2 cell lines.
Except for X. americana, all species exhibited in vivo or in vitro antimalarial activity,
inhibiting parasitic growth by up to 79%. Extracts exhibited moderate toxicity with dose-
dependent kinetics. In this sense, ethnopharmacological and chemosystematic approaches
were shown to be useful and promising tools in the search of new drugs. These findings
represent a significant contribution to scientific knowledge of the antimalarial potential of
Brazilian flora, thereby opening perspectives for the development of new antimalarials.
Keywords: Medicinal plants. Schizonticide activity. Cytotoxicity. Malaria.

LISTA DE FIGURAS
Figura 01 - Número de casos de malária de 1999-2007 notificados no Brasi. 17
Figura 02 - Índice Parasitário Anual da Amazônia Legal 16
Figura 03 - Proporção de espécie de Plasmodium na Amazônia Legal 17
Figura 04 - Ciclo biológico de Plasmodium falciparum 20
Figura 05 - Resistência de Cloroquina e Sulfadoxina-pirimetamina no mundo 26
Figura 06 - Ximenia americana (tronco e folhas) 39
Figura 07 - Maytenus rigida (folhas) 30
Figura 08 - Sideroxylon obtusifolium (tronco e folhas) 31
Figura 09 - Stryphnodendron coriaceum (folhas) 32
Figura 10 - Bowdichia virgilioides (aspecto geral, folhas e flores) 33
Figura 11 - Schinopis brasiliensis (aspecto geral e folhas) 34
Figura 12 - Picrolemma sprucei (folhas) 35
Figura 13 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato etanólico de Ximenia americana (ameixeira)
45
Figura 14 - Viabilidade celular de diferentes linhagens celulares após tratamento com Ximenia americana (ameixeira) por 24 horas
46
Figura 15 - Viabilidade celular de diferentes linhagens celulares após tratamento com Ximenia americana (ameixeira) por 48 horas
47
Figura 16 - Redução de parasitemia do Plasmodium berghei em camundongos tratados com o extrato etanólico de Maytenus rigida (bom nome)
48
Figura 17 - Viabilidade celular de diferentes linhagens celulares após tratamento com Maytenus rigida por 24h
49
Figura 18 - Viabilidade celular de diferentes linhagens celulares após tratamento com Maytenus rigida por 48h
50

Figura 19 - Redução da parasitemia Plasmodium berghei em camundongos tratados com o extrato etanólico de Sideroxylon obtusifolium (Quixabeira)
51
Figura 20 - Viabilidade celular de diferentes linhagens celulares após a incubação de 24h com diferentes concentrações do extrato de Sideroxylon
obtusifolium (Quixabeira)
52
Figura 21 - Viabilidade celular de diferentes linhagens celulares após a incubação de 48h com diferentes concentrações do extrato de Sideroxylon
obtusifolium (Quixabeira)
53
Figura 22 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato etanólico de Stryphnodendron coriaceum (Barbatimão)
54
Figura 23 - Viabilidade celular de diferentes linhagens celulares após a incubação de 24h com diferentes concentrações do extrato de Stryphnodendron
coriaceum (Barbatimão)
55
Figura 24 - Viabilidade celular de diferentes linhagens celulares após a incubação de 48h com diferentes concentrações do extrato de Stryphnodendron
coriaceum (Barbatimão)
56
Figura 25 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato etanólico de Bowdichia virgilioides (Sucupira)
57
Figura 26 - Viabilidade celular de diferentes linhagens celulares após a incubação de 24h com diferentes concentrações do extrato de Bowdichia virgilioides (Sucupira)
58
Figura 27 - Viabilidade celular de diferentes linhagens celulares após a incubação de 48h com diferentes concentrações do extrato de Bowdichia virgilioides (Sucupira)
59
Figura 28 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato etanólico de Schinopsis brasiliensis (Baraúna)
60
Figura 29 - Viabilidade celular da célula HeLa após a incubação por diferentes tempos e concentrações com o extrato de Schinopis brasiliensis
(Baraúna)
61
Figura 30 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato aquoso do caule de Picrolemma sprucei
(Caferana)
63
Figura 31 - Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato aquoso da raiz de Picrolemma sprucei (Caferana)
64

Figura 32 - Viabilidade celular de diferentes linhagens celulares após a incubação de 24h com diferentes concentrações do extrato de aquoso do caule Picrolemma sprucei (Caferana)
65
Figura 33 - Viabilidade celular de diferentes linhagens celulares após a incubação de 48h com diferentes concentrações do extrato de aquoso do caule Picrolemma sprucei (Caferana)
65
Figura 34 - Viabilidade celular de diferentes linhagens celulares após a incubação de 24h com diferentes concentrações do extrato de aquoso da raiz Picrolemma sprucei (Caferana)
66
Figura 35 - Viabilidade celular de diferentes linhagens celulares após a incubação de 48h com diferentes concentrações do extrato de aquoso da raiz Picrolemma sprucei (Caferana)
66
Figura 36 - Inibição do crescimento in vitro da cepa K1 de Plasmodium falciparum 68

LISTA DE TABELAS
Tabela 1 - Extratos e frações de diferentes espécies vegetais brasileiras utilizados nos testes antimaláricos in vivo e in vitro.
37
Tabela 2 - Atividade antimalárica da Ximenia americana (ameixeira). 45
Tabela 3 - Atividade antimalárica in vivo do extrato de Maytenus rigida (Bom nome). 48
Tabela 4 - Teste antimalárico in vivo de S. obtusifolium 51
Tabela 5 - Atividade antimalárica in vivo do extrato de Stryphnodendro coriaceum (Barbatimão)
54
Tabela 6 - Atividade antimalárica in vivo do extrato de Bowdichia virgiliodes (Sucupira)..
57
Tabela 7 - Atividade antimalárica in vivo do extrato de Schinopis brasiliensis (Baraúna).
60
Tabela 8 - Atividade antimalárica in vivo do extrato de caule de Picrolema sprucei
(caferana) 62
Tabela 9 - Atividade antimalárica in vivo do extrato de raiz de Picrolema sprucei (caferana)
63
Tabela 10 Quadro sinóptico dos resultados relativo à citotoxicidade e das análises feitas in vivo e in vitro das espécies vegetais estudadas
69

SUMÁRIO
1 INTRODUÇÃO 13 1.1 O PARASITO E O CICLO EVOLUTIVO 17
1.2 MANIFESTAÇÕES CLÍNICAS 21 1.3 DIAGNÓSTICO 22 1.4 CONTROLE 24 1.5 A ETNOFARMACOLOGIA 27
2 OBJETIVOS 36
3 MATERIAL E MÉTODOS 37 3.1 ESPÉCIES VEGETAIS E PREPARO DE SEUS EXTRATOS E FRAÇÕES 37 3.2 MANUTENÇÃO DO Plasmodium berghei 38 3.3 ANIMAIS 39
3.4 TESTE DE TOXICIDADE AGUDA 39
3.5 TESTES ANTIMALÁRICO IN VIVO 40
3.5.1 Administração de extratos aos camundongos 40
3.5.2 Avaliação da atividade antimalárica in vivo 40
3.5.3 Determinação da percentagem de parasitemia em esfregaços sanguíneos 41
3.6 TESTES ANTIMALÁRICOS IN VITRO CONTRA Plasmodium falciparum 41
3.7 TESTES DE CITOCICIDADE (IN VITRO) 42
3.8 ANÁLISE ESTATÍSTICA 43
4 RESULTADOS 44
4.1 Ximenia americana (AMEIXEIRA) 44
4.1.1 Teste de toxicidade aguda 44
4.1.2 Testes antimaláricos in vivo 44
4.1.3 Citotoxicidade 46
4.2 Maytenus rigida (BOM NOME) 47
4.2.1 Testes antimalárico in vivo 47
4.2.2 Teste de citotoxicidade 49
4.3 Sideroxylon obtusifolium (QUIXABEIRA) 50
4.3.1 Testes antimaláricos in vivo 50
4.3.2 Citotoxicidade 52
4.4 Stryphnodendro coriaceum (BARBATIMÃO) 53
4.4.1 Testes antimalárico in vivo 53
4.4.2 Citotoxicidade 55
4.5 Bowdichia virgiliodes (SUCUPIRA) 56

4.5.1 Teste antimalárico in vivo 56
4.5.2 Citotoxicidade 58
4.6 Schinopis brasiliensis (BARAÚNA) 59
4.6.1 Teste antimalárico in vivo 59
4.6.2 Citotoxicidade 61
4.7 Picrolemma sprucei (CAFERANA) 62
4.7.1 Teste antimalárico in vivo 62
4.7.2 Citotoxicidade 64
4.8 AVALIAÇÃO DA ATIVIDADE ANTIMALÁRICA EM ENSAIO IN VITRO COM O Plasmodium falciparium
67
5 DISCUSSÃO 70
5.1 TOXICIDADE AGUDA 73
5.2. ATIVIDADE ANTIMALÁRICA IN VIVO 74
5.3. ATIVIDADE ANTIMALÁRIA IN VITRO 79
5.4. CITOXICIDADE 82
6. CONCLUSÃO 85
REFERÊNCIAS 86 ANEXO A – Artigo Avaliação citotóxica e antimalárica de Maytenus rigida (Celastraceae) Sideroxylon obtusifolium (Sapotaceae) e Sideroxylon obtusifolium
(Anacardiaceae)
112

13
1. INTRODUÇÃO
A malária, popularmente conhecida por impaludismo, paludismo ou febre
intermitente, é uma doença parasitária que apresenta grande morbidade, acarretando sérios
problemas em saúde pública e considerável impacto econômico. Esta parasitose acomete
milhões de pessoas nas áreas tropicais e subtropicais do planeta. O número de casos de
malária ocorridos no mundo no ano de 2008 foi estimado em 243 milhões, sendo que 85% do
total ocorreram na África sub-saariana, 10% no sudeste da Ásia, 4% na região leste do
Mediterrâneo e o restante dos casos distribuídos em outros países, onde a malária é endêmica.
Essa doença se não for tratada pode ser mortal, matando mais de 1 milhão de indivíduos por
ano (WORLD HEALTH ORGANIZATION-WHO, 1997). Segundo dados epidemiológicos
de 2009 da Organização Mundial de Saúde, a África, a Ásia, as Américas e Euroásia/Oriente
Médio detêm, respectivamente, 50, 20, 22 e 17 países de ocorrência da doença, números esses
a serem reduzidos até 2015 através dos investimentos internacionais em medidas de controle
(WORLD HEALTH ORGANIZATION, 2009).
Os relatos sobre a malária datam do século IV e III a.C. Escritos de febres
sazonais ou intermitentes são encontrados em textos médicos e religiosos das antigas
civilizações suméria, assíria, babilônica, chinesa, egípcia e indiana. Os acessos febris no
período da doença foram atribuídos, principalmente, a ira dos deuses ou demônios que
governavam o destino da humanidade e da natureza, cuja maldição poderia ser afastada com
rituais, sacrifícios e uso de diferentes de infusões de ervas.
No ano de 2700 a.C., a medicina tradicional chinesa associou o aumento do baço
aos vários acessos de febre, atribuindo à doença a três demônios que eram responsáveis, cada
um deles, pela dor de cabeça, pelos calafrios e pela própria febre; tais sintomas eram curados
na época com exorcismos, acupuntura, cauterização da pele com uso de determinadas ervas e
com preparações a base de plantas medicinais; provavelmente já era utilizado o “Qinghaousu”
(Arthemisia annua) descrito pelos textos chineses em 168 a.C. (CROTTI, 2005).
Os gregos e os romanos coabitaram com a malária em toda a sua história. Foi a
partir de Hipócrates (século V a.C.) que a origem da doença deixou de ser atribuída a castigo
dos deuses, sendo, portanto, nesse novo ponto de vista, proveniente de causa natural, ligada a
ambientes pantanosos e insalubres. Deste modo, é compreensível a origem da palavra malária
a partir das expressões "mal'aria", "mal aere" ou "mal aire" cuja tradução destacava a crença
da origem da doença a partir do ar insalubre, ou ar "corrupto" dessa época.

14
Antes do período das grandes navegações, a malária, era causada na maioria dos
casos pelas espécies Plasmodium vivax e P. malariae; todavia, foi com o comércio entre
países africanos e o Oriente (século XVI) que ocorreu o surgimento da forma perniciosa da
doença causada pelo Plasmodium falciparum.
Em meados do século XIX, a abordagem experimental para o estudo de doenças
infecciosas, se tornou mais específica, através da microscopia e de metodologias inovadoras.
Um grande marco na história da malária foi à observação feita pelo cientista francês Laveran,
médico militar na Argélia, que visualizou os parasitas no sangue de pacientes não tratados
com quinino. No final, veio à confirmação da associação definitiva entre malária e estes
protozoários por parte dos italianos Celli, Marchiafava e Golgi, bem como as diferenças
morfológicas das espécies responsáveis pelas febres terçã "maligna" (causada pelo
Plasmodium falciparum), terçã "benigna" (causadas pelas espécies Plasmodium vivax e
Plasmodium ovale) e a febre quartã (causada pelo Plasmodium malariae). Outros
pesquisadores também foram importantes nessa fase inicial de descobertas sobre a malária,
mas foram os italianos Grassi, Bignami e Bastianelli, os responsáveis pelas maiores
descobertas; a descrição das fases do ciclo de vida do protozoário da malária no ser humano e
sua relação com a sintomatologia da doença (CROTTI, 2005).
Em nosso país, a malária já existia, desde o início do século XX, disseminada por
quase todo o território brasileiro, inclusive em estados do Nordeste, como o Rio Grande do
Norte e Ceará (MOTA, 2009). A criação do Instituto Soroterápico Federal no Rio de Janeiro –
que daria origem ao Instituto Oswaldo Cruz, em 1908 – e do Instituto Butantã em São Paulo,
marcariam um movimento institucional de reorganização dos serviços de higiene,
oficializando propostas de medidas de combate as endemias (LIMA; FONSECA;
HOCHMAN, 2005). Na década de 20, foi aprovado o regulamento do Departamento Nacional
de Saúde Pública do Ministério da Justiça e Negócios Interiores que estabelecia medidas para
profilaxia e controle da malária, tendo por base o diagnóstico parasitológico e a determinação
de índices endêmicos, medidas antilarvárias e tratamento de doentes, bem como a ênfase às
investigações epidemiológicas (ORGANIZAÇÃO PAN-AMERICANA DA SAÚDE, 1998).
Em 1965 foi iniciada a Campanha de Erradicação e Controle da Malária (CEM)
que, privilegiando medidas de controle vetorial, foi capaz, segundo Loiola, Silva & Tauil
(2002), de eliminar a malária em extensas áreas do país, permanecendo sua ocorrência
basicamente restrita à Amazônia. Porém, o grande fluxo de migrantes a partir dos anos 70,
causou um aumento da incidência da malária, de 52.469 casos observados nesta década para
169.871 nos anos 80. Decorrente dessas constatações, a CEM foi desativada e a coordenação

15
das atividades de controle em âmbito nacional passa para a Superintendência de Campanhas
de Saúde Pública (SUCAM), inaugurando um percurso de mudanças institucionais e de
medidas de intervenção (LOIOLA; SILVA; TAUIL, 2002; BARATA, 1995).
Atualmente, a malária continua ainda sendo um dos principais problemas de saúde
pública e de considerável impacto econômico no Brasil e no mundo. No ano 2000, o
Ministério da Saúde lançou o Plano de Intensificação das Ações de Controle da Malária na
Amazônia Legal (PIACM). Porém, esse plano acabou sendo continuado pelo Programa
Nacional de Controle da Malária (PNCM), de criação do Ministério da Saúde em 2003, que
então, procurou manter os ganhos obtidos pelo PIACM, fortalecendo ainda mais as estruturas
dos serviços de saúde, para melhorar e ampliar o atendimento aos pacientes de malária.
Contudo, não foi isso que se observou nos anos de 2003 a 2005, períodos esses marcados pelo
aumento no número de casos de malária, chegando a 600 mil casos de malária, incluindo
adultos e crianças. Números esses diminuídos nos anos subseqüentes (Figura 1)
(MINISTÉRIO DA SAÚDE, 2007).
Figura 01- Número de casos de malária de 1999-2007 notificados no Brasil.
Fonte: Sismal e Sivep- Malária/SVS/MS
No âmbito mundial.
Os investimentos nas medidas de controle da malária são bastante elevados, no
ano de 2007 o valor gasto foi de US$ 0.65 bilhões, quantia ainda pequena diante da real
necessidade de um controle mais efetivo (US$ 5 bilhões) (WORLD HEALTH
ORGANIZATION, 2009).
No Brasil, os investimentos para o controle da malária passaram de US$ 100
milhões em 2006. O controle da malária vem sendo dificultado, principalmente, por fatores
relativos às questões relacionadas à população suscetível, ao agente etiológico, ao vetor, além

16
das condições ecológicas, geográficas, econômicas, sociais e culturais (MINISTÉRIO DA
SAÚDE, 2005); sendo a região da Amazônia Legal (Amazonas, Pará, Acre, Roraima,
Rondônia, Amapá, Mato Grosso, Tocantins e Maranhão) responsável pela maioria dos casos
de malária no Brasil.
Os índices de ocorrência para a malária são classificados de acordo com a
Incidência Parasitária Anual (IPA), como de alto risco (IPA≥50/1.000 hab.), médio risco (IPA
entre 10 49/1.000 hab.) e baixo risco (IPA<10/1.000 hab.) (Figura 2), na qual a espécie P.
vivax é a predominante, como pode ser observado na figura 3.
Figura 02- Índice Parasitário Anual da Amazônia Legal. (MINISTÉRIO DA SAÚDE,
2008).
FONTE: Sivep-Malaria/SVS/MS

17
Figura 03- Distribuição da proporção de espécie de Plasmodium na Amazônia Legal no ano de 2009.
Fonte: Sismal e Sivep- Malária/SVS/MS
1.1 O PARASITO E CICLO EVOLUTIVO
O Plasmodium é um parasito unicelular que acomete várias espécies de animais,
infecta seu hospedeiro e causa-lhes várias complicações, principalmente em humanos. É um
gênero composto de 150 espécies, sendo 4 delas com capacidade de infectar humanos. São
agrupadas na seguinte taxonomia (Levine, 1980; disponível em www.ncbi.nlm.nih.gov):
Reino: Protista
Filo: Apicomplexa
Classe: Aconoidasida
Ordem: Haemosporida
Família: Plasmodiidae
Gênero: Plasmodium
Espécies: Plasmodium falciparum
Plasmodium vivax
Plasmodium malariae
Plasmodium ovale
Os esporozoítos, entre 10 e 100 (PHILLIPSON, 1986 apud ROSENBERG et al.,
1990), são liberados das glândulas salivares da fêmea do mosquito do gênero Anopheles,
durante o repasto sanguíneo, e, dentro de 30 a 45 minutos invadem os hepatócitos. Para

18
atingir os hepatócitos, os esporozoítos necessitam do reconhecimento e interação com o seu
receptor de modo a invadir a célula hospedeira. Os peptídeos que fazem parte da proteína de
superfície do esporozoíto, as proteínas circunsporozoíto (CSP) e a TRAP (Thrombospondin-
Related Adhesive Protein), foram apontadas como os responsáveis pela interação com os
receptores do hepatócito (PHILLIPSON, 1986 apud ROSENBERG et al., 1990; ROBSON et
al., 1995).
Em P. vivax e P. ovale, alguns dos esporozoítos se desenvolvem em hipnozoítos,
permanecendo por meses e até anos, de modo latente no hepatócito, reativando-se após um
período, o que resulta numa recaída do paciente à doença e que serve de vantagem para o
parasito, uma vez que o ciclo hepático pode se completar, fato importante para o sucesso
reprodutivo do parasito (PHILLIPSON, 1986 apud ROSENBERG et al., 1990).
Os esporozoítos, após a invasão nos hepatócitos, ficam contidos no vacúolo
parasitófago, formado pela membrana do hepatócito. Ali se multiplicam e se desenvolvem em
milhares de merozoítos até um pouco antes de serem liberados para a corrente sanguínea.
(PRUDÊNCIO; RODRIGUEZ; MOTA, 2006).
O crescimento e a esquizogonia no fígado para parasitas da malária humana levam
cerca de 6 a 15 dias dependendo das espécies, no qual após esse ciclo pré-eritrocítico,
milhares de merozoítos são liberados (merossomos) para o sangue através dos sinusóides e,
dentro de 15 a 20 segundos invadem os eritrócitos. O reconhecimento e a ligação são devidos
a interação receptor-ligante, e pelo menos para o P. vivax e P. falciparum, as moléculas
envolvidas são diferentes, a Duffy, para P. vivax e glicoforinas e EBA 174 para P. falciparum,
Esta fase proliferativa intracelular sanguínea conduz a todos os sinais e manifestações clínicas
da doença (BECKER et al., 2004 apud BUCHHOLZ et al., 2008). Durante o
desenvolvimento intra-eritrocítico são produzidas tanto formas assexuadas quanto as formas
sexuadas imaturas. As formas assexuadas são os trofozoítos, conhecidas como anéis, e os
esquizontes; já as formas sexuadas são os gametócitos (macro e microgametócito), formados
em pequena proporção quando comparadas às formas assexuadas e correspondendo aos
únicos responsáveis pela continuação do ciclo do Plasmodium no vetor - ciclo esporogônico.
(AKOMPONG et al., 2000). Na fêmea do mosquito Anopheles, os gametócitos ingeridos no
repasto sanguíneo sofrem maturação no meio circulante, ocorrendo à fusão do microgameta
(gametócito masculino) com o macrogameta (gametócito feminino) com conseqüente
formação do zigoto. Este após um período de maturação se diferencia em oocineto, célula
móvel, que atravessa a parede do intestino médio, escapando da matriz peritrófica, e
encistando-se na lâmina basal, camada que separa a hemocele do intestino médio. Uma vez

19
formado os oocistos, se iniciam as divisões mitóticas (esporogonia), resultando na formação
dos esporozoítos. Após a ruptura dos oocistos, os esporozoítos migram através da hemocele
para as glândulas salivares (aproximadamente 7 a 18 dias após a ingestão dos gametócitos),
onde ficam alojados até que sejam inoculados num novo hospedeiro susceptível durante o
repasto sanguíneo. Todas as etapas do ciclo de vida se dão por mecanismos de divisão
haplóide, exceto a etapa de fertilização que é diplóide, onde imediatamente após a fecundação
ocorrem duas etapas de divisão meiótica e que resulta numa célula que contém um núcleo
com quatro genomas haplóides (PHILLIPSON, 1986 apud ROSENBERG et al., 1990). A
divisão meiótica, após a fusão dos gametas, permite a ocorrência de recombinação genética,
que se reflete na composição genética dos esporozoítos, associados a menor ou maior
sensibilidade aos antimaláricos usuais. A figura 04 resume o ciclo evolutivo do Plasmodium.

20
Figura 04- Ciclo biológico de Plasmodium falciparum
Fonte: Adaptado de PHILLIPSON (1986).

21
1.2 MANIFESTAÇÕES CLÍNICAS
O período de incubação da malária varia de acordo com a espécie de plasmódio,
sendo de 8 a 12 dias para P. falciparum, 13 a 17 para P. vivax e 28 a 30 dias para P. malariae.
Uma fase sintomática inicial, caracterizada por mal-estar, cefaléia, cansaço e
mialgia, geralmente precede a clássica febre da malária. O ataque paroxístico inicia-se com
calafrio que dura entre quinze minutos e uma hora, sendo seguido por uma fase febril, com
temperatura corpórea em torno de 40oC. Após um período de duas a seis horas, ocorre
defervescência da febre e o paciente apresenta sudorese profusa e fraqueza intensa.
Após a fase inicial, a febre assume um caráter intermitente, dependente do tempo
de duração dos ciclos eritrocíticos de cada espécie de plasmódio: 36-48 horas para P.
falciparum e 48 horas para P. vivax (malárias terçãs); 72 horas para P. malariae (malária
quartã). Entretanto, a constatação desta regularidade é pouco comum nos dias atuais, em
decorrência do tratamento precoce realizado ainda na fase de assincronismo das
esquizogonias sanguíneas; de infecções por populações distintas de plasmódios e/ou, por
primoinfecção, no qual se constata o retardo da resposta imune específica em pessoas
acometidas nessas circunstâncias (MINISTÉRIO DA SAÚDE, 2001).
Dentre os indivíduos com a doença, a maioria apresenta apenas um quadro febril,
embora possam ocorrer problemas bem mais graves, tais como anemia grave, angústia
respiratória aguda e até malária cerebral, quantificando elevados números de mortes por ano
de cerca de 1-3 milhões de casos, principalmente em crianças (HALDAR et al., 2007). Na
África sub-saariana, onde a transmissão é intensa por P. falciparum, estima-se que 100.000
mortes de fetos ocorram a cada ano de um total de mais de 50 milhões de gestações.
(GUYATT; SNOW, 2004).

22
1.3 DIAGNÓSTICO
O controle da malária depende da rapidez e da forma correta de realização dos
métodos diagnósticos, bem como seu imediato tratamento, uma vez que aspectos clínicos,
como a febre típica, podem ainda não estar presente. O procedimento diagnóstico
frequentemente adotado é a realização de esfregaços com gota espessa e fina de sangue,
coloração pelo Giemsa - corante bastante difundido no Brasil devido a sua alta sensibilidade e
especificidade no diagnóstico da doença - e subsequente leitura em microscópico por
especialista capacitado em identificar a espécie de Plasmodium envolvido na infecção
(LALLOO et al., 2007; AVILA; FERREIRA, 2000). Os esfregaços são obtidos diretamente
de punção digital, tanto a gota espessa quanto a gota fina podem ser feitas, sendo que a gota
espessa aumenta 20-30 vezes a probabilidade de encontrar parasita quando comparado com o
esfregaço, tornando-se assim muito mais sensível que esta última (WARHURST;
WILLIAMS, 1996). Mais recentemente, o diagnóstico da malária também tem sido feito
através testes imunocromatográficos (WORLD HEALTH ORGANIZATION, 2009). Contudo
o “Padrão ouro” continua sendo a identificação do parasito em esfregaços sanguíneos corados.
A imunocromatografia rápida baseada na detecção de antígeno de P. falciparum e
P. vivax está disponível em alguns lugares do mundo, inclusive no Brasil, onde tal
metodologia para o diagnóstico da malária foi conseguida a partir de 1993, com o
desenvolvimento de ensaios rápidos baseados na captura qualitativa de um antígeno de P.
falciparum, a proteína 2 rica em histidina (PfHRP2), conhecidos comercialmente como
ParaSight-F® (Becton & Dickinson) e ICT Malária Pf® (ICT Diagnostics). Estes testes
consistem em fitas de papel de nitrocelulose contendo anticorpo monoclonal específico contra
peptídeos da PfHRP2 são a base do teste. Uma gota de sangue é lisada com detergente e a ela
aplica-se a fita, sobre a qual o lisado é lentamente absorvido. A reação da PfHRP2 com o seu
anticorpo monoclonal é revelada pela adição de anticorpo policlonal anti-HRP2 conjugado
com lipossomas contendo o corante sulfo-rodamina. Para controle do teste, a PfHRP2 é
também fixada à fita, 8 mm acima do anticorpo monoclonal. Tanto a fita quanto os reagentes
são extremamente estáveis, sendo adequados para trabalhos de campo e levantamentos
epidemiológicos, onde a temperatura e a umidade são geralmente elevadas e muito variáveis.
A sensibilidade superior a 95% são observadas quando o ParaSight-F® é comparado à gota
espessa e com parasitemias superiores a 60 parasitos/µl. Entretanto, com parasitemias
menores, a sensibilidade diminui drasticamente, sendo inferior a 60% nos casos de
parasitemia iguais ou inferiores a 10 parasitos/µl. Contudo, pela sua praticidade e facilidade

23
de realização, tanto o ParaSight-F® como o ICT Malária Pf® têm sido considerados testes
úteis para a triagem e mesmo para a confirmação diagnóstica, principalmente em situações
onde é complicado processar o exame da gota espessa, como áreas de difícil acesso, ou em
situações de baixa parasitemia. No entanto, a PfHRP2 só é produzida pelo P. falciparum, não
sendo possível, portanto, diagnosticar outras espécies de plasmódios com esses testes. Isto
representa um inconveniente para a nossa realidade, já que o P. vivax é a espécie mais
prevalente no Brasil. Mais recentemente, outro método de diagnóstico rápido foi
desenvolvido, com a vantagem de poder capturar antígenos de P. falciparum e P. vivax
simultaneamente, trata-se do OpitMAL®, um teste também baseado em fitas de detecção por
imunocromatografia, o qual utiliza anticorpos policlonais e monoclonais marcados com ouro
e dirigidos contra a enzima lactato desidrogenase específica para as duas espécies (pLDH),
presente no sangue total do paciente. Esta é uma enzima intracelular produzida em
abundância pelos plasmódios vivos, o que permite diferenciar entre fase aguda e
convalescença da infecção. Apesar de promissores, os estudos de campo que foram feitos até
o momento para determinar a sua efetividade no diagnóstico da malária não são suficientes
para recomendar a sua utilização no Brasil (MINISTÉRIO DA SAÚDE, 2001).
Três técnicas usando fluorescência para diagnosticar o parasito da malária têm
sido descritas: o QBC11 – Quantitative Buffy Coat Test – um kit comercial (CLENDEMAN,
LONG; BAIRD, 1995), o laranja de acridina de Kawamoto (GAY et al., 1996) e o uso do
BCP - benzotiocarboxipurina (COOKE et al., 1992). Estas três técnicas são fáceis de
manipulação e relativamente rápidas, demonstrando sensibilidade e especificidade equivalente
àquela da gota espessa.
Ambos os métodos QBC e Kawamoto usam laranja de acridina como fluorocromo
para corar os ácidos nucléicos de qualquer amostra de parasitas da malária; já o BCP é outro
fluorocromo. Apesar de ser o corante laranja de acridina um corante fluorescente forte, ele é
inespecífico e cora ácidos nucléicos de células de todos os tipos, consequentemente a
microscopia usando o laranja de acridina tem de ser cautelosa (DELACOLLETT;
VANDERSTUYFT, 1994). Outro fator importante é o elevado custo desta técnica devido aos
aparatos necessários, como por exemplo, o microscópio de imunofluorescência.
O diagnóstico da malária baseado na detecção de ácido nucléico mostrou grande
progresso em termos de eficácia. Além disso, com a extensa utilização da PCR (Polymerase
Chain Reaction - Reação em Cadeia da Polimerase) para o diagnóstico de outras doenças, as
técnicas de extração e purificação de DNA foram aprimoradas e simplificadas. Contudo, o
diagnóstico de malária através da PCR ainda é restrito aos grandes laboratórios, em virtude do

24
custo elevado, reagentes necessários e alta complexidade técnica no campo. (SMEGO; BEG,
2001).
1.4 CONTROLE
A situação da malária no mundo pode ser dividida em três categorias. Em
primeiro lugar, há áreas onde a transmissão da malária é intensa e que até agora tem sido
profundamente afetadas em relação aos programas de controle de vetores, como a África
tropical. A segunda categoria representa os países da Ásia e das Américas, onde programas de
controle de vetores tenham sido interrompidos ou levados a um colapso gradual devido à falta
de compromisso e ingerências por parte dos gestores, ocasionando focos da doença. Na
terceira categoria estão colocadas as áreas de rápido desenvolvimento econômico e os países
gravemente afetados por perturbações sociais, os quais podem conduzir às perturbações
ambientais, migração de população, e a ausência de infra-estrutura nos cuidados com a saúde.
A malária é uma doença focal, que difere em suas características de país para país e mesmo
dentro do mesmo país (PHILLIPSON, 1986 apud ROSENBERG et al., 1990).
Vários fatores devem ser considerados quanto ao controle da malária: (i) a
presença de mudanças sociais e ecológicas; (ii) características biológicas, antropológicas,
culturais e sociais da população; (iii) intensidade e periodicidade da transmissão da malária;
(iv) espécies dos mosquitos vetores, seu comportamento e sua susceptibilidade aos
inseticidas; (v) características dos serviços de saúde existentes e, naturalmente, (vi) espécies
dos parasitos e sua sensibilidade aos antimaláricos usuais (PHILLIPS, 2001). Para tanto, o
controle da malária exige uma abordagem integrada, compreendendo a prevenção e controle
vetorial, bem como o tratamento com antimaláricos (WORLD HEALTH ORGANIZATION,
2006).
As estratégias atuais para reduzir a malária consistem basicamente do controle de
vetores e a quimioterapia, uma vez que não existem ainda vacinas eficientes.
O controle do vetor é obtido através da redução da densidade vetorial,
interrompendo seu ciclo de vida, e criando uma barreira entre o hospedeiro humano e os
mosquitos (TAYLOR; WHITE, 2004). Dessa forma, faz-se necessária a adoção de medidas
de prevenção individual e coletiva, tais como: (i) evitar aproximação das áreas de risco após o
entardecer e logo ao amanhecer, (ii) utilizar repelentes em áreas expostas do corpo, usar
mosquiteiros e ainda, (iii) vedar portas e janelas com telas. Medidas de controle de vetores

25
também devem ser adotadas. Nesse sentido, métodos como borrifação com inseticidas de ação
residual nas paredes, uso de larvicidas, medidas de saneamento básico a fim de evitar os
criadouros do mosquito e educação sócio-ambiental, com objetivo de provocar mudanças de
atitude da população em relação aos fatores que facilitam a exposição à transmissão, devem
fazer parte da realidade da população a fim de que se tenha um controle mais eficaz contra a
malária (MINISTÉRIO DA SAÚDE, 2001).
No tocante às principais estratégias no combate à malária quanto ao paciente com
suspeita de malária, faz-se necessário a identificação precoce do plasmódio (MINISTÉRIO
DA SAÚDE, 2006) e o tratamento medicamentoso, feito com a associação de pelo menos
dois antimaláricos diferentes, com o intuito de intervir em pelo menos duas das fases do ciclo
do parasito (figura 04) (SHAPIRO; GOLDBERG in BRUNTON, LAZO; PARKER, 2006).
Portanto, uma associação medicamentosa deve incluir: esquizonticidas sangüíneos,
gametocitocidas, e, para o P. vivax, hipnozoiticidas.
Os objetivos de uma política de tratamento com antimaláricos são: assegurar
rapidamente a cura da infecção, reduzir a morbidade e mortalidade, incluindo a malária
relacionada com anemia; impedir a progressão da malária não complicada em grave e
potencialmente fatal doença; reduzir o impacto da infecção sobre o feto durante a gravidez;
reduzir o reservatório de infecção; impedir o aparecimento e disseminação de resistência a
drogas; e prevenir a malária em viajantes (WORLD HEALTH ORGANIZATION, 2008).
Em virtude da sua estrutura química e atividade nas diferentes fases do ciclo de
vida do Plasmodium, Warhurst (1986) classificou as drogas antimaláricas em três grupos:
antimetabólitos, 8-Aminoquinolinas e o grupo dos esquizonticidas sanguíneos. Já o Manual
de Terapêutica da Malária (2001) preconiza a classificação das drogas antimaláricas, segundo:
a) O grupo químico em: arilaminoálcoois (quinina, mefloquina e halofantrina), 4-
aminoquinolinas (cloroquina e amodiaquina), 8-aminoquinolinas (primaquina), peróxido de
lactona sesquiterpênica (derivados da artemisinina), naftoquinonas (atovaquona) e antibióticos
(tetraciclina, doxiciclina e clindamicina);
b) Alvo de ação no ciclo biológico em: esquizonticidas teciduais ou
hipnozoiticidas (cura radical do P. vivax), esquizonticidas sanguíneos (promovem a cura
clínica), gametocitocidas (bloqueia a transmissão) e esporonticidas (impede a infecção pelos
esporozoítos). Infelizmente, até o momento, nenhuma droga deste último grupo é disponível
para uso em humanos.
A cloroquina, droga esquizonticida caracterizada pelo preço acessível e bastante
disponível, foi no passado a droga mais promissora no controle da malária, mas se tornou

26
ineficaz na maioria dos casos de malária P. falciparum em áreas endêmicas, fato que
aconteceu também com outras drogas, como a sulfadoxina-pirimetamina. A descoberta da
artemisinina e o desenvolvimento de seus derivados é o que se tem de mais eficiente para o
combate da malária. As combinações terapêuticas a base de artemisinina (ACTs) são agora
geralmente consideradas como o melhor curso de tratamento de malária por P. falciparum não
complicada no mundo (WORLD HEALTH ORGANIZATION, 2006). Neste caso, a terapia
de combinação se torna uma alternativa que “atrasa” a aquisição da resistência do parasito aos
antimaláricos, já que a probabilidade de desenvolvimento de resistência simultânea a dois
agentes quimioterápicos com independentes mecanismos de ação é extremamente baixa, da
ordem de 1012 para cada droga do tratamento (WHITE, 1998). A demora e a pouca precisão
diagnóstica, o emprego de terapêutica inapropriada, a qualidade dos fármacos e o uso
indiscriminado de antimaláricos contribuem para a alta incidência e disseminação de cepas de
Plasmodium falciparum multi-resistentes. Na figura 05 pode-se observar a dispersão mundial
da resistência à cloroquina e sufadoxina-pirimetamina (NORONHA et al., 2000).
Figura 05- Resistência de Cloroquina e Sulfadoxina-pirimetamina no mundo.
Fonte: NORONHA et al., 2000.
Até o momento não existe uma droga ideal, com as características da cloroquina:
baixo custo, pouca toxicidade e eficácia. Em geral, todas as drogas causam toxicidade e
efeitos adversos, principalmente quando a dose da droga administrada é maior que a
recomendada, fato também visto para as drogas utilizadas na profilaxia da malária, efeitos que

27
podem variar de leves, graves e às vezes fatais (CROFT et al., 2002). Além disso, os custos
desses medicamentos são bastante elevados. Devido ao restrito arsenal profilático e
terapêutico, torna-se urgente o desenvolvimento de novos antimaláricos.
1.5 A ETNOFARMACOLOGIA
A abordagem etnofarmacológica refere-se a uma triagem racional baseada na
seleção de plantas usadas na medicina popular; é uma ferramenta que tem se mostrado
bastante promissora e satisfatória no descobrimento de novos medicamentos. Um bom
exemplo disto é a experiência chinesa na descoberta do potente antimalárico artemisinina
(ginghaosu), como parte do programa de pesquisas de novos medicamentos a partir de plantas
medicinais (revisado por PHILLIPSON, 1986 apud ROSENBERG et al., 1990).
A flora brasileira possui uma imensa fonte inexplorada de moléculas com grande
potencial farmacológico. Alguns exemplos de substâncias têm sido largamente utilizados
pelos povos indígenas, como extratos de folhas de jaborandi (Pilocarpus jaborandi), sementes
de guaraná (Paullinia cupana) ricas em cafeína usadas como estimulante e curares extraídos
de folhas de Chondondendrom spp. usados em técnicas anestésicas (ANDRADE-NETO et al.,
2007).
O surgimento de cepas de Plasmodium falciparum resistentes à cloroquina na
década de 60, principalmente na América do Sul e no sudeste asiático, foi um fator que
desencadeou uma preocupação em todo o mundo, tornando o desenvolvimento de novas
drogas para o tratamento da malária uma prioridade (FANDEUR; MORETTI; POLONSKY,
1985 apud SARAIVA et al., 2003; PETERS 1985). Além é claro da emergência real,
referente ao acometimento de centenas de milhões de pessoas pela malária em todo o mundo e
às limitações apresentadas pelas drogas antimaláricas conhecidas atualmente. Assim, é crucial
a introdução de novas drogas, que possuam mecanismos de ação diferentes dos existentes, ao
arsenal quimioterapêutico disponível (O’NEILL et al., 1986 apud DEHARO et al., 2001;
RIDLEY, 2002).
Estudos etnobotânicos revelaram o uso de várias espécies vegetais na medicina
tradicional no tratamento da malária, como Bidens pilosa, Cantharanthus roseus, Cassia
fistula, Cinchona calisaya, Cuphea ingrata, Geissospermum sericeum, Jateorrhiza palmata,
Quassia amara, Simaba ferruginea, e Strychnos pseudoquina. Outras, já tiveram inclusive

28
suas atividades antimaláricas testadas contra o Plasmodium, destacando-se as espécies:
Bathysa cuspidata, Cosmos sulphureus, Cecropia hololeuca, Erisma calcaratum, Gomphrena
arborescens, Musa paradisiaca, Ocotea odorifera, e Pradosia lactescens, algumas plantas são
utilizadas em associação com objetivo de aumentar a potência do tratamento. A maioria das
plantas antimaláricas pertence às famílias - Asteraceae, Rubiaceae, Apocynaceae e
Simaroubaceae (BOTSARIS, 2007).
Como um dos países de maior biodiversidade do mundo, contando com 20% do
número total de espécies do planeta, muitas dessas endêmicas, o Brasil tem um promissor
caminho utilizando a abordagem etnofarmacológica em busca de novas drogas mais ativas
contra a malária.
Diante do exposto, segue abaixo breves comentários sobre as espécies vegetais
testadas neste estudo tendo por base a abordagem etnofarmacológica para algumas das
espécies estudadas, bem como a utilização de outras espécies tendo em vista a indicações/ou
amenização de sintomas ocorridos na malária.
Ximenia americana Lineu, Oleacaceae
Esta planta é conhecida popularmente como ameixeira (Fig. 06) e é pertencente à
família das Oleacaceae, a qual tem inúmeras plantas com propriedades farmacológicas. É
uma planta originada na África, mas pode ser encontrada nas mais diversas regiões do mundo,
como na Índia, ilhas do Pacífico, Nova Zelândia, América Central e do Sul (ARBONNIER,
2004).
A Ximenia americana é uma árvore muito variável em tamanho, sendo encontrada
medindo até 5 m de altura. É, muitas vezes, semiparasita, com fortes espinhos, tronco fino e
dispersas nos trópicos, em regiões da África, América e Ásia. Uma pesquisa realizada entre a
população do Mali verificou o seu uso para diversos males, dentre eles: contra infecção na
garganta, malária e dismenorréia, sendo as raízes e as folhas as partes da planta mais
frequentemente usadas (GRØNHAUG et. al., 2008).
Diversos estudos têm demonstrado que esta planta tem propriedades anti-
neoplásicas, atividade microbicida e ação antiinflamatória (OGUNLEYE; IBITOYE, 2003;
JAMES et al., 2007). A despeito de sua grande utilização, pouco tem se relatado na literatura
acerca da toxicidade de diferentes partes desta planta.

29
A X. americana também é utilizada para outros fins, como úlcera hansênica,
infecções cutâneas (OGUNLEYE; IBITOYE, 2003) e dores de cabeça. Também pode ser
usado para resfriados e como laxante. A infusão das folhas é usada como colírio, e para a
constipação, dor de dente, e angina. As raízes também podem ser usadas para tratar doenças
venéreas, edema e também como um antídoto contra o veneno de cobras. Na África tropical, a
folha é utilizada para remédio contra tosse e febre.
No Brasil, a espécie ocorre na caatinga, na medicina popular é usada como
adstringente em banhos de mulheres com menstruação excessivas, em lavagem de feridas e na
forma de pó para cicatrização de feridas e úlceras (MAIA, 2004). Dentre os muitos usos, está
ainda à maceração de raízes e a aplicação de uma espécie de “emplastro” obtida daquela em
feridas de crianças doentes com sarampo (OMER; ELNIMA, 2003).
Figura 06- Ximenia americana (tronco e folhas)
Fonte: http://www.zimbabweflora.co.zw/speciesdata/image-display.php?species_id=121580&image_id=2
Maytenus rigida Martius, Celastraceae
A espécie Maytenus rigida pertence à família Celastraceae. É uma família de
árvores e arbustos, compreendendo cerca de aproximadamente 89 gêneros e 1300 espécies,
com o gênero Maytenus como um dos mais importantes, com cerca de 270 espécies
distribuídas por todo o territorio brasileiro (STEVENS, 2001).
A espécie Maytenus rigida Mart. (Fig. 07), popularmente conhecida como bom-
nome ou espinheira-santa, é considerada nativa no nordeste do Brasil, encontrada em

30
ambientes de caatinga e cerrado tem distribuição em todo o território brasileiro, com registros
para os estados do Rio Grande do Norte, Piauí, Ceará, Paraíba, Pernambuco, Alagoas,
Sergipe, Bahia, Maranhão, Minas Gerais, Espírito Santo, São Paulo e Rio de Janeiro (SALES,
MAYO; RODAL, 1998). Tradicionalmente utilizadas pelos indígenas, às folhas, em infusão,
de diversas espécies de Maytenus existentes no Brasil, são utilizadas contra afecções gástricas
(hiperacidez, úlceras gástricas, duodenais e gastrite crônica); diversos relatos científicos
comprovaram essas atividades (CARLINI, 1988). Entretanto, estudos etnobotânicos mostram
a indicação popular também para outras finalidades, como, por exemplo, depurativo do
sangue e no tratamento de diabetes, problemas no sistema urinário e problemas intestinais
(MARIOT, 2005), além de ser usada na medicina tradicional da América do Sul como
infusões ou decocções, devido à sua atividade analgésica, antiinflamatória e antiulcerogênica
(CORRÊA, 1984 apud MOTA et al., 2008; BUENO et al., 2005). Outros trabalhos citam o
uso tradicional da espinheira-santa como anticonceptivo, abortivo, vulnerário (para curar
feridas), anti-séptico, anti-espasmódico, diurético, anti-asmático, anti-tumoral, laxativo, cura
do vício da bebida e enfermidades do fígado, para tratar a hidropisia devido ao abuso do
álcool e para reduzir a produção de leite em quem amamenta (SCHEFFER, 2001 in REIS;
SILVA, 2004).
Figura 07- Maytenus rigida (folhas)
Fonte: http://caatingafaunaeflora.blogspot.com/2009/04/bom-nome-maytenus-rigida.html

31
Sideroxylon obtusifolium Penn., Sapotaceae
A espécie Sideroxylon obtusifolium, conhecida como “quixabeira” (Fig. 08), é
uma planta medicinal pertencente à família Sapotaceae. Sua ocorrência vai desdeo Sudoeste
até o Nordeste do Brasil, possuindo propriedades antiinflamatórias e hipoglicêmicas
(BELTRÃO et al., 2008). Essa ideia também é defendida por Lorenzi (2002), que além de
citar o amplo uso de cascas de caule e raízes de Sideroxylon obtusifolium na medicina caseira
no interior do Nordeste, relata que a mesma possui propriedades adstringentes, tônicas,
antiinflamatórias e antidiabéticas. Segundo o mesmo autor, a planta é usada na forma de
decocto, infusão e a maceração da casca do caule ou raiz.
Além dos usos já citados, os frutos da “quixabeira” são consumidos na dieta
alimentar de forma ocasional, e seu uso não parece representarem risco de extinção para a
espécie, diferentemente de outras espécies, onde a excessiva exploração, principalmente para
fins de queima (produção de combustível pelo carvão) ou de fonte de madeiramento para a
construção de casas, representa risco iminente para a biodiversidade brasileira (LUCENA et.
al., 2007).
Figura 08- Sideroxylon obtusifolium (tronco e folhas)
Fonte:http://w3.ufsm.br/herbarioflorestal/especie_detalhes.php?nome_filtrado=espinheiro

32
Stryphnodendron coriaceum Benth, Leguminosae
Popularmente conhecido como "barbatimão" (Fig. 09), esta planta tem sido
utilizado no tratamento de diversas doenças e particularmente em casos de úlceras de pele e,
também como anti-séptico. O gênero Stryphnodendron, pertence à família Leguminosae-
Mimosoideae que comporta 32 variedades, sendo uma espécie tipicamente brasileira
(OCCHIONI, 1990 apud MOTA et al., 2008). Nesse gênero encontram-se algumas espécies
que são tóxicas para animais (BRITO et al., 2001).
Figura 09- Stryphnodendron coriaceum (folhas)
Fonte: http://zipcodezoo.com/Key/Plantae/Stryphnodendron_Genus.asp
Bowdichia virgilioides Kunth, Leguminosae
A planta Bowdichia virgilioides Kunth (Leguminosae-Papilonoideae) (Fig. 10) é
uma árvore de tamanho médio encontrada em florestas tropicais da América do Sul.
Bowdichia virgiloides é comumente encontrada ao longo da Amazônia brasileira e boliviana.
No nordeste do Brasil ela é conhecida na medicina popular com o nome de “sucupira”, e sua
casca é tradicionalmente usada contra úlceras e anti-diabéticas (MACEDO; FERREIRA,
2004). As sementes são usadas no tratamento de reumatismo, artrite e doenças de pele

33
(CRUZ, 1965 apud THOMAZZI et al., 2010). O seu uso se dá na forma de chás preparados
com uma quantidade pequena de folhas secas, cascas ou sementes para cada 1 litro de água
fervente, sendo recomendado um copo do chá três vezes ao dia. Outra forma do uso da planta
é maceração de suas partes para aplicação tópica (BRANDÃO et al, 2006).
Figura 10- Bowdichia virgilioides Kunt.
Fonte:http://www.fiocruz.br/ccs/cgi/cgilua.exe/sys/start.htmfrom_info_index=631&infoid=2352&sid=9&tpl=printerview
Schinopis brasiliensis Engler, Anacardiaceae
A espécie Schinopis brasiliensis Engler (Fig. 11) é popularmente conhecida como
“baraúna” ou “braúna”, pertence à família Anacardiaceae e ocorre na Caatinga desde a Bahia
até a Paraíba (BRAGA, 1976 apud DANTAS et al., 2008), tendo poucos representantes do
Rio Grande do Norte ao Piauí (ANDRADE–LIMA, 1989 apud OLIVEIRA; OLIVEIRA,
2008). É uma grande árvore conhecida no Nordeste brasileiro devido a sua utilização no
tratamento de verminoses em animais e sua importância madeireira, motivo pelo qual esta
espécie encontra-se em extinção. Trabalhos anteriores com os extratos orgânicos do caule de
S. brasiliensis permitiram isolar triterpenos, esteróides, derivados do ácido gálico, depsídeos e
um n-alquil-fenol inédito (CARDOSO et al., 2005), provavelmente importantes para a sua
atividade biológica.
Algumas espécies de Anacardiaceae são estudadas quanto ao seu caráter
alergênico evidenciado por intensa dermatite de contato em indivíduos sensíveis. A literatura

34
científica atribui esse caráter a presença de alquil e alquenil fenóis, catecóis, resorcinóis,
hidroquinonas ou à mistura destes (conhecidas como urushiol).
Figura 11- Aspecto geral e detalhe foliar de Schinopis brasiliensis.
Fonte: Arizona State University Herbarium. Disponível em: < http://swbiodiversity.org/seinet/taxa/imgdetails.php?imgid=219701>.
Picrolemma sprucei Hook, Simaroubaceae
A espécie Picrolemma sprucei Hook (sin. P. pseudocoffea Ducke), conhecida
popularmente pelo nome “caferana” (Fig. 12) e da família Simaroubaceae, é um arbusto que
atinge até 2,5 m de altura, e está amplamente distribuída na Amazônia Ocidental (LE
COINTE, 1947 in SARAIVA et al., 2003; GRENAND, MORETTI; JACQUEMIN, 1987 in
SARAIVA et al., 2003).
Tanto no Brasil como no Chile, o chá das folhas e das raízes é usado no
tratamento de gastrite, como febrífugo e anti-helmíntico (LE COINTE, 1947 in SARAIVA et
al., 2003; DUKE; VASQUEZ, 1994 in SARAIVA et al., 2003).
Na Guiana Francesa, o estudo químico dos caules, raízes e folhas P. sprucei
resultou no isolamento dos quassinóides sergeolida, isobruceína B e 15-desacetil-sergeolida
(MORETTI et al., 1982 in SILVA et al., 2009; POLONSKY et al., 1984 in SILVA et al.,
2009). Do caule e raízes de espécimes nativas da Amazônia brasileira, foram isolados os
quassinóides neosergeolida e isobruceína B (SCHPECTOR et al., 1994 ; SARAIVA, 2001 in
SARAIVA et al., 2003). Algumas dessas substâncias apresentaram atividade in vitro contra

35
leucemia e malária (MORETTI et al., 1982 in SILVA et al., 2009; FANDEUR et al., 1985 in
SARAIVA et al., 2003; POLONSKY et al., 1984,SILVA et al., 2009).
Quimicamente, os quassinóides são derivados da degradação de compostos
triterpênicos, que são altamente oxigenados. Muitos quassinóides apresentam uma ampla
gama de atividades biológicas in vitro e/ou in vivo, incluindo antitumoral, antimalárica,
antiviral, anti-inflamatória, anti-inseticida, amebicida, antiúlcera e atividades herbicidas.
Bruceantina, simalikalactona D, quassine, brusatol e glaucarubinona são alguns dos
quassinóides mais bem estudados e apresentam uma grande variedade de atividades
biológicas (GUO et al. 2005).
A atividade biológica de alguns dos compostos encontrados em P. sprucei, como
toxicidade usando a Artemia franciscana, citotoxicidade em células tumorais humanas,
atividade larvicida em Aedes aegypti e atividade antimalárica in vitro e in vivo foram
descritas pelo nosso grupo (Silva et al. 2009).
Figura 12- Picrolemma sprucei
Fonte: http: <//f m2.fmnh.org/plantguides/results.asp?lang=br&intPerPage=50&pagenum=248>.

36
2 OBJETIVOS
Geral
Avaliar a atividade biológica de extratos e frações de plantas medicinais dos
Biomas da Caatinga e Amazônico a partir da abordagem etnofarmacológica.
Específicos
� Testar a atividade esquizonticida in vivo dos extratos brutos das espécies vegetais
estudadas em camundongos infectados com o Plasmodium berghei;
� Testar a atividade esquizonticida in vitro de extratos e frações das contra o
Plasmodium falciparum;
� Avaliar a citotoxicidade dos extratos vegetais estudados.

37
3 MATERIAL E MÉTODOS
3.1 ESPÉCIES VEGETAIS E PREPARO DE SEUS EXTRATOS E FRAÇÕES
A tabela 1 apresenta as espécies vegetais utilizadas neste estudo.
Tabela 1- Extratos e das frações das diferentes espécies vegetais brasileiras utilizados nos
testes antimaláricos in vivo e in vitro.
Espécie Parte da
planta
Extrato /Frações Bioma
Ximenia americana Caule Alcoólico Caatinga -RN
Maytenus rigida Entrecasca Alcoólico Caatinga - SE
Sideroxylon
obtusifolium
Entrecascas Alcoólico/clorofórmico/
hexânico/aquoso/acetato
de etila/hidrometanólico
Caatinga -SE
Stryphnodendron
coriaceum
Entrecasca Alcoólico/clorofórmica/
hexânica/aquoso/acetato
de etila/hidrometanólico
Caatinga- SE
Bowdichia virgiliodes Folhas Alcoólico Caatinga-SE
Schinopis brasiliensis Caule Alcoólico/clorofórmica/
hexânica/aquoso/acetato
de etila/hidrometanólico
Caatinga-SE
Picrolemma sprucei Caule/Raiz Aquoso Amazônico
Os extratos de Picrolema sprucei e de Arrabidaea chica foram fornecidos pelo
grupo do Prof. Adrian M. Pohlit, Laboratório de Princípios Ativos da Amazônia/AM, sendo as
exsicatas depositadas no herbário do INPA/AM.
Os demais extratos utilizados das espécies: Stryphnodendron coriaceum,
Sideroxylon obtusifolium, Bowdichia virgiliodes, Maytenus rigida, Schinopis brasiliensis

38
foram fornecidos pelo grupo do Prof. Charles dos S. Estevam, Departamento de Bioquímica
da Universidade Federal de Sergipe – UFS.
As amostras da X. americana foram coletadas no Centro de lançamento da
Barreira do Inferno, no município de Parnamirim/RN e divididas em duas partes: uma parte
seguiu para a identificação com a conseqüente produção da exsicata e registro no herbário da
UFRN; e outra parte foi destinada à produção do extrato bruto etanólico a serem utilizados
nos testes de toxicidade, citotoxicidade e antimaláricos. O extrato de Ximenia americana foi
preparado no LABMAT seguindo a metodologia padrão: após seco e macerado o material
vegetal foi posto em erlenmeyer e coberto pelo solvente etanol P.A. e deixado em ultrassom
por 45 minutos em tempo de infusão por 24 horas. Seguido o tempo para extração dos
compostos presentes naquela massa vegetal, o conteúdo foi filtrado em papel fitro e colocado
em rotaevaporador (Laboratório de Microbiologia) a uma temperatura de 40°C até que o
solvente evaporasse e então, obtido o extrato bruto.
3.2 MANUTENÇÃO DO Plasmodium berghei
O Plasmodium berghei da linhagem NK-65 foi doado gentilmente pela Dra. Luzia
Helena de Carvalho, do Centro de Pesquisas René Rachou/FIOCRUZ. A cepa é mantida em
camundongos suíços por passagens sangüíneas semanais de animal para animal. A
determinação da percentagem de parasitemia é feita após se confeccionar esfregaços
sangüíneos de camundongos infectados com formas eritrocíticas do P. berghei, fixar com
metanol, corar com solução de Giemsa, e examinar em microscópio ótico (1000x). O animal
doador selecionado, com 10 a 20% de parasitemia, foi sangrado por incisão na ponta da cauda
ou excepcionalmente, pelo plexo orbital (usando pipeta de Pasteur). O sangue foi colhido
com citrato de sódio (anticoagulante) a 3,8%. O número de hemácias por ml de sangue foi
determinado com auxílio de câmara de Neubauer (hemocitométrica), e a partir da
percentagem de parasitemia e do número total de hemácias foi feita a padronização do inóculo
com diluições adequadas do sangue parasitado em PBS-citratado, de modo que cada animal
recebesse 1 x105 hemácias parasitadas, via intraperitoneal, em um volume final de 0,2 ml.
Além disso, a cepa de P. berghei foi mantida congelada em nitrogênio líquido (-
1960C) usando-se 10% de glycerolyte como crioprotetor.

39
3.3 ANIMAIS
Camundongos suíços de 20 ± 2g de peso, provenientes do biotério do
LABMAT/CB/UFRN, foram inoculados com o número de parasita padrão e divididos
aleatoriamente em grupos de 3-5 camundongos por gaiola. Em cada experimento foram
incluídos um grupo controle tratado com placebo e um grupo controle tratado com dose sub-
curativa de cloroquina, além dos grupos testes. A utilização dos animais seguiu protocolo
aprovado pela Comissão de Ética no Uso Animal (CEUA 043/2010) da Universidade Federal
do Rio Grande do Norte.
3.4 TESTE DE TOXICIDADE AGUDA
Devido a pouca quantidade do material vegetal, o teste de toxicidade aguda foi
realizado apenas com o extrato de Ximenia americana. Grupos de 3 camundongos receberam
doses de 10.000mg/kg, 5.000mg/kg, 2.500mg/kg, 1.250mg/kg, 650mg/kg e 312,5mg/kg com
o objetivo de determinar a dose letal (DL-50). Os extratos foram administrados em uma só
dose, no qual cada animal recebeu por via oral um volume de 0,2 ml. Paralelamente, foi
utilizado um grupo controle negativo, consistindo apenas do solvente utilizado (água
destilada). Os animais foram monitorados diariamente após a administração das doses até o
oitavo dia do experimento. Foram verificados os seguintes parâmetros: peso, aspecto do pêlo
(normal ou eriçado), comportamento (sonolentos, agitados, quietos e normais), resposta a
estímulos, frequência cardíaca, frequência respiratória e mortalidade. A mortalidade
cumulativa foi expressa em percentagem, determinando a dose letal para 50% dos animais
(OKOKON & NWAFOR, 2008). A DL50 foi determinada seguindo protocolo padrão
(LORKE 1983; JAMES et al. 2008), por meio da média geométrica da dose que resultou em
100% de mortalidade, bem como daquelas que não resultou em mortalidade completa dentre
os animais do grupo. Os animais sobreviventes foram mantidos em suas respectivas gaiolas e
re-submetidos às mesmas dosagens para mais quatro dias consecutivos, bem como
monitorando os aspectos já descritos.

40
3.5 TESTES ANTIMALÁRICOS IN VIVO
3.5.1 Administracão de extratos aos camundongos infectados
As drogas foram trituradas em gral de massa e diluídas em água destilada.
Devido à baixa solubilidade da maior parte dos extratos brutos de plantas, foram
ressuspendidos com auxílio de DMSO ou tween 20 (1 a 3% concentração final). Nestas
concentrações o DMSO ou tween 20 não interfere significativamente no organismo dos
animais, conforme foi verificado previamente em nosso laboratório. Os camundongos foram
então tratados pela via oral por quatro dias consecutivos. Este teste denomina-se "teste de
supressão de quatro dias" descrito por Peters (1965; 1967).
3.5.2 Avaliação da atividade antimalárica in vivo.
A avaliação da atividade dos extratos e frações das plantas foi feita através da
determinação da percentagem da parasitemia em esfregaços sanguíneos dos camundongos
confeccionados nos 50 e 70 dias após inoculação. Os animais nos diversos grupos (tratados e
não tratados) tiveram seus esfregaços codificados, e contados em teste duplo cego a fim de
evitar tendenciosidade e reduzir a margem de erro. O cálculo da percentagem de redução da
parasitemia pelas drogas testadas foi feita em relação aos controles não tratados após
decodificação dos esfregaços, feita preferencialmente por um segundo observador.
Estabeleceu-se como critério de atividade da droga in vivo a inibição de 30% ou mais do
crescimento do P. berghei (CARVALHO; KRETTLI, 1991).
A mortalidade dos animais foi acompanhada diariamente em todos os
experimentos, até que todos os animais do grupo controle não tratado fossem a óbito.

41
3.5.3 Determinação da percentagem de parasitemia em esfregaços sanguíneos
Os esfregaços sangüíneos, corados pelo Giemsa, foram examinados e as
parasitemias determinadas através da contagem do número de hemácias infectadas em 2000-
3000 hemácias; no caso de parasitemias entre 5 a 20% ou de cerca de 1000 hemácias para
parasitemias maiores. No caso de infecções baixas, menores que 5%, se avaliou o número de
hemácias parasitadas em 6000 células ou 100 campos microscópicos. Essa avaliação foi feita
pela estimativa do número total de hemácias por campo microscópico, e, a partir desta, foi
calculado o número de campos a serem contados. O resultado foi expresso em termos de
percentagem de hemácias parasitadas. Os cálculos de redução (em porcentagem) da
parasitemia basearam-se na fórmula:
Onde: GC é a média de hemácias parasitadas do grupo controle negativo e GT é a
grupo teste.
3.6 TESTES ANTIMALÁRICOS IN VITRO CONTRA P. falciparum
Os ensaios in vitro foram realizados usando a cepa de P. faciparum
multirresistente (K1) doada pelo MR4 (Malaria Research e Reagente de Referência Resource
Center, Manassas, Virgínia, E.U.A.). Os parasitos foram mantidos em cultivo contínuo
segundo método de Trager e Jensen (1976), usando eritrócitos humanos - grupo sanguíneo
A+, em meio RPMI 1640 suplementado com 10% de soro humano.
Os extratos testes foram inicialmente preparados em solução estoque 5mg/mL,
utilizando meio de cultura incompleto, isto é, sem plasma e sem hipoxantina, contendo 0,1-
0,2% de DMSO. A partir daí, foram preparadas diferentes diluições das drogas a serem
testadas (5 e 50ug/mL). Em paraelo foram utilizados, como controle, antimaláricos de
referência.
Para avaliação da susceptibilidade do P. falciparum às drogas, hemácias
parasitadas com aproximadamente 1% de parasitemia e predomínio de formas jovens, foram
ressuspendidas em meio completo para hematócrito de 3%. Esta suspensão foi distribuída em
microplacas (96 poços) de polietileno de fundo chato, colocando-se 100 µl por poço. Após
R= GC –GT x 100 GC

42
aproximadamente 2 horas a 37ºC em estufa úmida de CO2 ou dessecador (tempo necessário
para a sedimentação das hemácias), o meio foi retirado de todos os poços e, esfregaços
sangüíneos foram confeccionados de 6 a 8 poços, escolhidos aleatoriamente, para obtenção da
parasitemia inicial. Isto foi feito apenas com o objetivo de garantir a uniformidade de
distribuição dos parasitos na microplaca. A seguir, acrescentou-se a cada poço meio de cultura
novo (controles) ou meio contendo as diferentes concentrações das drogas testes (extratos ou
frações) ou controles com cloroquina.
Após 24 horas de incubação dos parasitos a 37ºC em atmosfera de CO2 , o meio
de cultura, com ou sem adição das drogas, foi individualmente substituído para cada poço.
Com 48 horas foram confeccionados esfregaços de todas as amostras os quais, após coloração
pelo Giemsa, foram examinados ao microscópio ótico (1000x) para determinação da
percentagem de hamácias parasitadas. A média da parasitemia calculada nos cultivos
controles sem droga foi utilizada na avaliação das taxas de crescimento do parasito nos poços
com as drogas testes e controles com cloroquina. A atividade das drogas foi expressa pela
percentagem de redução da parasitemia em relação aos controles sem drogas. Os
experimentos foram realizados em duplicatas e a leitura feita em lâminas codificadas.
Como critério de atividade in vitro, estabeleceu-se que as drogas que inibiram em
80 a 100% o crescimento dos parasitos foram consideradas ativas (A); parcialmente ativas
(PA) quando a inibição foi de 50 a 79%; e inativas (I) quando a inibição foi menor que 50%,
na concentração mais alta da droga testada.
3.7 TESTES DE CITOTOXICIDADE (IN VITRO)
Os testes de citotoxicidade foram realizados no laboratório de cultura celular do
Departamento de Bioquímica da UFRN, sob a supervisão do Prof. Hugo Alexandre de
Oliveira Rocha. Para os bioensaios foram utilizadas as linhagens celulares: HeLa (carcinoma
cervical humano), HEPG2 (hepatoma humano), 3T3 (linhagem de fibroblasto de embrião de
rato) e CHO (proveniente de ovário de hamster chinês) e Raw (macrófagos de camundongos),
sendo as quatro primeiras linhagens utilizadas para testar a citotoxicidade dos extratos de:
Stryphnodendron coriaceum (barbatimão), Sideroxylon obtusifolium (Quixabeira), Bowdichia
virgilioides (Sucupira), Maytenus rigida (Bom-nome), Ximenia americana (ameixa) e
R= GC –GT x 100 GC

43
Picrolemma sprucei (caferana) nas suas duas formas (extrato aquoso do caule e da raiz), todos
nas concentrações de 100, 200, 400, 800 e 1000 µg/ml; a Raw foi utilizada apenas para o teste
de citotoxicidade de: Stryphnodendron coriaceum (barbatimão), Sideroxylon obtusifolium
(Quixabeira), Bowdichia virgiliodes (Sucupira), Maytenus rigida (Bom-nome), esta última foi
descartada devido à sua insolubilidade no meio, mesmo quando adicionado solventes como o
Tween (emulsificante – polisorbato) ou DMSO (Dimetil sulfóxido). No teste com a Raw
foram utilizadas as concentrações de 720, 576, 432, 288, 144, 72 e 35 µg/ml. O teste com as
amostras dos extratos foram realizados em triplicata.
Inicialmente, as células foram mantidas em culturas contínuas em meio DMEM
(Dulbecco’s Modified Eagle’s Medium), suplementado com 10% de soro fetal bovino, em
atmosfera de microaerofilia (5% CO2, 2% O2 e balanço de N2) a 37º C. A citotoxicidade dos
extratos vegetais foi determinada usando o ensaio colorimétrico methylthiazoletetrazolium
(MTT) (MOSMAT, 1983). Para os ensaios, as células foram tripsinadas, lavadas, suspendidas
em meio DMEM e distribuídas em placas de 96 poços (5 x 103 células por poço), sendo
incubadas por 18 h a 37º C.
Para efeito de comparação com os grupos testes, foram formados grupos controles
positivos: um constituído de meio DMEM com 10% de soro fetal bovino, outro com DMEM
sem soro fetal bovino e ainda um terceiro quando os extratos necessitavam de agente
solubilizante (DMSO) + DMEM a 10% de soro fetal.
Após a aplicação dos extratos, as placas foram incubadas pelos seus tempos (24h
e 48h), retiradas da incubadora para a seqüência de aplicação de MTT (100µL a 1mg/mL em
meio DMEM) – 4h de incubação a 37°C - lavagem com 100 µL de álcool isopropílico a 0,04
M em ácido clorídrico, em cada poço - subseqüente leitura no espectrofotômetro (562nm).
A viabilidade celular foi avaliada através das curvas da concentração dose-
resposta. Resultados foram expressos em média ± desvio padrão.
3.8 ANÁLISE ESTATÍSTICA Os dados obtidos das avaliações antimaláricas in vivo e in vitro foram submetidos
ao programa computacional GraphPad Prisma 4.0 software.

44
4 RESULTADOS
4.1 Ximenia americana (Ameixeira)
4.1.1 Teste de toxicidade aguda
Os testes mostraram que a maior mortalidade ocorreu nas doses mais altas. A
mortalidade acumulativa foi de 33,3% para a dose de 10.000mg/kg (2° dia após a
administração da dose única), 66% para 5.000mg/kg (3° dia da administração das doses
seguidas, etapa que só ocorre se o animal não tiver morrido na dose única) e, de 33.3% para a
dose de 2.500mg/kg ( 23° dia). A DL50 foi de 5000mg/kg, e além das mortes ocorridas outros
aspectos foram vistos entre os animais que permaneceram vivos, como, por exemplo,
agitação, taquicardia, aumento da freqüência respiratória, piloereção e perda de peso entre os
animais que receberam as maiores doses. Alguns desses aspectos, como agitação, taquicardia
e aumento da frequência respiratória foram normalizados após cerca de três dias da
administração da dose.
4.1.2 Testes antimaláricos in vivo
Os resultados revelaram que o extrato etanólico de X. americana não reduziu a
parasitemia nas três doses testadas: 250, 500 e 1000mg/Kg. Sendo, esta planta, inativa contra
a malária (Tabela 2 e Fig. 13).
A média de sobrevivência dos animais para as doses de 250, 500 e 1000mg/Kg foi
de 32,5; 26,7 e 26 dias respectivamente (Tabela 2). Os animais tratados com a dose de
250mg/kg apresentaram maior sobrevida que aqueles tratados com as demais doses; o grupo
controle não tratado apresentou sobrevida média de 28 dias, contudo, esta diferença não teve
significado estatístico.

45
Tabela 2- Atividade antimalárica in vivo da Ximenia americana (ameixeira)
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida média
dos animais foi > 60 dias.
Figura 13- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Ximenia americana (ameixeira).
5 7 5 7 5 7 0
10
20
30
40
50
500mg/kg1000mg/kg
Tempo em dias
250mg/kg
Red
ução
de
para
site
mia
(%
)
Espécie/Parte planta Doses
(mg/Kg)
% Inibição Parasitemia
( Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Ximenia americana
250
3,81 (0,018)
10,5 (0,05)
32,5 ± 6,6
500
8 (0,017)
0 (0,05)
26,7 ± 2,5
1000 17,1 (0,003)
11,4 (0,05)
26 ± 4,0
0* 0 (0,005)
0 (0,04)
28 ± 2,7
Dias após infecção

46
4.1.3 Citotoxicidade
Foi observado um padrão de resposta dose-dependente (quanto maior foi a
concentração testada, menor foi a viabilidade das células usadas) nas linhagens tumorais 3T3,
HeLa e HEPG2 no período de 24 horas, e nas linhagens tumorais 3T3, CHO, HeLa e HEPG2
durante o período de 48 horas.
Dentre os padrões de viabilidade observados, a linhagem CHO apresentou
crescimento ≥ 88% para todas as concentrações testadas após 24h de incubação. Na menor
concentração testada (0,1mg/ml), para as três linhagens celulares (3T3, CHO, HeLa), a
viabilidade celular foi superior ao do grupo controle (Fig. 14), o que já era esperado; contudo
sem significado estatístico. Foi possível observar que em todas as linhagens celulares,
incluindo ambos os períodos analisados, apresentaram viabilidade ≥50% na menor
concentração (0,1mg/ml), exceto para HEPG2 no tempo de 48h (Fig. 15).
A linhagem HEPG2 foi a que apresentou os menores valores de viabilidade dentre
as demais para os dois períodos testados (Fig. 14 e 15).
No período de 24h os valores de IC50 determinados foram: 0,654; 2,950;
0,500mg/ml para as células 3T3, CHO, e HeLa. Já para o período de 48h foram obtidos
0,1410; 0,566; 0,554mg/ml. O valor de IC50 não foi possível ser calculado para HEPG2.
Figura 14- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de Ximenia americana (ameixeira).
0
25
50
75
100
125
150
1753T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

47
Figura 15- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de Ximenia americana (ameixeira).
4.2 Maytenus rigida (Bom nome)
4.2.1 Teste antimalárico in vivo
O extrato etanólico de Maytenus rigida promoveu redução da parasitemia nas três
doses testadas (250, 500 e 1000mg/kg), com uma atividade antimalárica parcial observada
apenas no 5º dia. A supressão da parasitemia foi de 30, 32,4 e 45%, nas respectivas doses
testadas (Tabela 3 e Figura 16). No sétimo dia não foi verificada atividade esquizonticida para
o extrato. No acompanhamento da sobrevida, os animais tratados com a maior dose
começaram a morrer no mesmo dia que àqueles que receberam apenas o solvente com o
agente solubilizador do extrato, neste caso o DMSO 3%. O tempo de sobrevivência para as
doses de 250, 500 e 1000mg/kg foi: 25,3; 24 e 28,3 dias. O grupo controle apresentou média
de sobrevivência de 25,3 dias (Tabela 3).
0
25
50
75
100
125
1503T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%)

48
Tabela 3- Atividade antimalárica in vivo do extrato de Maytenus rigida (Bom nome).
Espécie/Parte planta Doses
(mg/Kg)
% Inibição Parasitemia
(Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Maytenus rigida
250
30 (0,006)
12,3 (0,052)
25,3 ± 3,8
500
32,4 (0,014)
31,5 (0,05)
24 ± 3,6
1000 45 (0,005)
23 (0,05)
28,3 ± 4,2
0* 0 (0,01)
0 (0,06)
25,3 ± 3,8
*Grupo controle negativo que recebeu água plus DMSO (concentração final de 3%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98%; a sobrevida média
dos animais foi > 60 dias.
Figura 16- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Maytenus rigida (bom nome).
5 7 5 7 5 7
0
20
40
60
80250 mg/kg
500 mg/kg
1000 mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%)
Dias após infecção

49
4.2.2 Teste de citotoxicidade
O extrato etanólico de Maytenus rigida apresentou baixa citotoxicidade. Quanto a
viabilidade das células frente ao extrato, um padrão de resposta dose-dependente foi
observado nas linhagens 3T3, HeLa e HEPG2. Não foi visto curva dose-resposta utilizando a
linhagem CHO, esta se manteve com valores de viabilidade superiores a 100%, apresentando
um crescimento maior do que o observado no grupo controle. Os valores achados para a
concentração inibitória de 50% das células no período de 24h (Fig. 17) para as células 3T3,
CHO, HeLa e HEPG2 foram de 1.178, 17.252, 0.933 e 1.327 mg/ml; respectivamente. Após
48h de incubação (Fig.18) a concentração inibitória de 50% do crescimento celular foi de
0.935, 5.003, 0.555 e 0.931mg/ml para estas mesmas células.
Figura 17- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de Maytenus rigida (bom-nome).
0
25
50
75
100
125
150
3T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

50
Figura 18- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de Maytenus rigida (bom-nome).
4.3 Sideroxylon obtusifolium (Quixabeira)
4.3.1 Teste antimalárico in vivo
O extrato de Sideroxylon obtusifolium apresentou atividade antimalárica parcial,
com redução de 56,2% no quinto dia para a dose de 500mg/Kg e de 33% na dose de
250mg/kg no sétimo dia (Tabela 4 e Figura 19).
Com relação ao atraso na mortalidade dos animais tratados, foi observado
similaridade com o grupo controle não tratado. As médias de sobrevida foram de 33.5 dias
para o grupo controle, 36.5 e 35 dias para as doses de 250 e 500mg/kg, respectivamente
(Tabela 4).
0
25
50
75
100
125
150
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
3T3
Tipos celulares
CHO HeLa HEPG2
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

51
Tabela 4- Teste antimalárico in vivo de Sideroxylon obtusifolium (Quixabeira)
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%)
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida
média dos animais foi > 60 dias..
Figura 19- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Sideroxylon obtusifolium (Quixabeira).
5 7 5 70
20
40
60
80250mg/kg
500mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%
)
Dias após infecção
Espécie/Parte
planta
Doses
(mg/Kg)
% Inibição de Parasitemia
(Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Sideroxylon
obtusifolium
250
2 (0,017)
33 (0,038)
36,5 ± 5,0
500
56,2 (0,0009)
-
35 ± 0
0* 0
(0,012)*
0 (0,06)
33,5 ± 9,2
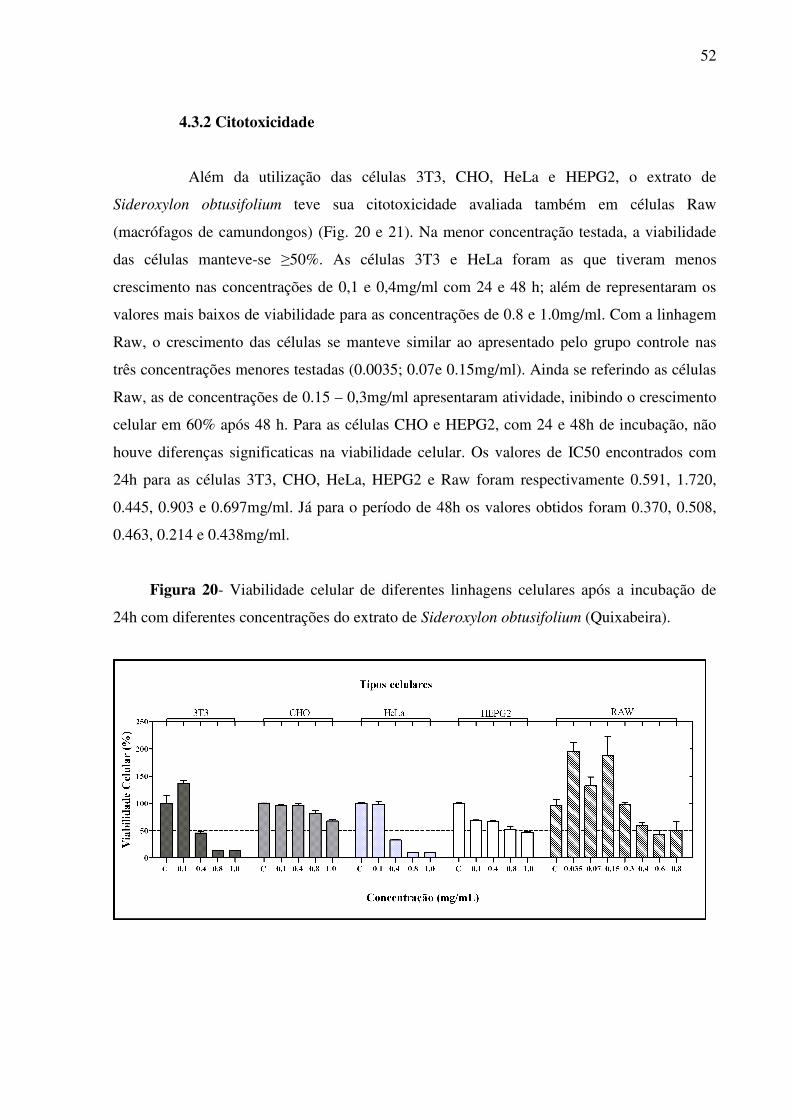
52
4.3.2 Citotoxicidade
Além da utilização das células 3T3, CHO, HeLa e HEPG2, o extrato de
Sideroxylon obtusifolium teve sua citotoxicidade avaliada também em células Raw
(macrófagos de camundongos) (Fig. 20 e 21). Na menor concentração testada, a viabilidade
das células manteve-se ≥50%. As células 3T3 e HeLa foram as que tiveram menos
crescimento nas concentrações de 0,1 e 0,4mg/ml com 24 e 48 h; além de representaram os
valores mais baixos de viabilidade para as concentrações de 0.8 e 1.0mg/ml. Com a linhagem
Raw, o crescimento das células se manteve similar ao apresentado pelo grupo controle nas
três concentrações menores testadas (0.0035; 0.07e 0.15mg/ml). Ainda se referindo as células
Raw, as de concentrações de 0.15 – 0,3mg/ml apresentaram atividade, inibindo o crescimento
celular em 60% após 48 h. Para as células CHO e HEPG2, com 24 e 48h de incubação, não
houve diferenças significaticas na viabilidade celular. Os valores de IC50 encontrados com
24h para as células 3T3, CHO, HeLa, HEPG2 e Raw foram respectivamente 0.591, 1.720,
0.445, 0.903 e 0.697mg/ml. Já para o período de 48h os valores obtidos foram 0.370, 0.508,
0.463, 0.214 e 0.438mg/ml.
Figura 20- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de Sideroxylon obtusifolium (Quixabeira).

53
Figura 21- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de Sideroxylon obtusifolium (Quixabeira).
4.4 Stryphnodendron coriaceum (Barbatimão)
4.4.1 Teste antimalárico in vivo
O extrato de Stryphndendron coriaceum, na dose de 1000mg/kg, reduziu em 60%
a parasitemia apenas no sétimo dia. Para as doses de 250 e 500mg/kg, o extrato foi
considerado inativo apresentando uma atividade “boderline” (Tabela 5 e figura 22).
Os animais do grupo tratado com as concentrações de 250, 500 e 1000mg/Kg
apresentaram média de sobrevivência de 26, 33.5 e 24 dias respectivamente. Apesar da
redução observada na dose de 1000mg/kg, a média de sobrevida dos animais desse grupo foi a
menor dentre as doses testadas.

54
Tabela 5- Atividade antimalárica in vivo do extrato de Stryphnodendron coriaceum
(Barbatimão).
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida média
dos animais foi > 60 dias.
Figura 22- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Stryphnodendron coriaceum (Barbatimão).
Espécie/Parte planta Doses
(mg/Kg)
% Inibição de Parasitemia
(Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Stryphnodendron
coriaceum
250
9,6 (0,002)
23 (0,0325)
26 ± 2,83
500
11,13 (0,014)
28 (0,05)
33,5 ± 6,4
1000 24 (0,026)
60 (0,025)
24 ± 0
0* 0 (0,01)
0 (0,045)
27,5 ± 0,8
5 7 5 7 5 70
20
40
60
80250mg/kg
500mg/kg
1000mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%
)
Dias após infecção

55
4.4.2 Citotoxicidade
Com 24h, o extrato etanólico de Stryphnodendron coriaceum apresentou uma
atividade dose-dependente para a linhagem CHO. Nas células 3T3, HeLa e HEPG2 o efeito
citotóxico do extrato foi similar, com maior crescimento celular da HEPG2 em relação as
anteriores. A linhagem Raw se mostrou menos susceptível ao extrato que as demais (Fig.23).
Quanto a concentração de extrato inibitória para metade das células foram
encontrados os valores de 0.380mg/ml e de 0,420mg/ml para as células CHO e Raw,
respectivamente. Os valores de IC50 para as demais células não foram determinados dado a
maior susceptibilidade frente ao extrato.
Após a incubação por 48h, as curvas de viabilidade celular foram bastante
similares (Fig.24). Não foi possível obtenção dos valores de IC50 para as concentrações
testadas.
Figura 23- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de Stryphnodendron coriaceum (Barbatimão).

56
Figura 24- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de Stryphnodendron coriaceum (Barbatimão).
4.5 Bowdichia virgilioides (Sucupira)
4.5.1 Teste antimalárico in vivo
Apenas a dose de 100 mg/kg foi ativa, reduzindo a parasitemia em 65 e 35% nos
dias 5 e 7 após infecção, respectivamente. As demais doses se mostraram inativas. A dose de
500 mg/Kg exibiu atividade apenas no 5º dia (Tabela 6 e Figura 25).
O tempo de sobrevivência dos grupos tratados nas doses de 100, 250 e 500mg/Kg
foi de 23, 40 e 32 dias, respectivamente. O grupo controle apresentou média de sobrevivência
de 30.5 dias (Tabela 6).

57
Tabela 6- Atividade antimalárica in vivo do extrato de Bowdichia virgilioides (Sucupira).
Espécie/Parte planta Doses
(mg/Kg)
% Inibição de Parasitemia
(Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Bowdichia
virgilioides
100
65 (0,021)
35 (0,06)
23 ± 0
250
0 (0,003)
0 (0,05)
40 ± 0
500 38 (0,012)
18 (0,05)
32 ± 10
0* 0 (0,014)
0 (0,06)
30,5 ± 13
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida média
dos animais foi > 60 dias..
Figura 25- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Bowdichia virgilioides (Sucupira).
5 7 5 7 5 70
20
40
60
80100mg/kg
250mg/kg
500mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%)
Dias após infecção

58
4.5.2 Citotoxicidade
As linhagens celulares, incubadas por 24 horas com o extrato da planta,
mostraram crescimento em até ≥ 100% nas concentrações testadas. Para as células 3T3, CHO,
HeLa, HEPG2 os valores determinados de IC50 foram 1.619, 1.453, 1.747 e 1.705mg/ml .
Não foi possível determinar o IC50 para a linhagem celular Raw.
Em relação ao tempo de incubação de 48 h, para as células HeLa e HEPG2,
quanto maior foi a concentração da droga testada, menor foi a percentagem de células viáveis.
Um padrão semelhante também pôde ser visto nas demais linhagens. Isso pôde ser observado
nas concentrações de 0.8mg/ml para 3T3 e CHO, e na de 0.3mg/ml para as células Raw (Fig.
27). Os valores de IC50 determinados foram 1.444, 1.929, 0.718, 0.934 e 1.080mg/ml,
respectivamente, para 3T3, CHO, HeLa, HEPG2 e Raw.
Figura 26- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de Bowdichia virgilioides (Sucupira).
0
50
100
150
200
2503T3
Tipos celulares
CHO HeLa HEPG2 RAW
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.035 0.07 0.15 0.3 0.4 0.6 0.8
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

59
Figura 27- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de Bowdichia virgilioides (Sucupira).
4.6 Schinopis brasiliensis (Baraúna)
4.6.1 Teste antimalárico in vivo
A espécie Schinopis brasiliensis mostrou atividade parcial apenas no 5° dia após
infecção nas doses de 250 e 500mg/kg, reduzindo a parasitemia em 86% e 95%,
respectivamente. Já no 7º dia essas doses foram inativas. A dose de 1000mg/kg monstrou-se
inativa nos dois dias avaliados (Tabela 7 e Fig. 28). Os grupos tratados apresentaram uma
sobrevida média entre 32-34 dias em relação ao grupo controle não-tratado.
0
25
50
75
100
125
150
175
200
225
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.035 0.07 0.15 0.3 0.4 0.6 0.8
3T3
Tipos celulares
CHO HeLa HEPG2 RAW
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

60
Tabela 7- Atividade antimalárica in vivo do extrato de Schinopsis brasiliensis
(Baraúna).
Espécie/Parte
planta
Doses
(mg/Kg)
% Inibição Parasitemia
( Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Schinopsis
brasiliensis 250
86 (0,001)
6,5 (0,0324)
32 ± 0
500 95,3
(0,00033)
11,5 (0,03)
34 ± 0
1000 0
(0,008)
0 (0,05)
32 ± 0
0* 0 (0,007)
0 (0,032)
24 ± 0
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida média
dos animais foi > 60 dias.
Figura 28- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato etanólico de Schinopsis brasiliensis (Baraúna).
5 7 5 7 5 7
0
50
100
150
250mg/kg
500mg/kg
1000mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%
)
Dias após infecção

61
4.6.2 Citotoxicidade
Dado a problemas técnicos no cultivo das linhagens celulares, o extrato de
Schinopsis brasiliensis teve a citotoxicidade testada apenas com a célula HeLa; sendo testado
com 24h, 48h e 72h de incubação.
A figura 35 apresenta os resultados com os 3 períodos de incubação. A maior
quantidade de células permaneceu viável por 24h, com crescente diminuição da viabilidade
após 48h e 72h. A concentração inibitória de 50% do crescimento celular foi determinada para
cada tempo de exposição do extrato, os valores encontrados foram de 0.584mg/ml,
0.018mg/ml com 24h e 48h, respectivamente (Fig. 29). Não foi possível obter o IC50 para
72h de incubação.
Figura 29- Viabilidade celular da célula HeLa após a incubação por diferentes
tempos e concentrações com o extrato de Schinopsis brasiliensis (Baraúna).
Controle 0,07 0,1 0,3 0,4 0,6 0,70
50
100
150 24H
48H
72H
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)

62
4.7 Picrolemma sprucei (Caferana)
4.7.1 Teste antimalárico in vivo
Foram testados os extratos aquosos do caule (C) e da raiz (R) da caferana nas
doses de 50, 100 e 200mg/Kg. No quinto dia após a infecção, os extratos inibiram a
parasitemia entre 35,4 e 79,3% (Figura 31 e 32).
A sobrevivência dos animais tratados com os extratos foram similares. (Tabela 8 e
9).
Tabela 8- Atividade antimalárica in vivo do extrato de caule de Picrolemma sprucei
(caferana).
Espécie/Parte
planta
Doses
(mg/Kg)
% Inibição Parasitemia
(Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Picrolemma
sprucei (Caule) 50
50,2 (0,008)
39,6 (0,054)
26,75 ± 11
100 35,4
(0,02)
20,7 (0,066)
25 ± 3,6
200 79,3
(0,006)
32,3 (0,044)
26,5 ± 5
0* 0 (0,022)
0 (0,068)
21,8 ± 3,6
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98%; a sobrevida média
dos animais foi > 60 dias.

63
Tabela 9- Atividade antimalárica in vivo do extrato de raiz de Picrolemma sprucei (caferana)
Espécie/Parte
planta
Doses
(mg/Kg)
% Inibição Parasitemia
( Parasitemia) Sobrevivência
Média ± D.P. 5° dia 7° dia
Picrolemma
sprucei (Raiz) 50
61 (0,004)
19,25 (0,043)
22 ± 1,4
100 60,5
(0,004)
42,4 (0,038)
24,75 ± 3,8
200
NT
NT
NT
Controle negativo 0 (0,022)
0 (0,07)
21,8 ± 3,6
*Grupo controle negativo que recebeu água plus tween 20 (concentração final de 2%).
D.P. = Desvio padrão; NT = Não testado.
A cloroquina (antimalárico de referência) testada em paralelo apresentou redução de 97-98 %; a sobrevida média
dos animais foi > 60 dias.
Figura 30- Redução da parasitemia do Plasmodium berghei em camundongos tratados
com o extrato aquoso do caule de Picrolemma sprucei.
5 7 5 7 5 7
0
20
40
60
80
100C 50mg/kg
C 100mg/kg
C 200mg/kg
Tempo em dias
Red
ução
de
para
site
mia
(%
)
Dias após infecção

64
Figura 31- Redução da parasitemia do Plasmodium berghei em camundongos tratados com o extrato aquoso da raiz de Picrolemma sprucei.
4.7.2 Citotoxicidade
Praticamente todas as concentrações testadas inibiram o crescimento das
linhagens celulares (Fig. 33). O único valor de IC50 que pôde ser determinado foi o relativo à
célula 3T3 e extrato do caule, valor igual a 0.097mg/ml.
Após 48 horas, o extrato do caule, inibiu o crescimento celular para todas as
células, destacando-se as células 3T3 e HeLa como as menos viáveis (Fig. 34). Em
contrapartida, comparando com o período de incubação de 24 horas, a linhagem CHO
apresentou uma variação de crescimento para duas das concentrações testadas 0,4 e 1,0mg/ml.
Para os resultados envolvendo o extrato de raiz, com as células para 3T3, CHO,
HEPG2, foi verificado o mesmo efeito tóxico (Fig. 35 e 36). O único valor de IC50 que pôde
ser determinado foi o relativo à célula HeLa (raiz), valor igual a 2.661mg/ml.
5 7 5 7
0
20
40
60
80
100R50
R100
Tempo em dias
Red
ução
de
para
site
mia
(%)
Dias após infecção

65
Figura 32- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de aquoso do caule Picrolemma sprucei
(Caferana).
Figura 33- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de aquoso do caule Picrolemma sprucei
(Caferana).
0
25
50
75
100
1253T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)
0
25
50
75
100
1253T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)
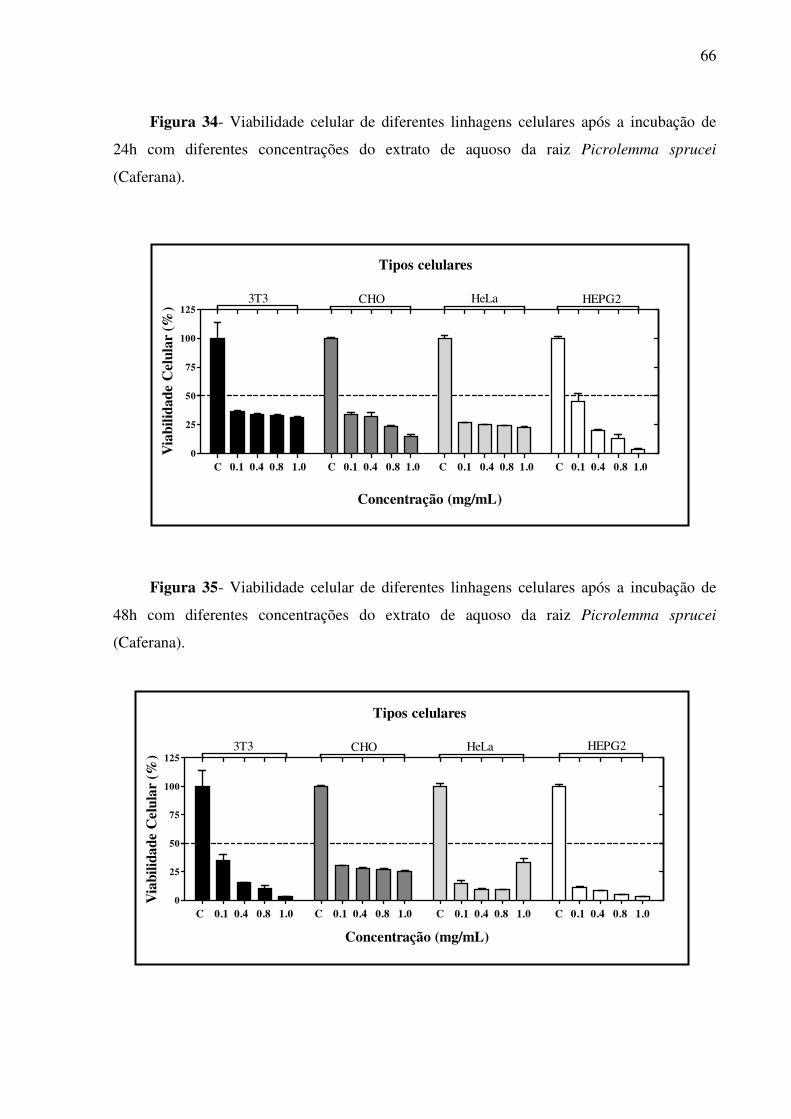
66
Figura 34- Viabilidade celular de diferentes linhagens celulares após a incubação de
24h com diferentes concentrações do extrato de aquoso da raiz Picrolemma sprucei
(Caferana).
Figura 35- Viabilidade celular de diferentes linhagens celulares após a incubação de
48h com diferentes concentrações do extrato de aquoso da raiz Picrolemma sprucei
(Caferana).
0
25
50
75
100
1253T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Concentração (mg/mL)
Via
bilid
ade
Cel
ular
(%
)
0
25
50
75
100
125
Concentração (mg/mL)
3T3
Tipos celulares
CHO HeLa HEPG2
C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0 C 0.1 0.4 0.8 1.0
Via
bilid
ade
Cel
ular
(%
)

67
4.8 AVALIAÇÃO DA ATIVIDADE ANTIMALÁRICA IN VITRO CONTRA O Plasmodium
falciparum
Testadas em cultura de Plasmodium falciparum (cepa K1), as frações clorofórmicas
obtidas das espécies S. obtusifolium, S. brasiliensis e M. rigida, assim como as frações
hexânicas obtidas da S. brasiliensis e M. rigida foram consideradas parcialmente ativas. Os
extratos etanólicos de B. virgiliodes e X. americana foram inativos e a atividade parcial de S.
coriaceum foi observada apenas com o extrato etanólico (Fig. 37). Como critério de atividade
in vitro foi estabelecido que as amostras que inibirem o crescimento dos parasitos de 80-100%
são consideradas ativas (A), 50 a 79% são considerados parcialmente ativas (PA) e <50% são
consideradas inativas (I).

68
Figura 37- Inibição do crescimento in vitro da cepa K1 de Plasmodium falciparum.
* Como critério de atividade in vitro foi estabelecido que as amostras que inibirem o crescimento dos parasitos de: 80-100%= ativas (A),
50 a 79%=parcialmente ativas (PA) e <50%= inativas (I).

69
Tabela 10- Quadro sinóptico dos resultados relativo à citotoxicidade e das análises feitas in vivo e in vitro das espécies vegetais estudadas. Espécie/Nome popular Atividade antimalárica in vivo Atividade antimalárica in vitro Citotoxicidade
Maytenus rigida (Bom nome)
250mg/kg – 5ºdia (30%) 500 mg/kg– 5º (32%) e 7º(31%)dias 1000mg/k -5º dia(45%)
Clorofórmica e hexânica Baixa
Sideroxylon obtusifolium (Quixabeira)
250mg/kg – 7º dia (33%) 500mg/kg – 5ºdia (56%)
Clorofórmica Média
Schinopsis brasiliensis (Baraúna)
250mg/kg – 5ºdia (86%) 500mg/kg – 5ºdia (95%)
Clorofórmica e hexânica Média
Stryphnodendron coriaceum (Barbatimão)
1000mg/kg – 7ºdia (45%) e Bordeline p 500mg/kg (28%)
EtOH Alta
Picrolemma sprucei (Caferana -C)
50mg/kg – 5º (50%)e 7ºdias (40%) 100mg/kh – 5ºdia (35%) 200mg/kg – 5º (79%) e 7ºdias (32%)
- Alta
Picrolemma sprucei (Caferana -R)
50mg/kg – 5ºdia (61%) 100mg/kg – 5º (60%)e 7ºdias (42%)
- Alta
Ximenia americana (Ameixa)
- - Média
Bowdichia virgilioides (Sucupira)
100mg/kg – 5º (65%)e 7ºdias (35%) 500mg/kg – 5ºdia (38%)
- Baixa

70
5 DISCUSSÃO
Em regiões com condições ambientais, econômicas e sociais favoráveis à
manutenção da transmissibilidade da malária, o controle da doença tem sido bastante difícil,
repercutindo em elevadas taxas de morbi-mortalidade principalmente causados pelo
Plasmodium falciparum. A existência de cepas de P. falciparum multi-resistente aos
antimaláricas usuais tem justificado a incessante busca por novas drogas antimaláricas que
tenham baixo custo, que sejam eficazes e com reduzida toxicidade (RIDDER et al., 2008). E é
nesse ponto que nosso trabalho se torna importante, uma vez que a busca a partir de uma
triagem baseada não só na etnofarmacologia, mas também no conhecimento de propriedades
biológicas de plantas medicinais usadas na medicina popular, podem representar uma
estratégia mais rápida ao se comparar a estratégia de descoberta por screening, ou seja, busca
em qualquer espécie vegetal.
A abordagem etnofarmacológica, isto é, a triagem racional baseada na seleção de
plantas usadas na medicina popular tem se mostrado promissora. Um bom exemplo disto é a
experiência chinesa na descoberta do potente antimalárico artemisinina (ginghaosu) como
parte do programa de pesquisas de novas drogas a partir de plantas medicinais
(PHILLIPSON, 1986 apud ROSENBERG et al., 1990). A artemisinina, um sesquiterpeno que
contém endoperóxidos em sua estrutura, foi uma das drogas obtidas apartir dessa abordagem
(PETERS, 1985; KLAYMANN, 1979 apud VENKATRAMAN; SITOLE; FRONCZEK,
2008).
O uso de plantas tem um papel de crucial importância para a medicina tradicional
devido a suas propriedades terapêuticas (MARTINS et al., 2003), sendo o uso de extratos
vegetais no tratamento de muitas doenças, um hábito bastante difundido no Brasil. Este hábito
pode ser explicado, pelo menos em parte, pelo pressuposto de que plantas exibem efeitos
terapêuticos sem causar efeitos tóxicos ou adveros no organismo. Por isso, a pesquisa de
plantas medicinais tem grande importância científica, haja vista que ela busca validar a
segurança de seu uso na medicina tradicional (BRANDÃO et al., 2006; CARVALHO et al.,
2008; VEIGA-JUNIOR et al., 2008).
A abordagem etnofarmacológica, portanto, pode contribuir para desmistificar o
uso de uma determinada espécie vegetal para uma dada finalidade, como visto no trabalho de
Andrade-Neto et al., 2003 e também no presente trabalho, no qual X. americana outrora usada
entre os povos da África para o tratamento da malária (OGUNLEYE; IBITOYE, 2003) foi

71
considerada sem atividade antimalárica in vivo contra P. berghei e in vitro contra P.
falciparum.
Os derivados da artemisinina em combinação com outras drogas constituem hoje
a principal estratégia preconizada pela Organização Mundial de Saúde (OMS) para o controle
global da malária (RIDDER et al., 2008; NGUYEN-POUPLIN et al., 2007). Segundo dados
da OMS, mais de 1 bilhão de pessoas em todo o mundo, cerca de 20%, vivem em condições
de extrema pobreza. Grande parte morre devido, principalmente, as condições precárias do
sistema público de saúde (WHO, 1993; 1997). Em 1996, a expectativa de vida média em
países como Malawi e Uganda foi de 47 anos contrastando com a média de 77 anos para a
Inglaterra e Estados Unidos (WHO, 1993; 1997). Neste contexto, grande parte da população
dos países em desenvolvimento, inclusive no Brasil, utilizam a medicina popular e fazem uso
de plantas medicinais como primeiro tratamento para doenças. Essas plantas são comumente
comercializadas em feiras livres, mercados populares e encontradas em quintais residenciais
(MACIEL et al., 2002), sendo amplamente utilizadas como remédios caseiros, na forma de
infusões e decocções, em decorrência do alto custo do medicamento industrializado
(BRANDÃO et al., 2008). Nos últimos anos as plantas medicinais vêm ocupando um papel
fundamental na medicina convencional e o uso de fitoterápicos vem aumentando tanto no
Brasil quanto em outros países (DUARTE et al., 2005). Contudo, diversos produtos
comercializados em mercados ou feiras são misturas de multicomponentes e sua eficácia pode
estar associada a mais de um composto tornando o seu controle da qualidade uma tarefa
bastante difícil. Apesar de grandes investimentos em pesquisa terem sido feitos para a
comprovação da eficácia e segurança dos medicamentos derivados de plantas medicinais,
poucos são os trabalhos científicos que dizem respeito à manutenção da qualidade desses
produtos. Extratos secos de plantas medicinais possuem efeito farmacológico devido à
presença de substâncias ativas que estão sujeitas a reações de degradação, de modo que se
torna importante o estudo da velocidade dessas reações, que por sua vez, pode ser previstas
através de estudo de estabilidade que envolve o armazenamento dos extratos em diversas
condições (diferentes temperatura e umidade relativa do ar) (BOTT, 2008). Tais condições,
bem como associação com outros fatores, podem influenciar o efeito farmacológico do
composto, e desta forma, comprometer a sua eficácia. Neste trabalho, foram observadas
variações entre os valores de redução de parasitemia para uma determinada espécie, o que
claro pode ser devido a biodegradação dos compostos presentes no extrato bruto ou pela
própria variação genética existente nos animais utilizados nesta pesquisa.

72
Algumas espécies utilizadas neste estudo, apesar de não ter indicação direta de seu
uso contra a malária, foram selecionadas por apresentarem indicações para sintomas também
existentes na malária, como ,por exemplo, antiinflamatórias, antitérmicas e antioxidante. A
mioria delas foram testadas pela primeira vez contra a malária e, algumas, se mostraram
ativas sendo de fundamental importância que elas passem por etapas de estudos adicionais
para que princípios ativos sejam isolados, ou até validadas em seu uso fitoterápico.
A medicina tradicionalmantém e difunde a prática do consumo de fitoterápicos,
tornando válidas informações terapêuticas acumuladas durante séculos. De maneira indireta,
este tipo de cultura medicinal desperta o interesse de pesquisadores em estudos envolvendo
áreas multidiciplinares, como botânica, farmacologia, fitoquímica, parasitologia e imunologia
que juntas enriquecem os conhecimentos sobre o potencial farmacológico da flora mundial
(MACIEL et al., 2002). Dessa forma, a medicina tradicicional não deve ser vista apenas como
uma alternativa mais rápida para a descoberta de fármacos ou fitoterápicos, mas estudada em
termos clínicos, principalmente, quando não resultam em avaliações in vitro e/ou in vitro
satisfatórias, pois apesar de não ser consideradas como promissoras em tais testes, podem ser
efetoras na estimulação do sistema imunológico e na atenuação de sintomas típicos da doença.
Outro ponto que merece ser ressaltado é o fato de não descartarmos uma dada espécie só
porque não foi considerada promissora nos testes in vitro ou in vivo, nesse sentido torna-se
necessária associações com outras espécies, de forma a verificar alguma ação sinérgica ou
aditiva, potencializando sua atividade biológica (BOTSARIS, 2007).
Phillipson et al. (1995) já postulavam esta hipótese, argumentando que uma
possível explicação para tal fato é que muitos dos componentes ativos de plantas funcionam
como antitérmicos ou estimulantes do sistema imunológico de forma que auxiliam no alívio
dos sintomas da doença, e não propriamente de uma atividade direcionada contra o
Plasmodium. Alternativamente, os precursores dos constituintes ativos podem estar presentes
nos extratos, mas que precisam passar por metabolização e modificação, normalmente in vivo,
antes da exibição de sua atividade (GESSLER et al., 1995). Compostos como esses são
chamados de pró-drogas e essa necessidade de metabolização poderiam explicar o efeito mais
proeminente in vivo do que in vitro, como observado nesse estudo; efeito mais tardio foi visto
para S. coreaceum, apresentando-se como espécie de atividade apenas no 7º dia, fato que para
outras plantas poderia apresentar uma menor atividade, sendo tal fato associado,
provavelmente, com o tempo de meia-vida do princípio ativo. A maioria das espécies
testadas neste trabalho apresentou maior atividade no 5º dia. Deste modo, antes de
invalidarmos o efeito antimalárico para uma determinada espécie vegetal, ensaios adicionais

73
devem ser realizados nas espécies consideradas inativas, principalmente, do ponto de vista
químico, farmacológico e biológico.
Como avaliado por Gathirwa et al. (2008), os extratos contêm diversos
constituintes que podem interferir antagonicamente na atividade dos metabólitos principais e,
deste modo, resultar em inatividade dos extratos para o Plasmodium. Além disso, fatores
como quimiotipos, parâmetros ambientais, colheita e condições de estocagem podem
influenciar coletivamente os metabólitos secundários de plantas antes e após a colheita,
refletindo em sua bioatividade.
O tempo de meia-vida do extrato (e como consequência, a biodisponibilidade), ou
seja, o tempo em que os compostos permanecem circulantes no plasma pode diminuir e afetar
seu efeito biológico. O que pode explicar alguns resultados, onde observamos variações entre
o efeito antimalárico no quinto e sétimo dia após infecção. Neste trabalho, nossos extratos
foram administrados aos animais pela rota oral, considerada como de primeira escolha devido
ser esta a mais preconizada pela facilidade de adesão ao tratamento, principalmente onde a
malária é endêmica, como também para a profilaxia ou para o tratamento curativo de viajantes
doentes (VIAL et al., 2004).
Foram utilizados neste estudo animais “outbred”, de constituição genética variada,
estratégia imprescindível quando se almeja a produção de um protótipo de novas drogas, e
que, portanto, seja um bom modelo para refletir a heterogeneidade existente na população
humana.
5.1 TOXICIDADE AGUDA
O ensaio de toxicidade aguda do extrato alcoólico de Ximenia americana sugere
toxicidade apenas nas doses mais elevadas, demonstrando uma moderada toxicidade segundo
Homburguer (1989). Por outro lado, Maikai et. al. (2008) estudando extratos de diferentes
partes de X. americana, não revelaram mortes até a dose de 5000mg/kg, no entanto, padrões
como agitação, inquietação, falta de apetite e, reduzida atividade durante as primeiras 24
horas de extrato administração também foram observados. No mesmo estudo, também foi
feita uma análise fitoquímica do extrato aquoso de cascas do caule desta planta, notificando a
presença de vários compostos, tais como glicosídeos, filobataninos, taninos, terpenóides, além
de menores quantidades de alcalóides, antraquinonas, flavonóides e saponinas (KUMAR-
ROINÉ; SINGH,1984). Estes autores também sugerem que tais compostos podem ter uma

74
outra ação, a de interferir com a absorção de nutrientes, tornando-os indisponíveis ao
metabolismo e, assim, acarretar perda de apetite nos animais, principalmente nas maiores
doses administradas. Isso, possivelmente, seja a causa da perda de peso observado nos
animais da nossa pesquisa.
5.2 ATIVIDADE ANTIMALÁRICA IN VIVO
A espécie, Ximenia americana, é usada para vários fins, dentre eles: dores de
cabeça, úlcera hansênica, infecções cutâneas, inclusive para o tratamento da malária entre os
povos da África (OGUNLEYE; IBITOYE, 2003). Estudos fitoquímicos em relação à X.
americana revelaram a presença de saponinas, carboidratos, glicosídeos, flavonóides, taninas,
alcalóides, antraquinonas e terpenos (OGUNLEYE; IBITOYE, 2003; JAMES et al, 2008;
MAIKAI et al., 2008). Carboidratos, glicosídeos cianogênicos, flavonóides e taninas têm
atividade bacteriana, o que valida o uso dessa planta no controle de Staphylococcus aureus,
Klebsiella pneumoniae e Shigella flexineri e, portanto, implica em uma propriedade de
tratamento para uma intoxicação alimentar, infecções de pele, úlceras, eczemas, diarréia e
pneumonia (JAMES et al., 2008); além de ação antimicrobiana.
Koné et al. (2004) investigaram a atividade antibacteriana do extrato bruto
etanólico de 50 plantas medicinais e verificaram que X. americana apresentou maior
atividade. Estudos farmacológicos realizados in vivo conduzidos por Soro et al. (2009)
avaliaram a atividade analgésica do extrato aquoso da casca do caule de Ximenia americana
L. (Olacaceae). Como resultado, foi verificado o efeito inibitório pronunciado (dose de
100mg/kg) em resposta às contorsões (cólicas abdominais) induzida por ácido acético em
camundongos e sobre a fase tardia do teste de formalina. Contudo, foi observada uma fraca
resposta pelo método de tail-flick (resposta de retirada de cauda do animal sobre o calor
depois de aplicada uma droga teste). Os autores apontam que a possível atividade analgésica
esteja relacionada com os flavonóides presentes no extrato, no entanto, eles sugerem mais
estudos a fim de identificar os compostos responsáveis por àquela, bem como compreender os
mecanismos de ação.
Os taninos e antraquinonas, presente nos extratos da X. americana podem tanto
ocasionar danos oxidativos, como atuarem como antioxidantes dentro do organismo

75
(KUMAR-ROINÉ; SINGH,1984) e deste modo interferir na resposta imune celular contra a
malária, baseado no óxido nítrico. Além disso, muitos dos componentes fitoquímicos
presentes no extrato podem atuar como modulador do sistema imunológico, sendo um
adjuvante potencializando a resposta imune contra outros patógenos, que não o Plasmodium
(RUKUNGA et al., 2009).
No que se refere a Maytenus rigida, os diversos trabalhos na literatura mostram
que as plantas desse gênero são ricas em metabólitos bioativos da classe dos terpenos ou
isoprenóides (SHIROTA et al., 1994; NIERO et al., 2006; REYES et al., 2006) e que
algumas de suas atividades biológicas são, entre outras, anti-inflamatória (SANOGO et al.,
2006; REYES et al., 2006; SOSA et al., 2007), antimalárica (MUTHAURA et al., 2007),
antioxidante (BUFFA et al., 2004), antinoceptiva (NIERO et al., 2006; SANOGO et al.,
2006) e antitumoral (SHIROTA et al., 1994; BUFFA et al., 2004).
Diversas espécies de Maytenus têm sua atividade biológica comprovada
experimentalmente, demonstrando atividade antinociceptiva em animais (DIAS et al., 2007),
antiinflamatória (JORGE et al., 2004), antioxidante (VELLOSO et al., 2006; MELO et al.,
2001) e antiulcerogênica (JORGE et al., 2004). Atividade antimicrobiana (KLOUCEK et al.,
2007) e antileishmanicida (PEREZ-VICTORIA et al., 1999) foram demonstradas com a
espécie M. macrocarpa. Outra espécie com atividade neste gênero é a M. krukovii, esta possui
atividade antimutagênica, antioxidante e antimicrobiana (BRUNI et al., 2006). Também têm
sido relatada propriedades analgésicas da espécie M. aquifolium (GONZÁLES et al., 2001)
bem como atividade antiulcerogênica de M. robusta (ANDRADE et al., 2007). A espécie M.
senegalensis demonstrou atividade antimalárica (GESSLER et al., 1995; EL TAHIR et al.,
1999), antibacteriana (MATU & VAN STADEN, 2003), antiviral (OTAKE et al., 1995) e
antitumoral (GESSLER et al., 1995).
Ensaios realizados por Muthaura et al. (2007) em modelo murino utilizando
extratos metanólicos e aquosos de Maytenus undata e Maytenus putterlickioides
demonstraram atividade parcial contra Plasmodium berghei. Reider e Roland (1984) in
Muthaura et al. (2007) atribuem as diversas atividades biológicas aos diversos metabólitos
secundários presentes neste gênero. Em outro estudo, realizado com M. ilicifolia, Buffa Filho
et al. (2002) detectaram como grupo mais diverso de metabólitos secundários os
triterpenóides quinonametídicos. Um triterpeno com esqueleto lupano e alcalóides, chamado
de rigidenol, foi observado em análises químicas com o extrato de Maytenus rigida (MARTA
et al., 1979 apud DIAS et al., 2007), considerado repelentes de insetos (DELLE et al., 1984
apud DIAS et al., 2007).

76
Nos últimos anos, diversas pesquisas in vivo demonstraram que o extrato etanólico
de Maytenus rigida Mart. apresentou efeito antiinflamatório, anti-ulcerogênico e anti-
diarréico (SANTOS et al., 2007).
No presente trabalho, os resultados obtidos com o extrato etanólico de M. rigida
reduziu a parasitemia de maneira similar ao estudo realizado por Dias et al. (2007) que
também demonstrou a atividade antimalárica in vivo desta espécie, encontrando uma redução
da parasitemia contra P. berghei na dose de 500mg/Kg.
Em se tratando da espécie Sideroxylon obtusifolium (Sapotaceae), pouco se sabe sobre
sua ação biológica e seus constituintes fitoquímicos. Contudo, alguns relatos apresentam uma
associação com atividade antiinflamatória e antioxidante (DESMARCHELIER et al, 1999;
DESMARCHELIER; SCHAUS, 2000; SPENCER et. al.,1988 apud DESMARCHELIER et
al, 1999; SREEJAYAN; RAO, 1996). Possivelmente a presença de flavonóides em sua
constituição poderiam agir contra radicais livres, diminuindo, assim, o estresse oxidativo
característico de doenças com processos inflamatórios (DESMARCHELIER et al., 1999).
Quanto a constituição química, em geral, assim como em X. americana e M. rigida, espécies
da família Sapotaceae apresentam alcalóides (ATA; FEJIR, 1975 apud MONTENEGRO et
al., 2006; YANG, et al., 1999 apud MONTENEGRO et al., 2006), flavonóides (MATHEW;
LAKSHMINAYANA, 1969 apud MONTENEGRO et al., 2006; MARANZ et al., 2003) e
terpenóides (MONTENEGRO, 2005), ocorrendo adicionalmente outros compostos, como os
benzenóides e os fenilpropanóides (DIXIT; SRIVASTAVA, 1990). Segundo Botsaris (2007)
a Pradosia lactescens outra Sapotaceae pode ser associada a qualquer outra droga utilizada
para o tratamento da malária.
Neste estudo investiga-se pela primeira vez o potencial esquizonticida de S.
obtusifolium. Montenegro et al (2006) demonstraram a atividade antimalárica do extrato
etanólico obtido de folhas de Puteria venosa, pertencente a família Sapotaceae. Estes dados
são animadores e corroboram com a atividade antimalárica descrita para a espécie S.
obtusifolium no presente trabalho e nos permitem confirmar mais uma espécie da família
Sapotaceae com atividade biológica importante.
A Stryphnodendron coriaceum (Leguminosae) apresentou atividade antimalárica,
fato possivelmente relacionado aos compostos presentes em seu extrato, tais como saponinas
(TURSCH et al., 1963, 1966 in LOPES et al., 2009), galactomananas (REICHER et al., 1993
in LOPES et al., 2009) e taninos (MELLO, PETEREIT, NAHRSTEDT, 1996a, 1996b, 1999
in ZOCOLER et al., 2009), seja na casca, folhas ou sementes, que podem estar agindo em
associação de forma a possibilitar efeito aditivo ou sinérgico.

77
As plantas que contêm saponinas apresentam sabor amargo, sendo essas
provenientes do metabolismo secundário vegetal, cuja característica química é de atuarem
como surfactante. Plantas com essa composição podem ser tóxicas em concentrações altas,
uma vez que destroem a membrana celular (GONSALVES, 1999). Esse dado é reforçado por
Kingsbury (1964) in Brito et al., (2001), este autor relata que as saponinas contidas em
algumas plantas podem causar desde irritação na parede do trato digestivo, muitas vezes
podendo acarretar grave gastrenterite. Nesse trabalho, as espécies: S. obovatum, S. coriaceum
e S. barbatimão ocasionaram problemas digestivos em animais que tinham essas plantas
como fonte alimentar, levando o autor a expecular a ação adversa associada às saponinas.
Apesar desse fato, não foi verificado, no presente trabalho, um efeito tóxico muito evidente
em camundongos.
Já as plantas que contêm taninos são usadas na medicina tradicional para tratar
doenças como diarréia, hipertensão arterial, reumatismo, hemorragias, feridas e queimaduras,
entre outros (SANTOS & MELLO, 2007). Várias espécies do gênero Stryphnodendron têm
sido descritas, como cicatrizante, antimicrobiano, anti-ulcerosos, hipotensores, anti-parasitária
contra Leishmania amazonensis (JORGE et al., 1996) e Trypanosoma cruzi (HERZOG-
SOARES et al., 2002), atividade antiviral (FELIPE et al., 2006), atividade antifúngica contra
Candida albicans (ISHIDA et al., 2006), atividade antinociceptiva (MELO et al., 2001), bem
como a capacidade antioxidante por meio de radicais livres (SANCHES et al., 2005 in
LOPES et al., 2009).
Diferente dos poucos relatos sobre a atividade biológica da S. coriaceum, as
informações referentes à Bowdichia virgiliodes são mais conhecidas e, portanto, mais
utilizada pela população na medicina tradicional. Pesquisas realizadas no Brasil estimam que
milhares de pessoas a utilizem em forma de decocção contra a malária (BRANDÃO et al.,
1992). Diversas atividades biológicas foram relatadas para esta espécie, tais como:
hipoglicêmicas (BARBOSA-FILHO et al., 2005), antimicrobianas (ALMEIDA et al., 2006) e
inibidoras da enzima acetil-colinesterase (BARBOSA-FILHO et al., 2006). Vários trabalhos
comprovaram o potencial antimicrobiano do óleo essencial de B. virgiliodes (ALMEIDA et
al., 2006) a as suas propriedades antiinflamatórias em modelos murinos (THOMAZZI et al.,
2010). Deharo et al. (2001) comprovaram a atividade antimalárica do extrato etanólico obtido
da casca desta espécie, sendo bastante similares ao obtidos em nosso trabalho.
Muito provavelmente, a atividade biológica vista por nosso grupo e aquelas
relatadas na literatura estejam associadas aos seus constituintes: flavonóides (VELOZO et al.,
1999), antocianinas (MELL, 1929 apud THOMAZZI et al., 2010), benzofuranóides (MELO

78
et al., 2001), óleo essencial (ARRIAGA et al., 1998), triterpenóides (MELO et al., 2001),
resinas (MACHADO, 1936 apud THOMAZZI et al., 2010), e alcalóides (TORRENEGRA et
al., 1989 apud THOMAZZI et al., 2010). Um tipo específico de flavonóide – os
Isoflavonóides glicosidados – foram considerados capazes de atuar em processos anti-
inflamatórios e antissupressores, um exemplo deles é a odoratina 7-O-β-D-glicopiranosídeo
(VELOZO et al, 1999). Os alcalóides, composto que conferem às plantas um gosto também
amargo, geralmente ligado ao sistema de proteção das plantas, dão as plantas um certo grau de
toxicidade. Para a espécie B. virgilioides foram descritos cinco tipos de alcalóides na casca
do caule (TORRENEGRA et al.; 1989 in THOMAZZI et al., 2010). Contudo, não
verificamos mortalidade expressiva nos grupos de camundongos avaliados quanto a
toxicidade aguda. Provavelmente isto deve-se às concentrações dos constituintes secundários
da planta, que podem variar de acordo com o tipo de solo, época do ano, ou outros fatores.
Diferente de B. virgilioides, relatos sobre a atividade biológica de Schinopis
brasiliensis são escassos na literatura, os poucos trabalhos existentes são de Cardoso et al.
(2005) e Lucena et al. (2007), porém não estão ligados a testes in vivo, tampouco in vitro.
Sendo assim, existem apenas alguns trabalhos relativos ao mesmo gênero e outros poucos
relativos à família. Atividades biológicas antibacteriana, antifúngica, leishmanicida e
cicatrizante são relatados para extratos de Schinus terebinthifolium em testes in vitro e in vivo
(LUCENA et al, 2007;. BRAGA et al, 2007). Já Pires et al. (2004) demonstraram sua baixa
toxicidade quando ele foi administrado por via oral em ratos. Diante disso, podemos sugerir
que S. brasiliensis também apresente baixa toxicidade, já que pertencem ao mesmo gênero
Schinopsis. Contudo, isto precisa ser avaliado em estudos complementares. Além disso, os
nossos resultados apresentaram uma quimiossupressão do Plasmodium berghei bastante
significativa, associada a uma taxa de sobrevida maior.
Neste trabalho foi estudada também a atividade biológica de Picrolemma sprucei,
planta conhecida na Amazônia brasileira como caferana e comumente usada no tratamento da
malária pela população. (GRENAND et al. 1987 apud SARAIVA et al., 2003; DUKE;
VASQUEZ; 1994 apud SARAIVA et al., 2003; MILLIKEN et al., 2005 apud POHLIT et al.,
2009). Os trabalhos etnofarmacológicos conduzidos na Guiana Francesa mostram que os
extratos brutos de raiz, caule e cascas usadas para o tratamento da malária, em associação com
Geissospermum spp. (Apocynaceae) e Quassia amara (Simaroubaceae) bem como com
drogas antimaláricas comerciais (VIGNERON et al., 2005) apresentam importante atividade
antimalárica. Diante disso, Bertani et al. (2005) avaliaram alguns desses extratos em testes in
vivo contra o Plasmodium yoelii. Os resultados mostraram que Picrolemma pseudocoffea

79
(sinônimia P. sprucei) e Quassia amara ambas Simaroubaceae foram capazes de inibir em
mais de 50% do crescimento do parasita. Os resultados encontraram atividades inibitórias
significativas de até 79% para os extratos testados, o que comprova sua atividade antimalárica
e valida o seu uso tradicional pelos habitantes da bacia amazônica.
5.3 ATIVIDADE ANTIMALÁRICA IN VITRO
Existem poucos estudos com Plasmodium falciparum acerca das espécies testadas
neste trabalho. Um exemplo desse fato ocorre com Maytenus rigida, Sideroxylon obtusifolium
e Schinopis brasiliensis, para essas plantas não há avaliação da atividade antimalárica in vitro.
Utilizando também um representante da mesma família que Maytenus rigida
(Celastraceae) Pavanand, Webster e Yongvanitchit (1989) avaliaram a atividade
antiplasmódica da pristimerina, triterpeno presente no extrato clorofórmico da planta
Celastrus paniculatus, contra as cepas U-1471, K-3, CH 12 e CH 150 do Plasmodium
falciparum; e não tiveram atividade antimalárica. Entretanto, avaliando outra espécie do
gênero – Maytenus senegalensis, os valores de inibição do crescimento do P. falciparum
foram bastante significativos. É importante acrescentar que essa planta possui indicação
etnofarmacolócica no Kenia (África), sendo usada para problemas de dores nas articulações e
tratamento da malária. Outras espécies de Maytenus também se mostraram ativas contra cepas
sensíveis e resistentes de P. falciparum, é o caso de M. undata e M. putterlickioides
consideradas ativas, cujos extratos metanólico e aquoso (exceto aquoso de M.
putterlickioides) apresentaram IC50 <10 µg/ml (MUTHAURA et al., 2007).
Entretanto, não tão raro, os resultados de atividade antimalárica in vitro e in vivo
podem não exibir similaridade ou co-relação. Ainda, uma dada espécie vegetal pode se
apresentar inativa em testes contra cepas de Plasmodium falciparum que apresentem
diferentes susceptibilidade. Isso foi observado no trabalho de Rukunga et al. (2009); o extrato
metanólico de M. senegalensis não foi ativo contra as cepas NF54 (sensível a cloroquina) e
ENT30 (resistente a cloroquina) de P. falciparum; espécie com atividade relatada em
trabalhos anteriores (GESSLER et al, 1995;. EL TAHIR et al, 1999). Essa diferença de
atividade entre as avaliações feitas por vários autores pode ser devida a fatores relacionados
ao tipo de solo, período do dia em que foi colhida a planta ou até o período do ano da coleta
podem interferir na atividade biológica dos compostos testados (BERO et al., 2009;

80
GATHIRWA et al., 2008; JONVILLE et al., 2008), ou ainda devido ao polimosfismo gênico
do parasito.
Por sua vez, M. rigida, espécies pertencente ao mesmo gênero e avaliada neste
estudo, se mostrou ativa para os extratos/frações testados contra P. falciparum.As diferenças
na atividade antimalárica foram observadas entre as diferentes extrações, sugerindo que os
compostos ativos, possam estar concentrados nas frações clorofórmicas e hexãnicas
(MUTHAURA et al., 2007).
Acreditamos que uma maior possibilidade de atividade contra P. falciparum esteja
mais associada quando as espécies vegetais avaliadas pertençam ao mesmo gênero que
quando pertencentes a nível de família; apesar de não podermos descartar que espécies de
diferentes gêneros pertencentes a mesma família possam apresentar atividade antimalárica.
Um exemplo disso são as espécies Sideroxylon obtusifolium e Omphalocarpum glomerata, de
mesma família (Sapotaceae) e que também apresentaram atividade parcial contra o P.
falciparum in vitro (MESIA et al., 2008). Contudo, o mesmo não pôde ser visto com a
Vitellaria paradoxa, cujos valores de IC50 foram considerados baixos para os extratos
diclorometânico, metanólico e aquoso (JANSEN et al., 2010), e portanto, considerada inativas
em testes in vitro.
Diversos estudos têm avaliado a atividade antimalárica in vitro de espécies da
família Leguminosae contra o Plasmodium falciparum (CONNELLY et al., 1996; GESSLER
et al., 1995; WEENEN et al., 1990). Dentro desta família, estão duas das espécies avaliadas
neste trabalho: a Stryphnodendron coriaceum e a Bowdichia virgiliodes, ambas pertencentes a
família Fabaceae.
Com relação as interpretações de atividade in vitro, adotando-se os valores de
IC50: ≤20µg/ml como ativo e de > 100 µg/ml como inativo. Bero et al. (2009) consideraram
Dialium guineense Willd. (Leguminosae) inativa para quase todos os extratos avaliados,
exceto para o diclorometano, no qual foi visto uma atividade moderada (IC50 = 42.1µg/ml).
Apesar de ter seguido outros padrões de valores para o IC50, desta vez os adotados pela WHO
(muito ativo: IC50 <5µg/ml, promissora: entre 5-15µg/ml, baixa: entre 15-50µg/ml, e inativos
quando IC50 > 50µg/ml apud JONVILLE et al., 2008), Esmaeili et al. (2009) não
encontraram alta atividade em Glycyrrhiza glabra (IC50=17,5 µg/ml), mas a espécie foi
considerada promissora para o desenvolvimento de novas drogas contra a malária, por
apresentar valor de IC50 bem próximo àquela faixa, segundo a WHO. Em nossa avaliação,
não seguimos os citados parâmetros, entretanto, foram adotados parâmetros de IC50 já
utilizados por nosso grupo e e referência em estudos de malária e produtos naturais

81
(ANDRADE-NETO et al., 2003, 2004 e 2007), nos quais espécies cujos valores de supressão
situaram entre 80 a 100% são consideradas ativas, entre 50 a 79% como parcialmente ativas, e
valores < 50% como inativas. Assim, dentre as espécies de Leguminosae aqui avaliadas,
apenas S. coriaceum apresentou atividade antimalárica, resultado visto aqui para o extrato
bruto, similarmente àquele encontrado para os testes in vivo Outro ponto a ser considerado é a
diferença de atividade entre as diferentes extrações para as mesmas espécies testadas.
Verificamos a atividade parcial para o extrato etanólico de S. coriaceum, diferindo de outros
trabalhos, nos quais extratos com mais atividade quase sempre são metanólicos ou
clorofórmicos (JASEN et al., 2010; GANSANÉ et al., 2010; RUKUNGA et al., 2009).
A Bowdichia virgiliodes, outra Leguminosae avaliada em nosso trabalho,
anteriormente testada contra as cepas D2 (resistente a cloroquina) e F32 (sensível a
cloroquina) de P. falciparum, mostrou-se bastante ativa (IC50= 1 µg /ml) para ambas as cepas
(DEHARO et al., 2001); atividade não observada em Cassia abbreviata (CONNELLY et al.
1996) e em Bauhinia rufescens (JANSEN et al., 2010). Estes resultados controversos podem
ser devidos as cepas utilizadas, uma vez que cada linhagem apresenta um grau de resistência
diferente às drogas utilizadas, bem como a fatores relacionados a época de coleta e solo
cultivado no qual a planta foi cultivada.
Dentre as espécies analisadas no presente estudo a espécie Schinopis brasiliensis
foi uma das que apresentaram melhor atividade antimalárica, sendo, portanto suas frações,
hexânica e clorofórmica, consideradas parcialmente ativas na maior concentração testada.
Estudos de Mesquita et al., (2009) já haviam avaliado atividade de S. brasiliensis em
associação com a Schinus terebenthifolius (Anacardiaceae), verificando-se desta vez, melhor
supressão do crescimento do parasito, sugerindo que uma dada espécie pode ser considerada
como parcialmente ativa quando testada isoladamente, mas que em associação pode ter um
efeito melhorado, possivelmente devido a efeitos sinérgicos e/ou aditivos.
De todas as espécies estudadas, a Picrolemma sprucei é a mais largamente
utilizada pela medicina popular e uma das espécies mais bem estudadas por nosso grupo
quanto a fitoquímica. Bertani et al. (2005) utilizando as espécies: Picrolemma pseudocoffea
(sinônimo de P. sprucei) e Quassia amara (Simaroubaceae), verificaram atividade para os
extratos aquosos testados contra a cepa W2 de P.falciparum (cepa resistente a cloroquina).
Wright et al. (2005) observaram que a família Simaroubaceae possui
quassinóides, compostos usados tradicionalmente no tratamento contra malária bem como
para outras doenças causadas por protozoários. Estes compostos quando degradados originam
triterpenos, que por sua vez são considerados metabólitos ativos tanto em testes in vitro como

82
in vivo. Isso pôde ser analisado em trabalhos com Simarrouba amara (PHILLIPSON et al.,
1995), no qual a glaucarubinona foi considerada parcialmente seletiva contra P. falciparum,
sugerindo uma possível correspondência de atividade contra P. falciparum entre espécies
dentro da família Simaroubaceae. Albernaz et al. (2010) verificaram elevada atividade para o
extrato diclorometânico de folhas de Simarouba versicolor contra a cepa multirresistente
FcB1 de P. falciparum. Utilizando a glaucarubinona, uma molécula isolada da casca da raiz,
também foi observada atividade similar (WRIGHT et al., 1993).
Diante disso, apesar de não termos avaliado a atividade antimalárica in vitro de
Picrolemma sprucei neste estudo, trabalhos anteriores do nosso grupo já o tinham realizado
um screenig in vitro. Dados preliminares mostraram que a espécie possui atividade biológica
e, que, portanto, é promissora para o desenvolvimento de protótipos para novos antimaláricos
Andrade-Neto et al. (2007).
5.4 CITOTOXICIDADE
No que se refere à avaliação de toxicidade aguda, a única espécie vegetal que
pôde ser avaliada in vivo foi a Ximenia americana. Os efeitos observados nos animais foram
piloereção e agitação. As demais, dado a quantidade disponível, só foram avaliados quanto a
toxicidade in vitro. Deste modo, partimos do referencial disponível no laboratório para testes
antimaláricos in vivo usando extrato bruto, sendo a dose mais alta a ser testada de 100mg/Kg
de peso. pelo fato de termos quantidade insuficientes de extrato para os testes nas doses
estabelecidas.
A Ximenia americana foi avaliada in vitro usando as várias linhagens: 3T3, CHO,
HeLa e HEPG2, contudo, foi com a célula HeLa que valores de mais baixa viabilidade pôde
ser observada. Voss et al. (2006) relatam que essa espécie é utilizada na medicina tradicional
africana para tratar alguns tipos de cancêres . Testes realizados com a riproximina, um de seus
componentes isolados, apresentou atividade anti-tumoral importante. Portanto, com base
nisso e em nossos resultados usando as células HeLa, verificamos seu efeito citotóxico nesta
linhagem celular. Apesar de não apresentar atividade antimalárica, a X. Americana, poderá ser
pensanda para estudos futuros com relação a sua potencial atividade anti-câncer.
Foi observada uma baixa citotoxicidade para o extrato alcoólico de Maytenus
rigida nas diversas linhagens celulares utilizadas. Entretanto, em estudos recentes, usando
células KS, foi verificada a citotoxicidade de triterpenos isolados da casca do caule desta

83
planta (MARTUCCIELO et al., 2010). Adicionalmente, Muthaura et al. (2007), relataram
diferentes níveis de citotoxicidade para extratos de M. undata e M. putterlickioides em células
Vero E6. Em M. undata, assim como em M. putterlickioides, o extrato metanólico foi mais
citotóxico que o aquoso.
Quanto a espécie Sideroxylon obtusifolium, não há relatos de sua citotoxicidade
na literatura. Nossos resultados apontam para uma toxicidade baixa a moderada. Um ponto de
destaque nessa avaliação foi observado para as células Raw. Nas maiores concentrações
testadas, foi verificada uma baixa citotoxicidade, inclusive com 48h de incubação.
Nas demais linhagens celulares a citotoxicidade foi dose e tempo-dependente, o
que já é esprado. Em relatos da literatura, a espécie Omphalocarpum glomerata pertencente a
mesma família da S. obtusifolium não foi citotóxica em linhagens de fibroblastos de pulmão
(MRC), mesmo na maior concentração testada (MESIA et al., 2008); fato que, mesmo sendo
espécie diferente daquela avaliada em nosso estudo, nos sugere que espécies pertencentes a
essa família podem apresentar moderada citotoxicidade.
Stryphnodendron coriaceum foi a única espécie que apresentou efeito citotóxico
em todas as concentrações testadas após 48h de incubação. Como não há estudos de
citotoxicidade relativa a essa espécie, sugerimos que esse fato seja ocasionados, talvez, por
metabólitos secundários presentes no extrato, como as saponinas, agentes que danificam a
membrana celular como descrito por Gonsalves (1999), bem como outros compostos ainda
não analisados nesta espécie. O que necessitam de estudos fitoquímicos futuros para um
melhor entendimento de seu real efeito citotóxico.
Por outro lado, a Bowdichia virgilioides, já estudada por Deharo et al. (2001),
apresentou tanto neste trabalho resultados similares. Estudos feitos por Benoit et al. (1995)
compararam a baixa citotoxicidade de B. virgilioides àquela encontrada em Artemisia annua.
Por outro lado, O’Neill et al. (1986) afirmam que B. virgilioides apresenta uma menor
citotoxicidade dentre as muitas espécies da família Simaroubaceae. Diante de sua atividade
antimalárica e de sua baixa toxicidade descrita neste trabalho, consideramos B. virgiloides
como uma espécie promissora para estudos posteriores quanto ao seu potencial como fonte de
compostos visando uma molécula líder na busca de novas drogas contra a malária.
Existem poucos relatos na literatura acerca da atividade biológica da Schinopis
brasiliensis (Anacardiaceae). A S. brasiliensis foi a única espécie estudada que foi avaliada
apenas com um tipo celular – HeLa; exibindo um moderada citotoxicidade, uma vez que a
viabilidade celular foi maior que 50% para todas concentrações testadas com 24h de
incubação; sendo ainda mais tóxica nos outros tempos de incubação. Outras espécies de

84
Anacardiaceae, como a Schinus terebinthifolius, cujas flores, caules e folhas são usados na
medicina tradicional para o tratamento de tumores e hanseníase (SCHMOURLO et al., 2005),
contudo, não se conhece seu efeito citotóxico. Por outro lado, a Anacardium occidentale,
amplamente encontrada na Caatinga nordestina (KUBO et al., 2010), é uma espécie produtora
de ácidos antioxidantes, e que já foi considerada inibidora de células cancerígenas. Assim,
pode-se sugerir que a atividade biológica S. brasiliensis seja similar àquelas espécies citadas.
A P. sprucei, conhecida pela população ribeirinha da região norte, que a compra
muitas vezes para o preparo de infusões concentradas, com finalidade de provocar abortos
(POHLIT, A. M., informações pessoais), foi a que apresentou a maior toxicidade in vitro.
Certamente, a citotoxicidade dos extratos de P. sprucei esteja relacionada com a presença dos
quassinóides (ANDRADE-NETO et al., 2007). Não podemos afirmar se estivessem sujeitas a
um metabolismo hepático, por exemplo, se seriam transformadas em metabólicos menos
tóxicos., bem como saber o mecanismo farmacocinético envolvido. Com relação à parte da
planta utilizada, a raiz foi a que apresentou maior citotoxicidade, apesar de ter inibido mais
ativamente a parasitemia in vivo. Maior citotoxicidade para a raiz já era esperada, pois
resultados in vivo também demonstraram toxicidade mais pronunciada para a raiz, observada
no tempo de sobrevivência dos animais. Estudos conduzidos por Mesquita et al., (2009)
consideraram esta espécie bastante tóxica contra várias linhagens celulares, apresentando
citotoxicidade ≥85% para menor concentração testada. Isso pode estar relacionado a uma
possível presença de glaucarubinona, presente em cascas de raízes de Simarouba versicolor,
neste caso, possivelmente a P. sprucei também possua, uma vez que são espécies bem
próximas. Além disso, não podemos descartar a possibilidade de uma ação em associação
com os quassinóides, que são altamente tóxicos.

85
6. CONCLUSÃO
Os nossos resultados apontam positivamente para utilização do uso de plantas
medicinais na medicina tradicional e na busca de novos medicamentos contra a malária. No
entanto, são necessários estudos conjuntos com botânicos, fitoquímicos, farmacologistas e
parasitologistas para elucidar os componentes presentes nas espécies promissoras, bem como
os prováveis mecanismos de ação dos constituintes de um extrato bruto. Salientamos a
importância de que mais pesquisas sejam conduzidas tendo em vista o importante problema
de saúde pública associado à malária e as limitações dos antimaláricos utilizados atualmente.
A abordagem etnofarmacológica é uma abordagem racional que proporciona ganho
substancial de tempo em relação ao “screening” aleatório, otimizando a busca de espécies
vegetais que possuam atividade biológica. Além disso, esta abordagem proporcionou
informações relevantes sobre a atividade biológica de espécies de dois biomas distintos –
principalmente da Caatinga - por ser bioma pouco explorado e com evidentes potencialidades.
O conhecimento da atividade biológica e sua validação científica podem contribuir para a
preservação de espécies e de sua disseminação como plantas medicinais, uma vez passam a
ser protegidas, preservadas para uma possível exploração sustentável. Por fim, este trabalho
proporcionou informações importantes sobre o efeito antimalárico de algumas espécies,
abrindo perspectivas para a continuação de pesquisas no campo da biodiversidade e
bioprospecção de espécies vegetais na busca de plantas promissoras como fonte para o
desenvolvimento de novos antimaláricos, fato de importante relevância uma vez que a
resistência do Plasmodium falciparum torna o arsenal profilático e terapêutico contra a
malária bastante restrito.

86
REFERÊNCIAS
AKOMPONG, T. et al. Gametocytocidal Activity and Synergistic Interactions of Riboflavin with Standard Antimalarial Drugs against Growth of Plasmodium falciparum in vitro. Antimicrobial Agents and Chemotherapy, [S.l.], v. 44, n. 11, p. 3107–3111, nov. 2000. ALBERNAZ,L. C. et al. Investigation of plant extracts in traditional medicine of the Brazilian Cerrado against protozoans and yeasts. Journal of Ethnopharmacology, [S.l.], v. 13, p. 116–121, 2010. ALMEIDA, J. R. G. S. et al. Antimicrobial activity of the essential oil of Bowdichia
virgilioides Kunt.Brazilian. Journal Pharmacognosy, [S.l.], v. 16, p. 638–641, 2006. AMINO, R. et al. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nature Medicine, [S.l.], v. 12, p. 220-224, jan. 2006. ANDRADE, C. F. et al. Thymic alterations in plasmodium berghei-infected mice. Cellular immunology, [S.l.], v. 253, p. 1-4, 2008. ANDRADE-LIMA, D. Plantas das caatingas. Rio de Janeiro: Academia Brasileira de Ciências, 1989. 243 p. Apud: OLIVEIRA, M.C.P.; OLIVEIRA, G. J. Superação da dormência de sementes de Schinopis brasiliensis. Ciência Rural, Santa Maria, v. 38, n. 1, p. 251-254, 2008. ANDRADE-NETO, V. F et al. Antimalarial activity of Cinchona like plants used to treat fever and malaria in Brazil. Journal Ethnopharmacology, [S.l.], v. 87, p.253-256, 2003. ANDRADE-NETO, V. F et al. Antimalarial activity of phenazines from lapachol, β-lapachone and its derivatives against Plasmodium falciparum in vitro and Plasmodium berghei in vivo. Bioorganic & Medicinal Chemistry Letters, [S.l.], v. 14, p.1145-1149, 2004. ANDRADE-NETO, V. F. et al. In vitro inhibition of Plasmodium falciparum by substances isolated from Amazonian antimalarial plants. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, v. 102, n. 3, p. 359-65, jun. 2007.
ARBONNIER, M. Trees, Shrubs and lianas of West African dryzones. [S.l.]: Margraf Publishers. 2004. 574 p.

87
ARRIAGA, A. M. C. et al. Volatile constituents from roots of Bowdichia virgilioides Kunt. Journal of Essential Oil Research, [S.l.], v. 10, p. 205–206, 1998. ATA, J.; FEJIR, D. Allantoin in shea kernel. Ghana Journal of Agricultural Science, [S.l.], v.8, p. 149, 1975. Apud: MONTENEGRO, L. H. M. et al. Terpenóides e avaliação do potencial antimalárico, larvicida, anti-radicalar e anticolinesterásico de Pouteria venosa
(Sapotaceae). Revista Brasileira de Farmacognosia, v. 16, p. 611-617, 2006. AVILA, S. L. M.; FERREIRA, A. W. An appraisal of laboratory methods addressing roll back malaria. Ciências e Cultura, [S.l.], v. 52, n. 4-5, p. 220-229, 2000. BAIRD, J. K. Host age as a determinant of naturally acquired immunity to Plasmodium falciparum. Parasitology Today, [S.l.], v. 11, n. 3, p. 105–111, 1995. BANCHEREAU, J.; STEINMAN, R. M. Dendritic cells and the control of immunity. Nature, New York, v. 392, p. 245-252, mar. 1998. BARATA, R. C. B. Malária no Brasil: panorama epidemiológico na última década. Caderno de Saúde Pública, Rio de Janeiro, v. 11, p. 128-136, 1995. BARBOSA-FILHO, J. M. et al. Natural Products inhibitors of the enzyme acetylcholinesterase. Revista Brasileira de Farmacognosia, [S.l.], v.16, p. 258–285, 2006. BARBOSA-FILHO, J. M. et al. Plants and their active constituents from South, Central, and North America with Hypoglycemic activity. Revista Brasileira de Farmacognosia, [S.l.], v. 15, p. 392–413, 2005. BARROSO, G. M. Sistemática de Angiospermas do Brasil. Viçosa: Imprensa Universitária, 1991. BECKER, K.; KONCAREVIC, S.; HUNT, N. H. Oxidative stress and antioxidant defense in malarial parasites. In: SHERMAN, I. Molecular Approaches to Malaria. Herndon, VA: American Society of Microbiology Press, 2005. p. 365-383. BECKER, K. et al. Oxidative stress in malaria parasite-infected erythrocytes: host-parasite interactions. International Journal for Parasitology, Australia, v. 34, n.2, p. 163-189, feb. 2004. Apud: BUCHHOLZ, K et al. Depletion of Plasmodium berghei Plasmoredoxin

88
Reveals a Non-Essential Role for Life Cycle Progression of the Malaria Parasite. Plos One, v. 3, n. 6, 2008. BELTRÃO, A. E. S. et al. In vitro biomass production of Sideroxylon obtusifolium (Roem & Schult). Brazilian Journal of Pharmacognosy, [S.l.], v. 18, p. 696-698, dez. 2008. Suplemento. BENOIT, F. et al. Antimalarial activity in vitro of Cochlospermum tinctorium tuberde extracts. Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 89, p. 217–218, 1995. BERO, J. et al. In vitro antiplasmodial activity of plants used in Benin in traditional medicine to treat malaria. Journal of Ethnopharmacology, [S.l.], v. 122, p. 439–444, 2009. BERTANI, S. et al. Evaluation of French Guiana traditional antimalarial remedies. Journal of Ethnopharmacology, [S.l.], v. 98, p. 45-54, 2005. BLOLAND, P. B.; ETTLING, M.; MEEK, S. Combination therapy for malaria in Africa: hype or hope? Bulletin of the World Health Organization, [S.l.], v. 78, n. 12, 2000. BOTT, R. F. Influência do processo de obtenção, das condições de armazenamento e das propriedades físico-químicas sobre a estabilidade de extratos secos padronizados de plantas medicinais. 2008. 181p. Tese (Doutorado em Ciências Farmacêuticas)- Faculdade de Ciências Farmacêuticas de Ribeirão Preto, Ribeirão Preto, 2008. BOTSARIS, Alexandros. Plants used traditionally to treat malaria in Brazil: the archives of Flora Medicinal. Journal of Ethnobiology and Ethnomedicine, Rio de Janeiro, v. 3, p. 18, maio. 2007. BOURDY, G.et al. Medicinal plants uses of the Tacana, an Amazonian Bolivian ethnic group. Journal of Ethnopharmacology, [S.l.], v. 70, p. 87–109, 2000. BRAGA, F. G. et al. Antileishmanial and antifungal activity of plants used in traditional medicine in Brazil. Journal of Ethnopharmacology, [S.l.], v. 111, p. 396–402, 2007. BRAGA, R. Plantas do Nordeste, especialmente do Ceará. 3. ed. Fortaleza: Escola Superior de Agricultura de Mossoró, 1976. 540 p. Apud: DANTAS, B. F. et al. Alterações bioquímicas durante a embebição de sementes de baraúna (Schinopsis brasiliensis Engl.). Revista Brasileira de Sementes, [S.l.], v. 30, n. 2, p.214-219, 2008.

89
BRANDÃO, M. G. L.; BOTELHO, M. G. A.; KRETTLI, A. U. Quimioterapia experimental antimalárica com produtos naturais: I. uma abordagem mais racional? Ciências e Cultura, São Paulo, v. 37, n. 7, p. 1152-1163, jul. 1985. BRANDÃO, M. G. L. et al. M. Medicinal plants and other botanical products from the Brazilian Official Pharmacopoeia. Revista Brasileira de Farmacognosia, [S.l.], v.16, p. 408–420, 2006. BRANDÃO, M. G. L. et al. Influence of the preparation method on the low efficacy of multi-herb commercial products: the example of João da Costa e Associações. Brazilian Journal of Pharmacognosy, [S.l.], v.20, n. 2, p. 250-255, 2008. BRANDÃO, M. G. L. et al. Survey of medicinal plants used as antimalarials in the Amazon. Journal of Ethnopharmacology, [S.l.], v. 36, n. 2, p. 175–182, 1992. BRASIL. Ministério da Saúde. Manual de Terapêutica da Malária. Brasília: FUNASA, 2001. Disponível em: <<http://portal.saude.gov.br/portal/arquivos/pdf/ manu_terapeutica_malaria.pdf>>. Acesso em: 10 dez. 2009. BRASIL. Ministério da Saúde. Programa nacional de prevenção e controle da malária.[S.l.], 2003. Disponível em: <<portal.saude.gov.br/portal/arquivos/pdf/ pncm.pdf>>. Acesso em: 10 dez. 2009.
BRASIL. Ministério da Saúde. Situação Epidemiológica da malária no Brasil em 2005. [S.l.], 2005. Disponível em: <<http://bvsms.saude.gov.br/bvs/publicacoes/ folder_malaria_2006_web.pdf>>. Acesso em: 20 fev. 2008. BRASIL. Ministério da Saúde. Situação Epidemiológica da malária no Brasil em 2006. [S.l.], 2007. Disponível em: <<http://portal.saude.gov.br /portal/arquivos/ pdf/folder_malaria_2007_web.pdf>>. Acesso em: 10 dez. 2008. BRASIL. Ministério da Saúde. Situação epidemiológica da malária no Brasil. Brasília: Ministério da Saúde, 2008. Disponível em: <<http://portal.saude.gov.br /portal/arquivos/pdf/folder_malaria_2008_final.pdf>>. Acesso em: 15 dez. 2009. BRITO, M. F. et al. Intoxicação experimental pelas favas de Stryphnodendron obovatum
(Leg. Mimosoideae) em bovinos: caracterização do quadro clínico. Pesquisa em Veterinária. Brasileira, [S.l.], v. 21, n.1, p. 9-17, 2001.

90
BRUMMIT, R. F. Vascular plant families and genera. London: Royal Botanic Gardens Kew, 1992. BRUNI, R. et al. Antimutagenic, antioxidant and antimicrobial properties of Maytenus krukovii bark. Fitoterapia, [S.l.], v. 77, p. 538-545, 2006. BUENO, N. R. et al. Medicinal plants used by Kaiowá and Guaranti indegenous populations in the Caarapó Reserve. Mato Grosso do Sul, Brazil. Acta Botânica Brasiliensis, [S.l.], v. 19, p. 39-44, 2005. BUFFA FILHO, W. et al. In vitro propagation of Maytenus ilicifolia (Celastraceae) as potential source for antitumoral and antioxidant quinomethide triterpenes production. A rapid quantitative method for their analysis by reverse-phase high-performance liquid chromatography. Arkivoc, [S.l.], v. 6, p.137-146, 2004. BUFFA FILHO, W. et al. Quantitative determination of cytotoxic Friedo-noroleanane derivatives from five morphological types of Maytenus ilicifolia (Celestraceae) by reverse-phase high-performance liquid chromatography. Phytochemistry Analysis, [S.l.], v. 13, p.75–78, 2002. CARDOSO, M. G. et al. Metabólitos secundários vegetais: visão geral química e medicinal. Lavras: Editora da UFLA, 2001. 81 p. CARDOSO, M. P. et al. Determinação Estrutural de um depsídeo clorado de Schinopsis
brasiliensis (ANACARDIACEAE). In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE QUÍMICA, 28. 2005, Poços de Caldas. Resumo. Poços de Caldas: Sociedade Brasileira de Química, 2005. CARDOSO, M. P.; DAVID, J. M.; DAVID, J. P. n- Alquil e n-alquenil fenóis identificados de Schinopsis brasiliensis. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE QUÍMICA, 29. 2006. São Paulo. Resumo. São Paulo: Sociedade Brasileira de Química, 2006. CARLINI, E. A. (Coord.). Estudo da ação antiúlcera gástrica de plantas brasileiras: Maytenus ilicifolia (Espinheira-santa) e outras. Brasília, DF: CEME/AFIP, 1988. 87 p. CARVALHO, A. C. B. et al. Situação do registro de medicamentos fitoterápicos no Brasil. Revista Brasileira de Farmacognosia, [S.l.], v. 18, p. 314-319, 2008.

91
CARVALHO, L. H.; KRETTLI, A. U. Antimalarial chemotherapy with natural products and chemically defined modeluces. Mem. Inst. Oswaldo Cruz, [S.l.], v. 86, n. 2, p. 181-184, 1991. CARVALHO, L. H. et al. Plantas brasileiras: alternativas no tratamento da malária. Ciências Hoje, [S.l.], v. 22, p. 62-68, 1997. CLENDEMAN, T. E.; LONG, G.; BAIRD, J. K. QBC1 and Giemsa stained thick blood films: diagnostic performance of laboratory technologists.Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 86, n. 2, p. 183- 184, mar-abr. 1995. CONNELLY, M. P. E. et al. Antimalarial activity in crude extacts of Malawian medicinal plants. Annals of Tropical Medicine and Parasitology, [S.l.], v. 90, p. 597-602, 1996. CONNER, E. M.; GRISHAM, M. B. Nitric oxide: biochemistry, physiology, pathophysiology. Methods: a Companion to Methods in Enzymology, [S.l.], v. 7, p. 3–13, 1995. COOKE, A. H. et al. Use of the Fluorochrome Benzothiocarboxypurine in malaria diagnosis. Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 86, n. 4, p. 378, jul-ago. 1992. CORRÊA, M. P. Dicionário das Plantas Úteis do Brasil e das Exóticas Cultivadas. Rio de Janeiro: IBDF-Ministério da Agricultura. 1984. v. 6, 149 p. CORSINO, J. et al. Bioactive sesquiterpene pyridine alkaloids from Maytenus aquifolium. Phytochemistry, [S.l.], v. 48, n. 1, p. 137-40, 1998b.
COWMAN, A. F.; CRAB, B. S. Invasion of red blood cells by malaria parasites. Cell, [S.l.], v. 124, n. 4, p. 755-766, fev. 2006. CROFT, A. M. et al. Safety evaluation of the drugs available to prevent malaria. Expert Opinion on Drug Safety, England, v. 1, n. 1, p. 19-27, maio 2002. CROTTI, D. La Malaria, ossia la malária: brevi note di una “storia sociale e popolare”. Le Infezioni in Medicina, [S.l.], v.13, n. 4, p. 265-270, 2005. CRUZ, G. L. Livro Verde das Plantas Medicinais e Industriais do Brasil. Belo Horizonte: Helmus, 1965. 779 p. Apud: THOMAZZI, M. et al. Antinociceptive and anti-

92
inflammatory activities of Bowdichia virgilioides (sucupira). Journal of Ethnopharmacology, v. 127, p. 451–456, 2010. DANTAS, B. F. et al. Alterações bioquímicas durante a embebição de sementes de baraúna (Schinopsis brasiliensis Engl.). Revista Brasileira de Sementes, [S.l.], v. 30, n. 2, p. 214-219, 2008. DEHARO, E. et al. A search fo rnatural bioactive compounds in Bolívia through a multidisciplinary approach. Part V. Evaluation of the antimalarial activity of plants used by the Tacana Indians. Journal of Ethnopharmacology, [S.l.], v. 77, p. 91-98, 2001. DELACOLLETT, D.; VANDERSTUYFT, P. Direct Acridine orange staining is not a`miracle' solution to the problems of malaria diagnosis in the field. Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 88, n. 2, p.187-188, mar./abr. 1994. DELLE, M. F.; MARINI-BETTOLO, G. B.; BERNAYS, E. Z. Isolation of insect antifeedant alkaloids from Maytenus rigida. (Celastraceae). Journal of Applied Entomology Zeitschrift Fur Angewandte Entomologie, v. 97, p.406, 1984. Apud: DIAS, K. S. et al. Antinociceptive activity of Maytenus rigida Stem Bark. Fitoterapia, [S.l.], v. 78, p. 460-464, 2007. DESMARCHELIER, C. et al. Antioxidant and free radical scavenging activities in extracts from medicinal trees used in the‘Caatinga’region in northeastern Brazil. Journal of Ethnopharmacology, [S.l.], v. 67, p. 69–77, 1999. DESMARCHELIER, C.; SCHAUS, F. W. Sixty medicinal plants from the Peruvian Amazon. Lima: Privately Publishe, 2000. 270 p. DI STASI, L. C. Aspectos químicos e farmacológicos da espinheira-santa: uma análise da utilidade dos dados. In: REIS, M. S.; SILVA, S. R. (Org.). Conservação e uso sustentável de plantas medicinais e aromáticas: Maytenus spp., espinheira-santa. Brasília: IBAMA, 2004. 204 p. DIAS, K. S. et al. Antinociceptive activity of Maytenus rigida stem bark. Fitoterapia, [S.l.], v. 78, p. 460-464, 2007. DIXIT, B.; SRIVASTAVA, S. Detection of tannins from the bark of Mimusops littoralis. Indian Journal of Natural Products, [S.l.], v. 6, p. 16-17, 1990.

93
DUARTE, M. C. T. et al. Anti-Candida activity of Brazilian medicinal plants. Journal of Ethnopharmacology, [S.l.], v. 97, p. 305-311, 2005. DUKE, J. A.; VASQUEZ, R. Amazonian Ethonobotanical Dictionary. USA: Ed. CRC
Press. 1994. 137 p. Apud: SARAIVA, R. C. G. et al. Anatomia foliar e caulinar de Picrolemma sprucei HOOK (Simaroubaceae). Acta Amazônica, v. 33, n. 2, p. 213-220, 2003. EL TAHIR, A.; SATTI, G. M. H.; KHALID, S.A. Anti-plasmodial activity of selected Sudanese medicinal plants with emphasis on Maytenus senegalensis (Lam.) Excell. Journal of Ethnopharmacology, [S.l.], v. 64, n.3, p. 227–233, 1999. ESMAEILI, S. et al. Screening of antiplasmodial properties among some traditionally used Iranian plants. Journal of Ethnopharmacology, [S.l.], v. 121, p. 400–404, 2009 FANDEUR, T., MORETTI, C.; POLONSKY, J. In vitro and in vivo assessment of the antimalarial activity of sergeolide. Planta Medica, [S.l.], v. 51, p. 20-23, 1985. FARNESWORTH, N. R.; SOEJARTO, D. D. Potential consequences of plant extinction in the United States in the current and future availability of prescription drugs. Society for Economic Botanic, New York, v.39, n.3, p. 231-240, 1991. FELIPE, A. M. M. et al. Antiviral effect of Guazuma ulmifolia and Stryphnodendron adstringens on poliovirus and bovine herpesvirus. Biologial & Pharmaceutical Bulletin,
Japão, v. 29, p.1092-1095, 2006. FRANCISCHETTI, I. M.; SEYDEL, K. B.; MONTEIRO, R. Q. Blood coagulation, inflammation, and malaria. Microcirculation, England, v. 15, n. 2, p.81-107, 2008. FREITAS, L. F. et al. Malária não complicada por Plasmodium vivax e P. falciparum no Brasil: evidências sobre fármacos isolados e associações medicamentosas empregados em esquemas terapêuticos recomendados pelo protocolo terapêutico oficial. Caderno de Saúde Pública, Rio de Janeiro, v. 23, n.10, p. 2285-2294, out. 2007. GANSANÉ, A. et al. Antiplasmodial activity and toxicity of crude extracts from alternatives parts of plants widely used for the treatment of malaria in Burkina Faso: contribution for their preservation. Parasitology Research, [S.l.], v. 106, p. 335–340, 2010:

94
GATHIRWA, J. W. et al. The in vitro anti-plasmodial and in vivo anti-malarial efficacy of combinations of some medicinal plants used traditionally for treatment of malaria by the Meru community in Kenya. Journal of Ethnopharmacology, [S.l.], v. 115, p. 223–231, 2008. GAY, F. et al. Direct Acridine orange Fluorescence examination of blood slides compared to current techniques for malaria diagnosis. Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 90, n. 5, p. 516-518, set./out. 1996. GESSLER, M. C. et al. Traditional healers in Tanzania: the treatment of malaria with plant remedies. Journal of Etnnophannacology, [S.l.], v. 48, p. 131-144, 1995. GESSLER, M. C. et al. Screening Tanzanian medicinal plants for antimalarial activity. Acta Tropica, [S.l.], v. 56, p. 65-77, 1994. GONÇALVES, A. D.; CASTELLO BRANCO JUNIOR, A. Kinetic of malaria caused by Plasmodium berghei in swiss mice. Hematologycal, parasitologycal and histopathogical studies. Unimar Ciências, [S.l.], v. 11, p. 11-18, 2002. GONSALVES, P. E. Medicinas alternativas: os tratamentos não-convencionais. [S.l.], 1989. Disponível em:< http://books.google.com/books?hl=pt-BR&id=WSSbQr8GrvMC&q=saponinas#v=snippet&q=saponinas&f=false>. Acesso em 11 de nov. 2010. GONZÁLEZ, F. G. et al. Antiulcerogenic and analgesic effects of Maytenus aquifolium, Sorocea bomplandii and Zolernia ilicifolia. Journal of Ethnopharmacology, [S.l.], v. 77, p. 41-7, 2001. GRENAND, P.; MORETTI, C. E.; JACQUEMIN, H. Pharmacopées Traditionelles em Guyane. Edições de l’Orstom, Institut Français de recherche scientifique pour Le développment en coopération, Paris, França, p. 397- 399, 1987. Apud: SARAIVA, R. et al. Anatomia foliar e caulinar de Picrolemma sprucei HOOK (Simaroubaceae). Acta Amazônica, v. 33, n. 2, p. 213-220, 2003. GRØNHAUG, E. et al. Ethnopharmacological survey of six medicinal plants from Mali,West-Africa. Journal of Ethnobiology and Ethnomedicine, [S.l.], v. 27, p. 24-26, 2008. GUO, Z. et al. Biologically active quassinoids and their chemistry: potentianl leads for drug design. Current Medical Chemistry, [S.l.], v. 12, p. 173-190, 2005.

95
GUYATT, H. L.; SNOW, R. W. Impact of malaria during pregnancy on low birth weight in sub-Saharan Africa. Clinical Microbiology Reviews, [S.l.], v. 17, n. 4, p. 760–769, out. 2004. HAKIZAMUNGU, E.; WERI, M. L’usage de plantes médicinales dans le traitement du paludisme en médicine traditionnelle Rwandaise. Bulletin de Médecine et Pharmacie, [S.l.], v. 2, 11–17, 1988. HALDAR, K. et al. Malaria: mechanisms of erythrocytic infection and pathological correlates of severe disease. Annual Review Pathology Mechanisms of Disease, [S.l.], v. 2, p.217-249, 2007. HAWKING, F.; WORMS, M. J.; GAMMAGE, K. 24- and 48-hour cycles of malaria parasites in the blood; their purpose, production and control. Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 62, p. 731–760, 1968. HERZOG-SOARES, J. et al, Atividade tripanocida in vivo de Stryphonodendron
adstringens (barbatimão verdadeiro) e Caryocar brasiliensis (pequi). Revista Brasileira de Farmacognosia, São Paulo, v.12, p.1-2, 2002. HOMBURGER, F. In vivo testing in the study of toxicity and safety evaluation. In: MARQUIS, J.K. (Ed.). A Guide to General Toxicology. New York: Karger, 1989. 198 p. INNOCENT, E. et al. African Journal of Traditional Complementary Alternative Medicine, [S.l.], v. 6, n. 2, p. 163 – 167, 2009. ISHIDA, K. et al. Influence of tannins from Stryphnodendron adstringens on growth and virulence factors of Candida albicans. J. Antimicrob. Chemother. Oxford, v. 58, p. 942-949, 2006. JAMES, D. B. et al. Assessment of the hepatic effects, heamatological effect and some phytochemical constituents of Ximenia americana (Leaves, stem and root) extracts. African Journal of Biotechnology, [S.l.], v.7, n.23, p. 4274-4278, 2008. JANSEN, O. et al. Evaluation of 13 selected medicinal plants from Burkina Faso for their antiplasmodial properties. Journal of Ethnopharmacology, [S.l.], v. 130, p. 143–150, 2010.

96
JOLLOW, D. J. et al. Acetaminophen-induced hepatic necrosis. VI. Metabolic disposition of toxic and nontoxic doses of acetaminophen. Pharmacology, [S.l.], v.12, n.4-5, p.251-271, 1974. JOLY, A. B. Botânica: introdução à taxonomia vegetal. São Paulo: Companhia Editora Nacional. 1993. JONVILLE, M. C.; et al. Screening of medicinal plants from Reunion Island for antimalarial and cytotoxic activity. Journal of Ethnopharmacology, [S.l.], v. 120, p. 382–386, 2008. JORGE, A. S. et al. Anti-leishmanial “barbatimão” (Stryphnodendron adstringens
(Martius) Coville, Mimosaceae) extract against promastigotes forms of Leishmania (Leishmania) amazonensis. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, v. 91. p. 1976. nov. 1996. Suplemento. JORGE, M. P. et al. Evaluation of wound healing properties of Arrabidaea chica Verlot extract. Journal of Ethnopharmacology, Campinas, v. 118, n. 3, p. 361-366, 2008. JORGE, R. M. et al. Evaluation of antinociceptive, anti-infl ammatory and antiulcerogenic activities of Maytenus ilicifolia. Journal of Ethnopharmacology, [S.l.], n. 94, p. 93-100, 2004. KENNEDY, M. L. et al. Chemo sensitization of a multidrugresistant Leishmania tropica line by new sesquiterpenes from Maytenus megellanica and Maytenus chubutensis. Journal of Medicinal Chemistry, [S.l.], v. 44, n. 26, p. 4668-76, 2001. KINGSBURY, J. M. Poisonous Plants of the Unites States and Canada. New York: Prentice-Hall, 1964.
KLAYMANN, D. L. et al. 2-Acetylpyridine thiosemicarbazones. 1. Uma nova classe de potenciais agentes antimaláricos. Journal of Medicinal Chemistry, v. 22, n. 7, p. 855-862, 1979. Apud: VENKATRAMAN, R.; SITOLE, L.; FRONCZEK, F. R.” Rhodamine 6G hexachloridostannate(IV) acetonitrile disolvate.” Acta Crystallographica, 2008.
KLOUCEK, P. et al. Antimicrobial activity of some medicinal barks used in Peruvian Amazon. Journal of Ethnopharmacology, [S.l.], v. 111, p. 427–429, 2007.

97
KONÉ, W. M. et al. Tradicional medicine in North Cote-d.Ivoire: screening of 50 medicinal plants for antibacterial activity. Journal of. Ethnopharmacology, [S.l.], v. 93, p. 43-9, 2004. KRETTLI, A. U. et al. The search for new antimalarial drugs from plants used to treat fever and malaria or plants ramdomly selected: a review. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, v. 96, n. 8, p. 1033-1042, nov. 2001. KUBO, I. et al. Multifunctional Cytotoxic Agents from Anacardium occidentale.
Phytotherapy Research, [S.l.], v. 25, n.1, p.38-45, 2011. KUMAR-ROINÉ, S.; SINGH, M. Tannins: their adverse role in ruminant. Nutrition. Journal of Agricultural and Food Chemistry, [S.l.], v. 32, p. 447-453, 1984. KUMAR-ROINÉ, S. et al. Ability of certain plant extracts traditionally used to treat ciguatera fish poisoning to inhibit nitric oxide production in RAW 264.7 macrophages. Journal of Ethnopharmacology, [S.l.], v.123, n. 3, p. 369-377, 25 Jun. 2009. LALLOO, D. G. et al. UK malaria treatment guidelines. The Journal of infection, Liverpool, v. 54, n. 2, p. 111-121, fev. 2007. LE COINTE, P. Árvores e plantas úteis (Amazônia brasileira III). São Paulo: Companhia Editora Nacional, 1947. 92 p. Apud: SARAIVA, R. C. G.; BARRETO, A. S.; SIANI, A. C.; FERREIRA, J. L. P.; POHLIT, A. M. Anatomia foliar e caulinar de Picrolemma sprucei HOOK (Simaroubaceae). Acta Amazônica, v. 33, n. 2, p. 213-220, 2003. LIMA, A. P. et al. Avaliação da atividade cicatrizante do extrato etanólico da casca da Maytenus rigida Mart. (Celastracea). Scientia Plena, [S.l.], v. 6, n. 3, p. 1-7, 2010. LIMA, N. T.; FONSECA, C. M. O.; HOCHMAN, G. A saúde na constituição do Estado Nacional no Brasil: reforma sanitária em perspectiva histórica. In: LIMA, N. T. et al. Saúde e democracia: historia e perspectivas do SUS. Rio de Janeiro: Fiocruz, 2005. p. 27-58. LOIOLA, C. C.; SILVA, C. J. M.; TAUIL, P. L. Controle da malária no Brasil: 1965 a 2001. Revista Panamericana de Salud Pública, Brasília, v. 11, n. 4, p. 235-243, 2002.

98
LONG, T. T. A. et al. Influence of CD4+CD25+ T cells on Plasmodium berghei NK65 infection in BALB/c mice. International Journal for Parasitology, [S.l.], v. 33, p. 175-183, 2003. LOPES, G. C. et al. Determinação quantitativa de taninos em três espécies de Stryphnodendron por cromatografia líquida de alta eficiência. Brazilian Journal of Pharmaceutical Sciences, [S.l.], v. 45, n. 1, p.135-143, 2009. LORKE, D. A new approach to practical acute toxicity testing. Archive of Toxicology, [S.l.], v. 54, n. 4, p. 275-287, 1983. LUCENA, R. F. P. et al. Useful Plants of the Semi-Arid Northeastern Region of Brazil: a Look at their Conservation and Sustainable Use. Environmental Monitoring Assessment, [S.l.], v. 125, p. 281–290, 2007. MACEDO, M.; FERREIRA, A. R.;. Plantas hipoglicemiantes utilizadas por comunidades tradicionais na Bacia do Alto Paraguai e Vale do Guaporé. Mato Grosso-Brasil. Revista Brasileira de Farmacognosia, [S.l.], v. 14, p. 45–47, 2004. Suplemento. MACHADO, A. Oleo (resin) of “sucupira” (Bowdichia virgilioides seeds). Revista da Associação Brasileira de Farmácia, [S.l.], v. 17, p. 117–118, 1936. Apud: THOMAZZI, M. et al. Antinociceptive and anti-inflammatory activities of Bowdichia virgilioides (sucupira). Journal of Ethnopharmacology, v. 127, p. 451–456, 2010. MACIEL, M. A. M.; PINTO, A. C.; VEIGA JUNIOR, V. F. Plantas medicinais: a necessidade de estudos multidisciplinares. Quimica Nova, [S.l.], v. 25, p. 429-438, 2002. MAIA, G. N. Caatinga: árvores e arbustos e suas utilidades. São Paulo: Leitura & Arte, 2004. MAIKAI, V. A.; KOBO, P. I.; ADAUD, A. O. Acute toxicity studies ofaqueous stem bark extract of Ximenia Americana. African Journal of. Biotechnology, [S.l.], v.7, n.10, p.1600-1603, 2008. MAIKAI, B. V. et al. Public health implications of soil contaminated with helminth eggs in the metropolis of Kaduna, Nigeria. Journal of Helminthology, [S.l.], v. 82, p. 113-118, 2008.

99
MARANZ, S.; WIESMAN, Z.; GARTI, N.. Phenolic constituents of Shea (Vitellaria paradoxa) kernels. Journal of Agricultural and Food Chemistry, [S.l.], v. 21, p. 6268-6273, 2003. MARIOT, M. P. Recursos genéticos de espinheira-santa (Maytenus ilicifolia e M.
aquifolium) no Rio Grande do Sul. 2005. 131 f. Tese (Doutorado em Fitomelhoramento)– Curso de Pós-graduação em Agronomia, Universidade Federal de Pelotas, Pelotas, 2005. MARIOT, M. P.; BARBIERI, R. L. Metabólitos secundários e propriedades medicinais da espinheira-santa (Maytenus ilicifolia Mart. ex Reiss. e M. aquifolium Mart.). Revista Brasileira de Plantas Medicinais, Botucatu, v.9, n.3, p.89-99, 2007. MARTA, M. et al. A new triterpene with a lupane skeleton from Maytenus rigida. Gazzetta Chimica Italiana, p. 109:61, 1979. Apud: DIAS, K. S. et al. Antinociceptive activity of Maytenus Rigida Stem Bark. Fitoterapia, v. 78, p. 460-464, 2007. MARTINS, E. R. et al. Plantas Medicinais. 5. ed. Viçosa: UFV, 2003. MARTUCCIELLO, S. et al. Effects of triterpene derivatives from Maytenus rigida on VEGF-induced Kaposi’s sarcoma cell proliferation. Chemico-Biological Interactions, [S.l], v. 183, p. 450–454, 2010. MATHEW, A.; LAKSHMINAYANA, S. Polyphenols of immature sapota fruit. Phytochemistry, [S.l.], v. 8, p. 507-509, 1969. Apud: MONTENEGRO, L. H. M. et al. Terpenóides e avaliação do potencial antimalárico, larvicida, anti-radicalar e anticolinesterásico de Pouteria venosa (Sapotaceae). Revista Brasileira de Farmacognosia, v. 16, p. 611-617, 2006. MATU, E. N.; VAN STADEN, J. Antibacterial and antiinfl ammatory activities of some plants used for medicinal purposes in Kenya. Journal of Ethnopharmacology, [S.l.], v. 87, p. 35-41, 2003. MCGREGOR, I. A. Studies in the acquisition of immunity to Plasmodium falciparum infection in Africa. Transactions of the Royal Society of Tropical Medicine and Hygiene, [S.l.], v. 58, p.80–87, 1964. MELL, C. D. Interesting sources of natural dyestus. Textile Colorist, [S.l.], v. 51, p. 453–455. 1929. Apud: THOMAZZI, M. et al. Antinociceptive and anti-inflammatory activities of Bowdichia virgilioides (sucupira). Journal of Ethnopharmacology, v. 127, p. 451–456, 2010.

100
MELLO, J. C. P. DE; PETEREIT, F.; NAHRSTEDT, A. Flavan-3-ols and prodelphinidins from Stryphnodendorn adstringens. Phytochemistry, [S.l.], v. 41, p. 807-813, 1996a.
MELLO, J. C. P. DE; PETEREIT, F.; NAHRSTEDT, A. Prorobinetinidins from Stryphnodendorn adstringens. Phytochemistry, [S.l.], v.42, p.857-862, 1996b. MELLO, J. C. P. DE; PETEREIT, F.; NAHRSTEDT, A. A dimeric proanthocyanidin from Stryphnodendron adstringens. Phytochemistry, [S.l.], v. 51, p.1105-1107, 1999. MELO, F. N. et al. Bowdenol, a 2,3-dihydrobenzofuran constituent from Bowdichia virgilioides. Natural Product Letters, [S.l.], v. 15, p. 261–266, 2001. MELO, S. F. et al. Effect of the Cymbopogon citrates, Maytenus ilicifolia and Baccharis genistelloides extracts against the stannous chloride oxidative damage in Escherichia coli. Mutation Research, [S.l.], v. 496, p. 33-38, 2001. MESIA, G. K. et al. Antiprotozoal and cytotoxic screening of 45 plant extracts from Democratic Republic of Congo. Journal of Ethnopharmacology, [S.l.], v. 115, p. 409–415, 2008. MESQUITA, M. L. et al. Cytotoxic activity of Brazilian Cerrado plants used in traditional medicine against cancer cell lines. Journal of Ethnopharmacology, [S.l.], v.123, p. 439–445, 2009. MILLIKEN, W. Plants for Malaria. Plants for Fever: medicinal Species in Latin America: a Bibliographic Survey. UK: Kew Publishing, 2005. Apud: POHLIT, A. M. et
al. LC-ESI-MS Determination of Quassinoids Isobrucein B and Neosergeolide in Picrolemma sprucei Stem Infusions. Journal of Brazilian Chemical Society, [S.l.], v. 20, n. 6, p.1065-1070, 2009. MONTENEGRO, L. H. M. Estudo químico e ensaios biológicos preliminares de Pouteria venosa (Mart.) Baehni e revisão dos terpenóides e das atividades biológicas de espécies de Sapotaceae. 2005. 141 f. Dissertação (Mestrado em química)–Universidade Federal de Alagoas, Maceió, 2005. MONTENEGRO, L. H. M. et al. Terpenóides e avaliação do potencial antimalárico, larvicida, anti-radicalar e anticolinesterásico de Pouteria venosa (Sapotaceae). Revista Brasileira de Farmacognosia, [S.l.], v. 16, p. 611-617, 2006.
MORETTI, C. et al. T. Isolation and structure of sergeolide, a potent cytotoxic quassinoid from Picrolemma pseudocoffea. Tetrahedron Lett., [S.l.], v. 23, p. 647-650, 1982. Apud: SILVA, E. C. C. et al. Biological activity of neosergeolide and isobrucein B (and two

101
semi-synthetic derivatives) isolated from the Amazonian medicinal plant Picrolemma
sprucei (Simaroubaceae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro, v. 104, n.1, p. 48-55, fev. 2009. MOTA, K. S. L. et al. Evaluation of the toxicity and antiulcerogenic activity of the ethanol extract of Maytenus obtusifolia Mart. leaves. Revista brasileira de farmacognonia, [S.l.], v. 18, n. 3, p. 441-446, 2008.
MOTA, M. L. Atividade antimalárica de plantas medicinais da biorregião do Araripe- CE em modelo murino – Plasmodium berghei. 2009. 147 f. Dissertação (Mestrado em Ciências Biológicas) - Universidade Federal do Rio Grande do Norte, Natal, 2009. MOODY, A. H.; CHIODINI, P. L. Methods for detection of blood parasites. Clinical & Laboratory Haematology, [S.l.], v. 22, n. 4, p. 189-201, 2000. MUGANGA, R. et al. Antiplasmodial and cytotoxic activities of Rwandan medicinal plants used in the treatment of malaria. Journal of Ethnopharmacology, [S.l.], v. 128, p. 52–57, 2010. MUTHAURA, C. N. et al. Antimalarial Activity of some plants traditionally used in treatment of malaria in Kwale district of Kenya. Journal of Ethnopharmacology, [S.l.], v. 112, p. 545-551, 2007. NGUYEN-POUPLIN, J. et al. Antimalarial and cytotoxic activities of ethnopharmacologically selected medicinal plants from South Vietnam. Journal of Ethnopharmacology, [S.l.], v. 109, p. 417–427, 2007. NIERO, R. et al. A New Triterpene with Antinociceptive Activity from Maytenus robusta. Journal of Asian Natural Products Research, [S.l.], v. 20, p. 1315-1320. 2006. NORONHA, E. et al. Resistência à mefloquina do tipo RIII em crianças com malária causada pó P. falciparum em Manaus, AM, Brasil. Revista da Sociedade Brasileira de Medicina Tropical, [S.l.], v.33, n. 2, p. 201-205, mar./abr. 2000. OCCHIONI, E. M. Considerações taxonômicas no gênero Stryphnodendron Mart. (Leguminosae Mimosoideae) e distribuição geográfica das espécies. Acta Botânica Brasílica, [S.l.], v. 4, n. 2, p.153-158, 1990. Apud:MOTA, K. S. L. et al. Evaluation of the toxicity and antiulcerogenic activity of the ethanol extract of Maytenus obtusifolia Mart. leaves. Revista brasileira de farmacognonia, [S.l.], v. 18, n. 3, p. 441-446, 2008.

102
OGUNLEYE, D. S.; IBITOYE, S. F. Studies of antimicrobial activity and chemical constituents of Ximenia americana. Tropical Journal of Pharmaceutical Researsh, [S.l.], v.2, n. 2, p. 239- 241, 2003. OKOKON, J. E.; NWAFOR, P. A. Antiplasmodial activity of root extract and fractions of Croton zambesicus. Journal of Etnopharmacology, [S.l.], v. 121, p. 74-78, 2009. OLIVEIRA, M. C. P.; OLIVEIRA, G. J. Superação da dormência de sementes de Schinopis brasiliensis. Ciência Rural, Santa Maria, v. 38, n. 1, p. 251-254, 2008. OMER, M. E. F. A.; ELNIMA, E. I. Antimicrobial activity of Ximenia Americana.
Fitoterapia, [S.l.], v. 74, n. 1, p. 122–126, fev. 2003. O'NEILL, M. J. et al. Plants as Sources of Antimalarial Drugs: in vitro antimalarial activities of some quassinoids. Antimicrobial Agents Chemotheraphy, [S.l.], v. 30, p.101-104, jul. 1986. O’NEILL, M. J. et al. Plants as source of antimalarial drugs. Part I, in vitro test method for the evaluation of crude extracts from plants. Planta Medica, [S.l.], v. 61, p. 394–398, 1985. Apud: DEHARO, E. et al. A search for natural bioactive compounds in Bolívia through a multidisciplinary approach. Part V. Evaluation of the antimalarial activity of plants used by the Tacana Indians. Journal of Ethnopharmacology, v. 77, p. 91-98, 2001. ORGANIZAÇÃO PAN-AMERICANA DA SAÚDE. Trajetória de um sanitarista: recompilação da produção intelectual do Dr. Agostinho Cruz Marques. Brasília: OPAS, 1998. OTAKE, T. et al. Screening of Indonesian plant extracts for anti-human immunodefi ciency virus type 1 (HIV-1) activity. Phytotherapy Reearch, [S.l.], v. 9, p. 6-10, 1995. PANIZZA, S. Fitoterapia. In: LORENZI, H. Plantas medicinais no Brasil. Nova Odessa: Instituto Plantarum, 2002. p.183-184. PAVANAND, K.; WEBSTER, H. K.; YONGVANITCHIT, K. Schizonticidal activity of Celastrus paniculatus Wiid. against Plasmodium falciparum in vitro. Phytotherapy research, [S.l.], v. 3, n. 4, p135-139, 1989.

103
PEREZ-VICTORIA, J. M.; et al. New natural sesquiterpenes as modulators of daunomycin resistance in a multidrug-resistant Leishmania tropica line. Journal of Medicinal Chemistry, [S.l.], v.42, p. 4388–4393, 1999. PETERS, W. I. Malaria Rational methods in the search for antimalarial drugs. Transactions of the Royal Society of Tropical Medicine and Hygiene, Netherlands, v. 61, p. 400-410, 1967. PETERS, W. The problem of drug resistence in malaria. Parasitology, [S.l.], v. 90, p. 705-715, abr.1985. PHILLIPS, R. Current status of malaria and potential for control. Clinical Microbiology Reviews, United Kingdom, v. 14, p. 208-226, jan. 2001. PHILLIPSON, J. D. The case for "Plant Medicines". Parasitology Today, [S.l.], v. 2, p. 327-328, 1986. Apud: ROSENBERG, R. et al. An estimation of the number of malaria sporozoites ejected by a feeding mosquito. Royal society of tropical medicine and hygiene, v. 84, p.209–212, 1990. PHILLIPSON, J. D. et al. Phytochemistry of some plants used in traditional medicine for the treatment of protozoal diseases. In: HOSTETTMANN, K. et al. (Eds.). Phytochemistry of Plants Used in Traditional Medicine. [S.l.]: Oxford Science Publications, 1995. p. 95–135. PIRES, O. C. et al. Preliminary comparative analysis of the acute toxicity and median lethal dose (LD50) of the fruit of the Brazilian black pepper (Schinus terebinthifolius
Raddi) and black pepper (Piper nigrum L.). Acta Farmaceutica Bonaerense, [S.l.], v. 23, p. 176–182, 2004. POHLIT, A. M. et al. LC-ESI-MS Determination of Quassinoids Isobrucein B and Neosergeolide in Picrolemma sprucei Stem Infusions. Journal of Brazilian Chemical Society, [S.l.], v. 20, n. 6, p.1065-1070, 2009. POLONSKY, J. Quassinoid bitter principles. Fortschr Chem Org Naturst, [S.l.], v. 30, p. 101-150, 1973. Apud: SILVA, E. C. C. et al. Biological activity of neosergeolide and isobrucein B (and two semi-synthetic derivatives) isolated from the Amazonian medicinal plant Picrolemma sprucei (Simaroubaceae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro, v. 104, n.1, p. 48-55, fev. 2009. POLONSKY, J., BHATNAGAR, S.; MORETTI, C. 15-deacety-lsergeolide, a potent antileukemic quassinoid from Picrolemma pseudocoffea. Journal of Natural Products,

104
[S.l.], v. 47, n. 6, p. 994-996, 1984. Apud: SILVA, E. C. et al. Biological activity of neosergeolide and isobrucein B (and two semi-synthetic derivatives) isolated from the Amazonian medicinal plant Picrolemma sprucei (Simaroubaceae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro, v. 104, n.1, p. 48-55, fev. 2009. PRUDÊNCIO, M.; RODRIGUEZ, A.; MOTA, M. M. The silent path to thousands of merozoites: the Plasmodium liver stage. Nature Reviews Microbiology, [S.l.], v. 4, p. 849-856, nov. 2006. RAMAZANI, A. et al. D. In vitro and in vivo anti-malarial activity of Boerhavia elegans and Solanum surattense. Malaria Journal, [S.l.], v. 9, p. 124, 2010. REICHER, F. et al. Properties of the seed gum of Stryphnodendron barbatiman
(barbatimão). Applied Biochemistry and Biotechnology, Clifton, v.34/35, p.349, 1992. REIDER, P.J.; ROLAND, D. M. The Alkaloids, New York: Academic Press, 1984. v. 23. Apud: MUTHAURA, C.N. et al. Antimalarial activity of some plants traditionally used in treatment of malaria in Kwale district of Kenya. Journal of Ethnopharmacology, [S.l.], v. 112, p. 545–551, 2007. REINACH, Fernando. Henrietta, a primeira mulher imortal. Jornal da Ciência. [S.l.], 2005. Disponível em: <http://www.jornaldaciencia.org.br/Detalhe.jsp? id=25519>. Acesso em: 09 ago. 2009. REY, Luís. Bases da parasitologia médica. Rio de Janeiro: Guanabara Koogan, 2002. REYES, C. P; NUNEZ, M. J,; JIMÉNEZ, I. A. Activity of lupine triterpenoides from Maytenus species as inhibitors of nitric oxide and prostaglandin E2. Bioorganic & Medicine Chemistry Letters, [S.l.], v. 14, p. 1573-1579, 2006. RIDDER, S.; KOOY, F. V.; VERPOORTE, R. Artemisia annua as a self-reliant treatment for malaria in developing countries. Journal of Ethnopharmacology, [S.l.], v. 120, p. 302–314, 2008. RIDLEY, R. Medical need, scientific opportunity and the drive for antimalarial drugs. Nature, [S.l.], v. 415, p. 686-693, 2002.

105
ROBSON, K. J, et al. Thrombospondin-related adhesive protein (TRAP) of Plasmodium falciparum: expression during sporozoite ontogeny and binding to human hepatocytes. The EMBO Journal, [S.l.], v. 14, n. 16, p. 3883-3894, ago. 1995.
RUKUNGA, G. M. et al. Anti-plasmodial activity of the extracts of some Kenyan medicinal plants. Journal of Ethnopharmacology, [S.l.], v. 121, p. 282–285, 2009. SALES, M. F.; MAYO, S. J.; RODAL, M. J. N. Plantas vasculares das florestas serranas de Pernambuco. Recife: APNE-UFRPE. 1998. 130 p. SANCHES, A. C. C. et al. Antioxidant and antifungal activities of extracts and condensed tannins from Stryphnodendron obovatum Benth. Revista Brasileira de Ciências Farmacêuticas, [S.l.], v. 41, p.101-107, 2005. SANOGO, R. et al. Analgesic and anti-inflammatory activities of the aqueous extracts of Maytenus senegalensis, Stereospermum kunthianum and Trichilia emetica used in the treatment of dysmenorrhoea in Mali. Planta Medica, [S.l.], v. 72, p.1059-1069, 2006. SANTOS, S. C.; MELLO, J. C. P. Taninos. In: SIMÕES, C. M. O. et al. Farmacognosia:
da planta ao medicamento. 6. ed. Porto Alegre: UFRGS; Florianópolis: UFSC, 2006. cap. 24, p. 615-656. SANTOS, V. L. et al. Pharmacological studies of ethanolic extracts of Maytenus rigida
Mart (Celastraceae) in animal models. Revista Brasileira de Farmacognosia, [S.l.], v. 17,
p. 336-342, 2007. SARAIVA, R. C. G. Estudo fitoquímico de Picrolemma sprucei Hook. (Simaroubaceae) e dosagem dos princípios antimaláricos nos chás do caule e raiz. 2001. 88p. Dissertação (Mestrado em Química de Produtos Naturais)- Universidade Federal do Amazonas (UFAM), Manaus, 2001. SARAIVA, R. C. G. et al. Anatomia foliar e caulinar de Picrolemma sprucei HOOK (Simaroubaceae). Acta Amazônica, [S.l.], v. 33, n. 2, p. 213-220, 2003. SCHEFFER, M. C. Uso Tradicional e atual de espécies de Maytenus. In: REIS, M. S.; SILVA, S. R. (Org.). Conservação e uso sustentável de plantas medicinais e aromáticas: Maytenus spp. espinheira-santa. Brasília: IBAMA, 2004. p. 53-66.

106
SCHMOURLO, G. et al. Screening of antifungal agents using ethanol precipitation and bioautography of medicinal and food plants. Journal of Ethnopharmacology, [S.l.], v. 96, p. 563–568, 2005. SCHPECTOR, J. Z. et al. A new quassinoid isolated from Picrolemma pseudocoffea. Acta crystallographica C. [S.l.], v. 50, p. 794-797, 1994. SEIXAS, E.; OSTLER, D. Plasmodium chabaudi chabaudi (AS): differential cellular responses to infection in resistant and susceptible mice. Experimental Parasitology, [S.l.], v. 110, n. 4, p. 394–405, ago. 2005. SHAPIRO, T. A.; GOLDBERG, D. E. Chemotherapy of protozoal infections: malaria. In: BRUNTON, L. L.; LAZO, J. S.; PARKER, K. L.(Org). Goodman & Gilman’s the pharmacological basis of therapeutics. New York: McGraw-Hill, 2006. cap. 39, p. 1021-1047. SHIROTA, O. et al. Cytotoxic Aromatic Triterpenes from Maytenus ilicifolia and Maytenus chuchuhuasca. Journal of Natural Products, [S.l.], v. 57, p.1675-81, 1994. SILVA, E. C. C. et al. Biological activity of neosergeolide and isobrucein B (and two semi-synthetic derivatives) isolated from the Amazonian medicinal plant Picrolemma sprucei (Simaroubaceae). Memórias do Instituto Oswaldo Cruz. Rio de Janeiro, v. 104, n.1, p. 48-55, fev. 2009. SIMONSEN, H. T. et al. In vitro screening of Indian medicinal plants for antiplasmodial activity. Journal of Ethnopharmacology, [S.l.], v. 74, p. 195–204, 2001. SMEGO, R. A.; BEG, A. Rapid diagnostic modalities for malaria. Journal Pakistan Medical Association, [S.l.], v. 50, n. 12, p. 398-399, 2001. SORO, T. Y.; TRAORE, F.; SAKANDE, J. Activité analgésique del’extrait aqueux de Ximena americana (Linné)(Olacaceae). Compters Rendus Biologies, [S.l.], v. 332, p. 371–377, 2009. SOSA, S. et al. Anti-inflammatory activity of Maytenus senegalensis root extracts and of maytenoic acid. Phytomedicine, [S.l.], v. 14, p. 109–114, 2007. SPENCER, C. et al. Polyphenol complexation: some thoughts and observations. Phytochemistry, [S.l.], v. 27, p. 2397–2409, 1988. Apud: DESMARCHELIER, C. et al. Antioxidant and free radical scavenging activities in extracts from medicinal trees used in

107
the‘Caatinga’region in northeastern Brazil. Journal of Ethnopharmacology, v. 67,p. 69–77, 1999. SREEJAYAN, N.; RAO, M. Free radical scavenging activity of curcuminoids. Drug Research, [S.l.], v. 46, p. 169–171, 1996. STEVENS, P. F. Angiosperm Phylogeny Website. Version 7. [S.l.], 2001. Disponível em: <http://www.mobot.org/MOBOT/ research/APweb/>. Acesso em maio 2007. SUTHERLAND, B. et al. Clustered DNA Damages Induced by X Rays in Human Cells. Radiation Research, [S.l.], v., 157, n. 6, p. 611-616, 2009. TAYLOR, W. R.; WHITE, N. J. Antimalarial drug toxicity. Drug Safety, [S.l.], v. 27, p. 25-61, 2004. THOMAZZI, M. et al. Antinociceptive and anti-inflammatory activities of Bowdichia
virgilioides (sucupira). Journal of Ethnopharmacology, [S.l.], v. 127, p. 451–456, 2010. TORRENEGRA, R.; BAUEREISS, P.; ACHENBACH, H. Homoormosanine-type alkaloids from Bowdichia virgilioides. Phytochemistry, [S.l.], v. 28, p. 2219–2221, 1989. Apud: THOMAZZI, M. et al. Antinociceptive and anti-inflammatory activities of Bowdichia virgilioides (sucupira). Journal of Ethnopharmacology, v. 127, p. 451–456, 2010. TSUJI, M.; ZAVALA, F. T cells as mediators of protective immunity against liver stages of Plasmodium. Trends in Parasitology. England, v. 19, n. 2, p 88–93, fev. 2003. TURSCH, B. et al. Terpenoids. LIII. Demonstration of ring conformational changes in triterpenes of the ß-amyrin class isolated from Stryphnodendron coriaceum. Journal of Organic Chemistry, [S.l.], v. 28, p. 2390-2394, 1963.
TURSCH, B. et al. Triterpenes II (*). Les sapogénines J et L de Stryphnodendron coriaceum. Bulletin des Sociétés Chimiques Belges, [S.l.], v.75, p.26-28, 1966. Apud: LOPES, G. C. et al. Determinação quantitativa de taninos em três espécies de Stryphnodendron por cromatografia líquida de alta eficiência. Brazilian Journal of Pharmaceutical Sciences, v. 45, n. 1, 2009.

108
VEIGA-JUNIOR, V. F. Estudo do consumo de plantas medicinais na Região Centro-Norte do Estado do Rio de Janeiro: aceitação pelos profissionais de saúde e modo de uso pela população. Revista Brasileira de Farmacognosia, [S.l.], v. 18, p. 308-313, 2008. VELLOSO, J. C. R. et al. Antioxidant activity of Maytenus ilicifolia root bark. Fitoterapia, [S.l.], v. 77, p 243-244, 2006. VELOZO, L. S. M. et al. Odoratin-7-O-β-d-glucopyranoside from Bowdichia virgilioides. Phytochemistry, [S.l.], v. 52, p. 1473–1477, 1999.
VIAL, H. J. et al. Prodrugs of bisthiazolium salts are orally potent antimalarials. Proceedings of the National Academy of Sciences, [S.l.], v. 101, n. 43, p.15458–15463, 2004.
VIGNERON, M. et al. Antimalarial remedies in French Guiana: A knowledge attitudes and practices study. Journal of Ethnopharmacology, [S.l.], v. 98, p.351-360, 2005. VILLALBA, M. A. et al. Atividades farmacológicas dos extratos de Zanthoxylum chiloperone (Rutaceae). Revista Brasileira de Farmacognonia, [S.l.], v. 17, n. 2, p. 236-241, 2007. VOSS, C. et al. Identification and characterization of riproximin, a new type II ribosome-inactivating protein with antineoplastic activity from Ximenia americana. FASEB Journal, [S.l.], v. 20, n. 8, p. 1194-6, 2006. VREDEN, S. S. The role of Kupffer cells in the clearance of malaria sporozoites from the circulation. Parasitology Today, [S.l.], v. 10, p.304–308, 1994. WARHURST, D. C. Antimalarial drugs: mode of action and resistance. Journal of Antimicrobial Chemotherapy, [S.l.], v. 18, p. 51-59, 1986. Suplemento B. WARHURST, D. C.; WILLIAMS, J. E. Laboratory diagnosis of malaria. Journal of Clinical Pathology, [S.l.], v. 49, n. 148, p. 533-538, jul. 1996. WEENEN, H. et al. Antimalarial compounds containing an alpha,beta-unsaturated carbonyl moiety from Tanzanian medicinal plants. Planta Medicinal, [S.l.], v. 56, p. 371-373, 1990.

109
WENIGER, B. et al. Evaluation ethnobotanically selected Benin medicinal plants for their in vitro antiplasmodial activity. Journal of Etnnonopharmacology, Benin, v. 90, n. 2-3, p. 279-284, fev. 2004. WHITE, N. J. Preventing antimalarial drug resistance through combinations. Drug Resistance Updates, [S.l.], v. 1, p. 3–9, 1998. WORLD HEALTH ORGANIZATION. A global strategy for malaria. [S.l.], 1993. WORLD HEALTH ORGANIZATION. Guidelines for the treatment of malaria. [S.l.], 2006. Disponível em: <http://apps.who.int/malaria/docs/TreatmentGuidelines2006.pdf>. Acesso em: 08 dez. 2008. WORLD HEALTH ORGANIZATION. Global Malaria Action Plan, [S.l.], 2009. Disponível em: <http://www.rbm.who.int/worldmalariaday/>. Acesso em: 14 jun. 2009. WORLD HEALTH ORGANIZATION. World malaria report 2008. [S.l.], 2008. Disponível em: <http://www.searo.who.int /LinkFiles/Reports_WMR_2008_Final.pdf>. Acesso em: 10 dez 2008. WORLD HEALTH ORGANIZATION. World malaria situation in 1994. Wkly Epidemiological Record, Geneva, v. 22, p. 161-167, set. 1997. WRIGHT, C. W. Traditional antimalarials and the development of novel antimalarial drugs. Journal of Ethnopharmacology, [S.l.], v. 100, p. 67–71, 2005. WRIGHT, E. M.; SNOPEK, B.; KOOPMAN, P. Seven new members of the Sox gene family expressed during mouse development. Nucleic Acids Research, [S.l.], v. 21, n. 3, 1993. YANG, S. et al. Synthesis and biological evaluation of analogues of cryptolepine, an alkaloid isolated from the Suriname rainforest. Journal of Natural Products, [S.l.], v. 62, p. 976-983, 1999. Apud: MONTENEGRO, L. H. M. et al. Terpenóides e avaliação do potencial antimalárico, larvicida, anti-radicalar e anticolinesterásico de Pouteria venosa
(Sapotaceae). Revista Brasileira de Farmacognosia, v. 16, p. 611-617, 2006.
ZORN, B. et al. 3-Desoxyanthocyanidins from Arrabidaea chica. Phytochemistry, [S.l.], v. 56, p. 831-835, 2001.

110
ZOCOLER, A. M. D. et al.. Antioxidant capacity of extracts and isolated compounds from Stryphnodendron obovatum Benth. Brazilian Journal of Pharmaceutical Sciences, [S.l.], v. 45, n. 3, 2009.

111
ANEXO

112
Artigo em preparação Avaliação citotóxica e antimalárica de Maytenus rigida (Celastraceae) Sideroxylon
obtusifolium (Sapotaceae) e Sideroxylon obtusifolium (Anacardiaceae)
Aline Mylena Gudes da Costa Oliveira 1; Valter Ferreira Andrade-Neto 2
1 Programa de Pós-graduação em Ciências Biológicas, Universidade Federal do Rio Grande
do Norte, UFRN, 59072-970,Natal, RN, Brazil; 2 Laboratório de Biologia da Malária e
Toxoplasmose - LABMAT, Departamento de Microbiologia e Parasitologia, Universidade
Federal do Rio Grande do Norte, UFRN, 59072-970, Natal, RN, Brazil.
* Corresponding author: [email protected] (VFAN), Tel.: +55 84 3215-3437, Fax : +55 84 3211- 9210.

113
Resumo O presente estudo investiga a atividade antimalárica de três espécies vegetais do Bioma da
Caatinga: Maytenus rigida (Celastraceae), Schinopsis brasiliensis (Anacardiaceae) e
Sideroxylon obtusifolium (Sapotaceae), através de avaliaçôes in vivo e in vitro. Nos testes in
vivo, camundongos Swiss foram infectados com a cepa NK65 de Plasmodium berghei e,
posteriormente, tratados com extrato alcoólico bruto (Teste de supressão de Peters, 1965); já
os testes in vitro consistiram em culturas sanguíneas com a cepa multirresistente K1 de P.
falciparum (Método de Trager e Jansen 1976), nos quais foi avaliada a ação esquizonticida
dos extratos ETHO e aquoso, além das frações clorofórmica, hexânica, acetato de etila e
hidrometanólica, cujas avaliações foram dadas de acordo com o protocolo de Andrade-Neto et
al. (2007). Os resultados da avaliação antimalárica in vivo mostraram que M. rigida foi
parcialmente ativa no quinto dia com reduções entre 30-45% para as doses testadas (250, 500
e 1000mg/kg) e sétimo dia na dose de 500mg/kg de 31,5%; Schinopsis brasiliensis foi
considerada ativa no quinto dia nas doses de 250 e 500mg/kg, com reduções de 86 e 95% e
Sideroxylon obtusifolium foi parcialmente ativa no sétimo dia na dose de 250mg/kg com
inibição de parasitemia de 33% e no quinto dia na dose de 500mg/kg de 56,2%. Já os
resultados in vitro mostraram inibições de parasitemia quando foram utilizadas as frações
clorofórmica e hexânica de M. rigida e S. brasiliensis, e apenas clorofórmica de S.
obtusifolium. Nesse contexto, pudemos observar que a busca de espécies cujas indicações
possam aliviar sintomas vistos também na malária foi uma ferramenta útil neste trabalho,
permitindo contribuir para o conhecimento científico do potencial antimalárico da flora
brasileira e deste modo, abrir perspectivas para o desenvolvimento de novos fármacos visando
melhor controle da doença.
Palavras-chaves: Plasmodium falciparum, Plasmodium berghei, malaria, etnofarmacologia,
produtos naturais.


















